Comparison of Promeristem Structure and Ontogeny of Procambium in Primary Roots of Zea mays ssp. Mexicana and Z. mays ‘Honey Bantam’ with Emphasis on Metaxylem Vessel Histogenesis

Abstract
:1. Introduction
2. Results
2.1. RNase Treatment before Staining
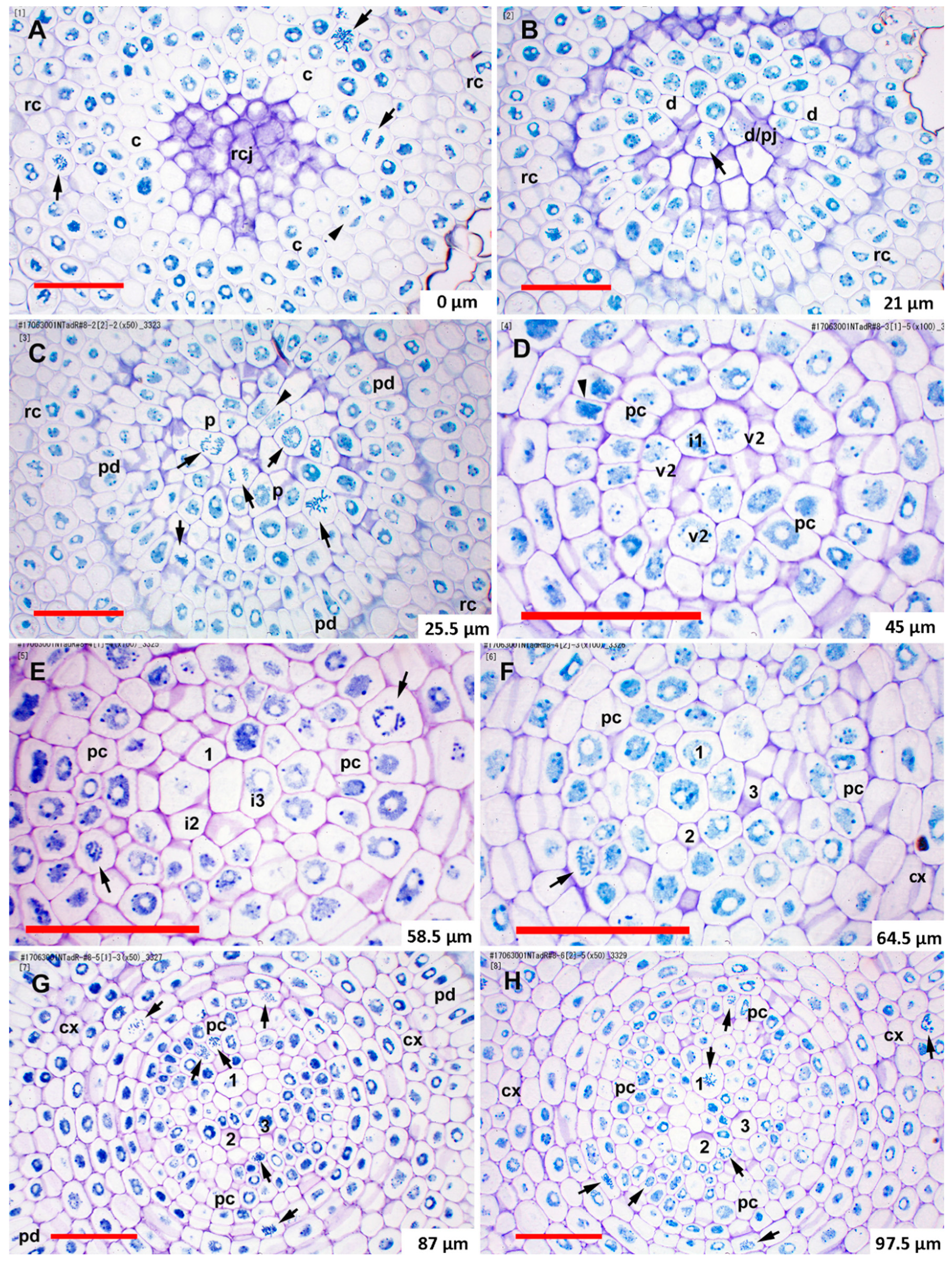
2.2. Histogen Organization in Teosinte and Sweetcorn
2.3. Mitotic Activity in the Promeristem
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Tissue Preparation for Light Microscopy (LM)
5.3. Cell Size Comparative Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LMX | late-maturing metaxylem vessel |
| QC | quiescent center |
| RCJ | root cap junction |
| RFLP | restriction fragment length polymorphism |
| RAM | root apical and primary meristem zones |
References
- Doebley, J. Molecular evidence and the evolution of maize. Econ. Bot. 1990, 44, 6–27. [Google Scholar] [CrossRef]
- Doebley, J. The genetics of maize evolution. Annu. Rev. Genet. 2004, 38, 37–59. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Diggle, P.K.; Friedman, W.E. Female gametophyte development and double fertilization in Balsas teosinte, Zea mays subsp. parviglumis (Poaceae). Sexual Plant Reprod. 2011, 24, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Burton, A.L.; Brown, K.M.; Lynch, J.P. Phenotypic diversity of root anatomical and architectural traits in Zea species. Crop Sci. 2013, 53, 1042–1055. [Google Scholar] [CrossRef]
- Koi, S.; Fujinami, R.; Kubo, N.; Tsukamoto, I.; Inagawa, R.; Imaichi, R.; Kato, M. Comparative anatomy of root meristem and root cap in some species of Podostemaceae and the evolution of root dorsiventrality. Am. J. Bot. 2006, 93, 682–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, L.J. Generation and elaboration of primary vascular tissue patterns in roots of Zea. Bot. Gaz. 1977, 138, 393–401. [Google Scholar] [CrossRef]
- Jiang, K.; Feldman, L.J. Regulation of root apical meristem development. Annu. Rev. Cell Dev. Biol. 2005, 21, 485–509. [Google Scholar] [CrossRef] [PubMed]
- Heimsch, C.; Seago, J.L., Jr. Organization of the root apical meristem in angiosperms. Am. J. Bot. 2008, 95, 1–21. [Google Scholar] [CrossRef]
- Guttenberg, H. von Grundzüge der histogenese höhere pflanzen. I. Die angiospermen. Handbuch der pflanzenanatomie. Band 8, Teil 3; Gebrüder Borntraeger: Berlin, Germany, 1960; pp. 72–103. [Google Scholar]
- Barlow, P.W. Towards an understanding of the behavior of root meristems. J. Theor. Biol. 1976, 57, 433–451. [Google Scholar] [CrossRef]
- Clowes, F.A.L. The difference between open and closed meristems. Ann. Bot. 1981, 48, 761–767. [Google Scholar] [CrossRef]
- Groot, E.P.; Doyle, J.A.; Nichol, S.A.; Rost, T.L. Phylogenetic distribution and evolution of root apical meristem organization in dicotyledonous angiosperms. Int. J. Plant Sci. 2004, 165, 97–105. [Google Scholar] [CrossRef]
- Von Hanstein, J. Die Scheitelzellegruppe im vegetationspunkt der phanerogamen. In Festschrift der Niederrheinischen Gesellschaft für Natur- und Heilkunde zum 50jährigen Jubiläum der Universität Bonn; Universität Bonn: Bonn, Germany, 1868; pp. 109–134. [Google Scholar]
- Clowes, F.A.L. The promeristem and the minimal constructional centre in grass root apices. New Phytol. 1954, 53, 108–116. [Google Scholar] [CrossRef]
- Armstrong, J.E.; Heimsch, C. Ontogenetic reorganization of the root meristem in the Compositae. Am. J. Bot. 1976, 63, 212–219. [Google Scholar] [CrossRef]
- Baum, S.F.; Dubrovsky, J.G.; Rost, T.L. Apical organization and maturation of the cortex and vascular cylinder in Arabidopsis thaliana (Brassicacea) roots. Am. J. Bot. 2002, 89, 908–920. [Google Scholar] [CrossRef] [PubMed]
- Dolan, L.; Janmaat, K.; Willemsen, V.; Linstead, P.; Poethig, S.; Roberts, K.; Scheres, B. Cellular organisation of the Arabidopsis thaliana root. Development 1993, 119, 71–84. [Google Scholar] [PubMed]
- Baum, S.F.; Rost, T.L. Root apical organization in Arabidopsis thaliana. I. Root cap and protoderm. Protoplasma 1996, 192, 178–188. [Google Scholar] [CrossRef]
- Ohashi-Ito, K.; Bergmann, D.C. Regulation of the Arabidopsis root vascular initial population by LONESOME HIGHWAY. Development 2007, 134, 2959–2968. [Google Scholar] [CrossRef]
- Miyashima, S.; Sebastian, J.; Lee, J.-Y.; Hellariuta, Y. Stem cell function during vascular development. EMBO J. 2013, 32, 178–193. [Google Scholar] [CrossRef]
- Ohashi-Ito, K.; Matsukawa, M.; Fukuda, H. An atypical bHLH transcription factor regulates early xylem development downstream of auxin. Plant Cell Physiol. 2013, 54, 398–405. [Google Scholar] [CrossRef]
- Smolarkiewicz, M.; Dhonukshe, P. Formative cell division: Principal determinants of plant morphogenesis. Plant Cell Physiol. 2013, 54, 333–342. [Google Scholar] [CrossRef]
- Zhang, Y.; Iakovidis, M.; Costa, S. Control of patterns of symmetric cell division in the epidermal and cortical tissues of the Arabidopsis root. Development 2016, 143, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Clowes, F.A.L. Development of quiescent centres in root meristems. New Phytol. 1958, 57, 85–88. [Google Scholar] [CrossRef]
- Clowes, F.A.L. Origin of the quiescent centre in Zea mays. New Phytol. 1978, 80, 409–419. [Google Scholar] [CrossRef]
- Morita, S.; Nemoto, K. Morphology and anatomy of rice roots with special reference to coordination in organo- and histogenesis. In Structure and Function of Roots: Proceedings of the Fourth International Symposium on Structure and Function of Roots; Baluska, F., Ciamporova, M., Gasparikova, O., Barlow, P., Eds.; Kluwer: Dordrecht, The Netherlands, 1995; pp. 75–86. [Google Scholar]
- Clowes, F.A.L. The growth fraction of the quiescent centre. New Phytol. 1982, 91, 129–135. [Google Scholar] [CrossRef]
- Heimsch, C. Development of vascular tissues in barley roots. Am. J. Bot. 1951, 38, 523–537. [Google Scholar] [CrossRef]
- Luxová, M. Some aspects of the differentiation of primary root tissues. In Structure and Function of Roots: Third Cabot Symposium; Torrey, J.G., Clarkson, D.T., Eds.; Academic Press: London, UK, 1975; pp. 73–89. [Google Scholar]
- Lux, A. A light microscopic study of the central metaxylem ontogenesis in the root of barley (Hordeum distichum L.). In Structure and Function of Roots: Proceedings of the 2nd International Symposium on Structure and Function of Roots; Brouwer, R., Gasparikova, O., Kolek, J., Loughman, B.C., Eds.; Martinus Nijhoff: The Hague, The Netherlands, 1981; pp. 71–76. [Google Scholar]
- Seago, J.L.; Wolniak, S.M. Cortical ontogeny in roots. I. Zea mays. Am. J. Bot. 1976, 63, 220–225. [Google Scholar] [CrossRef]
- Luxová, M. Growth region of the primary root of maize (Zea mays L.). In Structure and Function of Roots: Proceedings of the 2nd International Symposium on Structure and Function of Roots; Brouwer, R., Gasparikova, O., Kolek, J., Loughman, B.C., Eds.; Martinus Nijhoff: The Hague, The Netherlands, 1981; pp. 9–14. [Google Scholar]
- Barlow, P.W. Experimental modification of cell division patterns in the root meristem of Zea mays L. New Phytol. 1987, 105, 27–56. [Google Scholar] [CrossRef]
- Barlow, P.W. Cellular packets, cell division, and morphogenesis in the primary root of Zea mays L. Ann. Bot. 1989, 64, 13–20. [Google Scholar] [CrossRef]
- Rost, T.L. The organization of roots of dicotyledonous plants and the positions of control points. Ann. Bot. 2011, 107, 1213–1222. [Google Scholar] [CrossRef]
- Esau, K. Origin and development of primary vascular tissues in seed plants. Bot. Rev. 1943, 9, 125–206. [Google Scholar] [CrossRef]
- Kellicott, W.E. The daily periodicity of cell-division in the root of Allium. Bull. Torrey Bot. Club 1904, 31, 529–550. [Google Scholar] [CrossRef]
- Jensen, W.A.; Kavaljian, L.G. An analysis of cell morphology and the periodicity of division in the root tip of Allium cepa. Am. J. Bot. 1958, 45, 365–372. [Google Scholar] [CrossRef]
- Barlow, P.W.; Macdonald, P.D.M. An analysis of the mitotic cell cycle in the root meristem of Zea mays. Proc. R. Soc. Lond. B 1973, 183, 385–398. [Google Scholar]
- Gladish, D.K.; Niki, T. Factors inducing cavity formation in the vascular cylinders of pea roots (Pisum sativum L., cv. ‘Alaska’). Environ. Exp. Bot. 2000, 43, 1–9. [Google Scholar] [CrossRef]
- Niki, T.; Gladish, D.K.; Lu, P.; Rost, T.L. Cellular changes precede cavity formation in the vascular cylinder of pea roots (Pisum sativum L. cv. ‘Alaska’). Int. J. Plant Sci. 1995, 156, 290–302. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Histogen Heights (μm) at Center | Stele Diameters (μm) | |||||
|---|---|---|---|---|---|---|---|
| Dermatogen/Periblem | Plerome | First Vascular Initials Layer | Total ht | Plerome | @60μm | @100μm | |
| Teosinte | 15.9 ± 5.7 | 11.6 ± 3.9 | 12.1 ± 6.5 | 39.6 ± 15.1 | 56.0 ± 6.5 | 99.0 ± 5.7 | 129.9 ± 5.8 |
| Sweetcorn | 14.1 ± 2.4 | 9.4 ± 2.5 | 9.2 ± 2.9 | 32.7 ± 4.8 | 65.7 ± 6.1 | 130.0 ± 15.2 | 182.4 ± 9.7 |
| t | 0.77 | 1.27 | 1.08 | 1.16 | −2.90 | −5.14 | −12.29 |
| df | 8 | 10 | 8 | 7 | 11 | 7 | 9 |
| P | 0.464 | 0.234 | 0.311 | 0.285 | 0.014 | 0.001 | 0.000 |
| Taxon | Initiation of LMX Cell Files (µm) | Vascular Initials1 to 1st LMX (μm) | Distance Between 1st and 2nd LMX Initiation (μm) | ||
|---|---|---|---|---|---|
| First | Second | Third | |||
| Teosinte | 50.1 ± 19.7 | 54.5 ± 21.2 | 59.9 ± 19.7 | 10.4 ± 6.3 | 4.4 ± 2.6 |
| Sweetcorn | 37.9 ± 3.8 | 45.4 ± 4.1 | 48.2 ± 5.5 | 5.1 ± 4.7 | 5.1 ± 4.7 |
| t | 1.611 | 1.110 | 1.510 | −1.77 | −1.56 |
| df | 6 | 6 | 7 | 11 | 9 |
| P | 0.155 | 0.307 | 0.175 | 0.105 | 0.154 |
| Teosinte | ‘Honey Bantam’ Sweetcorn | ||||
|---|---|---|---|---|---|
| Teo 1 | Teo 2 | Teo 3 | HB 1 | HB 2 | HB 3 |
| 225.4 ± 56.0 | 194.6 ± 6.0 | 239 ± 46.7 | 268.3 ± 75.6 | 325.5 ± 46.0 | 318.1 ± 101.8 |
| n = 3 | n = 3 | n = 4 | n = 5 | n = 5 | n = 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saito, S.; Niki, T.; Gladish, D.K. Comparison of Promeristem Structure and Ontogeny of Procambium in Primary Roots of Zea mays ssp. Mexicana and Z. mays ‘Honey Bantam’ with Emphasis on Metaxylem Vessel Histogenesis. Plants 2019, 8, 162. https://doi.org/10.3390/plants8060162
Saito S, Niki T, Gladish DK. Comparison of Promeristem Structure and Ontogeny of Procambium in Primary Roots of Zea mays ssp. Mexicana and Z. mays ‘Honey Bantam’ with Emphasis on Metaxylem Vessel Histogenesis. Plants. 2019; 8(6):162. https://doi.org/10.3390/plants8060162
Chicago/Turabian StyleSaito, Susumu, Teruo Niki, and Daniel K. Gladish. 2019. "Comparison of Promeristem Structure and Ontogeny of Procambium in Primary Roots of Zea mays ssp. Mexicana and Z. mays ‘Honey Bantam’ with Emphasis on Metaxylem Vessel Histogenesis" Plants 8, no. 6: 162. https://doi.org/10.3390/plants8060162