1. Introduction
Since the resolution of the ANA grade [Amborellales, Nymphaeales, Austrobaileyales] as sister to all other flowering plants, most studies of anatomical traits that might be relevant to understanding the evolutionary development and phylogeny of angiosperms have been limited to multiple floral reproductive characters and to certain leaf and xylem traits [
1,
2]. However, it has been suggested that certain root traits might also be important to elucidate their elusive origin and rapid diversification [
3,
4,
5].
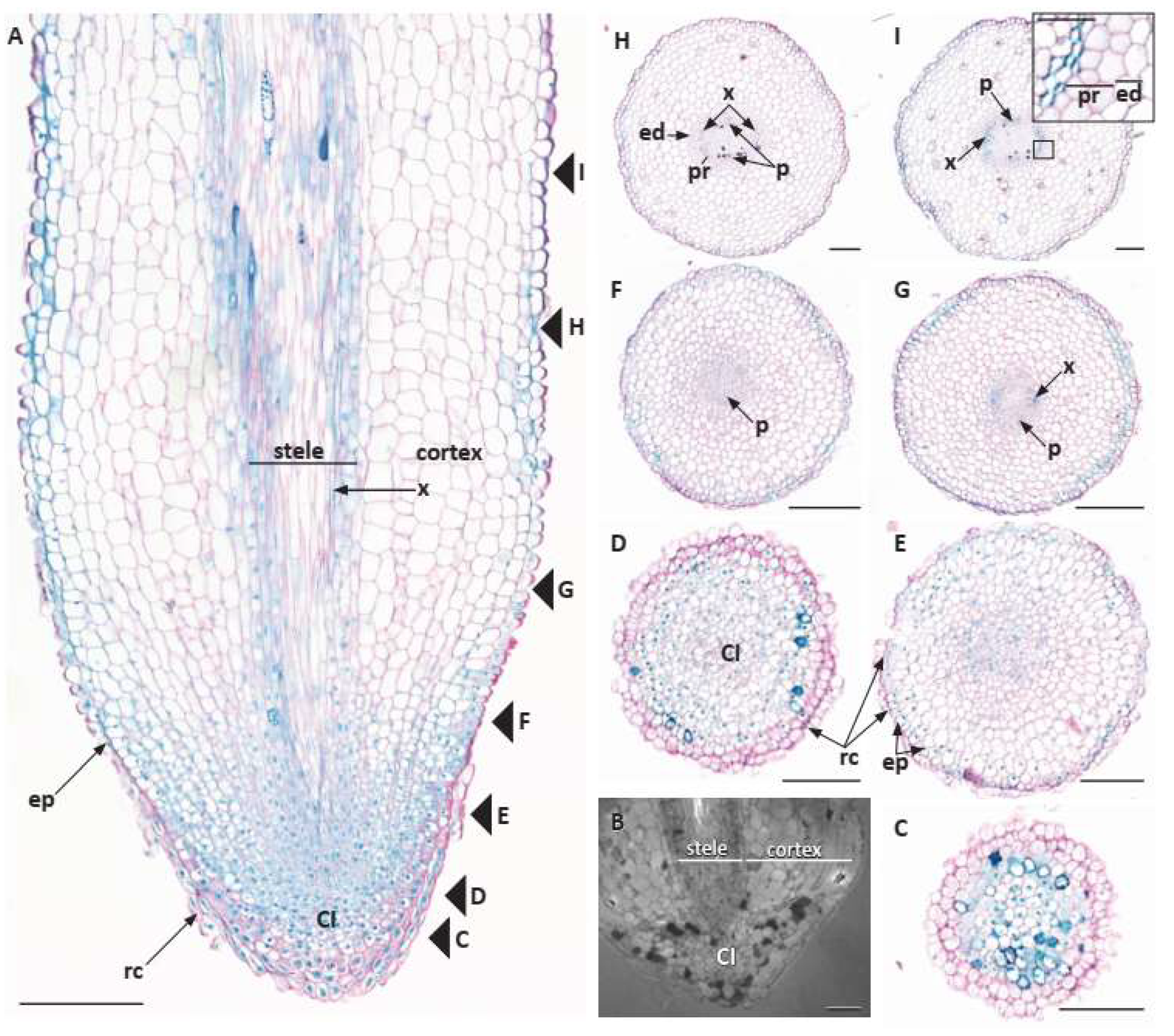
One significant root character is the occurrence of common initials, or CI, in root apical meristems, or RAM. In CIs, there is no clear and distinct boundary among the meristematic cells and tissues at the base of the root cap and the tips of the main root tissues, protoderm, ground meristem, and procambium, in the open RAMs of Amborellales and other woody species of the ANA grade (Austrobaileyales) and magnoliids (Laurales, Magnoliales, Canellales) [
4]. Furthermore, the RAMs of basal angiosperms turn out to be very similar to those in many gymnosperms, which are also known for having common initials for most tissues of the root apex [
6,
7,
8]. Indeed, Heimsch and Seago [
4] noted that gymnosperm CI RAMs, as reported by Pillai [
8], are also similar, when present, to those found in the more variable RAMs of Amborellales and Schisandraceae of the Austrobaileyales. However, with the exception of
Illicium, there is relatively little or no observation on the RAM and root developmental anatomy of other members of the Schisandraceae or Austrobaileyales, despite its position as sister to all other members of the order.
Austrobaileya scandens roots have received little attention. Bailey and Swamy [
9] studied
A. scandens but not its roots. Metcalfe and Chalk [
10] included
A. scandens as a member of the Schisandraceae and reported very little on it and nothing on its roots. Dickison and Endress [
11], Baranova [
12,
13], Feild et al. [
14], and Carpenter [
15,
16] dealt with various aspects of shoot anatomy, and Srivastava [
17] characterized gymnospermous features in its stem secondary phloem. It was not until 2001 that Carlquist [
18] reported on root wood, with vessel elements, tracheids, and fiber-tracheids, and bark structure. He touched upon the cortex, which he stated had compound starch grains and some oil cells, and secondary phloem, which was comprised of sieve cells and parenchyma, but lacked sieve tube elements and sclerenchyma.
Since nothing is known about Austrobaileya early root development and structure and RAM organization, our goals were to characterize them and the differentiation of primary tissues and secondary tissues during primary and early secondary root growth, and to evaluate if Austrobaileya shares more traits with other members of early-diverging lineages like Amborella, the closely related Illicium (Austrobaileyales), or the magnoliids. We were also interested to ascertain if they may still have some potentially new ancestral traits shared with extant and extinct gymnosperms (or other vascular plant lineages).
2. Materials and Methods
Material—About 30 adventitious and lateral roots of Austrobaileya scandens were collected from three 12-year old plants, which had been grown from seeds and cultivated in pots at the Weld Hill research building greenhouse in the Arnold Arboretum of Harvard University, Boston, Massachusetts. All roots were fixed in summer 2012 with 4% acrolein (Polysciences Inc., Warrington, PA, USA) in a modified PIPES buffer adjusted to pH 6.8 (50 mM PIPES and 1 mM MgSO4, both from BDH, London, UK; and 5 mM EGTA from Research Organics Inc., Cleveland, OH, USA) for 24 h. After rinsing with the same buffer, root material was dehydrated using an increasing ethanol dilution series and stored in 70% ethanol.
Scanning Electron microscopy (SEM)—Five roots were rehydrated and treated with 2% Osmium tetroxide for 2 h before complete re-dehydration using an increasing ethanol dilution series up to 100% and a transfer in acetone overnight. All material was further processed with an AutoSAMDRI-815 automatic critical point dryer (Tousimis, Rockville, MD, US) and specimens were mounted on aluminum stubs before observations.
Histological sections—About half of the roots were sectioned by hand using double-edged Wilkinson razor feather blades, while 10 were further dehydrated using dilutions up to a 100% ethanol solution, infiltrated and embedded in methacrylate resins (JB-4, Polysciences, Warrington, PA, USA, or Kulzer’s Technovit 7100, Electron Microscopy Sciences, Hatfield, PA, USA), and cut in 4–10 µm thick serial sections using a rotary microtome (Thermo Fisher Scientific, Waltham, MA, USA) and disposable blades (Leica Biosystems, Wetzlar, Germany). All sections were stained generally following Jensen [
19], either with saturated phloroglucinol-HCl for detection of lignin, berberine hemisulfate counterstained with 0.05% toluidine blue O (TBO) for detection of suberin, fluorol yellow for detection of lipids, calcofluor (i.e., Fluorescent Brightener 28) for detection of plant cell walls and fungal hyphae, Iodine Potassium (IKI) for detection of starch grains, or 0.05% TBO and 1.0% ruthenium red for general staining (all Sigma-Aldrich, Saint Louis, MO, USA). Resin sections were permanently mounted on glass slides using Citrisolv cleaning solution (Decon Labs, Inc., King of Prussia, PA, USA) and Permount (also Sigma-Aldrich) or Histomount (Agar Scientific, Stansted, United Kingdom) mounting media, and stored permanently at FU Berlin.
Image acquisition, processing and plates editing—Digital pictures of the roots in
Figure 1 were taken using the camera of a Samsung Galaxy S7 cell phone (Samsung Inc., Maetan-dong, South Korea). In SEM images, acquired with an Hitachi100 Scanning Electron Microscope (Hitachi Ltd, Tokyo, Japan), a black background has been edited to provide better contrast around the smooth contour of the roots. All hand and microtome sections were studied using a Zeiss LSM700 confocal microscope mounted with a Zeiss MRc Axiocam camera, and using brightfield, Differential Interference Contrast (DIC), or epifluorescence illumination, and the 405 nm and 488 nm solid-state lasers (Carl Zeiss AG, Oberkochen, Germany). All pictures and plates were edited and arranged using Adobe Creative Suite (Adobe, San Jose, CA, USA).
4. Discussion
We started this project because we were interested in possible pleisiomorphic characteristics in this representative of the ANA grade and basal angiosperms after the broad comparative studies of Clowes [
20] with 157 angiosperm species (including five Nymphaeales [Nymphaeaceae:
Nymphaea], two Austrobaileyales [Schisandraceae:
Schisandra,
Kadsura], 15 magnoliids, 65 monocots, and 70 eudicots), Groot et al. [
21] with 66 species (including two Nymphaeales [Nymphaeaceae:
Nymphaea], three magnoliids, and 61 eudicots), and Heimsch and Seago [
4] with 425 species (including
Amborella of the Amborellales, three Nymphaeales [Nymphaeaceae:
Nuphar and
Nymphaea], two Austrobaileyales [Schisandraceae:
Illicium,
Schisandra], eight magnoliids, 45 monocots, and 368 eudicots).
RAMs were consistently comprised of common initials in
A. scandens, whereas considerable variability was reported in roots of
Amborella and in
Illicium (Schisandraceae), the only other member of the Austrobaileyales that has been investigated to date [
4]. Nevertheless,
Amborella and
Illicium share some characteristics with
Austrobaileya in that some of their roots with open RAMs occasionally have a stronger resemblance to coniferous gymnosperm RAMs [
6,
8] than to other gymnosperms [
7]. Manifestations of the RAM with common initials in gymnosperms and some early-diverging lineages angiosperms, including members of the ANA grade and magnoliids, which also exhibit RAM common initials, must reflect the ancestral condition of this structural feature of roots and may be pleisiomorphic for seed plants as a whole [
4,
22].
In many respects,
A. scandens roots also exhibited primary growth generally similar to
Amborella and
Illicium roots [
5], although
A. scandens, along with
Illicium but unlike
Amborella, had tracheids and vessel elements [
18,
23,
24]. In
A. scandens, primary phloem with sieve cells was more prominent, and there was no triarchy or tetrarchy as observed in some
Amborella or
Illicium roots, respectively. Yet, all exhibit a cortex illustrative of differentiation from common initials and endodermis and exodermis [
5]. Although it has vessel elements, unlike
Amborella, the presence of common initials in the RAM and of phloem with sieve cells [
18] provides evidence of a pleisiomorphic state for
A. scandens.
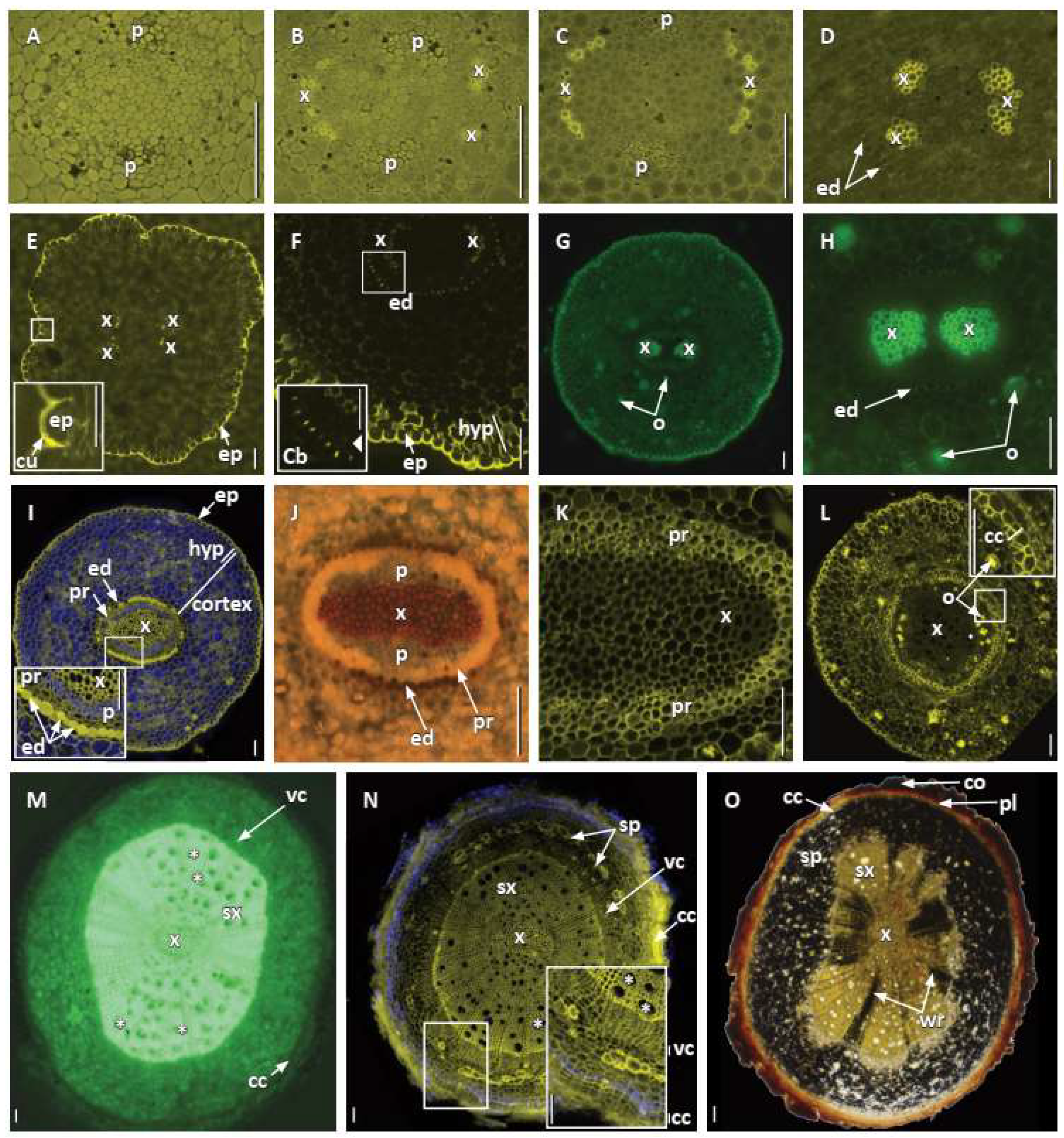
Early secondary growth in
A. scandens had vessel elements somewhat restricted to regions opposite the broad swath of the phloem poles [
18], whereas in
I. floridanum, solitary vessel elements were scattered throughout [
5]. In some ways, roots of
A. scandens were similar to the roots of
Amborella trichopoda, with a simple diarchy in which some protoxylem in each pole is slightly separated from other protoxylem of the same pole, probably by means of delayed maturation. Phloem was characterized by sieve cells and not sieve tube elements as found in most angiosperms, and this trait in
A. scandens was similar to the phloem in
Amborella [
18]. Additionally, the endodermis had similar Casparian bands and early production of suberin lamellae opposite broad phloem poles [
5]. Carlquist [
18] had previously identified tracheids and vessel elements in these roots, and as Bailey and Swamy [
9] had positively identified sieve cells. While Carlquist [
18] did not describe any sclerenchymatous cells in root secondary phloem in
A. scandens, we found scattered fibrous cells in the secondary phloem, similar to those found in stems. Similar to our findings, Carlquist [
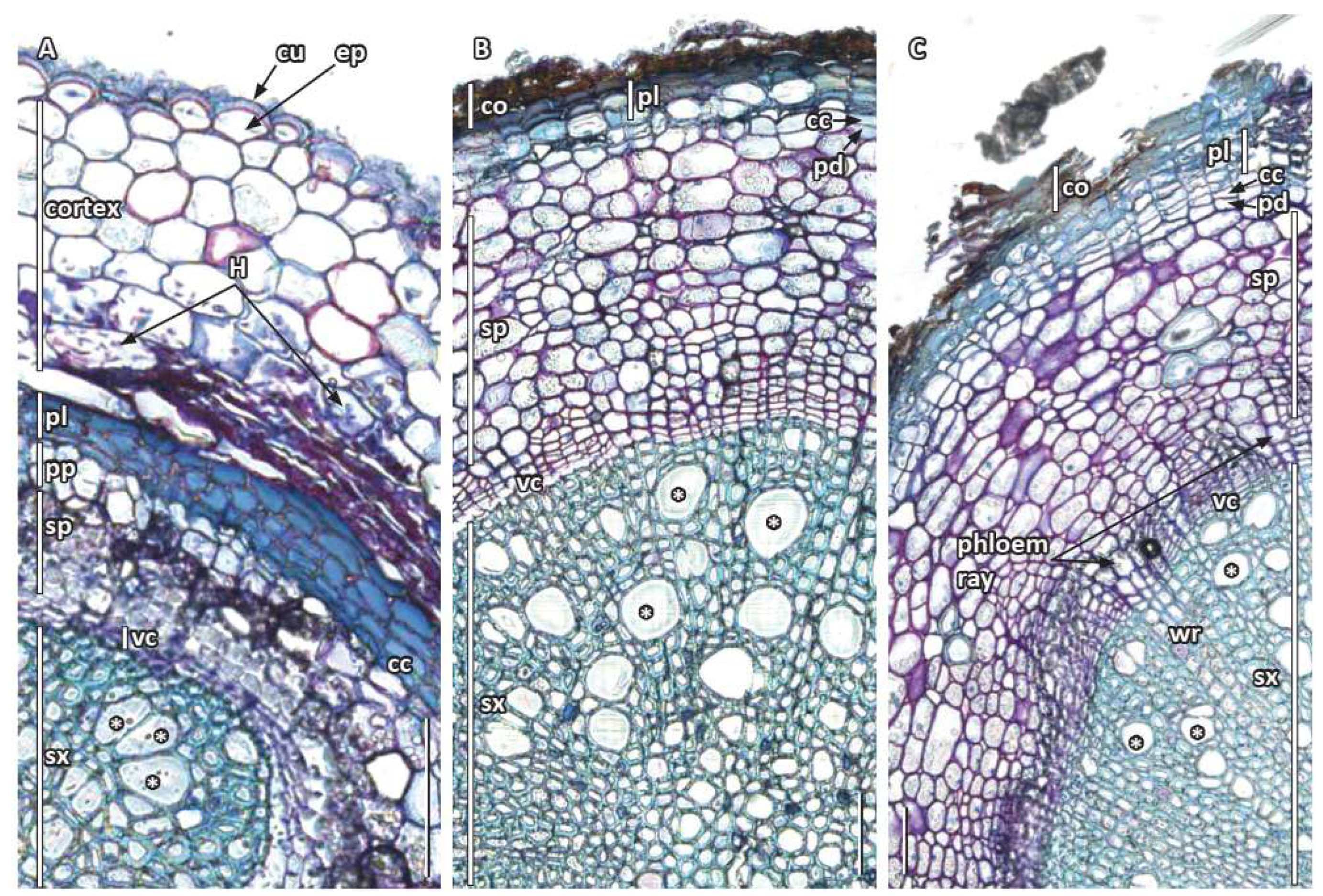
18] demonstrated a multiple-layered phelloderm under the phellogen and phellem.
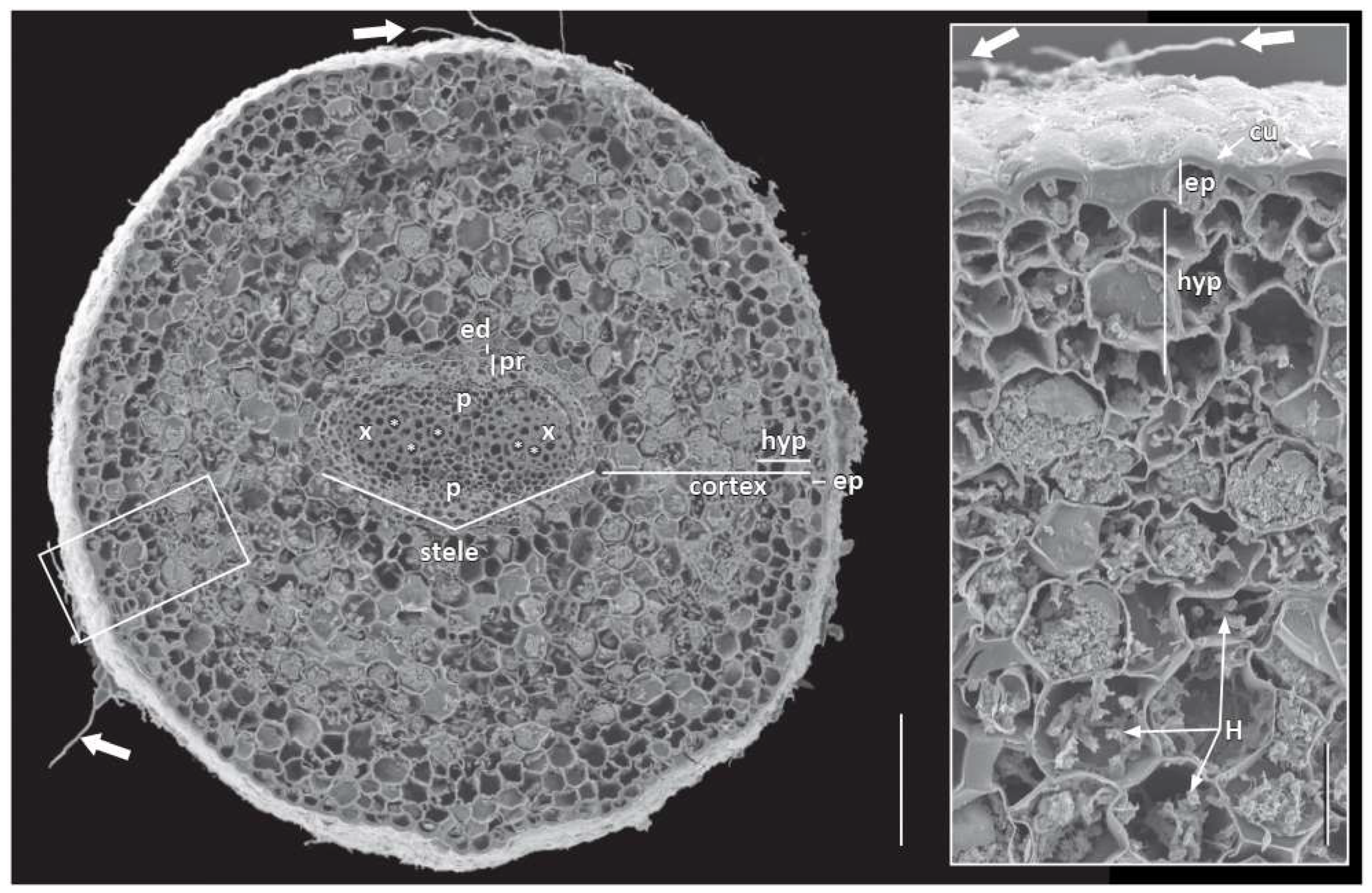
Another fascinating feature of adventitious roots in
A. scandens was the presence and thickness of a cuticle over the outer walls of epidermis and even its precursor protoderm and total absence of root hairs. The thick cuticle, as well as the presence of mycorrhizae, probably precluded the production of root hairs [
20,
25] and maybe a distally produced cuticle in roots contributed to or was a result of slow growth. However, that there were no trichoblasts does not mean that root hairs cannot be produced, like in roots of magnoliids where as Baylis [
26] noted, they often tend to be absent or, if present, appear to be slow-growing or stunted. Our study also revealed that starch grains accumulated during early primary root growth in the cortex and in secondary phloem, including phloem rays of older roots. Furthermore, since the cells of the middle cortex were pushed out beyond the secondary growth periderm, they in effect deposited outside the root any remnant starch, oil, and mycorrhizae still in those cells. While the presence of starch in roots of
A. scandens had already been reported in the field [
14], the presence of mycorrhizae was reported and illustrated here for the first time and needs to be confirmed in its natural habitat. Oil cells were found in the cortex of young roots, and Srivastava [
17] and Carlquist [
18] reported them in periderm of older roots. Oil droplets were also recently documented in the starchy endosperm of the seeds of
A. scandens [
27], which are by far the largest of the ANA grade and among the largest of magnoliids. Their composition and function remain unclear, although the presence of ethereal oils has been reported earlier in
Illicium and many other magnoliids, such as Aristolochiaceace or Piperaceae, and were easily perceived when crushing the roots of
A. scandens.
Two final points should be made. The distally thick cuticle may also prevent organisms in the rhizosphere from access to substances in the root during early root growth, and one of the possible outcomes of the disintegration of cortical cells with their contents of starch, oil, and mycorrhizae could be their availability to microorganisms in the soil. Lastly, as per Baylis [
14,
25], maybe this developmental trait of early angiosperms facilitated their survival and evolution.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




