
Tumor Biology and Microenvironment of Vestibular Schwannoma-Relation to Tumor Growth and Hearing Loss

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Methods
3. Peripheral Nerve Repair and Schwannoma Development
4. Involvement of Other Genes in the Biology of VS
5. First Proteomic Analyses of VSs
6. First Single-Cell RNA Sequencing of VS Tumor Cells
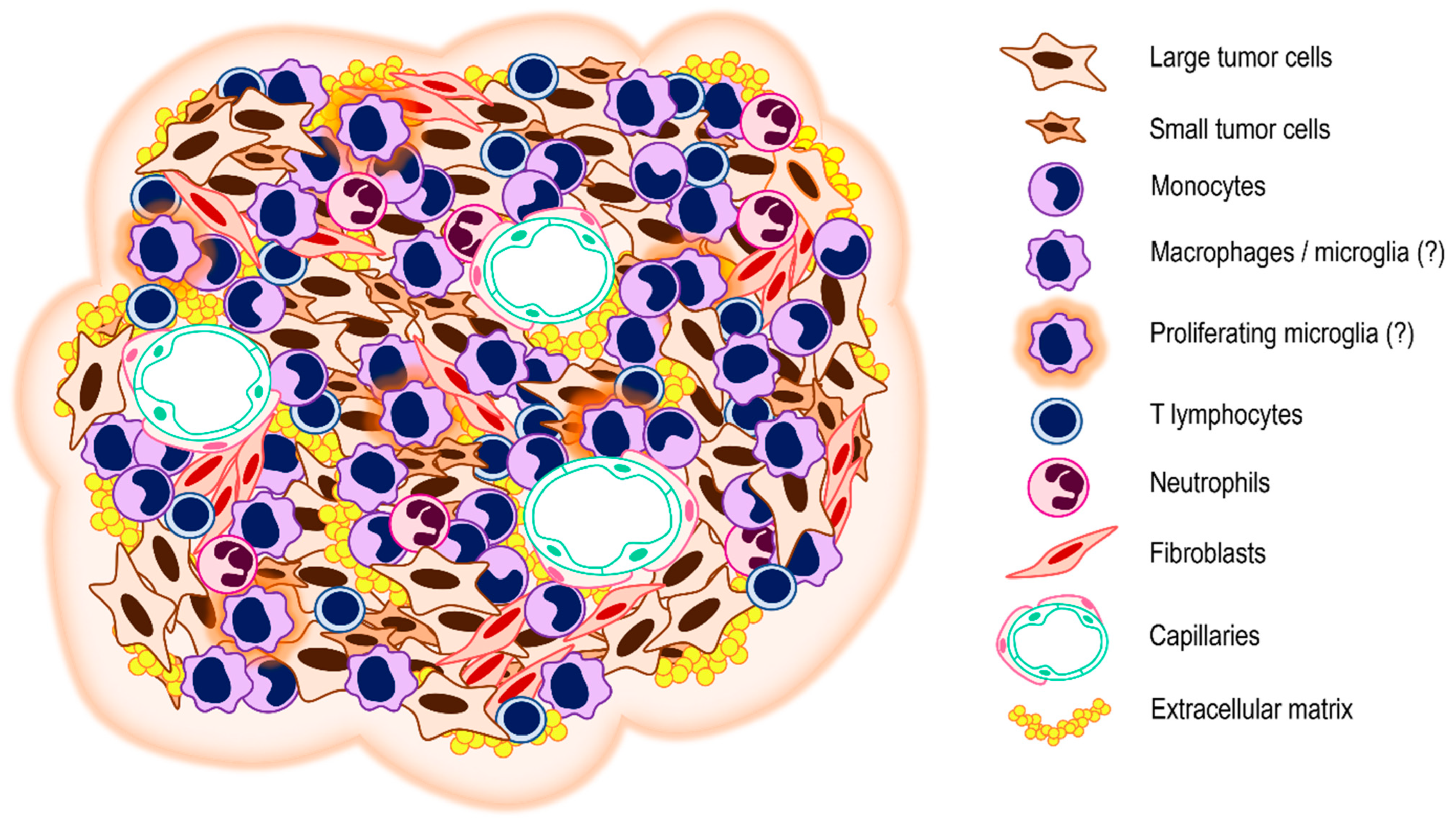
7. Tumor Microenvironment
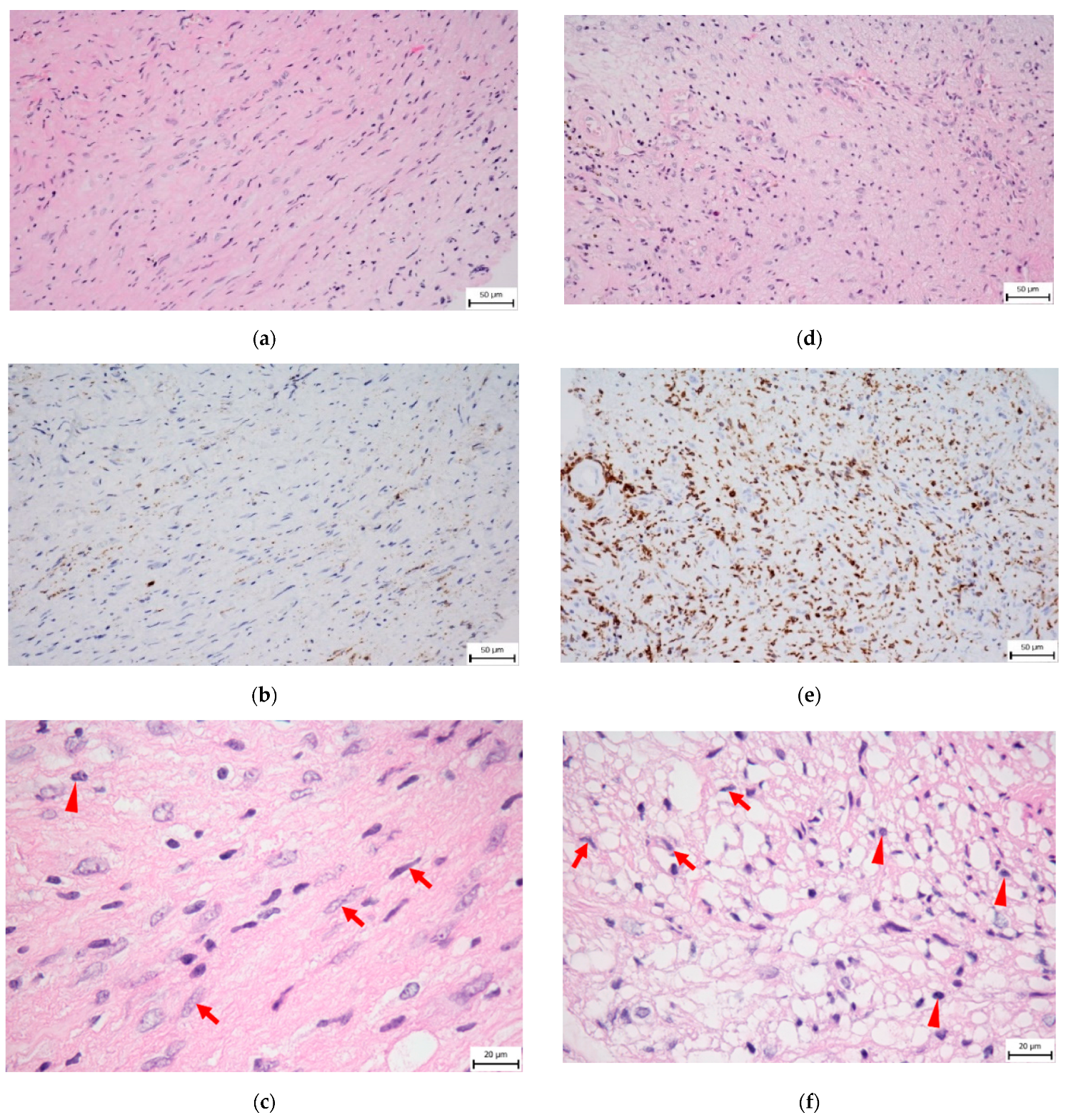
8. Cellular Components of the VS Stroma
9. Non-Cellular Components of the VS Stroma
10. Relation of the VS Biology and Microenvironment to Tumor Growth
10.1. NF2 and Other Modifications in Gene Expression
10.2. Differences in the Cellular Tumor Microenvironment
10.3. Differences in the Non-Cellular Tumor Microenvironment
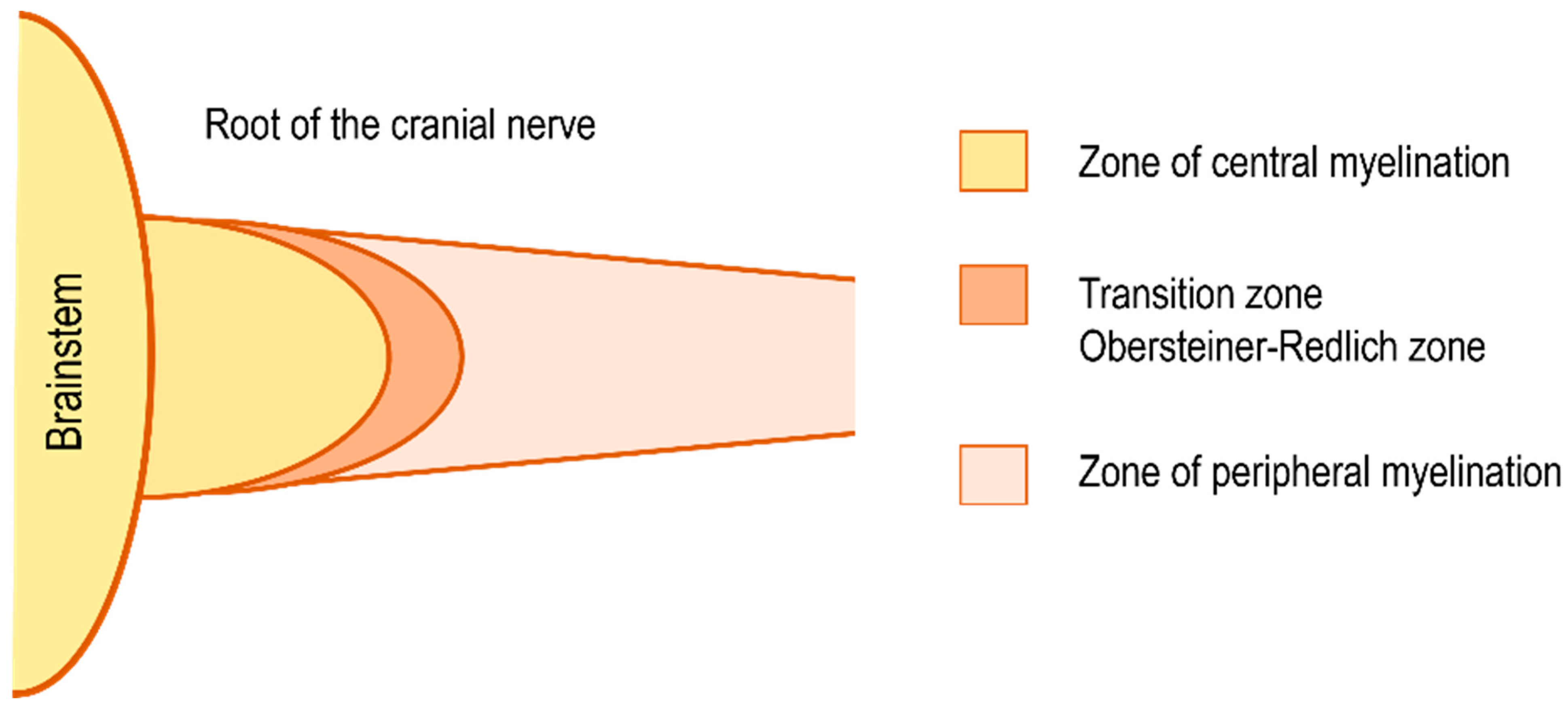
11. A New Concept of Hearing Loss in Patients with VS
12. Relation of the Biology and Microenvironment of VS to Hearing Loss
12.1. NF2 and Other Modifications in Gene Expression
12.2. Differences in the Cellular Tumor Microenvironment
12.3. Differences in the Non-Cellular Tumor Microenvironment
13. Clinical Application of the Knowledge of the Biology and Microenvironment of VS-Diagnosis and Targeted Therapy
14. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Constanzo, F.; Teixeira, B.C.D.A.; Sens, P.; Escuissato, D.; Ramina, R. Cerebellopontine Angle Schwannomas Arising from the Intermediate Nerve: A Scoping Review. Neurosurg. Rev. 2020, 43, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Koen, N.; Shapiro, C.; Kozin, E.D.; Cunnane, M.E.; Remenschneider, A.K.; McKenna, M.J.; Jung, D.H. Location of Small Intracanalicular Vestibular Schwannomas Based on Magnetic Resonance Imaging. Otolaryngol.-Head Neck Surg. 2020, 162, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Stangerup, S.E.; Tos, M.; Thomsen, J.; Caye-Thomasen, P. True Incidence of Vestibular Schwannoma? Neurosurgery 2010, 67, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Cutfield, S.W.; Wickremesekera, A.C.; Mantamadiotis, T.; Kaye, A.H.; Tan, S.T.; Stylli, S.S.; Itineang, T. Tumour Stem Cells in Schwannoma: A Review. J. Clin. Neurosci. 2019, 62, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Cho, Y.S. Growth of Vestibular Schwannoma: Long-Term Follow-up Study Using Survival Analysis. Acta Neurochir. 2021, 163, 2237–2245. [Google Scholar] [CrossRef]
- Lees, K.A.; Tombers, N.M.; Link, M.J.; Driscoll, C.L.; Neff, B.A.; van Gompel, J.J.; Lane, J.I.; Lohse, C.M.; Carlson, M.L. Natural History of Sporadic Vestibular Schwannoma: A Volumetric Study of Tumor Growth. Otolaryngol.-Head Neck Surg. 2018, 159, 535–542. [Google Scholar] [CrossRef]
- Hannan, C.J.; Lewis, D.; O’Leary, C.; Donofrio, C.A.; Evans, D.G.; Stapleton, E.; Freeman, S.R.; Lloyd, S.K.; Rutherford, S.A.; Hammerbeck-Ward, C.; et al. Beyond Antoni: A Surgeon’s Guide to the Vestibular Schwannoma Microenvironment. J. Neurol. Surg. Part B Skull Base 2022, 83, 1–10. [Google Scholar] [CrossRef]
- Nisenbaum, E.; Misztal, C.; Szczupak, M.; Thielhelm, T.; Peña, S.; Mei, C.; Goncalves, S.; Bracho, O.; Ma, R.; Ivan, M.E.; et al. Tumor-Associated Macrophages in Vestibular Schwannoma and Relationship to Hearing. OTO Open 2021, 5, 2473974X211059111. [Google Scholar] [CrossRef]
- Myrseth, E.; Pedersen, P.H.; Møller, P.; Lund-Johansen, M. Treatment of Vestibular Schwannomas. Why, When and How? Acta Neurochir. 2007, 149, 647–660. [Google Scholar] [CrossRef]
- Betka, J.; Chovanec, M.; Zverina, E.; Profant, O.; Lukes, P.; Skrivan, J.; Kluh, J.; Fik, Z. Minimally Invasive Endoscopic and Endoscopy-Assisted Microsurgery of Vestibular Schwannoma. In Advances in Endoscopic Surgery; IntechOpen: Rijeka, Croatia, 2011. [Google Scholar]
- Plotkin, S.R.; Merker, V.L.; Halpin, C.; Jennings, D.; McKenna, M.J.; Harris, G.J.; Barker, F.G., 2nd. Bevacizumab for Progressive Vestibular Schwannoma in Neurofibromatosis Type 2: A Retrospective Review of 31 Patients. Otol. Neurotol. 2012, 1046–1052. [Google Scholar] [CrossRef]
- Kim, G.; Hullar, T.E.; Seo, J.H. Comparison of Balance Outcomes According to Treatment Modality of Vestibular Schwannoma. Laryngoscope 2020, 130, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Yim, A.K.Y.; Wang, P.L.; Bermingham, J.R.; Hackett, A.; Strickland, A.; Miller, T.M.; Ly, C.; Mitra, R.D.; Milbrandt, J. Disentangling Glial Diversity in Peripheral Nerves at Single-Nuclei Resolution. Nat. Neurosci. 2022, 25, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, S.; Jiang, Y.; Wang, J.; Xiong, Y.; Dong, W.; Yao, Q.; Xing, Y.; Liu, F.; Chen, Z.; et al. Single-Cell RNA-Seq Reveals the Heterogeneity of Cell Communications between Schwann Cells and Fibroblasts within the Microenvironment in Vestibular Schwannoma. Am. J. Pathol. 2022, 192, 1230–1249. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Piao, X.; Bonaldo, P. Role of Macrophages in Wallerian Degeneration and Axonal Regeneration after Peripheral Nerve Injury. Acta Neuropathol. 2015, 130, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, R.E.; Echevarria, F.D. Macrophage Biology in the Peripheral Nervous System after Injury. Prog. Neurobiol. 2019, 173, 102–121. [Google Scholar] [CrossRef]
- Gomez-Sanchez, J.A.; Carty, L.; Iruarrizaga-Lejarreta, M.; Palomo-Irigoyen, M.; Varela-Rey, M.; Griffith, M.; Hantke, J.; Macias-Camara, N.; Azkargorta, M.; Aurrekoetxea, I.; et al. Schwann Cell Autophagy, Myelinophagy, Initiates Myelin Clearance from Injured Nerves. J. Cell Biol. 2015, 210, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Lutz, A.B.; Chung, W.S.; Sloan, S.A.; Carson, G.A.; Zhou, L.; Lovelett, E.; Posada, S.; Zuchero, J.B.; Barres, B.A. Schwann Cells Use TAM Receptor-Mediated Phagocytosis in Addition to Autophagy to Clear Myelin in a Mouse Model of Nerve Injury. Proc. Natl. Acad. Sci. USA 2017, 114, E8072–E8080. [Google Scholar] [CrossRef] [Green Version]
- Arthur-Farraj, P.J.; Latouche, M.; Wilton, D.K.; Quintes, S.; Chabrol, E.; Banerjee, A.; Woodhoo, A.; Jenkins, B.; Rahman, M.; Turmaine, M.; et al. C-Jun Reprograms Schwann Cells of Injured Nerves to Generate a Repair Cell Essential for Regeneration. Neuron 2012, 75, 633–647. [Google Scholar] [CrossRef] [Green Version]
- Hobson, M.I.; Green, C.J.; Terenghi, G. VEGF Enhances Intraneural Angiogenesis and Improves Nerve Regeneration after Axotomy. J. Anat. 2000, 197, 591–605. [Google Scholar] [CrossRef]
- Cattin, A.L.; Burden, J.J.; van Emmenis, L.; MacKenzie, F.E.; Hoving, J.J.A.; Garcia Calavia, N.; Guo, Y.; McLaughlin, M.; Rosenberg, L.H.; Quereda, V.; et al. Macrophage-Induced Blood Vessels Guide Schwann Cell-Mediated Regeneration of Peripheral Nerves. Cell 2015, 162, 1127–1139. [Google Scholar] [CrossRef]
- Birchmeier, C.; Nave, K.A. Neuregulin-1, a Key Axonal Signal That Drives Schwann Cell Growth and Differentiation. Glia 2008, 56, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Fry, E.J.; Ho, C.; David, S. A Role for Nogo Receptor in Macrophage Clearance from Injured Peripheral Nerve. Neuron 2007, 53, 649–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helbing, D.L.; Schulz, A.; Morrison, H. Pathomechanisms in Schwannoma Development and Progression. Oncogene 2020, 39, 5421–5429. [Google Scholar] [CrossRef] [PubMed]
- Wippold, F.J.; Lubner, M.; Perrin, R.J.; Lämmle, M.; Perry, A. Neuropathology for the Neuroradiologist: Antoni A and Antoni B Tissue Patterns. Am. J. Neuroradiol. 2007, 28, 1633–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, M.; Kawase, T.; Urano, M.; Mizoguchi, Y.; Kuroda, M.; Kasahara, M.; Suzuki, H.; Kanno, T. Analyses of Proliferative Potential in Schwannomas. Brain Tumor Pathol. 2000, 17, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Sian, C.S.; Ryan, S.F. The ultrastructure of neurilemoma with emphasis on antoni B tissue. Hum. Pathol. 1981, 12, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.G.R. Neurofibromatosis Type 2 (NF2): A Clinical and Molecular Review. Orphanet J. Rare Dis. 2009, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Petrilli, A.M.; Fernández-Valle, C. Role of Merlin/NF2 Inactivation in Tumor Biology. Oncogene 2016, 35, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Neff, B.A.; Bradley Welling, D.; Akhmametyeva, E.; Chang, L.-S. The Molecular Biology of Vestibular Schwannomas: Dissecting the Pathogenic Process at the Molecular Level. Otol. Neurotol. 2006, 27, 197–208. [Google Scholar] [CrossRef]
- Morrison, H.; Sherman, L.S.; Legg, J.; Banine, F.; Isacke, C.; Haipek, C.A.; Gutmann, D.H.; Ponta, H.; Herrlich, P. The NF2 Tumor Suppressor Gene Product, Merlin, Mediates Contact Inhibition of Growth through Interactions with CD44. Genes Dev. 2001, 15, 968–980. [Google Scholar] [CrossRef]
- Morrison, H.; Sperka, T.; Manent, J.; Giovannini, M.; Ponta, H.; Herrlich, P. Merlin/Neurofibromatosis Type 2 Suppresses Growth by Inhibiting the Activation of Ras and Rac. Cancer Res. 2007, 67, 520–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Yang, Z.; Chen, Y.; Zhou, Q.; Zhang, J.; Liu, J.; Wang, B.; He, Q.; Zhang, L.; Yu, Y.; et al. Deregulation of the Hippo Pathway Promotes Tumor Cell Proliferation Through YAP Activity in Human Sporadic Vestibular Schwannoma. World Neurosurg. 2018, 117, e269–e279. [Google Scholar] [CrossRef] [PubMed]
- Hilton, D.A.; Hanemann, C.O. Schwannomas and Their Pathogenesis. Brain Pathol. 2014, 24, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Schulz, A.; Büttner, R.; Hagel, C.; Baader, S.L.; Kluwe, L.; Salamon, J.; Mautner, V.F.; Mindos, T.; Parkinson, D.B.; Gehlhausen, J.R.; et al. The Importance of Nerve Microenvironment for Schwannoma Development. Acta Neuropathol. 2016, 132, 289–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, A.; Kyselyova, A.; Baader, S.L.; Jung, M.J.; Zoch, A.; Mautner, V.F.; Hagel, C.; Morrison, H. Neuronal Merlin Influences ERBB2 Receptor Expression on Schwann Cells through Neuregulin 1 Type III Signalling. Brain 2014, 137, 420–432. [Google Scholar] [CrossRef] [Green Version]
- Carlson, M.L.; Smadbeck, J.B.; Link, M.J.; Klee, E.W.; Vasmatzis, G.; Schimmenti, L.A. Next Generation Sequencing of Sporadic Vestibular Schwannoma: Necessity of Biallelic NF2 Inactivation and Implications of Accessory Non-NF2 Variants. Otol. Neurotol. 2018, 39, E860–E871. [Google Scholar] [CrossRef]
- Cayé-Thomasen, P.; Borup, R.; Stangerup, S.-E.; Thomsen, J.; Cilius Nielsen, F. Deregulated Genes in Sporadic Vestibular Schwannomas. Otol. Neurotol. 2010, 31, 256–266. [Google Scholar] [CrossRef] [Green Version]
- Welling, D.B.; Lasak, J.M.; Akhmametyeva, E.; Ghaheri, B.; Chang, L.-S. CDNA Microarray Analysis of Vestibular Schwannomas. Otol. Neurotol. 2002, 23, 736–748. [Google Scholar] [CrossRef]
- Aarhus, M.; Bruland, O.; Sætran, H.A.; Mork, S.J.; Lund-Johansen, M.; Knappskog, P.M. Global Gene Expression Profiling and Tissue Microarray Reveal Novel Candidate Genes and Down-Regulation of the Tumor Suppressor Gene CAV1 in Sporadic Vestibular Schwannomas. Neurosurgery 2010, 67, 998–1019. [Google Scholar] [CrossRef] [Green Version]
- Torres-Martin, M.; Lassaletta, L.; San-Roman-Montero, J.; de Campos, J.M.; Isla, A.; Gavilan, J.; Melendez, B.; Pinto, G.R.; Burbano, R.R.; Castresana, J.S.; et al. Microarray Analysis of Gene Expression in Vestibular Schwannomas Reveals SPP1/MET Signaling Pathway and Androgen Receptor Deregulation. Int. J. Oncol. 2013, 42, 848–862. [Google Scholar] [CrossRef]
- Casalino, L.; Verde, P. Multifaceted Roles of DNA Methylation in Neoplastic Transformation, from Tumor Suppressors to EMT and Metastasis. Genes 2020, 11, 922. [Google Scholar] [CrossRef] [PubMed]
- Madakashira, B.P.; Sadler, K.C. DNA Methylation, Nuclear Organization, and Cancer. Front. Genet. 2017, 8, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Gomez, P.; Bello, M.J.; Alonso, M.E.; Lomas, J.; Arjona, D.; de Campos, J.M.; Vaquero, J.; Isla, A.; Lassaletta, L.; Gutierrez, M.; et al. CpG Island Methylation in Sporadic and Neurofibromatis Type 2-Associated Schwannomas. Clin. Cancer Res. 2003, 9, 5601–5606. [Google Scholar] [PubMed]
- Lassaletta, L.; Bello, M.J.; Río, L.D.; Alfonso, C.; Roda, M.; Rey, A.; Gavilan, J. DNA Methylation of Multiple Genes in Vestibular Schwannoma: Relationship With Clinical and Radiological Findings. Otol. Neurotol. 2006, 27, 1180–1185. [Google Scholar] [CrossRef]
- Sass, H.; Cayé-Thomasen, P. Contemporary Molecular Biology of Sporadic Vestibular Schwannomas: A Systematic Review and Clinical Implications. J. Int. Adv. Otol. 2018, 14, 322–329. [Google Scholar] [CrossRef]
- Ahmed, S.G.; Abdelnabi, A.; Maguire, C.A.; Doha, M.; Sagers, J.E.; Lewis, R.M.; Muzikansky, A.; Giovannini, M.; Stemmer-Rachamimov, A.; Stankovic, K.M.; et al. Gene Therapy with Apoptosis-Associated Speck-like Protein, a Newly Described Schwannoma Tumor Suppressor, Inhibits Schwannoma Growth in Vivo. Neuro-Oncol. 2019, 21, 855–866. [Google Scholar] [CrossRef]
- Håvik, A.L.; Bruland, O.; Myrseth, E.; Miletic, H.; Aarhus, M.; Knappskog, P.M.; Lund-Johansen, M. Genetic Landscape of Sporadic Vestibular Schwannoma. J. Neurosurg. 2018, 128, 911–922. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Lu, D.; Gu, R.; Xu, Y.; Pan, R.; Bo, F.; Zhang, Y. Identification of Key Biomarkers and Immune Infiltration in Sporadic Vestibular Schwannoma Basing Transcriptome-Wide Profiling. World Neurosurg. 2022, 160, e591–e600. [Google Scholar] [CrossRef]
- Seo, J.-H.; Park, K.-H.; Jeon, E.-J.; Chang, K.-H.; Lee, H.; Lee, W.; Park, Y.-S. Proteomic Analysis of Vestibular Schwannoma: Conflicting Role of Apoptosis on the Pathophysiology of Sporadic Vestibular Schwannoma. Otol. Neurotol. 2015, 36, 714–719. [Google Scholar] [CrossRef]
- Taurone, S.; Bianchi, E.; Attanasio, G.; di Gioia, C.; Ierinó, R.; Carubbi, C.; Galli, D.; Pastore, F.S.; Giangaspero, F.; Filipo, R.; et al. Immunohistochemical Profile of Cytokines and Growth Factors Expressed in Vestibular Schwannoma and in Normal Vestibular Nerve Tissue. Mol. Med. Rep. 2015, 12, 737–745. [Google Scholar] [CrossRef]
- Sagers, J.E.; Sahin, M.I.; Moon, I.S.; Ahmed, S.G.; Stemmer-Rachamimov, A.; Brenner, G.J.; Stankovic, K.M. NLRP3 Inflammasome Activation in Human Vestibular Schwannoma: Implications for Tumor-Induced Hearing Loss. Hear. Res. 2019, 381, 107770. [Google Scholar] [CrossRef] [PubMed]
- Breun, M.; Schwerdtfeger, A.; Daniel Martellotta, D.; Kessler, A.F.; Perez, J.M.; Monoranu, C.M.; Ernestus, R.-I.; Matthies, C.; Löhr, M.; Hagemann, C. CXCR4: A New Player in Vestibular Schwannoma Pathogenesis. Oncotarget 2018, 9, 9940–9950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhang, Y.; Shi, Y.; Yin, D.; Dai, P.; Zhao, W.; Zhang, T. Identification of Predictive Proteins and Biological Pathways for the Tumorigenicity of Vestibular Schwannoma by Proteomic Profiling. Proteom. Clin. Appl. 2019, 13, e1800175. [Google Scholar] [CrossRef] [PubMed]
- Valach, J.; Fík, Z.; Strnad, H.; Chovanec, M.; Plzák, J.; Čada, Z.; Szabo, P.; Šáchová, J.; Hroudová, M.; Urbanová, M.; et al. Smooth Muscle Actin-Expressing Stromal Fibroblasts in Head and Neck Squamous Cell Carcinoma: Increased Expression of Galectin-1 and Induction of Poor Prognosis Factors. Int. J. Cancer 2012, 131, 2499–2508. [Google Scholar] [CrossRef]
- Fík, Z.; Dvořánková, B.; Kodet, O.; Bouček, J.; Betka, J.A.; Betka, J.; André, S.; Gabius, H.-J.; Šnajdr, P.; Smetana, K.; et al. Towards Dissecting Molecular Routes of Intercellular Communication in the Tumour Microenvironment: Phenotypic Plasticity of Stem Cell-Associated Markers in Co-Culture (Carcinoma Cell/Fibroblast) Systems. Folia Biol. 2014, 60, 205–212. [Google Scholar]
- Kolář, M.; Szabo, P.; Dvořánková, B.; Lacina, L.; Gabius, H.J.; Strnad, H.; Šáchová, J.; Vlček, Č.; Plzák, J.; Chovanec, M.; et al. Upregulation of IL-6, IL-8 and CXCL-1 Production in Dermal Fibroblasts by Normal/Malignant Epithelial Cells in Vitro: Immunohistochemical and Transcriptomic Analyses. Biol. Cell 2012, 104, 738–751. [Google Scholar] [CrossRef]
- Gál, P.; Varinská, L.; Fáber, L.; Novák, Š.; Szabo, P.; Mitrengová, P.; Mirossay, A.; Mučaji, P.; Smetana, K. How Signaling Molecules Regulate Tumor Microenvironment: Parallels to Wound Repair. Molecules 2017, 22, 1818. [Google Scholar] [CrossRef] [Green Version]
- Novák, Š.; Bandurová, V.; Mifková, A.; Kalfeřt, D.; Fík, Z.; Lukeš, P.; Szabo, P.; Plzák, J.; Smetana, K., Jr. Nádorové Mikroprostředí. Otoronolarygologie a Foniatrie 2019, 68, 41–51. [Google Scholar]
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-Associated Macrophages (TAM) as Major Players of the Cancer-Related Inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef]
- Chen, Y.; Song, Y.; Du, W.; Gong, L.; Chang, H.; Zou, Z. Tumor-Associated Macrophages: An Accomplice in Solid Tumor Progression. J. Biomed. Sci. 2019, 26, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Khodadoust, M.S.; Liu, C.L.; Newman, A.M.; Alizadeh, A.A. Profiling Tumor Infiltrating Immune Cells with CIBERSORT. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2018; Volume 1711, pp. 243–259. [Google Scholar]
- Filippou, P.S.; Karagiannis, G.S.; Constantinidou, A. Midkine (MDK) Growth Factor: A Key Player in Cancer Progression and a Promising Therapeutic Target. Oncogene 2020, 39, 2040–2054. [Google Scholar] [CrossRef] [PubMed]
- Nwabo Kamdje, A.H.; Seke Etet, P.F.; Kipanyula, M.J.; Vecchio, L.; Tagne Simo, R.; Njamnshi, A.K.; Lukong, K.E.; Mimche, P.N. Insulin-like Growth Factor-1 Signaling in the Tumor Microenvironment: Carcinogenesis, Cancer Drug Resistance, and Therapeutic Potential. Front. Endocrinol. 2022, 13, 927390. [Google Scholar] [CrossRef] [PubMed]
- Grassivaro, F.; Menon, R.; Acquaviva, M.; Ottoboni, L.; Ruffini, F.; Andrea Bergamaschi, X.; Muzio, L.; Farina, C.; Gianvito Martino, X. Convergence between Microglia and Peripheral Macrophages Phenotype during Development and Neuroinflammation. J. Neurosci. 2020, 40, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Jin, N.; Gao, L.; Fan, X.; Xu, H. Friend or Foe? Resident Microglia vs Bone Marrow-Derived Microglia and Their Roles in the Retinal Degeneration. Mol. Neurobiol. 2017, 54, 4094–4112. [Google Scholar] [CrossRef] [PubMed]
- Lisi, L.; Ciotti, G.M.P.; Braun, D.; Kalinin, S.; Currò, D.; dello Russo, C.; Coli, A.; Mangiola, A.; Anile, C.; Feinstein, D.L.; et al. Expression of INOS, CD163 and ARG-1 Taken as M1 and M2 Markers of Microglial Polarization in Human Glioblastoma and the Surrounding Normal Parenchyma. Neurosci. Lett. 2017, 645, 106–112. [Google Scholar] [CrossRef]
- Held-Feindt, J.; Rehmke, B.; Mentlein, R.; Hattermann, K.; Knerlich, F.; Hugo, H.H.; Ludwig, A.; Mehdorn, H.M. Overexpression of CXCL16 and Its Receptor CXCR6/Bonzo Promotes Growth of Human Schwannomas. Glia 2008, 56, 764–774. [Google Scholar] [CrossRef]
- Hong, B.; Krusche, C.A.; Schwabe, K.; Friedrich, S.; Klein, R.; Krauss, J.K.; Nakamura, M. Cyclooxygenase-2 Supports Tumor Proliferation in Vestibular Schwannomas. Neurosurgery 2011, 68, 1112–1117. [Google Scholar] [CrossRef]
- Hannan, C.J.; Lewis, D.; O’Leary, C.; Donofrio, C.A.; Evans, D.G.; Roncaroli, F.; Brough, D.; King, A.T.; Coope, D.; Pathmanaban, O.N. The Inflammatory Microenvironment in Vestibular Schwannoma. Neurooncol. Adv. 2020, 2, vdaa023. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.R.; Stemmer-Rachamimov, A.O.; Barker, F.G.; Halpin, C.; Padera, T.P.; Tyrrell, A.; Sorensen, A.G.; Jain, R.K.; di Tomaso, E. Hearing Improvement after Bevacizumab in Patients with Neurofibromatosis Type 2. New Engl. J. Med. 2009, 361, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.; Donofrio, C.A.; O’Leary, C.; Li, K.L.; Zhu, X.; Williams, R.; Djoukhadar, I.; Agushi, E.; Hannan, C.J.; Stapleton, E.; et al. The Microenvironment in Sporadic and Neurofibromatosis Type II–Related Vestibular Schwannoma: The Same Tumor or Different? A Comparative Imaging and Neuropathology Study. J. Neurosurg. 2021, 134, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Sass, H.C.R.; Borup, R.; Alanin, M.; Nielsen, F.C.; Cayé-Thomasen, P. Gene Expression, Signal Transduction Pathways and Functional Networks Associated with Growth of Sporadic Vestibular Schwannomas. J. Neurooncol. 2017, 131, 283–292. [Google Scholar] [CrossRef] [PubMed]
- de Vries, M.; Hogendoorn, P.C.W.; de Bruyn, I.B.; Malessy, M.J.A.; van der Mey, A.G.L. Intratumoral Hemorrhage, Vessel Density, and the Inflammatory Reaction Contribute to Volume Increase of Sporadic Vestibular Schwannomas. Virchows Arch. 2012, 460, 629–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vries, M.d.; Briaire-De Bruijn, I.; Malessy, J.A.; de Brubne, F.T.; van der Mey, G.L.; Hogendoorn, P.C.W. Tumor-Associated Macrophages Are Related to Volumetric Growth of Vestibular Schwannomas. Otol. Neurotol. 2013, 34, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.; Roncaroli, F.; Agushi, E.; Mosses, D.; Williams, R.; Li, K.L.; Zhu, X.; Hinz, R.; Atkinson, R.; Wadeson, A.; et al. Inflammation and Vascular Permeability Correlate with Growth in Sporadic Vestibular Schwannoma. Neuro-Oncol. 2019, 21, 314–325. [Google Scholar] [CrossRef]
- Graffeo, C.S.; Perry, A.; Raghunathan, A.; Kroneman, T.N.; Jentoft, M.; Driscoll, C.L.; Neff, B.A.; Carlson, M.L.; Jacob, J.; Link, M.J.; et al. Macrophage Density Predicts Facial Nerve Outcome and Tumor Growth after Subtotal Resection of Vestibular Schwannoma. J. Neurol. Surg. Part B Skull Base 2018, 79, 482–488. [Google Scholar] [CrossRef]
- Perry, A.; Graffeo, C.S.; Carlstrom, L.P.; Raghunathan, A.; Driscoll, C.L.W.; Neff, B.A.; Carlson, M.L.; Parney, I.F.; Link, M.J.; van Gompel, J.J. Predominance of M1 Subtype among Tumor-Associated Macrophages in Phenotypically Aggressive Sporadic Vestibular Schwannoma. J. Neurosurg. 2020, 133, 1637–1645. [Google Scholar] [CrossRef]
- Gonçalves, V.M.; Suhm, E.M.; Ries, V.; Skardelly, M.; Tabatabai, G.; Tatagiba, M.; Schittenhelm, J.; Behling, F. Macrophage and Lymphocyte Infiltration Is Associated with Volumetric Tumor Size but Not with Volumetric Growth in the Tübingen Schwannoma Cohort. Cancers 2021, 13, 466. [Google Scholar] [CrossRef]
- de Vries, W.M.; Briaire-de Bruijn, I.H.; van Benthem, P.P.G.; van der Mey, A.G.L.; Hogendoorn, P.C.W. M-CSF and IL-34 Expression as Indicators for Growth in Sporadic Vestibular Schwannoma. Virchows Arch. 2019, 474, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Behling, F.; Suhm, E.; Ries, V.; Gonçalves, V.M.; Tabatabai, G.; Tatagiba, M.; Schittenhelm, J. COX2 Expression Is Associated with Preoperative Tumor Volume but Not with Volumetric Tumor Growth in Vestibular Schwannoma. Neurol. Res. Pr. 2021, 3, 11. [Google Scholar] [CrossRef]
- Cayé-Thomasen, P.; Werther, K.; Nalla, A.; Bøg-Hansen, T.C.; Jørgen Nielsen, H.; Stangerup, S.-E.; Thomsen, J. VEGF and VEGF Receptor-1 Concentration in Vestibular Schwannoma Homogenates Correlates to Tumor Growth Rate. Otol. Neurotol. 2005, 26, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Cayé-Thomasen, P.; Baandrup, L.; Jacobsen, G.K.; Thomsen, J.; Stangerup, S.-E. Immunohistochemical Demonstration of Vascular Endothelial Growth Factor in Vestibular Schwannomas Correlates to Tumor Growth Rate. Laryngoscope 2003, 113, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Møller, M.N.; Werther, K.; Nalla, A.; Stangerup, S.E.; Thomsen, J.; Bøg-Hansen, T.C.; Nielsen, H.J.; Cayé-Thomasen, P. Angiogenesis in Vestibular Schwannomas: Expression of Extracellular Matrix Factors MMP-2, MMP-9, and TIMP-1. Laryngoscope 2010, 120, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Graamans, K.; van Dijk, J.E.; Janssen, L.W. Hearing Deterioration in Patients with a Non-Growing Vestibular Schwannoma. Acta Otolaryngol. 2003, 123, 51–54. [Google Scholar] [CrossRef]
- Fisher, L.M.; Doherty, J.K.; Lev, M.H.; Slattery, W.H. Concordance of Bilateral Vestibular Schwannoma Growth and Hearing Changes in Neurofibromatosis 2: Neurofibromatosis 2 Natural History Consortium. Otol. Neurotol. 2009, 30, 835–841. [Google Scholar] [CrossRef]
- Lassaletta, L.; Calvino, M.; Morales-Puebla, J.M.; Lapunzina, P.; Rodriguez-de la Rosa, L.; Varela-Nieto, I.; Martinez-Glez, V. Biomarkers in Vestibular Schwannoma–Associated Hearing Loss. Front. Neurol. 2019, 10, 978. [Google Scholar] [CrossRef] [Green Version]
- Asthagiri, A.R.; Vasquez, R.A.; Butman, J.A.; Wu, T.; Morgan, K.; Brewer, C.C.; King, K.; Zalewski, C.; Kim, H.J.; Lonser, R.R. Mechanisms of Hearing Loss in Neurofibromatosis Type 2. PLoS ONE 2012, 7, e46132. [Google Scholar] [CrossRef]
- Karch-Georges, A.; Veillon, F.; Vuong, H.; Rohmer, D.; Karol, A.; Charpiot, A.; Meyer, N.; Venkatasamy, A. MRI of Endolymphatic Hydrops in Patients with Vestibular Schwannomas: A Case-Controlled Study Using Non-Enhanced T2-Weighted Images at 3 Teslas. Eur. Arch. Oto-Rhino-Laryngol. 2019, 276, 1591–1599. [Google Scholar] [CrossRef]
- Gan, J.; Zhang, Y.; Wu, J.; Lei, D.; Zhang, F.; Zhao, H.; Wang, L. Current Understanding of Hearing Loss in Sporadic Vestibular Schwannomas: A Systematic Review. Front. Oncol. 2021. [Google Scholar] [CrossRef]
- Carlson, M.L.; Vivas, E.X.; McCracken, D.J.; Sweeney, A.D.; Neff, B.A.; Shepard, N.T.; Olson, J.J. Congress of Neurological Surgeons Systematic Review and Evidence-Based Guidelines on Hearing Preservation Outcomes in Patients with Sporadic Vestibular Schwannomas. In Proceedings of the Clinical Neurosurgery; Oxford University Press: Oxford, UK, 2018; Volume 82, pp. E35–E39. [Google Scholar]
- Lassaletta, L.; Patrón, M.; del Río, L.; Alfonso, C.; Maria Roda, J.; Rey, J.A.; Gavilan, J. Cyclin D1 Expression and Histopathologic Features in Vestibular Schwannomas. Otol. Neurotol. 2007, 28, 939–941. [Google Scholar] [CrossRef]
- Stankovic, K.M.; Mrugala, M.; Martuza, L.; Silver, K.; Betensky, A.; Nadol, B.; Stemmer-Rachamimov, O.; Stankovic, K.M. Genetic Determinants of Hearing Loss Associated With Vestibular Schwannomas. Otol. Neurotol. 2009, 30, 661–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassaletta, L.; Torres-Martín, M.; Peña-Granero, C.; Roda, J.M.; Santa-Cruz-Ruiz, S.; Castresana, J.S.; Gavilan, J.; Rey, J.A. NF2 Genetic Alterations in Sporadic Vestibular Schwannomas: Clinical Implications. Otol. Neurotol. 2013, 34, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Dilwali, S.; Lysaght, A.; Roberts, D.; Barker, F.G., 2nd; Mckenna, J.; Stankovic, M. Sporadic Vestibular Schwannomas Associated With Good Hearing Secrete Higher Levels of Fibroblast Growth Factor 2 Than Those Associated With Poor Hearing Irrespective of Tumor Size. Otol. Neurotol. 2013, 34, 748–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilwali, S.; Landegger, L.D.; Soares, V.Y.R.; Deschler, D.G.; Stankovic, K.M. Secreted Factors from Human Vestibular Schwannomas Can Cause Cochlear Damage. Sci. Rep. 2015, 5, 18599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotkin, S.R.; Duda, D.G.; Muzikansky, A.; Allen, J.; Blakeley, J.; Rosser, T.; Campian, J.L.; Clapp, D.W.; Fisher, M.J.; Tonsgard, J.; et al. Multicenter, Prospective, Phase II and Biomarker Study of High-Dose Bevacizumab as Induction Therapy in Patients With Neurofibromatosis Type 2 and Progressive Vestibular Schwannoma. J. Clin. Oncol. 2019, 37, 3446–3454. [Google Scholar] [CrossRef] [PubMed]
- Soares, V.Y.R.; Atai, N.A.; Fujita, T.; Dilwali, S.; Sivaraman, S.; Landegger, L.D.; Hochberg, F.H.; Oliveira, C.A.P.C.; Bahmad, F.; Breakefield, X.O.; et al. Extracellular Vesicles Derived from Human Vestibular Schwannomas Associated with Poor Hearing Damage Cochlear Cells. Neuro Oncol. 2016, 18, 1498–1507. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Hyakusoku, H.; Sagers, J.E.; Landegger, L.D.; Welling, D.B.; Stankovic, K.M. MMP-14 (MT1-MMP) Is a Biomarker of Surgical Outcome and a Potential Mediator of Hearing Loss in Patients With Vestibular Schwannomas. Front. Cell. Neurosci. 2020, 14, 191. [Google Scholar] [CrossRef] [PubMed]
- Breun, M.; Monoranu, C.M.; Kessler, A.F.; Matthies, C.; Löhr, M.; Hagemann, C.; Schirbel, A.; Rowe, S.P.; Pomper, M.G.; Buck, A.K.; et al. [68Ga]-Pentixafor PET/CT for CXCR4-Mediated Imaging of Vestibular Schwannomas. Front. Oncol. 2019. [Google Scholar] [CrossRef]
- Fujii, M.; Ichikawa, M.; Iwatate, K.; Bakhit, M.; Yamada, M.; Kuromi, Y.; Sato, T.; Sakuma, J.; Saito, K. Bevacizumab Therapy of Neurofibromatosis Type 2 Associated Vestibular Schwannoma in Japanese Patients. Neurol. Med. Chir. 2020, 60, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, Q.; Ma, H.; Sun, T.; Xiang, R.; Di, F. Therapeutic Effect and Side Effects of Bevacizumab Combined with Irinotecan in the Treatment of Paediatric Intracranial Tumours: Meta-Analysis and Systematic Review. J. Clin. Pharm. Ther. 2020, 45, 1363–1371. [Google Scholar] [CrossRef]
- Fujii, M.; Kobayakawa, M.; Saito, K.; Inano, A.; Morita, A.; Hasegawa, M.; Mukasa, A.; Mitsuhara, T.; Goto, T.; Yamaguchi, S.; et al. Rationale and Design of Beatnf2 Trial: A Clinical Trial to Assess the Efficacy and Safety of Bevacizumab in Patients with Neurofibromatosis Type 2 Related Vestibular Schwannoma. Curr. Oncol. 2021, 28, 726–739. [Google Scholar] [CrossRef] [PubMed]
- Tamura, R.; Toda, M. A Critical Overview of Targeted Therapies for Vestibular Schwannoma. Int. J. Mol. Sci. 2022, 23, 5462. [Google Scholar] [CrossRef] [PubMed]
- Kandathil, C.K.; Cunnane, M.E.; McKenna, M.J.; Curtin, H.D.; Stankovic, K.M. Correlation Between Aspirin Intake and Reduced Growth of Human Vestibular Schwannoma: Volumetric Analysis. Otol. Neurotol. 2016, 37, 1428–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinelli, J.P.; Lees, K.A.; Tombers, N.M.; Lohse, C.M.; Carlson, M.L. Impact of Aspirin and Other NSAID Use on Volumetric and Linear Growth in Vestibular Schwannoma. Otolaryngol. Neck Surg. 2019, 160, 1081–1086. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author | Year | Number of Patients | Analytical Method | Conclusions |
|---|---|---|---|---|
| NF2 and other modifications in gene expression | ||||
| Sass et al. [74] | 2017 | 16 sVS | Microarray analysis |
|
| Differences in the cellular tumor microenvironment | ||||
| De Vries et al. [75] | 2012 | 67 sVS | CD45, CD68 (IHC) |
|
| De Vries et al. [76] | 2013 | 10 slow-growing sVS 10 fast-growing sVS | CD163 (IHC) |
|
| Graffeo et al. [78] | 2018 | 46 sVS after STR 33 stable 13 progressive | CD68 (IHC) |
|
| Lewis et al. [77] | 2019 | 5 growing sVS 3 static sVS | Iba-1 (IHC) |
|
| Perry et al. [79] | 2019 | 44 sVS after STR 32 stable 12 progressive | CD163 (IHC) |
|
| Goncalves et al. [80] | 2021 | 923 sVS volumetric growth analysis in 189 sVS | CD3, CD8, CD68, CD163 (IHC) |
|
| Differences in the non-cellular tumor microenvironment | ||||
| Cayé-Thomasen et al. [84] | 2003 | 15 sVS | VEGF (IHC) |
|
| Cayé-Thomasen et al. [83] | 2005 | 27 sVS | VEGF, VEGFR-1 (concentrations) |
|
| Møller et al. [85] | 2010 | 34 sVS | MMP-2, MMP-9, TIMP-1 (IHC and concentrations) |
|
| De Vries et al. [81] | 2019 | 10 fast-growing sVS 10 slow-growing sVS | M-CSF, IL-34, CD163 (IHC) |
|
| Behling et al. [82] | 2021 | 898 sVS volumetric growth analysis 171 sVS | COX2 (IHC) |
|
| Author | Year | Number of Patients | Analytical Method | Conclusions |
|---|---|---|---|---|
| NF2 and other modifications in gene expression | ||||
| Lassaletta et al. [46] | 2006 | 21 sVS, 1 NF2-VS | Methylation-specific PCR |
|
| Lassaletta et al. [94] | 2007 | 21 sVS | Cyclin D1 (IHC) |
|
| Stankovic et al. [95] | 2009 | 9 sVS—good hearing 4 sVS—poor hearing | Microarray analysis |
|
| Lassaletta et al. [96] | 2013 | 51 sVS | PCR/dHPLC direct sequencing |
|
| Differences in the cellular tumor microenvironment | ||||
| Sagers et al. [53] | 2019 | 15 sVS—good hearing 15 sVS—poor hearing 7 GAN controls | qRT-PCR IL-1β, NLRP3, CD68 (IHC) |
|
| Nisenbaum et al. [8] | 2021 | 12 sVS—good hearing 15 sVS—poor hearing 3 NF2-VS—poor hearing | CD80, CD163 (IHC) |
|
| Differences in the non-cellular tumor microenvironment | ||||
| Dilwali et al. [97] | 2013 | 16 sVS—good hearing 19 sVS—poor hearing 7 GAN controls | Cytokine array ELISA of secretions |
|
| Dilwali et al. [98] | 2015 | 3 sVS—good hearing 10 sVS—poor hearing 2 GAN controls | ELISA of secretions Murine cochlear explant culture |
|
| Soares et al. [100] | 2016 | 3 sVS—good hearing 3 sVS—poor hearing | Extracellular vesicle analysis Murine cochlear explant culture |
|
| Ren et al. [101] | 2020 | 4 sVS—good hearing 19 sVS—poor hearing 6 GAN controls | MMP-14 (IHC, activity in VS secretions, activity in plasma) Murine cochlear explant culture |
|
| Breun et al. [102] | 2018 | 10 NF2-VS—good hearing 8 NF2-VS—moderate hearing 12 NF2-VS—poor hearing 8 sVS—good hearing 14 sVS—moderate hearing 8 sVS—poor hearing | CXCR4 (mRNA-PCR, IHC, Western blotting) |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tesařová, M.; Peterková, L.; Šťastná, M.; Kolář, M.; Lacina, L.; Smetana, K., Jr.; Hynek, R.; Betka, J.; Vlasák, A.; Lukeš, P.; et al. Tumor Biology and Microenvironment of Vestibular Schwannoma-Relation to Tumor Growth and Hearing Loss. Biomedicines 2023, 11, 32. https://doi.org/10.3390/biomedicines11010032
Tesařová M, Peterková L, Šťastná M, Kolář M, Lacina L, Smetana K Jr., Hynek R, Betka J, Vlasák A, Lukeš P, et al. Tumor Biology and Microenvironment of Vestibular Schwannoma-Relation to Tumor Growth and Hearing Loss. Biomedicines. 2023; 11(1):32. https://doi.org/10.3390/biomedicines11010032
Chicago/Turabian StyleTesařová, Michaela, Lenka Peterková, Monika Šťastná, Michal Kolář, Lukáš Lacina, Karel Smetana, Jr., Radovan Hynek, Jan Betka, Aleš Vlasák, Petr Lukeš, and et al. 2023. "Tumor Biology and Microenvironment of Vestibular Schwannoma-Relation to Tumor Growth and Hearing Loss" Biomedicines 11, no. 1: 32. https://doi.org/10.3390/biomedicines11010032