
Physiological, Morphological and Antioxidant Responses of Pediococcus pentosaceus R1 and Lactobacillus fermentum R6 Isolated from Harbin Dry Sausages to Oxidative Stress


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions and Colony Counts
2.2. Intracellular ROS Levels
2.3. Intracellular ATP Concentration
2.4. Electron Microscopy Analysis
2.5. Electric Conductivity
2.6. Assessment of Antioxidant Activity
2.6.1. Intact Cell Suspension and Intracellular Cell-Free Extract Preparation
2.6.2. 2′,2′-Diphenyl-1-Picrylhydrazyl Radical Scavenging Assay
2.6.3. Hydroxyl Radical Scavenging Assay
2.6.4. Superoxide Radical Scavenging Rates Assay
2.6.5. Reducing Power Assay
2.6.6. Measurement of Antioxidant Enzyme Activities
2.7. Statistical Analysis
3. Results and Discussion
3.1. Survival of the Strains under Oxidative Stress
3.2. Intracellular ROS Levels
3.3. Intracellular ATP Concentrations
3.4. SEM Analysis
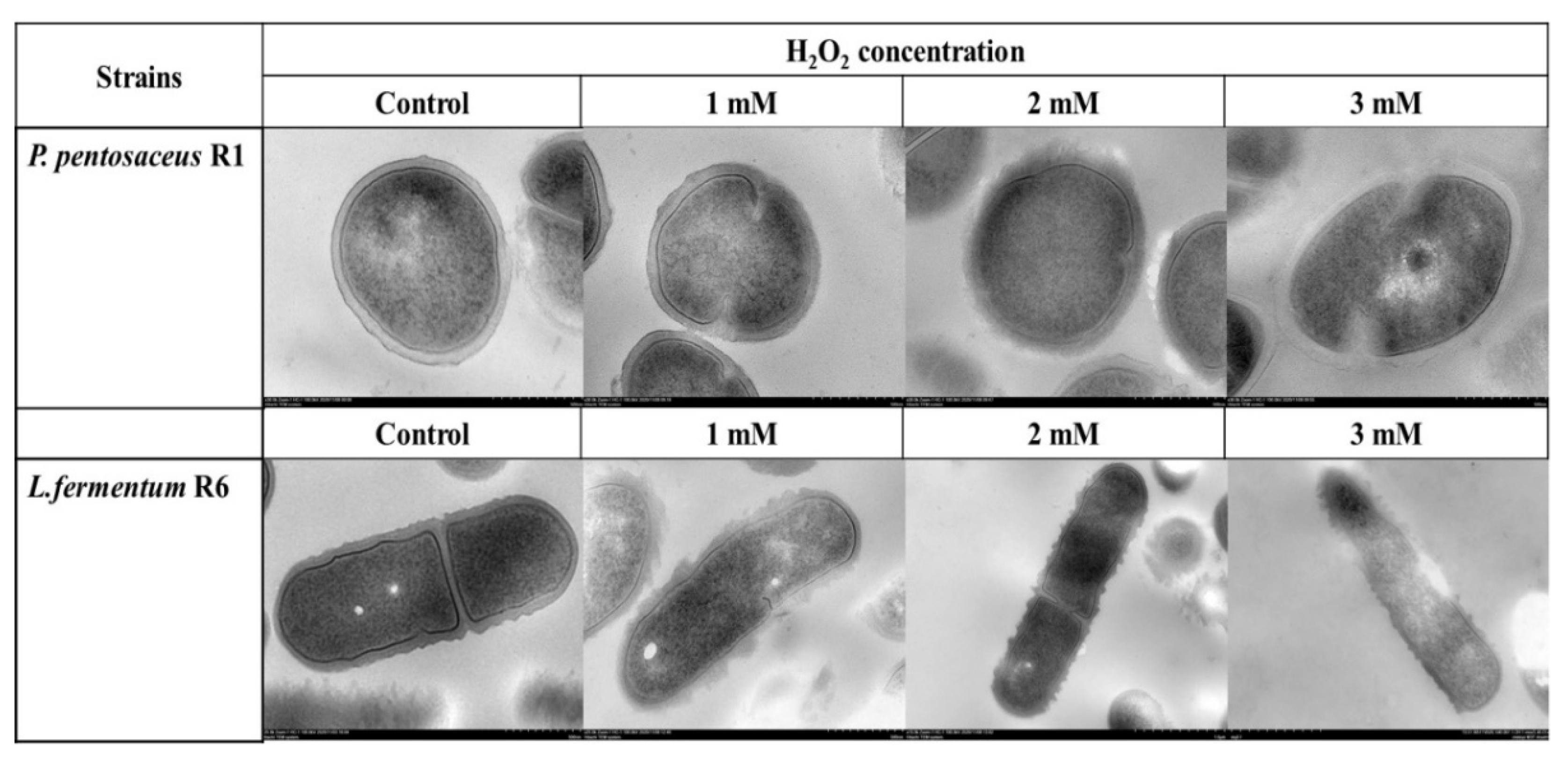
3.5. TEM Analysis
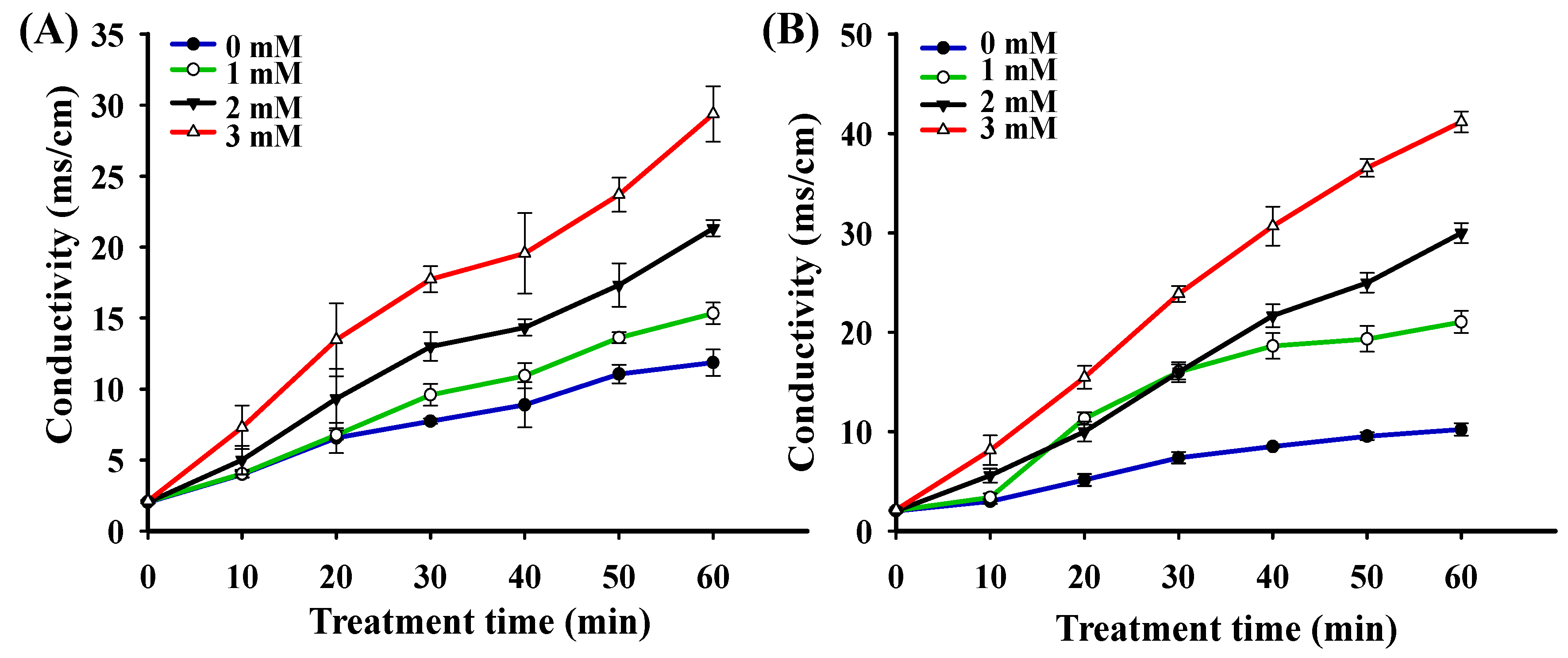
3.6. Extracellular Electrolytes
3.7. Free Radical Scavenging Rates and Reducing Power
3.8. Antioxidant Enzyme Activities Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bekhit, M.; Arabtehrany, E.; Kahn, C.J.F.; Cleymand, F.; Fleutot, S.; Desobry, S.; Sanchezgonzalez, L. Bioactive films containing alginate-pectin composite microbeads with Lactococcus lactis subsp. lactis: Physicochemical characterization and antilisterial activity. Int. J. Mol. Sci. 2018, 19, 574. [Google Scholar] [CrossRef] [Green Version]
- Evivie, S.E.; Huo, G.; Igene, J.O.; Bian, X. Some current applications, limitations and future perspectives of lactic acid bacteria as probiotics. Food Nutr. Res. 2017, 61, 1318034. [Google Scholar] [CrossRef] [Green Version]
- Chávarri, M.; Marañón, I.; Ares, R.; Ibáñez, F.C.; Marzo, F.; Villarán, M.C. Microencapsulation of a probiotic and prebiotic in alginate-chitosan capsules improves survival in simulated gastro-intestinal conditions. Int. J. Food Microbiol. 2010, 142, 185–189. [Google Scholar] [CrossRef]
- Chen, Q.; Kong, B.; Sun, Q.; Dong, F.; Liu, Q. Antioxidant potential of a unique LAB culture isolated from Harbin dry sausage: In vitro and in a sausage model. Meat Sci. 2015, 110, 180–188. [Google Scholar] [CrossRef]
- Peng, K.; Koubaab, M.; Balsa, O.; Vorobieva, E. Recent insights in the impact of emerging technologies on lactic acid bacteria: A review. Food Res. Int. 2020, 137, 109544. [Google Scholar] [CrossRef]
- Chen, Q.; Kong, B.; Han, Q.; Xia, X.; Xu, L. The role of bacterial fermentation in lipolysis and lipid oxidation in Harbin dry sausages and its flavour development. LWT 2017, 77, 389–396. [Google Scholar] [CrossRef]
- Neffeskocinska, K.; Jaworska, D.; Kolozynkrajewska, D.; Dolatowski, Z.J.; Jachaczjowko, L. The effect of lab as probiotic starter culture and green tea extract addition on dry fermented pork loins quality. Biomed. Res. Int. 2015, 2015, 452757. [Google Scholar] [CrossRef]
- Spyropoulos, B.G.; Misiakos, E.P.; Fotiadis, C.; Stoidis, C.N. Antioxidant properties of probiotics and their protective effects in the pathogenesis of radiation-induced enteritis and colitis. Digest. Dis. Sci. 2011, 56, 285–294. [Google Scholar] [CrossRef]
- Amaretti, A.; Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, V.; Quiberoni, A.; Reinhemer, J.; Suarez, V. Resistance of functional lactobacillus plantarum strains against food stress conditions. Food Microbiol. 2015, 48, 63–71. [Google Scholar] [CrossRef]
- Mills, S.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Enhancing the stress responses of probiotics for a lifestyle from gut to product and back again. Microb. Cell Fact. 2011, 10, S19. [Google Scholar] [CrossRef] [Green Version]
- Ianni, F.; Altomare, A.A.; Cenci-Goga, B.T.; Blasi, F.; Cossignani, L. Chromatographic characterization and in vitro bioactivity evaluation of Lactobacillus helveticus hydrolysates upon fermentation of different substrates. Appl. Sci. 2021, 11, 811. [Google Scholar] [CrossRef]
- Grispoldi, L.; Giglietti, R.; Traina, G.; Cenci-Goga, B. How to assess in vitro probiotic viability and the correct use of neutralizing agents. Front. Microbiol. 2020, 11, 204. [Google Scholar] [CrossRef] [Green Version]
- Akhova, A.V.; Sekatskaya, P.A.; Tkachenko, A.G. Formation of associated oxidative stress in cells of Escherichia coli exposed to different environmental stressors. Appl. Biochem. Microbiol. 2019, 55, 582–587. [Google Scholar] [CrossRef]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Alegria, A.; Bron, P.A.; De Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress physiology of lactic acid bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef] [Green Version]
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic metabolism in the genus Lactobacillus: Impact on stress response and potential applications in the food industry. J. Appl. Microbiol. 2017, 122, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zotta, T.; Ricciardi, A.; Ciocia, F.; Rossano, R.; Parente, E. Diversity of stress responses in dairy thermophilic streptococci. Int. J. Food Microbiol. 2008, 124, 34–42. [Google Scholar] [CrossRef]
- Leite, M.C.T.; Troxell, B.; Bruno-Bárcena, J.M.; Hassan, H.M. Biology of Reactive Oxygen Species, Oxidative Stress, and Antioxidants in Lactic Acid Bacteria; Caister Academic Press: Wymondham, UK, 2015; pp. 205–218. [Google Scholar]
- Zhao, Y.C.; Li, J.L.; Chen, Y.R.; Hang, H.X. Response to oxidative stress of Coriolus versicolor induced by exogenous hydrogen peroxide and paraquat. Ann. Microbiol. 2009, 59, 221–227. [Google Scholar] [CrossRef]
- Bai, Z.H.; Harvey, L.M.; McNeil, B. Physiological responses of chemostat cultures of Aspergillus niger (B1-D) to simulated and actual oxidative stress. Biotechnol. Bioeng. 2003, 82, 691–701. [Google Scholar] [CrossRef]
- Han, Q.; Kong, B.; Chen, Q.; Sun, F.; Zhang, H. In vitro comparison of probiotic properties of lactic acid bacteria isolated from harbin dry sausages and selected probiotics. J. Funct. Foods. 2017, 32, 391–400. [Google Scholar] [CrossRef]
- Sanhueza, E.; Paredes-Osses, E.; Gonzalez, C.L.; Garcia, A. Effect of pH in the survival of Lactobacillus salivarius strain UCO_979C wild type and the pH acid acclimated variant. Electron. J. Biotechnol. 2015, 18, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Ngamdee, W.; Tandhavanant, S.; Wikraiphat, C.; Reamtong, O.; Wuthiekanun, V.; Salje, J.; Low, D.A.; Peacock, S.J.; Chantratita, N. Competition between Burkholderia pseudomallei and B. thailandensis. BMC Microbiol. 2015, 15, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Qian, L.H.; Xie, J. Effect of chitosan on membrane permeability and cell morphology of Pseudomonas aeruginosa and Staphyloccocus aureus. Carbohyd. Polym. 2011, 86, 969–974. [Google Scholar] [CrossRef]
- Wang, A.N.; Yi, X.W.; Yu, H.F.; Dong, B.; Qiao, S.Y. Free radical scavenging activity of Lactobacillus fermentum in vitro and its antioxidative effect on growing-finishing pigs. J. Appl. Microbiol. 2009, 107, 1140–1148. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.; Pang, X.; Yao, W.; Gao, X. Characterization and antioxidant activity of two low-molecular-weight polysaccharides purified from the fruiting bodies of Ganoderma lucidum. Int. J. Biol. Macromol. 2010, 46, 451–457. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction--antioxidative activities of products of browning reaction prepared from glucosamine. J. Acad. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Arcanjo, N.O.; Andrade, M.J.; Padilla, P.; Rodriguez, A.; Madruga, M.S.; Estevez, M. Resveratrol protects Lactobacillus reuteri against H2O2-induced oxidative stress and stimulates antioxidant defenses through upregulation of the dhaT gene. Free Radic. Biol. Med. 2019, 135, 38–45. [Google Scholar] [CrossRef]
- Qian, J.; Wang, C.; Zhuang, H.; Zhang, J.; Yan, W. Oxidative stress responses of pathogen bacteria in poultry to plasma-activated lactic acid solutions. Food Control 2020, 118, 107355. [Google Scholar] [CrossRef]
- Serrazanetti, D.I.; Guerzoni, M.E.; Corsetti, A.; Vogel, R. Metabolic impact and potential exploitation of the stress reactions in lactobacilli. Food Microbiol. 2009, 26, 700–711. [Google Scholar] [CrossRef]
- Liu, J.; Wisniewski, M.; Droby, S.; Norelli, J.; Hershkovitz, V.; Tian, S.P.; Farrell, R. Increase in antioxidant gene transcripts, stress tolerance and biocontrol efficacy of Candida oleophila following sublethal oxidative stress exposure. FEMS Microbiol. Ecol. 2012, 80, 578–590. [Google Scholar] [CrossRef]
- Nie, F.; Zhang, X.; Qi, Q.; Yang, L.; Yang, Y.; Liu, W.; Lu, N.; Wu, Z.; You, Q.; Guo, Q. Reactive oxygen species accumulation contributes to gambogic acid-induced apoptosis in human hepatoma SMMC-7721 cells. Toxicology 2009, 260, 60–67. [Google Scholar] [CrossRef]
- Bignucolo, A.; Appanna, V.P.; Thomas, S.C.; Auger, C.; Han, S.W.; Omri, A.; Appanna, V.D. Hydrogen peroxide stress provokes a metabolic reprogramming in Pseudomonas fluorescens: Enhanced production of pyruvate. J. Biotechnol. 2013, 167, 309–315. [Google Scholar] [CrossRef]
- Jozefczuk, S.; Klie, S.; Catchpole, G.; Szymanski, J.; Cuadros-Inostroza, A.; Steinhauser, D.; Selbig, J.; Willmitzer, L. Metabolomic and transcriptomic stress response of Escherichia coli. Mol. Syst. Biol. 2010, 6, 364. [Google Scholar] [CrossRef]
- Balamurugan, S. Growth temperature associated protein expression and membrane fatty acid composition profiles of Salmonella enterica serovar Typhimurium. J. Basic. Microb. 2010, 50, 507–518. [Google Scholar] [CrossRef]
- Gray, M.J.; Jakob, U. Oxidative stress protection by polyphosphate—New roles for an old player. Curr. Opin. Microbiol. 2015, 24, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavida, P.; Jitlada, M.; Ornicha, P.; Mayumi, K.; Mamitaro, O. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [Green Version]
- Gallegos-Monterrosa, R.; Kankel, S.; Götze, S.; Barnett, R.; Stallforth, P.; Kovács, Á.T. Lysinibacillus fusiformis M5 induces increased complexity in Bacillus subtilis 168 colony biofilms via hypoxanthine. J. Bacteriol. 2017, 199, e00204-17. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Dong, Y.; Hong, X.; Pang, X.; Chen, D.; Chen, X. Directed evolution of dunaliella salina Ds-26-16 and salt-tolerant response in Escherichia coli. Int. J. Mol. Sci. 2016, 17, 1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijima, N.; Goyal, D.; Takada, A.; Wachi, M.; Nagai, K. Induction of only limited elongation instead of filamentation by inhibition of cell division in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 1998, 50, 227–232. [Google Scholar] [CrossRef]
- Mattick, K.L.; Jørgensen, F.; Legan, J.D.; Cole, M.B.; Porter, J.; Lappinscott, H.M.; Humphrey, T.J. Survival and filamentation of Salmonella enterica Serovar Enteritidis PT4 and Salmonella enterica Serovar Typhimurium DT104 at low water activity. Appl. Environ. Microbiol. 2000, 66, 1274–1279. [Google Scholar] [CrossRef] [Green Version]
- Ultee, E.; Ramijan, K.; Dame, R.T.; Briegel, A.; Claessen, D. Stress-induced adaptive morphogenesis in bacteria. Adv. Microb. Physiol. 2019, 74, 97–141. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Wang, Z.; Ren, M.; Huang, G.; Fang, B.; Bu, X.; Liu, Y.; Guan, S. Antibacterial effect of gallic acid against Aeromonas hydrophila and Aeromonas sobria through damaging membrane integrity. Curr. Pharm. Biotechnol. 2016, 17, 1153–1158. [Google Scholar] [CrossRef]
- Zhang, F.; Yao, X.; Sun, S.; Wang, L.; Wang, J. Effects of mesotrione on oxidative stress, subcellular structure, and membrane integrity in Chlorella vulgaris. Chemosphere 2019, 247, 125668. [Google Scholar] [CrossRef] [PubMed]
- Amanatidou, A.; Smid, E.J.; Bennik, M.H.J.; Gorris, L.G.M. Antioxidative properties of Lactobacillus sake upon exposure to elevated oxygen concentrations. FEMS Microbiol. Lett. 2001, 203, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Collins, E.B.; Aramaki, K. Production of hydrogen peroxide by Lactobacillus acidophilus. J. Dairy Sci. 1980, 63, 353–357. [Google Scholar] [CrossRef]
- Toscano, S.; Farieri, E.; Ferrante, A.; Romano, D. Physiological and biochemical responses in two Ornamental Shrubs to drought stress. Front. Plant Sci. 2016, 7, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urushihara, Y.; Kawasumi, K.; Endo, S.; Tanaka, K.; Hirakawa, Y.; Hayashi, G.; Sekine, T.; Kino, Y.; Kuwahara, Y.; Suzuki, M.; et al. Analysis of plasma protein concentrations and enzyme activities in cattle within the ex-evacuation zone of the fukushima daiichi nuclear plant accident. PLoS ONE 2016, 11, e0155069. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Xu, J.; Chen, Q.; Wang, H.; Kong, B. Physiological, Morphological and Antioxidant Responses of Pediococcus pentosaceus R1 and Lactobacillus fermentum R6 Isolated from Harbin Dry Sausages to Oxidative Stress. Foods 2021, 10, 1203. https://doi.org/10.3390/foods10061203
Zhang H, Xu J, Chen Q, Wang H, Kong B. Physiological, Morphological and Antioxidant Responses of Pediococcus pentosaceus R1 and Lactobacillus fermentum R6 Isolated from Harbin Dry Sausages to Oxidative Stress. Foods. 2021; 10(6):1203. https://doi.org/10.3390/foods10061203
Chicago/Turabian StyleZhang, Huan, Jianhang Xu, Qian Chen, Hui Wang, and Baohua Kong. 2021. "Physiological, Morphological and Antioxidant Responses of Pediococcus pentosaceus R1 and Lactobacillus fermentum R6 Isolated from Harbin Dry Sausages to Oxidative Stress" Foods 10, no. 6: 1203. https://doi.org/10.3390/foods10061203