3.3. Taxonomy
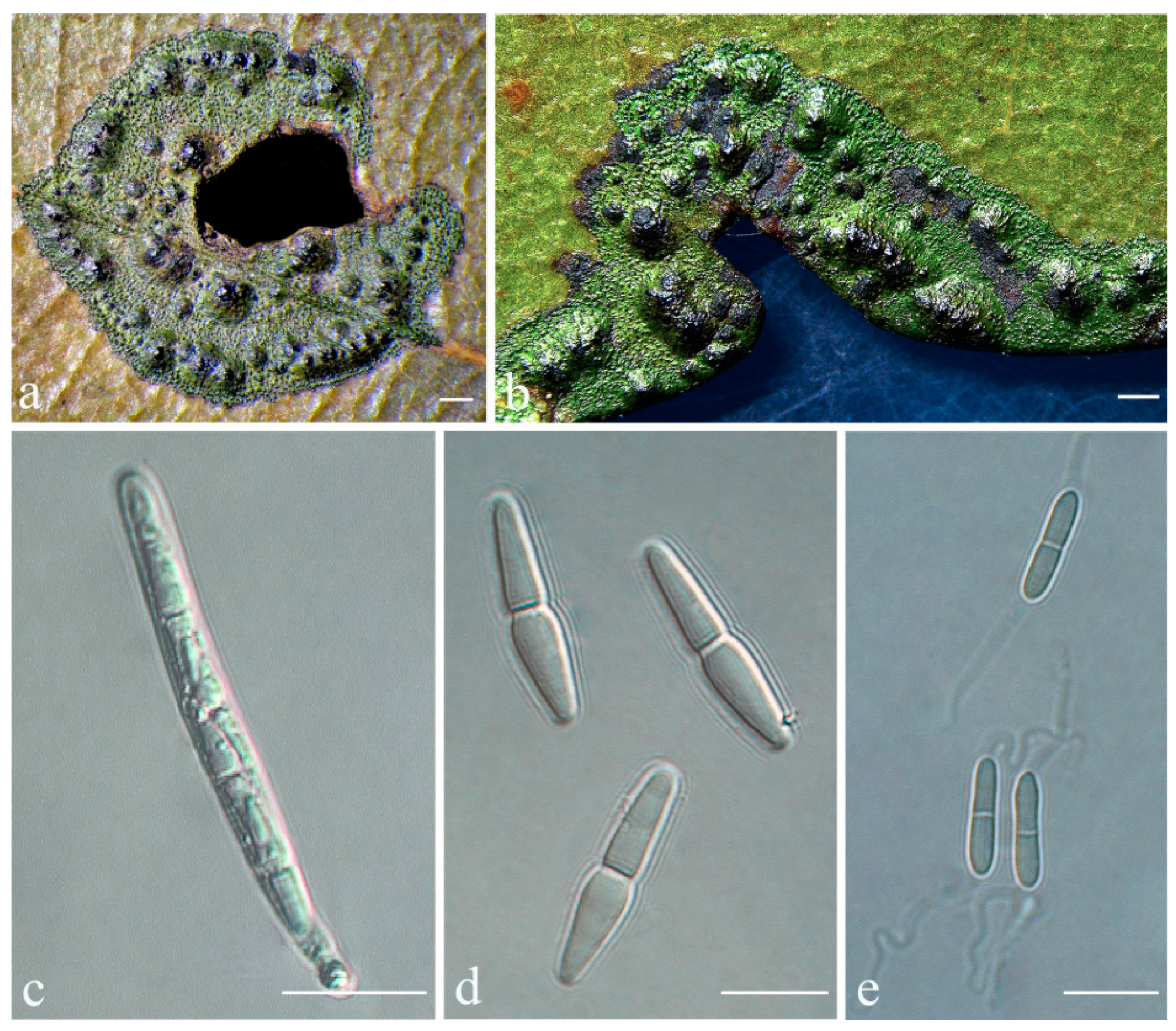
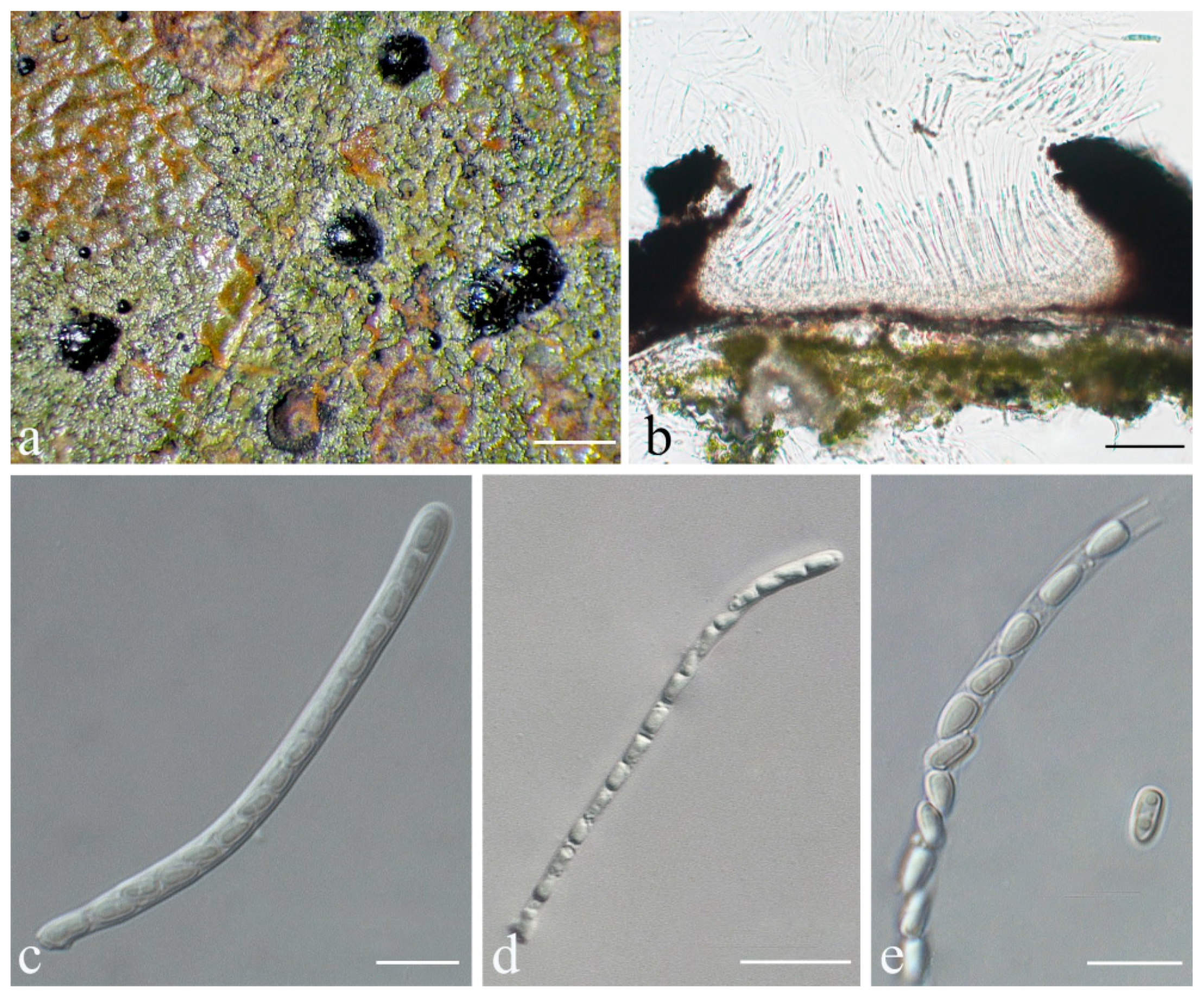
Phylloporis palmae (Cavalc. & A.A. Silva) S.H. Jiang, J.C. Wei, Xavier-Leite & Lücking,
comb. nov. (
Figure 4).
Basionym: Manaustrum palmae Cavalc. & A.A. Silva, in Cavalcante et al., Publicações. Instituto de Micologia da Universidade de Pernambuco 647: 14. 1972.
MycoBank MB 838157.
Typus: BRAZIL, Amazonas, Manaus, Rodovia Manaus-Itacoatiara, 67 km; May 1961, Garnier s.n. (URM 23296/Exs. 15761, holotype!; INPA, isotype, not seen). Amapá, Floresta Nacional do Amapá, Municipio de Porto Grande, by bus 112 km from Macapá (145 NNW from Macapá), and then by boat 57 km from Porto Grande (70 km WNW from Porto Grande), S 01°02′32″, W 51°56′32″, Amazon rainforest, on leaves, August 2015, A.B. Xavier-Leite 2670 (ISE–33670, epitype, here designated; ITS Genbank No. MW344138; MBT 395040).
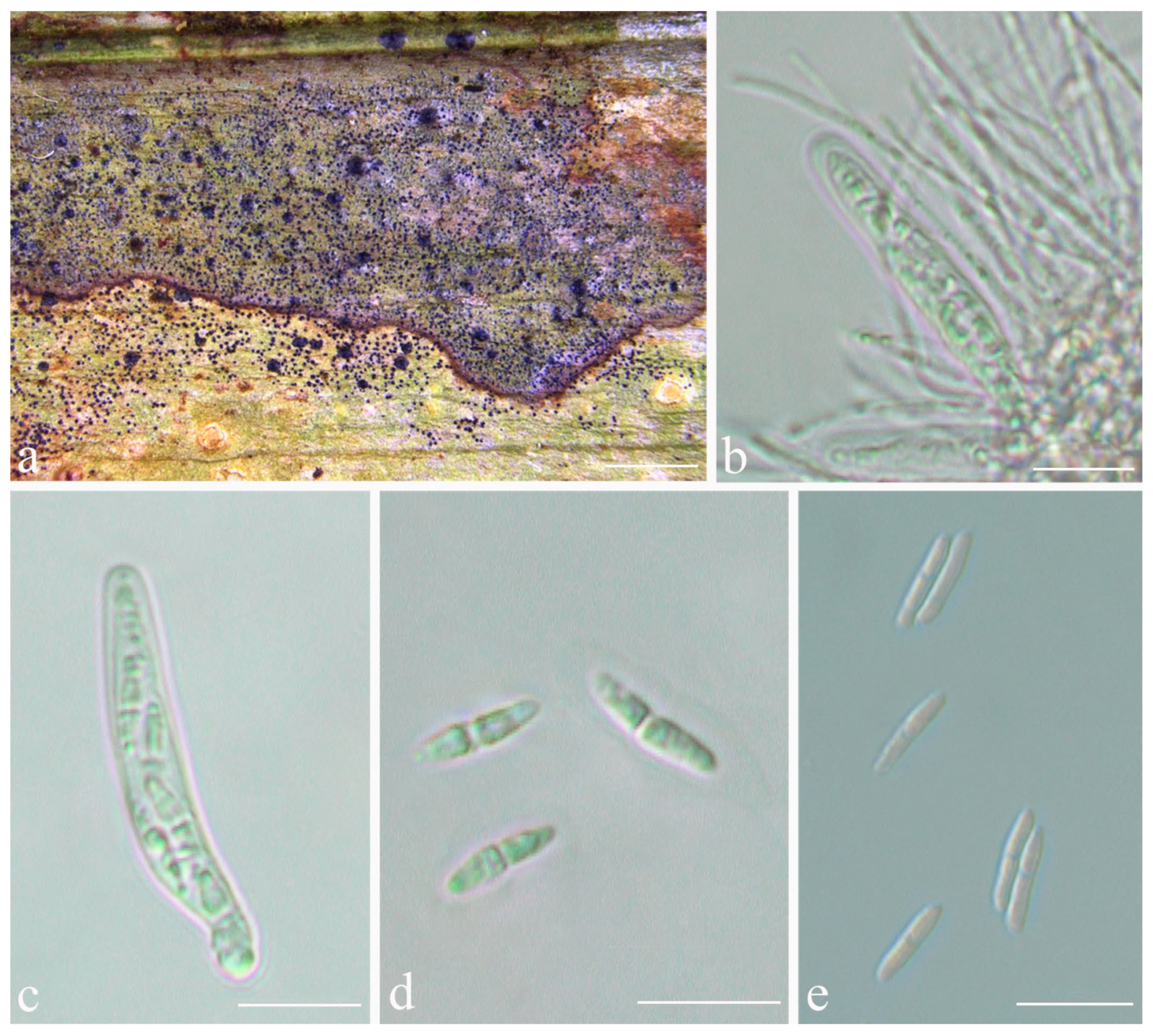
Description: Thallus supracuticular, continuous or marginally dispersed, 1–10 mm across and 10–15 μm thick, with entire margins, olive-green with metallic glance, densely furnished with minute, blackish verrucae. Photobiont Phycopeltis sp., cells rectangular, forming net-like plates with interspaces, 5–10 × 4–6 μm. Perithecia erumpent, hemispherical to wart-shaped, 0.2–0.3 mm diameter and 50–70 μm high, black or greyish black due to a thin thallus cover lacking algae. Excipulum prosoplectenchymatous, 7.5–10 μm thick, colourless to brownish. Involucrellum carbonaceous, 15–35 μm thick. Paraphyses unbranched, c. 1–1.5 μm thick. Asci cylindrical to narrowly obclavate, 28–45 × 5–7.5 μm. Ascospores eight per ascus, biseriate, fusiform-ellipsoid, one-septate, with slight constriction at septum, 8.5–10 × 2–3 μm, 3.5–4.5 times as long as broad. Pycnidia producing macroconidia present in the type material but not observed in the sequenced specimens, greyish black, wart-shaped, 0.1–0.15 mm in diameter. Macroconidia one-septate, 8–12 × 1.5–2 μm. Microconidia not seen.
Chemistry: No substances detected by TLC.
Additional specimens examined: BRAZIL, Amapá, Floresta Nacional do Amapá, Municipio de Porto Grande, by bus 112 km from Macapá, and then by boat 57 km from Porto Grande, S 01°02′32″, W 51°56′32″, Amazon rainforest, on leaves, August 2015, A.B. Xavier-Leite 2374 (ISE–33374), 2667 (ISE–33667).
Notes: The sequenced material corresponds morphologically to
Phylloporis multipunctata (G. Merr. ex R. Sant.) Vězda, described from Indonesia and recently synonymised under
P. cinefaciens (Nyl.) S.H. Jiang, Lücking & Sérus. [
41]. However, microscopic examination revealed that the foliicolous specimens from Brazil have much smaller ascospores than the type material of
P. cinefaciens and
P. multipunctata (12–16 × 3.5–4.5 µm in the latter two [
41]). We, therefore, take up the epithet introduced by Cavalcante et al. [
70], originally synonymised with
P. multipunctata by Lücking (1998), for this material [
71]. Since the holotype only bears pycnidia, we designate one of the sequenced specimens with numerous perithecia as epitype.
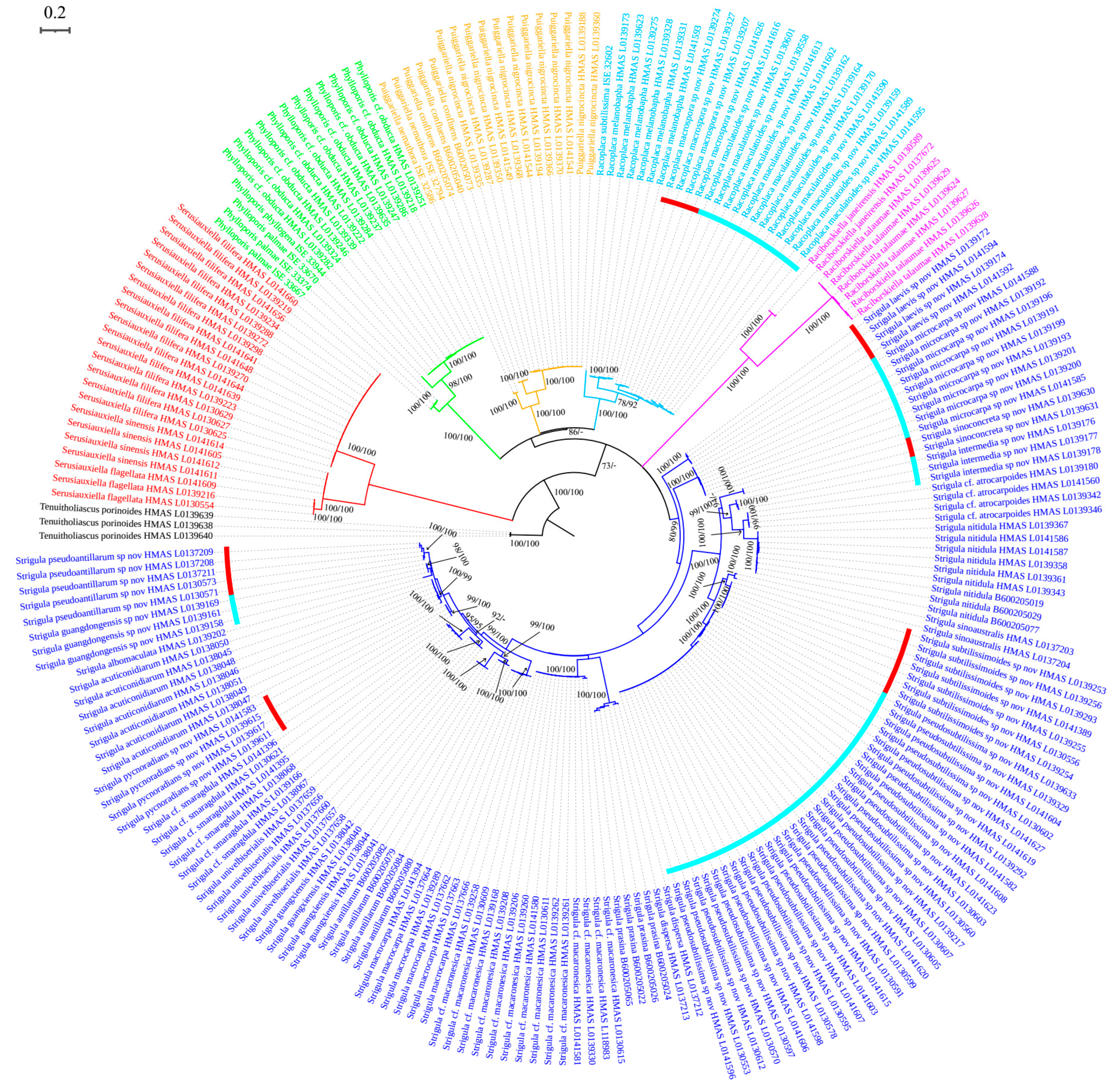
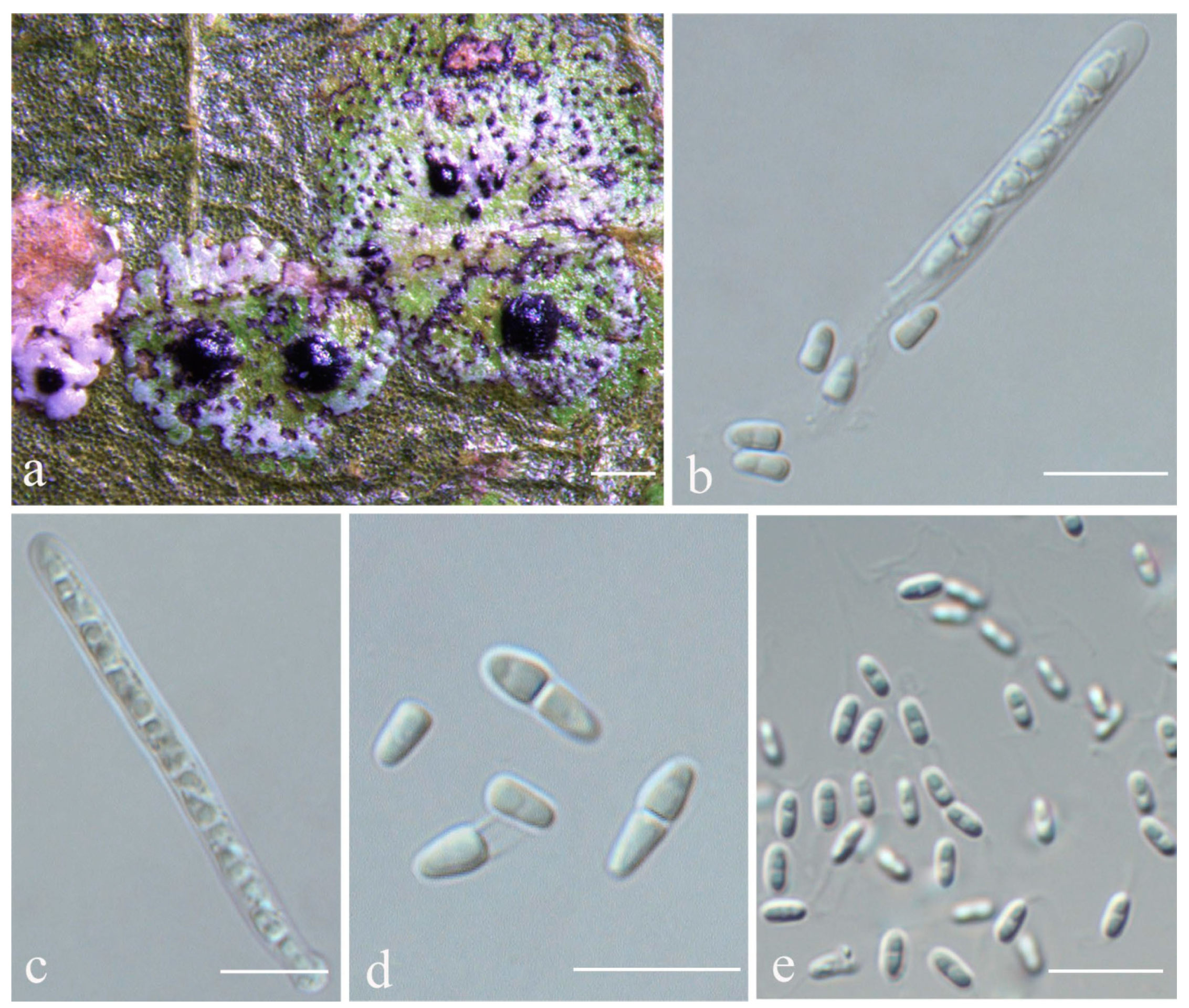
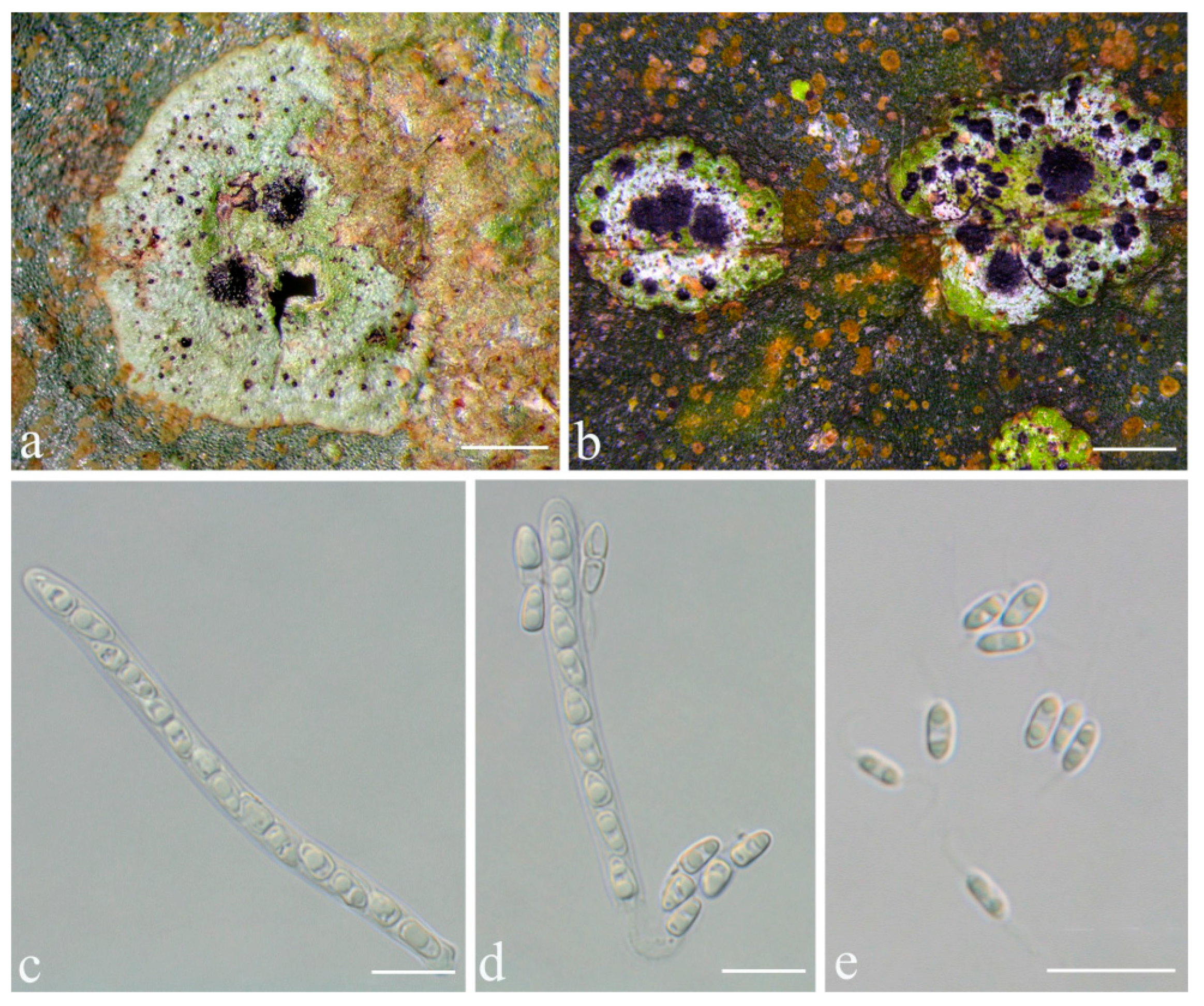
Racoplaca macrospora S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 5).
MycoBank MB 838159.
Etymology: The epithet macrospora refers to the large ascospores, thus far the largest known in the genus, strongly contrasting with the typically small thalli.
Typus: CHINA, Guangxi, Shangsi County, Shiwan Mountain National Nature Reserve, N 21°53′50″, E 107°54′18″, alt. 411 m, on living leaves, 25 May 2015, X.L. Wei & J.H. Wang GX20150306 (HMAS–L0139274, holotype; ITS Genbank No. MW344166).
Diagnosis: Differing from Racoplaca maculata (Cooke & Massee) S.H. Jiang, Lücking & J.C. Wei in the much larger, fusiform ascospores and the much larger, one-septate macroconidia.
Description: Thallus subcuticular, continuous, growing along the margins and scars of leaves, lobes small, not very long, forming irregular lines, margin crenulate-laciniate, bordered by a thin, black line, dark (olive-)green, nitidous, 5–10 mm across and 10–20 μm thick. Photobiont a species of Cephaleuros, cells angular-rounded, 8–13 × 4–7 μm. Perithecia covered by thin thallus layer up to ostiole, wart-shaped, 0.2–0.45 mm diameter and 100–200 μm high, greyish to greenish black. Excipulum prosoplectenchymatous, 10–15 μm thick, colourless to brown. Involucrellum carbonaceous, 15–30 μm thick, black. Paraphyses unbranched, thin, about 1 μm. Asci narrowly obclavate, 75–100 × 7.5–10 μm, eight-spored. Ascospores biseriate or irregularly orientated, fusiform, one-septate, with distinct constriction at septum, 22.5–27.5 × 4–5 μm. Pycnidia exposed, wart-shaped, black and shiny, those producing macroconidia 0.1–0.2 mm, those producing microconidia 0.07–0.15 mm diameter. Macroconidia bacillar, one-septate, 10–15 × 1.8–2.5 μm, with 17–35 μm long appendage at both ends. Pycnidia producing microconidia appearing as tiny black points, or included in pycnidia producing macroconidia. Microconidia ellipsoid, 3–5 × 1 μm.
Chemistry: No substances detected by TLC.
Habitat and distribution: The new species was found in humid, semi-exposed forest habitats in southern China.
Additional specimens examined: CHINA, Guangxi, Shangsi County, Shiwan Mountain National Nature Reserve, N 21°53′50″, E 107°54′18″, alt. 411 m, on living leaves, 25 May 2015, X.L. Wei & J.H. Wang GX20150316 (HMAS–L0139327); Hainan, Ledong County, Jianfengling, Mingfenggu, N 18°44′5′′, E 108°52′8′′, alt. 920 m, on living leaves, 11 December 2014, J.H. Wang & R.D. Liu HN2014144_3 (HMAS–L0141626); 28 July 2009, J.C. Wei WJC065 (HMAS–L0139206), WJC069 (HMAS–L0139207).
Notes: In terms of thallus morphology,
Racoplaca macrospora resembles
R. maculata in the relatively compact, more greenish thallus lacking distinct laciniae. The latter, however, has much smaller (12–18 × 2.5–3.5 μm), oblong ascospores and much smaller, non-septate macroconidia [
45]. The ascospores and macroconidia point to a closer relationship with
R. melanobapha (Kremp.) S.H. Jiang, Lücking & J.C. Wei; however, that species has still smaller ascospores (14–22 × 3–5 μm), and also differs in the strongly and regularly laciniate, more brownish thallus [
45]. This closer relationship is supported by the molecular data (
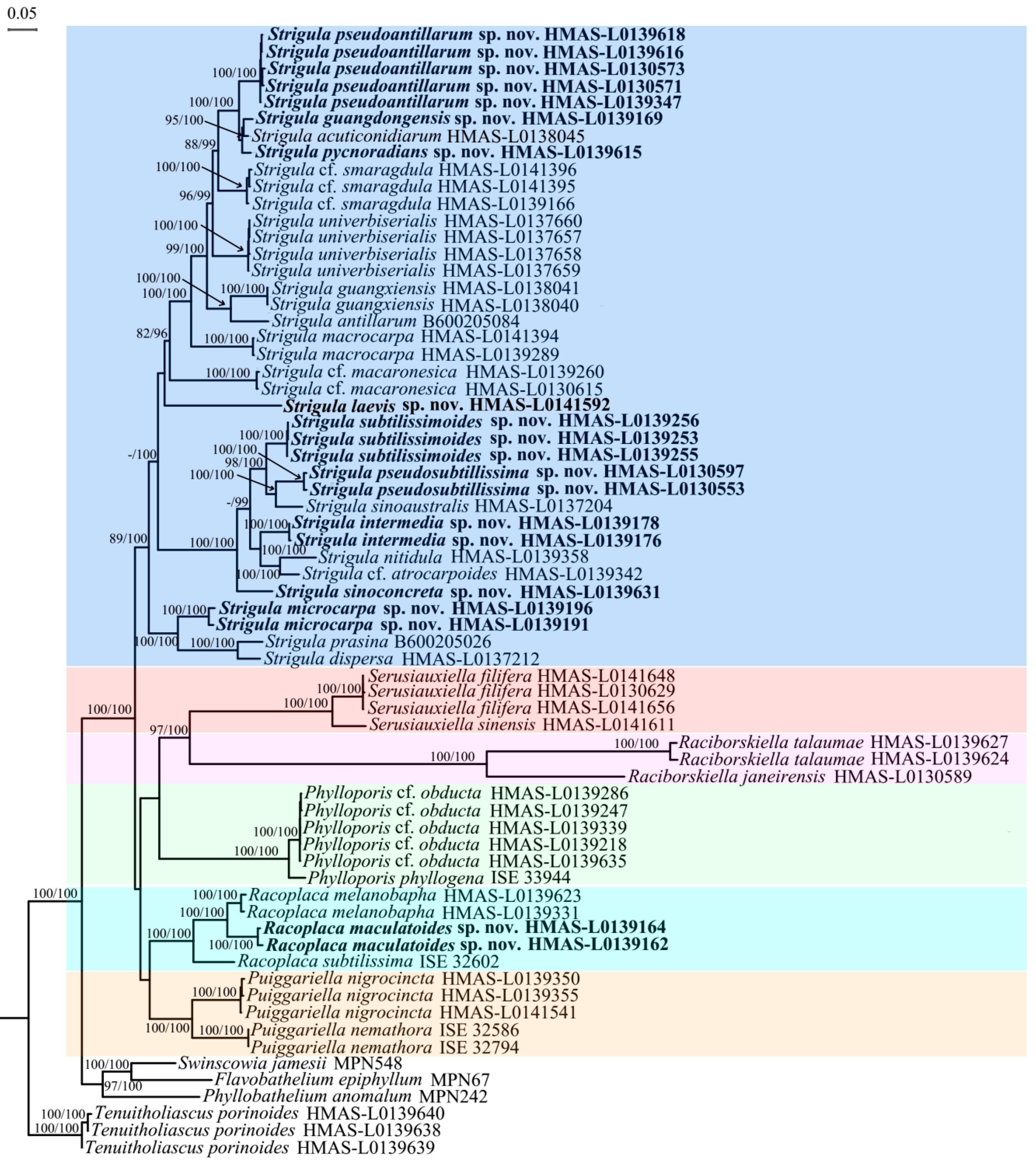
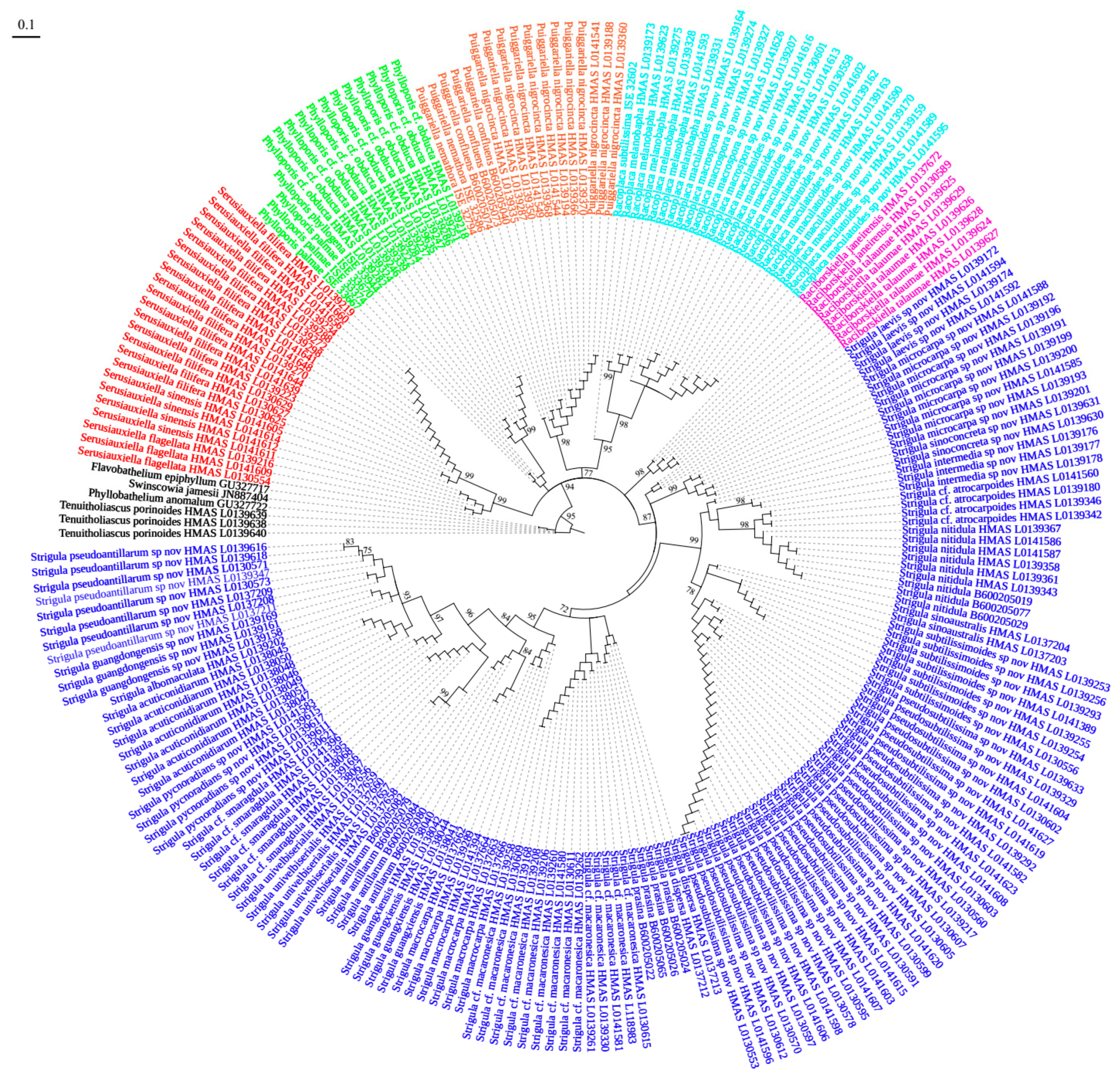
Figure 1).
Based on the ITS data,
Racoplaca macrospora was found nested within another newly recognised species,
R. maculatoides (see below). However, there are between 18 consistent base call differences between the two species (substitutions and indels), corresponding to a mean identity value of 96% (
Table S2), substantially below what could be accepted for within-species variation. Given that
R. maculatoides has distinctly shorter (15–25 μm vs. 22.5–27.5 μm long) ascospores and macroconidia with shorter appendages (10–18 μm vs. 17–35 μm), we consider
R. macrospora a recently emerging, yet distinct species. Species evolving from paraphyletic residuals are now broadly accepted [
36], but an artifactually paraphyletic topology could also result from the most closely related species not having been sequenced yet. Another possible explanation is that
R. maculatoides itself represents a species complex. Indeed, separate analysis of the ITS only for
Racoplaca revealed three subclades in this clade: one supported subclade (75%), including the type of
R. maculatoides, with a basally emerging individual with 63% support; another shallow subclade with four individuals currently assigned to
R. maculatoides; and a strongly supported subclade (100%) on a long branch, representing
R. macrospora (
Figure S1).
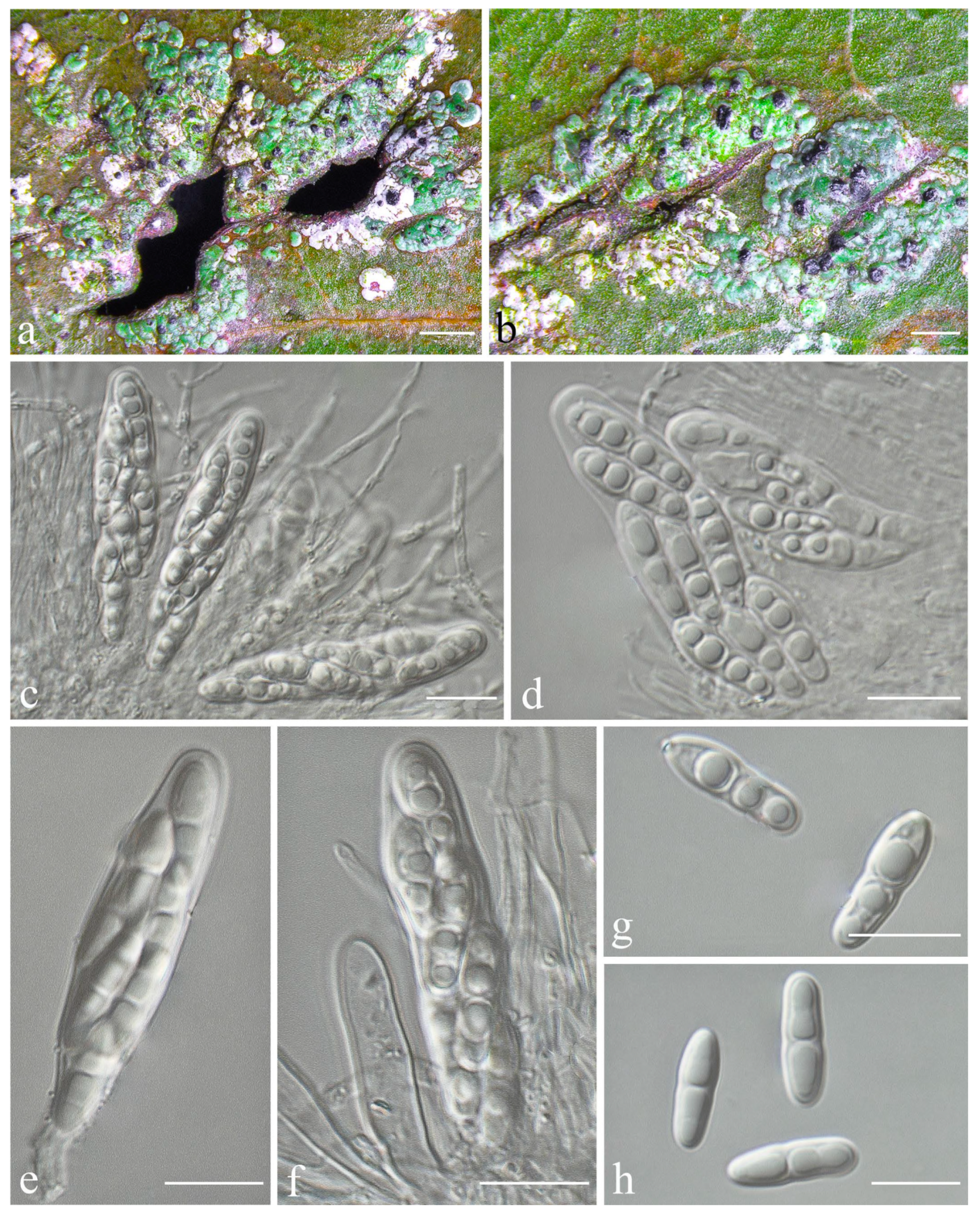
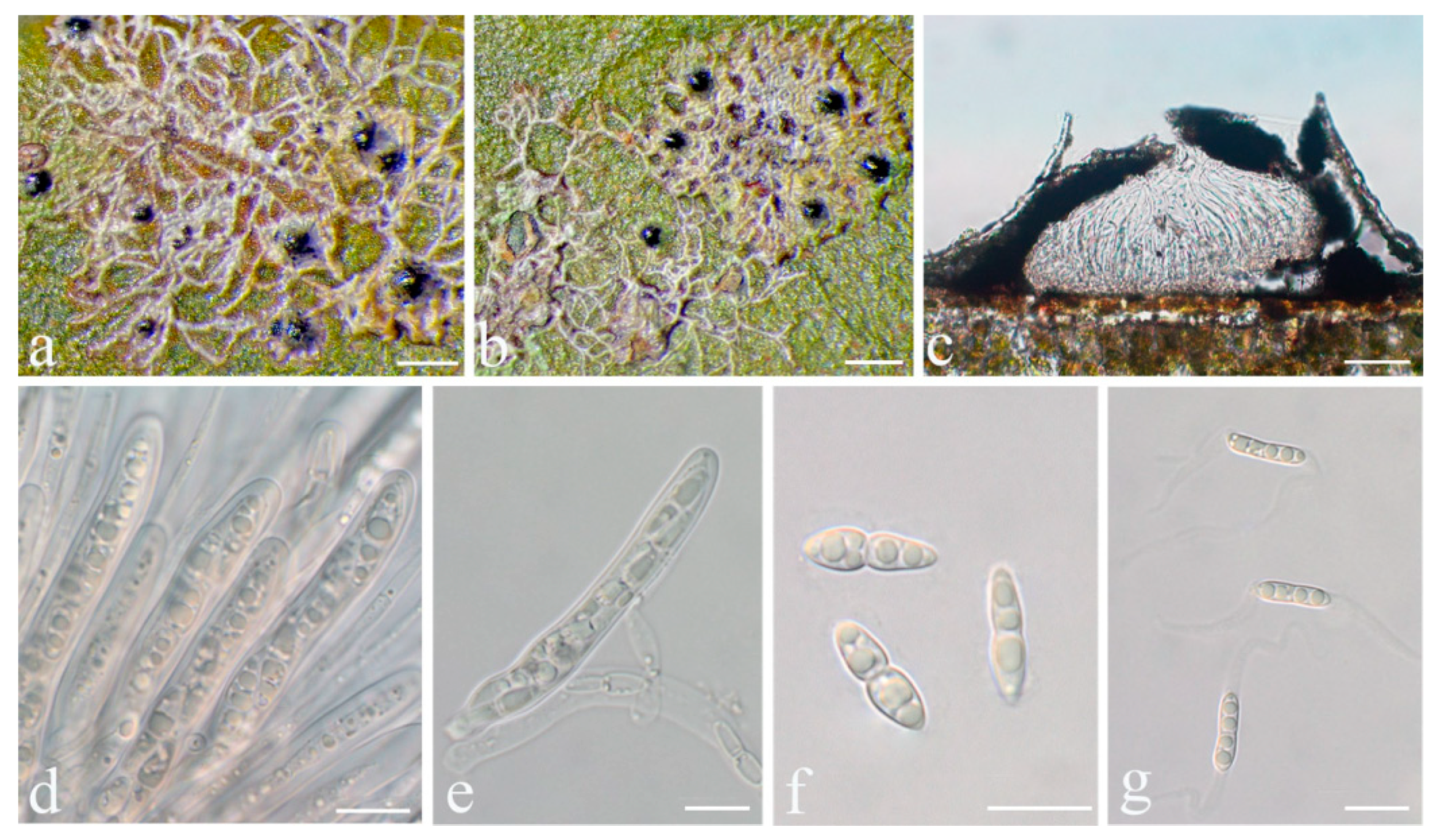
Racoplaca maculatoides S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 6).
MycoBank MB 838161.
Etymology: The epithet maculatoides refers to the morphological similarity with Racoplaca maculata.
Typus: CHINA, Guangdong, Shixing County, Chebaling National Nature Reserve, N 24°43′27″, E 114°15′22″, alt. 345 m, on living leaves, 14 May 2015, X.L. Wei & J.H. Wang GD2015020 (HMAS–L0139170, holotype; ITS Genbank No. MW344174).
Diagnosis: The new species externally resembles Racoplaca maculata but differs by the longer asci, larger, fusiform ascospores, and larger, one-septate macroconidia.
Description: Thallus subcuticular, continuous, sometimes occurring along the nerves, margins, and scars of leaves, lobes confluent, indicated by irregular, dark lines and dots on the thallus surface, margin entire or slightly crenulate, bordered by thin, black line, dark green, nitidous, with a metallic glance, 5–15 mm across and 10–20 μm thick. Photobiont a species of Cephaleuros, cells angular-rounded, 8–13 × 4–7 μm. Perithecia covered by thin thallus layer up to ostiole, wart-shaped, 0.25–0.5 mm diameter and 100–200 μm high, greyish to greenish black. Excipulum prosoplectenchymatous, 10–15 μm thick, colourless to brown. Involucrellum carbonaceous, 15–30 μm thick, black. Paraphyses unbranched or slightly branched, thin. Asci narrowly obclavate, 50–90 × 7.5–10 μm, eight-spored. Ascospores biseriate or irregularly orientated, fusiform, one-septate, with slight constriction at the septum, 15–25 × 3–5 μm. Pycnidia exposed, wart-shaped, black and shiny, those producing macroconidia 0.1–0.2 mm diameter, those producing microconidia 0.07–0.15 mm diameter. Macroconidia bacillar, one-septate, 10–12.5 × 2.5 μm, with 10–18 μm long appendage at both ends that may be terminated by a hook. Pycnidia producing microconidia appearing as tiny black points, or included in pycnidia producing macroconidia. Microconidia ellipsoid, 3–5 × 1 μm.
Chemistry: No substances detected by TLC.
Habitat and distribution: The new species was found in humid, semi-exposed forest habitats in southern China.
Additional specimens examined: CHINA, Guangdong, Shixing County, Chebaling National Nature Reserve, N 24°43′27″, E 114°15′22″, alt. 345 m, on living leaves, 14 May 2015, X.L. Wei & J.H. Wang GD2015025_2 (HMAS–L0139159), GD2015025_8 (HMAS–L0139162), GD2015025_9 (HMAS–L0139163), GD2015025_10 (HMAS–L0139164); N 24°42′35″, E 114°13′37″, alt. 398 m, on living leaves, 14 May 2015, X.L. Wei & J.H. Wang GD2015031_7_1 (HMAS–L0141589); N 24°42′39″, E 114°13′30″, alt. 387 m, on living leaves, 15 May 2015, X.L. Wei & J.H. Wang GD2015038_2_1 (HMAS–L0141590); Hainan, Ledong County, Jianfengling, Mingfenggu, N 18°44′5″, E 108°52′8″, alt. 920 m, on living leaves, 11 December 2014, J.H. Wang & R.D. Liu HN2014123 (HMAS–L0130586), HN2014123_5 (HMAS–L0141595), HN2014144 (HMAS–L0130598), HN2014177 (HMAS–L0130558), HN2014134 (HMAS–L0130591), HN2014148 (HMAS–L0130601), HN2014136 (HMAS–L0130592); N 18°44′36″, E 108°50′39″, alt. 962 m, on living leaves, 12 December 2014, J.H. Wang & R.D. Liu HN2014221 (HMAS–L0130606), HN2014221_4 (HMAS–L0141602); N 18°44′32″, E 108°50′32″, alt. 985 m, on living leaves, 12 December 2014, J.H. Wang & R.D. Liu HN2014286_2 (HMAS–L0141616), HN2014281_1 (HMAS–L0141613); Qiongzhong County, Limu mountain, N 19°10′29″, E 109°44′53″, alt. 800 m, on living leaves, 10 September 2017, S.H. Jiang HN20170656 (HMAS–L0139637).
Notes:
Racoplaca maculatoides strongly resembles
R. maculata in morphology but can be distinguished by the longer asci; larger, fusiform ascospores; and larger, one-septate macroconidia (asci 40–60 × 5–8 μm, ascospores 12–18 × 2.5–3.5 μm, macroconidia aseptate, 4–6 × 1.5–2 μm in
R. maculata [
45]).
Racoplaca maculatoides is most similar and most closely related to
R. macrospora (
Figure 1). It differs by the shorter ascospores and the narrower macroconidia with shorter appendages.
Racoplaca melanobapha resembles
R. maculatoides in ascospores and macroconidia, but differs in the strongly and regularly laciniate, more brownish thallus [
45]. Both are also phylogenetically distinct (
Figure 1 and
Figure 2).
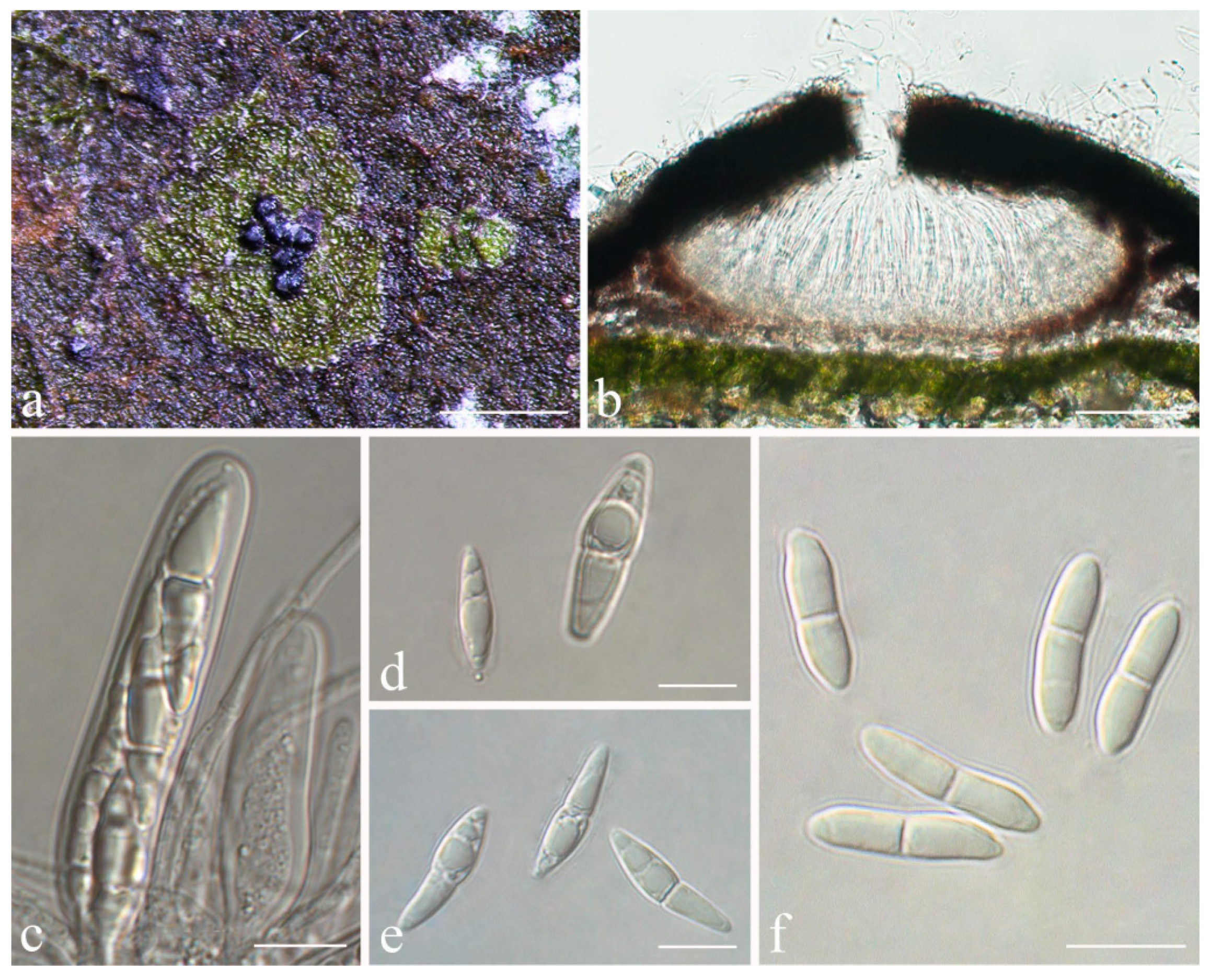
Racoplaca melanobapha (Kremp.) S.H. Jiang, Lücking & J.C. Wei, comb. nov.
MycoBank MB 842243.
Basionym: Verrucaria melanobapha Kremp., Lichenes Foliicolae Quos legit O. Beccari Annis 1866–1867 in Insula Borneo: 18 (1874); Verrucaria melanobapha Kremp., Nuovo Giornale Botanico Italiano 7: 51 (1875) (nom. inval., ICN Art. 6.3 Note 2); Strigula melanobapha (Kremp.) R. Sant., Symb. Bot. Upsal. 12(1): 188 (1952).
Typus: Malaysia (Borneo), Beccari 219 (M, holotype!).
Notes: This combination was first proposed by Jiang et al. [
47] but was unfortunately invalid as the basionym was not cited. It is, therefore, validated here.
Strigula guangdongensis S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 7).
MycoBank MB 838162.
Etymology: The specific epithet is derived from the name of the type locality of the species, Guangdong province.
Typus: CHINA, Guangdong, Shixing County, Chebaling National Nature Reserve, N 24°43′27″, E 114°15′22″, alt. 345 m, on living leaves, 14 May 2015, X.L. Wei & J.H. Wang GD2015019 (HMAS–L0139169, holotype; ITS Genbank No. MW344202).
Diagnosis: The new species can be distinguished by the beaked, more elongate pycnidia than in other species of the genus. The phylogenetic trees also indicate it forms an independent clade.
Description: Thallus subcuticular, 0.5–4 mm across and 15–25 μm thick, continuous or dispersed into rounded to irregular, partly confluent patches, with entire to crenulate or lobulate margins, bright green mottled with white. Photobiont a species of Cephaleuros, cells angular-rounded, 5–15 × 4–10 μm. Perithecia few, 0.1–0.3 mm diameter and 120–180 μm high, postmature; no asci or ascospores observed. Pycnidia numerous, conical to shortly beaked, 40–60 μm diameter, black, the beak paler and somewhat translucent. Macroconidia bacillar, one-septate, sometimes breaking into halves, 12.5–20 × 3–4 μm. Microconidia not seen.
Chemistry: No substances detected by TLC.
Habitat and distribution: It was found on the surface of living leaves in humid, semi-exposed forests of south China.
Additional specimens examined: CHINA, Guangdong, Shixing County, Chebaling National Nature Reserve, N 24°43′27″, E 114°15′22″, alt. 345 m, on living leaves, 14 May 2015, X.L. Wei & J.H. Wang GD2015025_7 (HMAS–L0139161), GD2015025_1 (HMAS–L0139158).
Notes: This is another newly recognised species that belongs in the morphologically defined
Strigula smaragdula complex. The latter has been described from Nepal and we have currently marked one phylogenetically distinct clade from China as a candidate for this species (
S. cf.
smaragdula;
Figure 1 and
Figure 2).
Strigula guangdongensis differs from this clade phylogenetically and is also set apart by its beaked pycnidia. Lücking also mentioned specimens of
S. smaragdula s.lat. with shortly beaked pycnidia from the Neotropics, but these likely represent another unrecognised taxon [
45]. A corticolous species from Madagascar,
S. rostrata R.C. Harris & Aptroot, has more strongly beaked pycnidia and submuriform macroconidia [
43]; that species is now placed in a different genus, as
Swinscowia rostrata (R.C. Harris & Aptroot) S.H. Jiang, Lücking & Sérus. [
41].
Strigula intermedia S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 8).
MycoBank MB 838163.
Etymology: The epithet refers to the thallus being intermediate between that of Strigula concreta and S. nitidula.
Typus: CHINA, Yunnan, Pu’er city, Simao District S214, alt. 829 m, on living leaves, 23 October 2016, X.Y. Liu YN20160161 (HMAS–L0139177, holotype; ITS Genbank No. MK206305).
Diagnosis: The new species is intermediate between Strigula concreta (Fée) R. Sant. s.str. and S. nitidula Mont. s.str. in having a slightly thickened thallus, with irregular, short black lines.
Description: Thallus subcuticular, 1.5–4.5 mm across and 7.5–22.5 μm thick, with entire to crenulate margins, bright green mottled with white, surface smooth, sometimes with black dots or lines. Photobiont a species of Cephaleuros, cells angular-rounded, 8–12 × 4–6 μm. Perithecia exposed, conical to wart-shaped, 0.25–0.5 mm in diameter and 100–190 μm high, black. Excipulum prosoplectenchymatous, 12.5–25 μm thick, brown. Involucrellum 25–62.5 μm thick, carbonaceous, black. Paraphyses unbranched. Asci cylindrical, 40–60 × 3–4 μm. Ascospores eight per ascus, uniseriate, ellipsoid, one-septate, sometimes with one to two oil droplets per cell when fresh, with distinct constriction at the septum and sometimes broken into parts outside the asci, 7.5–10 × 2–2.5 μm. Pycnidia wart-shaped, those producing macroconidia 0.1–0.15 mm, those producing microconidia 0.05–0.1 mm diameter, black. Macroconidia bacillar, zero- to one-septate, 4–6 × 1.5–2.5 μm, with appendage at one end or both ends c. 5–10 μm long. Microconidia fusoid, aseptate, 4–5 × 1.5–2 μm.
Chemistry: No substances detected by TLC.
Habitat and distribution: The new species grows on living leaves in wet tropical forest in China.
Additional specimens examined: CHINA, Yunnan, Pu’er city, Simao District S214, alt. 829 m, on living leaves, 23 October 2016, X.Y. Liu YN20160160 (HMAS–L0139176), YN20160162 (HMAS–L0139178).
Notes:
Strigula intermedia conforms to the morphology of
S. concreta, in particular the rather thin thallus with crenulate margins, the exposed, black perithecia, and the uniseriate asci with short ascospores sometimes breaking into halves [
45]. There is at least one other new species belonging to this morphodeme,
S. sinoconcreta (see below). Given that both are phylogenetically distinct in both the ITS and the four-marker phylogeny (
Figure 1 and
Figure 2), we have to assume that
S. concreta s.lat. forms a species complex, similar to
S. smaragdula and intermingled with lineages representing the
S. nitidula morphodeme (see below). The latter agrees with
S. concreta anatomically, including the uniseriate asci with short ascospores often breaking into halves, but has a very thin thallus often bordered by a black line, as in
Racoplaca [
42,
45]. The two thallus morphologies are sometimes difficult to distinguish, a feature attributed to leaf characteristics [
45], but it appears that numerous lineages are involved.
Strigula intermedia represents an intermediate thallus type, thinner than in typical
S. concreta but thicker than in typical
S. nitidula, and with short, irregular black lines not as distinct as in
S. nitidula.
Strigula concreta has been described from the Caribbean, with one current synonym from Brazil (
S. rugulosa Müll. Arg.), one from Africa (
S. atrocarpa Vain.), and two from the Philippines (
S. sulcata Vain.,
Porina crenulata Vain. [
42,
45]). These names are, therefore, potentially available for cryptic or near-cryptic lineages in the corresponding regions. However, considering the striking diversification of the
S. smaragdula complex in Asia alone, with no apparent overlap in species distributions between (sub)tropical China and Korea or Thailand ([
48,
49,
50], this paper), it seems unlikely that the Philippine material of
Strigula sulcata or
Porina crenulata is conspecific with any of the two Chinese lineages detected here, and, therefore, we introduce new species for the latter.
Strigula laevis S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 9).
MycoBank MB 838164.
Etymology: Epithet of the new species “laevis” is a Latin word, and means smooth and bright thallus.
Typus: CHINA, Guangdong, Ruyuan county, Nanling National Forest Park, N 24°54′59″, E 113°2′5″, alt. 848 m, on living leaves, 17 May 2015, X.L. Wei & J.H. Wang GD2015066_2 (HMAS–L0141592, holotype; ITS Genbank No. MW344203).
Diagnosis: The species is morphologically similar to Strigula nigrocarpa Lücking, but can be distinguished by the small, thin thallus, not sharply delimited perithecia, and biseriate, smaller ascospores.
Description: Thallus subcuticular, continuous, 2–5 mm across and 10–20 μm thick, with entire margins, bright to dark green, smooth. Photobiont a species of Cephaleuros, cells angular-rounded, 6–15 × 6–10 μm. Perithecia covered by a thin thallus layer, applanately hemispherical, 0.3–0.5 mm diameter and 100–180 μm high, dark green but uppermost part exposed and black. Excipulum prosoplectenchymatous, 5–10 μm thick, colourless to brown. Involucrellum carbonaceous, 12.5–32.5 μm thick, black. Paraphyses unbranched. Asci obclavate, 52–80 × 6–10 μm, eight-spored. Ascospores biseriate, oblong-ellipsoid, one-septate, with slight constriction at septum, 16–20 × 3–5 μm. Pycnidia not seen.
Chemistry: No substances detected by TLC.
Habitat and distribution: This species was found on living leaves in tropical to subtropical rainforests of southern China.
Additional specimens examined: CHINA, Guangdong, Ruyuan county, Nanling National Forest Park, N 24°54′59″, E 113°2′5″, alt. 848 m, on living leaves, 17 May 2015, X.L. Wei & J.H. Wang GD2015066 (HMAS–L0139172), GD2015066_4 (HMAS–L0141594). Hunan, Mangshan National Forest Park, N 24°56′59″, E 112°56′2″, alt. 1257 m, on living leaves, 17 May 2015, X.L. Wei & J.H. Wang GD2015077 (HMAS–L0139174).
Notes:
Strigula laevis belongs in the
S. smaragdula complex and is somewhat similar to the neotropical
S. nigrocarpa, due to the blackish perithecia. However, in
S. nigrocarpa, the perithecia are sharply delimited from the thallus, and the ascospores are uniseriate in longer, cylindrical asci, and the ascospores are somewhat larger [
45]. It is also similar to the neotropical
S. minuta Lücking, but that species has much smaller ascospores (9–12 × 2–3 μm [
45]).
Strigula microcarpa S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 10).
MycoBank MB 838165.
Etymology: Epithet of the new species “microcarpa” is a Latin word, and means small perithecia.
Typus: CHINA, Yunnan, Xishuangbanna, Jinghong city, N 22°19′15″, E 100°47′13″, alt. 968 m, on living leaves, 28 October 2016, X.Y. Liu YN20160099 (HMAS–L0139196, holotype; ITS Genbank No. MW344208).
Diagnosis: The new species was similar with Strigula microspora Lücking in the thallus, but can be distinguished by the shorter ascus and the biseriate ascospores.
Description: Thallus subcuticular, continuous, 3–7 mm across and 20–30 μm thick, typically with crenulate or shortly lobulate margins, pale greyish green to whitish. Photobiont a species of Cephaleuros, cells angular-rounded, 5–17 × 4–11 μm. Perithecia immersed-erumpent, hemispherical, 0.2–0.3 mm diameter and 100–180 μm high, exposed part black. Excipulum prosoplectenchymatous, 5–10 μm thick, blackish brown. Involucrellum confluent with excipulum, carbonaceous, 20–40 μm thick, black. Paraphyses unbranched or slightly branched, thin. Asci cylindrical, 35–55 × 8.5–10 μm. Ascospores eight per ascus, biseriate to irregularly arranged, fusiform to ellipsoid, one-septate, with slight constriction at septum, 12.5–15 × 3.5–4.5 μm, 3–5 times as long as broad. Pycnidia not seen.
Chemistry: No substances detected by TLC.
Habitat and distribution: Collected on living leaves in humid, semi-exposed forests of southern China.
Additional specimens examined: CHINA, Yunnan, Mengla county, N 21°42′10″, E 101°37′28″, alt. 669 m, on living leaves, 24 October 2016, X.Y. Liu YN20160154_2 (HMAS–L0141585); Pu’er city, Hani and Yi Autonomous County of Jiangcheng, N 22°38′56″, E 101°24′27″, alt. 829 m, on living leaves, 23 October 2016, X.Y. Liu YN20160161_1 (HMAS–L0141588); Pu’er city, Simao district S214, alt. 1316 m, on living leaves, 23 October 2016, X.Y. Liu YN20160044 (HMAS–L0139175); Xishuangbanna, Jinghong city, N 22°19′15″, E 100°47′13″, alt. 968 m, on living leaves, 28 October 2016, X.Y. Liu YN20160097 (HMAS–L0139191), YN20160098 (HMAS–L0139192), YN20160102 (HMAS–L0139199), YN20160103 (HMAS–L0139193), YN20160105 (HMAS–L0139200), YN20160106 (HMAS–L0139201).
Notes:
Strigula microcarpa bears some resemblance to the neotropical species
S. microspora, but the latter has longer asci (50–70 × 5–6 μm) and uniseriate ascospores [
45].
Strigula wandae M. Cáceres & Lücking from the Valdivian Forest in Chile also resembles
S. microcarpa in morphology, but has larger ascospores (15–23 × 4–5 μm [
45]). While the ITS resolved the new species as an early diverging lineage within
Strigula s.str. (
Figure 1), the four-marker tree revealed a strongly supported relationship with the
S. prasina Müll. Arg. complex (
Figure 2).
Strigula pseudoantillarum S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 11).
MycoBank MB 838166.
Etymology: The epithet refers to the potential confusion with Strigula antillarum.
Typus: CHINA, Guangxi, Nanning, Long’an County, Longhu Mountain Natural Reserve, N 22°57′42″, E 107°37′40″, alt. 147 m, on living leaves, 1 December 2015, S.H. Jiang GX201511110 (HMAS–L0137209, holotype; ITS Genbank No. KX216696).
Diagnosis: Strigula pseudoantillarum is similar to S. antillarum (Fée) Müll. Arg. regarding the ascospores and aggregated pycnidia producing macroconidia, but it has a thin thallus and is also phylogenetically different from the latter.
Description: Thallus subcuticular, medium green and with a somewhat metallic glance, 0.5–7.5 mm diameter and 12.5–27.5 μm thick, with entire to crenulate margins, green. Photobiont a species of Cephaleuros, cells angular-rounded, 6–15 × 6–10 μm. Perithecia covered by thallus, hemispherical, 0.3–0.6 mm diameter and 120–180 μm high. Excipulum prosoplectenchymatous, 5–12.5 μm thick, colourless to brown. Involucrellum carbonaceous, 10–63 μm thick, black. Paraphyses unbranched. Asci obclavate, 62–75 × 10–12.5 μm. Ascospores eight per ascus, biseriate, fusiform, with acute ends, 15–28 × 4–6 μm. Pycnidia producing macroconidia often aggregated and confluent in the centre of thallus patches, wart-shaped, 0.1–0.15 mm diameter, black. Macroconidia bacillar, one-septate, 15–20 × 3–4 μm. Pycnidia producing microconidia not seen.
Chemistry: No substances detected by TLC.
Distribution and ecology: The new species was found on living leaves in humid, semi-exposed forest habitats of southern China.
Additional specimens examined: CHINA, Guangxi, Nanning, Long’an County, Longhu Mountain Natural Reserve, N 22°57′42″, E 107°37′40″, alt. 147 m, on living leaves, 1 December 2015, S.H. Jiang GX201511112 (HMAS–L0137208), GX201511114 (HMAS–L0137207), GX201511137 (HMAS–L0137667), GX201511166 (HMAS–L0137669), GX201511168 (HMAS–L0137668); Hainan, Dongfang city, Nanlang village, E’xian Ling, N 19°00′18″, E 109°04′09″, alt. 160 m, on living leaves, 13 December 2014, J.H. Wang & R.D. Liu HN2014376 (HMAS–L0130571), HN2014354 (HMAS–L0130622); N 19°00′07″, E 109°04′20″, alt. 126 m, J.H. Wang & R.D. Liu HN2014377 (HMAS–L0130572); Yunnan, Xishuangbanna, Mengla County, Tropical Botanical Garden of Chinese Academy of Sciences, East area, N 21°55′39″, E 101°15′52″, alt. 560 m, on living leaves, 18 November 2015, X.L. Wei & S.H. Jiang XTBG2015051 (HMAS–L0137211); Lvshilin, N 21°54′35″, E 101°16′52″, alt. 626 m, 18 November 2015, X.L. Wei & S.H. Jiang XTBG2015119 (HMAS–L0137210).
Notes:
Strigula pseudoantillarum is characterised by its aggregated, confluent pycnidia in the centre of the thallus. Together with the similar asci and ascospores, it thus resembles
S. antillarum, and the material was originally reported under this name from China [
48]. However, inclusion of authentic material of
S. antillarum from the Caribbean, the type region, showed that both taxa are phylogenetically distinct and not even closely related (
Figure 1 and
Figure 2). A consistent morphological difference is found in the thin thallus in the new species, compared to the thicker, somewhat bulging thallus patches in typical
S. antillarum [
48].
Strigula lacericola P.M. McCarthy, described from Australia, can be distinguished by the pseudostromatic groups of pycnidia and by the smaller ascospores and macroconidia [
72].
Strigula pseudosubtilissima S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 12).
MycoBank MB 838167.
Etymology: The epithet pseudosubtilissima refers to the similarity with the unrelated Racoplaca subtilissima.
Typus: CHINA, Hainan, Ledong County, Jianfengling, Mingfenggu, N 18°44′5″, E 108°52′8″, alt. 920 m, on living leaves, 11 December 2014, J.H. Wang & R.D. Liu HN2014130 (HMAS–L0130553, holotype; ITS Genbank No. MW344227).
Diagnosis: Strigula pseudosubtilissima is similar to Racoplaca subtilissima Fée in the thin, laciniate thallus bordered by a black line, but can be distinguished by the uniseriate, short ascospores, as well as the green thallus colour and fully exposed, black perithecia. It differs from the closely related S. nitidula in the rather long, free lobes.
Description: Thallus subcuticular, marginally with rather long, free laciniae, bordered by a thin, irregular, black line, 6–10 mm across and 7.5–12.5 μm thick, bright green with metallic glance. Photobiont a species of Cephaleuros, cells 8–14 × 4–6 μm. Perithecia completely exposed, almost conical, 0.2–0.6 mm diameter and 100–200 μm high, black and shiny. Excipulum prosoplectenchymatous, 10–20 μm thick, colourless to brown. Involucrellum carbonaceous, black, 15–30 μm thick. Paraphyses unbranched. Asci cylindrical or almost thread-like, very numerous and compact, 75–90 × 4–5 μm. Ascospores eight per ascus, uniseriate, ellipsoid, one-septate, with distinct constriction at septum and often broken into halves within the asci, 8–12 × 2.5–3 μm, 3–4 times as long as broad. Pycnidia usually rather few, often empty, 0.05–0.15 mm diameter. Pycnidia producing macroconidia and those producing microconidia are difficult to distinguish from their exterior. Macroconidia not seen. Microconidia fusiform, non-septate, 4–5 × 1.5–2 μm.
Chemistry: No substances detected by TLC.
Habitat and distribution: The new species was found on living leaves in humid, semi-exposed forest in southern China.
Additional specimens examined: CHINA, Guangxi, Shangsi County, Shiwan Mountain National Nature Reserve, N 21°54′13″, E 107°54′13″, alt. 264 m, on living leaves, 5 December 2015, S.H. Jiang GX201511356 (HMAS–L0139292); N 21°54′16″, E 107°54′09″, alt. 271 m, on living leaves, 26 May 2015, X.L. Wei & J.H. Wang GX20150341 (HMAS–L0139329), GX20150341_3 (HMAS–L0141582); Hainan, Ledong County, Jianfengling, Mingfenggu, N 18°44′49″, E 108°51′44″, alt. 828 m, on living leaves, 12 December 2014, J.H. Wang & R.D. Liu HN2014312_2 (HMAS–L0141619), HN2014256 (HMAS–L0130570), HN2014224 (HMAS–L0130607); N 18°44′5″, E 108°52′8″, alt. 920 m, on living leaves, 11 December 2014, J.H. Wang & R.D. Liu HN2014134 (HMAS–L0130591), HN2014141 (HMAS–L0130597); N 18°44′32″, E 108°50′32″, alt. 985 m, on living leaves, 12 December 2014, J.H. Wang & R.D. Liu HN2014284 (HMAS–L0130614); N 18°44′22″, E 108°51′58″, alt. 845 m, on living leaves, 11 December 2014, J.H. Wang & R.D. Liu HN2014009 (HMAS–L0130578); Lingshui, Diaoluo Mountain National Nature Reserve, N 18°43′36″, E 109°52′06″, alt. 930 m, on living leaves, 31 October 2016, X.Y. Liu HN2016003 (HMAS–L0139217); Qiongzhong County, Limu mountain, N 19°10′29″, E 109°44′53″, alt. 800 m, on living leaves, 10 September 2017, S.H. Jiang HN20170609 (HMAS–L0139633).
Notes: At first, this new species resembles
Racoplaca subtilissima, but it differs in the green thallus, the fully exposed, black perithecia, and the uniseriate, short ascospores breaking into halves. These latter features point to a close relationship with
S. nitidula, which is supported by the molecular data.
Strigula nitidula s.lat. often forms marginal black lines, akin to
R. subtilissima, but differs in the above features from the latter [
45].
Strigula nitidula was described from the Caribbean (Cuba), and the ITS data suggest that Cuban and Chinese material form a homogeneous clade representing a single species (
Figure 1), thus far the only species within the family demonstrated to be genuinely pantropical. Both the ITS data and the four-marker tree show that
S. pseudosubtilissima is phylogenetically distinct (
Figure 1 and
Figure 2), also differing from typical
S. nitidula morphologically in the rather long, partly free marginal laciniae.
Strigula pycnoradians S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 13).
MycoBank MB 838169.
Etymology: Epithet of the new species “pycnoradians” is a Latin word, and conveys the more striking radiating pycnidia.
Typus: THAILAND, Nakorn Nayok, Wang Takrai Park, N 14°13′15″, E 101°23′34″, alt. 120 m, on living leaves, 22 August 2017, W.C. Wang WWC342_1 (HMAS–L0139611, holotype; ITS Genbank No. MW344257).
Diagnosis: Strigula pycnoradians is closely related and morphologically similar to S. acuticonidiarum, but can be distinguished by the shorter asci and smaller ascospores.
Description: Thallus subcuticular, dispersed into rounded to partly confluent patches, 1–7 mm across and 15–65 μm thick, with more or less an entire margin, bright to dark green. Photobiont a species of Cephaleuros, cells angular-rounded, 5–15 × 4–10 μm. Perithecia immersed-erumpent, hemispherical, 0.2–0.35 mm diameter and 85–115 μm high, dark green due to a thallus cover but uppermost part often exposed and black. Excipulum prosoplectenchymatous, 7.5–12.5 μm thick, colourless to brown. Involucrellum carbonaceous, 12.5–37.5 μm thick, black. Paraphyses unbranched, c. 1–2 μm thick. Asci obclavate, 32–38 × 5–10 μm. Ascospores 8 per ascus, biseriate, fusiform, 1-septate but often appearing three-septate due to oil droplets when fresh, with slight constriction at the septum, 10–12.5 × 2.5–3.5 μm, 3–4 times as long as broad. Pycnidia producing macroconidia numerous, black, wart-shaped, clustered or in radiating rows, 0.1–0.15 mm in diameter. Macroconidia one-septate, 15–20 × 2.5–4 μm, with large mucilaginous cap at the distal end. Microconidia not seen.
Chemistry: No substances detected by TLC.
Habitat and distribution: The new species was found in humid, semi-exposed tropical forest in Thailand.
Additional specimens examined: THAILAND, Nakorn Nayok, Wang Takrai Park, N 14°13′15″, E 101°23′34″, alt. 120 m, 22 August 2017, W.C. Wang WWC353_1 (HMAS–L0139615), WWC360_1 (HMAS–L0141583).
Notes: The new species is mostly similar to the closely related
Strigula acuticonidiarum S.H. Jiang, X.L. Wei & J.C. Wei [
49]. Both share the general morphology of somewhat grouped or confluent pycnidia also seen in the neotropical
S. antillarum, which is, however, phylogenetically separate (
Figure 1).
Strigula pycnoradians differs from
S. acuticonidiarum in the often radiating rows of pycnidia and the larger mucilaginous cap of the macroconidia. In addition, the ascospores are much smaller (12.5–20 × 3.7–5 µm in
S. acuticonidiarum), leading also to much shorter asci (50–65 × 8–12 µm in
S. acuticonidiarum [
49]). The radiating pycnidia are reminiscent of those of
S. novae-zelandiae (Nag Raj) Sérus. and
S. oleistrata M. Ford, D.J. Blanchon & de Lange, both known from New Zealand [
73]. However, in these two species, the macroconidia are shorter (7–15 μm) and the thalli are typically much larger.
Strigula sinoconcreta S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 14).
MycoBank MB 838170.
Etymology: The epithet “sinoconcreta” refers to the region of the type material and the similarity with Strigula concreta.
Typus: CHINA, Hainan, Changjiang county, Bawangling National Nature Reserve, N 19°04′57″, E 109°07′29″, alt. 550 m, on living leaves, 5 September 2017, S.H. Jiang HN20171454 (HMAS–L0139630, holotype; ITS Genbank No. MW344261).
Diagnosis: The new species differs from Strigula concreta in the bright green thallus and the larger, less prominent, basally spreading perithecia.
Description: Thallus subcuticular, continuous or dispersed into rounded to irregular, partly confluent patches, typically with crenulate or shortly lobulate margins, pale greyish green to green, 1.5–5 mm across and 10–15 μm thick, sometimes occurring along the nerves. Photobiont a species of Cephaleuros, cells 8–14 × 4–6 μm. Perithecia wart-shaped to hemispherical, basally spreading, 0.4–0.7 mm diameter and 100–180 μm high, black. Excipulum prosoplectenchymatous, 7.5–12.5 μm thick, colourless to brown. Involucrellum carbonaceous, 50–75 μm thick, black. Paraphyses unbranched, thin. Asci cylindrical, 52.5–62.5 × 5–7.5 μm. Ascospores eight per ascus, uniseriate, ellipsoid, one-septate, with distinct constriction at septum and often broken into halves outside asci, 9–12.5 × 2–3 μm, 3–4 times as long as broad. Pycnidia wart-shaped, those producing macroconidia 0.1–0.15 mm, those producing microconidia 0.05–0.1 mm diameter, black. Macroconidia bacillar, non-septate, 4–5 × 1.5–2.5 μm, with appendage at one end or both ends c. 5–10 μm long. Microconidia fusiform, aseptate, 4–5 × 1.5–2 μm.
Chemistry: No substances detected by TLC.
Habitat and distribution: The new species was discovered in humid forest in southern China. Individuals mostly grew on living leaves with a rigid texture.
Additional specimens examined: CHINA, Hainan, Changjiang county, Bawangling National Nature Reserve, N 19°04′57″, E 109°07′29″, alt. 550 m, on living leaves, 5 September 2017, S.H. Jiang HN20171477 (HMAS–L0139631).
Notes:
Strigula sinoconcreta agrees with
S. concreta in general habit and in the uniseriate, short ascospores partly breaking into halves, but can be recognized by the bright green thallus (grey–green in
S. concreta [
45]) and the larger, flatter, basally somewhat spreading perithecia. The new species was phylogenetically distinct from
S. intermedia (
Figure 1 and
Figure 2) and differs from the latter in the thicker thallus not bordered by black lines.
Strigula stenoloba S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 15).
MycoBank MB 838171.
Etymology: The epithet “stenoloba” refers to the thin, meandering lobes.
Typus: CHINA, Hunan, Chenzhou city, Mangshan National Nature Reserve, N 24°56′59″, E 112°56′02″, alt. 1300 m, on living leaves, 13 September 2017, S.H. Jiang HN20170463 (HMAS–L0139622, holotype).
Diagnosis: Strigula stenoloba differs from other species in the genus by its very narrow, irregular lobes.
Description: Thallus subcuticular, continuous, occurring or not along the nerves, margins, and scars of leaves, formed of dichotomously branched, narrow, convex lobes that can fuse laterally and form a reticulate pattern, margins crenulate, 3–7 mm across and 10–15 μm thick. Photobiont a species of Cephaleuros, cells angular-rounded, 8–13 × 4–7 μm. Perithecia covered by thin, thallus layer up to ostiole, wart-shaped, 0.4–0.6 mm diameter and 150–230 μm high, greyish to greenish black. Excipulum prosoplectenchymatous, 5–7.5 μm thick, colourless to brown. Involucrellum carbonaceous, 35–50 μm thick, black. Paraphyses unbranched, thin. Asci obclavate, 50–75 × 7.5–10 μm. Ascospores eight per ascus, biseriate, fusiform-ellipsoid to fusiform, with acute or rather rounded ends, one-septate, with constriction at septum, 12.5–17.5 × 3–5 μm. Pycnidia exposed, wart-shaped, black and shiny, those producing macroconidia 0.1–0.2 mm, those producing microconidia 0.07–0.15 mm diameter. Macroconidia bacillar, one-septate, 12.5–15 × 2.5–3.7 μm, with 17–35 μm long appendage at both ends that may be terminated by a hook. Pycnidia producing microconidia not seen.
Chemistry: No substances detected by TLC.
Habitat and distribution: The new species was found on living leaves in humid, semi-exposed forest habitats in southern China.
Additional specimens examined: CHINA, Hunan, Chenzhou city, Mangshan National Nature Reserve, N 24°56′59″, E 112°56′02″, alt. 1300 m, on living leaves, 13 September 2017, S.H. Jiang HN20170451 (HMAS–L0139621).
Notes: Unfortunately, we were unable to obtain molecular data for this material. Yet, the morphological features are so unique that we decided to describe it formally as a new species. At first glance, one could consider this a barely lichenised form or an early ontogenetic stage, but the morphology was consistent across all individuals seen in the type collection and different from anything we have observed in other species. The formation of fully mature perithecia also contradicts the interpretation of this material as an aberrant form of a known species. The new species is characterised by its very thin, widely separated, partly anastomosing lobes. It should not be confused with
S. delicata Sérus., a species described from New Zealand [
74]. The latter has a much better developed thallus with narrow but regular lobes, although it agrees in ascospore and macroconidial size. Species of
Racoplaca also differ in the regular thallus lobes, which are flatter and typically olive-green to brownish [
45]. Somewhat similar is
R. tremens (Müll. Arg.) S.H. Jiang, Lücking & J.C. Wei, known from Brazil, which agrees in the somewhat irregular, narrow, more or less free laciniae, but the laciniae are completely flat and bordered by a thin, black line, the ascospores are longer (17–23 × 3–5 μm), and the involucrellum is thinner (15–25 μm thick); macroconidia are unfortunately not known [
45].
Strigula subtilissimoides S.H. Jiang, J.C. Wei & Lücking,
sp. nov. (
Figure 16).
MycoBank MB 838172.
Etymology: The thallus feature of this species is its similarity to Racoplaca subtilissima, although not related to the latter, so we decided to name it Strigula subtilissimoides.
Typus: CHINA, Guangxi, Shangsi County, Shiwan Mountain Forest Park, N 21°54′13″, E 107°54′13″, alt. 264 m, on living leaves, 5 December 2015, S.H. Jiang GX201511373 (HMAS–L0139253, holotype; ITS Genbank No. MK206352).
Diagnosis: Strigula subtilissimoides is intermediate between S. maculata and Racoplaca subtilissima in thallus, but the ascospores belong to the S. nitidula type. Phylogenetically different from the other S. nitidula clade.
Description: Thallus subcuticular, 5–25 mm across and 12.5–27.5 μm thick, with entire to crenulate margins to irregularly laciniate, pale to dark olive to brownish, sometimes with tiny verrucae on the surface, nitidous. Photobiont a species of Cephaleuros, cells angular-rounded, 8–12 × 4–6 μm. Perithecia exposed, conical to wart-shaped, 0.4–0.6 mm diameter and 120–200 μm high, black. Excipulum prosoplectenchymatous, 12.5–22.5 μm thick, brown. Involucrellum 22–60 μm thick, carbonaceous, black. Paraphyses unbranched and thin. Asci cylindrical or thread like, numerous and dense, 40–90 × 3–5 μm. Ascospores eight per ascus, uniseriate, ellipsoid, one-septate, sometimes with 1–2 oil droplets per cell when fresh, with distinct constriction at the septum and sometimes broken into halves outside the asci, 8.5–12.5 × 1.5–2.5 μm. Pycnidia producing macroconidia not seen. Pycnidia producing microconidia wart-shaped, 0.05–0.1 mm diameter, black. Microconidia fusiform, aseptate, 4–5 × 1.5–2 μm.
Chemistry: No substances detected by TLC.
Habitat and distribution: The new species was found on living leaves in humid, semi-exposed forest in southern China.
Additional specimens examined: CHINA, Guangxi, Shangsi County, Shiwan Mountain Forest Park, N 21°54′13″, E 107°54′13″, alt. 264 m, on living leaves, 5 December 2015, S.H. Jiang GX201511376 (HMAS–L0139293), GX201511384 (HMAS–L0139254), GX201511392 (HMAS–L0139256), GX201511393 (HMAS–L0139255); N 21°54′03″, E 107°54′26″, alt. 342 m, on living leaves, 25 May 2015, X.L. Wei & J.H. Wang GX20150270 (HMAS–L0139277); Hainan, Ledong County, Jianfengling, Mingfenggu, N 18°44′5″, E 108°52′8″, alt. 920 m, on living leaves, 11 December 2014, J.H. Wang & R.D. Liu HN2014149 (HMAS–L0130556), HN2014149_2 (HMAS–L0141629), J.H. Wang & R.D. Liu HN2014160_3 (HMAS–L0141599), J.H. Wang & R.D. Liu HN2014240_2 (HMAS–L0141389); 27 July 2009, J.C. Wei WJC034f11 (HMAS–L0139204).
Notes:
Strigula subtilissimoides is another new species besides
S. pseudosubtilissima (see above) that is morphologically similar to
Racoplaca subtilissima. In this case, the resemblance is even more striking, and superficial examination may place this material under the latter species. However, the more or less exposed, black perithecia, and particularly the uniseriate, cylindrical asci with short ascospores breaking into halves reveal a close relationship with
S. nitidula, which is supported by the molecular data, which also show that
S. pseudosubtilissima and
S. subtilissimoides are phylogenetically distinct (
Figure 1 and
Figure 2). Morphologically, both differ chiefly in thallus colour.
Strigula subtilissimoides is sister to
S. sinoaustralis S.H. Jiang, X.L. Wei & J.C. Wei, another taxon with
S. nitidula morphology but which features numerous small, white papillae on the thallus surface [
48].


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}















