
Novel Species and Records of Dictyosporiaceae from Freshwater Habitats in China and Thailand
by
 , , and
, , and
Hongwei Shen
1,2,3,
Danfeng Bao
1,3,4,
Dhanushka N. Wanasinghe
5 ,
,
Saranyaphat Boonmee
2,3,
Jiankui Liu
6 and
Zonglong Luo
1,* 1
College of Agriculture and Biological Science, Dali University, Dali 671003, China
2
School of Science, Mae Fah Luang University, Chiang Rai 57100, Thailand
3
Center of Excellence in Fungal Research, Mae Fah Luang University, Chiang Rai 57100, Thailand
4
Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai 50200, Thailand
5
Honghe Center for Mountain Futures, Kunming Institute of Botany, Chinese Academy of Sciences, Honghe 654400, China
6
Center for Informational Biology, School of Life Science and Technology, University of Electronic Science and Technology of China, Chengdu 611731, China
*
Author to whom correspondence should be addressed.
J. Fungi 2022, 8(11), 1200; https://doi.org/10.3390/jof8111200
Submission received: 9 October 2022
/
Revised: 4 November 2022
/
Accepted: 7 November 2022
/
Published: 14 November 2022
(This article belongs to the Special Issue Ascomycota: Diversity, Taxonomy and Phylogeny)
Abstract
:China and Thailand are rich in fungal diversity with abundant freshwater resources that are favorable for numerous fungal encounters. Resulting from this, the majority of the Dictyosporiaceae species reported were from these two countries. During the investigation on the diversity of lignicolous freshwater fungi in the Greater Mekong Subregion, eleven collections of cheirosporous species on submerged wood were collected from lentic and lotic habitats in China and Thailand. Phylogenetic analysis that combined nuclear small-subunit ribosomal RNA (SSU), internal transcribed spacer region (ITS), nuclear large subunit ribosomal RNA (LSU) and translation elongation factor 1α (tef 1-α) loci revealed six new species: Dictyocheirospora chiangmaiensis, D. multiappendiculata, D. suae, Digitodesmium aquaticum, Vikalpa grandispora and V. sphaerica. In addition, four known species were also identified and reported based on morphological and phylogenetic evidence. The detailed descriptions and illustrations of these taxa are provided with an updated phylogenetic tree of Dictyosporiaceae.
1. Introduction
Several studies investigating the ecology of aquatic fungi in early 2000 [1,2,3,4] recorded numerous species of cheiroid hyphomycetes, many in Dictyosporium and several other genera of Dictyosporiaceae. Dictyosporiaceae (Pleosporales) comprises 20 genera, the species of these genera are distributed worldwide and most taxa are saprobes on plant litter, especially dead or decaying wood in freshwater and terrestrial habitats [5,6,7,8,9,10,11,12,13,14]. Most of the asexual morphic members of the Dictyosporiaceae are hyphomycetous and characterized by the production of cheiroid, digitate, palmate and/or dictyosporous, and pale brown to brown conidia, viz., Aquadictyospora [8], Aquaticheirospora [7], Cheirosporium [6], Dictyocheirospora [5], Dictyopalmispora [5], Dictyosporium [15], Digitodesmium [16], Jalapriya [5], Neodigitodesmium [12], Pseudodictyosporium [17] and Vikalpa [5]. A few genera have coelomycetous asexual morphs, viz., Immotthia [18], Pseudocoleophoma [19], Pseudoconiothyrium [20], Pseudocyclothyriella [11], Sajamaea [21] and Verrucoccum [14]. Sexual morphs are known only from four genera Dyctiosporium [15], Gregarithecium [19], Immotthia [18], Pseudocoleophoma [19] and Verrucoccum [14].
Dictyosporium typified by D. elegans Corda [15] is characterized by effuse or compact granular colonies; micronematous, mononematous and branched conidiophores; monoblastic, terminal, determinate, doliiform, spherical or subspherical conidiogenous cells; and cheiroid, solitary, branched and complanate conidia, with or without appendages, comprising multiple rows of cells. The sexual morphs are characterized by subglobose ascomata, cylindrical asci and hyaline, fusiform, uniseptate ascospores, with or without a sheath [5,9,10,22]. Dictyosporium was placed in Pleosporales based on a phylogenetic analysis of LSU/SSU [23,24]. Liu et al. [25], Tanaka et al. [19] and Boonmee et al. [5] determined the phylogenetic position of Dictyosporium in Dictyosporiaceae (Pleosporales) based on multi-gene phylogeny analysis. Prasher and Verma [26] and Silva et al. [27] provided an account of Dictyosporium species with profiles and comparisons, including 48 accepted species. Since this publication, ten further species were recorded in this genus, and 13 species were relocated to Dictyocheirospora, Jalapriya and Vikalpa [5,9,10,19,26,28,29,30,31,32,33]. Currently, 54 species are accepted in Dictyosporium, of which, 17 were collected from the freshwater habitat, while 37 were collected from terrestrial habitats.
Dictyocheirospora was established by Boonmee et al. [5] to accommodate species with dark sporodochial colonies that produce aeroaquatic cheiroid, non-complanate or cylindrical conidia, mostly with conidial arms that are closely gathered together at the apex, and with Dictyocheirospora rotunda as the type species. Boonmee et al. [5] introduced three new species and transferred four species of Dictyosporium to Dictyocheirospora based on phylogenetic results. The difference between Dictyocheirospora and Dictyosporium is that the former’s conidia arms are on different planes and closely located at the apex, while in the latter, the conidia arms are closely arranged on the same plane. Based on these two morphological features, members of Dictyocheirospora and Dictyosporium are easily distinguished. Therefore, based on morphological and phylogenetic analysis, 23 species are accepted in Dictyocheirospora, including seven species that were transferred from Dictyosporium and one species from Cattanea [5,9,10,34,35,36,37,38,39,40].
Digitodesmium was introduced by Kirk [16] to accommodate D. elegans P.M. Kirk and it is characterized by the euseptate, cheiroid (digitate) conidia produced in sporodochia, the arms of conidia scattered in different planes and an apical gelatinous cap [12,16,35,41]. Currently, nine epithets of Digitodesmium are listed in Index Fungorum (accessed on 22 September 2022), except for Digitodesmium bambusicola L. Cai, K.Q. Zhang, McKenzie, W.H. Ho & K.D. Hyde; D. chiangmaiense Q.J. Shang & K.D. Hyde; D. polybrachiatum T.F. Nóbrega, B.W. Ferreira & R.W. Barreto; and D. tectonae (Rajeshk., Rajn. K. Verma, Boonmee, K.D. Hyde, Chandrasiri & Wijayaw.) W.H. Tian & Maharachch., where the remaining five species lack molecular sequence data [12,16,41,42,43,44]. Members of Digitodesmium are mainly distributed in terrestrial and freshwater environments in Asia and Europe [20,35,41,42,43,45]. Two species were reported in China, with both from submerged wood in freshwater environments, and one in Thailand reported from terrestrial environments [35,41,45].
Vikalpa was established by Boonmee et al. [5] to accommodate species characterized by sporododochial conidiomata, septate conidia and three rows of cells in different planes. Vikalpa is a sparse group that contains only four species, of which Vikalpa australiensis (B. Sutton) D’souza, Boonmee & K.D. Hyde; V. freycinetiae (McKenzie) D’souza, Boonmee & K.D. Hyd; and V. micronesiaca (Matsush.) D’souza, Bhat & K.D. Hyde were collected from decaying wood and leaves in terrestrial habitats of Australia, New Zealand and Micronesia [46,47,48], and only V. lignicola D’souza, Bhat, H.Y. Su & K.D. Hyde was from a lotic freshwater habitat in Yunnan, China [5]. The sequence data (ITS) is only available for the type species.
The Greater Mekong Subregion includes Yunnan Province in China and Thailand and is a highly diverse region for plants and fungi [49,50,51,52,53]. We continuously studied the lignicolous freshwater fungi along a north/south gradient of the region [54] and eleven fresh collections were collected. The aim was to (1) clarify the phylogenetic positions and taxonomy of the eleven collections based on morphological and phylogenetic analysis; (2) provide a comparison of conidial morphological characteristics of Dictyocheirospora, Digitodesmium and Vikalpa species; and (3) provide insights into the lignicolous freshwater taxa and increase knowledge of microfungi in the Greater Mekong Subregion.
2. Materials and Methods
2.1. Specimen Collection, Examination and Isolation
The specimens of submerged decaying wood were collected from lentic (lakes; Fuxian, Erhai and Luguhu) and lotic (streams; Mae sai, Pong Pham and three unknown) habitats in China and Thailand. The samples were selected from wood substrates submerged in water with a diameter of 1–2 cm and a length of about 20 cm, including tree branches, twigs and rotten branches of grasses. The collected samples were placed in plastic ziplock bags and were taken to the laboratory for processing.
Morphological observations were done following Luo et al. [55] and Senanayake et al. [56] with a few modifications. The samples were incubated in a plastic box at room temperature for one week. Macromorphological characters of samples were observed using an Optec SZ 760 compound stereomicroscope. Temporarily prepared microscope slides were placed under a Nikon ECLIPSE Ni-U compound stereomicroscope for observation and micro-morphological photography. The morphologies of colonies on native substrates were photographed with a Nikon SMZ1000 stereo zoom microscope.
Single spore isolations were performed as follows: the tip of a sterile toothpick dipped in sterile water was used to capture the conidia of the target colony directly from the specimen; the conidia were then streaked on the surface of water agar (WA) or potato dextrose agar (PDA) and incubated at room temperature overnight. The single germinated conidia were transferred to fresh PDA plates and incubated at room temperature. A few of the remaining germinated spores in the media plate were separated along with agar by using a needle and transferred onto water-mounted glass slides for photographs to capture the germination position of the germ tubes.
After finalizing the observation and isolation, the specimens were dried under natural light, wrapped in absorbent paper and placed in a ziplock bag with mothballs. Specimens were deposited in the herbarium of Mae Fah Luang University (MFLU), Chiang Rai, Thailand, and the Kunming Institute of Botany, Chinese Academy of Sciences (KUN-HKAS), Kunming, China. The cultures were deposited with the Mae Fah Luang University Culture Collection (MFLUCC), China General Microbiological Culture Collection Center (CGMCC) and Kunming Institute of Botany Culture Collection (KUNCC). Facesoffungi numbers and MycoBank numbers were obtained as described in Jayasiri et al. [57] and MycoBank (https://www.mycobank.org; accessed on 8 October 2022).
2.2. DNA Extraction, PCR Amplification and Sequencing
DNA extraction, PCR amplification, sequencing and phylogenetic analysis were done in accordance with the methods of Dissanayake et al. [58]. Mycelia for DNA extraction from each isolate were grown on PDA for 3–4 weeks at room temperature. Total genomic DNA was extracted from 100–300 mg axenic mycelium via scraping from the edges of the growing culture using a sterile scalpel and transferred to a 1.5 mL microcentrifuge tube using sterilized inoculum needles. The mycelium was ground to a fine powder with liquid nitrogen or quartz sand to break the cells for DNA extraction. When the cultures could not be maintained with some of the collected samples, fruiting structures (20–50 mg) were removed from the natural substrate using a sterile scalpel and placed on sterile paper, and then transferred to a 1.5 mL microcentrifuge tube. DNA was extracted with the TreliefTM Plant Genomic DNA Kit (TSP101) following the manufacturer’s guidelines.
Four gene regions, viz., ITS, LSU, SSU and tef1-α, were amplified using ITS5/ITS4 [59], LR0R/LR5 [60], NS1/NS4 [59] and EF1-983F/EF1-2218R [61] primer pairs, respectively. Primer sequences are available in the WASABI database on the AFTOL website (aftol.org). The PCR mixture contained 12.5 μL of 2 × GS Taq PCR MasterMix (mixture of DNA polymerase, dNTPs, Mg2+ and optimized buffer; Genesand Biotech, Beijing, China); 1 μL of each primer, including forward primer and reverse primer (10 μM); 1 μL template DNA extract; and 9.5 μL double-distilled water [55]. The PCR thermal cycling conditions of ITS and SSU were as follows: 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 56 °C for 50 s, elongation at 72 °C for 1 min and a final extension at 72 °C for 10 min; the LSU and TEF1-α thermal cycling conditions were as follows: 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 50 s, elongation at 72 °C for 1 min and a final extension at 72 °C for 10 min. PCR products were then purified using minicolumns, purification resin and buffer according to the manufacturer’s protocols. The sequences were carried out at the Beijing Tsingke Biological Engineering Technology and Services Co., Ltd. (Beijing, China).
2.3. Phylogenetic Analysis
BLAST searches using the BLASTn algorithm were performed to retrieve similar sequences from GenBank (http://www.ncbi.nlm.nih.gov, accessed on 05 May 2022) and relevant publications [10,40]. The sequences were aligned using the MAFFT online service: multiple alignment program MAFFT v.7 [62] (http://mafft.cbrc.jp/alignment/server/index.html, accessed on 11 May 2022), and sequence trimming was performed with trimAl v1.2 with default parameters (http://trimal.cgenomics.org (accessed on 11 May 2022) for specific operation steps) [63]. The sequence dataset was combined using SquenceMatrix v.1.7.8 [64]. FASTA alignment formats were changed to PHYLIP and NEXUS formats using the website Alignment Transformation Environment (ALTER) (http://sing.ei.uvigo.es/ALTER/, accessed on 21 September 2022).
Maximum likelihood (ML) analysis was performed by setting RAxML-HPC2 on XSEDE (8.2.12) [65,66] in the CIPRES Science Gateway [67] (http://www.phylo.org/portal2, accessed on 25 January 2022) using the GTR+GAMMA model with 1000 bootstrap repetitions. Bayesian analyses were performed in MrBayes 3.2.6 [68] and the best-fitting model of sequences evolution was estimated via MrModeltest 2.2 [69,70,71]. The Markov Chain Monte Carlo (MCMC) sampling approach was used to calculate posterior probabilities (PP) [72]. Bayesian analyses of six simultaneous Markov chains were run for 5 M generations and trees were sampled every thousand generations.
Phylogenetic trees were visualized using FigTree v1.4.0 (http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 13 May 2022)), while editing and typesetting were achieved using Adobe Illustrator (AI) (Adobe Systems Inc., the United States). The new sequences were submitted in GenBank and the strain information used in this study is provided in Table 1.
3. Results
3.1. Phylogenetic Analysis
The dataset was composed of the combined SSU, ITS, LSU and tef 1-α sequence data of 124 taxa in Dictyosporiaceae, including 4041 characters (including gaps) with Periconia igniaria (CBS 379.86 and CBS 845.96) as the outgroup taxon (Figure 1). Maximum likelihood (ML) analysis and Bayesian analysis produced similar topologies that were consistent across the major clades. The likelihood of the final tree is evaluated and optimized using GAMMA. The best RAxML tree with a final likelihood value of −32,818.079790 is presented in Figure 1. The matrix had 1738 distinct alignment patterns, with 32.59% undetermined characters or gaps. The estimated base frequencies were as follows: A = 0.241143, C = 0.262390, G = 0.283430 and T = 0.213037; substitution rates AC = 1.312486, AG = 3.111299, AT = 1.426646, CG = 1.005458, CT = 6.740140, GT = 1.000000, α = 0.275558 and tree-length = 2.908192. The Bayesian analyses generated 1408 trees (average standard deviation of split frequencies: 0.009910) from which 1056 were sampled after 25% of the trees were discarded as burn-in. Bootstrap support values with an ML greater than 75%, and Bayesian posterior probabilities (PP) greater than 0.97 are given above the nodes.
The multigene phylogenetic analysis showed that eleven of our new strains were nested in Dictyocheirospora, Dictyosporium, Digitodesmium and Vikalpa. Dictyocheirospora heptaspora (MFLUCC 22-0096) was clustered with two strains of D. heptaspora (CBS 396.59 and DLUCC 1992), D. aquadulcis (MFLUCC 22-0095) and D. nabanheensis (MFLUCC 22-0094), and Dictyosporium tubulatum (KUN-HKAS 115789) was clustered with their ex-type strains. A new taxon, viz., Dictyocheirospora chiangmaiensis (MFLUCC 22-0097), formed a distinct clade with D. clematidis (MFLUCC 17-2089), D. metroxylonis (MFLUCC 15-0028a and MFLUCC 15-0028b), D. taiwanense (MFLUCC 17-2654) and D. thailandica (MFLUCC 18-0987). Dictyocheirospora suae (KUNCC 22-12424) and D. multiappendiculata (KUNCC 22-10734 and KUNCC 22-10736) aggregated to form a separate clade in the genus Dictyocheirospora. Digitodesmium aquaticum (MFLU 22-0203) clustered as a sister clade with Di. bambusicola (CBS 110279) and Digitodesmium sp. (TBRC 10037 and BRC 10038) with strong support (100% ML/1.00 PP). Vikalpa grandispora (KUNCC 22-12425) clustered with the new species V. sphaerica (CGMCC 3.20682) with low support (84% ML), and clustered as a sister clade with V. australiensis (HKUCC 10304).
3.2. Taxonomy
Dictyocheirospora M.J. D’souza, Boonmee & K.D. Hyde, Fungal Diversity 80: 465 (2016).
Notes: The current study found three new species in Dictyocheirospora. We also re-collected Dictyocheirospora aquadulcis, D. heptaspora and D. nabanheensis from freshwater habitats in Thailand. These taxa are subsequently illustrated and described below.
Dictyocheirospora chiangmaiensis H.W. Shen, S. Boonmee & Z.L. Luo sp. nov., Figure 2.
MycoBank number: MB 846309.
Etymology: “chiangmaiensis” refers to the Chiang Mai Province, Thailand, where the species was collected.
Holotype: MFLU 22-0199.
Saprotrophic on submerged decaying wood in freshwater habitats. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on a natural substrate were punctiform, sporodochial, scattered and brown. Mycelium was composed of immersed or partly superficial, pale brown, septate, smooth, thin-walled and branched hyphae. Conidiophores were micronematous, mononematous, septate, cylindrical, pale brown to brown, smooth and thin-walled. Conidiogenous cells were holoblastic, cylindrical, septate, hyaline to pale brown, smooth and thin-walled. Conidia were (40–)42–46(–48) × (–14)16–18(–20) μm ( = 44 × 17 μm, n = 30), solitary, cheiroid, ellipsoid to cylindrical, brown, not complanate, composed of 4–6 rows of cells, euseptate, unseparated, each row with 9–10 cells, slightly bent inward at the apex and sometimes with hyaline subglobose appendages at the subapical. Conidial secession was schizolytic.
Culture characteristics: Conidia germinating on PDA within 12 h and germ tubes produced at the base (Figure 2i). Colony growth was slow, reaching 2 cm after 8 weeks at room temperature. Mycelium was loose, flocculent, smooth-edged, yellow on the forward, middle mastoid, edge brown to brown-red and middle black on the reverse side.
Material examined: Thailand, Chiang Mai Province, around the Mushroom Research Center (MRC); 19°07′05″ N, 98°45′40″ E, (680 m); on submerged decaying wood in a stream; 9 July 2020; H.W. Shen; and SHW 22 (MFLU 22-0199, holotype) and ex-type culture (MFLUCC 22-0097).
Notes: Phylogenetically, Dictyocheirospora chiangmaiensis forms a distinct lineage basal to four Dictyocheirospora species, i.e., D. clematidis (MFLUCC 17-2089) [32], D. metroxylonis (MFLUCC 15-0028a, MFLUCC 15-0028b) [37], D. taiwanense (MFLUCC 17-2654) [35] and D. thailandica (MFLUCC 18-0987) [38] with good support (97% ML/1.00 PP, Figure 1). Dictyocheirospora chiangmaiensis can be distinguished from other species in this clade by its smaller conidia with fewer rows (4–6) of cells (Table 2).
Dictyocheirospora suae H.W. Shen & Z.L. Luo, sp. nov., Figure 3.
MycoBank number: MB 846010.
Etymology: “suae” (Lat.) in memory of the Chinese mycologist Prof. Hong-Yan Su (4 April 1967–3 May 2022).
Holotype: KUN-HKAS 121703.
Saprotrophic on submerged decaying wood in a freshwater lake. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on a natural substrate were punctiform, sporodochial, scattered and brown. Mycelium was composed of immersed or partly superficial, brown to dark brown, septate, smooth, thin-walled and branched hyphae. Conidiophores were micronematous, mononematous, septate, cylindrical, pale brown, smooth and thin-walled. Conidiogenous cells were holoblastic, cylindrical, septate, hyaline to pale brown, smooth, thin-walled and sometimes lacked conidiogenous cells. Conidia were (65–)72–79 × (–17)20–25(–29) μm ( = 76 × 23 μm, n = 30), solitary, cheiroid, ellipsoid to cylindrical, not complanate, brown, composed of 5–7 rows of cells, unseparated, euseptate, tightly clustered at the apex of rows, each row with 12–15 cells and with globose to subglobose apical appendages. Conidial secession was schizolytic.
Culture characteristics: Conidia germinated on PDA within 12 h and germ tubes were produced at the base of cells (Figure 3i). Colonies on PDA reached about 4 cm in one month at room temperature. Mycelium was loose, flocculent and white with flaxen spots on the forward.
Material examined: China, Yunnan Province, Dali City, Erhai Lake; 25°49′04″ N, 100°08′46″ E, (1790 m); on submerged decaying wood; 1 April 2021; S.P. Huang; and 3EH GSC 8-6-2 (KUN-HKAS 121703, holotype) and ex-type cultures (KUNCC 22-12424).
Notes: Phylogenetic analysis showed that Dictyocheirospora suae clustered as a sister taxon to D. multiappendiculata with good support (95% ML/1.00 PP, Figure 1). Dictyocheirospora suae and D. multiappendiculata shared similar morphology in having solitary, cheiroid, ellipsoid to cylindrical and not complanate conidia with apical appendages. However, D. suae had larger conidia (72–79 × 20–25 μm vs. 36–46 × 13–18 μm) and more cells in each row (12–15 vs. 9–13). In addition, the conidia of D. multiappendiculata had more appendages than D. suae. A comparison of the ITS sequences of D. suae and D. multiappendiculata showed 2.35% (11/469 bp) nucleotide differences.
Dictyocheirospora multiappendiculata H.W. Shen & Z.L. Luo, sp. nov., Figure 4.
MycoBank number: MB 846014.
Etymology: “multiappendiculata” refers to the conidia of this species containing multiple appendages.
Holotype: KUN-HKAS 122866.
Saprotrophic on submerged decaying wood in a freshwater lake. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on a natural substrate were punctiform, sporodochial, scattered and brown. Mycelium was composed of immersed or partly superficial, pale brown, septate, smooth, thin-walled and branched hyphae. Conidiophores were micronematous, mononematous, septate, cylindrical, pale brown, smooth and thin-walled. Conidiogenous cells were (10–)11–18(–20) × (–3)4–8(–10) ( = 14 × 6 μm, n = 20) μm holoblastic, cylindrical to subglobose, hyaline to pale brown, smooth, thin-walled and easy to break. Conidia were (50–)55–62(–68) × (–16)19–22(–24) μm ( = 59 × 20 μm, n = 50), solitary, cheiroid, cylindrical, pale brown, non-complanate, smooth, closely appressed, composed of (5–)7 rows of cells, euseptate, each row with 9–13 cells, slightly bent inward at the apex, straight or slightly curved arms were inserted in different planes, with 1–5(–6) hyaline, globose to subglobose subapical appendages, and appendages were thin and easy to rupture and shrink. Conidial secession was schizolytic.
Culture characteristics: Conidia germinated on PDA within 12 h and germ tubes were produced at the basal cell (Figure 4k). Colonies on PDA reached about 4 cm in 1 month at room temperature. Mycelium was loose, flocculent, smooth-edged, white to light yellow on the surface and pale brown to brown in reverse.
Material examined: China, Yunnan Province, Yuxi City; on submerged decaying wood in Fuxian lake; 24°30′33″ N, 102°54′36″ E, (1700 m); 10 July 2021; H.W. Shen and Q.X. Yang; and L-963 (KUN-HKAS 122866, holotype) and ex-type cultures (KUNCC 22-10734). Furthermore, ibid.; 24°37′14″ N 102°51′03″ E, (1700 m); 12 July 2021; Y.K. Jiang and S. Luan; and L-987 (KUN-HKAS 122870) and living cultures (KUNCC 22-10736).
Notes: Dictyocheirospora multiappendiculata shared similar morphological characters with other Dictyocheirospora species in having cheiroid, cylindrical, euseptate and non-complanate conidia that were densely clustered at the apex. The globose to subglobose apical or subapical appendages were similar to D. hydei, D. indica, D. musae, D. nabanheensis, D. pseudomusae, D. suae and D. tetraploides. However, D. multiappendiculata was distinguished from these species by the numbers of appendages (Table 2). In the phylogenetic analysis, D. multiappendiculata and D. suae formed a sister lineage with strong support (95% ML/1.00 PP, Figure 1). For the nucleotide comparison, see the Notes section of D. suae. Following Jeewon and Hyde’s [75] recommendations for establishing species boundaries and new taxa among fungi, we introduce D. multiappendiculata as a new species here.
Dictyocheirospora aquadulcis Sorvongxay, S. Boonmee & K.D Hyde, Fungal Diversity 96: 23 (2019), Figure 5.
Index Fungorum number: IF556308; Facesoffungi number: FoF05963.
Material examined: Thailand, Chiang Rai Province, Muang Chiang Rai District; 20°00′14″ N, 99°43′01″ E; on submerged decaying wood in a stream; 2 January 2021; H.W. Shen; and CR 8-29 (MFLU 22-0201) and living culture (MFLUCC 22-0095).
Notes: Phylogenetic analysis showed that our new collection MFLUCC 22-0095 clustered with the type strain of Dictyocheirospora aquadulcis (MFLUCC 17-2571) with low support (0.97% PP, Figure 1). Our taxon fit well with the morphological characteristics of D. aquadulcis in having cheiroid, ellipsoid to cylindrical and euseptate conidia consisting of seven rows of cells. However, our strain (MFLUCC 22-0095) comprised hyaline globose to subglobose appendages at the conidia apical or subapical, which are lacking in D. aquadulcis [35]. There were 3 bp nucleotide (491/494, including 2 bp of gaps) differences of ITS between D. aquadulcis (MFLUCC 22-0095) and D. aquadulcis (MFLUCC 17-2571). Based on the morphological and phylogenetic analysis, we therefore identified our new collected strain as D. aquadulcis.
Dictyocheirospora nabanheensis Tibpromma & K.D. Hyde, Fungal Diversity 93: 10 (2018), Figure 6.
Index Fungorum number: IF554474; Facesoffungi number: FoF04483.
Material examined: Thailand, Chiang Rai Province, Muang Chiang Rai District; 20°00′1″ N, 99°43′01″ E; on submerged decaying wood in a stream; 2 Jan 2021; H.W. Shen; and CR 8-23 (MFLU 22-0200) and living culture (MFLUCC 22-0094).
Notes: Based on the phylogenetic analysis of the combined SSU, ITS, LSU and tef 1-α sequence data, our new collection (MFLUCC 22-0094) clustered with the ex-type strain of Dictyocheirospora nabanheensis (KUMCC 16-0152) with high support (99% ML/1.00 PP, Figure 1). Morphologically, this new collection resembled the holotype of D. nabanheensis in having hyaline globose to subglobose appendages in the apical region [9] but with a greater number of appendages (1–5 vs. 1–2). We identified our collection as D. nabanheensis. Dictyocheirospora nabanheensis was found on dead leaves of Pandanus sp. in a terrestrial habitat in Yunnan, China, whereas our new collection was found on submerged decaying wood in a freshwater habitat in Thailand. Our study expanded the geographical distribution of this species and showed that it adapted to different ecological niches.
Dictyocheirospora heptaspora (Garov.) M.J. D’souza, Boonmee & K.D. Hyde, Fungal Diversity 80: 469 (2016), Figure 7.
Index Fungorum number: IF 551589.
Material examined: Thailand, Chiang Rai Province, Muang Chiang Rai District; 20°00′14″ N, 99°43′01″ E; on submerged decaying wood in a stream; 2 January 2021; H.W. Shen; and CR 8-30 (MFLU 22-0202) and living culture (MFLUCC 22-0096).
Notes: Multigene phylogenetic analysis showed that Dictyocheirospora heptaspora was related to D. aquadulcis. Dictyocheirospora heptaspora was transferred from Dictyosporium to Dictyocheirospora by Boonmee et al. [5] based on the phylogenetic evidence of ITS sequences data. Tsui et al. [24] provided ITS and SSU sequence data for strain CBS 395.59 but there is no morphological description support for this strain. Hongsanan et al. [76] provided the ITS, LSU and TEF sequence data for D. heptaspora (DLUCC 1992) collected from submerged decaying wood in Yunnan, China. Morphologically, D. aquadulcis is almost identical to D. heptaspora. Goh et al. [22], Prasher and Verma [26], and Hongsanan et al. [76] provided a detailed morphological description for D. heptaspora, where the conidia are characterized by being cheiroid, ellipsoid to cylindrical, consisting of mostly 7 (5–7) rows of cells closely appressed together, each row composed of 10–18 cells, 50–86 µm long and 19–30 µm wide. Based on the comparative analysis of the ITS gene region, the nucleotide difference between each strain of D. heptaspora and D. aquadulcis strains was less than 1.5%, and thus, they should be merged into one species following the recommendations of Jeewon and Hyde [75]. However, we consider that the description of the type strain of D. heptaspora is vague and there is no sequence data. Therefore, the holotype of D. heptaspora needs to be examined to determine whether they belong to the same species.
The morphological characteristics of our new collection fit well with D. heptaspora [5,22]. In addition, the phylogenetic analysis showed that a new collection MFLUCC 22-0096 clustered with isolates of D. heptaspora (CBS 396.59 and DLUCC 1992). Although Dictyocheirospora heptaspora was isolated several times from submerged wood in Thailand, none of the previous strains were sequenced [22]; we provide the sequences for this species.
Dictyosporium Corda, Beiträge zur gesammten Natur- und Heilwissenschaften: 87 (1836).
Notes: Members of the Dictyosporium are distributed worldwide and are mainly saprotrophic on dead wood, decaying leaves and plant litter from terrestrial and aquatic habitats [10,23,24,25,26,27]. Dictyosporium currently contains 54 species, but there are still no molecular sequence data for most species. In this study, we discovered Dictyosporium tubulatum for the first time in China on submerged decaying wood in a small stream in Yiliang County, Yunnan Province, and provided ITS, LSU, SSU and tef 1-α sequence data.
Dictyosporium tubulatum J. Yang, K.D. Hyde & Z.Y. Liu, MycoKeys 36: 94 (2018), Figure 8.
Index Fungorum number: IF450470; Facesoffungi number: FoF04677.
Saprotrophic on submerged decaying wood in a freshwater habitat. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on a natural substrate were punctiform, sporodochial, scattered and dark brown to black. Mycelium was composed of partly immersed, partly superficial, hyaline to pale brown, septate and branched hyphae. Conidiophores were micronematous, mononematous, septate, cylindrical, pale brown to brown, smooth and thin-walled. Conidiogenous cells were (3–)5–10(–11) × 2–5(–7) μm ( = 8 × 3 μm, n = 30), monoblastic, cylindrical, terminal, determinate, septate, pale brown to brown, smooth, thin-walled and sometimes swollen. Conidia were (18–)21–26(–27) × (7–)12–15(–18) μm ( = 23 × 14 μm, n = 50), acrogenous, solitary, brown to dark brown, cheiroid, complanate, composed of four arms close together, 8–11 euseptate in each arm, side arms lighter than the middle arm, with hyaline, tubular, elongated appendages, (12–)14–20(–27) × (3–)4–5(–6) μm ( = 17 × 4 μm, n = 40) and attached at the apical part of two outer arms. Conidial secession was schizolytic.
Material examined: China, Yunnan Province, Kunming City, Yiliang County; 24°39′27″ N, 103°08′05″ E, (2100 m); on submerged decaying wood in a small stream; 16 May 2021; H.W. Shen; YL 3-72-1 (KUN-HKAS 115789). We tried to obtain pure cultures on common fungal isolation media (PDA, MEA, WA, CMA), but its conidia did not germinate on any of the media. We therefore obtained DNA sequences directly from clean colonies on natural substrates.
Notes: Multigene phylogenetic analysis showed that our new collection Dictyosporium tubulatum (KUN-HKAS 115789) clustered with isolates of D. tubulatum (MFLUCC 15-0631 and MFLUCC 17-2056). Morphologically, our collection fit well with the description of a holotype (MFLU 15-1166) in having four-armed, cheiroid and complanate conidia with hyaline, tubular and elongated appendages [10]. Based on the morphological characteristics and phylogenetic support, we identified the new collection as D. tubulatum. The type strain of Dictyosprium tubulatum (MFLUCC 15-0631) was collected from a freshwater habitat in Thailand, while our new collection was collected from freshwater habitat in China. It is a new record for China; therefore, our study suggested that this species was distributed geographically.
Digitodesmium P.M. Kirk, Transactions of the British Mycological Society 77: 284 (1981).
Notes: Digitodesmium accommodated a group of species with divergent/closely gathered conidial rows, including D. bambusicola, whose conidia produced only three rows on the same plane [42]. Based on morphological features and phylogenetic analysis, we introduce another species here, viz., Digitodesmium aquaticum, that produced three rows in the same plane.
Digitodesmium aquaticum H.W. Shen, S. Boonmee & Z.L. Luo, sp. nov., Figure 9.
MycoBank number: MB 846307.
Etymology: “aquaticum” refers to the aquatic environment where this species was collected.
Saprotrophic on submerged decaying wood in a freshwater habitat. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on a natural substrate were punctiform, sporodochial, scattered, dark brown and white, and flaky. Mycelium were composed of partly immersed, partly superficial, hyaline to pale brown, septate and branched hyphae. Conidiophores were micronematous, mononematous, septate, cylindrical, hyaline to pale brown, thin-walled and unbranched. Conidiogenous cells were (3–)5–7(–9) × 3–4(–5) μm ( = 6 × 4 μm, n = 35), monoblastic, holoblastic, cylindrical sometimes flat at the base, septate, hyaline to pale brown, smooth and thin-walled. Conidia were (39–)41–44(–46) × (17–)19–21(–22) μm ( = 43 × 20 μm, n = 35), acrogenous, solitary, cheiroid, brown, complanate, composed of three arms, 6–9 euseptate in each arm, truncated at the basal cell, and slightly bent inward and hyaline at the apex cell of each arm. Conidial secession was schizolytic.
Material examined: Thailand, Chiang Rai Province, Muang Chiang Rai District; 19°57′55″ N, 99°41′10″ E; on submerged decaying wood in a stream; 16 January 2021; H.W. Shen; and CR 9-6 (MFLU 22-0203, holotype). We tried to obtain pure cultures on common fungal isolation media (PDA, MEA, WA, CMA) but its conidia did not germinate on any of the media. We therefore obtained DNA sequences directly from clean colonies on natural substrates.
Notes: The phylogenetic analysis showed that our collection Digitodesmium aquaticum (MFLU 22-0203) clustered with a type strain of Di. bambusicola (CBS 110279) and unidentified species Di. sp. (TBRC 10,037 and TBRC 10038) with strong support (100% ML and 1.00 PP, Figure 1) but formed a distinct lineage. Comparison of the ITS and LSU sequences data of Di. aquaticum and Di. bambusicola showed 99.39% (489/492 bp) and 99.24% (521/525 bp, including one gap) sequence identity, respectively. Morphologically, Digitodesmium aquaticum resembled Di. bambusicola in having cheiroid, euseptate and complanate conidia. However, Di. bambusicola (CBS 110279) had smaller conidia (24–32.5 × 12.5–23 μm vs. 41–44 × 19–21 μm), with hyaline, thin-walled, globose to subglobose or clavate appendages, and apical or subapical on the arms, which was distinct from our new species [42]. Digitodesmium bambusicola (PDD 74494) was collected from the submerged bamboo culms in a river in Philippines, while our new species was collected from submerged decaying wood in a stream in Thailand. Therefore, Digitodesmium aquaticum can be recognized as a phylogenetically distinct species and described in this study.
Vikalpa D’souza, Boonmee, Bhat & K.D. Hyde, Fungal Diversity 80: 479 (2016).
Notes: Vikalpa has four species, and currently only the type species Vikalpa australiensis HKUCC 8797 has unique ITS sequence data. This study contributed two additional novel species from freshwater habitats, viz., V. grandispora and V. sphaerica. Detailed descriptions and colored photoplates are given below.
Vikalpa grandispora H.W. Shen, S. Boonmee & Z.L. Luo, sp. nov., Figure 10.
MycoBank number: MB 846015.
Etymology: “grandispora” refers to the large conidia.
Holotype: KUN-HKAS 122867.
Saprotrophic on submerged decaying wood in a freshwater habitat. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on a natural substrate were punctiform, sporodochial, scattered and brown. Mycelium were composed of immersed, septate, branched and hyaline to pale brown hyphae. Conidiophores were micronematous, mononematous, septate, subglobose to cylindrical, unbranched, hyaline to pale brown, smooth, thin-walled and sometimes reduced to conidiogenous cells. Conidiogenous cells were holoblastic, subglobose, smooth and thin-walled. Conidia were (33–)38–49(–53) × (9–)13–18(–21) μm ( = 44 × 16 μm, n = 50), solitary, cheiroid, pale brown to brown, not complanate, with 3 rows in different planes, arms closely appressed when young, separated at maturity, 9–12 cells in each row (5–)6 × 7 μm wide, euseptate, irregular, constricted at the septa and guttulate; appendages were more inflated than the apical cell, (4–)5–6(–7) × (5–)6–7(–9) μm ( = 5 × 7 μm, n = 50), hyaline and globose to subglobose. Conidial secession was schizolytic.
Culture characteristics: Conidia germinated on PDA within 12 h and germ tubes were produced at the basal cell (Figure 10j). Colonies grew on PDA; mycelium grew slowly, reaching about 2.5 cm in 1 month at room temperature and were loose, flocculent, smooth, white to creamy-yellow on the surface, orange on the edge and gray in the middle on the reverse side.
Material examined: China, Yunnan Province, Lijiang City, Yadong village; 26°58′08″ N, 100°24′29″ E, (2160 m); on submerged decaying wood in a small stream; 20 October 2021; H.W. Shen; and H593 (KUN-HKAS 122867, holotype) and ex-type culture (KUNCC 22-12425).
Notes: Vikalpa grandispora resembled V. australiensis, V. freycinetiae, V. micronesiaca and V. sphaerica in having solitary, cheiroid, non-complanate and three-armed conidia [46,47,48]. Vikalpa grandispora could be easily distinguished from other species by its larger conidia (Table 2). The phylogenetic analysis of the combined SSU, ITS, LSU and tef1-α sequence data showed that V. grandispora clustered as a sister lineage to V. sphaerica with low support (84% ML, Figure 1). The nucleotide comparison of the SSU, ITS, LSU and tef1-α genes of V. grandispora and V. sphaerica revealed 9 bp (2%), 5 bp (0.6%, including gaps), 5 bp (0.9%, including gaps) and 18 bp (2%) nucleotide differences, respectively. Therefore, we identified V. grandispora as a new species in Vikalpa based on the distinguished morphology and phylogenetic evidence.
Vikalpa sphaerica H.W. Shen & Z.L. Luo, sp. nov., Figure 11.
MycoBank number: MB 846016.
Etymology: “sphaerica” refers to the spherical appendages.
Holotype: KUN-HKAS 115805.
Saprotrophic on submerged decaying wood in a freshwater habitat. Sexual morph: undetermined. Asexual morph: hyphomycetous. Colonies on a natural substrate were punctiform, sporodochial, scattered and dark brown. Mycelium were composed of immersed or partly superficial, septate, branched and hyaline to pale brown hyphae. Conidiophores were micronematous, mononematous, septate, cylindrical, unbranched, hyaline to pale brown, smooth-walled and sometimes reduced to conidiogenous cells. Conidiogenous cells were 4–7(–9) × (2–)3–4(–5) (M = 5 × 4 μm, n = 15), holoblastic, cylindrical, sometimes flat at the base, septate, hyaline to pale brown and smooth-walled. Conidia were (23–)26–30(–34) × (11–)16–19(–20) μm ( = 28 × 17 μm, n = 50), solitary, cheiroid, pale brown to brown, not complanate, usually composed of 3–4 rows, rarely with 3 rows, rows separated from each other, 5–9 cells in each row, euseptate, irregular, constricted at the septa, guttulate, with hyaline, globose to subglobose appendages (6–)7–8(–9) × (5–)7–8(–9) μm ( = 8 × 8 μm, n = 50) at the apical cells and a few rows had either no appendages or they had fallen off. Conidial secession schizolytic.
Culture characteristics: Conidia germinated on PDA within 12 h and germ tubes were produced at both ends (Figure 11m). Colony growth on PDA; growth was slow, reaching about 4 cm in 1 month at room temperature. Mycelium was loose, flocculent, smooth edged, brownish-red on the forward, orange on the edge and gray in the middle on the reverse side.
Material examined: China, Yunnan Province, Lijiang City, Luguhu Lake; 27°42′11″ N, 100°48′18″ E, (2700 m); on submerged decaying wood in a small stream; 04 March 2021; Z.Q. Zhang and L. Sha; and L185 (KUN-HKAS 115805, holotype) and ex-type cultures (CGMCC3.20682 = KUNCC 21-10711).
Notes: Phylogenetic analysis placed our new taxon Vikalpa sphaerica (CGMCC 3.20682) close to V. grandispora with low support (84% ML, Figure 1). Morphologically, V. sphaerica resembled V. grandispora, V. australiensis and V. freycinetiae in having solitary and cheiroid conidia with apical appendages [48]. However, V. grandispora had longer conidia (38–49 vs. 26–30) and more cells in each row (9–12 vs. 5–9). The rows of V sphaerica were separated from each other and had fewer cells in each row than V. australiensis (5–9 vs. 7–11), while V. freycinetiae had a greater number of cells (9–13 vs. 5–9) that were larger (31–43 μm vs. 26–30 μm) than V. sphaerica [46]. Therefore, V. sphaerica is introduced here as a new species based on the morphological and phylogenetic analysis.
4. Discussion
Dictyosporiaceae accommodates a holomorphic group of Dothideomycetes that can produce cheiroid (digitate) and septate conidia [5]. Dictyosporaceae was recently well-studied based on the combination of morphological and phylogenetic analysis. However, due to the lack of valuable cultures and molecular sequence data, the taxonomic status of several earlier-described genera with fewer members remains confusing and ambiguous. Therefore, morphological identification continues to play a crucial role in the identification of species in Dictyosporiaceae.
DNA sequence data are available for most cheirosporous genera, except Dictyopalmispora. Therefore, the distinction between Dictyopalmispora and other cheirosporous genera is mainly based on unique hair-like appendages produced on all arms of Dictyopalmispora [5]. Four species are accepted in Vikalpa, including V. lignicola from freshwater environments, and only V. australiensis has ITS sequence data. The species of Vikalpa are distinguished from other genera mainly by the three rows of conidia on different planes produced in sporododochial conidiomata [5]. In this study, V. grandispora and V. sphaerica were introduced from freshwater lotic and lentic environments to enrich the habitat and distribution area of Vikalpa species.
The species of Digitodesmium is morphologically confused with Dictyosporium, but some morphological features can be used to distinguish them from each other: the arms of Digitodesmium are separated at the apex, while those in Dictyosporium are not separated. In addition, the conidial secession of Digitodesmium is schizolytic, while it is rhexolytic in Dictyosporium [6,42,43]. Digitodesmium polybrachiatum is easily confused with the Dictyocheirospora species in having non-complanate, cheiroid conidia and the arms being closely compacted at the apex. Therefore, using morphological characteristics alone, they cannot be properly distinguished [44]. Multigene phylogeny showed that members of Digitodesmium are divided into two main clades: Clade 1 is composed of Di. bambusicola and Di. aquaticum, with the main feature being that the conidia are produced in sporodochia, consisting of three rows of cells on one plane and they are euseptate. Clade 2 is composed of Di. chiangmaiense and Di. polybrachiatum, which are mainly characterized by their conidia arms being closely gathered at the apex, and they are euseptate, with or without appendages. There is a clear phylogenetic distance between these two clades, which strongly suggests that Di. bambusicola and Di. aquaticum belong to a different genus. However, there are no valuable culture and sequence data for the five species in the genus, including type species Di. elegans, to ensure which clade has been adapted as the type species; therefore, we agree with the suggestions of Nobrega et al. [44] and still maintain the taxonomic status of the species in the two clades.
Cheirosporous hyphomycetes are widely distributed as saprotrophic fungi on various plant debris substrates in freshwater and terrestrial habitats worldwide [5,6,7,8,9,10,11,12,13,14]. Although cheirosporous hyphomycetes are distributed globally, the main species contribution is from the Greater Mekong Subregion (GMC, Thailand and Yunnan, China) in Asia, accounting for more than 50% of the total. The Greater Mekong Subregion is rich in cheirosporous hyphomycetes resources, and species on wood substrates in freshwater habitats account for more than 50% of the total in this region. We are conducting research on lignicolous freshwater fungi in this region, which provides insights into the lignicolous freshwater taxa and increases knowledge of microfungi in the Greater Mekong Subregion.
Author Contributions
Conceptualization, Z.L., S.B. and D.N.W.; methodology, H.S.; formal analysis, H.S.; investigation, H.S. and D.B.; resources, Z.L. and S.B.; data curation, H.S.; writing—original draft preparation, H.S.; writing—review and editing, D.B., D.N.W., S.B., J.L. and Z.L.; funding acquisition, Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (project IDs: 32060005 and 31970021) and the Yunnan Fundamental Research Project (grant nos. 202101AU070137 and 202201AW070001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Shaun Pennycook for checking the nomenclature. Hong-Wei Shen thanks Long-Li Li, Qiu-Xia Yang, Sha Luan, Si-Ping Huang, Yan Tao and Zheng-Quan Zhang for their help with the sample collection, DNA extraction and PCR amplification. Thanks to Rong-Ju Xu for his help with the specimen and culture preservation. We thank Abhaya Balasuriya under the Reinventing Visiting Professor Program 2022, Mae Fah Luang University, for his valuable correction and suggestion.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ho, W.H.; Hyde, K.D.; Hodgkiss, I.J. Yanna Fungal communities on submerged wood from streams in Brunei, Hong Kong, and Malaysia. Mycol. Res. 2001, 105, 1492–1501. [Google Scholar] [CrossRef]
- Ho, W.H.; Yanna; Hyde, K.D.; Hodgkiss, I.J. Seasonality and sequential occurrence of fungi on wood submerged in Tai Po Kauforest stream, Hong Kong. Fungal Divers. 2002, 10, 21–43. [Google Scholar]
- Cai, L.; Zhang, K.Q.; McKenzie, E.H.C.; Hyde, K.D. Freshwater fungi from bamboo and wood submerged in the Liput River in the Philippines. Fungal Divers. 2003, 13, 1–12. [Google Scholar]
- Luo, J.; Yin, J.F.; Cai, L.; Zhang, K.Q.; Hyde, K.D. Freshwater fungi in lake Dianchi, a heavily polluted lake in Yunnan, China. Fungal Divers. 2004, 16, 93–112. [Google Scholar]
- Boonmee, S.; D’souza, M.J.; Luo, Z.; Pinruan, U.; Tanaka, K.; Su, H.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Taylor, J.E.; et al. Dictyosporiaceae fam. nov. Fungal Divers. 2016, 80, 457–482. [Google Scholar] [CrossRef]
- Cai, L.; Guo, X.Y.; Hyde, K.D. Morphological and molecular characterisation of a new anamorphic genus Cheirosporium from freshwater in China. Persoonia 2008, 20, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Kodsueb, R.; Lumyong, S.; Ho, W.H.; Hyde, K.D.; Mckenzie, E.H.; Jeewon, R. Morphological and molecular characterization of Aquaticheirospora and phylogenetics of Massarinaceae (Pleosporales). Bot. J. Linn. Soc. 2007, 155, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Li, W.L.; Luo, Z.L.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Su, H.Y.; Hyde, K.D. Lignicolous freshwater fungi from China I: Aquadictyospora lignicola gen. et sp. nov. and new record of Pseudodictyosporium wauense from northwestern Yunnan Province. Mycosphere 2017, 8, 1587–1597. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 92, 1–160. [Google Scholar] [CrossRef]
- Yang, J.; Liu, J.-K.; Hyde, K.D.; Jones, E.B.G.; Liu, Z.-Y. New species in Dictyosporium, new combinations in Dictyocheirospora and an updated backbone tree for Dictyosporiaceae. MycoKeys 2018, 36, 83–105. [Google Scholar] [CrossRef]
- Jiang, H.B.; Jeewon, R.; Karunarathna, S.C.; Phukhamsakda, C.; Doilom, M.; Kakumyan, P.; Suwannarach, N.; Phookamsak, R.; Lumyong, S. Reappraisal of Immotthia in Dictyosporiaceae, Pleosporales: Introducing Immotthia bambusae sp. nov. and Pseudocyclothyriella clematidis comb. et gen. nov. based on morphology and phylogeny. Front. Microbiol. 2021, 12, 656235. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.H.; Chen, Y.P.; Maharachchikumbura, S.S.N. Neodigitodesmium, a novel genus of family Dictyosporiaceae from Sichuan Province, China. Phytotaxa 2022, 559, 176–184. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Atienza, V.; Hawksworth, D.L.; Pérez-Ortega, S. Verrucoccum (Dothideomycetes, Dictyosporiaceae), a new genus of lichenicolous fungi on Lobaria s. lat. for the Dothidea hymeniicola species complex. Mycologia 2021, 113, 1233–1252. [Google Scholar] [CrossRef]
- Corda, A.J.C. Mykologische Beobachtungen. In Beitrage zur Gesammtem Natur-und Heilwissenschaften, 1st ed.; Weitenweber, W.R., Ed.; Commission bei Kronberger und Weber: Prague, Czech, 1836; Volume 1, pp. 80–88. [Google Scholar]
- Kirk, P.M. New or interesting microfungi II. Dematiaceous hyphomycetes from Esher Common, Surrey. Trans. Br. Mycol. Soc. 1981, 77, 279–297. [Google Scholar] [CrossRef]
- Kobayasi, Y. Mycological reports from New Guinea and the Solomon Islands (1–11). Bull. Nat. Sci. Mus. Tokyo 1971, 14, 367–551. [Google Scholar]
- Barr, M.E. New taxa and combinations in the loculoascomycetes. Mycotaxon 1987, 29, 501–505. [Google Scholar]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Schumache, R.K.; Akulov, A.; Thangavel, R.; Hernández Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfeld, M.J.; Summerel, B.A.; Quaedvlieg, W.; et al. New and interesting fungi. 2. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Piątek, M.; Rodriguez-Flakus, P.; Domic, A.; Palabral-Aguilera, A.N.; Gómez, M.I.; Flakus, A. Phylogenetic placement of Leptosphaeria polylepidis, a pathogen of Andean endemic Polylepis tarapacana, and its newly dis-covered mycoparasite Sajamaea mycophila gen. et sp. nov. Mycol. Prog. 2020, 19, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Goh, T.; Hyde, K.; Ho, W.; Yanna. A revision of the genus Dictyosporium, with descriptions of three new species. Fungal Divers. 1999, 2, 65–100. [Google Scholar]
- Crous, P.W.; Braun, U.; Wingfield, M.J.; Wood, A.R.; Shin, H.D.; Summerell, B.A.; Alfenas, A.C.; Cumagun, C.J.R.; Groenewald, J.Z. Phylogeny and taxonomy of obscure genera of microfungi. Persoonia 2009, 22, 139–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsui, C.; Berbee, M.; Jeewon, R.; Hyde, K. Molecular phylogeny of Dictyosporium and allied genera inferred from ribosomal DNA. Fungal Divers. 2006, 21, 157–166. [Google Scholar]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Prasher, I.B.; Verma, R.K. Two new species of Dictyosporium from India. Phytotaxa 2015, 204, 193–202. [Google Scholar]
- Silva, C.R.; Gusmão, L.F.P.; Castañeda-Ruiz, R.F. Dictyosporium amoenum sp. nov from Chapada, Diamantina, Bahia, Brazil. Mycotaxon 2016, 130, 1125–1133. [Google Scholar] [CrossRef]
- Abdel-Aziz, F.A. Two new cheirosporous asexual taxa (Dictyosporiaceae, Pleosporales, Dothideomycetes) from freshwater habitats in Egypt. Mycosphere 2016, 7, 448–457. [Google Scholar] [CrossRef]
- Santos da Silva, S.; Castañeda Ruiz, R.F.; Pascholati Gusmão, L.F. New species and records of Dictyosporium on Araucaria angustifolia (Brazilian pine) from Brazil. Nova Hedwig 2016, 102, 523–530. [Google Scholar] [CrossRef]
- Alves-Barbosa, M.; Costa, P.M.O.; Malosso, E.; Castañeda-Ruiz, R.F. Two new species of Dictyosporium and Helminthosporium (Ascomycota) from the Brazilian Atlantic Forest. Nova Hedwig 2017, 105, 65–73. [Google Scholar] [CrossRef]
- Zhang, Y.; Cai, C.S.; Zhao, G.Z. Dictyosporium wuyiense sp. nov. from Wuyi Mountain China. Phytotaxa 2017, 314, 251–258. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Thilini Chethana, K.W.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.-Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; et al. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild Seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Gareth Jones, E.B.; Jayarama Bhat, D.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Wang, R.-X.; Luo, Z.-L.; Hyde, K.D.; Bhat, D.J.; Su, X.-J.; Su, H.-Y. New species and records of Dictyocheirospora from submerged wood in north-western Yunnan, China. Mycosphere 2016, 7, 1357–1367. [Google Scholar] [CrossRef]
- Ho, W.-H.; Hyde, K.D.; Hodgkiss, I.J. Digitodesmium recurvum, a new species of chirosporous hyphomycete from Hong Kong. Mycologia 1999, 91, 900–904. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Zhang, K.; Mc Kenzie, E.H.C.H.; Hyde, K.D. Acrodictys liputii sp. nov. and Digitodesmium bambusicola sp. nov. from bamboo submerged in Liput River in the Philippines. Nova Hedwig 2002, 75, 525–532. [Google Scholar] [CrossRef]
- Silvera-Simón, C.; Mercado-Sierra, A.; Mena-Portales, J.; Gene, J.; Guarro, J. Two new species of Digitodesmium from Euskadi. Cryptogam. Mycol. 2010, 31, 211–218. [Google Scholar]
- Nobrega, T.F.; Ferreira, B.W.; Barreto, R.W. Digitodesmium polybrachiatum sp. nov., a new species of Dictyosporiaceae from Brazil. Mycol. Prog. 2021, 20, 1135–1144. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, K.; McKenzie, E.H.C.; Hyde, K.D. New species of Dictyosporium and Digitodesmium from submerged wood in Yannan, China. Sydowia 2003, 55, 129–135. [Google Scholar]
- McKenzie, E.H.C. Two new dictyosporous hyphomycetes on Pandanaceae. Mycotaxon 2008, 104, 23–28. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs 2; Matsushima Fungus Collection: Kobe, Japan, 1981; pp. 1–68. [Google Scholar]
- Sutton, B.C. Notes on some deuteromycete genera with cheiroid or digitate brown conidia. Proc. Indian Acad. Sci. Sect. B 1985, 94, 229–244. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Feng, B.; Yang, Z.L. Studies on diversity of higher fungi in yunnan, southwestern China: A review. Plant Divers. 2018, 40, 165–171. [Google Scholar] [CrossRef]
- Chaiwan, N.; Gomdola, D.; Wang, S.; Monkai, J.; Tibpromma, S.; Doilom, M.; Wanasinghe, D.N.; Mortimer, P.E.; Lumyong, S.; Hyde, K.D. https://gmsmicrofungi.org: An online database providing updated information of microfungi in the Greater Mekong Subregion. Mycosphere 2021, 12, 1513–1526. [Google Scholar] [CrossRef]
- Monkai, J.; Wanasinghe, D.N.; Jeewon, R.; Promputtha, I.; Phookamsak, R. Morphological and phylogenetic characterization of fungi within Bambusicolaceae: Introducing two new species from the Greater Mekong Subregion. Mycol. Prog. 2021, 20, 721–732. [Google Scholar] [CrossRef]
- Mortimer, P.E.; Jeewon, R.; Xu, J.C.; Lumyong, S.; Wanasinghe, D.N. Morpho-Phylo Taxonomy of novel dothideomycetous fungi associated with dead woody twigs in Yunnan Province, China. Front. Microbiol. 2021, 12, 654683. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Fryar, S.; Tian, Q.; Bahkali, A.H.; Xu, J. Lignicolous freshwater fungi along a north-south latitudinal gradient in the Asian/Australian region; can we predict the impact of global warming on biodiversity and function? Fungal Ecol. 2016, 19, 190–200. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Li, W.L.; Su, H.Y. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Indunil Chinthani, S.; Achala, R.R.; Diana, S.M.; Mark, S.C.; Eleni, G.; Hyang Burm, L.; Vedprakash, G.H.; Dhandevi, P.; Lakmali, S.D.; Subodini, N.W.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.-C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.; Liu, J.-K.J. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2652–2676. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 1990, 18, 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest 2.0; Program Distributed by the Author; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Photita, W.; Lumyong, P.; McKenzie, E.H.C.; Hyde, K.D.; Lumyong, S. A new Dictyosporium species from musa acuminata in Thailand. Mycotaxon 2002, 82, 415–419. [Google Scholar]
- Rajeshkumar, K.C.; Verma, R.K.; Boonmee, S.; Chandrasiri, S.; Hyde, K.D.; Ashtekar, N.I.; Lad, S.; Wijayawardene, N.N. Paradictyocheirospora tectonae, a novel genus in the family Dictyosporiaceae from India. Phytotaxa 2021, 509, 257–271. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; Mckenzie, E.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
Figure 1.
The maximum likelihood (ML) tree is based on the combined SSU, ITS, LSU and tef 1-α sequence data. Bootstrap support values with an ML greater than 75% and Bayesian posterior probabilities (PP) greater than 0.97 are given above the nodes, shown as “ML/PP”. The tree was rooted to Periconia igniaria (CBS 379.86 and CBS 845.96). New species are indicated in blue and type strains are in bold.
Figure 1.
The maximum likelihood (ML) tree is based on the combined SSU, ITS, LSU and tef 1-α sequence data. Bootstrap support values with an ML greater than 75% and Bayesian posterior probabilities (PP) greater than 0.97 are given above the nodes, shown as “ML/PP”. The tree was rooted to Periconia igniaria (CBS 379.86 and CBS 845.96). New species are indicated in blue and type strains are in bold.


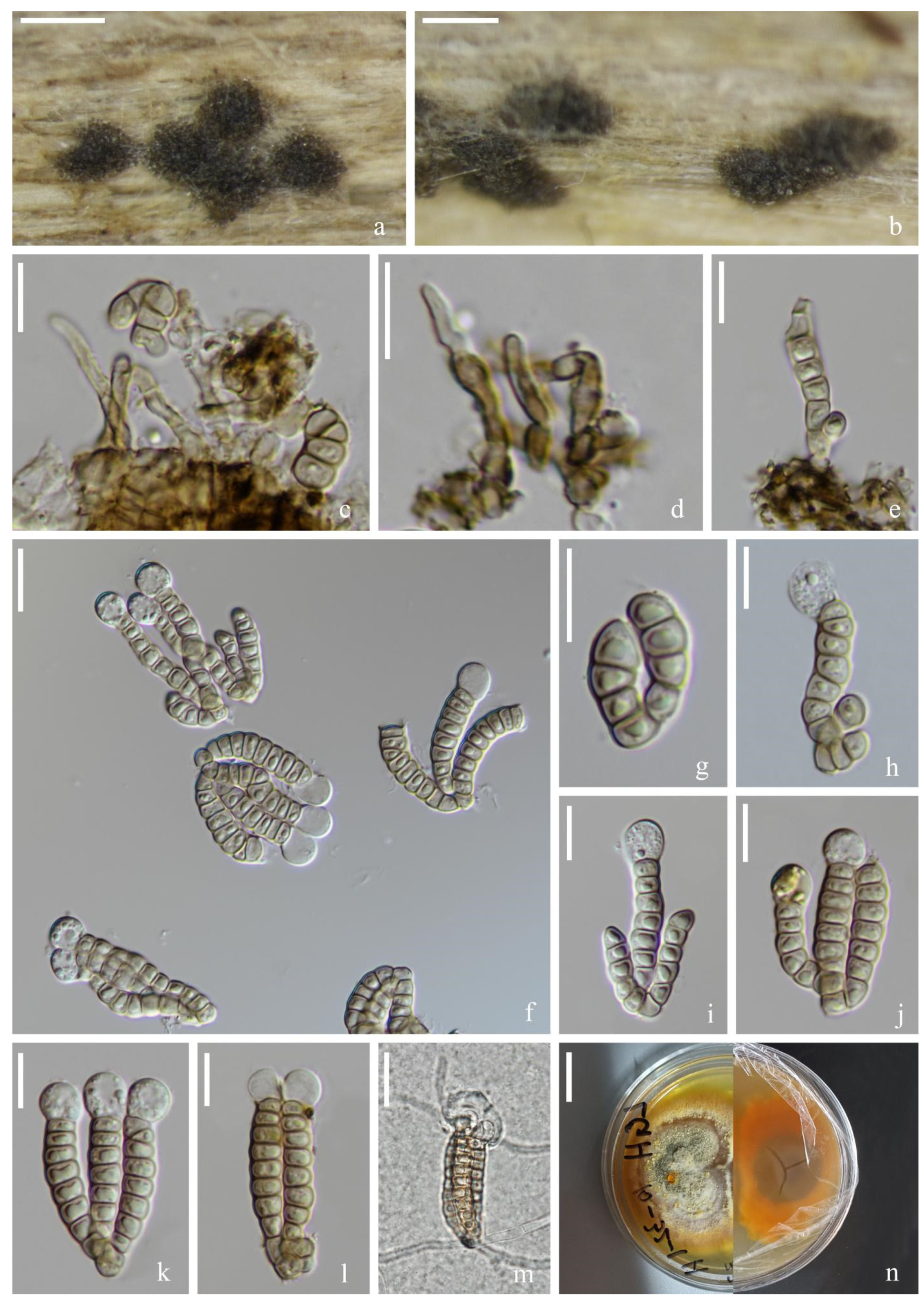
Figure 2.
Dictyocheirospora chiangmaiensis (MFLU 22–0199, holotype). (a,b) Colonies on the substratum. (c) Conidiophores. (d) Conidiomata with conidiophores. (e–g) Conidiogenous cells with conidia. (h) Conidia. (i) Germinating conidium. (j,k) Culture colonies on MEA, reverse (left) and obverse (right). Scale bars: (a) 200 μm, (b) 100 μm, (c,e–i) 10 μm, (d) 20 μm and (j,k) 1 cm.
Figure 2.
Dictyocheirospora chiangmaiensis (MFLU 22–0199, holotype). (a,b) Colonies on the substratum. (c) Conidiophores. (d) Conidiomata with conidiophores. (e–g) Conidiogenous cells with conidia. (h) Conidia. (i) Germinating conidium. (j,k) Culture colonies on MEA, reverse (left) and obverse (right). Scale bars: (a) 200 μm, (b) 100 μm, (c,e–i) 10 μm, (d) 20 μm and (j,k) 1 cm.

Figure 3.
Dictyocheirospora suae (KUN-HKAS 121703, holotype). (a,b) Colonies on the substratum. (c) Conidiomata. (d,e) Conidiophores and conidiogenous cells. (f) Conidia. (g) Conidiogenous cells with conidia. (h) Conidia with appendages. (i) Germinating conidium. (j) Culture colonies on PDA, reverse (upper) and obverse (lower). Scale bars: (a) 200 μm, (b) 100 μm, (c) 30 μm, (d–i) 20 μm and (j) 2 cm.
Figure 3.
Dictyocheirospora suae (KUN-HKAS 121703, holotype). (a,b) Colonies on the substratum. (c) Conidiomata. (d,e) Conidiophores and conidiogenous cells. (f) Conidia. (g) Conidiogenous cells with conidia. (h) Conidia with appendages. (i) Germinating conidium. (j) Culture colonies on PDA, reverse (upper) and obverse (lower). Scale bars: (a) 200 μm, (b) 100 μm, (c) 30 μm, (d–i) 20 μm and (j) 2 cm.

Figure 4.
Dictyocheirospora multiappendiculata (KUN-HKAS 122866, holotype). (a–c) Colonies on the substratum. (d) Conidiomata and conidia. (e) Conidiophores and conidiogenous cells. (f–h) Conidiogenous cells, conidia and appendages. (i,j) Conidia with appendages. (k) Germinating conidium. (l,m) Culture colonies on PDA, reverse (upper) and obverse (lower). Scale bars: (a) 2000 μm, (b) 200 μm, (c) 100 μm, (d) 30 μm, (e–k) 20 μm and (l,m) 1 cm.
Figure 4.
Dictyocheirospora multiappendiculata (KUN-HKAS 122866, holotype). (a–c) Colonies on the substratum. (d) Conidiomata and conidia. (e) Conidiophores and conidiogenous cells. (f–h) Conidiogenous cells, conidia and appendages. (i,j) Conidia with appendages. (k) Germinating conidium. (l,m) Culture colonies on PDA, reverse (upper) and obverse (lower). Scale bars: (a) 2000 μm, (b) 200 μm, (c) 100 μm, (d) 30 μm, (e–k) 20 μm and (l,m) 1 cm.

Figure 5.
Dictyocheirospora aquadulcis (MFLU 22-0201). (a) Colonies on the substratum. (b) Conidiomata. (c) Conidiophores. (d,e) Conidiophores and conidia. (f) Conidiophores, conidia and appendages. (g–i) Conidiogenous cells and conidia. (j) Conidia. (k) Culture colonies on PDA, reverse (left) and obverse (right). Scale bars: (a) 100 μm, (b–j) 20 μm and (k) 2 cm.
Figure 5.
Dictyocheirospora aquadulcis (MFLU 22-0201). (a) Colonies on the substratum. (b) Conidiomata. (c) Conidiophores. (d,e) Conidiophores and conidia. (f) Conidiophores, conidia and appendages. (g–i) Conidiogenous cells and conidia. (j) Conidia. (k) Culture colonies on PDA, reverse (left) and obverse (right). Scale bars: (a) 100 μm, (b–j) 20 μm and (k) 2 cm.

Figure 6.
Dictyocheirospora nabanheensis (MFLU 22-0200). (a,b) Colonies on the substratum. (c,d) Conidiophores. (e) Conidia heap. (f,g) Conidiogenous cells, conidia and appendages. (h) Conidia. (i,j) Conidia with appendages. (k) Germinating conidium. (l) Culture colonies on PDA, reverse (left) and obverse (right). Scale bars: (a) 100 μm, (b) 200 μm, (c,d,g–j) 10 μm, (e,f,k) 20 μm and (l) 2 cm.
Figure 6.
Dictyocheirospora nabanheensis (MFLU 22-0200). (a,b) Colonies on the substratum. (c,d) Conidiophores. (e) Conidia heap. (f,g) Conidiogenous cells, conidia and appendages. (h) Conidia. (i,j) Conidia with appendages. (k) Germinating conidium. (l) Culture colonies on PDA, reverse (left) and obverse (right). Scale bars: (a) 100 μm, (b) 200 μm, (c,d,g–j) 10 μm, (e,f,k) 20 μm and (l) 2 cm.

Figure 7.
Dictyocheirospora heptaspora (MFLU 22-0202). (a) Colonies on the substratum. (c–f) Conidiogenous cells with conidia. (b,g,h) Conidia. Scale bars: (a) 200 μm, (b) 40 μm and (c–h) 20 μm.
Figure 7.
Dictyocheirospora heptaspora (MFLU 22-0202). (a) Colonies on the substratum. (c–f) Conidiogenous cells with conidia. (b,g,h) Conidia. Scale bars: (a) 200 μm, (b) 40 μm and (c–h) 20 μm.

Figure 8.
Dictyosprium tubulatum (KUN-HKAS 115789, new geographical record). (a) Colonies on the substratum. (b–d) Conidiophores and conidia. (e) Conidia heap. (f,i,m) Conidiogenous cells, conidia and appendages. (g,h) Conidiogenous cells with conidia. (j–l) Conidia with appendages. Scale bars: (a) 200 μm, (b–e) 20 μm and (f–m) 10 μm.
Figure 8.
Dictyosprium tubulatum (KUN-HKAS 115789, new geographical record). (a) Colonies on the substratum. (b–d) Conidiophores and conidia. (e) Conidia heap. (f,i,m) Conidiogenous cells, conidia and appendages. (g,h) Conidiogenous cells with conidia. (j–l) Conidia with appendages. Scale bars: (a) 200 μm, (b–e) 20 μm and (f–m) 10 μm.

Figure 9.
Digitodesmium aquaticum (MFLU 22-0203, holotype). (a,b) Colonies on the substratum. (c–f) Conidiophores with conidia. (g–j) Conidia. Scale bars: (a) 1000 μm, (b) 100 μm, (c) 40 μm and (d–j) 20 μm.
Figure 9.
Digitodesmium aquaticum (MFLU 22-0203, holotype). (a,b) Colonies on the substratum. (c–f) Conidiophores with conidia. (g–j) Conidia. Scale bars: (a) 1000 μm, (b) 100 μm, (c) 40 μm and (d–j) 20 μm.

Figure 10.
Vikalpa grandispora (KUN-HKAS 122867, holotype). (a,b) Colonies on the substratum. (c) Conidiomata. (d) Conidiogenous cells. (e,g–i) Conidia with appendages. (f) Conidia. (j) Germinating conidium. (k) Culture colonies on PDA, reverse (upper) and obverse (lower). Scale bars: (a) 1000 μm, (b) 200 μm, (c–j) 10 μm and (k) 1 cm.
Figure 10.
Vikalpa grandispora (KUN-HKAS 122867, holotype). (a,b) Colonies on the substratum. (c) Conidiomata. (d) Conidiogenous cells. (e,g–i) Conidia with appendages. (f) Conidia. (j) Germinating conidium. (k) Culture colonies on PDA, reverse (upper) and obverse (lower). Scale bars: (a) 1000 μm, (b) 200 μm, (c–j) 10 μm and (k) 1 cm.

Figure 11.
Vikalpa sphaerica (KUN-HKAS 115805, holotype). (a,b) Colonies on the substratum. (c,d) Conidiophores and conidiogenous cells. (e) Conidia. (f–l) Conidia with appendages. (m) Germinating conidium. (n) Culture colonies on PDA, reverse (left) and obverse (right). Scale bars: (a,b) 200 μm, (c–m) 10 μm and (n) 1 cm.
Figure 11.
Vikalpa sphaerica (KUN-HKAS 115805, holotype). (a,b) Colonies on the substratum. (c,d) Conidiophores and conidiogenous cells. (e) Conidia. (f–l) Conidia with appendages. (m) Germinating conidium. (n) Culture colonies on PDA, reverse (left) and obverse (right). Scale bars: (a,b) 200 μm, (c–m) 10 μm and (n) 1 cm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains/specimens used for phylogenetic analysis and their GenBank accession numbers.
| Species 1 | Source 2 | GenBank Accession Number 3 | |||
|---|---|---|---|---|---|
| SSU | ITS | LSU | tef 1-α | ||
| Aquadictyospora clematidis | MFLUCC 17-2080 T | MT226664 | MT310592 | MT214545 | MT394727 |
| Aquadictyospora lignicola | MFLUCC 17-1318 T | – | MF948621 | MF948629 | MF953164 |
| Aquaticheirospora lignicola | HKUCC 10304 T | AY736377 | AY864770 | AY736378 | – |
| Cheirosporium triseriale | HMAS 180703 T | – | EU413953 | EU413954 | – |
| Dendryphiella phitsanulokensis | MFLUCC 17-2513 T | MG754402 | MG754400 | MG754401 | – |
| Dendryphiella variabilis | CBS 584.96 T | – | LT963453 | LT963454 | – |
| Dendryphiella eucalyptorum | CBS 137987 T | – | KJ869139 | KJ869196 | – |
| Dendryphiella fasciculata | MFLUCC 17-1074 T | – | MF399213 | MF399214 | – |
| Dendryphiella paravinosa | CBS 141286 T | – | KX228257 | KX228309 | – |
| Dictyocheirospora aquadulcis | MFLUCC 17-2571 T | – | MK634545 | MK634542 | – |
| Dictyocheirospora aquadulcis * | MFLUCC 22-0095 | OP526625 | OP526634 | OP526644 | OP542236 |
| Dictyocheirospora aquatica | KUMCC 15-0305 T | – | KY320508 | KY320513 | – |
| Dictyocheirospora bannica | KH 332 T | AB797223 | LC014543 | AB807513 | AB808489 |
| Dictyocheirospora bannica | MFLUCC 16-0874 | – | MH381765 | MH381774 | – |
| Dictyocheirospora cheirospora | KUMCC 17-0035 T | MF928073 | MF177035 | MF177036 | – |
| Dictyocheirospora clematidis | MFLUCC 17-2089 T | MT226665 | MT310593 | MT214546 | MT394728 |
| Dictyocheirospora chiangmaiensis * | MFLUCC 22-0097 T | OP526621 | OP526630 | OP526640 | OP542232 |
| Dictyocheirospora garethjonesii | MFLUCC 16-0909 T | – | KY320509 | KY320514 | – |
| Dictyocheirospora garethjonesii | DLUCC 0848 | – | MF948623 | MF948631 | MF953166 |
| Dictyocheirospora gigantica | BCC 11346 | – | DQ018095 | – | – |
| Dictyocheirospora heptaspora | CBS 396.59 | DQ018082 | DQ018090 | – | – |
| Dictyocheirospora heptaspora | DLUCC 1992 | – | MT756244 | MT756243 | MT776563 |
| Dictyocheirospora heptaspora * | MFLUCC 22-0096 | – | OP526635 | OP526645 | OP542237 |
| Dictyocheirospora indica | MFLUCC 15-0056 | MH381757 | MH381763 | MH381772 | MH388817 |
| Dictyocheirospora lithocarpi | MFLUCC 17-2537 T | MK347888 | MK347781 | MK347999 | – |
| Dictyocheirospora metroxylonis | MFLUCC 15-0028a T | MH742317 | MH742321 | MH742313 | – |
| Dictyocheirospora metroxylonis | MFLUCC 15-0028b T | MH742318 | MH742322 | MH742314 | MH764301 |
| Dictyocheirospora multiappendiculata * | KUNCC 22-10734 T | OP526623 | OP526632 | OP526642 | OP542234 |
| Dictyocheirospora multiappendiculata * | KUNCC 22-10736 | OP526624 | OP526633 | OP526643 | OP542235 |
| Dictyocheirospora nabanheensis | MFLUCC 17-0562 T | – | MH388340 | MH376712 | MH388375 |
| Dictyocheirospora nabanheensis * | MFLUCC 22-0094 | OP526627 | OP526637 | OP526647 | OP542239 |
| Dictyocheirospora pandanicola | MFLUCC 16-0365 T | MH388309 | MH388341 | MH376713 | MH388376 |
| Dictyocheirospora pseudomusae | yone 234 T | AB797230 | LC014550 | AB807520 | AB808496 |
| Dictyocheirospora rotunda | MFLUCC 14-0293 T | KU179101 | KU179099 | KU179100 | – |
| Dictyocheirospora rotunda | MFLUCC 17-0222 | – | MH381764 | MH381773 | MH388818 |
| Dictyocheirospora suae * | KUNCC 22-12424 T | OP526622 | OP526631 | OP526641 | OP542233 |
| Dictyocheirospora subramanianii | BCC 3503 | – | DQ018094 | – | – |
| Dictyocheirospora taiwanense | MFLUCC 17-2654 T | – | MK495821 | MK495820 | – |
| Dictyocheirospora thailandica | MFLUCC 18-0987 T | – | MT627734 | MN913743 | – |
| Dictyocheirospora vinaya | MFLUCC 14-0294 T | KU179104 | KU179102 | KU179103 | – |
| Dictyocheirospora xishuangbannaensis | KUMCC 17-0181 T | MH388310 | MH388342 | MH376714 | MH388377 |
| Dictyocheirospora xishuangbannaensis | MFLUCC 17-2087 | MT226666 | MT310594 | MT214547 | MT394729 |
| Dictyosporium appendiculatum | KUMCC 17-0311 T | – | MH388343 | MH376715 | – |
| Dictyosporium hongkongensis | KMUCC 17-0268 T | MH388313 | MH388346 | MH376718 | MH388380 |
| Dictyosporium krabiense | MFLU 16-1890 T | MH388314 | – | MH376719 | MH388381 |
| Dictyosporium marinum | GJ357 T | – | – | MN017841 | – |
| Dictyosporium muriformis | GZCC 20-0006 T | MN901117 | MT002304 | MN897834 | MT023011 |
| Dictyosporium pandanicola | MFLU 16-1886 T | – | MH388347 | MH376720 | MH388382 |
| Dictyosporium alatum | ATCC 34953 T | DQ018080 | DQ018088 | DQ018101 | – |
| Dictyosporium aquaticum | MF 1318 T | – | KM610236 | – | – |
| Dictyosporium bulbosum | yone 221 | AB797221 | LC014544 | AB807511 | AB808487 |
| Dictyosporium digitatum | KH 401 | AB797225 | LC014545 | AB807515 | AB808491 |
| Dictyosporium digitatum | yone 280 | AB797228 | LC014547 | AB807512 | AB808488 |
| Dictyosporium elegans | NBRC 32502 T | DQ018079 | DQ018087 | DQ018100 | – |
| Dictyosporium guttulatum | MFLUCC 16-0258 T | MH388312 | MH388345 | MH376717 | MH388379 |
| Dictyosporium hughesii | KT 1847 | AB797227 | LC014548 | AB807517 | AB808493 |
| Dictyosporium meiosporum | MFLUCC 10-0131 T | KP710946 | KP710944 | KP710945 | – |
| Dictyosporium nigroapice | BCC 3555 | – | DQ018085 | – | – |
| Dictyosporium nigroapice | MFLUCC 17-2053 T | – | MH381768 | MH381777 | MH388821 |
| Dictyosporium olivaceosporum | KH 375 T | AB797224 | LC014542 | AB807514 | AB808490 |
| Dictyosporium sexualis | MFLUCC 10-0127 T | KU179107 | KU179105 | KU179106 | – |
| Dictyosporium sp. | MFLUCC 15-0629 | – | MH381766 | MH381775 | MH388819 |
| Dictyosporium stellatum | CCFC 241241 T | – | NR_154608 | JF951177 | – |
| Dictyosporium strelitziae | CBS 123359 T | – | NR_156216 | FJ839653 | – |
| Dictyosporium tetrasporum | KT 2865 | AB797229 | LC014551 | AB807519 | AB808495 |
| Dictyosporium thailandicum | MFLUCC 13-0773 T | – | KP716706 | KP716707 | – |
| Dictyosporium tratense | MFLUCC 17-2052 T | MH381761 | MH381767 | MH381776 | MH388820 |
| Dictyosporium tubulatum | MFLUCC 15-0631 T | – | MH381769 | MH381778 | MH388822 |
| Dictyosporium tubulatum | MFLUCC 17-2056 | – | MH381770 | MH381779 | – |
| Dictyosporium tubulatum * | KUN-HKAS 115789 | OP749878 | OP749871 | OP749876 | OP756063 |
| Dictyosporium wuyiense | CGMCC 3.18703 T | – | KY072977 | – | – |
| Dictyosporium zhejiangense | MW-2009a T | – | FJ456893 | – | – |
| Digitodesmium aquaticum * | MFLU 22-0203 T | OP749879 | OP749872 | OP749877 | OP756064 |
| Digitodesmium bambusicola | CBS 110279 T | – | DQ018091 | DQ018103 | – |
| Digitodesmium chiangmaiense | KUN-HKAS 102163 T | MK571775 | – | MK571766 | – |
| Digitodesmium polybrachiatum | COAD 3174 T | MW879325 | MW879318 | MW879316 | – |
| Digitodesmium polybrachiatum | COAD 3175 | MW879326 | MW879319 | MW879317 | – |
| Digitodesmium sp. | TBRC 10037 | – | MK405234 | MK405232 | MK405230 |
| Digitodesmium sp. | TBRC 10038 | – | MK405235 | MK405233 | MK405231 |
| Gregarithecium curvisporum | KT 922 | AB797257 | AB809644 | AB807547 | – |
| Gregarithecium curvisporum | MFLUCC 13-0853 T | KX364283 | KX364281 | KX364282 | – |
| Immotthia atrograna | ZT-Myc-64283 | – | MW489540 | – | – |
| Immotthia bambusae | KUN-HKAS 112012A1 T | MW489461 | MW489455 | MW489450 | MW504646 |
| Immotthia bambusae | KUN-HKAS 112012B T | – | MW489457 | MW489452 | – |
| Immotthia bambusae | KUN-HKAS 112012C T | MW489463 | MW489458 | MW489453 | MW504648 |
| Immotthia bambusae | KUN-HKAS 112012D T | MW489464 | MW489459 | MW489454 | MW504649 |
| Jalapriya inflata | NOU 3855 | JQ267361 | JQ267362 | JQ267363 | – |
| Jalapriya pulchra | MFLUCC 15-0348 T | KU179110 | KU179108 | KU179109 | – |
| Jalapriya pulchra | MFLUCC 17-1683 | – | MF948628 | MF948636 | MF953171 |
| Jalapriya toruloides | CBS 209.65 | DQ018081 | DQ018093 | DQ018104 | – |
| Neodendryphiella mali | FMR 16561 T | – | LT906655 | LT906657 | – |
| Neodendryphiella mali | FMR 17003 | – | LT993734 | LT993735 | – |
| Neodendryphiella michoacanensis | FMR 16098 T | – | LT906660 | LT906658 | – |
| Neodendryphiella tarraconensis | FMR 16234 T | – | LT906659 | LT906656 | – |
| Neodendryphiella tarraconensis | GZCC20_0002 | – | MN999922 | MN999927 | – |
| Periconia igniaria | CBS 379.86 | – | LC014585 | AB807566 | AB808542 |
| Periconia igniaria | CBS 845.96 | – | LC014586 | AB807567 | AB808543 |
| Pseudocoleophoma bauhiniae | MFLUCC 17-2586 T | MK347844 | MK347736 | MK347953 | MK360076 |
| Pseudocoleophoma bauhiniae | MFLUCC 17-2280 | MK347843 | MK347735 | MK347952 | MK360075 |
| Pseudocoleophoma flavescen | CBS_178.93 | GU238216 | – | GU238075 | – |
| Pseudocoleophoma rusci | MFLUCC 16-1444 T | MT214983 | MT185549 | MT183514 | – |
| Pseudocoleophoma zingiberacearum | NCYUCC 19-0052 T | – | MN615939 | MN616753 | MN629281 |
| Pseudocoleophoma zingiberacearum | NCYUCC 19-0053 | – | MN615940 | MN616754 | MN629282 |
| Pseudocoleophoma calamagrostidis | KT 3284 T | LC014604 | LC014592 | LC014609 | LC014614 |
| Pseudocoleophoma polygonicola | KT 731 T | AB797256 | AB809634 | AB807546 | AB808522 |
| Pseudocoleophoma typhicola | MFLUCC 16-0123 T | – | KX576655 | KX576656 | – |
| Pseudoconiothyrium broussonetiae | CBS 145036 T | – | MK442618 | MK442554 | – |
| Pseudocyclothyriella clematidis | MFLUCC 17-2177 T | – | MT310596 | MT214549 | MT394730 |
| Pseudocyclothyriella clematidis | MFLUCC 17-2177A T | MT226667 | MT310595 | MT214548 | – |
| Pseudodictyosporium elegans | CBS 688.93 T | DQ018084 | DQ018099 | DQ018106 | – |
| Pseudodictyosporium indicum | CBS 471.95 | – | DQ018097 | – | – |
| Pseudodictyosporium thailandica | MFLUCC 16-0029 T | KX259524 | KX259520 | KX259522 | KX259526 |
| Pseudodictyosporium wauense | NBRC 30078 T | DQ018083 | DQ018098 | DQ018105 | – |
| Pseudodictyosporium wauense | DLUCC 0801 | – | MF948622 | MF948630 | MF953165 |
| Verrucoccum coppinsii | E00814291 conidioma T | MT918778 | MT918784 | MT918770 | – |
| Verrucoccum coppinsii | E00814291 ascoma T | MT918777 | MT918785 | MT918769 | – |
| Verrucoccum spribillei | SPO2343 T | MT918773 | MT918780 | MT918765 | – |
| Verrucoccum spribillei | SPO1154 T | MT918772 | MT918781 | MT918764 | – |
| Vikalpa australiensis | HKUCC 8797 | – | DQ018092 | – | – |
| Vikalpa grandispora * | KUNCC 22-12425 T | OP526628 | OP526638 | OP526648 | OP542240 |
| Vikalpa sphaerica * | CGMCC3.20682 T | OP526629 | OP526639 | OP526649 | OP542241 |
1 The newly generated sequences show “*” after the species name; 2 type specimens/ex-type strains show “T” after the number; 3 missing sequences are indicated with “–”.
Table 2.
Comparison of conidia characteristics and habitats of Dictyocheirospora, Digitodesmium and Vikalpa species.
Table 2.
Comparison of conidia characteristics and habitats of Dictyocheirospora, Digitodesmium and Vikalpa species.
| Species | Conidia Size (μm) | Conidia Septate | No. of Rows | Appendages | Habitat | References |
|---|---|---|---|---|---|---|
| D. aquadulcis | 60–80 × 17–29 | Euseptate | 7 | No appendages | Freshwater | [35] |
| D. aquatica | 34–42 × 12.5–19.5 | Euseptate | 8–10 | No appendages | Freshwater | [40] |
| D. bannica | 73–86(–90) × 21–26(–31) | - | (5–)–7 | No appendages | Terrestrial | [5] |
| D. cheirospora | 54–63 × 15–26 | - | 5–7 | No appendages | Terrestrial | [34] |
| D. clematidis | 42–60 × 15–30 | Distosepta | 6–7 | No appendages | Terrestrial | [37] |
| D. chiangmaiensis | (40–)42–46 × 16–18 | Euseptate | 4–6 | No appendages | Freshwater | This study |
| D. suae | (65–)72–79 × (–17)20–25(–29) | Euseptate | Appendages | Freshwater | This study | |
| D. garethjonesii | 45.5–54.5 × 15.5–24.5 | Euseptate | 6–7 | No appendages | Freshwater | [40] |
| D. gigantica | 105–121 × 25–32 | - | 7 | No appendages | Freshwater | [22] |
| D. heptaspora | 50–80 × 20–30 | - | 7 | No appendages | Terrestrial | [22] |
| D. hydei | (26–)30–33(–35) × 14–17 | - | 7 | Supra-basal appendages | Terrestrial | [26] |
| D. indica | (33–)36–46(–48) × 13–18 | - | 6–7 | Subapical appendages | Terrestrial | [26] |
| D. lithocarpi | 35–40 × 12–18 | Euseptate | 6 | No appendages | Terrestrial | [36] |
| D. metroxylonis | 45–69 × 15–29 | Distoseptate | 4–6 | No appendages | Terrestrial | [37] |
| D. multiappendiculata | (65–)72–79 × (–17)20–25(–29) | Euseptate | 5–7 | Subglobose appendages | Freshwater | This study |
| D. musae | 45–65 × 20–27 | - | 7 | Appendages | Terrestrial | [73] |
| D. nabanheensis | 35–40 × 18–21 | - | 6 | Appendages | Terrestrial | [9] |
| D. pandanicola | 60–75 × 18.5–35.5 | - | 5–7 | No appendages | Terrestrial | [9] |
| D. pseudomusae | (58–)61–78(–81) × 19–29(–33) | - | (6–)7 | Globose to subglobose appendages | Terrestrial | [19] |
| D. rotunda | 42–58 × 19–38 | Distoseptate | 5–7 | No appendages | Freshwater | [5] |
| D. subramanianii | 33–42 × 16–20 | - | 7 | - | Terrestrial | [48] |
| D. taiwanense | (72–)74–84(–86) × 16–20(–24) | - | 5 | No appendages | Terrestrial | [35] |
| D. tetraploides | 52.5–72.5 × 18.5– 26.5 | Euseptate | 5 | Subapical appendages | Freshwater | [45] |
| D. thailandica | 42–65 × 20–45 | - | 6–7 | No appendages | Freshwater | [38] |
| D. vinaya | 58–67 × 15.5–26.5 | Distoseptate | 6–7 | No appendages | Freshwater | [5] |
| D. xishuangbannaensis | 35–50 × 17–25 | - | 6 | No appendages | Terrestrial | [9] |
| Digitodesmium aquaticum | (39–)41–44(–46) × (17–)19–21(–22) | Euseptate | 3 | No appendages | Freshwater | This study |
| Di. bambusicola | 24–29–32.5 × 12.5–17–23 | Euseptate | 3 | Appendages | Freshwater | [42] |
| Di. chiangmaiense | 33–42 × 15–18 | Euseptate | 3 | No appendages | Terrestrial | [35] |
| Di. elegans | 45–60 × 12–21 | Euseptate | (2–)3–4(–6) | No appendages | Terrestrial | [33] |
| Di. heptasporum | 50–75 × 32.5–70 | Euseptate | (6)–7 | No appendages | Freshwater | [45] |
| Di. intermedium | 39–76 × 25–35 | Euseptate | 3–11 | No appendages | Terrestrial | [43] |
| Di. macrosporum | 130–145 × 19–26 | Euseptate | 5–8 | No appendages | Terrestrial | [43] |
| Di. polybrachiatum | 35–54 × 15–19 | Euseptate | 6–9 | Appendages | Terrestrial | [44] |
| Di. recurvum | 30–45 × 12–21 | Euseptate | 4–7 | No appendages | Freshwater | [41] |
| Di. tectonae | 28.5–41 × 14–17 | - | 3–6 | Subglobose appendages | Terrestrial | [74] |
| Vikalpa grandispora | (33–)38–49(–53) × (9–)13–18(–21) | Euseptate | 3 | Subglobose appendages | Freshwater | This study |
| V. australiensis | 36–43 × 11.5–12 | Euseptate | 3 | - | Terrestrial | [48] |
| V. freycinetiae | (27–)31–43 × 10– 20 | Euseptate | 3 | Subglobose appendages | Terrestrial | [46] |
| V. lignicola | 25–40.5 × 10–18 | Distoseptate | 3 | No appendages | Freshwater | [5] |
| V. micronesiaca | 20–30 × 10–12 | - | 2–4 | No appendages | Terrestrial | [47] |
| V. sphaerica | (23–)26–30(–34) × (11–)16–19 | Euseptate | 3–4 | Subglobose appendages | Freshwater | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shen, H.; Bao, D.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.; Luo, Z. Novel Species and Records of Dictyosporiaceae from Freshwater Habitats in China and Thailand. J. Fungi 2022, 8, 1200. https://doi.org/10.3390/jof8111200
AMA Style
Shen H, Bao D, Wanasinghe DN, Boonmee S, Liu J, Luo Z. Novel Species and Records of Dictyosporiaceae from Freshwater Habitats in China and Thailand. Journal of Fungi. 2022; 8(11):1200. https://doi.org/10.3390/jof8111200
Chicago/Turabian StyleShen, Hongwei, Danfeng Bao, Dhanushka N. Wanasinghe, Saranyaphat Boonmee, Jiankui Liu, and Zonglong Luo. 2022. "Novel Species and Records of Dictyosporiaceae from Freshwater Habitats in China and Thailand" Journal of Fungi 8, no. 11: 1200. https://doi.org/10.3390/jof8111200
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.