



A Comprehensive Account of the Rust Genus Skierka (Skierkaceae)
 , ,
, ,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. The Genus Skierka
2.1. General History
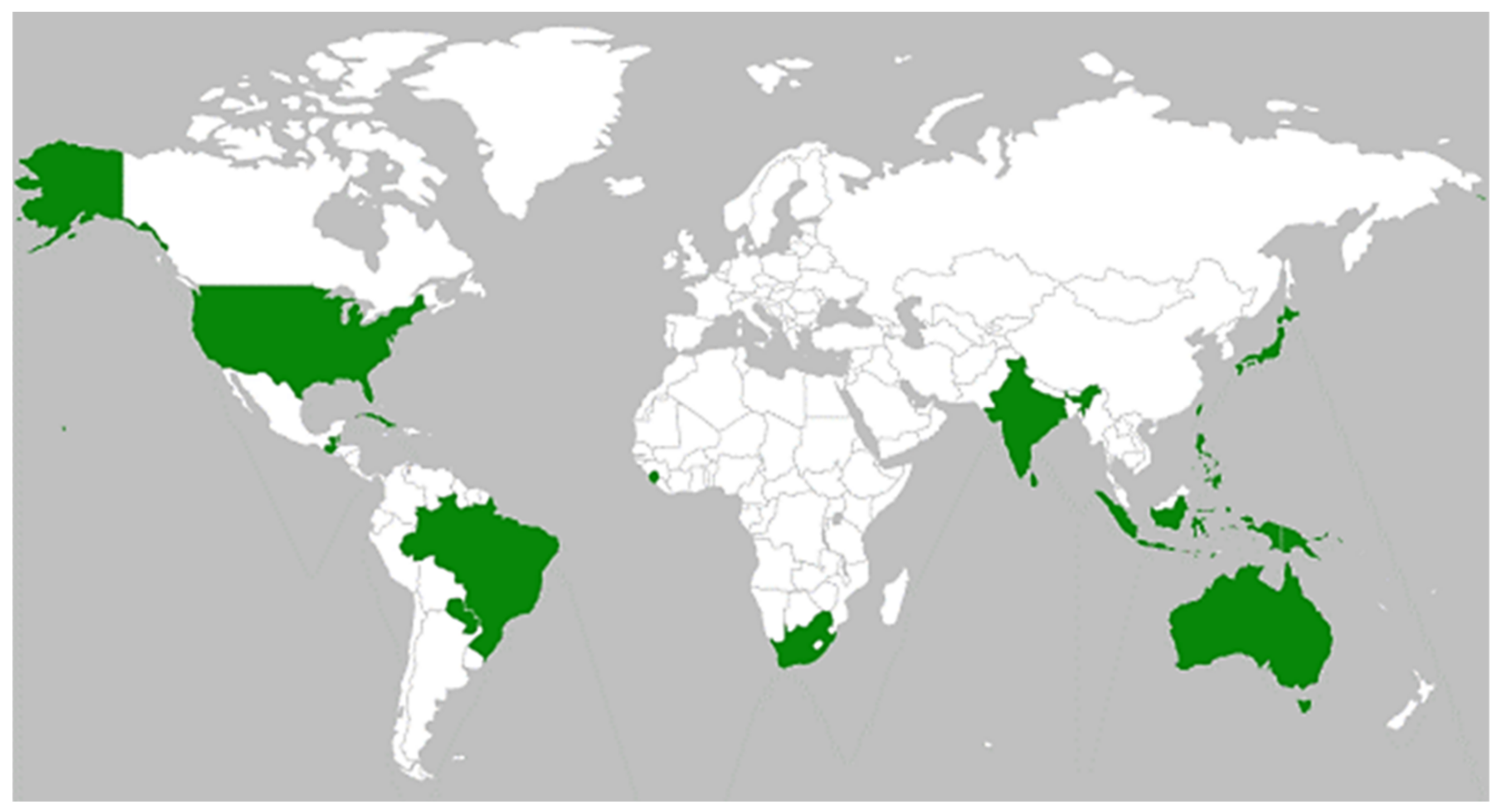
2.2. Diversity and Distribution
2.3. Ecology
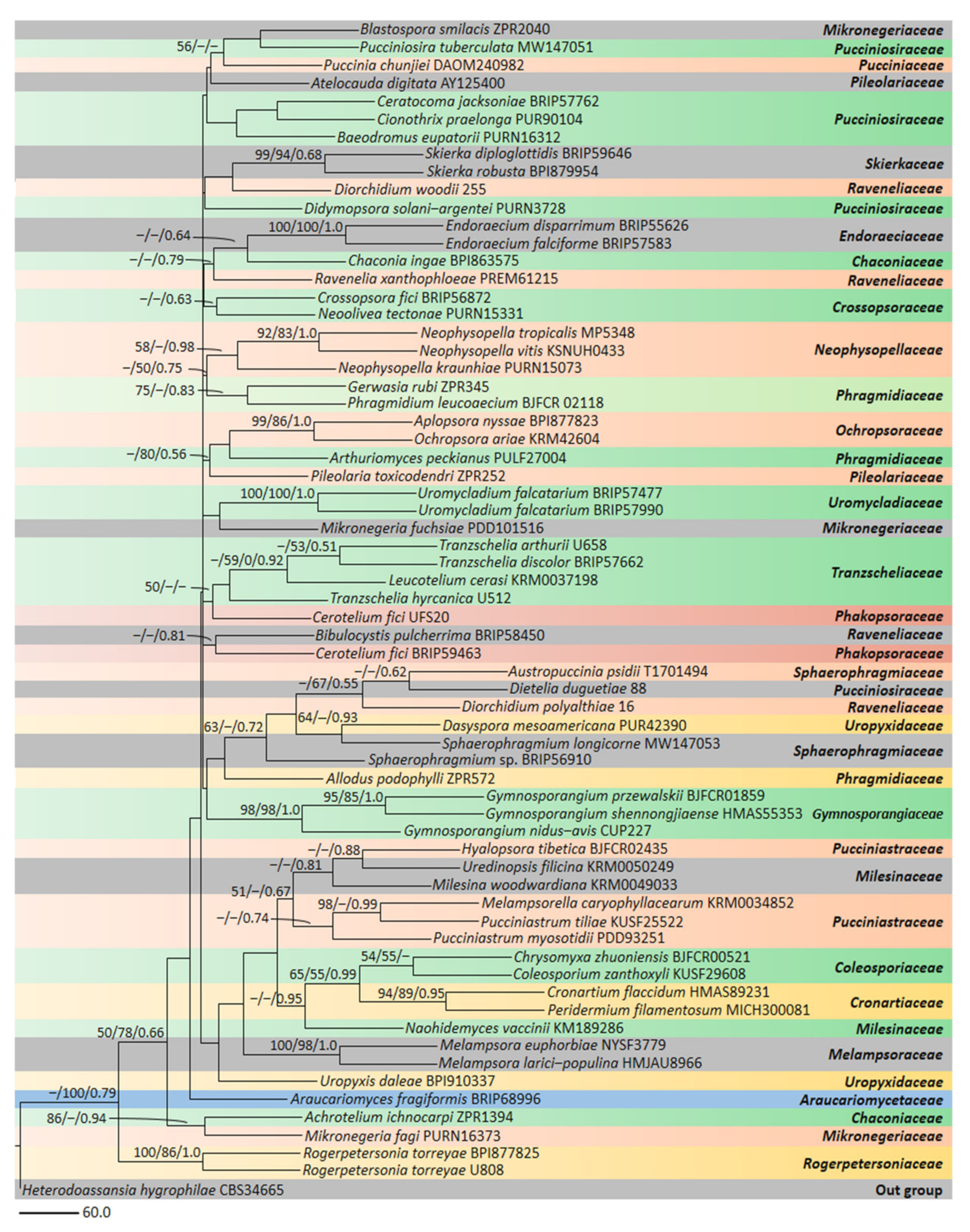
3. Phylogeny and Molecular Diversity
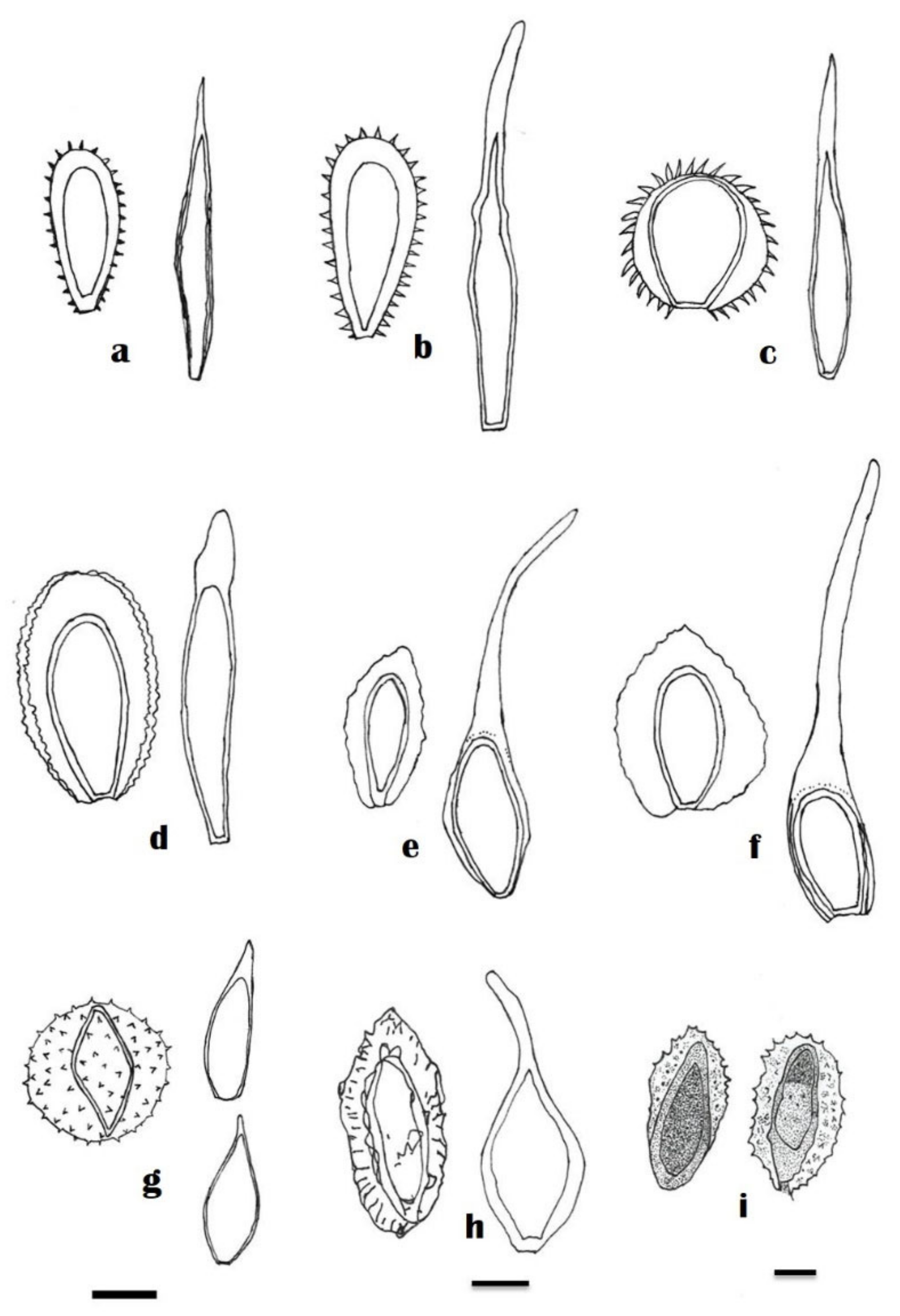
4. Taxonomy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cummins, G.B.; Hiratsuka, Y. Illustrated Genera of Rust Fungi, 3rd ed.; American Phytopathological Society (APS Press): Saint Paul, Minnesota, USA, 2003. [Google Scholar]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Cheek, M.; Nic Lughadha, E.; Kirk, P.; Lindon, H.; Carretero, J.; Looney, B.; Douglas, B.; Haelewaters, D.; Gaya, E.; Llewellyn, T.; et al. New scientific discoveries: Plants and fungi. Plants People Planet 2020, 2, 371–388. [Google Scholar] [CrossRef]
- Hyde, K.D. The numbers of fungi. Fungal Divers. 2022, 114, 1. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Aime, M.C.; McTaggart, A.R. A higher-rank classification for rust fungi, with notes on genera. Fungal Syst. Evol. 2021, 7, 21–47. [Google Scholar] [CrossRef] [PubMed]
- Carefoot, G.L.; Sprott, E.R. Famine on the Wind: Plant Diseases and Human History; Rand McNally & Company: Chicago, IL, USA, 1967. [Google Scholar]
- Raciborski, M. Parasitische Algen und Pilze Java’s; Staatsdruckerei: Vienna, Austria, 1900; Volume 2, pp. 1–46. [Google Scholar]
- Hennings, P. Fungi. Annales du Musée du Congo (Belge). Botanique 1907, 2, 85–106. [Google Scholar]
- Raciborski, M. Über einige javanische Uredineae (Moose). Bull. Int. L’academie Sci. Crac. Cl. Sci. Math. Nat. 1909, 3, 266–280. [Google Scholar]
- Arthur, J.C. Uredinales of Guatemala based on collections of E.W.D. Holway, II. Am. J. Bot. 1918, 5, 420–446. [Google Scholar] [CrossRef]
- Doidge, E.M. A preliminary study of the South African rust fungi. Bothalia 1926, 2, 1–228. [Google Scholar] [CrossRef]
- Sydow, P.; Sydow, H. Monographia Uredinearum. vol. 3. Pucciniaceae—Melampsoraceae Zaghouaniaceae—Coleosporiaceae. F. Borntraeger Leipz. Ger. 1915, 3, 726. [Google Scholar]
- Cunningham, G.H. The Rust Fungi of New Zealand, together with the Biology, Cytology and Therapeutics of the Uredinales; John McIndoe: Duedin, New Zealand, 1931. [Google Scholar]
- Arthur, J.C. Order Uredinales. North. Am. Flora 1907–1931, 7, 83–969. [Google Scholar]
- Dietel, P. Hemibasidii (Ustilagiales und Uredinales). In Die Natürlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1928; Volume 2, pp. 24–98. [Google Scholar]
- Hiratsuka, Y.; Cummins, G.B. Morphology of the spermogonia of the rust fungi. Mycologia 1963, 55, 487–507. [Google Scholar] [CrossRef]
- Cummins, G.B.; Hiratsuka, Y. Illustrated Genera of Rust Fungi; Rev. ed.; APS Press: Saint Paul, MN, USA, 1983. [Google Scholar]
- Koorders, S.H. Morophologische-Systematisch Notizen iiber einige Mittel-javanische, vorwiegend Blatter bewohnende Pilze. Verh. Konink. Akad. Wetenschappen 1907, 13, 161–264. [Google Scholar]
- Mains, E.B. The Genera, Skierka and Ctenoderma. Mycologia 1939, 31, 175–190. [Google Scholar] [CrossRef]
- Aime, M.C. Toward resolving family-level relationships in rust fungi (Uredinales). Mycoscience 2006, 47, 112–122. [Google Scholar] [CrossRef]
- Dietel, P. Kleine Beitrage zur Systematik der Uredineen V. Ann. Myc. 1925, 23, 182–185. [Google Scholar]
- Hiratsuka, N. Materials for a Rust-Flora of Riukiu Islands. II. Bot. Mag. LIV 1940, 646, 373–377. [Google Scholar] [CrossRef]
- Cummins, G.B. Uredinales of New Guinea—III. Mycologia 1941, 33, 143–154. [Google Scholar] [CrossRef]
- Ito, S.; Murayama, D. Notae mycologicae Asiae orientalis. IV. Trans. Sapporo Nat. Hist. Soc. 1943, 17, 160–172. [Google Scholar]
- Hiratsuka, N.; Shimabukuro, S. Uredinales of the Ryukyu Islands in Contributions to the rust flora of Eastern Asia, 8. Sci. Bull. Fac. Agric. Univ. Ryukyus 1954, 1, 1–56. [Google Scholar]
- Hijwegen, T. Fungi as plant taxonomists. II. Affinities of the Rosiflorae. Acta Bot. Neerl. 1981, 30, 479–491. [Google Scholar] [CrossRef]
- Dianese, J.C.; Medeiros, R.B.; Santos, L.T.P.; Furalnetto, C.; Sanchez, M.; Dianese, A.C. Batistopsora gen. nov. and new Phakopsora, Ravenelia, Cerotelium and Skierka species from the Brazilian Cerrado. Fitopatol. Bras. 1993, 18, 436–450. [Google Scholar]
- Purkayastha, R.; Pal, A. SEM studies on a mangrove rust of Sundarbans, Eastern India. Mycol. Res. 1998, 102, 692–694. [Google Scholar] [CrossRef]
- Gautam, A.K.; Avasthi, S. Fungi associated with Pistacia integerrima with a description of a new species and one new record from India. Acta Mycol. 2017, 52, 1100. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.Y.; Ling, J.F.; Xu, B. Additional report on rust fungi from subtropical China. Mycosystema 2021, 40, 912–919. [Google Scholar] [CrossRef]
- Dianese, J.C.; Inácio, C.A.; de Carvalho-Junior, A.A.; dos Santos, M.D.M.; Cantillo-Pérez, T.; Pinho, D.B. Exploring the Overlooked Diversity of Plant-Associated Cerrado Microfungi. Revisão Anual Patologia Plantas 1922, 28, 69–101. [Google Scholar] [CrossRef]
- Boedijn, K.B. The Uredinales of Indonesia. Nova Hedwig. 1960, 1, 463–494. [Google Scholar]
- Chavan, P.B. Critical Notes on Rust Fungi of Maharashtra (India). Sydowia Ann. Mycol. 1968, 22, 292–294. [Google Scholar]
- Spaulding, P. Foreign Diseases of Forest Trees of the World: An Annotated List; Agriculture Handbook No. 197; US Department of Agriculture: Washington, DC, USA, 1961; Volume 197, pp. 1–361. [Google Scholar]
- Shaw, D.E. Microorganisms in Papua New Guinea. Dept. Primary Ind. Res. Bull. 1984, 33, 1–344. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, USA, 14 November 2010. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree Version 1.4.0. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 26 September 2022).
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony, Version 4.0 b10; Sinauer Associates: Sunderland, Minnesota, USA, 2002. [Google Scholar]
- Kishino, H.; Hasegawa, M. Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data, and the branching order in Hominoidea. J. Mol. Evol. 1989, 29, 170–179. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. BMC Genom. 2002, 3, e4. [Google Scholar] [CrossRef] [Green Version]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Arther, J.C.; Cummins, G.B. Philippine rusts in the Clemens collection 1923–1926. II. The Philippine Jour. Sci. 1936, 61, 463–488. [Google Scholar]
- Sydow, H.; Sydow, P. Beitrag zur Kenntnis der Pilzflora der Philippinen Inseln. Ann. Myc. 1917, 15, 165–268. [Google Scholar]
- Sydow, H.; Petrak, F. Micromycetes philiippinenses. I. Ann. Myc. 1928, 26, 414–446. [Google Scholar]
- Sydow, H.; Petrak, F. Micromycetes philiippinenses. II. Ann. Myc. 1931, 29, 145–279. [Google Scholar]
- Sydow, H.; Sydow, P. Über einige Uredineen mit quellbaren Membranen und erhöhter Keimporenzahl. Ann. Myc. 1919, 17, 101–107. [Google Scholar]
- Cooke, M.C. Australian fungi. Grevillea 1889, 17, 55–56. [Google Scholar]
- Kuntze, O. Revisio generum plantarum. Leipzig 1898, 3, 1–576. [Google Scholar]
- Sydow, H. Über einige wenig bekannte Uredineen aus dem Kew herbar. Ann. Myc. 1922, 20, 54–60. [Google Scholar]
- Sydow, H. Novae fungorum species. XVIII. Ann. Myc. 1923, 21, 342–344. [Google Scholar]
- Gautam, A.K.; Verma, R.K.; Avasthi, S.; Sushma; Bohra, Y.; Devadatha, B.; Niranjan, M.; Suwannarach, N. Current insight into traditional and modern methods in fungal diversity estimates. J. Fungi 2022, 8, 226. [Google Scholar] [CrossRef]
- Gautam, A.K.; Avasthi, S.; Verma, R.K.; Sushma; Niranjan, M.; Devadatha, B.; Jayawardena, R.S.; Suwannarach, N.; Karunarathna, S.C. A global overview of diversity and phylogeny of the rust genus Uromyces. J. Fungi 2022, 8, 633. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Research Detail | Reference |
|---|---|---|
| 1900 | The genus Skierka was proposed based on the species Skierka cannarii | [8] |
| 1907 | Skierka congensis was described as the second species of this genus | [9] |
| 1907 | Skierka was placed in the Melampsoraceae family | [19] |
| 1907 | A subfamily Skierkatae of the Aecidiaceae (Pucciniaceae) was proposed and the genus Skierka, along with Ctenoderma, Sphenospora, and Chaconia, were included | [15] |
| 1909 | Skierka agallocha was described as a new species from Java | [10] |
| 1915 | Skierka was placed in Pucciniaceae based on the fusoid teliospores that resemble Uromyces-like rust | [13] |
| 1918 | Skierka holwayi was described as a new species from Central America | [11] |
| 1925 | Skierka was included as the oldest group of the Pucciniaceae | [22] |
| 1926 | Skierka robusta was described as a new species from Africa | [12] |
| 1928 | Skierka was placed in the tribe Skierkeae of the Pucciniaceae | [16] |
| 1939 | Skierka cristata was described. This species was derived from the re-examination of the rust sample of Ctenoderma cristata (Speg.) Sydow (Uredo cristata Speg.) | [20] |
| 1939 | Skierka diploglottidis was described as a new species | [20] |
| 1939 | Skierka petchii was described as a new species | [20] |
| 1939 | Skierka philippinensis was described as a new species | [20] |
| 1940 | Skierka agallocha was described from the Ryukyu Islands | [23] |
| 1941 | Skierka clemensiae was described as a new species | [24] |
| 1943 | Skierka nephelii was described as a new species | [25] |
| 1943 | Skierka toddaliae was described as a new species | [25] |
| 1954 | Skierka agallocha was reported from the Ryukyu Islands | [26] |
| 1981 | The host range Skierka was described with Sapindaceae, Rutaceae, Burseraceae, and Euphorbiaceae | [27] |
| 1993 | Skierka divinopolensis was described as a new species | [28] |
| 1998 | SEM studies of Skierka agallocha on mangrove rust of Sundarbans, Eastern India | [29] |
| 2017 | Skierka himalayensis was described as a new species | [30] |
| 2021 | Telial stage of Skierka nephelii was reported as a new rust addition to subtropical China | [31] |
| 2022 | Reviewed the contributions to Cerrado mycology (including Skierka) from the early 19th century to date | [32] |
| Name | Host (Family) | Biogeographical Region (Country) | References |
|---|---|---|---|
| Skierka agallochae Racib. | Excoecaria agallocha (Euphorbiaceae) | Batavia, Java (Indonesia), Okinawa Islands (Japan), and Maharashtra (India) | [10,23,26,33,34] |
| Skierka canarii Racib. | Canarium commune, Canarium moluccanum, Canarium villostttn, and Canarium sp. (Burseraceae) | Java (Indonesia), New Guinea, and the Philippines (Asia) | [8] |
| Skierka clemensiae Cummins | Canarium sp. (Burseraceae) | Papua New Guinea | [24] |
| Skierka congensis Henn. | Dombeya sp. (Sterculiaceae), Alchornea cordifolia, and Macaranga sp. (Euphorbiaceae) | The Democratic Republic of the Congo, and Sierra Leone | [9,20] |
| Skierka cristata Mains | Cupania americana, Cupania belizonsis, Cupania macrophylla, and Cupania sp. (Sapindaceae) | Cuba, Paraguay, Trinidad, and Belize | [20] |
| Skierka diploglottidis (Cooke and Massee) Mains | Dictyoneura obtuse and Diploglottis sp.(Sapindaceae) | Queensland and Bailey (Texas) | [20] |
| Skierka divinopolensis Dianese | Cupania rugosa and Matayba guianensis (Sapindaceae) | Minas Gerais (Brazil) | [28] |
| Skierka himalayensis A.K. Gautam and S. Avasthi | Pistacia integerrima (Pistaceaceae) | Himachal Pradesh (India) | [30] |
| Skierka holwayi Arthur | Thouinidium decandrum, and Thouinidium sp. (Sapindaceae) | Guatemala | [11] |
| Skierka nephelii S. Ito and Muray. | Litchi chinensis (Sapindaceae) | Taiwan (China) | [25,31] |
| Skierka petchii (Syd.) Mains | Sapindus bifoliolatus (Sapindaceae) | Sri Lanka | [20] |
| Skierka philippinensis Mains | Canarium luzonicum and Canarium sp. (Burseraceae) | Philippines and Papua New Guinea | [20,35,36] |
| Skierka robusta Doidge | Rhoicissus rhomboidea (Vitaceae) | South Africa | [12] |
| Skierka toddaliae (Petch) Hirats. | Toddalia aculeate (Rutaceae) | Sri Lanka | [25] |
| Taxon | Voucher Number (Collection Number) | GenBank Accession No. | ||
|---|---|---|---|---|
| LSU | ITS | SSU | ||
| Achrotelium ichnocarpi | ZPR1394 | MK518684 | MK518985 | MK488400 |
| Allodus podophylli | ZPR572 | MK518482 | MK518834 | MK488140 |
| Aplopsora nyssae | BPI877823 | MW049244 | NA | NA |
| Araucariomyces fragiformis | BRIP68996 | MW049245 | NA | MW049292 |
| Araucariomyces fragiformis | BRIP68996 | NG074475 | NA | NG073560 |
| Arthuriomyces peckianus | PULF27004 | NA | MW448622 | NA |
| Atelocauda digitate | - | NA | NA | AY125400 |
| Austropuccinia psidii | T1701494 | NA | MK020421 | NA |
| Baeodromus eupatorii | PURN16312 | MW049246 | NA | NA |
| Bibulocystis pulcherrima | BRIP58450 | MW049247 | NA | NA |
| Blastospora smilacis | ZPR2040 | MK518730 | MK519028 | MK488480 |
| Ceratocoma jacksoniae | BRIP57762 | KT199394 | NA | KT199382 |
| Cerotelium fici | BRIP59463 | MH047210 | NA | MW049299 |
| Cerotelium fici | UFS20 | NA | MZ047090 | NA |
| Chaconia ingae | BPI863575 | MW049249 | NA | NA |
| Chrysomyxa zhuoniensis | BJFCR00521 | MZ444061 | NR153462 | NA |
| Cionothrix praelonga | PUR90104 | MW049252 | NA | NA |
| Coleosporium zanthoxyli | KUSF29608 | MH460677 | MH465095 | NA |
| Cronartium flaccidum | HMAS89231 | MK208289 | MK193822 | NA |
| Crossopsora fici | BRIP56872 | MH047208 | NA | MH047213 |
| Dasyspora mesoamericana | PUR42390 | NA | NR136010 | NG064973 |
| Didymopsora solani argentei | PURN3728 | MW049254 | NA | NA |
| Dietelia duguetiae | 88 | NA | KM217365 | KM217382 |
| Diorchidium woodii | 255 | KM217352 | NA | KM217370 |
| Gerwasia rubi | ZPR345 | MK518735 | NA | MK488442 |
| Gymnosporangium przewalskii | BJFCR01859 | NG060667 | NR154073 | NA |
| Heterodoassansia hygrophilae | CBS34665 | NG064047 | NR160101 | NA |
| Hyalopsora tibetica | BJFCR02435 | NG081469 | MK795976 | NA |
| Leucotelium cerasi | KRM0037198 | KX228776 | KX228771 | NA |
| Melampsora euphorbiae | NYSF3779 | MK518509 | MK518852 | MK488184 |
| Melampsora larici populina | HMJAU8966 | MT757879 | MT759611 | NA |
| Melampsorella caryophyllacearum | KRM0034852 | ON063363 | ON063389 | NA |
| Mikronegeria fagi | PURN16373 | MW049267 | NA | MW049298 |
| Mikronegeria fuchsiae | PDD101516 | NA | KX985772 | NA |
| Milesina woodwardiana | KRM0049033 | NA | NR163315 | NA |
| Naohidemyces vaccinii | KM189286 | NA | MZ159489 | NA |
| Neoolivea tectonae | PURN15331 | NG074476 | NA | NG073561 |
| Neophysopella vitis | KSNUH0433 | OM420271 | OM423812 | NA |
| Ochropsora ariae | KRM42604 | KX228778 | KX228773 | NA |
| Peridermium filamentosum | MICH300081 | MK208299 | MK193831 | NA |
| Phragmidium leucoaecium | BJFCR02118 | MN264737 | MN264719 | NA |
| Pileolaria toxicodendri | ZPR252 | MK518537 | MK518871 | MK488231 |
| Pucciniastrum myosotidii | PDD93251 | NA | KJ716347 | KJ746815 |
| Pucciniastrum tiliae | KUSF25522 | OL519197 | OL519191 | NA |
| Puccinia chunjiei | DAOM240982 | NA | NR111548 | NA |
| Pucciniosira tuberculate | - | MW147051 | NA | NA |
| Ravenelia xanthophloeae | PREM61215 | MG946017 | MG945985 | NA |
| Rogerpetersonia torreyae | BPI877825 | NG075238 | NA | NG073502 |
| Rogerpetersonia torreyae | U808 | MG907207 | NA | NA |
| Skierka diploglottidis | BRIP59646 | MW049278 | NA | MW049304 |
| Skierka robusta | BPI879954 | MW049279 | NA | MW049305 |
| Sphaerophragmium longicorne | - | MW147053 | NA | MW147077 |
| Sphaerophragmium sp. | BRIP56910 | KJ862350 | NA | KJ862429 |
| Tranzschelia arthurii | U658 | MG948659 | MG947386 | NA |
| Tranzschelia discolor | BRIP57662 | KR994891 | NA | KR994969 |
| Tranzschelia hyrcanica | U512 | NA | MG948663 | NA |
| Uredinopsis filicina | KRM0050249 | MK302213 | MH908488 | NA |
| Uromycladium falcatarium | BRIP57477 | KJ632973 | NR138392 | KJ633013 |
| Uropyxis daleae | BPI910337 | NA | KY798364 | NA |
| Species | Taxonomic Characteristics | Reference | |||
|---|---|---|---|---|---|
| Uredia (mm) | Uredospores (µm) | Telia (mm) | Teliospores (µm) | ||
| Skierka agallochae Racib. | 0.1–0.4 | 20–43 × 47–90 | 0.1–0.8 × 0.5–0.8 | 8–12 × 60–100 | [10] |
| Skierka canarii Racib. | - | 14–16 × 24–30 | - | 7–10 × 65–75 | [8] |
| Skierka clemensiae Cummins | 0.5–2.0 | 10–15 × 23–30 | - | - | [24] |
| Skierka congensis Henn. | - | 8–16 × 23–38 | - | 6–8 × 50–85 | [9] |
| Skierka cristata Mains | - | 16–20 × 30–40 | - | 10–15 × 60–96 | [20] |
| Skierka diploglottidis (Cooke and Massee) Mains | - | 12–16 × 32–42 | - | 15–18 × 70–90 | [20] |
| Skierka divinopolensis Dianese | 21.6–26 | (24–) 28–32 (–34) × (50–) 55–77 | - | (15–) 17–20 × (35–) 43–48 (–50) | [28] |
| Skierka himalayensis A. K. Gautam and S. Avasthi | - | - | - | 10.5–18.9 × 35.7–48.3 | [30] |
| Skierka holwayi Arthur | - | 14–20 × 30–65 | - | 11–14 × 28–38 | [11] |
| Skierka nephelii S. Ito and Muray. | - | - | - | 12−20 × 23−75 | [25] |
| Skierka petchii (Syd.) Mains | - | 8–12 × 24–50 | - | 12–18 × 38–44 | [20] |
| Skierka philippinensis Mains | 0.2–0.5 | 12–16 × 28–54 | 0.2–0.5 | 11–19 × 64–110 | [20] |
| Skierka robusta Doidge | - | - | - | 20–27 × 120–180 | [12] |
| Skierka toddaliae (Petch) Hirats. | - | 28–36 × 40–70 | - | - | [25] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balkrishna, A.; Gautam, A.K.; Avasthi, S.; Niranjan, M.; Verma, R.K.; Arya, V.; Kumar, A.; Karunarathna, S.C.; Suwannarach, N. A Comprehensive Account of the Rust Genus Skierka (Skierkaceae). J. Fungi 2022, 8, 1243. https://doi.org/10.3390/jof8121243
Balkrishna A, Gautam AK, Avasthi S, Niranjan M, Verma RK, Arya V, Kumar A, Karunarathna SC, Suwannarach N. A Comprehensive Account of the Rust Genus Skierka (Skierkaceae). Journal of Fungi. 2022; 8(12):1243. https://doi.org/10.3390/jof8121243
Chicago/Turabian StyleBalkrishna, Acharya, Ajay Kumar Gautam, Shubhi Avasthi, Mekala Niranjan, Rajnish Kumar Verma, Vedpriya Arya, Ashwani Kumar, Samantha C. Karunarathna, and Nakarin Suwannarach. 2022. "A Comprehensive Account of the Rust Genus Skierka (Skierkaceae)" Journal of Fungi 8, no. 12: 1243. https://doi.org/10.3390/jof8121243