

Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China

1
Department of Plant Pathology, Agriculture College, Guizhou University, Guiyang 550025, China
2
Key Laboratory of Plant Resources Conservation and Germplasm Innovation in Mountainous Region (Ministry of Education), College of Life Sciences/Institute of Agro-Bioengineering, Guizhou University, Guiyang 550025, China
3
Private Bag 92170, Auckland Mail Centre, Auckland 1142, New Zealand
4
Faculdade de Ciências, Biosystems and Integrative Sciences Institute (BioISI), Universidade de Lisboa, Campo Grande, 1749-016 Lisbon, Portugal
*
Author to whom correspondence should be addressed.
J. Fungi 2022, 8(12), 1301; https://doi.org/10.3390/jof8121301
Submission received: 4 November 2022
/
Revised: 8 December 2022
/
Accepted: 12 December 2022
/
Published: 15 December 2022
(This article belongs to the Special Issue Polyphasic Identification of Fungi 2.0)
Abstract
:In a study of fungi isolated from plant material in Guizhou Province, China, we identified 23 strains of Diaporthales belonging to nine species. These are identified from multigene phylogenetic analyses of ITS, LSU, rpb2, tef1, and tub2 gene sequence data coupled with morphological studies. The fungi include a new genus (Pseudomastigosporella) in Foliocryphiaceae isolated from Acer palmatum and Hypericum patulum, a new species of Chrysofolia isolated from Coriaria nepalensis, and five new species of Diaporthe isolated from Juglans regia, Eucommia ulmoides, and Hypericum patulum. Gnomoniopsis rosae and Coniella quercicola are newly recorded species for China.
1. Introduction
Diaporthales is an important and species-rich ascomycetous order in the subclass Diaporthomycetidae (Sordariomycetes). Despite its cosmopolitan distribution and high diversity with distinctive morphology, this order has received relatively little attention. Currently, the existing classification lists 31 accepted families within the order Diaporthales [1], including Foliocryphiaceae, Diaporthaceae, Gnomoniaceae, and Schizoparmaceae. Members of Diaporthales have a wide range of ecological habitats and numerous modes of nutrition [2]. Excepting the members of Tirisporellaceae, most taxa in Diaporthales occur in terrestrial habitats. Species in Diaporthales form solitary or aggregated, immersed to erumpent, rarely superficial, and orange, brown, or black perithecial ascomata, with short or long necks that are located in stromatic tissues or substrates and with a lack of hamathecium or with few paraphyses [2,3,4,5]. Their asci are unitunicate with a conspicuous refractive ring [5,6]. Their ascospores are diverse in shape, size, and color. The asexual morphs of Diaporthales are generally coelomycetous [6], producing acervuli, pycnidial, or synnematal conidiomata and with or without a well-developed stroma. Conidiogenesis is phialidic or rarely annellidic, and conidia are usually unicellular or one-septate [6].
In China, the first monograph for Diaporthales referred to Phomopsis (=Diaporthe), which introduced 133 morphological taxa (including two specialized forma) isolated from 74 familial plants [7]. One diaporthalean pathogen that causes a devastating wilt disease for Cyathea lepifera was reported in Taiwan [8]. Pustulomyces accommodated in Diaporthaceae was revealed by morphology and molecular analyses [9]. Two novel families, Melansporellaceae and Diaporthosporellaceae, were introduced to accommodate the monotypic genera, Melanosporella and Diaporthosporella, based on both holomorphic morphology and phylogenetic analysis [10,11], and Foliocryphiaceae was established by Jiang et al. [1] to retain Chrysofolia, Foliocryphia, and Neocryphonectria. Fan et al. [12] reported families and genera of diaporthalean fungi associated with canker and dieback of tree hosts. In addition, dozens of Diaporthales taxa were first described in China [13,14,15,16].
Morphological comparisons and phylogenetic analyses have been commonly used to describe the taxa of Diaporthales and to confirm their taxonomic placement. Genealogical concordance phylogenetic species recognition (GCPSR) has also been used for the delineation of Foliocryphiaceae, Diaporthe, Gnomoniopsis, and Coniella species. GCPSR relies on performing a pairwise homoplasy index coupled with phylogenetic relatedness in a multi-locus dataset and the interpretation of nucleotide differences [17,18].
The present study follows a recently revised classification [1] combined with molecular data, morphology, and pairwise homoplasy index (PHI) test results and introduces seven novel taxa and two newly recorded taxa within the family Diaporthales found in Guizhou, China.
2. Materials and Methods
2.1. Sample Collection and Fungal Strain Isolation
The live plant samples were collected from Wengan, Longli, and Dejiang counties in Guizhou Province, China, in June and September 2021 and March 2022. They were placed in envelopes, taken back to the laboratory, and photographed. Before single-spore isolation, the surfaces of the samples were disinfected by spraying 2 or 3 times with 75% ethanol. The single spore isolation procedure of Chomnunti et al. [19] was followed in order to obtain pure cultures. Each pure culture was spread onto 90 mm diam. Petri dishes containing either potato dextrose agar (PDA) or oatmeal agar (OA) [20]. Diaporthe spp. were induced to sporulate by plating them on 2% water agar (WA) [21] containing sterilized pine needles. These dishes were cultured at a constant temperature (25 °C) under a 12 h light/dark regime in a light incubator. Dried holotype specimens were conserved in the Herbarium of the Department of Plant Pathology, Agricultural College, Guizhou University (HGUP). Ex-type cultures were conserved in the Culture Collection at the Department of Plant Pathology, Agriculture College, Guizhou University, China (GUCC).
2.2. Morphological Description
The pure cultures were grown on PDA and OA media in a constant-temperature incubator (25 °C) under a 12 h light/dark regime. Culture characteristics were recorded and examined using a stereo microscope (LEICA S9i, Wetzlar, Germany). Morphological observations were made with a Zeiss Scope 5 (Axioscope 5, Shanghai, China) equipped with an AxioCam 208 color camera (ZEN 3.0), and measurements were made with program (ZEN 3.0). Adobe Photoshop CC 2017 was used to make the photoplates. All new taxa were registered in MycoBank [22].
2.3. DNA Extraction, PCR Amplification, and Sequencing
Fresh mycelium was scraped from cultures using a sterilized scalpel, and genomic DNA was extracted using Fungal gDNA Kit (Biomiga #GD2416, San Diego, CA, USA) in accordance with the manufacturer’s instructions. Five genes were selected: internal transcribed spacers (ITS), 28S subunit rDNA (LSU), RNA polymerase II subunit 2 (rpb2), translation elongation factor 1 (tef1), and the β-tubulin gene region (tub2). Polymerase chain reactions (PCR) were carried out in 20 μL reaction volume, which contained 10 μL 2 × PCR Master Mix, 7 μL of ddH2O, 1 μL of each primer, and 1 μL of template DNA. The PCR thermal cycle program and primers are shown in Table 1. Purification and sequencing of PCR products were carried out by the Sangon Biotech Company (Shanghai, China). All isolates of all analyzed genes were deposited in GenBank.
2.4. Phylogeny
Sequences used in this study (Table 2, Table 3, Table 4 and Table 5) were assembled based on the closest matches from BLASTn search results (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and previous publications [1,29,30,31,32,33,34,35,36,37]. Alignments were conducted with the online version of MAFFT v. 7.505 [38], checked visually, and improved manually where necessary using BioEdit 7.1.3.0 [39]. Sequence matrix v. 1.7.8 [40] was used to concatenate the aligned sequences of the different loci. Ambiguous areas were excluded from the analysis using Aliview [41], and gaps were viewed as missing data. The final alignments were deposited in TreeBASE (www.treebase.org) under accession number 29787.
Phylogenetic analyses were carried out by maximum likelihood (ML), maximum parsimony (MP), and Bayesian inference (BI). The ML analysis was performed using RAxML-HPC BlackBox(8.2.12) [42] partial and general time reversible model (GTR) using the discrete gamma distribution as the evolution model by CIPRES Science Gateway version 3.3 [43]. Non-parametric bootstrap analysis was implemented with 1000 iterations. The resulting duplicates were plotted onto the best-scoring tree previously obtained.
Maximum parsimony (MP) analyses were performed with PAUP on XSEDE (4.a168) on CIPRES Science Gateway v. 3.3 using the heuristic search option with 1000 random sequence addition replicates and tree bisection and reconnection (TBR) with reconnection limit (=8) as the branch-swapping algorithm. Maxtrees were set to 5000 (and not increased). Branches were collapsed, creating polytomies if maximum branch length was zero. Tree length (TL), consistency index (CI), retention index (RI), rescaled consistency index (RC), and homoplasy index (HI) were calculated for each tree generated.
Bayesian inference (BI) analysis was performed by MrBayes 3.2.7a [44] in the CIPRES Science Gateway version 3.3. The optimal substitution model with gamma rates and dirichlet base frequencies for ITS, LSU, rpb2, tef1, and tub2 sequences was decided by modelGUI for each locus [45]. The Markov chain Monte Carlo (MCMC) sampling approach was used to calculate posterior probabilities (PP) [46]. Six simultaneous Markov chains were run for 50 million generations and trees were sampled every 1000th generation; thus, 50,000 trees were obtained. The first 25% of trees, representing the burn-in phase of the analyses, were discarded, and the remaining trees were used for calculating posterior probabilities (PP) in the majority rule consensus tree.
The phylogenetic trees were viewed with FigTree v. 1.4.3 [47] and processed with Adobe Illustrator CS5. ML bootstrap support (MLBS) and MP bootstrap support (PBS) equal or greater than 70% [48] and Bayesian posterior probabilities (PP) equal or greater than 0.95 [49] are displayed in the first, second, and third positions on the edited phylogenetic tree, respectively.
2.5. Genealogical Phylogenetic Species Recognition (GCPSR) Analysis
Morphologically and phylogenetically related species were analyzed using GCPSR as described by Taylor et al. [17] by the pairwise homogeneity index test (PHI) [50]. The PHI tests were performed in SplitsTree v. 4.17.1 [18,51] as described by Quaedvlieg et al. [52] to determine the level of recombination within phylogenetically closely related species. This test determines the null hypothesis probability (p-value) of no recombination within the dataset. When the p-value is less than 0.05, we reject the null hypothesis and accept the alternate hypothesis that there is evidence of the presence of recombination. The results were visualized by constructing a split graph using LogDet conversion and Splits options.
3. Results
3.1. Phylogenetic Analyses
To reveal the phylogenetic position of the family Foliocryphiaceae, genera Diaporthe, Gnomoniopsis, and Coniella, within the order Diaporthales, phylogenetic analyses were performed with ITS, LSU, rpb2, tef1, and tub2 sequence data.
The first sequence dataset of ITS, LSU, rpb2, tef1, and tub2 was analyzed to focus on Cryphonectriaceae and Foliocryphiaceae. The alignment included 43 taxa, including representatives of Cryphonectriaceae and Foliocryphiaceae and outgroup sequences of Dwiroopa lythri (CBS 109755, ex-type strain) and Dw. punicae (CBS 143163, ex-type strain) (Table 2). The aligned five-locus datasets comprised 3596 characters of the family Foliocryphiaceae, viz. ITS: 1–769, LSU: 770–1617, rpb2: 1618–2442, tef1: 2443–2952, and tub2: 2953–3596. Of these, 2145 characters were constant, 1131 characters were parsimony-informative, and 320 were parsimony-uninformative (gaps were treated as missing). The parameter settings used are shown in Table 6. A RAxML tree was selected to show the topology (Figure 1), and MP and Bayesian analyses resulted in similar topology to ML.
Two new strains of Chrysofolia coriariae sp. nov. (GUCC 416.4, ex-type strain and GUCC 416.14) collected during this study in Guizhou Province shared the same branch length with 100% MLBS/99% MPBS/1 PP support and were grouped with the type strains of Ch. colombiana (CPC 24986) and Ch. barringtoniae (TBRC 5647) with high statistical support, being (76% MLBS/99% MPBS)/(100% MLBS/99% MPBS/1 PP) (Figure 1), respectively. A comparison of the DNA base composition (Table 7) indicated that between our two strains and Ch. colombiana (CPC 24986), there were seven different bases in the ITS region, two different bases in the LSU region, and 149 different bases in the tef1 region. Between GUCC 416.4, GUCC 416.14, and Ch. barringtoniae (TBRC 5647), there were 31 different bases in the ITS region and four different bases in the LSU region. Unfortunately, Ch. colombiana did not have rpb2 or tub2 sequence data, and Ch. barringtoniae did not have rpb2, tef1, or tub2 sequence data.
Four new strains of Pseudomastigosporella guizhouensis sp. nov. (GUCC 406.6, ex-type strain, GUCC 405.3, GUCC 405.4, and GUCC 405.8) from China formed a stable subclade, clustered with the genera Neocryphonectria, Chrysofolia, and Foliocryphia. They formed a well-resolved clade (99% MLBS/94% MPBS/1 PP) within Foliocryphiaceae. Our four strains were closer to N. carpini (CFCC 53027, ex-type strain) and N. chinensis (CFCC 53025, ex-type strain and CFCC 53029) with high support in their respective branches (97% MLBS/1 PP) (Figure 1). A comparison of the DNA bases (Table 7) revealed 116/118/32 base pair differences in ITS, 22/21/21 base pair differences in LSU, no data and 75/76 base pair differences in rpb2, and no data and 130/130 base pair differences in tef1 between our four strains, N. carpini (CFCC 53027, ex-type strain), and N. chinensis (CFCC 53025, ex-type strain and CFCC 53029) but no tub2 sequences data for comparison.
The pairwise homoplasy index (PHI) (Figure 2a) test revealed that there was no significant recombination (p-value = 1.0) between our strains (GUCC 416.4, GUCC 416.14, GUCC 406.6, GUCC 405.3, GUCC 405.4, and GUCC 405.8) and three other genera in Foliocryphiaceae (Chrysofolia, Foliocryphia, and Neocryphonectria).
The second sequence dataset of ITS, tef1, and tub2 was analyzed in combination to infer the interspecific relationships within Diaporthe. The alignment included 55 taxa, including the outgroup sequences of Diaporthella corylina (CBS 121124, ex-neotype strain) and Di. cryptica (CBS 140348, ex-neotype strain) (Table 3). The aligned three-locus datasets comprised 1770 characters of Diaporthe, viz. ITS: 1–640, tef1: 641–1116 and tub2: 1117–1770. Of these, 959 characters were constant, 644 characters were parsimony-informative, and 167 were parsimony-uninformative (gaps were treated as missing). The parameter settings that were used are shown in Table 6. A RAxML tree was selected to show the topology (Figure 3), and MP and Bayesian analyses resulted in similar topology to ML.
Two new strains of Diaporthe juglandigena sp.nov. (GUCC 422.16, ex-type strain and GUCC 422.161) from China had a close relationship to D. chimonanthi (HGUP191001 and HGUP192087) and D. caryae (CFCC 52563, ex-epitype strain and PSCG520), supported by MLBS (96%), PBS (88%), PP (1) and MLBS (100%), PBS (100%), and PP (1), respectively (Figure 3). A comparison of the DNA bases (Table 7) showed that our strains kept some distinction from D. chimonanthi (HGUP191001 and HGUP192087) and D. caryae (CFCC 52563, ex-type strain and PSCG520) with 18/23 and 2/4 base pair differences in ITS, no data and 6/6 base pair differences in tef1, and 5/2 and 17/30 base pair differences in tub2. The pairwise homoplasy index (PHI) test (Figure 2b) revealed that there was no significant recombination (p-value = 0.5412) among these two strains (GUCC 422.16 and GUCC 422.161) to D. chimonanthi (HGUP191001 and HGUP192087) and D. caryae (CFCC 52563, ex-epitype strain and PSCG520).
Strains of Diaporthe eucommiigena sp.nov. (GUCC 420.9, ex-type strain and GUCC 420.19) shared the same branch length with 100% MLBS/100% MPBS/1 PP support and were grouped with the ex-type strains of D. passiflorae (CBS 132527, ex-type strain) and D. malorum (CAA734, ex-type strain, CAA740, and CAA752) with high statistical support (100% MLBS/100% MPBS/1 PP) (Figure 3). A comparison of the DNA bases (Table 7) revealed 11/15 bp differences in ITS, 23/19 bp differences in tef1, and 13/13 bp differences in tub2 between the two strains and D. passiflorae and D. malorum. The PHI test (Figure 2c) did not find statistically significant evidence of recombination (p-value = 1.0) between the strains (GUCC 420.9 and GUCC 420.19) and related taxa D. passiflorae (CBS 132527, ex-type strain) and D. malorum (CAA734, ex-type strain, CAA740, and CAA752).
Strains of Diaporthe dejiangensis sp.nov. (GUCC 421.2, ex-type strain and GUCC 421.21) have a close relationship with D. eres (CBS 138594 ex-type strain, CAA801) with high support (98% ML, 93% MP, 1 PP) and formed a well-resolved clade sister to D. eres. A comparison of the DNA bases (Table 7) showed 4/3/7 bp differences in ITS, 6/7/5 bp differences in tef1, and 6/7/13 bp differences in tub2 between GUCC 421.2 and GUCC 421.21 and the other three strains in the clade. Based on the PHI test (Figure 2d), there was no significant recombination (p-value = 1.0) between our strains (GUCC 421.2 and GUCC 421.21) and the sister taxon D. eres (CBS 138594, ex-type strain and CAA801).
Strains of Diaporthe tongrensis sp.nov. (GUCC 421.10, ex-type strain and GUCC 421.101) formed a highly supported subclade (83% ML, 0.99 PP) with D. phragmitis (CBS 138897, ex-type strain). There were eight base pair differences in the ITS and 13 base pair difference in the tub2. Unfortunately, D. phragmitis (CBS 138897, ex-type strain) did not have tef1 sequences data for comparison. The PHI test (Figure 2e) did not find statistically significant evidence of recombination (p-value = 1.0) between our Diaporthe strains (GUCC 421.10, ex-type strain and GUCC 421.101) and related taxa D. ellipicola (CGMCC 3.17084, ex-type strain and CGMCC 3.17085) and D. phragmitis (CBS 138897, ex-type strain).
Two new strains of Diaporthe hyperici sp. nov. (GUCC 414.4, ex-type strain and GUCC 414.41) formed a high-support subclade (100% ML, 100% MP, 1.00 PP) with D. caulivora (CBS 127268, ex-type strain, Dip1, and Dpc11). There were 17 base pair differences in ITS, 30 base pair differences in tef1, and 11 base pair differences in the tub2 from our strains based on a DNA base comparison (Table 7). The PHI test (Figure 2f) did not find any statistically significant evidence of recombination (p-value = 1.0) between our two strains (GUCC 414.4, GUCC 414.41) and strains of D. caulivora.
The third sequence dataset of ITS, LSU, rpb2, tef1, and tub2 was analyzed in combination to infer the interspecific relationships within Gnomoniopsis. The alignment included 29 taxa, including the outgroup sequences of Sirococcus tsugae (CBS 119626) (Table 4). The aligned five-locus datasets comprised 3354 characters of Gnomoniopsis, viz. ITS: 1–572, LSU: 573–1423, rpb2: 1424–2460, tef1: 2461–2861, and tub2: 2862–3354. Of these, 2431 characters were constant, 615 characters were parsimony-informative, and 308 were parsimony-uninformative (gaps were treated as missing). The parameter settings that were used are shown in Table 6. A RAxML tree was selected to show the topology (Figure 4), and MP and Bayesian analyses resulted in similar topology to RAxML.
Our strains of Gnomoniopsis rosae (GUCC 408.7 and GUCC 408.17) clustered in the same subclade (100% ML, 100% MP, 1.00 PP) with G. rosae (CBS 145085), and there were identical sequences in the ITS, LSU, and rpb2 regions. The phi test results (p-value = 1.0) (Figure 2g) of our strains (GUCC 408.7 and GUCC 408.17), G. rosae (CBS 145085), G. angolensis (CBS 145057), and G. clavulata (AR 4313) showed no statistically significant recombination.
The fourth sequence dataset of ITS, LSU, and tef1 was analyzed in combination to infer the interspecific relationships within Coniella. The alignment included 32 taxa, including the outgroup sequences of C. fragariae (CBS 172.49, ex-type strain) and C. nigra (CBS 165.60, ex-type strain) (Table 5). The aligned three-locus datasets comprised 2165 characters of Coniella, viz. ITS: 1–595, LSU: 596–1767, and tef1: 1768–2165. Of these, 1717 characters were constant, 385 characters were parsimony-informative, and 63 were parsimony-uninformative (gaps were treated as missing). The parameter settings that were used are shown in Table 6. A RAxML tree was selected to show the topology (Figure 4), and MP and Bayesian analyses resulted in similar topology to RAxML.
Strains of Coniella quercicola (GUCC 414.2, GUCC 414.21, GUCC 412.3, GUCC 405.6, and GUCC 405.16) clustered very close to C. quercicola (CBS 904.69, ex-type strain, CBS 283.76, and CPC 12133) with only MP support (95% MPBS) (Figure 4). A comparison of the DNA base composition (Table 7) indicated that between C. quercicola (CBS 904.69, ex-type strain, CBS 283.76, and CPC 12133) and our five strains (GUCC 414.2, GUCC 414.21, GUCC 412.3, GUCC 405.6, and GUCC 405.16), there were identical sequences in the ITS and LSU regions, but 1/5/10/10/18/7/6 bases were different in the tef1 region. The PHI test (Figure 2h) did not find statistically significant evidence of recombination (p-value = 0.2264) between our five strains and related taxa C. quercicola (CBS 904.69, ex-type strain, CBS 283.76, and CPC 12133).
3.2. Taxonomy
Pseudomastigosporella S.Y. Wang, Yong Wang bis, and Y. Li, gen. nov.
MycoBank Number: MB846026
Etymology: In reference to Mastigosporella, to which this genus is morphologically similar.
Classification: Foliocryphiaceae, Diaporthales, Sordariomycetes.
Description: Life style: Parasitic, leaves of Hypericum patulum and Acer palmatum. Asexual morph: Conidiomata pycnidial, globose or subglobose, base immersed, separate to aggregated, mycelium superficial, fluffy, granular, white or gray-white to pale yellow, exuding light brown-orange to medium brown-orange to deep brown-orange conidial masses, bright yellow or light orange in lactic acid, 2–5 wall layers of olive to gray-green textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells arising from base, central cushion of hyaline cells, densely aggregated, slightly thicker, cylindrical to ampulliform, simple, lining the inner cavity of base, mostly hyaline, sometimes pale olive, smooth, cylindrical to ampulliform, straight to curved, sometimes wider at the base. Conidia solitary, hyaline, smooth, guttulate, fusoid to ellipsoidal, sometimes long bubble-shaped, straight to curved, aseptate, base tapering with flattened scar, apex with 1 tubular appendage. Sexual morph: Unknown.
Type species: Pseudomastigosporella guizhouensis S.Y. Wang, Yong Wang bis & Y. Li.
Notes: In Foliocryphiaceae, the important morphological characters of asexual morph was to produce dimorphic conidia. The microconidia were minute, cylindrical, aseptate, hyaline to pale brown; macroconidia were fusoid, aseptate, hyaline [1]. However, Pseudomastigosporella only had macroconidia but like species in Mastigosporellaceae with an apical appendage developing as continuation of conidium body. This feature contradicted the root of “key to genera in Cryphonectriaceae, Foliocryphiaceae, and Mastigosporellaceae” provided by Jiang et al. [1]. However, following our phylogenetic analyses we still proposed that Pseudomastigosporella should be placed in Foliocryphiaceae family.
Pseudomastigosporella guizhouensis S.Y. Wang, Yong Wang bis, and Y. Li, sp. nov.
MycoBank Number: MB846027, Figure 5.
Etymology: In reference to the location where the fungus was found, being isolated from Guizhou Province.
Type: China, Guizhou Province, Wengan County, on leaves of Hypericum patulum and Acer palmatum, June 2021, S.Y. Wang (HGUP 406, holotype; HGUP 405, ex-type living culture GUCC 406.6).
Description: Life style: Parasitic, leaves of Hypericum patulum and Acer palmatum. Asexual morph: Conidiomata pycnidial, globose or subglobose, base immersed, separate to aggregated, mycelium superficial, fluffy, granular, white or gray-white to pale yellow, producing light brown-orange to medium brown-orange to deep brown-orange conidial masses, up to 570 μm diam., bright yellow or light orange in lactic acid, 2–5 wall layers of olive to gray-green textura angularis, 50–570 µm diam. Conidiophores reduced to conidiogenous cells. Conidiogenous cells arising from base, central cushion of hyaline cells, densely aggregated, slightly thicker, cylindrical to ampulliform, simple, lining the inner cavity of base, mostly hyaline, sometimes pale olive, smooth, cylindrical to ampulliform, straight to curved, sometimes wider at the base, 5–20 × 1.5–4.5 µm ( = 12 × 3 µm; n = 20). Conidia solitary, hyaline, smooth, guttulate, fusoid to ellipsoidal or fish-shaped, sometimes long bubble-shaped, straight to curved, aseptate, 15–31 × (3.5–)5.5–8.5 µm ( = 25 × 6.5 µm; n = 30), base tapering with flattened scar, 1.5–4 µm diam., with 1 apical, tubular appendage, 3.5–14.5 µm long. Sexual morph: Unknown.
Material examined: China, Guizhou Province, Wengan County, on leaves of Hypericum patulum and Acer palmatum, June 2021, S.Y. Wang (HGUP 406, holotype; HGUP 405); culture ex-type GUCC 406.6, additional living culture: GUCC 405.3, GUCC 405.4, GUCC 405.8.
Culture characteristics: Colonies covering 9 cm Petri dish after 2 weeks at 25 °C and under a 12 h light/dark regime. On PDA, white or gray-white, fluffy, granular, effuse surface, reverse white or beige; on OA, white or gray-white to pale yellow, fluffy, granular, effuse surface, exuding light brown-orange to medium brown-orange to deep brown-orange conidial masses, reverse white or beige to pale yellow.
Notes: Although Ps. guizhouensis produces macroconidia with one tubular apical appendage, the conidia of this species differs in shape and size from those of Chrysofolia and Foliocryphia. The conidia of Chrysofolia are ellipsoidal and measure (4–)6–7.5(–10) × (2–)2.5(–3) µm [53], while those of Foliocryphia are also ellipsoidal and measure (5–)6–8(–9) × (2–)2.5(–3) µm [54]. The results of the DNA base comparisons (Table 7) showed that there were striking differences in each gene among our four Pseudomastigosporella strains and adjacent genera. Based on its distinct morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results, Pseudomastigosporella was described here as a new genus in Foliocryphiaceae with Ps. guizhouensis as the type species.
Chrysofolia coriariae S.Y. Wang, Yong Wang bis, and Y. Li, sp. nov.
MycoBank Number: MB845958, Figure 6.
Etymology: In reference to the host plant Coriaria nepalensis, from which this fungus was collected.
Type: China, Guizhou Province, Longli County, on leaves of Coriaria nepalensis, June 2021, S.Y. Wang (HGUP 416, holotype; ex-type living culture GUCC 416.4).
Description: Life style: Parasitic, leaves of Coriaria nepalensis. Asexual morph: Conidiomata pycnidial, globose or subglobose, separate to aggregated, mycelium superficial and immersed, exuding yellow to bright orange to brown-orange wet conidial masses, green-brown in lactic acid, but bright yellow or light orange in sterile water, 2–6 wall layers of green-brown to brown textura angularis, 50–400 µm diam.; neck 15–60 µm long, 50–200 µm diam. where attached to the globose, terminating in an obtusely rounded apex. Conidiophores reduced to conidiogenous cells. Conidiogenous cells arising from base, central cushion of hyaline cells, densely aggregated, slightly thicker, tapering or cylindrical to ampulliform, simple, lining the inner cavity of base, hyaline, smooth, cylindrical to ampulliform, straight to curved, wider at the base, 5–20 × 1–3.5 µm. Conidia solitary, hyaline, smooth, ellipsoidal, or crescent-shaped, straight to allantoid, apex obtuse, base tapering with flattened scar, 0.5 µm diam., 5.5–9 × 2–4 µm ( = 7 × 2.7 µm; n = 30). Sexual morph: Unknown.
Material examined: China, Guizhou Province, Longli County, on leaves of Coriaria nepalensis, June 2021, S.Y. Wang (HGUP 416, holotype); culture ex-type GUCC 416.4, additional living culture: GUCC 416.14.
Culture characteristics: Colonies culturing under a controlled temperature light incubator at 25 °C and under a 12 h light/dark regime for 2 weeks. Colonies on PDA 75–90 mm diam. after 2 weeks at 25 °C, light brown to white or gray-white, felty, effuse surface, with white fluffy even mycelium margin, reverse brown to light brown to white edge. Colonies on OA 65–85 mm diam. after 2 weeks at 25 °C, light brown to white or gray-white, flat surface, exuding orange or brown conidial masses, reverse light brown to white or gray-white.
Notes: The conidiogenous cells of Ch. coriariae (5–20 × 1–3.5 µm) are longer than those of Ch. colombina (5–8 × 2–3 µm) [53] and Ch. barringtoniae (3–7.5 × 2–3 µm) [55], and the conidiomata of Ch. coriariae (5–400 µm) are larger than those of Ch. barringtoniae (5–110 µm) [55]. Chrysofolia coriariae was phylogenetically distinct from other known species (Figure 1) and displayed some differences in the DNA base comparison with Ch. colombina and Ch. barringtoniae (Table 7). The new isolates were described here as a new species based on their distinct morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results.
Diaporthe juglandigena S.Y. Wang, Yong Wang bis, and Y. Li, sp. nov.
MycoBank Number: MB845959, Figure 7.
Etymology: Name refers to the plant host genus (Juglans) from which this fungus was collected.
Type: China, Guizhou Province, Dejiang County, on the peel of Juglans regia, September 2021, S.Y. Wang (HGUP 422, holotype; ex-type living culture GUCC 422.16).
Description: Life style: Parasitic, peels of Juglans regia. Asexual morph: Conidiomata pycnidial, scattered, immersed or superficial, irregular globose and subglobose to slightly erumpent, black conidial masses surrounded by white mycelium, up to 2 mm diam, exuding transparent drops of water. Conidiophores reduced to conidiogenous cells. Conidiogenous cells densely aggregated, slightly thicker, subulate, simple, rarely branched above, simple, tapering, hyaline, smooth, 19–34 × 1–2.5 µm ( = 27 × 1.7 µm; n = 20), wider at base, rarely branched, densely aggregated, cylindrical, straight to sinuous. Alpha conidia hyaline, fusoid to ellipsoidal, asymmetrical, frequently guttulate, smooth-walled, 0–1-septate, tapering towards both ends, mostly straight, 5–8 × 2–3 µm ( = 6.4 × 2.3 µm; n = 30). Beta conidia infrequent, hyaline, filiform, aseptate, smooth, eguttulate, apex acute, mostly curved, 23–36 × 1–2 µm ( = 31 × 1.3 µm; n = 10). Gamma conidia not observed. Sexual morph: Not observed.
Material examined: China, Guizhou Province, Dejiang County, on the peel of Juglans regia, September 2021, S.Y. Wang (HGUP 422, holotype); culture ex-type GUCC 422.16, additional living culture: GUCC 422.161.
Culture characteristics: Colonies covering 9 cm Petri dish after 2 weeks at 25 °C and under a 12 h light/dark regime; spreading with uneven aerial mycelium. On PDA, surface with abundant aerial mycelium, white or gray-white to pale brown; reverse white to pale yellow to light brown. On OA surface with white or pale white to pale yellow thin aerial mycelium, with black conidial masses surrounded by white or gray-white mycelium; reverse white or beige to light yellow. On pine needles with irregular dark green to black subglobose conidial masses surrounded by thick white mycelium.
Notes: The conidiomata of D. juglandigena (2 mm diam.) are larger than those of D. chimonanthi (= Phomopsis chimonanthi) (150–238 µm wide, 130–230 µm high) [56] and D. caryae (310–325 µm diam.) [11]. The conidiogenous cells and beta conidia of D. juglandigena (19–34 × 1–2.5 µm, 23–36 × 1–2 µm) are longer than those of Phomopsis chimonanthi (13–25 × 1.6–2.5 µm, 15–18 × 1–1.5 µm) and D. caryae (7–11 × 1.4–2.2 µm, 15–34 × 1.1–1.4 µm), while the alpha conidia of D. juglandigena (5–8 × 2–3 µm) are shorter than those of P. chimonanthi (6.6–8.8 × 1.8–2.2 µm) and D. caryae (7–8.5 × 2.1–2.5 µm). Diaporthe juglandigena was phylogenetically distinct from the species presently known from DNA analyses (Figure 3). The results of DNA base comparisons (Table 7) showed that there were significant differences in three loci between our two D. juglandigena strains and sister species (D. chimonanthi and D. caryae). Based on its distinct morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results, D. juglandigena was described here as a new species.
Diaporthe eucommiigena S.Y. Wang, Yong Wang bis, and Y. Li, sp. nov.
MycoBank Number: MB845961, Figure 8.
Etymology: eucommiigena, in reference to plant host (Eucommia ulmoides), from which the fungus was isolated.
Type: China, Guizhou Province, Guiyang, Huaxi District, South Campus of Guizhou University, on dead woods of Eucommia ulmoides, March 2022, S.Y. Wang (HGUP 420, holotype; ex-type living culture GUCC 420.9).
Description: Life style: Saprobic, dead woods of Eucommia ulmoides. Asexual morph: Conidiomata pycnidial, separated but sometimes aggregated, immersed, sometimes superficial, irregular globose or subglobose, forming dark olive or dull green to black conidial masses, up to 2 mm diam., sometimes surrounded by white mycelium. Conidiophores reduced to conidiogenous cells. Conidiogenous cells densely aggregated, slightly thicker, subulate, hyaline, simple, rarely branched above, simple, hyaline, smooth, 12–27.5 × 1.5–3 µm ( = 19 × 2.2 µm; n = 20), wider at base, tapering at apex, rarely branched, densely aggregated, cylindrical, slightly bent. Alpha conidia hyaline, fusoid to ellipsoidal, frequently guttulate, asymmetrical, smooth-walled, 0–1-septate, tapering towards both ends, mostly straight, 5.5–8 × 1.5–3 µm ( = 7 × 2.3 µm; n = 30). Beta conidia hyaline, filiform, aseptate, smooth, eguttulate, apex acute, mostly curved, 27–37 × 1–2 µm ( = 32 × 1.3 µm; n = 10). Gamma conidia hyaline, fusoid to ellipsoidal, frequently guttulate, smooth, aseptate, straight, tapering at apex, 7.5–10 × 1.5–2.5 µm ( = 8.6 × 2.1 µm; n = 20). Sexual morph: Not observed.
Material examined: China, Guizhou Province, Guiyang, Huaxi District, South Campus of Guizhou University, on dead wood of Eucommia ulmoides, March 2022, S.Y. Wang (HGUP 420, holotype); culture ex-type GUCC 420.9, additional living culture: GUCC 420.19.
Culture characteristics: Colonies covering 9 cm Petri dish after 2 weeks at 25 °C and under a 12 h light/dark regime; spreading with uneven aerial mycelium. On PDA, surface with abundant white to pale yellow, uneven zonated aerial mycelium and margin, distinctly imbricated like a flower; reverse with pale yellow to light brown and pale pink, uneven zonated aerial mycelium and margin, exuding abundant dark green to black spots with age. On OA surface with uneven white to olive aerial mycelium, forming black conidial masses surrounded by white or gray-white mycelium; reverse white to olive, irregular. On pine needles with irregular dark green to black subglobose conidial masses surrounded by thick gray-white mycelium.
Notes: The conidiomata of Diaporthe eucommiigena (2 mm diam.) are larger than those of D. passiflorae (300 µm diam.) [57]. The conidiogenous cells of D. passiflorae are 2–3-septate, while those of D. eucommiigena are aseptate. The conidiogenous cells of D. eucommiigena (12–27.5 × 1.5–3 µm) are smaller than those of D. passiflorae (20–30 × 2.5–4 µm), and the beta conidia of D. eucommiigena (27–37 × 1–2 µm) are longer than those of D. passiflorae (14–20 × 1.5–2 µm) and D. malorum (17.4–26.6 × 0.8–2 µm) [21]. Gamma conidia were not observed in D. passiflorae but are present in D. eucommiigena. Diaporthe eucommiigena was phylogenetically distinct from the species presently known based on the DNA data (Figure 3). A comparison of the DNA bases (Table 7) showed significant difference between D. eucommiigena and adjacent species (D. passiflorae and D. malorum). Based on its distinct morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results, D. eucommiigena was described here as a new species.
Diaporthe dejiangensis S.Y. Wang, Yong Wang bis, and Y. Li, sp. nov.
MycoBank Number: MB845962, Figure 9.
Etymology: Name refers to the location (Dejing), from where the host plant was collected.
Type: China, Guizhou Province: Dejiang County, on the peel of Juglans regia, September 2021, S.Y. Wang (HGUP 421, holotype; ex-type living culture GUCC 421.2).
Description: Life style: Parasitic, peels of Juglans regia. Asexual morph: Conidiomata pycnidial, solitary to aggregated, immersed or superficial, irregular globose or subglobose to depressed, exuding white to dark brown to black conidial masses, with age surrounded by thin white mycelium, up to 2 mm diam., 4–7 wall layers of olive textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells densely aggregated, hyaline, smooth, cylindrical, wider at base, mostly straight, phialidic, simple, subcylindrical, tapering towards apex, hyaline, smooth, 9.5–17 × 1–3 µm ( = 13 × 1.8 µm; n = 30), mostly straight, rarely branched. Alpha conidia hyaline, fusiform to ellipsoidal, frequently guttulate, asymmetrical, smooth-walled, 0–1-septate, rounded towards both ends, mostly straight, 6–8.5 × 1.5–3 µm ( = 7 × 2.3 µm; n = 30). Beta conidia and gamma conidia not observed. Sexual morph: Not observed.
Material examined: China, Guizhou Province, Dejiang County, on the peel of Juglans regia, September 2021, S.Y. Wang (HGUP 421, holotype); culture ex-type GUCC 421.2, additional living culture: GUCC 421.21.
Culture characteristics: Colonies covering 9 cm diam. Petri dish after 2 weeks at 25 °C under a 12 h light/dark regime. On PDA surface with thick aerial mycelium, flat, velvet, white and beige; reverse white to pale yellow. On OA surface with white or pale white thin aerial mycelium, exuding black conidial masses, surrounded by white mycelium; reverse white or beige. On pine needles, irregular, black, globose conidial masses surrounded by thick white mycelium.
Notes: The conidiomata of D. dejiangensis (2 mm diam.) are larger than those of D. cotoneastri (1.5 mm diam.) [58], while its alpha conidia (6–8.5 × 1.5–3 µm) are smaller than those of D. cotoneastri (6–10 × 2–3 µm). Neither beta nor gamma conidia were observed for D. dejiangensis, while D. cotoneastri produced beta conidia (18–25 × 1 µm). Diaporthe dejiangensis was phylogenetically distinct from the species presently known based on the DNA data (Figure 3). The results of the DNA base comparisons are shown in Table 7 and indicate that there were many base differences among three genes. Based on its distinct morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results, D. dejiangensis was described here as a new species.
Diaporthe tongrensis S.Y. Wang, Yong Wang bis, and Y. Li, sp. nov.
MycoBank Number: MB845963, Figure 10.
Etymology: tongrensis, in reference to the city (Tongren) where the fungus was isolated.
Type: China, Guizhou Province, Tongren City, Dejiang County, on the peel of Juglans regia, September 2021, S.Y. Wang (HGUP 421, holotype; ex-type living culture GUCC 421.10).
Description: Life style: Parasitic, peels of Juglans regia. Asexual morph: Conidiomata pycnidial, separated, immersed or superficial, irregular globose or subglobose to depressed, exuding black conidial masses surrounded by white mycelium, up to 2.5 mm diam., 7–10 wall layers of olive textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells densely aggregated, slightly thicker, subulate, simple, rarely branched above, tapering, hyaline, smooth, 12–24 × 1.5–2.5 µm ( = 16 × 1.8 µm; n = 20), wider at base, cylindrical, straight to sinuous. Alpha conidia hyaline, fusoid to ellipsoidal, asymmetrical, smooth-walled, 1-septate, rounded towards both ends, mostly straight, 5.5–7.5 × 2–3 µm ( = 6.5 × 2.5 µm; n = 30). Beta conidia infrequent, hyaline, filiform, aseptate, eguttulate, smooth, apex acute, base slightly truncate, mostly straight, sometimes curved, 20–30 × 1–2 µm ( = 25.5 × 1.5 µm; n = 15). Gamma conidia not observed. Sexual morph: Not observed.
Material examined: China, Guizhou Province, Dejiang County, on the peel of Juglans regia, September 2021, S.Y. Wang (HGUP 421, holotype); culture ex-type GUCC 421.10, additional living culture: GUCC 421.101.
Culture characteristics: Colonies covering 9 cm diam. Petri dish after 2 weeks at 25 °C and a 12 h light/dark regime; spreading with aerial mycelium and uneven zonation. On PDA, surface with abundant aerial mycelium, with white uneven zonated aerial mycelium in the middle; reverse with white to pale yellow to light brown, uneven zonated aerial mycelium. On OA, surface with white or pale white, thin aerial mycelium, exuding black conidial masses surrounded by white mycelium; reverse white or beige. On pine needles, irregular black subglobose conidial masses surrounded by white mycelium.
Notes: The conidiomata of D. tongrensis (2.5 mm diam.) are larger than those of D. phragmitis (250 µm diam.) [59], and its conidiogenous cells are aseptate, while those of D. phragmitis are 1–3-septate. The conidiogenous cells of D. tongrensis (12–24 × 1.5–2.5 µm) are smaller than those of D. phragmitis (20–30 × 3–4 µm). The alpha conidia of D. tongrensis (5.5–7.5 × 2–3) are shorter than those of D. phragmitis (7–8 × 2–3 µm). Diaporthe tongrensis formed beta conidia, while neither beta nor gamma conidia were observed in D. phragmitis. Diaporthe tongrensis is phylogenetically distinct from the presently known species based on the DNA data (Figure 3). D. tongrensis showed some differences in the DNA base comparison with D. phragmitis, as shown in Table 7. Diaporthe tongrensis was described as a new taxon based on the high phylogenetic support for the clade, distinct morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results with adjacent species.
Diaporthe hyperici S.Y. Wang, Yong Wang bis, and Y. Li, sp. nov.
MycoBank Number: MB845965, Figure 11.
Etymology: Name refers to Hypericum patulum, the host genus from which this fungus was collected.
Type: China, Guizhou Province, Longli County, on leaves of Hypericum patulum, June 2021, S.Y. Wang (HGUP 416, holotype; ex-type living culture GUCC 416.4).
Description: Life style: Parasitic, leaves of Hypericum patulum. Asexual morph: Conidiomata pycnidial, separated or aggregated, immersed or superficial, globose or subglobose, deep brown to black, exuding black droplets from central ostioles with age, up to 3 mm diam., 6–10 wall layers of brown-green or brown textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells densely aggregated, smooth, cylindrical, subulate, straight, phialidic, simple, cylindrical, hyaline, smooth, 11–21 × 1–3 µm ( = 17 × 1.9 µm; n = 30), slightly tapered towards apex, apex with inconspicuous periclinal thickening. Alpha conidia hyaline, fusoid to ellipsoidal, asymmetrical, smooth-walled, 0–1-septate, tapering towards both ends, mostly straight, 5–9.5 × 1.5–3 µm ( = 7 × 2.2 µm; n = 30). Beta conidia hyaline, spindle-shaped, aseptate, smooth, apex subacutate, base slightly truncate, sometimes straight, mostly curved, 10–20 × 1–2.5 µm ( = 15 × 1.5 µm; n = 30). Gamma conidia not observed. Sexual morph: Not observed.
Material examined: China, Guizhou Province, Longli County, on leaves of Hypericum patulum, June 2021, S.Y. Wang (HGUP 416, holotype); culture ex-type GUCC 416.4, additional living culture: GUCC 416.14.
Culture characteristics: Colonies covering 9 cm diam. Petri dish after 2 weeks at 25 °C and a 12 h light/dark regime. On PDA, surface with thick, white uneven zonated aerial mycelium, exuding brown to dark orange conidial masses; reverse white to pale yellow to light brown, uneven zonated aerial mycelium and margin. On OA, surface with white or gray-white to pale yellow, fluffy, granular aerial mycelium, exuding white to dark green to black conidial masses; reverse white or beige to pale yellow.
Notes: The conidiomata of D. hyperici (3 mm diam.) are larger than those of D. caulivora (230–310 µm diam.) [60], but the alpha conidia of D. hyperici (5–9.5 × 1.5–3 µm) are shorter than those of D. caulivora (8.9–9.2 × 2.4–2.5 µm). Diaporthe caulivora produces a sexual morph with unitunicate asci, while D. hyperici has no known sexual morph. Diaporthe hyperici was phylogenetically distinct from the presently known species based on the DNA data (Figure 3). A comparison of the DNA bases (Table 7) showed significant difference between D. hyperici and sister species (D. caulivora). Based on its distinct morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results, D. hyperici was described here as a new species.
Gnomoniopsis rosae Crous, Persoonia 41: 305 (2018).
MycoBank Number: MB 828203, Figure 12.
Description: Life style: Parasitic, leaves of Rosa sp. (Rosaceae). Asexual morph: Conidiomata erumpent, separated, immersed or superficial, globose to depressed, initially appearing deep brown to black, slowly oozing transparent white or pale-yellow oily spheres with age, up to 600 µm diam., 5–7 wall layers of olive brown to brown textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells lining the inner cavity, hyaline, smooth, subcylindrical, branched at base or not, frequently branched above, simple, tapering, hyaline, smooth, subcylindrical, tapering towards apex, 6–15 × 1–3.5 µm ( = 11 × 2 µm; n = 20). Conidia solitary, aseptate, fusoid, hyaline, asymmetrical, guttulate, smooth-walled, rounded to acute apex, 6–12 × 2–4 µm ( = 8.5 × 3 µm; n = 30). Sexual morph: Unknown.
Materials examined: China, Guizhou Province, Wengan County, on leaves of Rosa sp. (Rosaceae), June 2021, S.Y. Wang (HGUP 408, living cultures GUCC 408.7 and GUCC 408.17) (new country record).
Culture characteristics: Colonies cultured at 25 °C and a 12 h light/dark regime for 2 weeks on PDA 60–85 mm diam., forming a circle of transparent mycelium in the center, followed by a circle of white or gray-white thick ridges, then uneven zonated aerial mycelium, slightly imbricated, thick, initially appearing white to pale yellow, slowly turning olive-gray with age outside the two concentric rings, with an uneven edge; reverse transparent to white or olive and white uneven imbricated zonated to white or light brown uneven edge. Colonies on OA covering the whole dish, pale white or light gray-white, flat surface, exuding deep brown to black conidial masses, slowly oozing transparent white or pale-yellow oily spheres with age, reverse pale white or light gray-white.
Notes: Gnomoniopsis represented a genus of mostly host-specific fungi [61,62]. Gnomoniopsis rosae (GUCC 408.7 and GUCC 408.17) was phylogenetically identical to the ex-type strain (CBS 145085) isolated by Crous et al. [31] in ITS, LSU, and rpb2, and we also supplemented the DNA sequences of this species with tef1 and tub2 genes. The DNA base comparison results are shown in Table 7; there were no DNA base differences among several genes. The isolates of G. rosae were newly recorded for China based on their morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results.
Coniella quercicola (Oudem.) L.V. Alvarez and Crous, Studies in Mycology 85: 27 (2016).
MycoBank Number: MB 817831, Figure 13.
Description: Life style: Parasitic, leaves of Hypericum patulum, Aralia chinensis and Acer palmatum. Asexual morph: Conidiomata pycnidial, separated, immersed or superficial, globose to depressed, initially appearing hyaline or light yellow becoming deep brown to black with age, up to 300 µm diam., 2–5 wall layers of dark brown textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells central cushion of hyaline cells, densely aggregated, slightly thicker, subulate, simple, frequently branched above, tapering, hyaline, smooth, 12–22 × 3–4.5 µm ( = 17 × 3.5 µm; n = 15), surrounded by a gelatinous coating, apex with visible periclinal thickening. Conidia hyaline, asymmetrical, smooth-walled, cylindrical, slightly curved to naviculate, aseptate, rounded to acute apex, tapered to a subtruncate base, germ slits absent, 10–23 × 2.5–4.5 µm ( = 17 × 3.5 µm; n = 30). Sexual morph: Unknown.
Materials examined: China, Guizhou Province, Longli and Wengan counties, on leaves of Hypericum patulum, Aralia chinensis and Acer palmatum, June 2021, S.Y. Wang (HGUP 414, HGUP 412 and HGUP 405, living cultures GUCC 414.2, GUCC 414.21, GUCC 412.3, GUCC 405.6 and GUCC 405.16). (new host and country record).
Culture characteristics: Colonies covering 9 cm diam. Petri dish after 2 weeks at 25 °C and a 12 h light/dark regime; spreading with sparse aerial mycelium and uneven zonation. On PDA, surface with thin, white uneven zonated aerial mycelium, distinctly imbricated like a flower, producing a few pale yellow to dark brown to black conidial masses; reverse with white and pale brown, uneven zonated, imbricated aerial mycelium. On OA, surface with white or gray-white to pale yellow, fluffy, granular aerial mycelium, producing pale yellow to dark brown to black conidial masses from the center; reverse white or beige to pale yellow.
Notes: Coniella quercicola was originally described as Macroplodia quercicola on the leaves of Quercus robur collected in the Netherlands. It was described as having pale-brown, cylindrical conidia, 24 × 4 µm [63]. Our strains (GUCC 414.2, GUCC 414.21, GUCC 412.3, GUCC 405.6, and GUCC 405.16) are phylogenetically identical to the neotype culture (CBS 904.69) of C. quercicola isolated and designated by Alvarez et al. [64] in ITS, LSU, and tef1 genes (Figure 14). According to the results of the DNA base comparison (Table 7), we note that base differences almost only occur in the tef1 region. The identification of C. quercicola was based on its morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test results.
4. Discussion and Conclusions
Families, genera, and species within Diaporthales are now characterized and separated based on a combination of morphology and molecular data [12,29,30,64,65,66,67,68,69,70,71,72]. The present study described and illustrated nine species (within five genera) of Diaporthales isolated from various host plants in Guizhou Province, China, including Gnomoniopsis mostly as host-specific fungi [61,62,65,73]. Based on their unique morphological characteristics, DNA phylogeny, DNA base differences, and pairwise homoplasy index (PHI) test evaluations, we described one new genus, seven new species, and two new fungal records for China. Only asexual morphology was observed for all the taxa described in this paper.
Foliocryphiaceae (Diaporthales) was established by Jiang et al. [1] based on the type genus Foliocryphia [54] and two allied genera, Chrysofolia [53] and Neocryphonectria [1]. Chrysofolia and Foliocryphia were originally placed in the family Cryphonectriaceae but they were transferred to Foliocryphiaceae by Jiang et al. [1]. Species of Chrysofolia usually exude a yellow slimy mass of conidia from a globose pycnidium with an immersed base. Only two species are listed in MycoBank (www.mycobank.org; accessed on 8 October 2022), Ch. colombiana [53], a pathogen of Eucalyptus urophylla from Colombia, and Ch. barringtoniae [55], an endophyte of Barringtonia acutangula from Thailand. Chrysofolia coriariae sp. nov. observed in the present study represents the first taxon of Chrysofolia in China.
Diaporthe is a large genus in Diaporthaceae with 1168 epithets listed in Index Fungorum (http://www.indexfungorum.org/; accessed on 4 July 2022) but only one-fifth of these taxa have been studied with molecular data [73,74,75]. The sexual morph of Diaporthe is characterized by immersed perithecial ascomata and an erumpent pseudostroma with more or less elongated perithecial necks; unitunicate clavate to cylindrical asci; and fusoid, ellipsoid to cylindrical, septate or aseptate, hyaline ascospores, which are biseriately to uniseriately arranged in the ascus, sometimes having appendages [29,30,76]. The asexual morph is characterized by ostiolate conidiomata, with cylindrical phialides producing three types of hyaline, aseptate conidia [76,77]. Type I α-conidia are hyaline, fusiform, straight, guttulate, or eguttulate; aseptate; and smooth-walled. Type II β-conidia are hyaline, filiform, straight or hamate, aseptate, smooth-walled, and eguttulate. Type III γ-conidia are rarely produced, and are hyaline, multiguttulate, and fusiform to subcylindrical with an acute or rounded apex, while the bases are sometimes truncate. Five new taxa of Diaporthe were introduced, which indicates that more potential novel and known taxa in this genus could be discovered because of the rich biodiversity in Guizhou Province.
Gnomoniaceae is a large family within Diaporthales, containing 38 accepted genera [65,78,79,80,81]. Among them, Gnomoniopsis is a well-delimited genus inhabiting the leaves, branches, and fruits of hosts in three families: Fagaceae, Onagraceae, and Rosaceae [62,65,73]. The sexual morph of Gnomoniaceae is characterized by ascomata that are generally immersed, solitary, or aggregated in an undeveloped stroma [6,61]. The perithecia are dark brown to black and pseudoparenchymatous with central, eccentric, or lateral necks [6,61]. The asci usually have an inconspicuous or distinct apical ring. Ascospores are generally small, hyaline, and uniseptate. The asexual morph is characterized by acervular or pycnidial conidiomata, phialidic conidiogenous cells, and non-septate conidia [82]. Gnomoniopsis rosae in our study was isolated as asexual morph from Rosa sp. and was newly recorded for China.
The family Schizoparmeaceae (Diaporthales) was introduced by Rossman et al. [6]. Historically, the family consisted of three genera, two of which only produce asexual morphs (Coniella and Pilidiella), while one can produce sexual morphs (Schizparme) [6]. This family was reassessed by Alvarez et al. [64], who proposed that Pilidiella is a taxonomic synonym of Coniella. Coniella was erected by Höhnel [83] and typified by C. pulchella, [84] who separated the genus into Euconiella (with dark conidia) and Pseudoconiella (with pale conidia) [64]. The key characteristics of Coniella are erumpent, brown, or black ascomata or conidiomata that later become superficial and an irregularly thickened peridium with plate-like ornamentation and one-celled ascospores, initially hyaline and later becoming pale to dark brown [30]. The present isolates of C. quercicola represent a new country record for China and new host records.
The molecular data provided evidence that our new genus belongs to Foliocryphiaceae, although in morphology it is similar to Mastigosporella in Mastigosporellaceae. In this molecular era, morphological conclusions are increasingly being reduced to a subordinate or even insignificant position. Thus, we accepted the phylogenetic conclusion to create the monotypic genus Pseudomastigosporella. Despite this, we still require additional strains of Diaporthales in order to compare the genome-wide information of members in this order due to the high level of similarities in morphology but measurable differences in molecular data. Currently, there are too few data for the adequate comparison of fungi.
Author Contributions
Sampling, molecular biology analysis, fungal isolation, description and phylogenetic analysis, genome sequencing, assembly and gene annotation: S.-Y.W.; writing original draft preparation: S.-Y.W.; writing–review, editing and producing the final version: S.-Y.W., Y.W., E.H.C.M., A.J.L.P. and Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research is supported by the following projects: National Natural Science Foundation of China (Nos. 31972222, 31660011), Program of Introducing Talents of Discipline to Universities of China (111 Program, D20023), Talent project of Guizhou Science and Technology Cooperation Platform ([2017]5788-5, [2019]5641, [2019]13), Guizhou Science, Technology Department of International Cooperation Base project ([2018]5806), the project of Guizhou Provincial Education Department ([2020]001), Guizhou Science and Technology Innovation Talent Team Project ([2020]5001), the Open Project of Key Laboratory of Plant Resources Conservation and Germplasm Innovation in Mountainous Region (Ministry of Education) (MOELP-201801), and Postgraduate Education Innovation Program in Guizhou Province (YJSKYJJ [2021] 058). Alan J. L. Phillips acknowledges the support from UIDB/04046/2020 and UIDP/04046/2020 Centre grants from FCT, Portugal (to BioISI).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and/or are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare that they have no competing interest.
References
- Jiang, N.; Fan, X.L.; Tian, C.M.; Crous, P.W. Reevaluating Cryphonectriaceae and allied families in Diaporthales. Mycologia 2020, 112, 267–292. [Google Scholar] [CrossRef] [PubMed]
- Barr, M.E. The Diaporthales in North America: With emphasis on Gnomonia and its segregates. Mycol. Mem. 1978, 7, 1–232. [Google Scholar]
- Alexopoulus, C.J.; Mims, C.W. Introductory Mycology, 3rd ed.; Wiley: New York, NY, USA, 1978. [Google Scholar]
- Castlebury, L.A.; Rossman, A.Y.; Jaklitsch, W.J.; Vasilyeva, L.N. A preliminary overview of the Diaporthales based on large subunit nuclear ribosomal DNA sequences. Mycologia 2002, 94, 1017–1031. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Kirk, P.M.; Sutton, B.C.; Pegler, D.N. Ainsworth & Bisby’s Dictionary of the Fungi, 8th ed.; CAB International: Oxford, UK, 1995. [Google Scholar]
- Rossman, A.Y.; Farr, D.F.; Castlebury, L.A. A review of the phylogeny and biology of the Diaporthales. Mycoscience 2007, 48, 135–144. [Google Scholar] [CrossRef]
- Qi, P.K.; Jiang, Z.D.; Xiang, M.M. Flora Fungorum Sinicorum: Phomopsis; Science Press: Beijing, China, 2007; Volume 34, pp. 1–185. [Google Scholar]
- Fu, C.H.; Hsieh, H.M.; Chen, C.Y.; Chang, T.T.; Huang, Y.M.; Ju, Y.M. Ophiodiaporthe cyatheae gen. et sp. nov., a diaporthalean pathogen causing a devastating wilt disease of Cyathea lepifera in Taiwan. Mycologia 2013, 105, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.Q.; Wijayawardene, N.N.; Bhat, D.J.; Chukeatirote, E.; Bahkali, A.H.; Zhao, R.L.; Xu, J.C.; Hyde, K.D. Pustulomyces gen. nov. accommodated in Diaporthaceae, Diaporthales, as revealed by morphology and molecular analyses. Cryptogam. Mycol. 2014, 35, 63–72. [Google Scholar] [CrossRef]
- Du, Z.; Hyde, K.D.; Yang, Q.; Liang, Y.M.; Tian, C.M. Melansporellaceae: A novel family of Diaporthales (Ascomycota). Phytotaxa 2017, 305, 191–200. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Guarnaccia, V.; Tian, C.M. High diversity of Diaporthe species associated with dieback diseases in China, with twelve new species described. MycoKeys 2018, 39, 97–149. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Bezerra, J.D.; Tian, C.M.; Crous, P.W. Families and genera of diaporthalean fungi associated with canker and dieback of tree hosts. Persoonia 2018, 40, 119–134. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Yang, Q.; Fan, X.; Tian, C. Micromelanconis kaihuiae gen. et sp. nov. a new diaporthalean fungus from Chinese chestnut branches in southern China. MycoKeys 2021, 79, 1–16. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Promputtha, I.; Liu, J. Dendrostoma covidicola sp. nov. (Erythrogloeaceae, Diaporthales) on Fagus sylvatica from Sichuan Province, China. Phytotaxa 2021, 483, 85–94. [Google Scholar] [CrossRef]
- Dong, Z.; Manawasinghe, I.S.; Huang, Y.; Shu, Y.; Luo, M. Endophytic Diaporthe associated with Citrus grandis cv. Tomentosa in China. Front. Microbiol. 2021, 11, 609387. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.T.; Xia, J.W.; Pan, Y.; Zhu, C.Y.; Li, Z. Morphological and molecular description of Diaporthe nannuoshanensis sp. nov. (Diaporthaceae, Diaporthales) in south-western China. Nova Hedwig. 2021, 112, 399–411. [Google Scholar] [CrossRef]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.; Liu, X.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Santos, L.; Phillips, A.J.L.; Crous, P.W.; Alves, A. Diaporthe species on Rosaceae with descriptions of D. pyracanthae sp. nov. and D. malorum sp. nov. Mycosphere 2017, 8, 485–511. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Rehner, S.A.; Samuels, G.J. Molecular systematics of the Hypocreales: A teleomorph gene phylogeny and the status of their anamorphs. Can. J. Bot. 1990, 73, 816–823. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G. Development of primer sets designed for use with PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Crous, P.W.; Groenewald, J.Z.; Maharachchikumbura, S.S.; Jeewon, R.; Phillips, A.J.; Bhat, J.D.; Perera, R.H.; Li, Q.R.; Li, W.J.; et al. Families of Diaporthales based on morphological and phylogenetic evidence. Stud. Mycol. 2017, 86, 217–296. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Jeewon, R.; Chomnunti, P.; Wanasinghe, D.N.; Norphanphoun, C.; Karunarathna, A.; Pem, D.; Perera, R.H.; Camporesi, E.; McKenzie, E.H.C.; et al. Taxonomic circumscription of Diaporthales based on multigene phylogeny and morphology. Fungal Divers. 2018, 93, 241–443. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-Ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal planet description sheets 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.; Huang, S.K.; Abdel-Wahab, M.A.; Daranagama, D.A.; Dayarathne, M.; D’souza, M.J.; Goonasekara, I.D.; et al. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Huang, S.-K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; et al. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Groenewald, J.Z.; Woodhall, J.; Armengol, J.; Cinelli, T.; Eichmeier, A.; Ezra, D.; Fontaine, F.; Gramaje, D.; Gutierrez-Aguirregabiria, A.; et al. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 2018, 40, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Raudabaugh, D.B.; Iturriaga, T.; Carver, A.; Mondo, S.; Pangilinan, J.; Lipzen, A.; He, G.; Amirebrahimi, M.; Grigoriev, I.V.; Miller, A.N. Coniella lustricola, a new species from submerged detritus. Mycol. Prog. 2018, 17, 191–203. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Kuo, C.-H.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I.; et al. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Sym. Series 1999, 41, 95–98. [Google Scholar]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 22, 3276–3278. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceeding of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Mark, V.D.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY: A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rannala, B.; Yang, Z.H. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree-Version 1.4.3. A Graphical Viewer of Phylogenetic Trees. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 5 October 2016).
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Hespanhol, L.; Vallio, C.S.; Costa, L.M.; Saragiotto, B.T. Understanding and interpreting confidence and credible intervals around effect estimates. Braz. J. Phys. Ther. 2019, 23, 290–301. [Google Scholar] [CrossRef]
- Bruen, T.C.; Philippe, H.; Bryant, D. A simple and robust statistical test for detecting the presence of recombination. Genetics 2006, 172, 2665–2681. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H. SplitsTree: Analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Restrepo, M.H.; Sutton, D.A.; Acharya, K.; Barber, P.; Boekhout, T.; Dimitrov, R.A.; Dueñas, M.; et al. Fungal planet description sheets: 320–370. Persoonia 2015, 34, 167–266. [Google Scholar] [CrossRef]
- Cheewangkoon, R.; Groenewald, J.Z.; Summerell, B.A.; Hyde, K.D.; To-anun, C.; Crous, P.W. Myrtaceae, a cache of fungal biodiversity. Persoonia 2009, 23, 55–85. [Google Scholar] [CrossRef] [Green Version]
- Suwannarach, N.; Kumla, J.; Sri-Ngernyuang, K.; Lumyong, S. A new endophytic fungus, Chrysofolia barringtoniae sp. nov. from Thailand. Mycoscience 2016, 57, 361–365. [Google Scholar] [CrossRef]
- Chang, C.Q.; Xi, P.G.; Xiang, M.M.; Jiang, Z.D.; Chi, P.K. New species of Phomopsis on woody plants in Hunan province. Mycosystema 2005, 24, 145–154. [Google Scholar]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Burgess, T.I.; Groenewald, J.Z. Fungal planet description sheets: 107–127. Persoonia 2012, 28, 138–182. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; Crous, P.W.; McKenzie, E.H.C.; Chukeatirote, E.; Hyde, K.D. A multi-locus phylogenetic evaluation of Diaporthe (Phomopsis). Fungal Divers. 2012, 56, 157–171. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Restrepo, M.H.; Sutton, D.A.; Acharya, K.; Barber, P.; Boekhout, T.; Dimitrov, R.A.; Dueñas, M.; et al. Fungal Planet Description Sheets: 281–319. Persoonia 2014, 33, 212–289. [Google Scholar] [CrossRef]
- Santos, J.; Vrandečić, K.; Ćosić, J.; Duvnjak, T.; Phillips, A. Resolving the Diaporthe species occurring on soybean in Croatia. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef]
- Sogonov, M.V.; Castlebury, L.A.; Rossman, A.Y.; Mejía, L.C.; White, J.F. Leaf-inhabiting genera of the Gnomoniaceae, Diaporthales. Stud. Mycol. 2008, 62, 1–77. [Google Scholar] [CrossRef]
- Walker, D.M.; Castlebury, L.A.; Rossman, A.Y.; Sogonov, M.V.; White, J.F. Systematics of genus Gnomoniopsis (Gnomoniaceae, Diaporthales) based on a three gene phylogeny, host associations and morphology. Mycologia 2010, 102, 1479–1496. [Google Scholar] [CrossRef]
- Saccardo, P.A.; Saccardo, D. Supplementum universale, Pars VII. Discomycetae-Deuteromycatae. Syll. Fung. 1906, 18, 1–838. [Google Scholar]
- Alvarez, L.V.; Groenewald, J.Z.; Crous, P.W. Revising the Schizoparmaceae: Coniella and its synonyms Pilidiella and Schizoparme. Stud. Mycol. 2016, 85, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Tian, C.M. An emerging pathogen from rotted chestnut in China: Gnomoniopsis daii sp. nov. Forests 2019, 10, 1016. [Google Scholar] [CrossRef] [Green Version]
- Voglmayr, H.; Castlebury, L.A.; Jaklitsch, W.M. Juglanconis gen. nov. on Juglandaceae, and the new family Juglanconidaceae (Diaporthales). Persoonia 2017, 38, 136–155. [Google Scholar] [CrossRef] [Green Version]
- Voglmayr, H.; Jaklitsch, W.M. Stilbosporaceae resurrected: Generic reclassification and speciation. Persoonia 2014, 33, 61–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voglmayr, H.; Rossman, A.Y.; Castlebury, L.A.; Jaklitsch, W.M. Multigene phylogeny and taxonomy of the genus Melanconiella (Diaporthales). Fungal Divers. 2012, 57, 1–44. [Google Scholar] [CrossRef]
- Suetrong, S.; Klaysuban, A.; Sakayaroj, J.; Preedanon, S.; Ruang-Areerate, P.; Phongpaichit, S.; Pang, K.-L.; Jones, E. Tiarisporellaceae, a new family in the order Diaporthales (Sordariomycetes, Ascomycota). Crypt. Mycol. 2015, 36, 319–330. [Google Scholar] [CrossRef]
- Norphanphoun, C.; Hongsanan, S.; Doilom, M.; Bhat, D.J.; Wen, T.-C.; Senanayake, I.C.; Bulgakov, T.; Hyde, K.D. Lamproconiaceae fam. nov. to accommodate Lamproconium desmazieri. Phytotaxa 2016, 270, 89–102. [Google Scholar]
- Yang, Q.; Fan, X.L.; Du, Z.; Tian, C.M. Diaporthosporellaceae, a novel family of Diaporthales (Sordariomycetes, Ascomycota). Mycoscience 2018, 59, 229–235. [Google Scholar] [CrossRef]
- Guterres, D.C.; Galvão-Elias, S.; Dos Santos, M.D.D.M.; de Souza, B.C.P.; de Almeida, C.P.; Pinho, D.B.; Miller, R.N.G.; Dianese, J.C. Phylogenetic relationships of Phaeochorella parinarii and recognition of a new family, Phaeochorellaceae (Diaporthales). Mycologia 2019, 111, 660–675. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, N.; Tian, C.M. Three new Diaporthe species from Shaanxi Province, China. MycoKeys 2020, 67, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.S.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef]
- Zapata, M.; Palma, M.A.; Aninat, M.J.; Piontelli, E. Polyphasic studies of new species of Diaporthe from native forest in Chile, with descriptions of Diaporthe araucanorum sp. nov., Diaporthe foikelawen sp. nov. and Diaporthe patagonica sp. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 3379–3390. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H.A.; Hyde, K.D. The genus Phomopsis: Biology, applications. species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejía, L.C.; Castlebury, L.A.; Rossman, A.Y.; Sogonov, M.V.; White, J.F. A systematic account of the genus Plagiostoma (Gnomoniaceae, Diaporthales) based on morphology, host-associations, and a four-gene phylogeny. Stud. Mycol. 2011, 68, 211–235. [Google Scholar] [CrossRef] [PubMed]
- Mejía, L.C.; Rossman, A.Y.; Castlebury, L.A.; White, J.F. New species, phylogeny, host-associations and geographic distribution of genus Cryptosporella (Gnomoniaceae, Diaporthales). Mycologia 2011, 103, 379–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejia, L.C.; Rossman, A.Y.; Castlebury, L.A.; Yang, Z.L.; White, J.F. Occultocarpon, a new monotypic genus of Gnomoniaceae on Alnus nepalensis from China. Fungal Divers. 2012, 52, 99–105. [Google Scholar] [CrossRef]
- Walker, D.M.; Castlebury, L.A.; Rossman, A.Y.; Mejía, L.C.; White, J.F. Phylogeny and taxonomy of Ophiognomonia (Gnomoniaceae, Diaporthales), including twenty-five new species in this highly diverse genus. Fungal Divers. 2012, 57, 85–147. [Google Scholar] [CrossRef]
- Monod, M. Taxonomic monograph of the Gnomoniaceae (Ascomycetes of the ordre Diaporthales). I. Beih. Zur Sydowia 1983, 9, 1–315. [Google Scholar]
- Höhnel, F.V. Dritte vorlaufige Mitteilung mycologischer Ergebnisse (Nr. 201–304). Ber. Der Dtsch. Bot. Ges. 1918, 36, 309–317. [Google Scholar]
- Petrak, F.; Sydow, H. Die Gattungen der Pyrenomyzeten, Sphaeropsideen and Melanconieen. I. Der phaeosporen Sphaeropsideen und die Gattung Macrophoma. Feddes Repert. Speciarum Nov. Regni Veg. Beih. 1927, 42, 1–551. [Google Scholar]
Figure 1.
Phylogram generated from RAxML analysis of a concatenated ITS-LSU-rpb2-tef1-tub2 sequence dataset to represent the phylogenetic relationships of taxa in Foliocryphiaceae and Cryphonectriaceae. The tree was rooted with Dwiroopa lythri (CBS 109755, ex-type strain) and Dw. punicae (CBS 143163, ex-type strain). Bootstrap support values for ML and MP equal to or greater than 70% and Bayesian posterior probabilities equal to or higher than 0.95 PP are indicated above the nodes as ML/MP/PP. Support values lower than 70% ML/MP and 0.95 PP are indicated by a hyphen (-). The newly generated sequences are indicated in red.
Figure 1.
Phylogram generated from RAxML analysis of a concatenated ITS-LSU-rpb2-tef1-tub2 sequence dataset to represent the phylogenetic relationships of taxa in Foliocryphiaceae and Cryphonectriaceae. The tree was rooted with Dwiroopa lythri (CBS 109755, ex-type strain) and Dw. punicae (CBS 143163, ex-type strain). Bootstrap support values for ML and MP equal to or greater than 70% and Bayesian posterior probabilities equal to or higher than 0.95 PP are indicated above the nodes as ML/MP/PP. Support values lower than 70% ML/MP and 0.95 PP are indicated by a hyphen (-). The newly generated sequences are indicated in red.

Figure 2.
Results of the pairwise homoplasy index (PHI) test of closely related species using both LogDet transformation and splits decomposition. Our strains are indicated in red, other involved strains are indicated in blue. (a) Foliocryphiaceae. (b–f) Diaporthe. (g) Gnomoniopsis. (h) Coniella.
Figure 2.
Results of the pairwise homoplasy index (PHI) test of closely related species using both LogDet transformation and splits decomposition. Our strains are indicated in red, other involved strains are indicated in blue. (a) Foliocryphiaceae. (b–f) Diaporthe. (g) Gnomoniopsis. (h) Coniella.

Figure 3.
Phylogram generated from RAxML analysis of a concatenated ITS-tef1-tub2 sequence dataset to represent the phylogenetic relationships of taxa in Diaporthe. The tree was rooted with Diaporthella corylina (CBS 121124, ex-type strain) and Di. cryptica (CBS 140348, ex-neotype strain). Bootstrap support values for ML and MP equal to or greater than 70% and Bayesian posterior probabilities equal to or higher than 0.95 PP are indicated above the nodes as ML/MP/PP. Support values lower than 70% ML/MP and 0.95 PP are indicated by a hyphen (-). The newly generated sequences are indicated in red.
Figure 3.
Phylogram generated from RAxML analysis of a concatenated ITS-tef1-tub2 sequence dataset to represent the phylogenetic relationships of taxa in Diaporthe. The tree was rooted with Diaporthella corylina (CBS 121124, ex-type strain) and Di. cryptica (CBS 140348, ex-neotype strain). Bootstrap support values for ML and MP equal to or greater than 70% and Bayesian posterior probabilities equal to or higher than 0.95 PP are indicated above the nodes as ML/MP/PP. Support values lower than 70% ML/MP and 0.95 PP are indicated by a hyphen (-). The newly generated sequences are indicated in red.

Figure 4.
Phylogram generated from RAxML analysis of a concatenated ITS-LSU-rpb2-tef1-tub2 sequence dataset to represent the phylogenetic relationships of taxa in Gnomoniopsis. The tree was rooted with Sirococcus tsugae (CBS 119626). Bootstrap support values for ML and MP equal to or greater than 70% and the Bayesian posterior probabilities equal to or higher than 0.95 PP are indicated above the nodes as ML/MP/PP. Support values lower than 70% ML/MP and 0.95 PP are indicated by a hyphen (-). The newly generated sequences are indicated in red.
Figure 4.
Phylogram generated from RAxML analysis of a concatenated ITS-LSU-rpb2-tef1-tub2 sequence dataset to represent the phylogenetic relationships of taxa in Gnomoniopsis. The tree was rooted with Sirococcus tsugae (CBS 119626). Bootstrap support values for ML and MP equal to or greater than 70% and the Bayesian posterior probabilities equal to or higher than 0.95 PP are indicated above the nodes as ML/MP/PP. Support values lower than 70% ML/MP and 0.95 PP are indicated by a hyphen (-). The newly generated sequences are indicated in red.

Figure 5.
Pseudomastigosporella guizhouensis (GUCC 406.6). Hosts: (a) Hypericum patulum; (b) Acer palmatum. (c) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (d) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (e) Mass of conidia. (f) Conidioma. (g,h) Conidiomata and conidiogenous cells. (i–l) Conidia. Scale bars: (f–i) = 50 µm; (j–l) = 10 µm.
Figure 5.
Pseudomastigosporella guizhouensis (GUCC 406.6). Hosts: (a) Hypericum patulum; (b) Acer palmatum. (c) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (d) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (e) Mass of conidia. (f) Conidioma. (g,h) Conidiomata and conidiogenous cells. (i–l) Conidia. Scale bars: (f–i) = 50 µm; (j–l) = 10 µm.

Figure 6.
Chrysofolia coriariae (GUCC 416.4). (a,b) Host: Coriaria nepalensis. (c) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (d) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (e,f) Conidial masses. (g,h) Conidiomata. (i,j) Conidiogenous cells. (k,l) Conidia. Scale bars: (g–i) = 50 µm; (j–l) = 10 µm.
Figure 6.
Chrysofolia coriariae (GUCC 416.4). (a,b) Host: Coriaria nepalensis. (c) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (d) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (e,f) Conidial masses. (g,h) Conidiomata. (i,j) Conidiogenous cells. (k,l) Conidia. Scale bars: (g–i) = 50 µm; (j–l) = 10 µm.

Figure 7.
Diaporthe juglandigena (GUCC 422.16). (a) Host: Juglans regia. (b) Colony on PDA after 2 wk (left: above, right: reverse). (c) Colony on OA after 2 wk (left: above, right: reverse). (d,e) Mass of conidia. (f,g) Conidiogenous cells. (h,i) Alpha conidia. (j,k) Beta conidia. Scale bars: (f–k) = 10 µm.
Figure 7.
Diaporthe juglandigena (GUCC 422.16). (a) Host: Juglans regia. (b) Colony on PDA after 2 wk (left: above, right: reverse). (c) Colony on OA after 2 wk (left: above, right: reverse). (d,e) Mass of conidia. (f,g) Conidiogenous cells. (h,i) Alpha conidia. (j,k) Beta conidia. Scale bars: (f–k) = 10 µm.

Figure 8.
Diaporthe eucommiigena (GUCC 420.9). (a,b) Host: Eucommia ulmoides. (c) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (d) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (e–g) Mass of conidia. (h) Conidiogenous cells. (i,j) Alpha conidia. (k,l) Beta conidia. (m,n) Gamma conidia. Scale bars: (h–n) = 10 µm.
Figure 8.
Diaporthe eucommiigena (GUCC 420.9). (a,b) Host: Eucommia ulmoides. (c) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (d) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (e–g) Mass of conidia. (h) Conidiogenous cells. (i,j) Alpha conidia. (k,l) Beta conidia. (m,n) Gamma conidia. Scale bars: (h–n) = 10 µm.

Figure 9.
Diaporthe dejiangensis (GUCC 421.2). (a) Host: Juglans regia. (b) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (c) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (d–f) Mass of conidia. (g,h) Conidiogenous cells. (i,j) Alpha conidia. Scale bars: (g–j) = 10 µm.
Figure 9.
Diaporthe dejiangensis (GUCC 421.2). (a) Host: Juglans regia. (b) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (c) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (d–f) Mass of conidia. (g,h) Conidiogenous cells. (i,j) Alpha conidia. Scale bars: (g–j) = 10 µm.

Figure 10.
Diaporthe tongrensis (GUCC 421.10). (a) Host: Juglans regia. (b) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (c) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (d,e) Mass of conidia. (f,g) Conidiogenous cells. h Alpha conidia. (i,j) Beta conidia. Scale bars: (f–j) = 10 µm.
Figure 10.
Diaporthe tongrensis (GUCC 421.10). (a) Host: Juglans regia. (b) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (c) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (d,e) Mass of conidia. (f,g) Conidiogenous cells. h Alpha conidia. (i,j) Beta conidia. Scale bars: (f–j) = 10 µm.

Figure 11.
Diaporthe hyperici (GUCC 414.4). (a,b) Host: Hypericum patulum. (c) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (d) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (e,f) Mass of conidia. (g–i) Conidiogenous cells. (j) Alpha conidia. (k) Alpha and beta conidia. (l,m) Beta conidia. Scale bars: (g,h) = 50 µm; (i–m) = 10 µm.
Figure 11.
Diaporthe hyperici (GUCC 414.4). (a,b) Host: Hypericum patulum. (c) Colony on PDA after 2 wk at 25 °C (left: above, right: reverse). (d) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (e,f) Mass of conidia. (g–i) Conidiogenous cells. (j) Alpha conidia. (k) Alpha and beta conidia. (l,m) Beta conidia. Scale bars: (g,h) = 50 µm; (i–m) = 10 µm.

Figure 12.
Gnomoniopsis rosae (GUCC 408.7). (a) Host: Rose sp. (b) Colonies on PDA after 2 wk at 25 °C (left: above, right: reverse). (c) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (d) Mass of conidia. (e) Conidiomata. (f,g) Conidiogenous cells. (h–j) Conidia. Scale bars: (e–j) = 10 µm.
Figure 12.
Gnomoniopsis rosae (GUCC 408.7). (a) Host: Rose sp. (b) Colonies on PDA after 2 wk at 25 °C (left: above, right: reverse). (c) Colony on OA after 2 wk at 25 °C (left: above, right: reverse). (d) Mass of conidia. (e) Conidiomata. (f,g) Conidiogenous cells. (h–j) Conidia. Scale bars: (e–j) = 10 µm.

Figure 13.
Coniella quercicola (GUCC 412.3). Hosts. (a) Acer palmatum; (b) Aralia chinensis; (c) Hypericum patulum. (d) Colony on PDA after 2 wk 25 °C (left: above, right: reverse). (e) Colony on OA after 2 wk 25 °C (left: above, right: reverse). (f) Mass of conidia. (g) Conidioma. (h–j) Conidiogenous cells. (k–n) Conidia. Scale bars: (g) = 50 µm; (h–n) = 10 µm.
Figure 13.
Coniella quercicola (GUCC 412.3). Hosts. (a) Acer palmatum; (b) Aralia chinensis; (c) Hypericum patulum. (d) Colony on PDA after 2 wk 25 °C (left: above, right: reverse). (e) Colony on OA after 2 wk 25 °C (left: above, right: reverse). (f) Mass of conidia. (g) Conidioma. (h–j) Conidiogenous cells. (k–n) Conidia. Scale bars: (g) = 50 µm; (h–n) = 10 µm.

Figure 14.
Phylogram generated from RAxML analysis of a concatenated ITS-LSU-tef1 sequence dataset to represent the phylogenetic relationships of taxa in Coniella. The tree was rooted with C. fragariae (CBS 172.49, ex-type strain) and C. nigra (CBS 165.60, ex-type strain). Bootstrap support values for ML and MP equal to or greater than 70% and the Bayesian posterior probabilities equal to or higher than 0.95 PP are indicated above the nodes as ML/MP/PP. Support values lower than 70% ML/MP and 0.95 PP are indicated by a hyphen (-). The newly generated sequences are indicated in red.
Figure 14.
Phylogram generated from RAxML analysis of a concatenated ITS-LSU-tef1 sequence dataset to represent the phylogenetic relationships of taxa in Coniella. The tree was rooted with C. fragariae (CBS 172.49, ex-type strain) and C. nigra (CBS 165.60, ex-type strain). Bootstrap support values for ML and MP equal to or greater than 70% and the Bayesian posterior probabilities equal to or higher than 0.95 PP are indicated above the nodes as ML/MP/PP. Support values lower than 70% ML/MP and 0.95 PP are indicated by a hyphen (-). The newly generated sequences are indicated in red.


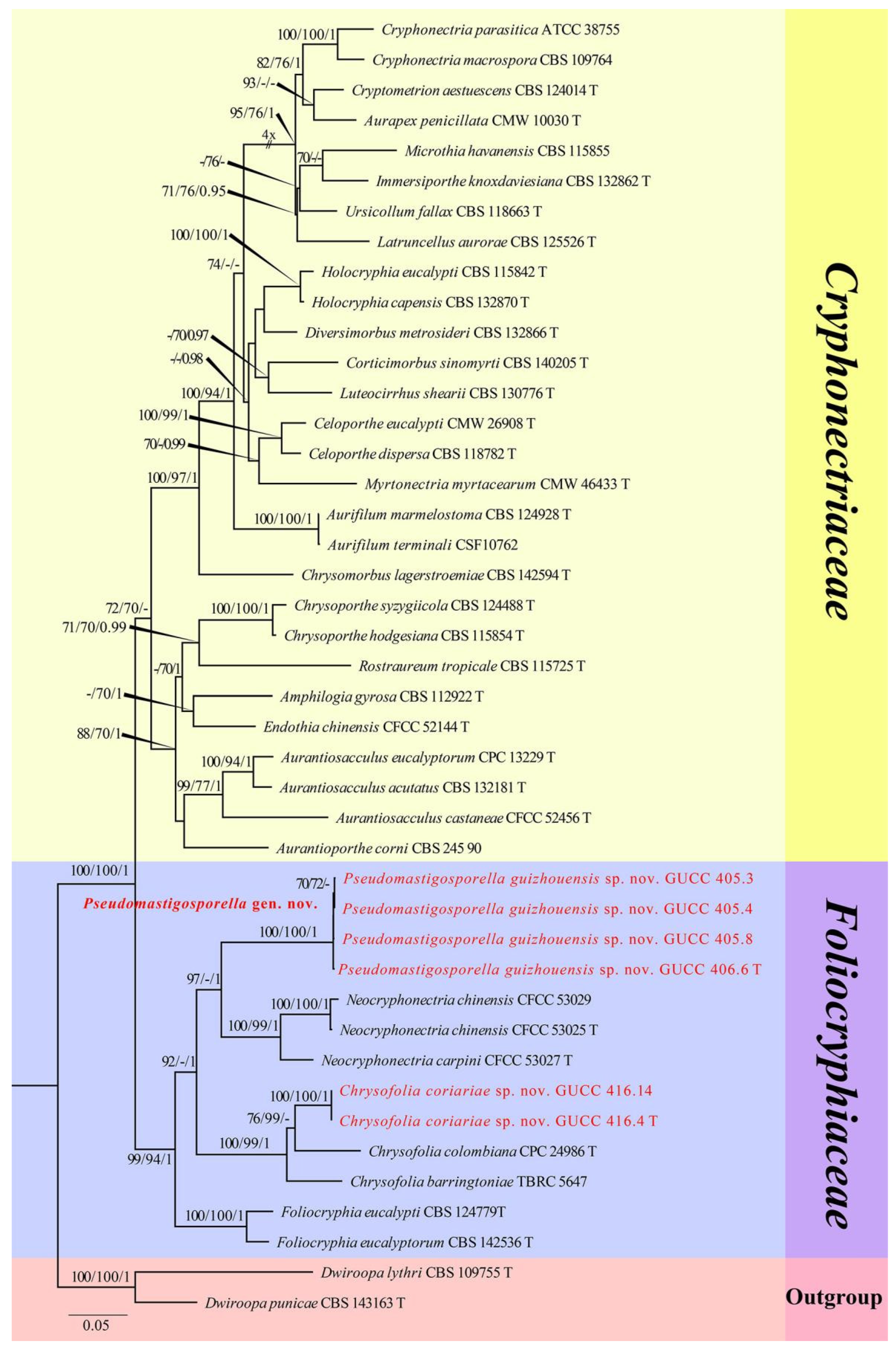{kind=link}
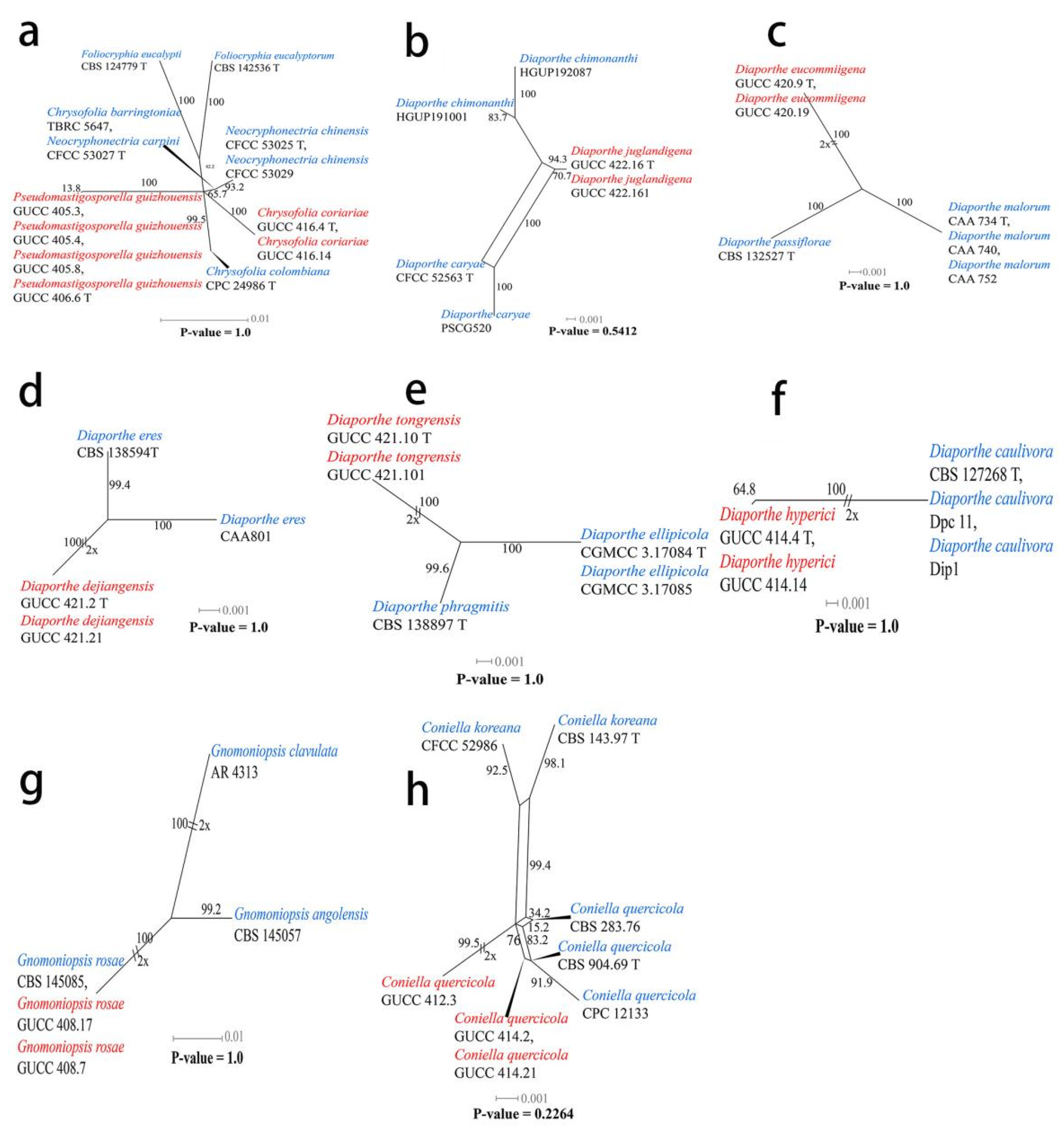{kind=link}
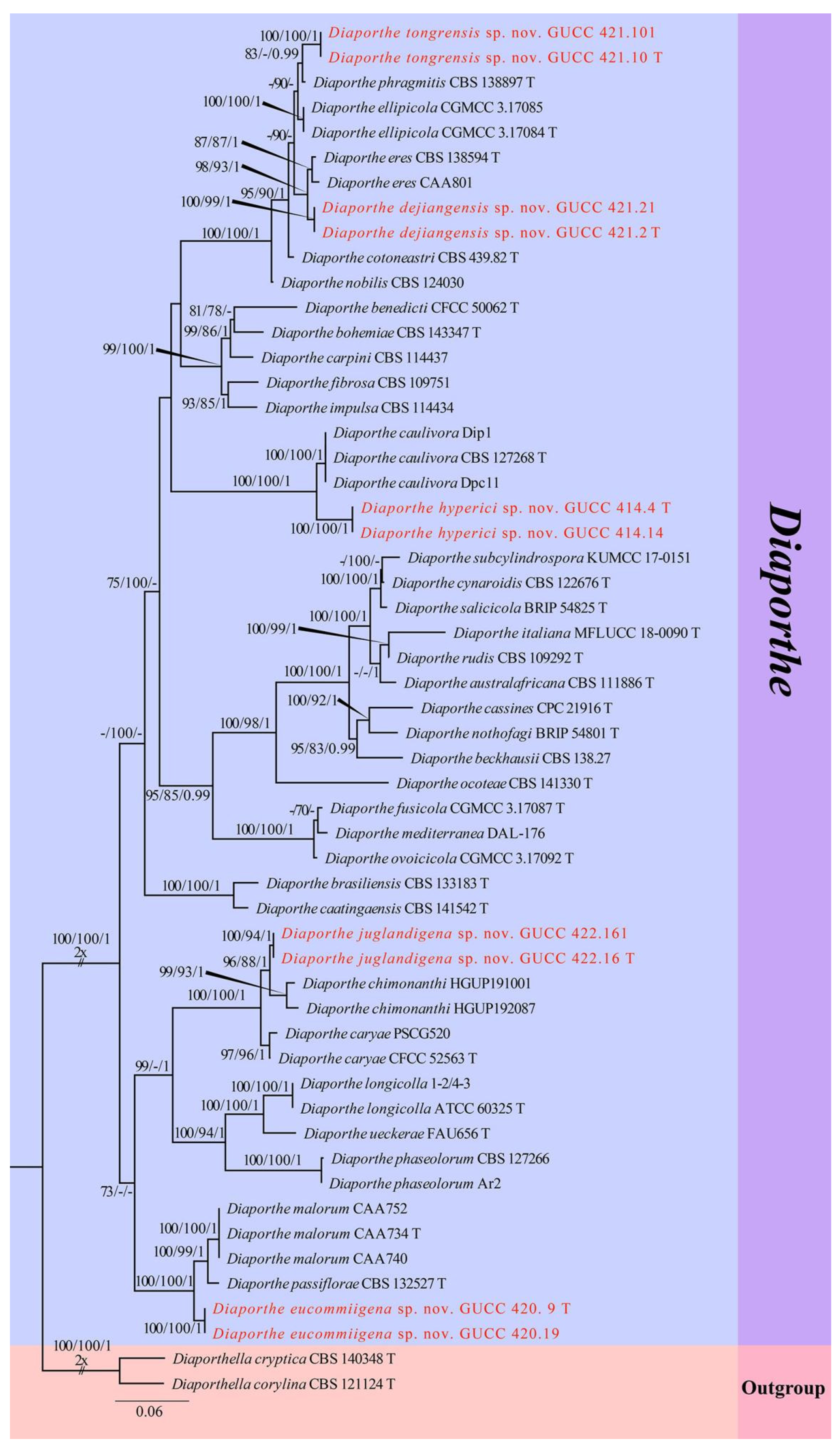{kind=link}
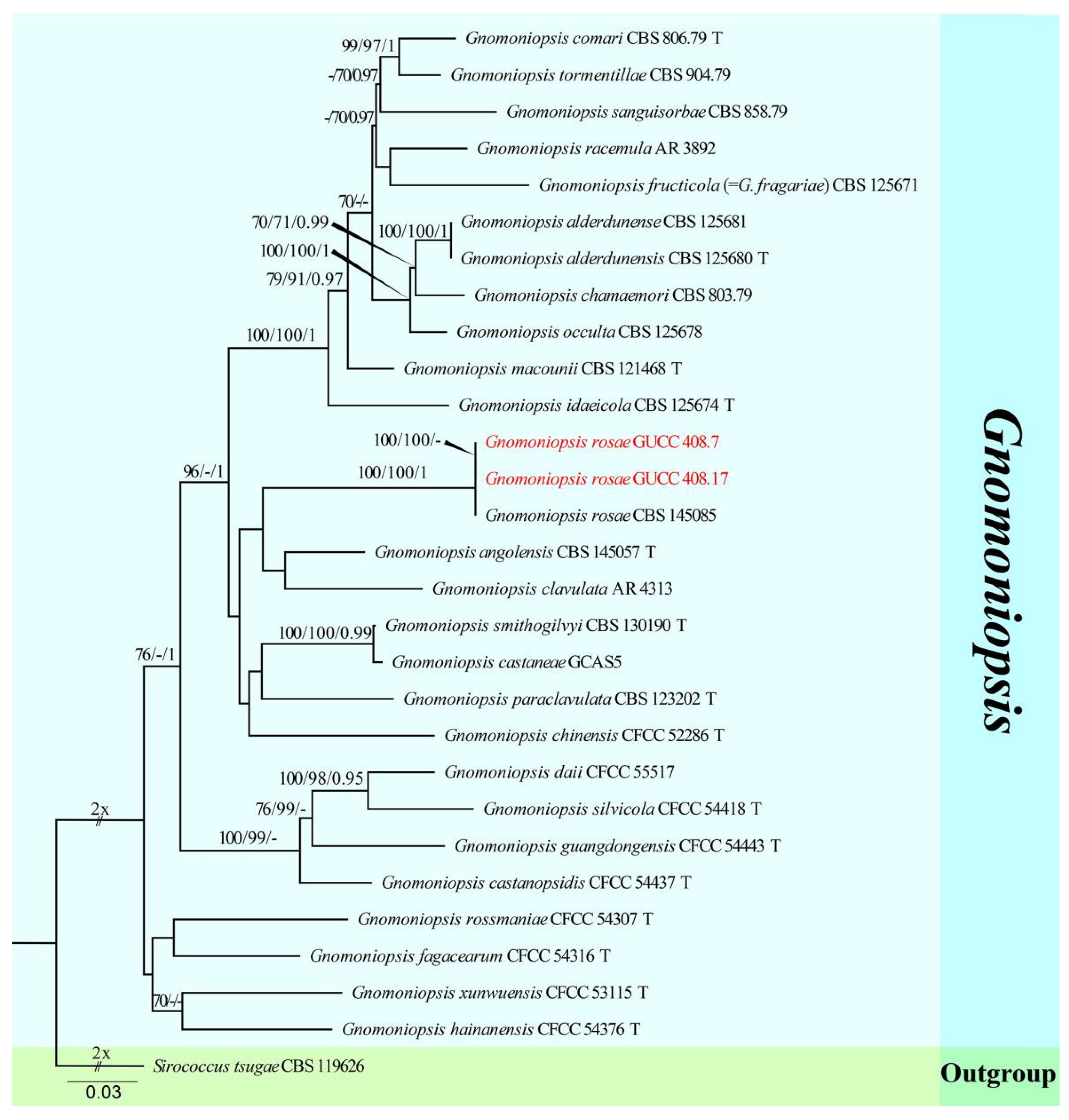{kind=link}
{kind=link}
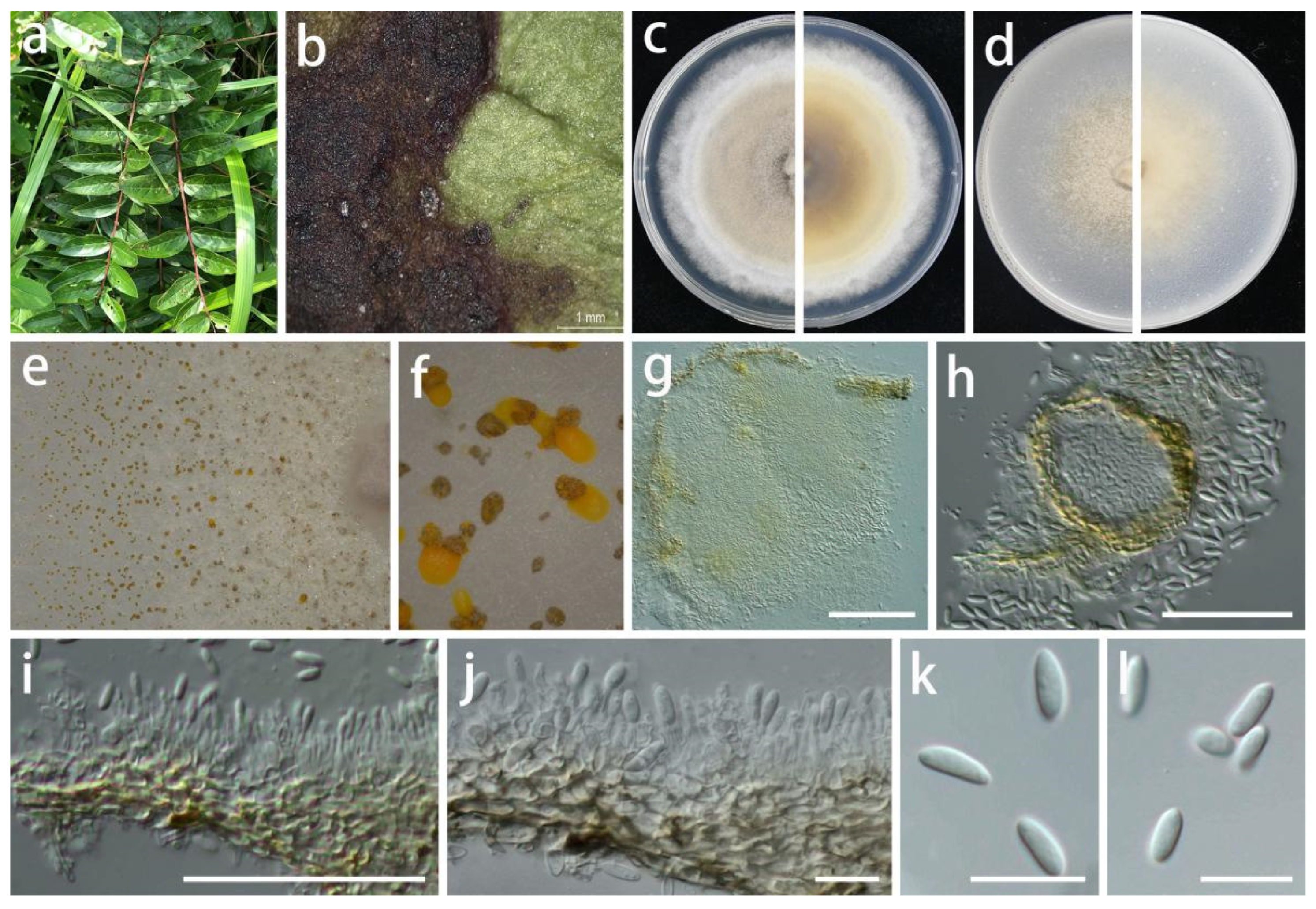{kind=link}
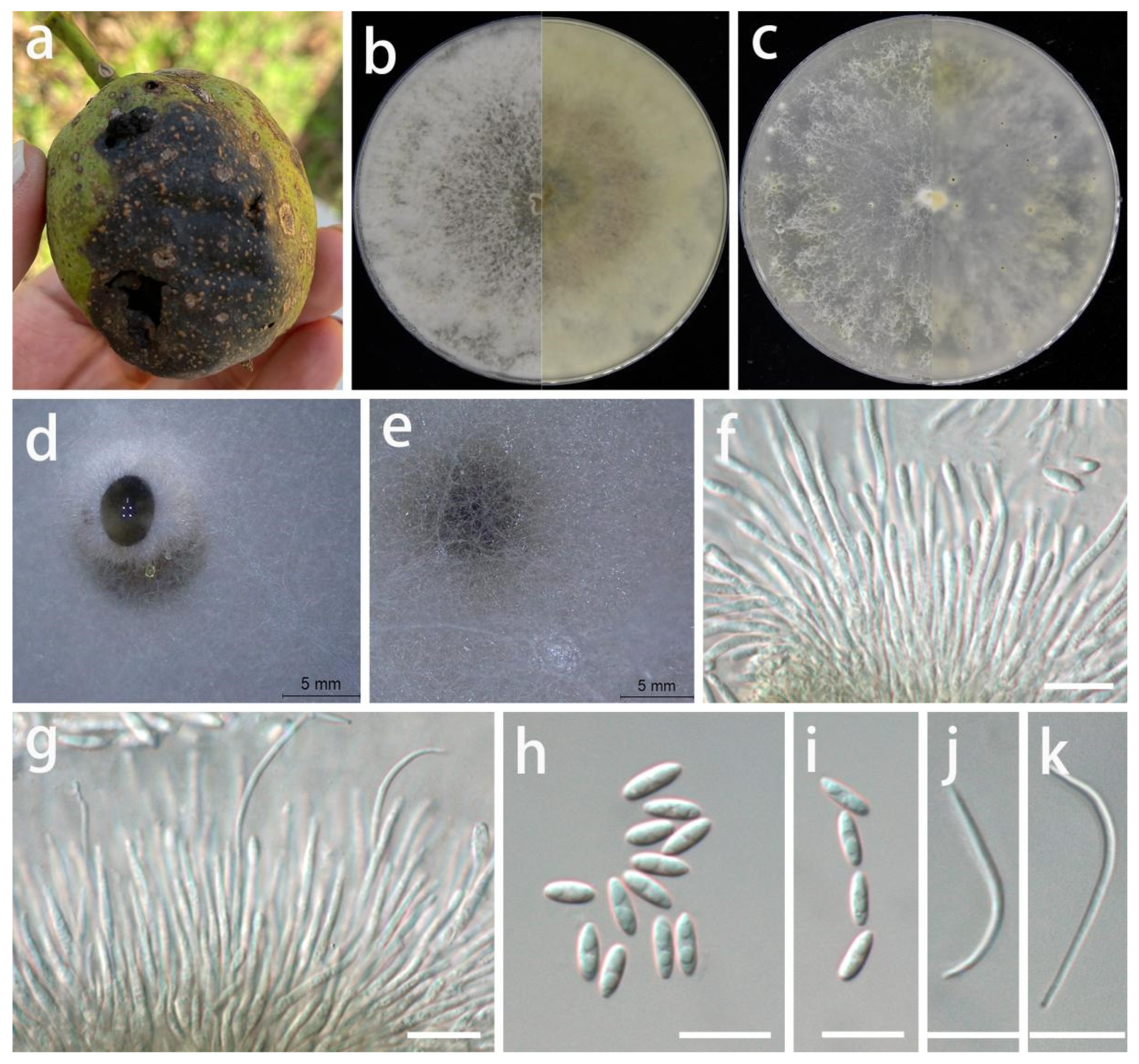{kind=link}
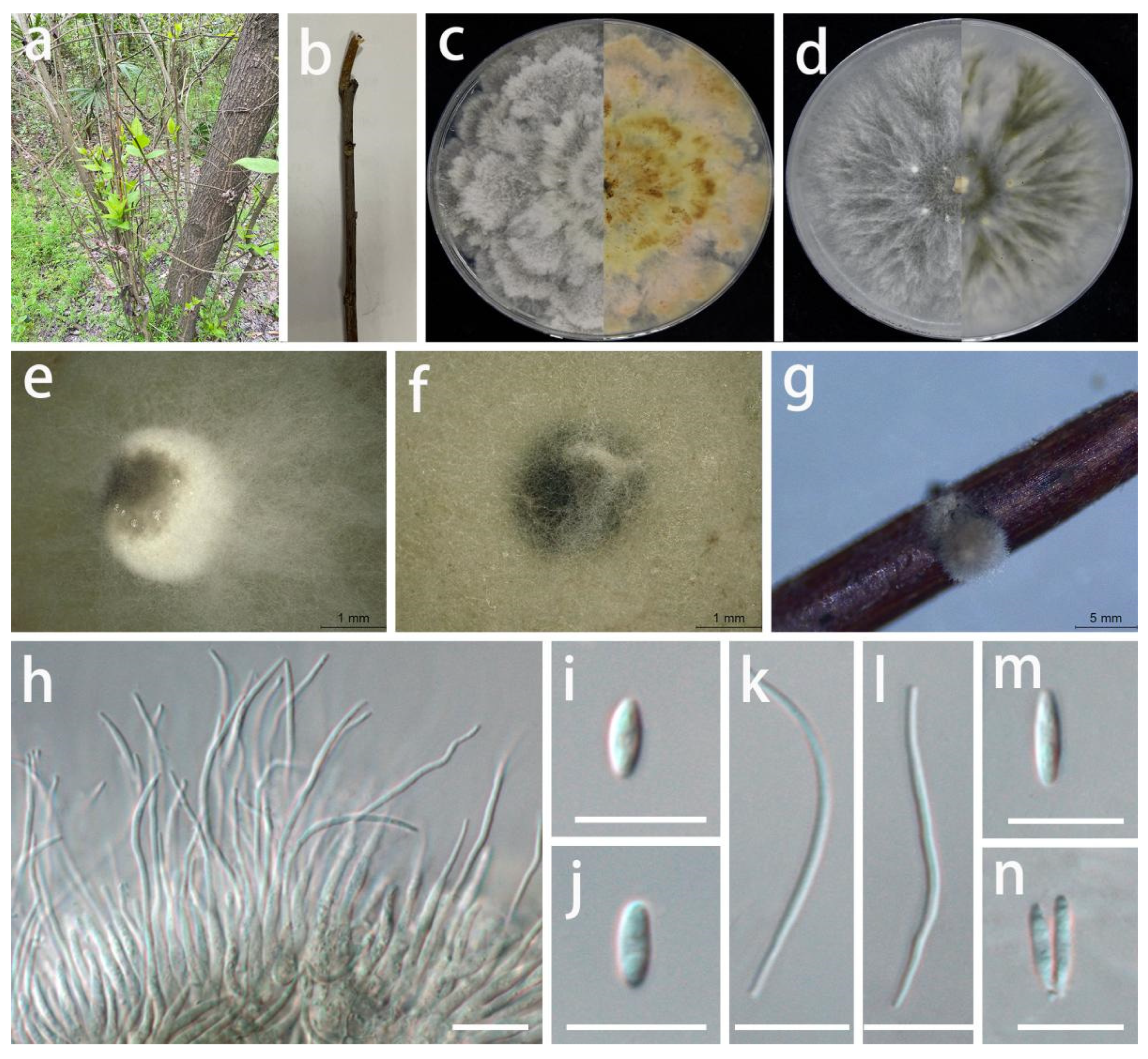{kind=link}
{kind=link}
{kind=link}
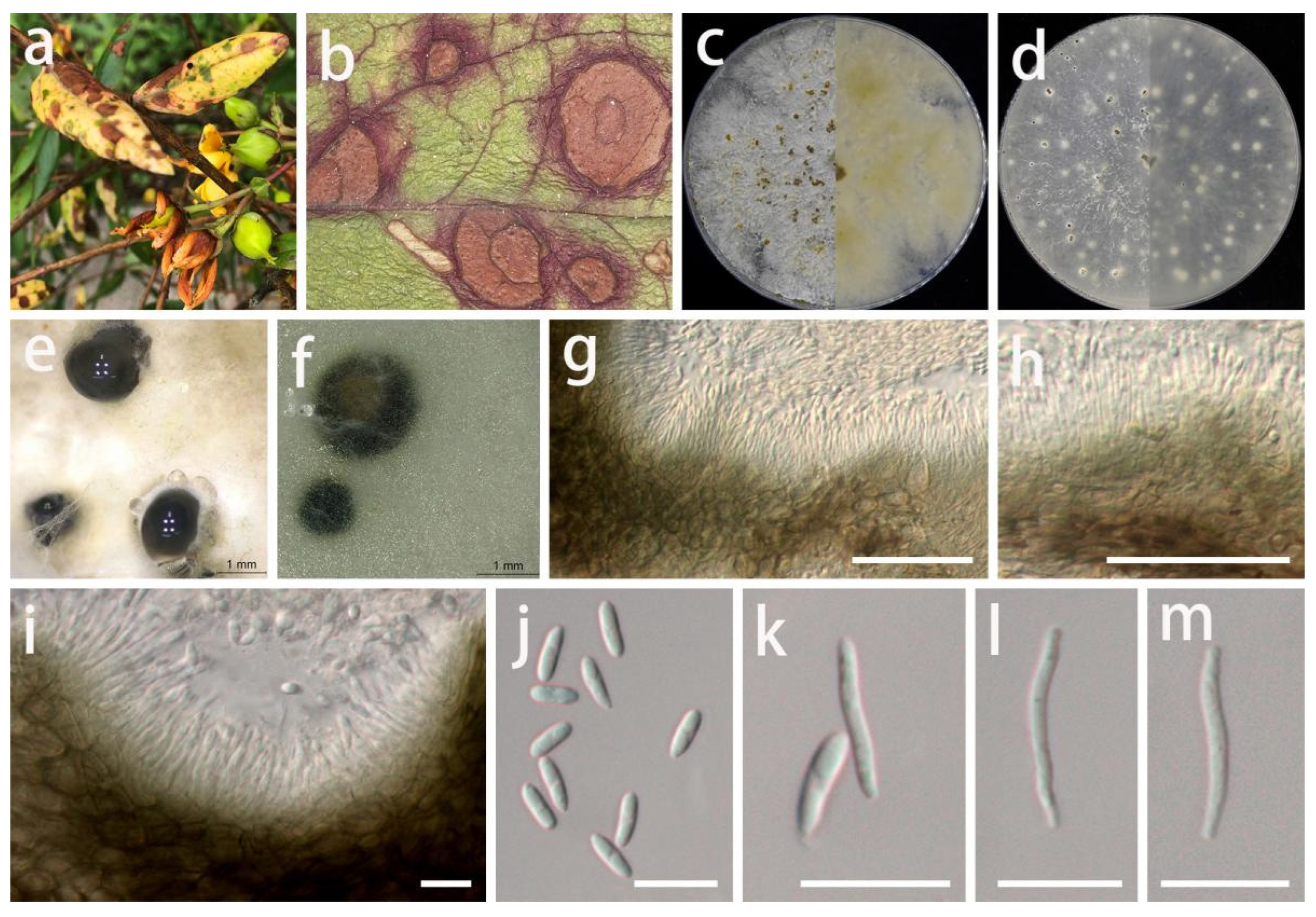{kind=link}
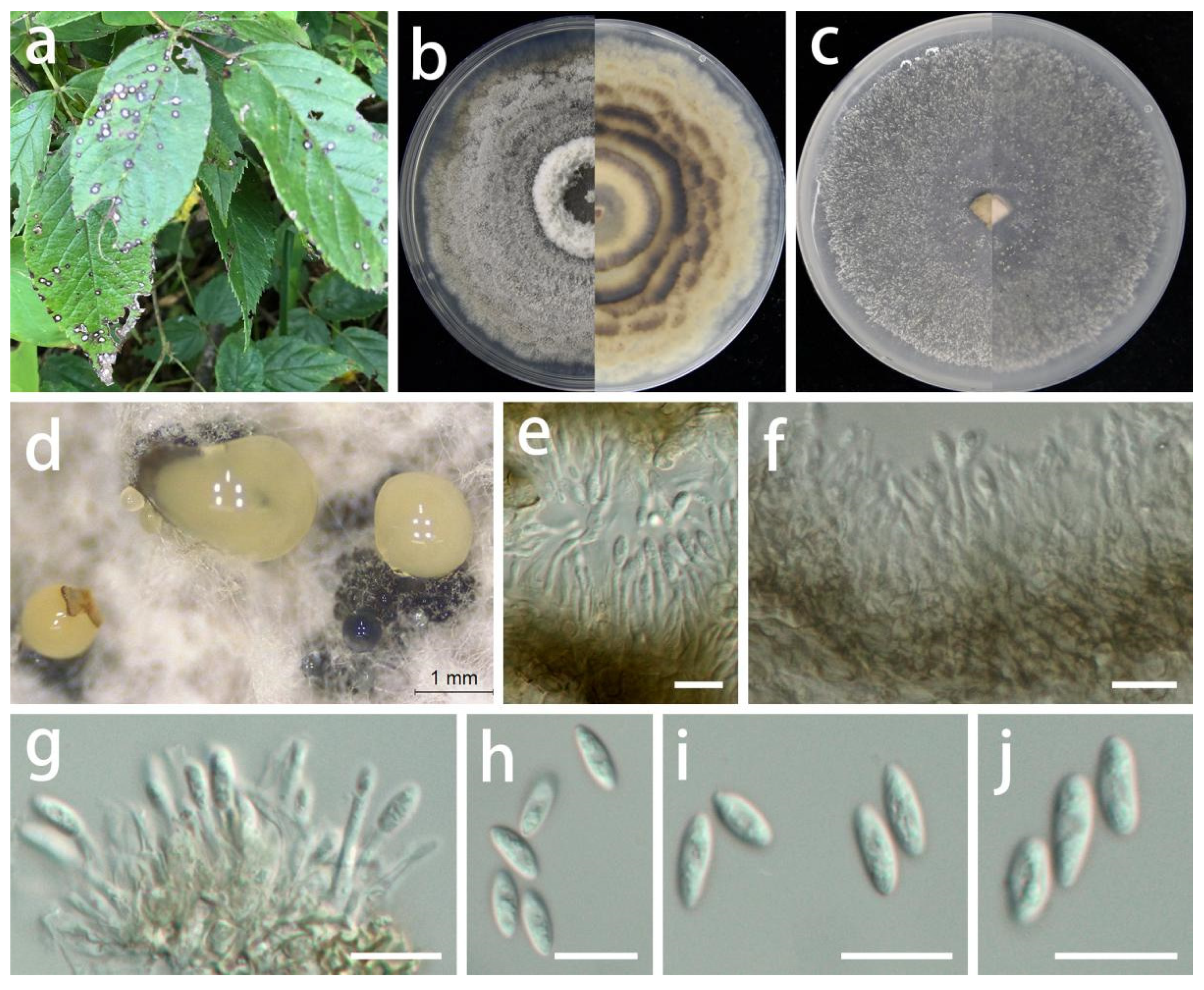{kind=link}
{kind=link}
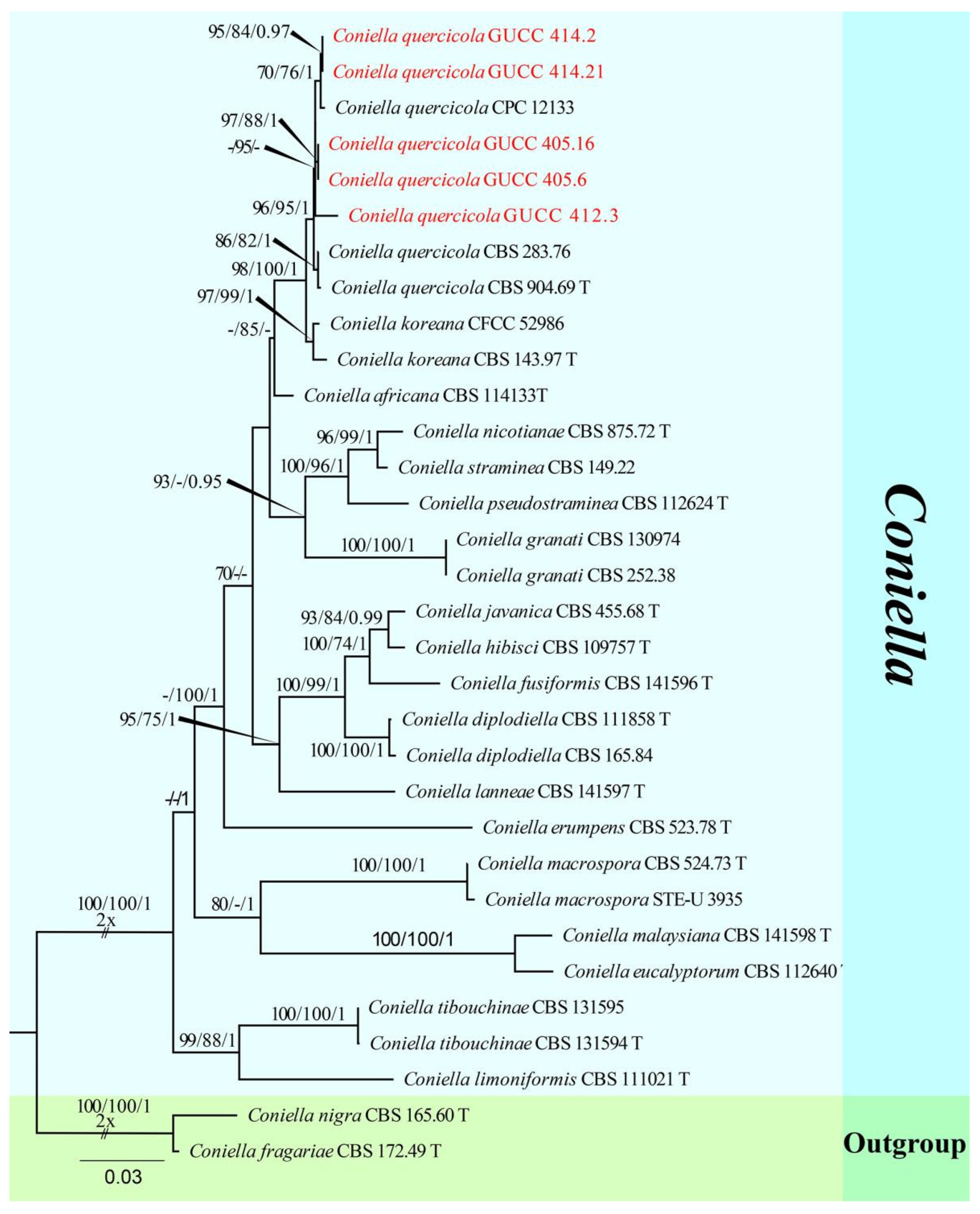{kind=link}
Table 1.
Primers and PCR procedures used in this study.
| Locus | Primers | Primer Sequence (5′–3′) | PCR Thermal Cycle Protocols | References |
|---|---|---|---|---|
| ITS | ITS5 | GGAAGTAAAAGTCGTAACAAGG | 94 °C–2 min; 94 °C–1 min; 58 °C–1 min; 72 °C–90 s; repeat 2–4 for 35 cycles; 72 °C–10 min; 4 °C on hold | White et al. [23] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| LSU | LR0R | ACCCGCTGAACTTAAGC | Rehner and Samuels [24]; Vilgalys and Hester [25] | |
| LR5 | TCCTGAGGGAAACTTCG | |||
| rpb2 | fRPB2-5F | GAYGAYMGWGATCAYTTYGG | 94 °C–2 min; 95 °C–45 s; 57 °C–50 s; 72 °C–90 s; repeat 2–4 for 35 cycles; 72 °C–10 min; 4 °C on hold | Liu et al. [26] |
| fRPB2-7cR | CCCATRGCTTGYTTRCCCAT | |||
| tef1 | EF1-728F | CATCGAGAAGTTCGAGAAGG | 94 °C–2 min; 95 °C–30 s; 58 °C–50 s; 72 °C–1 min; repeat 2–4 for 35 cycles; 72 °C–10 min; 4 °C on hold | Carbone and Kohn [27] |
| EF1-986R | TACTTGAAGGAACCCTTACC | |||
| tub2 | Bt2a | GGTAACCAAATCGGTGCTGCTTTC | 95 °C–5 min; 95 °C–30 s; 60 °C–30 s; 72 °C–30 s; repeat 2–4 for 34 cycles; 72 °C–10 min; 4 °C on hold | Glass and Donaldson [28] |
| Bt2b | ACCCTCAGTGTAGTGACCCTTGGC |
Table 2.
Strains and their GenBank accession numbers used in the molecular phylogenetic analyses of Cryphonectriaceae and Foliocryphiaceae. Newly generated sequences are in bold. (T) = ex–type strain.
Table 2.
Strains and their GenBank accession numbers used in the molecular phylogenetic analyses of Cryphonectriaceae and Foliocryphiaceae. Newly generated sequences are in bold. (T) = ex–type strain.
| Species | Strain Number | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | rpb2 | tef1 | tub2 | ||
| Amphilogia gyrosa | CBS 112922T | AF452111 | AY194107 | MN271782 | MN271818 | AF525714 |
| Aurantioporthe corni | CBS 245.90 | MN172403 | MN172371 | MN271784 | MN271822 | – |
| Aurantiosacculus acutatus | CBS 132181T | JQ685514 | JQ685520 | – | MN271823 | – |
| Aurantiosacculus castaneae | CFCC 52456T | MH514025 | MH514015 | MN271786 | – | MH539688 |
| Aurantiosacculus eucalyptorum | CPC 13229T | JQ685515 | JQ685521 | MN271785 | MN271824 | – |
| Aurapex penicillata | CMW 10030T | AY214311 | AY194103 | MN271787 | – | AY214239 |
| Aurifilum marmelostoma | CBS 124928T | MH863426 | MH874934 | MN271788 | MN271827 | MN987000 |
| Aurifilum terminali | CSF10762 | MN199838 | MN258786 | – | MN258781 | MN258771 |
| Celoporthe dispersa | CBS 118782T | DQ267130 | HQ730853 | – | HQ730840 | AY214280 |
| Celoporthe eucalypti | CMW 26908T | HQ730837 | HQ730863 | MN271790 | HQ730850 | MN263386 |
| Chrysofolia barringtoniae | TBRC 5647 | KU948046 | KU948045 | – | – | – |
| Chrysofolia colombiana | CPC 24986T | KR476738 | KR476771 | – | MN271829 | – |
| Chrysofolia coriariaesp. nov. | GUCC 416.4T | OP581211 | OP581237 | – | OP688516 | OP688542 |
| Chrysofolia coriariaesp. nov. | GUCC 416.14 | OP581212 | OP581238 | – | OP688517 | OP688543 |
| Chrysomorbus lagerstroemiae | CBS 142594T | KY929338 | KY929328 | – | MN271830 | KY929348 |
| Chrysoporthe hodgesiana | CBS 115854T | AY692322 | MN172380 | MN271793 | MN271836 | – |
| Chrysoporthe syzygiicola | CBS 124488T | FJ655005 | MN172383 | – | MN271839 | FJ805236 |
| Corticimorbus sinomyrti | CBS 140205T | KT167169 | KT167179 | MN271794 | MN271841 | KT167183 |
| Cryphonectria macrospora | CBS 109764 | EU199182 | AF408340 | EU220029 | KC465405 | AH014594 |
| Cryphonectria parasitica | ATCC 38755 | MH843497 | MH514021 | DQ862017 | MN271848 | MW086477 |
| Cryptometrion aestuescens | CBS 124014T | MH863335 | MH874864 | MN271798 | MN271851 | GQ369456 |
| Diversimorbus metrosideri | CBS 132866T | JQ862871 | JQ862828 | – | MN271857 | JQ862953 |
| Dwiroopa lythri | CBS 109755T | MN172410 | MN172389 | MN271801 | MN271859 | – |
| Dwiroopa punicae | CBS 143163T | MK510676 | MK510686 | MK510692 | MH020056 | MK510714 |
| Endothia chinensis | CFCC 52144T | MH514027 | MH514017 | – | MN271860 | MH539690 |
| Foliocryphia eucalypti | CBS 124779T | GQ303276 | GQ303307 | MN271802 | MN271861 | JQ706128 |
| Foliocryphia eucalyptorum | CBS 142536T | KY979772 | KY979827 | MN271803 | MN271862 | KY979936 |
| Holocryphia capensis | CBS 132870T | JQ862854 | JQ862811 | – | MN271883 | JQ862948 |
| Holocryphia eucalypti | CBS 115842T | MN172411 | MN172391 | MN271804 | MN271882 | JQ862772 |
| Immersiporthe knoxdaviesiana | CBS 132862T | JQ862765 | JQ862755 | MN271805 | MN271886 | JQ862785 |
| Latruncellus aurorae | CBS 125526T | GU726947 | HQ730872 | – | MN271888 | GU726959 |
| Luteocirrhus shearii | CBS 130776T | KC197021 | KC197019 | MN271807 | MN271890 | KC197006 |
| Microthia havanensis | CBS 115855 | DQ368735 | MN172393 | MN271811 | – | – |
| Myrtonectria myrtacearum | CMW 46433T | MG585736 | MG585750 | – | – | MG585734 |
| Neocryphonectria carpini | CFCC 53027T | MN172413 | MN172396 | – | – | – |
| Neocryphonectria chinensis | CFCC 53025T | MN172414 | MN172397 | MN271812 | MN271893 | – |
| Neocryphonectria chinensis | CFCC 53029 | MN172415 | MN172398 | MN271813 | MN271894 | – |
| Pseudomastigosporella guizhouensis sp. nov. | GUCC 406.6T | OP581233 | OP581246 | OP688514 | OP688538 | OP688563 |
| Pseudomastigosporella guizhouensis sp. nov. | GUCC 405.3 | OP581234 | OP581247 | OP688515 | OP688539 | OP688564 |
| Pseudomastigosporella guizhouensis sp. nov. | GUCC 405.4 | OP581235 | OP581248 | – | OP688540 | OP688565 |
| Pseudomastigosporella guizhouensis sp. nov. | GUCC 405.8 | OP581236 | OP581249 | – | OP688541 | OP688566 |
| Rostraureum tropicale | CBS 115725T | AY167435 | MN172399 | MN271814 | MN271895 | AY167431 |
| Ursicollum fallax | CBS 118663T | DQ368755 | EF392860 | MN271816 | MN271897 | AH015658 |
Table 3.
Strains and their GenBank accession numbers used in the molecular phylogenetic analyses of Diaporthe. Newly generated sequences are in bold. (T) = ex–type strain.
Table 3.
Strains and their GenBank accession numbers used in the molecular phylogenetic analyses of Diaporthe. Newly generated sequences are in bold. (T) = ex–type strain.
| Species | Strain Number | GenBank Accession Number | ||
|---|---|---|---|---|
| ITS | tef1 | tub2 | ||
| Diaporthe australafricana | CBS 111886T | KC343038 | KC343764 | KC344006 |
| D. beckhausii | CBS 138.27 | KC343041 | KC343767 | KC344009 |
| D. benedicti | CFCC 50062T | KP208847 | KP208853 | KP208855 |
| D. bohemiae | CBS 143347T | MG281015 | MG281536 | MG281188 |
| D. brasiliensis | CBS 133183T | KC343042 | KC343768 | KC344010 |
| D. caatingaensis | CBS 141542T | KY085927 | KY115603 | KY115600 |
| D. carpini | CBS 114437 | KC343044 | KC343770 | KC344012 |
| D. caryae | CFCC 52563T | MH121498 | MH121540 | MH121580 |
| D. caryae | PSCG520 | MK626952 | MK654895 | MK691315 |
| D. cassines | CPC 21916T | KF777155 | KF777244 | – |
| D. caulivora | CBS 127268T | KC343045 | KC343771 | KC344013 |
| D. caulivora | Dip1 | HM347703 | HM347687 | – |
| D. caulivora | Dpc11 | HM347704 | HM347688 | – |
| D. chimonanthi | HGUP191001 | MZ724752 | – | MZ724033 |
| D. chimonanthi | HGUP192087 | MZ724753 | – | MZ724034 |
| D. cotoneastri | CBS 439.82T | MH861511 | GQ250341 | JX275437 |
| D. cynaroidis | CBS 122676T | KC343058 | KC343784 | KC344026 |
| D. dejiangensis sp. nov. | GUCC 421.2T | OP581221 | OP688526 | OP688551 |
| D. dejiangensis sp. nov. | GUCC 421.21 | OP581222 | OP688527 | OP688552 |
| D. ellipicola | CGMCC 3.17084T | KF576270 | KF576245 | KF576294 |
| D. ellipicola | CGMCC 3.17085 | KF576271 | KF576246 | KF576295 |
| D. eres | CBS 138594T | KJ210529 | KJ210550 | KJ420799 |
| D. eres | CAA801 | KY435644 | KY435631 | KY435672 |
| D. eucommiigenasp. nov. | GUCC 420.9T | OP581223 | OP688528 | OP688553 |
| D. eucommiigenasp. nov. | GUCC 420.19 | OP581224 | OP688529 | OP688554 |
| D. fibrosa | CBS 109751 | KC343099 | KC343825 | KC344067 |
| D. fusicola | CGMCC 3.17087T | KF576281 | KF576256 | KF576305 |
| D. hypericisp. nov. | GUCC 414.4T | OP581227 | OP688532 | OP688557 |
| D. hypericisp. nov. | GUCC 414.14 | OP581228 | OP688533 | OP688558 |
| D. impulsa | CBS 114434 | KC343121 | KC343847 | KC344089 |
| D. italiana | MFLUCC 18-0090T | MH846237 | MH853686 | MH853688 |
| D. juglandigena sp. nov. | GUCC 422.16T | OP581229 | OP688534 | OP688559 |
| D. juglandigena sp. nov. | GUCC 422.161 | OP581230 | OP688535 | OP688560 |
| D. longicolla | ATCC 60325T | KJ590728 | KJ590767 | KJ610883 |
| D. longicolla | 1-2/4-3 | HM347711 | HM347682 | – |
| D. malorum | CAA734T | KY435638 | KY435627 | KY435668 |
| D. malorum | CAA740 | KY435642 | KY435629 | KY435670 |
| D. malorum | CAA752 | KY435643 | KY435630 | KY435671 |
| D. mediterranea | DAL-176 | MT007496 | MT006996 | MT006693 |
| D. nobilis | CBS 124030 | KC343149 | KC343875 | KC344117 |
| D. nothofagi | BRIP 54801T | JX862530 | JX862536 | KF170922 |
| D. ocoteae | CBS 141330T | KX228293 | – | KX228388 |
| D. ovoicicola | CGMCC 3.17092T | KF576264 | KF576239 | KF576288 |
| D. passiflorae | CBS 132527T | JX069860 | KY435633 | KY435674 |
| D. phaseolorum | Ar2 | HM347705 | HM347679 | – |
| D. phaseolorum | CBS 127266 | HM347707 | HM347672 | HQ333513 |
| D. phragmitis | CBS 138897T | KP004445 | – | KP004507 |
| D. rudis | CBS 109292T | KC343234 | KC343960 | KC344202 |
| D. salicicola | BRIP 54825T | JX862531 | JX862537 | KF170923 |
| D. subcylindrospora | KUMCC 17-0151 | MG746629 | MG746630 | MG746631 |
| D. tongrensissp. nov. | GUCC 421.10T | OP581225 | OP688530 | OP688555 |
| D. tongrensissp. nov. | GUCC 421.101 | OP581226 | OP688531 | OP688556 |
| D. ueckerae | FAU656T | KJ590726 | KJ590747 | KJ610881 |
| Diaporthella corylina | CBS 121124T | KC343004 | KC343730 | KC343972 |
| Diaporthella cryptica | CBS 140348T | MN172409 | MN271854 | – |
Table 4.
Strains and their GenBank accession numbers used in the molecular phylogenetic analyses of Gnomoniopsis. Newly generated sequences are in bold. (T) = ex–type strain.
Table 4.
Strains and their GenBank accession numbers used in the molecular phylogenetic analyses of Gnomoniopsis. Newly generated sequences are in bold. (T) = ex–type strain.
| Species | Strain Number | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | rpb2 | tef1 | tub2 | ||
| Gnomoniopsis alderdunense | CBS 125681 | GU320827 | MH875098 | – | GU320802 | GU320789 |
| G. alderdunensis | CBS 125680T | MH863625 | MH875097 | – | GU320801 | GU320787 |
| G. angolensis | CBS 145057T | MK047428 | MK047479 | MK047539 | – | – |
| G. castaneae | GCAS5 | MH107830 | MZ682110 | – | MH213486 | MH213481 |
| G. castanopsidis | CFCC 54437T | MZ902909 | – | – | MZ936385 | |
| G. chamaemori | CBS 803.79 | EU254808 | EU255107 | – | GU320809 | EU219155 |
| G. chinensis | CFCC 52286T | MG866032 | – | – | MH545370 | MH545366 |
| G. clavulata | AR 4313 | EU254818 | – | EU219251 | EU221934 | EU219211 |
| G. comari | CBS 806.79T | EU254821 | EU255114 | EU219286 | GU320810 | EU219156 |
| G. daii | CFCC 55517 | MZ902911 | – | – | MZ936387 | MZ936403 |
| G. fagacearum | CFCC 54316T | MZ902916 | – | – | MZ936392 | MZ936408 |
| G. fructicola (=G. fragariae) | CBS 125671 | MH863616 | MH875088 | – | GU320793 | GU320776 |
| G. guangdongensis | CFCC 54443T | MZ902918 | – | – | MZ936394 | MZ936410 |
| G. hainanensis | CFCC 54376T | MZ902921 | – | – | MZ936397 | MZ936413 |
| G. idaeicola | CBS 125674T | MH863619 | MH875091 | – | GU320796 | GU320780 |
| G. macounii | CBS 121468T | MH863110 | MH874666 | – | GU320804 | – |
| G. occulta | CBS 125678 | MH863623 | MH875095 | – | GU320800 | GU320786 |
| G. paraclavulata | CBS 123202T | GU320830 | – | – | GU320815 | GU320775 |
| G. racemula | AR 3892 | EU254841 | EU255122 | EU219241 | EU221889 | EU219125 |
| G. rosae | CBS 145085 | MK047451 | MK047501 | MK047547 | – | – |
| G. rosae | GUCC 408.7 | OP581231 | OP581244 | OP688512 | OP688536 | OP688561 |
| G. rosae | GUCC 408.17 | OP581232 | OP581245 | OP688513 | OP688537 | OP688562 |
| G. rossmaniae | CFCC 54307T | MZ902923 | – | – | MZ936399 | MZ936415 |
| G. sanguisorbae | CBS 858.79 | GU320818 | KY496735 | – | GU320805 | GU320790 |
| G. silvicola | CFCC 54418T | MZ902926 | – | – | MZ936402 | MZ936418 |
| G. smithogilvyi | CBS 130190T | MH865607 | MH877031 | JQ910648 | JQ910645 | JQ910639 |
| G. tormentillae | CBS 904.79 | EU254856 | EU255133 | – | GU320795 | EU219165 |
| G. xunwuensis | CFCC 53115T | MK432667 | MK429910 | – | MK578141 | MK578067 |
| Sirococcus tsugae | CBS 119626 | EF512472 | – | – | EF512534 | EU219140 |
Table 5.
Strains and their GenBank accession numbers used in the molecular phylogenetic analyses of Coniella. Newly generated sequences are in bold. (T) = ex–type strain.
Table 5.
Strains and their GenBank accession numbers used in the molecular phylogenetic analyses of Coniella. Newly generated sequences are in bold. (T) = ex–type strain.
| Species | Strain Number | GenBank Accession Number | ||
|---|---|---|---|---|
| ITS | LSU | tef1 | ||
| Coniella africana | CBS 114133T | AY339344 | AY339293 | KX833600 |
| C. diplodiella | CBS 111858T | MH862886 | KX833335 | KX833603 |
| C. diplodiella | CBS 165.84 | KX833529 | KX833354 | KX833622 |
| C. erumpens | CBS 523.78T | KX833535 | KX833361 | KX833630 |
| C. eucalyptorum | CBS 112640T | AY339338 | AY339290 | KX833637 |
| C. fusiformis | CBS 141596T | KX833576 | KX833397 | KX833674 |
| C. granati | CBS 130974 | JN815312 | KX833398 | KX833675 |
| C. granati | CBS 252.38 | KX833581 | AY339291 | KX833681 |
| C. hibisci | CBS 109757T | KX833589 | AF408337 | KX833689 |
| C. javanica | CBS 455.68T | KX833583 | KX833403 | KX833683 |
| C. koreana | CBS 143.97T | KX833584 | AF408378 | KX833684 |
| C. koreana | CFCC 52986 | MK432612 | MK429882 | MK578112 |
| C. lanneae | CBS 141597T | KX833585 | KX833404 | KX833685 |
| C. limoniformis | CBS 111021T | KX833586 | KX833405 | KX833686 |
| C. macrospora | CBS 524.73T | KX833587 | AY339292 | KX833687 |
| C. macrospora | STE-U 3935 | AY339343 | – | AY339363 |
| C. malaysiana | CBS 141598T | KX833588 | KX833406 | KX833688 |
| C. nicotianae | CBS 875.72T | KX833590 | KX833407 | KX833690 |
| C. pseudostraminea | CBS 112624T | KX833593 | KX833412 | KX833696 |
| C. quercicola | CBS 904.69T | KX833595 | KX833414 | KX833698 |
| C. quercicola | CBS 283.76 | KX833594 | KX833413 | KX833697 |
| C. quercicola | CPC 12133 | KX833596 | – | KX833699 |
| C. quercicola | GUCC 414.2 | OP581213 | OP581239 | OP688518 |
| C. quercicola | GUCC 414.21 | OP581214 | OP581240 | OP688519 |
| C. quercicola | GUCC 412.3 | OP581215 | OP581241 | OP688520 |
| C. quercicola | GUCC 405.6 | OP581216 | OP581242 | OP688521 |
| C. quercicola | GUCC 405.16 | OP581217 | OP581243 | OP688522 |
| C. straminea | CBS 149.22 | AY339348 | AY339296 | KX833704 |
| C. tibouchinae | CBS 131594T | JQ281774 | KX833418 | JQ281778 |
| C. tibouchinae | CBS 131595 | JQ281775 | KX833419 | JQ281779 |
| C. fragariae | CBS 172.49T | AY339317 | AY339282 | KX833663 |
| C. nigra | CBS 165.60T | AY339319 | KX833408 | KX833691 |
Table 6.
Parameters of maximum parsimony and Bayesian methods in this study.
| Datasets | Maximum Parsimony (MP) | |||||
|---|---|---|---|---|---|---|
| TL | PT | CI | RI | RC | HI | |
| Foliocryphiaceae | 4515 | 9 | 0.5132 | 0.6746 | 0.3462 | 0.4868 |
| Diaporthe | 2261 | 9 | 0.5657 | 0.8362 | 0.4730 | 0.4343 |
| Gnomoniopsis | 2423 | 3 | 0.5712 | 0.5930 | 0.3387 | 0.4288 |
| Coniella | 1292 | 8 | 0.5735 | 0.7112 | 0.4079 | 0.4265 |
| Bayesian | ||||||
| Model | ASDSF | |||||
| ITS | LSU | rpb2 | tef1 | tub2 | ||
| Foliocryphiaceae | GTR+I+G | HKY+I+G | HKY+G | 0.009523 | ||
| Diaporthe | SYM+I+G | n/a | n/a | HKY+I+G | 0.009906 | |
| Gnomoniopsis | GTR+I+G | GTR+I | GTR+G | HKY+I+G | GTR+I+G | 0.009908 |
| Coniella | SYM+I+G | GTR+I+G | n/a | GTR+I+G | n/a | 0.009408 |
TL: Tree length; PT: Parsimonious tree; CI: Consistency indices; RI: Retention indices; RC: Rescaled consistency indices; HI: Homoplasy index; Model: the best nucleotide substitution model used for the different partitions; ASDSF: average standard deviation of split frequencies.
Table 7.
DNA base differences between our strains and related taxa in the five-locus regions. Asterisks (*) denote our material. (T) = ex–type strain.
Table 7.
DNA base differences between our strains and related taxa in the five-locus regions. Asterisks (*) denote our material. (T) = ex–type strain.
| Species | Strain Number | Gene Region and Alignment Positions | ||||
|---|---|---|---|---|---|---|
| ITS (1–769 Characters) | LSU (770–1617 Characters) | rpb2 (1618–2442 Characters) | tef1 (2443–2952 Characters) | tub2 (2953–3596 Characters) | ||
| (with gap: 151 characters) | (with gap: 41 characters) | (no data) | (no data) | (no data) | ||
| Chrysofolia barringtoniae | TBRC 5647 | – | – | n/a | n/a | n/a |
| Chrysofolia coriariae sp. nov. * | GUCC 416.4T | 31 | 4 | n/a | n/a | n/a |
| Chrysofolia coriariae sp. nov. * | GUCC 416.14 | 31 | 4 | n/a | n/a | n/a |
| (with gap: 158 characters) | (with gap: 41 characters) | (no data) | (with gap: 156 characters) | (no data) | ||
| Chrysofolia colombiana | CPC 24986 | – | – | n/a | – | n/a |
| Chrysofolia coriariae sp. nov. * | GUCC 416.4T | 7 | 2 | n/a | 149 | n/a |
| Chrysofolia coriariae sp. nov. * | GUCC 416.14 | 7 | 2 | n/a | 149 | n/a |
| (with gap: 152 characters) | (with gap: 34 characters) | (no data) | (no data) | (no data) | ||
| Neocryphonectria carpini | CFCC 53027T | – | – | n/a | n/a | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 406.6T | 116 | 22 | n/a | n/a | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.3 | 116 | 21 | n/a | n/a | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.4 | 116 | 21 | n/a | n/a | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.8 | 116 | 21 | n/a | n/a | n/a |
| (with gap: 139 characters) | (with gap: 34 characters) | (with gap: 53 characters) | (with gap: 177 characters) | (no data) | ||
| Neocryphonectria chinensis | CFCC 53025T | – | – | – | – | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 406.6T | 118 | 21 | 75 | 130 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.3 | 118 | 20 | 76 | 131 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.4 | 118 | 20 | n/a | 129 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.8 | 118 | 20 | n/a | 132 | n/a |
| (with gap: 333 characters) | (with gap: 34 characters) | (with gap: 200 characters) | (with gap: 177 characters) | (no data) | ||
| Neocryphonectria chinensis | CFCC 53029 | – | – | – | – | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 406.6T | 32 | 21 | 66 | 130 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.3 | 32 | 20 | 67 | 131 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.4 | 32 | 20 | n/a | 129 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.8 | 32 | 20 | n/a | 132 | n/a |
| ITS (1–640 characters) | (no data) | (no data) | tef1 (641–1116 characters) | tub2 (1117–1770 characters) | ||
| (with gap: 146 characters) | (no data) | (no data) | (with gap: 163 characters) | (with gap: 140 characters) | ||
| Diaporthe juglandigena sp. nov. * | GUCC 422.16T | – | n/a | n/a | – | – |
| Diaporthe juglandigena sp. nov. * | GUCC 422.161 | 0 | n/a | n/a | 0 | 0 |
| Diaporthe chimonanthi | HGUP191001 | 18 | n/a | n/a | n/a | 5 |
| Diaporthe chimonanthi | HGUP192087 | 23 | n/a | n/a | n/a | 2 |
| Diaporthe caryae | CFCC 52563T | 2 | n/a | n/a | 6 | 17 |
| Diaporthe caryae | PSCG520 | 4 | n/a | n/a | 6 | 30 |
| (with gap: 98 characters) | (no data) | (no data) | (with gap: 275 characters) | (with gap: 151 characters) | ||
| Diaporthe eucommiigena sp. nov. * | GUCC 420.9T | – | n/a | n/a | – | – |
| Diaporthe eucommiigena sp. nov. * | GUCC 420.19 | 0 | n/a | n/a | 0 | 0 |
| Diaporthe passiflorae | CBS 132527T | 11 | n/a | n/a | 23 | 13 |
| Diaporthe malorum | CAA734T | 15 | n/a | n/a | 19 | 13 |
| Diaporthe malorum | CAA740 | 15 | n/a | n/a | 19 | 13 |
| Diaporthe malorum | CAA752 | 15 | n/a | n/a | 19 | 13 |
| (with gap: 128 characters) | (no data) | (no data) | (with gap: 147 characters) | (with gap: 255 characters) | ||
| Diaporthe dejiangensis sp. nov. * | GUCC 421.2T | – | n/a | n/a | – | – |
| Diaporthe dejiangensis sp. nov. * | GUCC 421.21 | 0 | n/a | n/a | 0 | 0 |
| Diaporthe eres | CBS 138594T | 4 | n/a | n/a | 6 | 6 |
| Diaporthe eres | CAA801 | 3 | n/a | n/a | 7 | 7 |
| Diaporthe cotoneastri | CBS 439.82T | 7 | n/a | n/a | 5 | 13 |
| (with gap: 67 characters) | (no data) | (no data) | (with gap: 105 characters) | (with gap: 138 characters) | ||
| Diaporthe tongrensis sp. nov. * | GUCC 421.10T | – | n/a | n/a | – | – |
| Diaporthe tongrensis sp. nov. * | GUCC 421.101 | 0 | n/a | n/a | 0 | 0 |
| Diaporthe phragmitis | CBS 138897T | 8 | n/a | n/a | n/a | 14 |
| (with gap: 98 characters) | (no data) | (no data) | (with gap: 145 characters) | (with gap: 216 characters) | ||
| Diaporthe hyperici sp. nov. * | GUCC 414.4T | – | n/a | n/a | – | – |
| Diaporthe hyperici sp. nov. * | GUCC 414.14 | 0 | n/a | n/a | 0 | 2 |
| Diaporthe caulivora | CBS 127268T | 17 | n/a | n/a | 30 | 11 |
| Diaporthe caulivora | Dip1 | 17 | n/a | n/a | 29 | n/a |
| Diaporthe caulivora | Dpc11 | 17 | n/a | n/a | 29 | n/a |
| ITS (1–572 characters) | LSU (573–1423 characters) | rpb2 (1424–2460 characters) | tef1 (2461–2861 characters) | tub2 (2862–3354 characters) | ||
| (with gap: 34 characters) | (with gap: 11 characters) | (with gap: 279 characters) | (no data) | (no data) | ||
| Gnomoniopsis rosae | CBS 145085 | – | – | – | (no data) | (no data) |
| Gnomoniopsis rosae * | GUCC 408.7 | 0 | 0 | 0 | n/a | n/a |
| Gnomoniopsis rosae * | GUCC 408.17 | 0 | 0 | 0 | n/a | n/a |
| ITS (1–595 characters) | LSU (596–1767 characters) | (no data) | tef1 (1768–2165 characters | (no data) | ||
| (with gap: 40 characters) | (with gap: 390 characters) | n/a | (with gap: 135 characters) | n/a | ||
| Coniella quercicola | CBS 904.69T | – | – | n/a | – | n/a |
| Coniella quercicola | CBS 283.76 | 0 | 0 | n/a | 1 | n/a |
| Coniella quercicola | CPC 12133 | 1 | n/a | n/a | 5 | n/a |
| Coniella quercicola * | GUCC 414.2 | 0 | 0 | n/a | 10 | n/a |
| Coniella quercicola * | GUCC 414.21 | 0 | 0 | n/a | 10 | n/a |
| Coniella quercicola * | GUCC 412.3 | 0 | 0 | n/a | 18 | n/a |
| Coniella quercicola * | GUCC 405.6 | 0 | 0 | n/a | 7 | n/a |
| Coniella quercicola * | GUCC 405.16 | 0 | 0 | n/a | 6 | n/a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, S.-Y.; McKenzie, E.H.C.; Phillips, A.J.L.; Li, Y.; Wang, Y. Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China. J. Fungi 2022, 8, 1301. https://doi.org/10.3390/jof8121301
AMA Style
Wang S-Y, McKenzie EHC, Phillips AJL, Li Y, Wang Y. Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China. Journal of Fungi. 2022; 8(12):1301. https://doi.org/10.3390/jof8121301
Chicago/Turabian StyleWang, Si-Yao, Eric H. C. McKenzie, Alan J. L. Phillips, Yan Li, and Yong Wang. 2022. "Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China" Journal of Fungi 8, no. 12: 1301. https://doi.org/10.3390/jof8121301
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.