
Transcriptome Analysis of Fusarium–Tomato Interaction Based on an Updated Genome Annotation of Fusarium oxysporum f. sp. lycopersici Identifies Novel Effector Candidates That Suppress or Induce Cell Death in Nicotiana benthamiana


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Fol Strain and Inoculations
2.3. Extraction of RNA for RNA-Seq and qRT-PCR Analysis
2.4. RNA-Seq-Guided Genome Annotation and Genome Alignment
2.5. Secretome Prediction
2.6. Gene Expression Analysis
2.7. Plant Transformation Vector Construction
2.8. Agrobacterium-Mediated Transient Assays in N. benthamiana
2.9. Yeast Secretion Trap Assay
3. Results
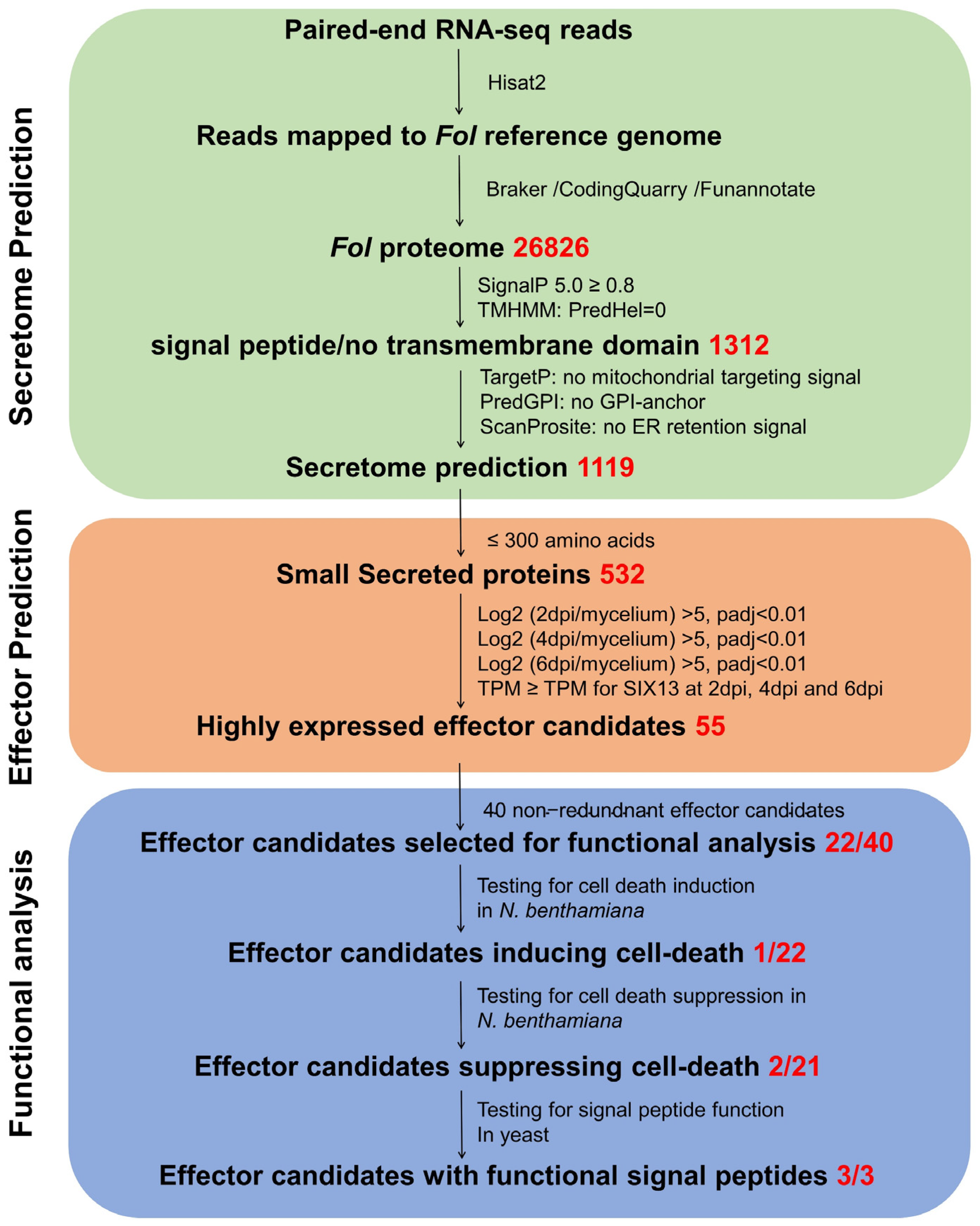
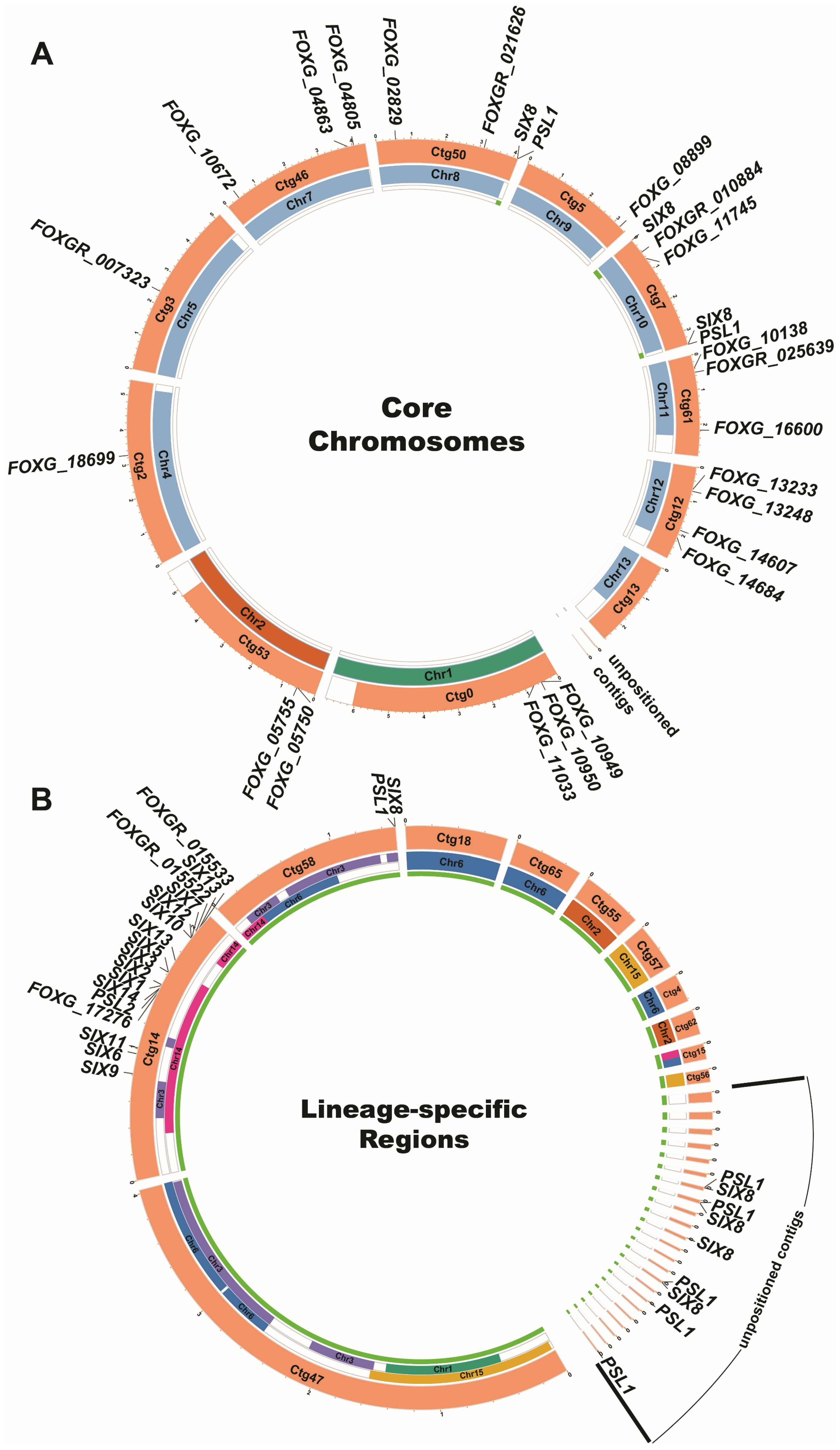
3.1. Identifying Candidate Effector Genes Expressed during Fol Infection
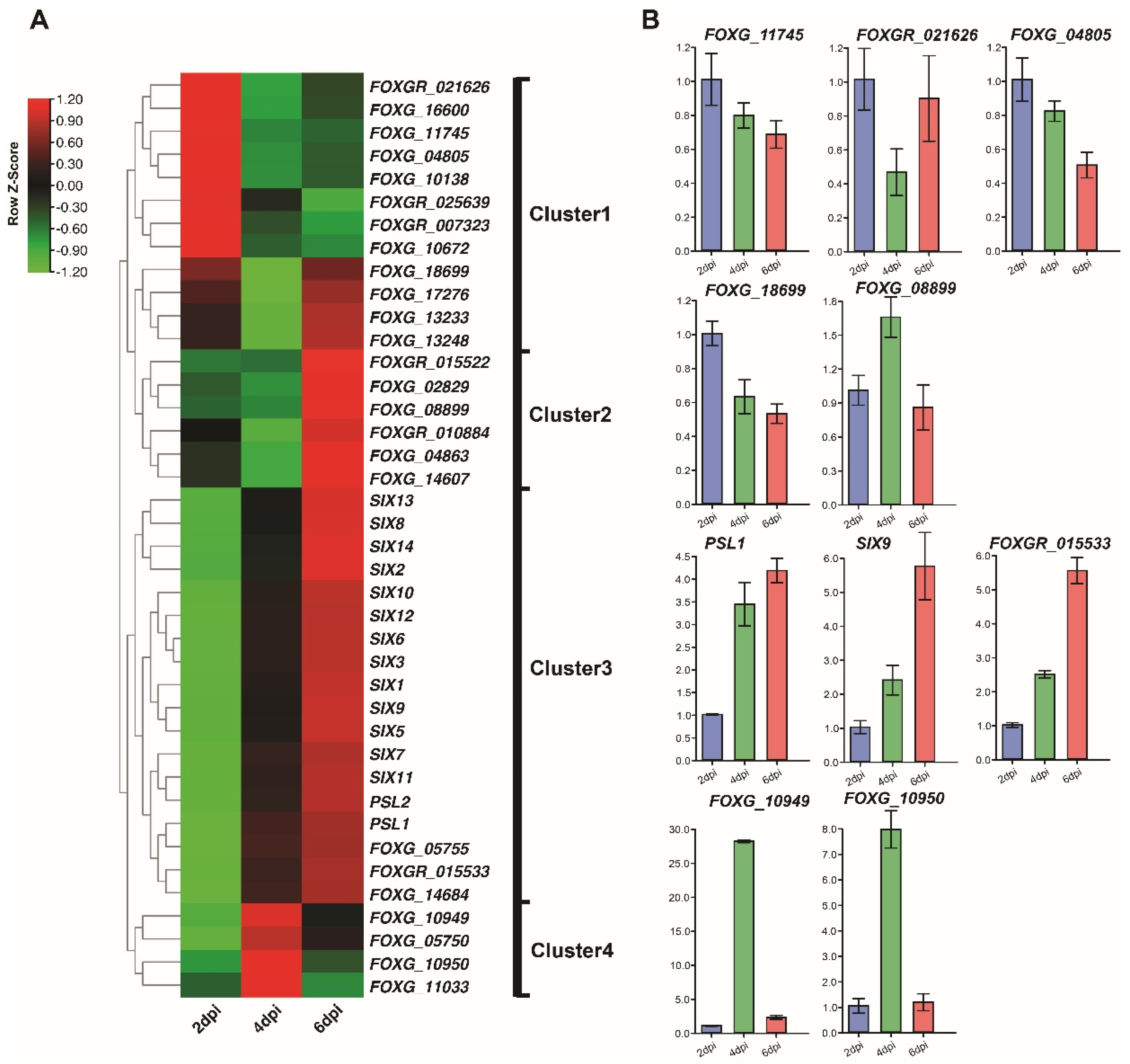
3.2. Expression Profiles of Fol Effector Candidates during Infection
3.3. Identification of Cell Death-Inducing Effector Candidates
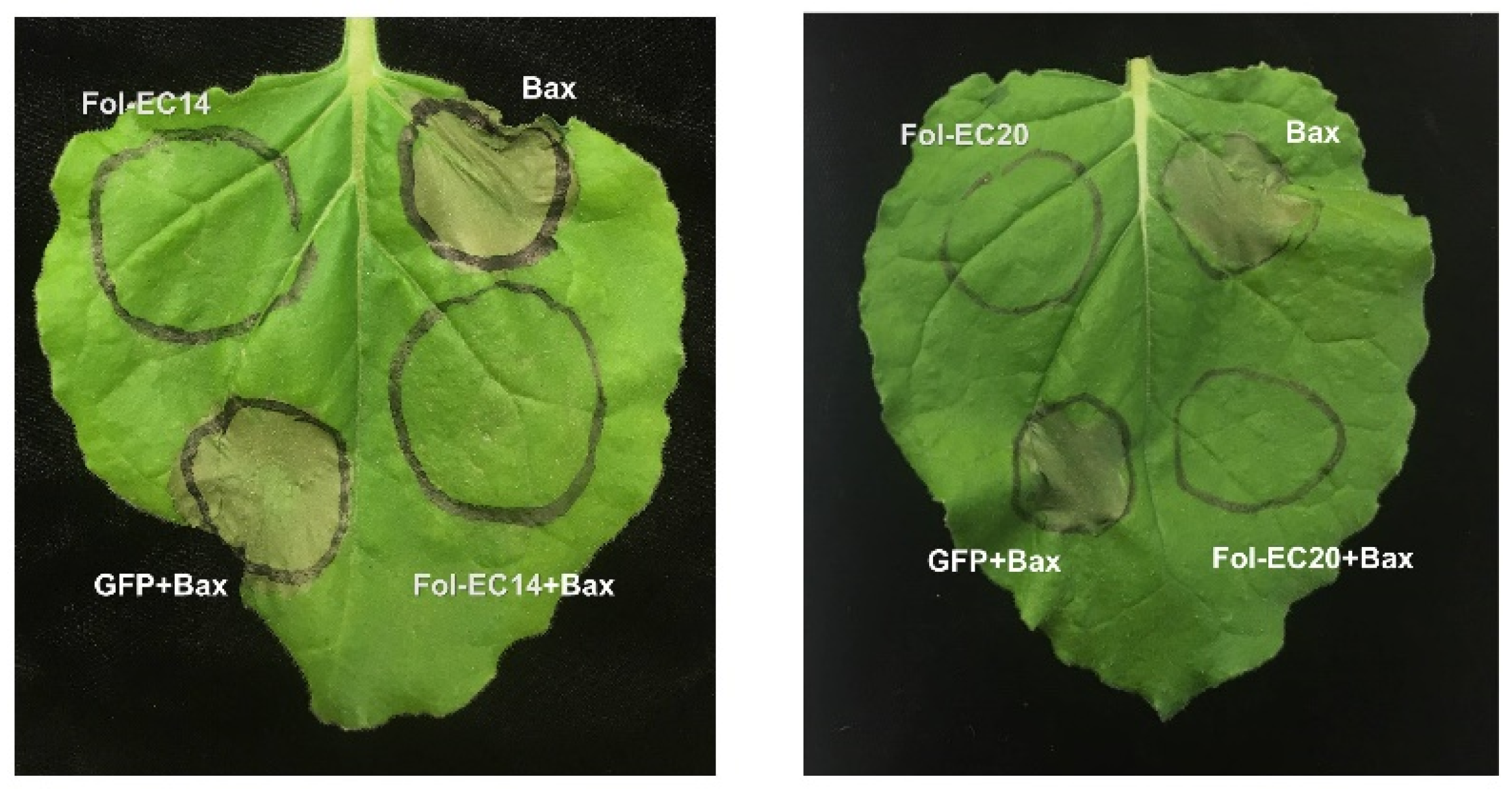
3.4. Identification of Effector Candidates That Suppress Bax-Induced Cell Death Using a Transient Expression Assay
3.5. Suppression of I/Avr1- and I-2/Avr2-Induced Cell Death
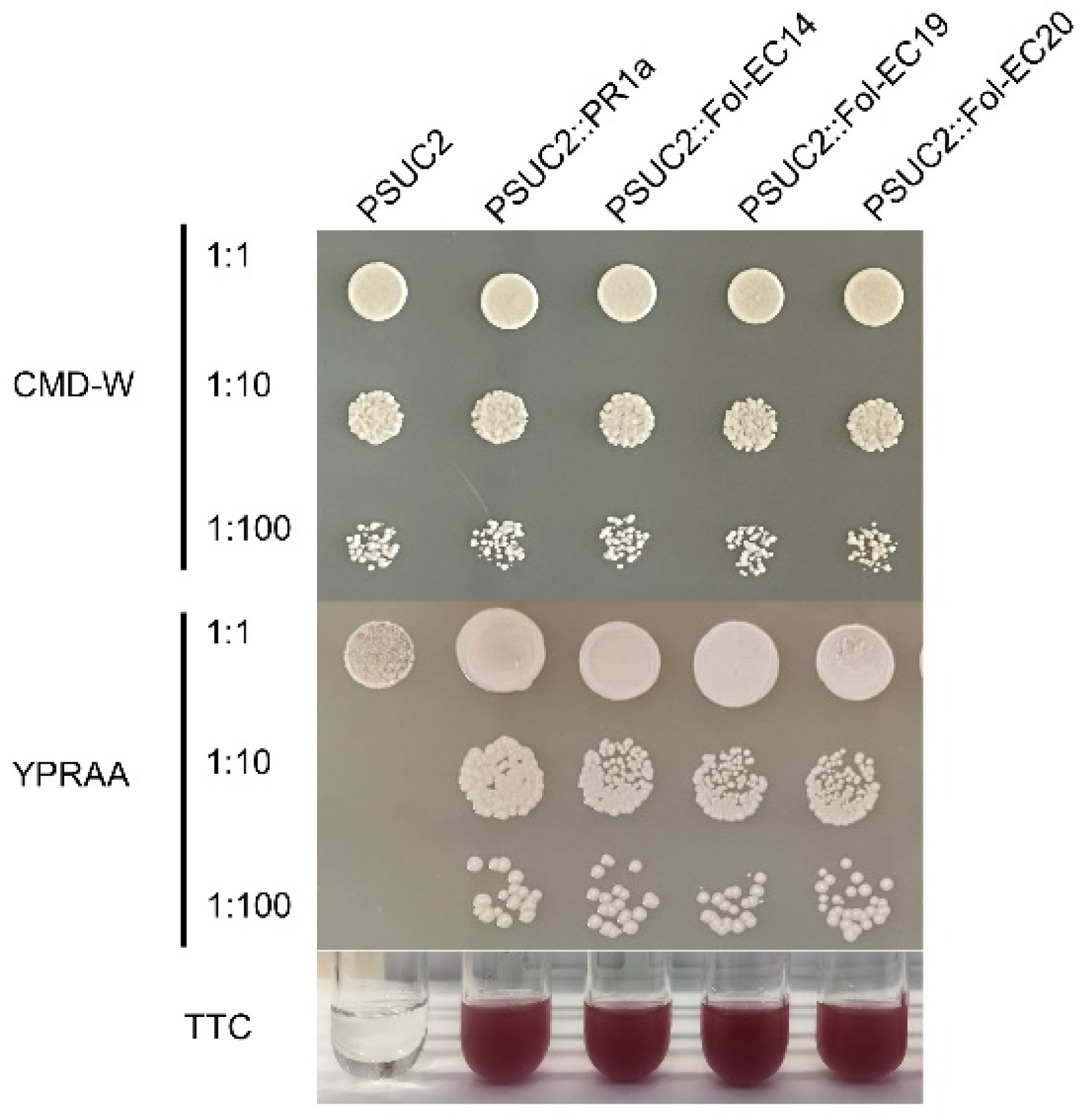
3.6. Functional Characterization of the Signal Peptides of the Fol-EC14, Fol-EC19, and Fol-EC20 Effectors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, J.-M.; Zhang, Y. Plant Immunity: Danger perception and signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, M.; Valent, B. Filamentous plant pathogen effectors in action. Nat. Rev. Genet. 2013, 11, 800–814. [Google Scholar] [CrossRef]
- van der Burgh, A.M.; Joosten, M. Plant Immunity: Thinking outside and inside the box. Trends Plant Sci. 2019, 24, 587–601. [Google Scholar] [CrossRef]
- Thomma, B.P.; Nurnberger, T.; Joosten, M.H. Of PAMPs and effectors: The blurred PTI-ETI dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondeau, K.; Blaise, F.; Graille, M.; Kale, S.D.; Linglin, J.; Ollivier, B.; Labarde, A.; Lazar, N.; Daverdin, G.; Balesdent, M.H.; et al. Crystal structure of the effector AvrLm4-7 of Leptosphaeria maculans reveals insights into its translocation into plant cells and recognition by resistance proteins. Plant J. 2015, 83, 610–624. [Google Scholar] [CrossRef]
- Plissonneau, C.; Daverdin, G.; Ollivier, B.; Blaise, F.; Degrave, A.; Fudal, I.; Rouxel, T.; Balesdent, M. A game of hide and seek between avirulence genes AvrLm4-7 and AvrLm3 in Leptosphaeria maculans. New Phytol. 2016, 209, 1613–1624. [Google Scholar] [CrossRef] [Green Version]
- Ghanbarnia, K.; Ma, L.; Larkan, N.J.; Haddadi, P.; Fernando, W.G.D.; Borhan, M.H. Leptosphaeria maculans AvrLm9: A new player in the game of hide and seek with AvrLm4-7. Mol. Plant Pathol. 2018, 19, 1754–1764. [Google Scholar] [CrossRef] [Green Version]
- Bourras, S.; McNally, K.E.; Ben-David, R.; Parlange, F.; Roffler, S.; Praz, C.R.; Oberhaensli, S.; Menardo, F.; Stirnweis, D.; Frenkel, Z.; et al. Multiple avirulence loci and allele-specific efector recognition control the Pm3 race-specific resistance of wheat to powdery mildew. Plant Cell 2015, 27, 2991–3012. [Google Scholar]
- Houterman, P.M.; Cornelissen, B.J.C.; Rep, M. Suppression of plant resistance gene-based immunity by a fungal effector. PLoS Pathog. 2008, 4, e1000061. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, S.R.; Yin, C.; Kud, J.; Tanaka, K.; Mahoney, A.K.; Xiao, F.; Hulbert, S.H. Effectors from wheat rust fungi suppress multiple plant defense responses. Phytopathology 2017, 107, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selin, C.; de Kievit, T.R.; Belmonte, M.F.; Fernando, W.G. Elucidating the role of effectors in plant-fungal interactions: Progress and challenges. Front. Microbiol. 2016, 7, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemri, A.; Saunders, D.G.; Anderson, C.; Upadhyaya, N.M.; Win, J.; Lawrence, G.J.; Jones, D.A.; Kamoun, S.; Ellis, J.G.; Dodds, P.N. The genome sequence and effector complement of the flax rust pathogen Melampsora lini. Front. Plant Sci. 2014, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- Guyon, K.; Balagué, C.; Roby, D.; Raffaele, S. Secretome analysis reveals effector candidates associated with broad host range necrotrophy in the fungal plant pathogen Sclerotinia sclerotiorum. BMC Genom. 2014, 15, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddadi, P.; Ma, L.; Wang, H.; Borhan, M.H. Genome-wide transcriptomic analyses provide insights into the lifestyle transition and effector repertoire of Leptosphaeria maculans during the colonization of Brassica napus seedlings. Mol. Plant Pathol. 2016, 17, 1196–1210. [Google Scholar] [CrossRef] [Green Version]
- Pérez-López, E.; Hossain, M.; Tu, J.; Waldner, M.; Todd, C.D.; Kusalik, A.J.; Wei, Y.; Bonham-Smith, P.C. Transcriptome analysis identifies Plasmodiophora brassicae secondary infection effector candidates. J. Eukaryot. Microbiol. 2020, 67, 337–351. [Google Scholar] [CrossRef] [Green Version]
- Kanja, C.; Hammond-Kosack, K.E. Proteinaceous effector discovery and characterization in filamentous plant pathogens. Mol. Plant Pathol. 2020, 21, 1353–1376. [Google Scholar] [CrossRef]
- Petre, B.; Saunders, D.G.O.; Sklenar, J.; Lorrain, C.; Krasileva, K.; Win, J.; Duplessis, S.; Kamoun, S. Heterologous expression screens in Nicotiana benthamiana identify a candidate effector of the wheat yellow rust pathogen that associates with processing bodies. PLoS ONE 2016, 11, e0149035. [Google Scholar] [CrossRef] [Green Version]
- Lacomme, C.; Santa Cruz, S. Bax-induced cell death in tobacco is similar to the hypersensitive response. Proc. Natl. Acad. Sci. USA 1999, 96, 7956–7961. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Han, C.; Ferreira, A.O.; Yu, X.; Ye, W.; Tripathy, S.; Kale, S.D.; Gu, B.; Sheng, Y.; Sui, Y.; et al. Transcriptional programming and functional interactions within the Phytophthora sojae RXLR effector repertoire. Plant Cell 2021, 23, 2064–2086. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, J.; Xiang, S.; Zhou, J.; Peng, X.; Hai, Y.; Wang, Y.; Li, S.; Wei, S. A putative effector UvHrip1 inhibits BAX-triggered cell death in Nicotiana benthamiana, and infection of Ustilaginoidea virens suppresses defense-related genes expression. PeerJ 2020, 8, e9354. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wu, K.; Yao, J.; Li, S.; Wang, X.; Huang, L.; Kang, Z. PSTha5a23, a candidate effector from the obligate biotrophic pathogen Puccinia striiformis f. sp. tritici, is involved in plant defense suppression and rust pathogenicity. Environ. Microbiol. 2017, 19, 1717–1729. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Tang, C.; Wang, X.; Sun, S.; Zhao, J.; Kang, Z.; Wang, X. An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat. Commun. 2019, 10, 5571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutra, D.; Agrawal, N.; Ghareeb, H.; Schirawski, J. Screening of Secreted Proteins of Sporisorium reilianum f. sp. zeae for Cell Death Suppression in Nicotiana benthamiana. Front. Plant Sci. 2020, 11, 95. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Yang, J.; Fang, A.; Wang, J.; Li, D.; Li, Y.; Wang, S.; Cui, F.; Yu, J.; Liu, Y.; et al. The essential effector SCRE1 in Ustilaginoidea virens suppresses rice immunity via a small peptide region. Mol. Plant Pathol. 2020, 21, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Derevnina, L.; Dagdas, Y.F.; de la Concepcion, J.C.; Białas, A.; Kellner, R.; Petre, B.; Domazakis, E.; Du, J.; Wu, C.-H.; Lin, X.; et al. Nine things to know about elicitins. New Phytol. 2016, 212, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Han, Y.; Qu, M.; Chen, J.; Chen, X.; Geng, X.; Wang, Z.; Chen, S. Apoplastic cell death-inducing proteins of filamentous plant pathogens: Roles in plant-pathogen interactions. Front. Genet. 2020, 11, 661. [Google Scholar] [CrossRef]
- Takken, F.; Rep, M. The arms race between tomato and Fusarium oxysporum. Mol. Plant Pathol. 2010, 11, 309–314. [Google Scholar] [CrossRef]
- Olivain, C.; Humbert, C.; Nahalkova, J.; Fatehi, J.; L’Haridon, F.; Alabouvette, C. Colonization of tomato root by pathogenic and nonpathogenic Fusarium oxysporum strains inoculated together and separately into the soil. Appl. Environ. Microbiol. 2006, 72, 1523–1531. [Google Scholar] [CrossRef] [Green Version]
- Rep, M.; Van Der Does, H.C.; Meijer, M.; Van Wijk, R.; Houterman, P.M.; Dekker, H.L.; De Koster, C.G.; Cornelissen, B.J.C. A small, cysteine-rich protein secreted by Fusarium oxysporum during colonization of xylem vessels is required for I-3-mediated resistance in tomato. Mol. Microbiol. 2004, 53, 1373–1383. [Google Scholar] [CrossRef]
- Houterman, P.M.; Speijer, D.; Dekker, H.L.; de Koster, C.G.; Cornelissen, B.J.C.; Rep, M. The mixed xylem sap proteome of Fusarium oxysporum-infected tomato plants. Mol. Plant Pathol. 2007, 8, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.M.; Houterman, P.M.; Schreiver, I.; Ma, L.; Amyotte, S.; Chellappan, B.; Boeren, S.; Takken, F.L.W.; Rep, M. MITEs in the promoters of effector genes allow prediction of novel virulence genes in Fusarium oxysporum. BMC Genom. 2013, 14, 119. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.-J.; Van Der Does, H.C.; Borkovich, K.A.; Coleman, J.J.; Daboussi, M.-J.; Di Pietro, A.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B.; et al. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 2010, 464, 367–373. [Google Scholar] [CrossRef]
- Li, J.; Fokkens, L.; Conneely, L.J.; Rep, M. Partial pathogenicity chromosomes in Fusarium oxysporum are sufficient to cause disease and can be horizontally transferred. Environ. Microbiol. 2020, 22, 4985–5004. [Google Scholar] [CrossRef]
- Gawehns, F.; Ma, L.; Bruning, O.; Houterman, P.M.; Boeren, S.; Cornelissen, B.J.C.; Rep, M.; Takken, F.L.W. The effector repertoire of Fusarium oxysporum determines the tomato xylem proteome composition following infection. Front. Plant Sci. 2015, 6, 967. [Google Scholar] [CrossRef] [PubMed]
- Houterman, P.M.; Ma, L.; van Ooijen, G.; de Vroomen, M.J.; Cornelissen, B.J.; Takken, F.L.; Rep, M. The effector protein Avr2 of the xylem-colonizing fungus Fusarium oxysporum activates the tomato resistance protein I-2 intracellularly. Plant J. 2009, 58, 970–978. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Houterman, P.M.; Gawehns, F.; Cao, L.; Sillo, F.; Richter, H.; Clavijo-Ortiz, M.J.; Schmidt, S.M.; Boeren, S.; Vervoort, J.; et al. The AVR2-SIX5 gene pair is required to activate I-2-mediated immunity in tomato. New Phytol. 2015, 208, 507–518. [Google Scholar] [CrossRef]
- Gawehns, F.; Houterman, P.M.; Ichou, F.A.; Michielse, C.B.; Hijdra, M.; Cornelissen, B.J.; Rep, M.; Takken, F.L.W. The Fusarium oxysporum effector Six6 contributes to virulence and suppresses I-2-mediated cell death. Mol. Plant Microbe Interact. 2014, 27, 336–348. [Google Scholar] [CrossRef] [Green Version]
- Vlaardingerbroek, I.; Beerens, B.; Rose, L.; Fokkens, L.; Cornelissen, B.J.C.; Rep, M. Exchange of core chromosomes and horizontal transfer of lineage-specific chromosomes in Fusarium oxysporum. Environ. Microbiol. 2016, 18, 3702–3713. [Google Scholar] [CrossRef]
- Ayukawa, Y.; Asai, S.; Gan, P.; Tsushima, A.; Ichihashi, Y.; Shibata, A.; Komatsu, K.; Houterman, P.M.; Rep, M.; Shirasu, K.; et al. A pair of effectors encoded on a conditionally dispensable chromosome of Fusarium oxysporum suppress host-specific immunity. Commun. Biol. 2021, 4, 707. [Google Scholar] [CrossRef]
- Gonzalez-Cendales, Y.; Catanzariti, A.-M.; Baker, B.; Mcgrath, D.J.; Jones, D.A. Identification of I-7 expands the repertoire of genes for resistance to Fusarium wilt in tomato to three resistance gene classes. Mol. Plant Pathol. 2015, 17, 448–463. [Google Scholar] [CrossRef]
- Simons, G.; Groenendijk, J.; Wijbrandi, J.; Reijans, M.; Groenen, J.; Diergaarde, P.; van der Lee, T.; Bleeker, M.; Onstenk, J.; de Both, M. Dissection of the fusarium I2 gene cluster in tomato reveals six homologs and one active gene copy. Plant Cell 1998, 10, 1055–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catanzariti, A.M.; Lim, G.T.T.; Jones, D.A. The tomato I-3 gene: A novel gene for resistance to Fusarium wilt disease. New Phytol. 2015, 207, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Catanzariti, A.M.; Do, H.T.; Bru, P.; de Sain, M.; Thatcher, L.F.; Rep, M.; Jones, D.A. The tomato I gene for Fusarium wilt resistance encodes an atypical leucine-rich repeat receptor-like protein whose function is nevertheless dependent on SOBIR1 and SERK3/BAK1. Plant J. 2017, 89, 1195–1209. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, S.; Taylor, A.; Jackson, A.C.; Armitage, A.D.; Bates, H.J.; Mead, A.; Harrison, R.J.; Clarkson, J.P. Identification and Expression of Secreted In Xylem Pathogenicity Genes in Fusarium oxysporum f. sp. pisi. Front. Microbiol. 2021, 12, 593140. [Google Scholar] [CrossRef]
- Vleeshouwers, V.G.; Oliver, R.P. Effectors as tools in disease resistance breeding against biotrophic, hemibiotrophic, and necrotrophic plant pathogens. Mol. Plant Microbe Interact. 2014, 27, 196–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domazakis, E.; Lin, X.; Aguilera-Galvez, C.; Wouters, D.; Bijsterbosch, G.; Wolters, P.J.; Vleeshouwers, V.G.A.A. Effectoromics-Based Identification of Cell Surface Receptors in Potato. In Plant Pattern Recognition Receptors: Methods and Protocols; Shan, L., He, P., Eds.; Springer: New York, NY, USA, 2017; pp. 337–353. [Google Scholar]
- Mes, J.J.; Wit, R.; Testerink, C.S.; de Groot, F.; Haring, M.A.; Cornelissen, B.J. Loss of avirulence and reduced pathogenicity of a gamma-irradiated mutant of Fusarium oxysporum f. sp. lycopersici. Phytopathology 1999, 89, 1131–1137. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2^(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinforma. Biomath. 2013, 3, 71–85. [Google Scholar]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Shao, M.; Kingsford, C. Accurate assembly of transcripts through phase-preserving graph decomposition. Nat. Biotechnol. 2017, 35, 1167–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testa, A.C.; Hane, J.K.; Ellwood, S.R.; Oliver, R.P. CodingQuarry: Highly accurate hidden Markov model gene prediction in fungal genomes using RNA-seq transcripts. BMC Genom. 2015, 16, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoff, K.J.; Lomsadze, A.; Borodovsky, M.; Stanke, M. Whole-Genome Annotation with BRAKER. Methods Mol. Biol. 2019, 1962, 65–95. [Google Scholar] [PubMed]
- Waterhouse, R.M.; Seppey, M.; Simão, F.A.; Manni, M.; Ioannidis, P.; Klioutchnikov, G.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO Applications from Quality Assessments to Gene Prediction and Phylogenomics. Mol. Biol. Evol. 2018, 35, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.P.; Tang, H.B.; DeBarry, J.D.; Tan, X.; Li, J.P.; Wang, X.Y.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Kurtz, S.; Phillippy, A.; Delcher, A.L.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S.L. Versatile and open software for comparing large genomes. Genome Biol. 2004, 5, R12. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Neu, E.; Debener, T. Prediction of the Diplocarpon rosae secretome reveals candidate genes for effectors and virulence factors. Fungal Biol. 2019, 123, 231–239. [Google Scholar] [CrossRef]
- Armenteros, A.J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- de Castro, E.; Sigrist, C.J.A.; Gattiker, A.; Bulliard, V.; Langendijk-Genevaux, P.S.; Gasteiger, E.; Bairoch, A.; Hulo, N. ScanProsite: Detection of PROSITE signature matches and ProRule-associated functional and structural residues in proteins. Nucleic Acids Res. 2006, 34, W362–W365. [Google Scholar] [CrossRef]
- Pierleoni, A.; Martelli, P.L.; Casadio, R. PredGPI: A GPI-anchor predictor. BMC Bioinform. 2008, 9, 392. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods 2005, 1, 13. [Google Scholar] [CrossRef] [Green Version]
- de Rybel, B.; van den Berg, W.; Lokerse, A.; Liao, C.Y.; van Mourik, H.; Moller, B.; Peris, C.L.; Weijers, D. A versatile set of ligation-independent cloning vectors for functional studies in plants. Plant Physiol. 2011, 156, 1292–1299. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Lukasik, E.; Gawehns, F.; Takken, F.L.W. The use of agroinfiltration for transient expression of plant resistance and fungal effector proteins in Nicotiana benthamiana leaves. Methods Mol. Biol. 2012, 835, 61–74. [Google Scholar]
- Ma, L.; Djavaheri, M.; Wang, H.; Larkan, N.J.; Haddadi, P.; Beynon, E.; Gropp, G.; Borhan, M.H. Leptosphaeria maculans effector protein AvrLm1 modulates plant immunity by enhancing MAP kinase 9 phosphorylation. iScience 2018, 3, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Wang, Y.; Chen, T.; Lin, Y.; Luo, C. Functional Evaluation of the Signal Peptides of Secreted Proteins. BIO Protoc. 2018, 8, e2839. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved prediction of fungal effector proteins from secretomes with EffectorP 2.0. Mol. Plant Pathol. 2018, 19, 2094–2110. [Google Scholar] [CrossRef] [Green Version]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2020, 18, 1247–1256. [Google Scholar] [CrossRef] [Green Version]
- Bernoux, M.; Ve, T.; Williams, S.; Warren, C.; Hatters, D.; Valkov, E.; Zhang, X.; Ellis, J.G.; Kobe, B.; Dodds, P.N. Structural and functional analysis of a plant resistance protein TIR domain reveals interfaces for self-association, signaling, and autoregulation. Cell Host Microbe 2011, 9, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Hogenhout, S.A.; Van der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging concepts in effector biology of plant-associated organisms. Mol. Plant Microbe Interact. 2009, 22, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Lievens, B.; Houterman, P.M.; Rep, M. Effector gene screening allows unambiguous identification of Fusarium oxysporum f. sp. lycopersici races and discrimination from other formae speciales. FEMS Microbiol Lett. 2009, 300, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Lafond, M.; Navarro, D.; Haon, M.; Couturier, M.; Berrin, J.G. Characterization of a broad-specificity beta-glucanase acting on beta-(1,3)-, beta-(1,4)-, and beta-(1,6)-glucans that defines a new glycoside hydrolase family. Appl. Environ. Microbiol. 2012, 78, 8540–8546. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Yoshida, M.; Tamura, M.; Tanaka, Y.; Umezawa, K.; Nishikawa, A.; Tonozuka, T. Crystal structure of the N-terminal domain of a glycoside hydrolase family 131 protein from Coprinopsis cinerea. FEBS Lett. 2013, 587, 2193–2198. [Google Scholar] [CrossRef] [Green Version]
- Anasontzis, G.E.; Lebrun, M.H.; Haon, M.; Champion, C.; Kohler, A.; Lenfant, N.; Martin, F.; O’Connell, R.J.; Riley, R.; Grigoriev, I.V.; et al. Broad-specificity GH131 beta-glucanases are a hallmark of fungi and oomycetes that colonize plants. Environ. Microbiol. 2019, 21, 2724–2739. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Jelsch, C.; Ostergaard, P.; Rypniewski, W.; Lamzin, V.S. Trypsin revisited: Crystallography AT (SUB) atomic resolution and quantum chemistry revealing details of catalysis. J. Biol. Chem. 2003, 278, 43357–43362. [Google Scholar] [CrossRef] [Green Version]
- Rypniewski, W.; Hastrup, S.; Betzel, C.; Dauter, M.; Dauter, Z.; Papendorf, G.; Branner, S.; Wilson, K. The sequence and X-ray structure of the trypsin from Fusarium oxysporum. Protein Eng. 1993, 6, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stork, I.; Gartemann, K.H.; Burger, A.; Eichenlaub, R. A family of serine proteases of Clavibacter michiganensis subsp. michiganensis: ChpC plays a role in colonization of the host plant tomato. Mol. Plant Pathol. 2008, 9, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Chalupowicz, L.; Barash, I.; Reuven, M.; Dror, O.; Sharabani, G.; Gartemann, K.H.; Eichenlaub, R.; Sessa, G.; Manulis-Sasson, S. Differential contribution of Clavibacter michiganensis ssp. michiganensis virulence factors to systemic and local infection in tomato. Mol. Plant Pathol. 2017, 18, 336–346. [Google Scholar] [CrossRef]
- Jahr, H.; Dreier, J.; Meletzus, D.; Bahro, R.; Eichenlaub, R. The endo-beta-1,4-glucanase CelA of Clavibacter michiganensis subsp. michiganensis is a pathogenicity determinant required for induction of bacterial wilt of tomato. Mol. Plant Microbe Interact. 2020, 13, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, L.M.; Ohno, Y.; Miura, Y.; Kawakita, K.; Doke, N. Host Selective Suppression by Water-Soluble Glucans from Phytophthora spp. of Hypersensitive Cell Death of Suspension-Cultures Cells from Some Solanaceous Plants Caused by Hyphal Wall Elicitors of the Fungi. Jpn. J. Phytopathol. 1992, 58, 664–670. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Magne, M.; Chen, S.; Côté, O.; Stare, B.G.; Obradovic, N.; Jamshaid, L.; Wang, X.; Bélair, G.; Moffett, P. Analysis of putative apoplastic effectors from the nematode, Globodera rostochiensis, and identification of an expansin-like protein that can induce and suppress host defenses. PLoS ONE 2015, 10, e0115042. [Google Scholar] [CrossRef]
- Carlile, A.J.; Bindschedler, L.V.; Bailey, A.M.; Bowyer, P.; Clarkson, J.M.; Cooper, R.M. Characterization of SNP1, a cell wall-degrading trypsin, produced during infection by Stagonospora nodorum. Mol. Plant. Microbe Interact. 2000, 13, 538–550. [Google Scholar] [CrossRef] [Green Version]
- Hao, G.; McCormick, S.; Vaughan, M.M.; Naumann, T.A.; Kim, H.-S.; Proctor, R.; Kelly, A.; Ward, T.J. Fusarium graminearum arabinanase (Arb93B) enhances wheat head blight susceptibility by suppressing plant immunity. Mol. Plant Microbe Interact. 2019, 32, 888–898. [Google Scholar] [CrossRef]
- Hao, G.; McCormick, S.; Usgaard, T.; Tiley, H.; Vaughan, M.M. Characterization of three Fusarium graminearum effectors and their roles during Fusarium head blight. Front. Plant Sci. 2020, 11, 579553. [Google Scholar] [CrossRef]
- Rose, J.K.; Ham, K.S.; Darvill, A.G.; Albersheim, P. Molecular cloning and characterization of glucanase inhibitor proteins: Coevolution of a counterdefense mechanism by plant pathogens. Plant Cell 2002, 14, 1329–1345. [Google Scholar] [CrossRef] [Green Version]
- Bozkurt, T.O.; Schornack, S.; Banfield, M.J.; Kamoun, S. Oomycetes, effectors, and all that jazz. Curr. Opin. Plant Biol. 2012, 15, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wang, Y.; Tian, M.; Dai, K.; Zheng, W.; Liu, Z.; Yang, S.; Liu, X.; Shi, D.; Zhang, H.; et al. Fg12 ribonuclease secretion contributes to Fusarium graminearum virulence and induces plant cell death. J. Integr. Plant Biol. 2021, 63, 365–377. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Lineage-Specific Region | Chromosome | 2020 Contig | Start | End | Strand | Introns | Protein Length | Cys Residues | Protein Domain/Homology |
|---|---|---|---|---|---|---|---|---|---|---|
| SIX9 | YES | 14 | 14 | 809,946 | 810,290 | - | 0 | 114 | 6 | |
| SIX6 | YES | 14 | 14 | 962,344 | 963,070 | - | 1 | 225 | 9 | |
| SIX11 | YES | 14 | 14 | 1,007,574 | 1,007,906 | - | 0 | 110 | 8 | |
| FOXG_17276 * | YES | 14 | 14 | 1,450,230 | 1,450,661 | - | 0 | 143 | 6 | LysM domain |
| PSL2 * | YES | 14 | 14 | 1,486,868 | 1,487,339 | - | 3 | 106 | 9 | PSE1 homologue |
| SIX14 | YES | 14 | 14 | 1,489,136 | 1,489,452 | + | 1 | 88 | 6 | |
| SIX1 | YES | 14 | 14 | 1,508,020 | 1,508,874 | - | 0 | 284 | 8 | |
| SIX2 | YES | 14 | 14 | 1,515,507 | 1516205 | + | 0 | 232 | 8 | |
| SIX3 | YES | 14 | 14 | 1,620,196 | 1,620,687 | - | 0 | 163 | 3 | |
| SIX5 | YES | 14 | 14 | 1,621,885 | 1,622,410 | + | 3 | 119 | 7 | |
| SIX13 | YES | 14 | 14 | 1,721,250 | 1,722,191 | + | 1 | 293 | 12 | |
| SIX10 | YES | 14 | 14 | 1,929,603 | 1,930,122 | - | 1 | 149 | 2 | |
| SIX12 | YES | 14 | 14 | 1,931,346 | 1,931,777 | - | 1 | 127 | 10 | |
| SIX7 | YES | 14 | 14 | 1,934,097 | 1,934,727 | + | 1 | 163 | 2 | |
| FOXGR_015522 | YES | 14 | 14 | 1,996,473 | 1,996,874 | - | 2 | 79 | 9 | |
| SIX13 | YES | 14 | 14 | 2,018,772 | 2,019,713 | + | 1 | 293 | 12 | |
| FOXGR_015533 * | YES | 14 | 14 | 2,033,852 | 2,034,196 | - | 0 | 114 | 3 | Fol-EC3 |
| FOXG_10949 * | NO | 1 | 0 | 498,344 | 498,719 | - | 1 | 107 | 8 | hydrophobin |
| FOXG_10950 * | NO | 1 | 0 | 500,457 | 501,074 | + | 3 | 152 | 9 | hydrophobin |
| FOXG_11033 * | NO | 1 | 0 | 722,540 | 723,369 | - | 3 | 226 | 0 | |
| FOXG_05750 | NO | 2 | 53 | 540,805 | 541,602 | + | 0 | 265 | 12 | LysM domain x2 |
| FOXG_05755 * | NO | 2 | 53 | 549,873 | 550,346 | + | 0 | 157 | 3 | |
| FOXG_18699 * | NO | 4 | 2 | 3,264,675 | 3,264,965 | + | 0 | 96 | 10 | |
| FOXGR_007323 * | NO | 5 | 3 | 2,372,326 | 2,372,640 | - | 1 | 86 | 0 | |
| FOXG_10672 | NO | 7 | 46 | 413,827 | 414,423 | + | 0 | 198 | 13 | PAN/Apple domain x2 |
| FOXG_04863 * | NO | 7 | 46 | 3,884,224 | 3,885,126 | + | 0 | 300 | 8 | |
| FOXG_04805 * | NO | 7 | 46 | 4,051,134 | 4,051,586 | + | 1 | 132 | 8 | |
| FOXG_02829 * | NO | 8 | 50 | 561,065 | 561,514 | - | 0 | 149 | 16 | |
| FOXGR_021626 * | NO | 8 | 50 | 3,119,908 | 3,120,893 | + | 2 | 294 | 3 | Glucanase—Fol-EC14 |
| SIX8 | YES | 8 | 50 | 4,122,866 | 4,123,541 | - | 2 | 141 | 2 | |
| PSL1 * | YES | 8 | 50 | 4,124,187 | 4,124,677 | + | 3 | 111 | 9 | |
| FOXG_08899 * | NO | 9 | 5 | 3,212,288 | 3,212,782 | + | 1 | 148 | 1 | |
| SIX8 | YES | 10 | 7 | 1363 | 1768 | + | 2 | 141 | 2 | |
| FOXGR_010884 | NO | 10 | 7 | 444,492 | 445,286 | + | 0 | 264 | 0 | |
| FOXG_11745 * | NO | 10 | 7 | 655,322 | 655,924 | + | 1 | 183 | 4 | phospholipase A2 |
| SIX8 | YES | 10 | 7 | 3,412,328 | 3,413,003 | - | 2 | 141 | 2 | |
| PSL1 * | YES | 10 | 7 | 3,413,649 | 3,414,139 | + | 3 | 111 | 9 | |
| FOXG_10138 | NO | 11 | 61 | 398,337 | 399,122 | + | 0 | 261 | 2 | peptidase G1 family |
| FOXGR_025639 * | NO | 11 | 61 | 507,487 | 507,728 | + | 1 | 61 | 2 | |
| FOXG_16600 * | NO | 11 | 61 | 2,173,630 | 2,174,176 | - | 1 | 164 | 0 | |
| FOXG_13233 * | NO | 12 | 12 | 668,427 | 668,929 | + | 2 | 131 | 4 | ribonuclease F1—Fol-EC19 |
| FOXG_13248 * | NO | 12 | 12 | 703,080 | 703,929 | + | 2 | 248 | 6 | Trypsin—Fol-EC20 |
| FOXG_14607 * | NO | 12 | 12 | 1,873,367 | 1,874,245 | + | 1 | 275 | 7 | metalloprotease MEP1 |
| FOXG_14684 * | NO | 12 | 12 | 2,089,656 | 2,090,270 | - | 2 | 168 | 14 | |
| PSL1 * | YES | - | 9 | 576 | 1066 | - | 3 | 111 | 9 | |
| SIX8 | YES | - | 10 | 224 | 783 | + | 2 | 141 | 2 | |
| PSL1 * | YES | - | 44 | 18,988 | 19,478 | - | 3 | 111 | 9 | |
| SIX8 | YES | - | 44 | 20,124 | 20,683 | + | 2 | 141 | 2 | |
| PSL1 * | YES | - | 45 | 6024 | 6515 | - | 3 | 111 | 9 | |
| PSL1 * | YES | - | 58 | 1,499,796 | 1,500,286 | - | 3 | 111 | 9 | |
| SIX8 | YES | - | 58 | 1,500,932 | 1,501,607 | + | 2 | 141 | 2 | |
| PSL1 * | YES | - | 59 | 3963 | 4453 | - | 3 | 111 | 9 | |
| SIX8 | YES | - | 59 | 5099 | 5658 | + | 2 | 141 | 2 | |
| PSL1 * | YES | - | 60 | 7085 | 7575 | - | 3 | 111 | 9 | |
| SIX8 | YES | - | 60 | 8221 | 8896 | + | 2 | 141 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Fang, X.; Wang, D.; Jones, D.A.; Ma, L. Transcriptome Analysis of Fusarium–Tomato Interaction Based on an Updated Genome Annotation of Fusarium oxysporum f. sp. lycopersici Identifies Novel Effector Candidates That Suppress or Induce Cell Death in Nicotiana benthamiana. J. Fungi 2022, 8, 672. https://doi.org/10.3390/jof8070672
Sun X, Fang X, Wang D, Jones DA, Ma L. Transcriptome Analysis of Fusarium–Tomato Interaction Based on an Updated Genome Annotation of Fusarium oxysporum f. sp. lycopersici Identifies Novel Effector Candidates That Suppress or Induce Cell Death in Nicotiana benthamiana. Journal of Fungi. 2022; 8(7):672. https://doi.org/10.3390/jof8070672
Chicago/Turabian StyleSun, Xizhe, Xiangling Fang, Dongmei Wang, David A. Jones, and Lisong Ma. 2022. "Transcriptome Analysis of Fusarium–Tomato Interaction Based on an Updated Genome Annotation of Fusarium oxysporum f. sp. lycopersici Identifies Novel Effector Candidates That Suppress or Induce Cell Death in Nicotiana benthamiana" Journal of Fungi 8, no. 7: 672. https://doi.org/10.3390/jof8070672