
Exploring Yeast Diversity to Produce Lipid-Based Biofuels from Agro-Forestry and Industrial Organic Residues



1
iBB—Institute for Bioengineering and Biosciences, Instituto Superior Técnico, Universidade de Lisboa, Av. Rovisco Pais, 1, 1049-001 Lisbon, Portugal
2
Department of Bioengineering, Instituto Superior Técnico, Universidade de Lisboa, Av. Rovisco Pais, 1, 1049-001 Lisbon, Portugal
3
i4HB—Institute for Health and Bioeconomy, Instituto Superior Técnico, Universidade de Lisboa, Av. Rovisco Pais, 1, 1049-001 Lisbon, Portugal
4
BIOREF—Collaborative Laboratory for Biorefineries, Rua da Amieira, Apartado 1089, São Mamede de Infesta, 4465-901 Matosinhos, Portugal
*
Author to whom correspondence should be addressed.
J. Fungi 2022, 8(7), 687; https://doi.org/10.3390/jof8070687
Submission received: 8 June 2022
/
Revised: 24 June 2022
/
Accepted: 27 June 2022
/
Published: 29 June 2022
(This article belongs to the Special Issue Yeast Biorefineries)
Abstract
:Exploration of yeast diversity for the sustainable production of biofuels, in particular biodiesel, is gaining momentum in recent years. However, sustainable, and economically viable bioprocesses require yeast strains exhibiting: (i) high tolerance to multiple bioprocess-related stresses, including the various chemical inhibitors present in hydrolysates from lignocellulosic biomass and residues; (ii) the ability to efficiently consume all the major carbon sources present; (iii) the capacity to produce lipids with adequate composition in high yields. More than 160 non-conventional (non-Saccharomyces) yeast species are described as oleaginous, but only a smaller group are relatively well characterised, including Lipomyces starkeyi, Yarrowia lipolytica, Rhodotorula toruloides, Rhodotorula glutinis, Cutaneotrichosporon oleaginosus and Cutaneotrichosporon cutaneum. This article provides an overview of lipid production by oleaginous yeasts focusing on yeast diversity, metabolism, and other microbiological issues related to the toxicity and tolerance to multiple challenging stresses limiting bioprocess performance. This is essential knowledge to better understand and guide the rational improvement of yeast performance either by genetic manipulation or by exploring yeast physiology and optimal process conditions. Examples gathered from the literature showing the potential of different oleaginous yeasts/process conditions to produce oils for biodiesel from agro-forestry and industrial organic residues are provided.
1. Introduction
The sustainable production and use of renewable fuels to enable the transition to a low-carbon and more sustainable economy have been promoted in recent years in response to the global climate crisis and the growing energy needs [1,2]. Liquid biofuels play a central role in that transition, in particular biodiesel which is considered an ideal candidate for the replacement of petroleum-derived diesel due to its high cetane number and flash point and the possibility to be used in any compression-ignition engine without the need for modification [3,4]. Biodiesel results from the reaction of one triacylglycerol molecule (consisting of three long-chain fatty acids attached to glycerol) with three alcohol molecules (usually methanol or ethanol) to produce three biodiesel molecules, i.e., methyl esters or ethyl esters, and one glycerol molecule [5,6]. Biodiesel is currently mainly produced through the transesterification of oils, in particular vegetable oils (including edible oils) or animal fats [7]. However, the use of these sources is of concern as they can compete with the food oil market and implicate increased deforestation and biodiversity loss by intensifying the use of land for cultivation [8]. To overcome these disadvantages, microbial lipids have gained attention in recent years. Besides not competing with food, their production is not susceptible to seasonal changes, the growth of microorganisms is much faster and microbial oils production can be optimised and controlled in bioreactors, with their composition being very similar to that of vegetable oils [9,10,11]. Among the best microbial candidates capable of producing lipids in high concentrations and with appropriate characteristics are oleaginous yeasts, described as those capable of accumulating more than 20% of cell dry weight in lipids [11,12]. In addition to the aforementioned advantages, oleaginous yeasts also have the potential to metabolise diverse carbon sources of difficult catabolism, can exhibit high tolerance to a wider range of relevant bioprocess-associated stresses and have an unusual and specialised metabolism producing a wide and diverse repertoire of lipids, proteins and metabolites with high commercialisation potential [13,14,15,16,17]. Although there are over 160 yeast species described as oleaginous, only a small set of these species are relatively well characterised, including Lipomyces starkeyi, Yarrowia lipolytica, Rhodotorula glutinis, Rhodosporidium/Rhodotorula toruloides, Cutaneotrichosporon oleaginosus (previously classified as Cryptococcus curvatus or Trichosporon oleaginosus) and Cutaneotrichosporon cutaneum (formerly known as Trichosporon cutaneum) [18]. These non-Saccharomyces yeast species are referred to as non-conventional yeasts. However, based on their biotechnological interest and current intense research, it is anticipated that they will soon, if not already, stop being considered as such.
The economic viability of industrial lipid production bioprocesses depends on the performance of the yeast strain under optimised conditions and the efficient use of low-cost renewable raw materials, such as lignocellulosic biomasses [19,20]. Extensive screenings of yeast strains of various species available in culture collections and of new strains isolated for this purpose were carried out. Studies to improve selected yeast strains for better performance using lignocellulosic biomass hydrolysates or other interesting feedstocks with emphasis on diverse wastes were also performed. However, the use of genetic engineering techniques requires the availability of appropriate tools to be applied to the non-conventional yeast species of interest. If they are not available or not efficient enough, the exploration of other more traditional genetic improvement techniques is a possible approach. The optimisation of conditions for the production process (e.g., pH, temperature, medium and other culture conditions, type of reactors) is equally required.
This literature review presents recent results that support the idea that there is a huge interest and potential in several oleaginous yeast species/strains to generate oils for biodiesel production, in particular when produced from lignocellulosic biomasses from forestry and agriculture and industrial organic by-products/residues/wastes. The challenges faced by yeasts when cultivated in these feedstocks are discussed, including the catabolism of different carbon sources (C-sources), lipid biosynthetic pathways, and tolerance to the toxic compounds present and to other relevant stresses. Results gathered from recent literature concerning the production of yeast oils from different feedstocks, from a circular bio-economy perspective, are also provided.
2. Diversity of Oleaginous Yeasts
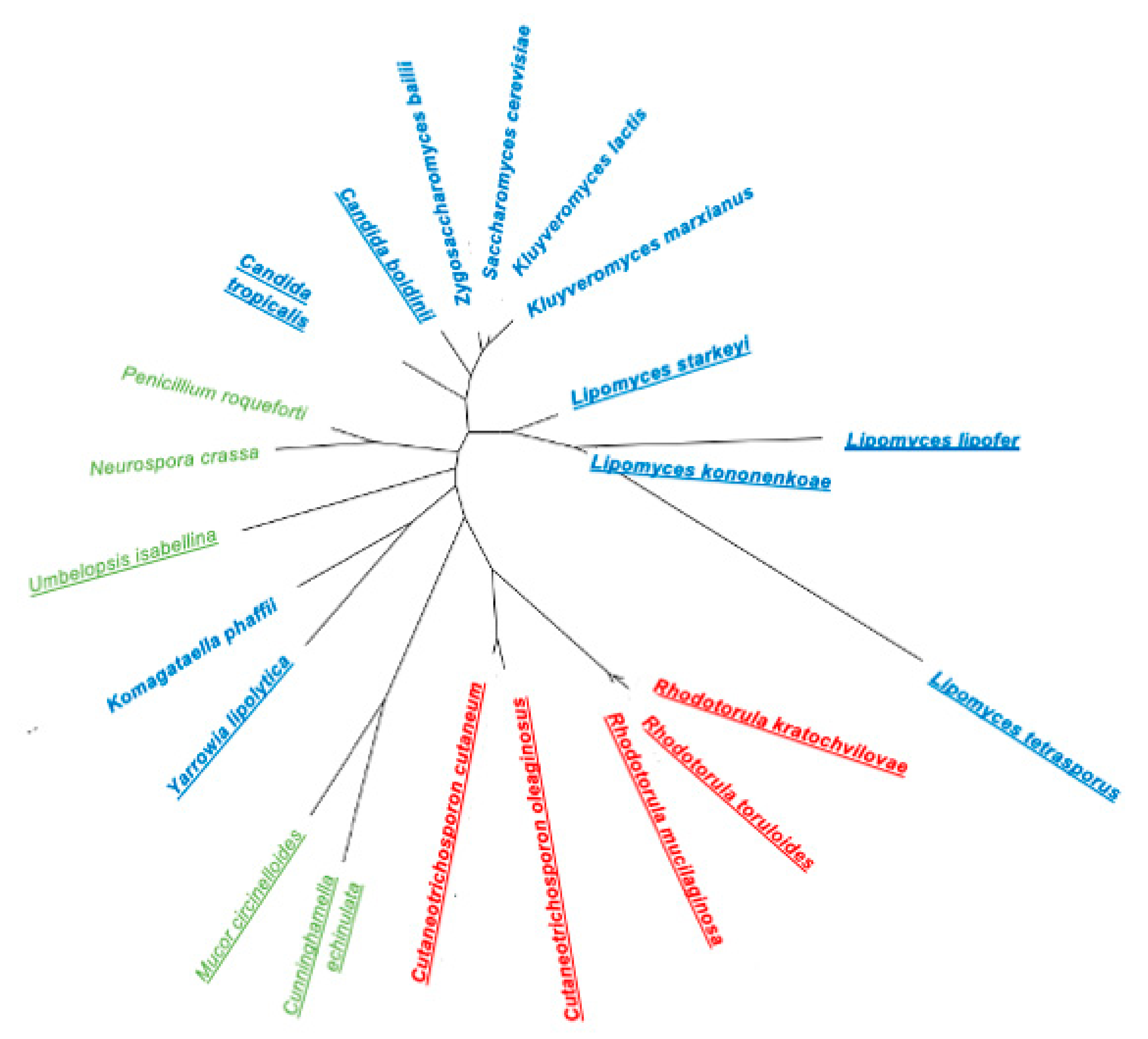
Oleaginous yeasts belong to the two phyla within the kingdom Fungi, Ascomycota and Basidiomycota. The diversity of oleaginous yeasts is observed in the phylogenetic tree prepared for biotechnologically relevant yeasts (Figure 1).
In the phylum Ascomycota, yeasts from the genera Lipomyces and Candida, and the species Yarrowia lipolytica were extensively studied due to their intrinsic lipid production potential [18,21,22,23]. There are sixteen species of the genus Lipomyces, with the Lipomyces starkeyi and Lipomyces tetrasporus species being isolated worldwide [24]. Besides being present in the soil, some species were also isolated from insect-associated habitats such as frass, decaying cactus tissues and tree fluxes [24]. L. starkeyi is the species of the Lipomyces genus with a larger number of published studies due to its high lipid productivity [11,25]. Y. lipolytica is a dimorphic yeast usually found in environments containing hydrophobic substrates, rich in alkanes and fats. It can be isolated from cheese, yoghurt, kefir, soy sauce, meat and shrimp salads [26]. The genome sequences of strains Y. lipolytica Po1f, commonly used for metabolic engineering, and the reference strain CLIB122, are available [27] and there are several synthetic biology tools for Y. lipolytica genetic manipulation [28]. They include DNA assembly techniques for synthetic biology, DNA parts for expression cassette construction, genome-editing techniques, and computational tools [28]. Regarding the Candida genus, the species Candida boidinii, Candida utilis, and Candida tropicalis were reported as oleaginous [22,29]. C. boidinii strains were isolated from natural environments (soil, seawater, sap fluxes of many sugar-rich tree species) or samples associated with human activities (wine fermentations or olive manufacturing), suggesting a biotechnological potential [30].
Regarding the phylum Basidiomycota, Rhodotorula (Rhodosporidium) toruloides, Cryptococcus curvatus and species of the genus Trichosporon are interesting lipid producers. Rhodotorula species are present in several habitats such as bark-beetles, tree exudates, plants and vegetables, soil, fresh water, coastal sediments and marine environments but were also isolated from clinical samples [31]. These yeasts, known as red yeasts, exhibit a red colour due to the production of carotenoids and can exist both in the yeast or in mycelial forms [18]. Remarkably, the species R. toruloides presents a huge potential as a workhorse for biotechnological applications [32]. One of the most extensively studied Cryptococcus species, C. curvatus (now, Cutaneotrichosporon oleaginosus), can accumulate up to 73% dry cell weight (DCW) in lipids [33]. It is distributed in nature and can be isolated from foodstuffs (raw milk, lettuce) and marine sediments [11]. The majority of Trichosporon strains were isolated from soil and milk whey samples [30] but some strains were also isolated from immunocompromised hosts. The potential pathogenicity may limit the use of this and other species for industrial applications.
The ability of oleaginous yeasts to grow in low-cost substrates can be related to the habitats from where these oleaginous yeasts are isolated and a considerable number of oleaginous yeasts are primarily found in soils, in particular L. starkeyi and L. tetrasporus, and some Cryptococcus and Trichosporon species [18]. The type of soil enhances the ability of the yeasts present there to consume a wide variety of substrates, such as lignocellulosic biomasses [24]. L. starkeyi, T. cutaneum and some species of Cryptococcus are tolerant to the major inhibitors present in lignocellulosic biomass hydrolysates, including acetic acid, furfural, and 5-hydroxymethylfurfural (HMF) [34,35,36,37,38]. Additionally, R. toruloides and R. mucilaginosa are able to consume the acid sugar galacturonic acid from hydrolysates from sugar beet pulp, a pectin-rich residue [39,40]. Xylose, the second most abundant sugar in lignocellulosic biomass hydrolysates, is used as a carbon source (C-source) for growth by all the aforementioned genus/species. Even though many Yarrowia isolates readily consume xylose [41], Y. lipolytica Po1-derived strains require genetic modifications to be able to use this pentose as a carbon source [42]. Glycerol can also be used as a C-source for lipid production by C. boidinii, C. curvatus, L. starkeyi, R. toruloides and Y. lipolytica [25,43,44]. Therefore, crude glycerol, a byproduct of the biodiesel industry is potentially an interesting substrate for oil production by all these yeast species, especially for those also capable of catabolising and tolerating methanol, a major contaminant in crude glycerol [45].
3. Oleaginous Yeasts for Sustainable Biodiesel Production
3.1. Biosynthesis of Yeast Oils
Biodiesel is produced through the transesterification of oils involving the conversion of triacylglycerols (TAGs) to fatty acid methyl (or ethyl) esters (FAMEs) [46]. The structure, including the chain length of the fatty acids produced, can vary and determines the quality of the biofuel. Oleaginous yeasts accumulate non-polar lipids, such as steryl esters (SEs) and triacylglycerols (TAGs), in particular mystiric acid [C14:0], palmitic acid [C16:0], stearic acid [C18:0], oleic acid [C18:1], linoleic acid [C18:2], and linolenic acid [C18:3] [19]. Fatty acid-lipid profiles can vary depending on culture conditions and yeast species/strains. Remarkably, the FAMEs derived from oleaginous yeast have similar properties to more traditional sources derived from food crops such as rapeseed, palm or sunflower oils [25,46,47]. Oleaginous yeasts can produce different oils, with oleic acid (18:1) being the oil that is produced in higher titers. For example, in L. starkeyi NBRC 10381, oleic acid content represents nearly 74% of the total produced oils [48]. Since oleic acid is the lipid that best meets the criteria to obtain biodiesel with the best properties [49,50], yeasts are highly interesting cell factories for sustainable biodiesel production.
3.2. Triacylglycerol (TAG) Metabolism in Yeasts
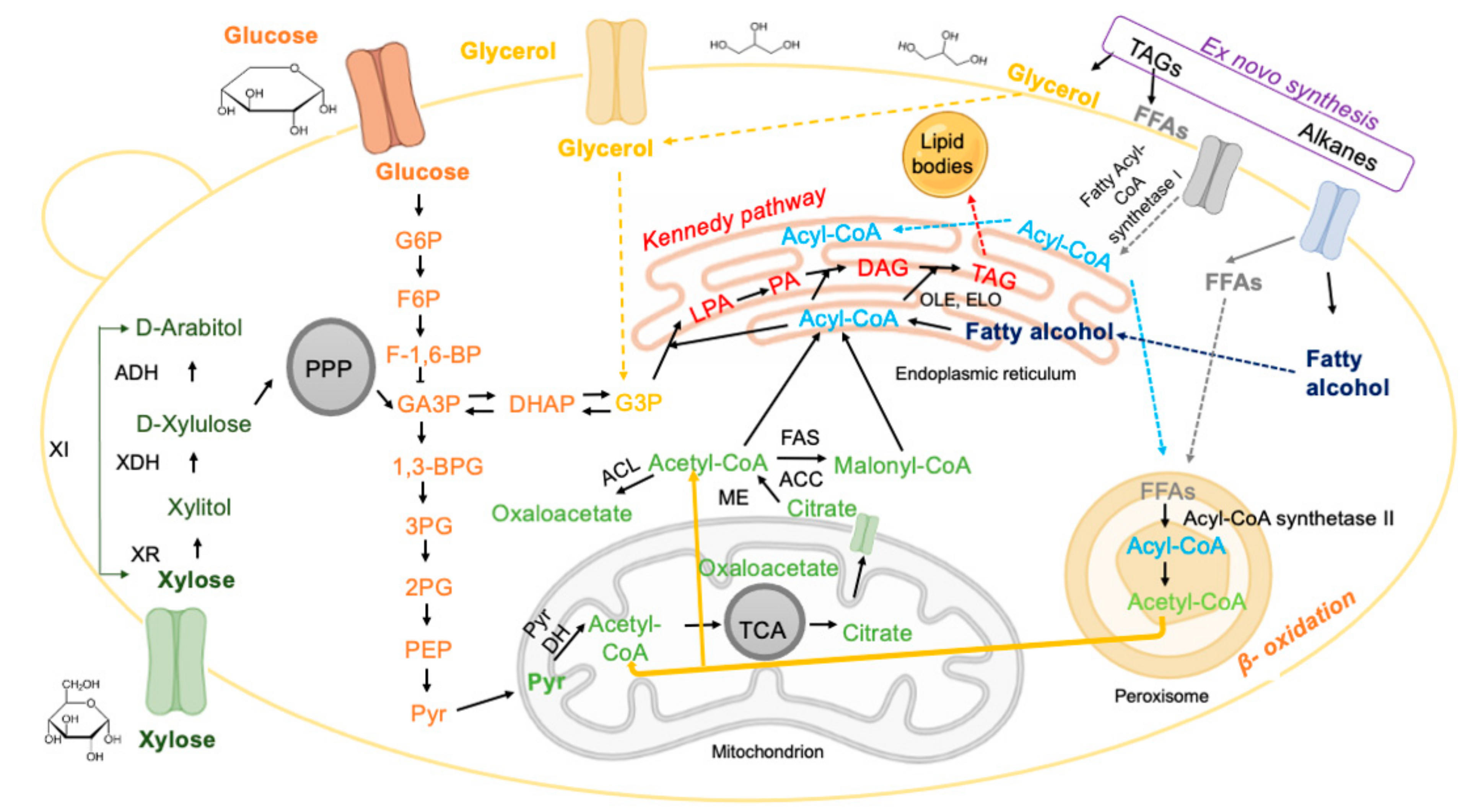
Triacylglycerols (TAGs) can be synthesised and accumulated by: (i) de novo synthesis, when the precursors of fatty acid biosynthesis are produced from different carbon sources (e.g., sugars, weak acids, glycerol), or (ii) ex novo synthesis, based on the fatty acids present in the culture medium (Figure 2).
3.2.1. De Novo Synthesis
The de novo synthesis pathway of TAGs is triggered by the limitation of the nitrogen source when the carbon source is in excess, i.e., in a culture medium with a high C/N ratio [19,42], as detailed in Section 6.2. The biochemical reactions involved in the de novo synthesis of lipids are schematised in Figure 2. Under nitrogen-limiting conditions, adenosine monophosphate deaminase (AMPD) is activated and catalyses the hydrolysis of adenosine monophosphate (AMP) to inosine monophosphate (IMP) and ammonia, thereby providing nitrogen to the cell [51]. At low AMP concentrations, isocitrate dehydrogenase activity decreases [52] and the tricarboxylic acid cycle (TCA) becomes dysregulated, leading to isocitrate accumulation. Through the action of the enzyme aconitase, isocitrate and citrate levels balance and citrate is transported from the mitochondria to the cytosol via malate/citrate antiport [53]. Once in the cytosol, citrate is converted into acetyl-CoA and oxaloacetate by ATP citrate lyase (ACL), a key enzyme during lipogenesis. The next steps include (i) the conversion of oxaloacetate to malate, and (ii) the cleavage of malate into pyruvate and NADPH. The pyruvate enters the pyruvate dehydrogenase complex (PDH cycle) where pyruvate is decarboxylated, producing acetyl-CoA, the key molecule for lipid production, as well as NADH and carbon dioxide. Fatty acid synthesis begins with cytosolic acetyl-CoA being condensed into malonyl-CoA, in a reaction catalysed by acetyl-CoA carboxylase. Acetyl-CoA and malonyl-CoA are condensed to acyl-CoA by the fatty acid synthase complex (FAS). NADPH is used as a reducing cofactor by fatty acid synthase and two molecules of NADPH are used in each step of acyl-CoA chain elongation. The most common chain length of naturally synthesised acyl-CoAs has 16 or 18 carbon atoms. The C16:0 and C18:0 molecules are routed to the endoplasmic reticulum (ER) in order to proceed to the elongation and desaturation steps [54]. The synthesis of TAGs is carried out via the Kennedy pathway, starting with glycerol-3-phosphate (G3P) from glycolysis and acyl-CoA [55]. Two fatty acids (FA) are added to the glycerol structure by two acetyltransferases. Glycerol-3-phosphate acyltransferase (GPAT) converts G3P to lysophosphatidic acid (LPA) [56] and lysophosphatidic acid is acetylated by LPA acyltransferase. The latter reaction produces phosphatidic acid (PA), which is dephosphorylated to diacylglycerol (DAG) in a reaction performed by phosphatidic acid phosphatase (PAP) [57]. The last step comprises the acylation of TAGs, at the sn-3 position, either by an acyl-CoA-dependent or an acyl-CoA-independent reaction, to form TAGs that are stored in the form of lipid droplets [11,23].
3.2.2. Ex Novo Synthesis
In the ex novo pathway, hydrophobic substrates such as esters, TAGs, alkanes, etc., present in the culture medium are hydrolysed and transported to the intracellular space by active transport systems. There are two alternatives regarding the fate of the released fatty acids: they can be stored in lipid droplets, or they can be used for growth after the beta-oxidation of fatty acids. In both alternatives, the following step consists of the conversion of free fatty acids into acyl-CoA, a reaction catalysed by acyl-CoA synthetase [19]. Acyl-CoA can be esterified with glycerol, producing reserve and structural lipids [22]. Microbial lipids produced via the ex-novo pathway contain lower amounts of TAGs and higher amounts of free fatty acids compared to lipids produced via the de novo process [22].
4. Production of Yeast Oils from Lignocellulosic Biomass Hydrolysates: Inherent Challenges
The results of the extensive screenings of several yeast species/strains for the utilisation of different types of lignocellulosic biomasses to produce lipids are summarised in Table 1. Corn-derived biomasses show promising results with the highest lipid content described for a corn cob hydrolysate, reaching values of about 73% [58] and the highest lipid concentration values for the bioconversion of corn straw (23.3 g/L in a bioreactor, produced by R. toruloides DSMZ 4444). The highest lipid concentration (39.6 g/L) was obtained from Jerusalem artichoke extract hydrolysate using R. toruloides Y4 [59]. These results reinforce the idea of the potential of R. toruloides for lipid production from different feedstocks.
The negative impact of growth inhibitors that arise from the pretreatment of lignocellulosic substrates or the challenges registered in the catabolism of several C-sources by several oleaginous yeast species are discussed below (Section 4.1, Section 4.2 and Section 4.3).
4.1. Inhibition of Yeast Growth and Metabolism by Toxic Compounds Generated during Pretreatment
Lignocellulosic biomass is the largest renewable resource in the world. It is composed of complex carbohydrate polymers consisting of cellulose, hemicellulose, lignin and, depending on the biomass, a more or less residual part that includes pectin, proteins, extracts and ash [78,79]. Due to the recalcitrant nature of this biomass to deconstruction, a combination of enzymatic and thermochemical pretreatment processes is required to release the sugar components that can be converted into different value-added bioproducts by yeasts or other microorganisms [11,20]. Lignocellulosic biomass pre-treatments depend on the nature, chemical composition and structure of the biomass (hardwood, softwood or herbaceous) [80]. The nature and concentration of the by-products generated depend on the pre-treatment but may include furans and aldehydes, 2-furaldehyde (furfural) and 5-(hydroxymethyl)furfural (HMF), aromatic compounds (vanillin, syringaldehyde and 4-hydroxybenzoic acid) and weak acids (acetic, formic and levulinic acids) [80,81]. Since hemicellulose and lignin are acetylated [20,82], acetic acid is frequently present in lignocellulosic biomass hydrolysates at concentrations that can reach toxic values [83,84,85,86] being considered one of the major inhibitory compounds in lignocellulosic biomass hydrolysates. However, several strategies were developed to minimise the toxic effect of acetic acid, either by implementing a pretreatment that tackles the deacetylation and mechanical refining (DMR) or by the conversion of acetate directly into lipids [87] or co-products [88]. For its importance, the role of acetic acid in this context is detailed in Section 4.3. The concentrations of the main compounds present in lignocellulosic hydrolysates obtained after different biomasses pre-treatments were compiled [20,89]. Depending on the concentrations attained, they can seriously compromise yeast growth and bioconversion performance [80].
The furan-derived compounds, 2-furaldehyde (furfural) and 5-(hydroxymethyl)furfural (HMF) are formed during biomass pretreatment by the dehydration of pentoses and hexoses, respectively, and affect the activity of key enzymes of cellular metabolisms, such as glycolytic enzymes [90]. Furthermore, due to the action of the reactive aldehyde groups of furfural and HMF [34,37,91], reactive oxygen species (ROS) accumulate and may oxidise proteins, lipids and nucleic acids, affecting the corresponding cellular structures and leading to increased mutagenesis, protein denaturation, and biomembrane damage [90,92]. For this reason, the intrinsic tolerance of oleaginous yeasts to furan-derived compounds is also considered critical.
For detoxification of furfural and HMF, yeasts use reductases and dehydrogenases, that reduce or oxidise them to less toxic alcohols (furfuryl alcohol and 2,5-bis-hydroxymethylfuran) or acids (furoic acid and 2,5-furan-dicarboxylic acid) [35,93,94]. Furoic acid showed a lower toxic effect in Trichosporon fermentans when compared with furfural or furfuryl alcohol, inhibiting sugar utilisation rate less markedly [35]. However, it is important to note that tolerance to these furan compounds, as to any other toxicant, is strain-dependent [91,94]. Most Rhodotorula species are able to tolerate furfural concentrations up to 0.5 g/L. Two notable examples are the R. graminis strain UCDFST 04-862, which tolerates more than 0.5 g/L of HMF [91] or the R. pacifica strain INDKK, which is able to survive to 0.5 g/L of HMF and 2 g/L of furfural [95]. The tolerance of the R. graminis strain to HMF was increased using adaptive laboratory evolution experiments (ALE) by incubation in a corn maceration liquor medium supplemented with HMF (0.4%) for 7 days [96]. A Pichia kudriavzevii strain, isolated from soil, was found to be able to tolerate exceptional levels of HMF, up to 7 g/L [97]. There is a wide range of robust non-conventional yeasts with a natural tolerance to furfural and HMF while maintaining the ability to accumulate lipids as shown in Table 2. Trichosporon cutaneum 2.1374 is a good example as it is able to grow and produce lipids in media containing up to 1 g/L furfural or 2 g/L HMF more efficiently when compared to other species under the same conditions [34]. Although there are no in-depth studies available on the tolerance mechanisms active in oleaginous yeast species to these furan derivatives [98], the knowledge obtained in model yeasts is useful to guide strategies for increasing their tolerance to these and other stresses associated to related bioprocesses [99,100]. The development of more tolerant strains is discussed in Section 7.
4.2. Limitations to the Efficient and Complete Use of All the C-Sources Present, in Particular Xylose
The complete and efficient use of the sugars and other potential C-sources present in lignocellulosic biomass hydrolysates or in hydrolysates from any other feedstocks is essential to make their conversion economically viable. In lignocellulosic biomass hydrolysates, the main challenge is the bioconversion of xylose, which, in general, is the second most abundant sugar [101,102]. In the case of pectin-rich biomasses and residues, the acid sugar galacturonic acid is another highly challenging C-source for catabolisation by yeasts but can be efficiently catabolised by some oleaginous species [39,40]. Native xylose metabolism is not common in the Saccharomycotina but fairly common throughout the non-conventional yeast species [103]. Among other factors, carbon catabolite repression (CCR) represses xylose utilisation if glucose is present, in particular, the transport of sugars into the cell. This species-specific regulation leads to the sequential, rather than simultaneous, use of these C-sources as a result of preferential use of glucose, or another repressing carbon source, over others also present [104,105,106]. For this reason, CCR negatively affects the performance of biotechnological processes, since it prolongs the production time and, consequently, increases the inherent costs. This means that CCR is a very important regulatory mechanism when the use of mixtures of different C-sources is envisaged, as it is the case of hydrolysates from biomass or organic by-products or residues/wastes. The systematic study of lipid accumulation and production kinetics in a variety of oleaginous ascomycetous and basidiomycetous yeast strains grown on glucose and xylose, followed by the use of the selected strains for the bioconversion of wheat straw hydrolysate, pointed out as promising strains of the species L. starkeyi, R. glutinis, Rhodotorula babjevae and R. toruloides [107].
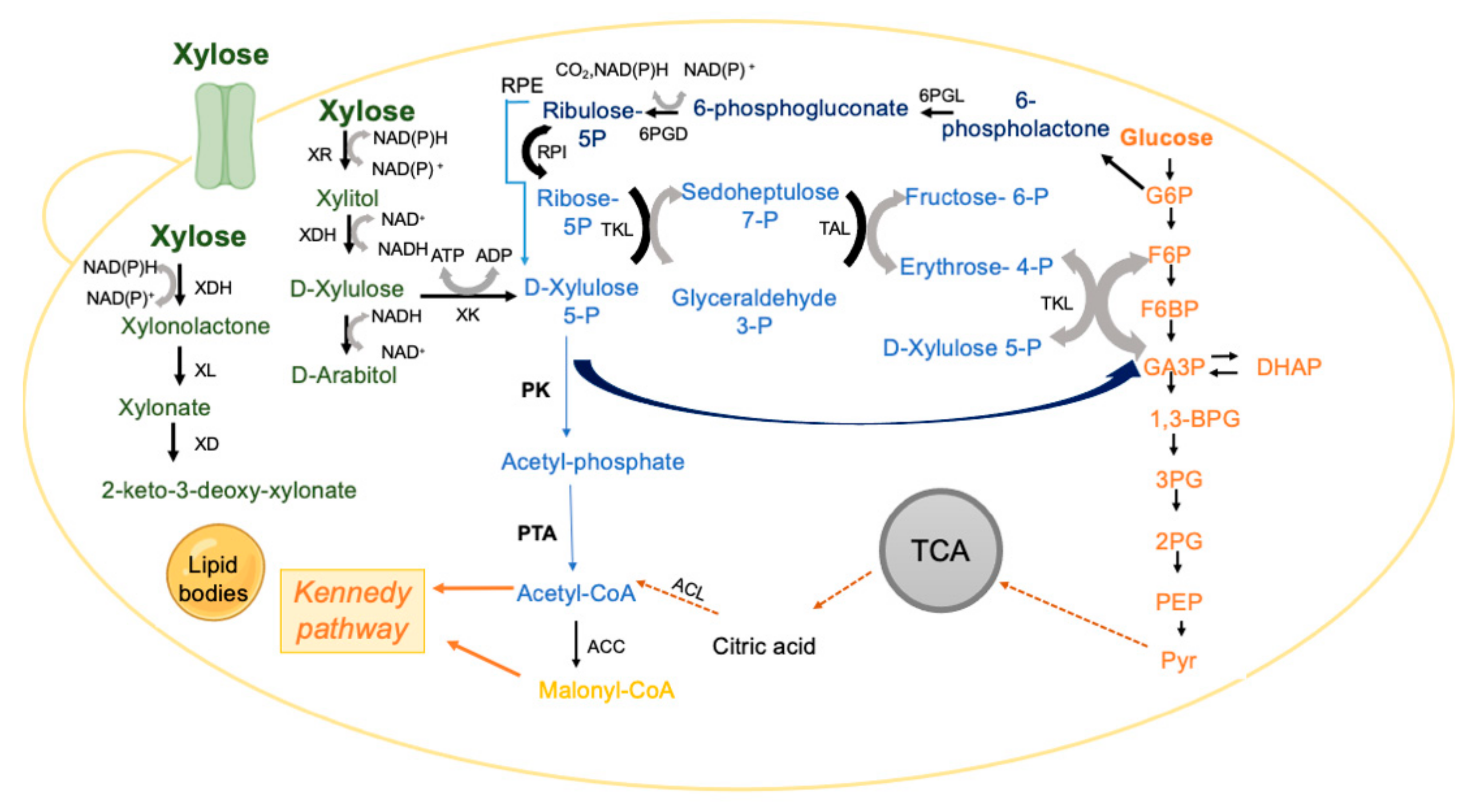
One of the limiting steps during xylose conversion, justified by CCR, is the xylose transport into the cell since xylose transporters are less efficient than those responsible for glucose transport, mainly due to low selectivity and/or affinity towards xylose [108,109]. Therefore, the identification of xylose transporters in L. starkeyi, R. toruloides and Y. lipolytica using molecular, bio-informatic, enzymatic, and transcriptomic analyses constitutes a starting point for the development of engineered strains for lipid production from xylose-rich substrates, [108,110,111,112,113]. The uptake of xylose into the yeast cell is followed by the activity of the xylose oxidoreductive pathway [102,114,115] (Figure 3). Briefly, xylose is reduced to xylitol, a reaction catalyzed by xylose reductase (XR) that uses NADH or NADPH as a cofactor [103,116]. The enzyme xylitol dehydrogenase (XDH) converts xylitol to D-xylulose by reducing NAD+ to NADH [117]. These two steps cause a redox imbalance that can be another limiting step in many yeasts. D-xylulose is then phosphorylated to xylulose-5-phosphate (X5P) by xylulose kinase (XK) [118]. The latter metabolite enters the phosphoketolase (PK) pathway, or the non-oxidative pentose phosphate pathway [102,103].
R. toruloides and L. starkeyi species can actively assimilate xylose [119,120]. R. toruloides tends to accumulate arabitol [114,121], produced through D-xylulose, in a reaction catalysed by the enzyme alcohol dehydrogenase (ADH) with consequent NAD+ production, which could be coupled to the reaction catalysed by XDH (Figure 3). Thus, these two reactions can contribute to the redox balance during xylose assimilation, with arabitol accumulation increasing under unbalanced conditions [121]. In addition, some Rhodotorula species, such as R. graminis, R. glutinis or R. toruloides metabolise X5P that can be transformed into glyceraldehyde-3-phosphate and acetyl-phosphate via the phosphoketolase (PK) pathway [122,123]. The PK pathway is more efficient if carbon economy is considered, as acetyl-phosphate can bypass pyruvate decarboxylation [102]. L. starkeyi was also shown to produce arabitol (about 1 g/L) when grown on corn stover hydrolysate, suggesting that it might also possess the arabitol production pathway [124].
Due to the limited genetic tools for both Rhodosporidium sp./Rhodotorula sp. and Lipomyces sp., genetic and metabolic engineering studies are scarce and have not yet allowed the detailed clarification of their xylose assimilation pathway. Y. lipolytica is known to possess in its genome genes encoding xylose reductase, xylitol dehydrogenase and xylulose kinase, but they are not sufficiently expressed to allow the efficient utilisation of xylose [125]. Thus, several genetic engineering strategies were explored to improve xylose assimilation in this species (detailed in Section 7.3).
4.3. The Dual Role of Acetic Acid as a Metabolism Inhibitor and C-Source
Acetate concentrations as high as 15 g/L can be found in lignocellulosic hydrolysates [126] and, depending on medium pH, significantly inhibit yeast growth and metabolism compromising sugar consumption rate and lipid yield [19,85,86]. At a pH below the pKa of this weak acid, (4.75 at 25 °C), acetic acid is essentially in the toxic undissociated form and enters the cell through the lipid bilayer of the plasma membrane by simple diffusion. In the cytosol, at a pH close to neutrality, acetic acid dissociates and the release of a proton (H+) leads to a decrease in intracellular pH and acetate accumulation [85,86]. Increased oxidative stress and turgor and inhibition of yeast growth and metabolism are among the detrimental effects of acetic acid toxicity [85,86]. Studies dedicated to the mechanisms of adaptation and tolerance to acetic acid in yeasts, in particular at the genome-scale in S. cerevisiae, are available in the literature and in several review papers [83,84,85,86,127,128,129]. Changes that occur in the molecular composition, structure and physical properties of the plasma membrane and cell wall are among the adaptive responses to this weak acid [130,131,132,133]. Because of such modifications, the permeability of the cell envelope in adapted cells is reduced, and so is the rate of passive diffusion of the acid form into the cell. This response, coordinated with the action of plasma membrane efflux pumps, such as Aqr1, Tpo2 and Tpo3, is reported to catalyse the active expulsion of intracellular acetate out of the cell, leading to the decrease in the internal concentration of the acid, and, consequently, its toxicity [134,135]. Regarding cell wall remodelling in response to acetic acid stress, a recent study reports that an adaptive response towards a more rigid and robust cell wall is also critical for acetic acid tolerance [130]. This response limits the futile cycle associated with the re-entry of the toxic acid form after the active expulsion of acetate from the cell interior [130]. The crosstalk between the ergosterol content of yeast plasma membrane and cell wall biophysical properties, involving the plasma membrane ABC transporter Pdr18, described as a determinant of acetic acid tolerance due to its involvement in ergosterol transport at the plasma membrane level, was also demonstrated [131]. Considering the high importance that acetic acid toxicity has in the performance of yeasts, in particular oleaginous yeasts, the exploration of yeast biodiversity and a better understanding of the molecular targets and pathways behind the increase in yeast efficiency and robustness under stress imposed by acetic acid is essential to the productivity and economic sustainability of lignocellulosic biorefineries [136].
Although toxic, acetic acid can also be an interesting C-source for oleaginous yeasts. However, it is important to use a cultivation medium pH leading to lower acetic acid toxicity and acetic acid concentrations below the threshold for each yeast strain tolerance [39,137,138]. Acetate can be converted into acetyl-CoA, which is a precursor for lipid biosynthesis, so most yeasts capable of assimilating acetic acid are oleaginous. In oleaginous fungi, a considerable percentage of acetate is directed to lipid biosynthesis in the presence of glucose and xylose, as in the case of lignocellulosic biomass hydrolysates [139]. Furthermore, acetate can be directly converted to acetyl-CoA in the cytosol by acetyl-CoA synthetase (ACS) and immediately used for fatty acid biosynthesis without the involvement of complex and energy-consuming metabolic and mitochondrial transport processes [139].
In the presence of glucose, acetic acid assimilation can be repressed in yeasts such as S. cerevisiae, Candida utilis, Torulaspora delbruecki and Dekkera anomala [98]. However, in other yeasts, acetate can be simultaneously catabolised, as in the case of Zygosaccharomyces bailii [140] and R. toruloides [39,141]. Several studies indicate that there is an increase in lipid production when the co-consumption of sugars and acetic acid occurs [19,126,142]. When acetic acid is co-consumed with xylose, sugar assimilation can be facilitated as well as lipid accumulation [19,126,142]. For example, the presence of acetate and its co-metabolism with glucose-enhanced lipid content to levels close to 70% in the presence of 7.2 g/L acetate, indicates that the excess acetate is used as building blocks in lipid biosynthesis by R. toruloides [143]. A Cryptococcus curvatus strain was able to simultaneously consume mixtures of (i) acetate and glucose, (ii) acetate and xylose, and (iii) acetate in rich corn hydrolysates, and produce lipids. Furthermore, the partial replacement of glucose by acetic acid in the same amount resulted in higher lipid concentration (6.8 g/L in medium with 30g/L glucose and 10 g/L acetic acid compared with 6.0 g/L lipid concentration, obtained in the medium with 40 g/L glucose) [142]. In the case of Trichosporon cutaneum 2.1374, this strain was able to slowly metabolise acetic acid simultaneously with glucose or xylose, and lipid productivity was also higher in acetic acid supplemented medium. Acetic acid can be used either as the sole C-source or as a C-source in the second stage of two-stage fermentation (Table 3). In the latter case, the increase in acetate concentration leads to a higher C/N ratio, suitable for lipid production (see Section 6.2) [143]. For example, with 20 g/L of acetic acid, R. toruloides AS 2.1389 cells accumulated 48.2% in lipids, whereas with 4 g/L of this acid, the lipid content was reduced to approximately one-third of that value [141]. Acetic acid can also exert a beneficial effect on lipid production when it is present in a mixture of volatile fatty acids (VFAs), a topic discussed in Section 5.2.
5. Production of Yeast Oils from Organic Industrial Byproducts/Wastes/Residues
5.1. From Crude Glycerol, a Biodiesel Production Byproduct
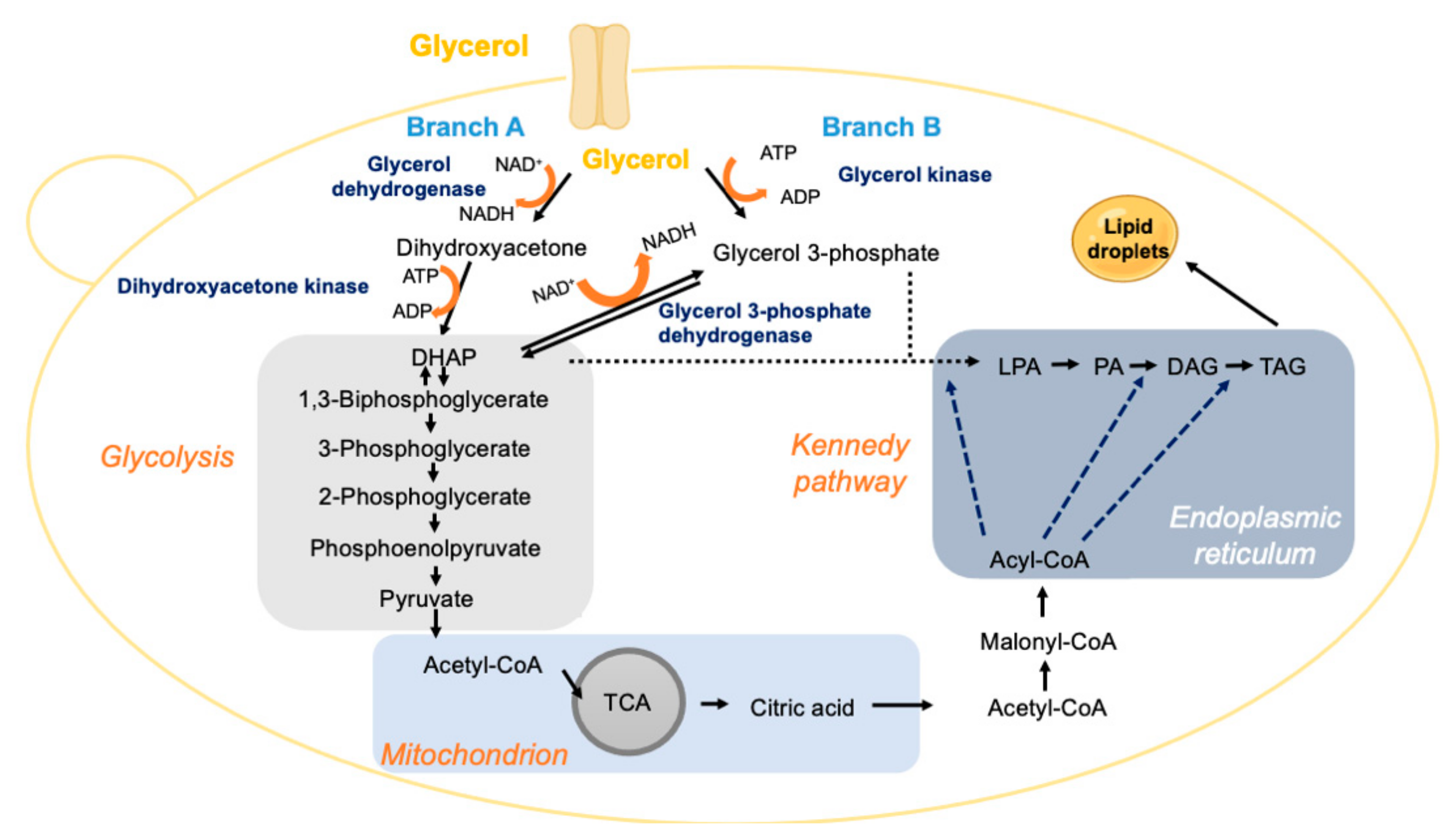
Crude glycerol is a byproduct of biodiesel manufacturing that can be used as feedstock for the production of yeast oils (Table 4). Crude glycerol produced in the biodiesel industry is composed of 70–80% glycerol that can be used as a C-source for lipid production by suitable oleaginous yeasts (Figure 4). However, although crude glycerol composition varies depending on the industrial process, it is contaminated with alcohols (mainly methanol), catalysts, dissolved salts, and water [146,147].
The use of glycerol as a carbon source by oleaginous species, as is the case for C. boidinii, C. curvatus, L. starkeyi, R. toruloides and Y. lipolytica [25,43,44], occurs through the phosphorylation and oxidative pathways. For S. cerevisiae and Y. lipolytica, the active transport mechanism primarily uses glycerol/H+ antiporters [44]. Regarding the oxidative pathway (Branch A, Figure 4), the first step consists of the oxidation of glycerol to dihydroxyacetone through FAD/NAD dehydrogenase. Dihydroxyacetone is phosphorylated by a dihydroxyacetone kinase, producing dihydroxyacetone phosphate [6]. The other alternative reaction involves the enzyme 3-phosphoglycerate dehydrogenase (NAD-dependent), which catalyses the conversion of glycerol-3-phosphate into dihydroxyacetone phosphate in the mitochondria [44]. Once in the cytosol, dihydroxyacetone enters the glycolytic pathway (Branch B, Figure 4) and follows the TCA and Kennedy pathways (Figure 4). Considering the phosphorylation pathway, glycerol is phosphorylated by glycerol kinase, generating glycerol-3-phosphate that can enter directly into the Kennedy pathway for lipid production.
Since most biodiesel manufacturers utilise high methanol-to-oil molar ratios, methanol is a major contaminant of crude glycerol residues and methanol toxicity affects the performance of oleaginous yeasts [45]. Although part of this alcohol can be removed by thermal treatment, the residual methanol concentration may be toxic to yeast cells and limit bioprocess productivity [161]. Lipid production by R. toruloides 32489 using crude glycerol supplemented with increasing methanol concentrations (from 2 to 20 g/L) was inversely proportional to methanol concentration: at 20 g/L, biomass, lipid content and lipid production dropped by 6.6%, 11.9% and 17.7%, respectively, compared with pure glycerol [155]. However, when mixed with other impurities present in crude glycerol substrates such as esters (e.g., methyl and sodium oleate), salts and soap, the negative effects of methanol can, apparently, be alleviated [155]. Moreover, methanol can be useful in avoiding bacterial contamination of non-sterilised crude glycerol used for lipid production [150,152].
Methanol can also be used as a C-source by yeasts but no study addressing methanol consumption and lipid production from crude glycerol could be found in the literature. Recent reports on the exploitation of different methodologies to optimise the utilisation of methanol as a C-source by methylotrophic and non-methylotrophic yeasts were published [161,162,163,164]. According to a metabolomics study, the methylotrophic yeast Ogataea methanolica responds to the presence/absence of methanol and also to its concentration [164]. The native capacity of S. cerevisiae for methylotrophy was examined as the first step towards the unraveling of methylotrophy in the model yeast [163]. Synthetic methylotrophy constitutes a challenging alternative that can be implemented in non-methylotrophic oleaginous hosts to increase the feasibility of bioprocesses that use crude glycerol as a substrate.
5.2. From Volatile Fatty Acids (VFAs), Intermediate Compounds from Anaerobic Digestion of Organic Wastes
Volatile fatty acids (VFAs) are intermediate compounds obtained from anaerobic digestion of organic wastes, for example, food wastes. VFAs are produced after the hydrolytic and acidogenic phases of anaerobic digestion. The most common VFAs are acetic (C2), propionic (C3), butyric (C4), isovaleric, valeric (C5) and caproic (C6) acids [165], and their ratio depends on the experimental conditions, substrate composition and the microorganisms present in the anaerobic digestion system [166]. VFAs are considered a promising alternative feedstock for lipid production by oleaginous yeasts in a circular bio-economy context [19,145,167,168]. The main studies available in the literature using a mixture of VFAs as carbon sources for lipid production are summarised in Table 5. When Y. lipolytica was grown on a mixture of VFAs, acetic acid was found to play a key role in the consumption of longer-chain VFAs (C5 and C6), increasing the availability of the C-sources suitable for lipid production [169]. Additionally, a higher proportion of acetic acid in the VFA mixture of acetic acid:propionic acid:butyric acid (in ratios of 8:1:1) led to higher lipid accumulation, lipid concentration and productivity when compared to lower proportions of acetic acid in the VFA mixture (e.g., acetic acid:propionic acid:butyric acid in ratios of 4:3:3 or 6:1:3) [170].
5.3. From Combinations of Residues/Wastes
Different combinations of crude glycerol and other carbon sources present in lignocellulosic biomasses or other organic industrial residues were also explored [43,151,157,175,176,177]. The addition of cellulosic hydrolysates to crude glycerol improved the lipid production rate of Rhodotorula species. The mixing of crude glycerol with 10% hydrolysate from wheat straw subjected to the acid-based steam explosion (composition: 2.6 g/L xylose, 0.6 g/L glucose and 0.8 g/L acetic acid) led to an enhanced lipid production rate and the reduction of the time for consumption of all the available carbon sources of R. toruloides and R. glutinis. The valorisation of crude glycerol and sunflower meal (SFM) resulting from biodiesel production plants was also examined using R. toruloides, L. starkeyi and C. curvatus. Among the aforementioned species, the lipid profile of R. toruloides oils was the closest to the palm oil used for biodiesel [157]. The strategy of fed-batch cultivation, using sugarcane top hydrolysate as a substrate in the first stage and crude glycerol in the second stage was found to have a strong influence on biomass and lipid production in Rhodosporidiobolus fluvialis DMKU–SP314 [175,177]. Genetically engineered microorganisms are also an alternative to enhance lipid production using low-cost residual substrates. A good example is the genetically manipulated Y. lipolytica JMY4086, which is able to successfully catabolise crude glycerol and molasses, producing lipids [176].
For feedstocks with a low C/N ratio, such as in wastewater sludge, the addition of crude glycerol as a promising C-source for lipid production can be considered an interesting strategy [178]. For example, the use of a combination of municipal sludge fortified with crude glycerol, allows Y. lipolytica SKY7 to produce higher concentrations of biomass and lipids when compared with unsupplemented crude glycerol, leading to the valorisation of these two byproducts/wastes [148].
A first study focusing on the combination of pumpkin peels with syrup from candied fruits processing was recently published, demonstrating that these wastes were sufficient to support yeast growth and enhance lipid accumulation in Rhodosporidiobolus azoricus and Cutaneotrichosporon oleaginosum [179].
6. Effect of Process Conditions in the Production of Yeast Oils
Several physiological and environmental factors affect the growth, lipid accumulation and lipid profile of oleaginous yeasts. These include yeast species/strain, growth phase, culture medium components (e.g., carbon (C)-source, nitrogen (N)-source, molar C/N ratio), and other macronutrients (e.g., phosphorus and sulphur) as well as micronutrients (trace metals in minimal media) and undefined micronutrients in complex media, and other cultivation conditions (e.g., inoculum size and physiological state, pH, temperature, dissolved oxygen (DO) level, type of bioreactor(s), cultivation time). The optimisation of these factors is essential to achieve high productivity and minimise production costs.
6.1. Nitrogen (N) Source
Lipid production is influenced by the nature and concentration of the carbon and nitrogen sources used by oleaginous yeasts. The influence of the C-source was discussed above (Section 4.2, Section 4.3, and Section 5.1, Section 5.2 and Section 5.3). Concerning the N-source, both organic (yeast extract, peptone or urea) and inorganic (ammonium chloride, ammonium sulphate and sodium nitrate, or a mixture of both) nitrogen were tested [180,181,182]. Since yeast extract is an expensive medium component, its replacement by other low-cost organic nitrogen sources (e.g., corn steep liquor, monosodium glutamate, soybean powder or urea) was explored [71]. The results revealed that when half of the yeast extract was replaced by urea, a significant decrease in lipid concentration occurred (from 6.6 g/L to 4.9 g/L) [71]. This could be the result of the alkalinisation of the culture medium by the ammonium ion resulting from urea hydrolysis, known to trigger L. starkeyi cell death [124]. However, the use of urea or even ammonia as major nitrogen sources is well documented for Y. lipolytica [183,184]. Inorganic sources, such as ammonia, are preferred in industrial processes due to the lower cost. However, if organic and inorganic nitrogen sources are compared, organic nitrogen sources are more favorable for lipid accumulation, as described for R. toruloides, with an oil content of 50% when grown on organic nitrogen compared to 18% when inorganic nitrogen was used [185]. This result may also be related to the likely presence of some nutrients such as amino acids and vitamins in organic sources that may enhance cell growth and lipid accumulation [186].
6.2. Carbon-to-Nitrogen (C/N) Ratio
Lipid production by oleaginous microorganisms requires a medium in which there is an excess of carbon and a limited amount of other nutrients such as phosphorus, sulphur or nitrogen [9]. The excess carbon is redirected to lipid synthesis, rather than to cell proliferation [187]. As previously described in Section 3.2.1, it is considered that nitrogen depletion triggers the activation of adenosine monophosphate deaminase and catalyses the conversion of AMP to inosine 5’-monophosphate and ammonium, initiating the TAG synthesis reaction [188]. Therefore, the carbon-to-nitrogen (C/N) ratio is critical during lipid biosynthesis. C/N ratios suitable for lipid production range from 50 to 150 [189]. However, it is essential to establish a suitable C/N ratio that favors lipid accumulation without compromising cell growth in the medium. A reported exception to the referred production profile is the case of Cryptococcus terricolus which accumulates lipids when there is still nitrogen in the culture medium [190].
Specific examples supporting the general conclusions stated above follow. When R. toruloides CCT 0783 was grown with four different C/N ratios (60, 80, 100 and 120) and three different C-sources (glycerol, acetic acid or xylose), the highest lipid yields in acetic acid and xylose were for a C/N ratio of 120 (0.6 g/g and 0.53 g/g, respectively) [191]. However, for glycerol, a C/N ratio as high as 120 caused a marked decrease in specific growth rate and lipid yield [191]. In a study using Trichosporon dermatis 32903 that compares the influence on lipid production of C/N ratios from 30 to 130, the ratio of 110 led to the highest lipid production (16.33 g/L) [75]. For R. taiwanensis AM2352, the highest amount of lipids accumulated was at a C/N ratio of 30 [69]. Collectively, these results emphasise the importance of choosing an appropriate C/N ratio according to the carbon source, the selected strain and other cultivation conditions.
In addition to the initial C/N ratio, the amount of nitrogen per se should also be considered [124]. In other words, increasing the amount of the initial carbon source may not always be sufficient to increase lipid production if cells only start accumulating lipids when nitrogen concentration is low enough [124]. As a strategy to achieve high lipid concentrations without compromising biomass production, two-stage batch processes were used in which cell proliferation occurs first, in a rich medium, and lipid accumulation occurs later, under nitrogen-limiting conditions [19,60]. Under such conditions, lipid production by L. starkeyi NRRL Y-1388 increased by 78% [192]. When L. tetrasporus Y-11562, L. kononenkoae Y-7042 and R. toruloides Y-1091 were used and a C/N 60 was present in the first phase and a C/N ~500 in the second phase, lipid productivity was three to seven times higher than was possible during the first growth phase [60]. Since the aim of the second phase is to produce lipids and not biomass that accumulates during the first phase, it is possible to use higher concentrations of C-sources that also act as growth inhibitors. This is a strategy used for yeast species with poor growth on acetic acid [145,193], or to enhance lipid yield in species capable of using acetic acid efficiently [141,170].
6.3. Dissolved Oxygen (DO) Concentration
Dissolved oxygen (DO) concentration affects both lipid accumulation and composition, although the results reported in the literature vary with the yeast species. Typically, oleaginous yeasts require oxygen for rapid growth; in bioreactors, agitation increases nutrient availability while maintaining uniformity of cell distribution in the medium [58]. However, in general, high aeration levels lead to a decrease in the lipid content but the optimum aeration level depends on the yeast strain [124,194,195,196]. For example, for DO levels of 25% and 60%, the higher lipid accumulation by R. glutinis was at the lower DO level while higher DO levels favor biomass production [196]. However, not all yeast species are equally affected by the DO concentration concerning lipid accumulation [197]. For example, Rhododosporium azoricum’s production of lipids was found to be more prone to low DO concentrations than Trichosporon oleaginous [197]. Finally, it is important to note that fatty acid desaturases use oxygen as a substrate to catalyse the unsaturation reaction [124], but there is no unequivocal association between higher saturation and dissolved oxygen levels [196,198].
6.4. Temperature and pH
The cultivation temperature also influences the composition of yeast oils and their degree of saturation and the optimum temperatures for which biomass production is favoured, may not be optimal for lipid accumulation [71,194,199]. Additionally, the activity of yeast desaturases is temperature-dependent, these enzymes being more stable at low temperatures, namely the Δ12-desaturase [188,200], thus, the saturation degree of yeast oils is also temperature-dependent [138,201].
The optimum pH for lipid production should be selected for specific substrates and strains [11,22,187]. When glucose is the main carbon source, acidic conditions, mainly in the range of pH 5 to 6, are employed in lipid production [137]. Remarkably, L. starkeyi is capable of growing and producing lipids in media with a very low pH, around 3.0, likely due to their intracellular buffering capacity [63,184]. In the case of R. mucilaginosa and R. toruloides, the presence of acetic acid in the hydrolysates (30–40 mM) adjusted to pH 5.0 did not compromise the rapid and full utilisation of D-glucose, D-galactose and acetic acid [39]. Nevertheless, at pH 3.5, yeast growth was fully abrogated [39].
Regarding the use of VFAs as a carbon source, there are two different perspectives. Some authors consider that slightly acidic conditions (pH 5.6–7) are beneficial compared to alkaline conditions, mainly for low concentrations of VFAs. Other authors report that alkaline pHs can be advantageous since they alleviate the toxic effect of the high content of the weak acids present in VFA mixtures, enhancing lipid production [137]. In the case of cultivation media with high content of VFAs, an initial pH of 8 was found to be the optimal pH condition for lipid production by Y. lipolytica [137].
6.5. Effect of the Inoculum
The size and physiological state of the inoculum are critical to the performance of stress-associated bioprocess, as is the case for most of the bioconversions of lignocellulosic biomass and other organic residues/wastes by yeasts. Additionally, the inoculum size influences biomass production, lipid titer and lipid content [202,203,204,205]. It is expected that an increase in the inoculum size (frequently associated with the initial culture OD600nm) may lead to the increase in the concentration of viable producing cells capable of initiating growth under stress conditions, in particular under the toxic effect of chemicals present in lignocellulosic biomass hydrolysates or in any other organic residues [65]. A higher active cell fraction elevates the probability of the cell population resuming growth after sudden exposure to a stressful environment and exhibiting an increased C-source consumption rate [202]. For example, the negative impact on R. toruloides performance of inhibitory concentrations of acetic acid present in sugar beet pulp (SBP) hydrolysates was negligible when higher concentrations of inoculum were used [39]. A similar increase in process performance concerning the consumption rate of a mixture of xylose and glucose was reported for L. starkeyi when the inoculum size was increased [202]. An inoculum ratio of 10% (v/v) was considered ideal for maximum biomass and lipid production, and lipid content by Phenoliferia glacialis (syn, Rhodotorula glacialis) DBVPG4875 [203] and Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 [204] but other values were found depending on the specific bioprocess conditions [119,205].
7. Strategies to Develop Superior Strains for the Production of Oils from Residual Feedstocks
7.1. Exploring Available Bioinformatics Tools
For guiding the development of superior yeasts by genetic and genome engineering, in particular for non-conventional yeasts, several web database resources provide a wealth of functional and transcription regulation information for the analysis of gene expression datasets. This is the case of the Saccharomyces Genome Database (SGD) (https://www.yeastgenome.org/, accessed on 7 June 2022), the major community resource for gene, genomic and protein information in yeast and the YeastIP database that compiles nucleotide sequences of the most common markers used for yeast taxonomy and phylogeny, allowing identification, taxonomy and phylogeny of yeasts species [206]. The YEASTRACT+ database and information system, a tool for the analysis of transcription regulatory associations in Saccharomyces cerevisiae, currently includes the N.C.Yeastract database (Non-Conventional Yeastract; http://yeastract-plus.org/ncyeastract/, accessed on 7 June 2022) [207,208]. N.C.Yeastract allows the (i) inference of orthologous genes, (ii) search for putative TF binding sites, and (iii) inter-species comparison of transcription regulatory networks and prediction of TF-regulated networks based on documented regulatory associations available in YEASTRACT + for well-studied species, especially S. cerevisiae [208]. For example, the prediction of the Haa1 regulon in R. toruloides (RtHaa1) in response to acetic acid stress was possible using YEASTRACT+ [209]. The outcome of such analysis can have an impact on the optimisation of R. toruloides robustness for the bioconversion of lignocellulosic and pectin-rich residue hydrolysates [209] given that the transcription regulator Haa1 is a major determinant of acetic and formic acids tolerance in yeasts [85].
Other in silico approaches are available to facilitate the development of superior yeasts. Genome-scale metabolic models available for several oleaginous yeasts such as L. starkeyi [210,211], R. toruloides [212], T. oleaginosus (Cutaneotrichosporon oleaginosus) [213], and Y. lipolytica [214], constitute useful tools to guide the manipulate of yeast metabolism [215,216]. Another advantage of in silico approaches is the identification of targets that may be relevant for increasing stress tolerance. For example, the entire transportome of Starmerella bombicola was unveiled using a bioinformatics tool that identifies putative transporters and the obtained results highlighted the role of the ABC transporters superfamily in the export of sophorolipids [217].
7.2. Genetic Engineering to Improve Lipid Biosynthesis
Enzymes involved in the lipid biosynthetic pathway are likely key molecular targets for the development of strategies to improve yeast oil accumulation. One of the most widely used approaches relies on the increase in the expression of genes encoding enzymes that directly influence oil accumulation (Table 6). Two of the most explored molecular targets are the diacylglycerol O-acyl-transferases (DGA) encoding genes. The enzyme Dga1 was characterised in different yeast species and its overexpression promotes lipid accumulation [218,219,220,221,222,223]. In Y. lipolytica, two different coding genes, DGA1 and DGA2, were identified, but YlDGA1 outperforms YlDGA2 [224]. Another approach involves the redirection of the central carbon metabolism to increase the availability of precursors by the over-expressing malic enzyme (ME) and acetyl-CoA carboxylase (ACC) that supply the pathway with the essential molecules, acetyl-CoA, NADPH, and malonyl-CoA, respectively (Table 6). Since the malic enzyme of R. toruloides is the main enzyme providing NADPH during synthesis, its expression is essential for lipid accumulation [225], whereas its overexpression in Y. lipolytica or L. starkeyi does not alter lipid accumulation [226,227]. In Y. lipolytica, the main source of NADPH is the pentose phosphate pathway [228]. To surpass this limitation, four biosynthetic pathways were designed to convert NADH into NADPH in Y. lipolytica. The best result was obtained for a Y. lipolytica strain co-overexpressing the glyceraldehyde-3-phosphate dehydrogenase GapC (catalyses the conversion of glyceraldehyde 3-phosphate to 1,3-diphosphoglycerate with the reduction of NAD to NADH) and malate dehydrogenase, the enzyme encoded by the Mucor circinelloides MCE2 gene; this enzyme is responsible for the decarboxylation of malate with reduction of NADP+ to NADPH (Table 6) [229]. The overexpression in Y. lipolytica and R. toruloides of acetyl-CoA carboxylase, encoded by ACC1 which catalyses the carboxylation of acetyl-CoA to malonyl-CoA, led to an increased lipid content [221,230]. On the other hand, the homologous or heterologous overexpression of ATP-citrate lyase (ACL) genes, responsible for the supply of acetyl-CoA from the cleavage of citrate, did not lead to an increase in lipid content [231]. Other genetic manipulations were also explored, including the manipulation of fatty acid synthesis to obtain lipids with longer chains or targeting the expression of acyl-CoA/acyl-ACP processing enzymes in other cellular compartments, such as the cytoplasm, the peroxisome or the endoplasmic reticulum, in order to minimise the effects of compartmentalisation on the accessibility to the downstream engineered biocatalysts [232].
The deletion of genes involved in lipid degradation, such as the acyl-CoA oxidases (POX) or peroxisomal biogenesis (PEX) genes, was also examined. The deletion of one of these genes, PEX10, to abolish peroxisome biogenesis and therefore lipid catabolism, enhanced lipid accumulation in Y. lipolytica [233], while in R. toruloides lipid accumulation was reduced [225]. The elimination of PEX10 also led to slower growth, corroborating previous studies that found that peroxisome biosynthesis is required for robust cell growth in basidiomycetes [234,235,236]. The deletion of genes related to by-product formation was also found to be beneficial to lipid accumulation. For example, this is the case of the elimination of genes of glycogen synthesis, indicating that this pathway competes with oil accumulation [237].
Transcription factor engineering also emerged as a promising strategy to increase yeast tolerance to different biotechnological relevant stresses to be used with the goal of enhancing lipid production. To assure a C/N ratio suitable for lipid production, high concentrations of glucose are usually present and consequently, the expression of genes required for the metabolism of alternative carbohydrates, gluconeogenesis and mitochondrial functions are repressed [238,239]. The deletion of the transcription factor MIG1 [239] or the mutation of SNF1, encoding a serine/threonine-protein kinase that regulates MIG1 [240], both involved in glucose repression, was found to increase lipid titers in Y. lipolytica. Furthermore, the deletion of MGA2, a regulator of the expression of desaturases, enhanced lipogenesis and the biosynthesis of unsaturated fatty acids [241]. The importance of fatty acid desaturases as potential targets for increasing lipid accumulation was widely examined. The expression of these membrane-bound proteins that catalyse the addition of a double bond in fatty acid hydrocarbon chains to produce unsaturated and polyunsaturated fatty acids, was found to enhance lipid production in several yeast species/strains as well as the accumulation of unsaturated lipids, which is beneficial for biodiesel production [223,225,242,243,244].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 6.
Genetic modifications performed in different oleaginous yeasts to increase lipid production. A brief description of the experimental conditions and genetic modification, as well as lipid production, is also presented. The species covered in the table are Yarrowia lipolytica, Rhodotorula toruloides, Rhodotorula fluvialis and Candida phangngensis. The underlined values correspond to the lipid production parameters obtained for the parental strain.
Table 6.
Genetic modifications performed in different oleaginous yeasts to increase lipid production. A brief description of the experimental conditions and genetic modification, as well as lipid production, is also presented. The species covered in the table are Yarrowia lipolytica, Rhodotorula toruloides, Rhodotorula fluvialis and Candida phangngensis. The underlined values correspond to the lipid production parameters obtained for the parental strain.
| Species and Strain | Experimental Condition | Genetic Modification | Lipid Content (%) Lipid Titer (g/L) Lipid Productivity (g/Lh) | Ref. |
|---|---|---|---|---|
| Y. lipolytica JMY3501, derived from Po1d | Bioreactor, 150 g/L Suc, 1.7 g/L YNB, 3.75 g/L NH4Cl, 0.7 g/L KH2PO4, and 1.0 g/L MgSO4.7H2O. C/N 60. | pox1- 6Δ and tgl4Δ. o/e of GDP1 and DGA2 | - 5.76 - | [245] |
| Y. lipolytica Y4086, derived from Po1d | pox1- 6Δ and tgl4Δ. o/e of GDP1, DGA2, and HXK1. h.e. of ScSUC2 | - 9.15 - | ||
| Y. lipolytica, derived from Po1f | Test-tube, 80 g/L Glu, 6.7 g/L YNB(−/−), 1.365 g/L NH4, 0.79 g/L CSM supplement. | pex10Δ and mfe1Δ. o/e of DGA1 | 74/16.8 6.0/0.41 - | [224] |
| Y. lipolytica, derived from Po1f | Bioreactor (batch), 160 g/L Glu, 13.4 g/L YNB−/−, 2.73 g/L NH4. | 70.6/- - - | ||
| Y. lipolytica MTYL065, derived from Po1g | Bioreactor, 90 g/L glucose, 1.5 g/L YNB−/−, 2 g/L (NH₄)₂SO₄, 1 g/L YE. | o/e of ACC1 and DGA1 | 61.7/11.7 - 0.143/- | [230] |
| Y. lipolytica MTYL065, derived from Po1g | Flask, 50 g/L Glu, 1.7 g/L YNB−/−, 1.5 g/L YE, C/N 20. | o/e of ACC1 and DGA1 | 41.4/8.77 - - | [230] |
| Y. lipolytica YL-ad9, derived from Po1g | Bioreactor, 150 g/L Glu, 3.4 g/L YNB−/−, 8.8 g/L (NH₄)₂SO₄, 2 g/L YE. | Replacement of the hp4d promoter with the TEFin promoter to co-overexpress ACC1 and DGA1 o/e of stearoyl-CoA desaturase (SCD) | 67/- 55/- 0.707/- | [223] |
| Y. lipolytica Ylgsy1Δ, derived from H222 | Bioreactor, 20 g/L Glu, 0.4 g/L (NH₄)₂SO₄. | gsy1Δ | >60% TAGs synthesis increase | [237] |
| Bioreactor, 20 g/L Gly, 0.4 g/L (NH₄)₂SO₄. | ||||
| Y. lipolytica snf1Δ, derived from ATCC 20362 | Flask, Growth phase: SD medium (2% Glu and 0.5% (NH₄)₂SO₄). | snf1Δ Expression of Δ- 9 elongase/Δ-8 desaturase pathway | 18.5/7.1 - - | [240] |
| Y. lipolytica snf1Δ, derived from ATCC 20362 | Flask, Oleaginous phase: high concentration of Glu (8% Glu without N source). | 18.5/12.6 - - | ||
| Y. lipolytica M25, derived from ACA-DC 50109 | Flask, 5.0% Glu, 0.7% KH2PO4, 0.25% Na2HPO4, 0.15% MgSO4·7H2O, 0.015% CaCl2, 0.015% FeCl3·6H2O, 0.002% ZnSO4·7H2O, 0.006% MnSO4·H2O, 0.05% YE. | mig1Δ | 48.7/36.0 - - | [239] |
| Y. lipolytica L36 DGA1, derived from PO1f | Bioreactor (fed-batch), 80 g/L Glu, 3.4 g/L YNB−/−, 4 g/L (NH₄)₂SO₄. | Mutation in the gene MGA2 (mga2-G643R). o/e of DGA1 | - 25/- 0.145/- | [241] |
| Y. lipolytica NS432, derived from YB-392 | Bioreactor (batch), 150 g/L Glu, 0.1 g/L corn peptone, 3 g/L YE. | o/e of DGA1 from R. toruloides h.e. of DGA2 from Claviceps purpurea tgl3Δ | 77/24 30.8/3.6 0.27/0.03 | [222] |
| Y. lipolytica NS432, derived from YB-392 | Bioreactor (fed-batch), 150 g/L Glu, 0.1 g/L corn peptone, 3 g/L YE. | 73/25 84.5/12.8 0.73/0.11 | ||
| Y. lipolytica YL10, derived from PO1f | Bioreactor, 40 g/L Glu, 1.7 g/L YNB−/−, 3.52 g/L (NH₄)₂SO₄, 2 g/L uracil. | h.e. of Δ-15 desaturase from flax o/e of ACC1, DGA1, SCD, Δ-12D pex10Δ and mfe1Δ | 77.8/- 50.0/- - | [244] |
| Y. lipolytica VHb, derived from Polh | Bioreactor, 90 g/L Glu, 1.5 g/L YNB−/−, 2 g/L (NH₄)₂SO₄, 1 g/L YE. | h.e. of Vitreoscilla haemoglobin (VHb) | 14.5/10.5 - - | [246] |
| Y. lipolytica YL-1292sp-ACL-6, derived from Polh | Flask., Modified K&R medium with 100 g/L Gly, 2 g/L C4H12N2O6. | h.e. of ACL from Mus musculus | 23.1/7.3 - - | [247] |
| Y. lipolytica AD-perCAT2, derived from Polg | Bioreactor, 100 g/L Glu, 3.4 g/L YNB−/−, 2.2 g/L (NH₄)₂SO₄. | h.e. of perCAT2 from Saccharomyces cerevisiae o/e ACC1 and DGA1 | - 66.4/- 0.565/- | [232] |
| Y. lipolytica Adgapc, derived from Polg | Bioreactor, 100 g/L Glu, 3.4 g/L YNB, 2.5 g/L YE, 8.8 g/L (NH₄)₂SO₄ | h.e. of GapC from Clostridium acetobutylicum | 62.5/54.7 63.3/47.8 - | [229] |
| Y. lipolytica Adme, derived from Polg | h.e. of MCE2 from Mucor circinelloides | 63.7/54.7 61.4/47.8 - | ||
| Y. lipolytica Adpp, derived from Polg | Co-expression of a phosphoketolase from Leuconostoc mesenteroides and a phosphate acetyltransferase from Clostridium kluyveri | 52.7/54.7 56.2/47.8 - | ||
| Y. lipolytica Adgy, derived from Polg | Co-expression of the heterologou Clostridium acetobutylicum GapC and Y. lipolytica YEF | 63.2/54.7 54.6/47.8 - | ||
| Y. lipolytica Adgm, derived from Polg | Co-expression of the heterologous Clostridium acetobutylicum GapC and the heterologous Mucor circinelloides MCE2 | 75.5/54.7 66.8/47.8 - | ||
| Y. lipolytica ALDH, derived from Polg | Bioreactor, 100 g/L Glu, 3.4 g/L YNB−/−, 4.4 g/L (NH₄)₂SO₄. | o/e of glutathione peroxidase (GPO), glutathione disulfide reductase (GSR), h.e. of glucose-6- phosphate dehydrogenase (S.cerevisiae Zwf1) and aldehyde dehydrogenase (Escherichia coli AldH) | 81.4/40.6 72.7/- 0.97/- | [248] |
| Y. lipolytica JMY5035, derived from Pold | Flask., 6% Soluble starch, 0.17% (w/v) YNB, 0.15% (w/v) NH4Cl, pH 6, C/N 60. | o/e of DGA2, GPD1 and h.e. of alpha-amylase from Oryza sativa + glucoamylase from Aspergillus niger; pox1-6Δ and tgl4Δ | 21.1/3.7 2.44/- - | [249] |
| Flask, 6% Soluble starch, 0.17% (w/v) YNB, 0.15% (w/v) NH4Cl, pH 6, C/N 90. | o/e of DGA2, GPD1 and h.e. of alpha-amylase from Oryza sativa + glucoamylase from Aspergillus niger; pox1-6Δ and tgl4Δ | 27.0/- 3.32/- - | ||
| R. toruloides RT880-AD, derived from IFO 0880 | Bioreactor (batch), 150 g/L Glu, 0.5 g/L (NH4)2SO4, 1 g/L KH2PO4, 1 g/L MgSO4, 8 g/LYE. | o/e of ACC1 and DGA1 | 53.9/36.0 24.8/14.2 0.25/0.09 | [225] |
| R. toruloides RT880-ADS, derived fromRT880-AD | o/e of ACC1, DGA1, and SCD | 51.1/36.0 27.4/14.2 0.31/0.09 | ||
| R. toruloides RT880-ADM, derived fromRT880-AD | Flask, N-limited medium supplemented with 70 g/L Glu. | o/e of ACC1, DGA1 and malic enzyme | - 18.6/16.5 - | |
| R. toruloides TK16 | Flask, 70 g/L Glu, 0.55 g/L (NH4)2SO4, 0.4 g/L KH2PO4, 2 g/L MgSO4.7H2O, 0.75 g/L YE | o/e of Δ12-FAD | 27.0/16.0 5.9/2.5 - | [243] |
| R. toruloides TK16 | Flask, 70 g/L Glu, 0.55 g/L (NH4)2SO4, 0.4 g/L KH2PO4, 2 g/L MgSO4.7H2O, 0.75 g/L YE | o/e of Δ9-FAD + Δ12-FAD | 26.0/16.0 3.5/2.5 - | [243] |
| R. toruloides L1-1 | o/e of Δ12-FAD | 24.0/14.8 6.7/4.5 - | ||
| R. toruloides L1-1 | o/e of Δ9-FAD + Δ12-FAD | 20.0/14.8 6.0/4.5 - | ||
| R. toruloides NP-Pta-15, derived from NP11 | Flask, 50 g/L Glu, 1.5 g/L Mg2SO4.7H2O, 0.1 g/L (NH4)2SO4, 0.75 g/L YE. | o/e of phosphotransacetylase (Pta) | 65.6/62.1 - 0.05/0.03 | [250] |
| R. fluvialis DMKU-RK253 | Flask, 70 g/L crude Gly, 0.55 g/L (NH4)2SO4, 1 g/L MSG, 2 g/L MgSO4.7H2O, 0.4 g KH2PO4. | o/e of DGA1 | 18.53/6.11 1.2/0.47 - | [220] |
| C. phangngensis JQCP03H, derived from PT1-17 | Flask, Lipid production medium. 50 g/L Glu, 4 g/L pep, 1.5 g/L YE. | h.e. of DGA1 from Y. lipolytica | 63.3/52.1 11.4/8.3 - | [219] |
Notes: -, no data available. h.e.: heterologous expression; o/e: overexpression; Δ: deletion. Lipid content: g of produced lipids/g dry weight (%). Lipid titer: g of produced lipids/L of culture. Lipid productivity: g of produced/L of culture per hour. Abbreviations: Glycerol (Gly); Glucose (Glu); Monosodium glutamate (MSG); Sucrose (Suc); References (Ref.); YE: yeast extract; YNB-/-: yeast nitrogen base without amino acids and without ammonium sulfate.
7.3. Genetic Engineering of Substrate Utilisation Pathways, in Particular of Xylose
Numerous genetic manipulations were performed to increase the efficiency of consumption of substrates of difficult catabolism. As referred to in Section 4.2, Y. lipolytica possesses in its genome genes encoding xylose reductase (XR), xylitol dehydrogenase (XDH) and xylulose kinase (XK), but they are not sufficiently expressed to allow the efficient catabolism of xylose [125]. To address this issue, the heterologous expression of xylose pathway genes from microorganisms that use this sugar as a C-source, such as the yeast Scheffersomyces stipitis, was attempted (Table 7). Interestingly, the most efficient xylose consumption was obtained using a lipid-accumulating strain, referred to as the obese strain (overexpresses the G3P dehydrogenase GPD1 and the diacylglycerol O-acyl-transferase DGA2 genes and has the genes that code for the acyl-CoA oxidases (POX1-6) and the triacylglycerol lipase, TGL4, deleted) overexpressing the genes encoding the xylose reductase and xylitol dehydrogenase from S. stipitis and the xylulose kinase from Y. lipolytica [54].
The expression of genes involved in xylose metabolism, namely the endogenous phosphoketolase (PK) and the expression of a heterologous phosphotransacetylase (PTA) in Rhodosporidium azoricum also led to an increase of 89% in lipid yield using a culture medium with a mixture of glucose and xylose, without compromising biomass production and improving xylose utilisation [251]. As discussed in Section 4.2, xylose transport constitutes a limiting step in xylose utilisation. Transporters that allow the co-consumption of glucose and xylose were identified in C. tropicalis [29] and L. starkeyi [110] and may be considered targets for future genetic engineering of oleaginous yeasts. A new family of transporters very abundant in plant genomes is of potential interest. They belong to the Sugars Will Eventually be Exported Transporter (SWEET) superfamily and present a wide-ranging specificity and affinity towards a variety of mono- and disaccharide sugars [110,252]. Due to the high affinity for glucose and xylose of the transporters of this superfamily, they are pointed out as promising regarding the co-utilisation of both sugars being considered good targets for genetic manipulation of S. cerevisiae and other biotechnologically relevant yeasts [252].
Another sugar of more difficult catabolism is the acid sugar D-galacturonic acid, a monomer of pectin abundant in residues rich in pectin. Species of the Rhodotorula genus are able to efficiently catabolise D-galacturonic acid [39] but this is not the case for most of the yeast species, in particular S. cerevisiae [40]. A recent study involved the engineering of D-galacturonic acid catabolism in an S. cerevisiae strain previously equipped with a NAD-dependent glycerol catabolic pathway [253]. Although this study has proved the capacity to produce bioethanol, not lipids, from D-galacturonic acid in S. cerevisiae, results can be considered a proof of concept for the use as feedstocks two industrial organic residues/by-products such as the pectin-rich residues sugar beet pulp from sugar refinery or citrus peels, and crude glycerol, from the biodiesel industry [253].
Table 7.
Genetic modifications performed in Yarrowia lipolytica to improve xylose consumption. Strains of Y. lipolytica that are not genetically manipulated and not able to use xylose as C-source are considered controls. The meaning of the abbreviations used is explained at the end of the table. The underlined values correspond to the lipid production parameters obtained for the parental strain.
Table 7.
Genetic modifications performed in Yarrowia lipolytica to improve xylose consumption. Strains of Y. lipolytica that are not genetically manipulated and not able to use xylose as C-source are considered controls. The meaning of the abbreviations used is explained at the end of the table. The underlined values correspond to the lipid production parameters obtained for the parental strain.
| Strain | Experimental Condition | Genetic Modification | Consumed Xylose (g/Lh) | Lipid Content (%) Lipid Titer (g/L) Lipid Productivity (g/Lh) | Ref. |
|---|---|---|---|---|---|
| XYL+, derived from Po1d | Bioreactor (fed-batch), 150 g/L Xyl, 1.6 g/L NH4Cl, 1 g/L YE, 1 g/L YNB, 1.0 g/L MgCl2.7H2O, 0.5 g/L H2PO4. | o/e of XK h.e. of XR and XDH from Scheffersomyces stipitis | 2.14 | - 5.9 0.06 | [54] |
| XYL+Obese, derived from Po1d | o/e XK, DGA2 and GPD1 h.e. of XDH and XR from Scheffersomyces stipitis pox1-6Δ and tgl4Δ | - 20.1 0.19 | |||
| XYL+, derived from Po1d | Bioreactor (fed-batch), co-feeding with Gly, 150 g/L Xyl, 1.6 g/L NH4Cl, 1 g/L YE, 1 g/L YNB, 1.0 g/L MgCl2.7H2O, 0.5 g/L H2PO4. | o/e of XK h.e. of XR and XDH from Scheffersomyces stipitis | - | - 7.3 0.03 | |
| XYL+Obese, derived from Po1d | Bioreactor (fed-batch), co-feeding with Gly, 150 g/L Xyl, 1.6 g/L NH4Cl, 1 g/L YE, 1 g/L YNB, 1.0 g/L MgCl2.7H2O, 0.5 g/L H2PO4. | o/e of DGA2, GDP1, XK h.e. of XR and XDH from Scheffersomyces stipitis pox1-6Δ and tgl4Δ | - | - 50.5 0.23 | |
| YlXYL+Obese-XA, derived from Po1d | Bioreactor (fed-batch), Lignocellulosic hydrolysate from agave with 18% Glu and 12% Xyl, C/N 15. | o/e of XDH, XR, XK, DGA2 and GPD1 h.e. XPKA and ACK from Aspergillus nidulans pox1-6Δ and tgl4Δ | 0.47 | 67 16.5/2.0 0.185 | [254] |
| E26 XUS, derived from E26 | Bioreactor (batch),160 g/L Xyl, YNB with 10 g/L (NH₄)₂SO₄. | h.e. of XYL1 and XYL2 from Scheffersomyces stipitis | 0.74 | - 15.1 0.19 | [255] |
| YSXID, derived from Po1f | Bioreactor (batch), 80 g/L Glu and 80 g/L Xyl, 0.69 g/L CSM Leu/Ura, 1.76 g/L YNB−/−, 3.52 g/L (NH₄)₂SO₄, C/N 100. | Genetic background YSX (obtained in an ALE experiment) o/e of DGA1 and XK h.e. of xylose isomerase mutant gene pex10Δ | 0.08 | 56.7/51.6 13.5/7.3 - | [256] |
| YSXID, derived from Polf | Bioreactor (fed-batch), Lignocellulosic hydrolysate from Miscanthus sacchariflorus supplemented with Glu and Xyl at a final concentration of 35 g/L. | YSX background (obtained in an ALE experiment) o/e of DGA1 and XK h.e. of xylose isomerase mutant gene pex10Δ | - | 42.4/- 12.01/- - | [256] |
| PSA02004PP, derived from PSA02004 | Bioreactor (batch), Minimal medium with 60g/L Xyl. | o/e of XR, XDH and XK. | 0.71 | - - - | [257] |
| Y14, derived from ATCC 201249 | Bioreactor (batch), YPX (2% Xyl). | ku70Δo/e. of XKS, tHMG1, ERG9, ERG20, TKL, TAL1, TX. h.e. of DS, PPDS, ATR1 h.e. of XYL1 and XYL2 from Scheffersomyces stipitis Adaptation step in xylose. | 0.56 | - - - | [258] |
| YBX08, derived from PDe1 | Flas, YP with 40 g/L Xyl. | o/e of XK, tLS, tNDPS. HMG1 and ERG12 h.e. of XR and XDH from Scheffersomyces stipitis | ~0.56 | - - - | [259] |
| Yl -nar05, derived from Po1f | Flask, YPX medium, with 40 g/L Xyl. | o/e of XDH, XKS and TAL2. | 0.56 | - - - | [260] |
Notes: -, no data available. h.e.: heterologous expression; o/e: overexpression; Δ: deletion. Lipid content: g lipids/g dry weight (%); Lipid concentration: g of produced lipids/L of culture; Lipid yield: g of produced lipids/L of culture per hour. Abbreviations: Acetate kinase (ACK); DS (DMD synthase); DGA2 (acyl-CoA: diacylglycerol acyltransferase); GPD1 (G3P dehydrogenase); ERG12 (mevalonate kinase); HMG1 (HMG-CoA reductase gene); Phosphoketolase (XPKA); POX1-6 genes (acyl-CoA oxidases); PPDS (PPD synthase) tLS (d-limonene synthase from Agastache rugosa); TAL1 (transaldolase); TAL2 (tyrosine ammonia lyase); TKL (transketolase); tNDPS (neryl di-phosphate synthase 1 from Solanum lycopersicum); TX (xylose transporter); TGL4 (Triacylglycerol lipase 4); Xylitol dehydrogenase (XDH); Xylulose kinase (XK), Xylose reductase (XR) and YNB−/− (yeast nitrogen base without amino acids and without ammonium sulphate).
7.4. Genetic Engineering to Increase Yeast Tolerance to Stress Factors
The unveiling of the molecular mechanisms and functional pathways involved in yeast cell response to toxicants is essential to guide the genetic manipulation of oleaginous yeasts to improve tolerance. The use of lignocellulosic and industrial organic residues biomass for the production of added-value chemicals is a challenging task since yeast cells need to cope with multiple bioprocess-related stresses, either individually or combined, emphasising the relevance of enhancing multiple stress tolerance to maximise their performance in industrial production [89,261,262,263,264]. Physical and chemical extracellular stresses include non-optimum ranges of temperature and pH, osmotic pressure and the presence of growth inhibitors [89,261,262,263,264]. Despite being considered synonyms in some contexts, the concepts of tolerance and robustness may not coincide. Tolerance is defined as the ability of a cell to grow in the presence of single or multiple perturbations whereas the robustness concept is based on the stability of specific phenotypic traits in a multi-stress environment [136]. Thus, having tolerant and robust strains is fundamental for developing the sustainable production of lipid-based biofuels by yeasts. Moreover, yeast resilience, i.e., yeast’s ability to recover from a large environmental perturbation [265], is also important in the biorefinery context.
Concerning oleaginous yeasts, the individual and combined effect of six inhibitors from three major groups of inhibitors (furaldehydes, aromatics and weak acids) was investigated in Y. lipolytica overexpressing the endogenous xylose reductase, xylitol dehydrogenase, and xylulose kinase grown in glucose and in xylose [94]. The obtained results were similar in both C-sources, being cinnamic acid and coniferyl aldehyde tolerated while furfural contributed to an extended lag phase and hydroxymethylfurfural was responsible for partial growth inhibition [94]. Formic acid only compromised growth at concentrations above 25 mM [94]. A recent study identified that the native furfural detoxification mechanism and furfural resistance were increased through the rational engineering of Y. lipolytica, by the overexpression of aldehyde dehydrogenase endogenous genes to enhance the conversion of furfural to furoic acid [266]. The most promising result was obtained for the aldehyde dehydrogenase FALDH2, leading to the highest conversion rate of furfural to furoic acid, as well as a two-fold increase in cell growth and lipid production in the presence of 0.4 g/L of furfural [266]. The thermotolerant L1–1 strain of R. toruloides, obtained by an adaptive breeding strategy [267], was also found to tolerate (i) oxidative stress (ethanol and hydrogen peroxide), (ii) osmotic stress (high glucose concentrations), and (iii) cell membrane disturbing reagent (DMSO) [268]. This strain, which produced high titers of lipids, was able to cope with the increase in ROS and presented a stronger cell wall and increased levels of unsaturated membrane lipids under various stresses [268].
It is known that growth inhibitors present in lignocellulosic hydrolysates may compromise the integrity, fluidity and selective permeability of yeast plasma membrane [269]. For this reason, the majority of membrane engineering attempts to increase tolerance to multiple stresses target the modulation of its lipid composition, in order to maintain the integrity and fluidity under stress, namely by altering lipid saturation or changing the length of lipid in biomembranes [264]. The genetic manipulation of oleaginous yeasts, comprising the degree of saturation of lipids [200,223,243,244,270,271,272] or the length of the lipidic chain [273,274] led to increased lipid titers. Additionally, membrane proteins including integral membrane proteins and transport proteins are also extremely relevant in stress tolerance. A remarkable example is overexpression in Y. lipolytica of the gene MFS1, a putative MFS transporter, that led to an increased propionate tolerance [275]. However, studies on the role of transporters in stress tolerance are scarce in oleaginous yeasts but this is a research topic explored in the yeast model S. cerevisiae. A relevant example regarding transport proteins and stress level involves TRK1, encoding the high-affinity potassium transporter and a major determinant of tolerance to acetic acid in S. cerevisiae [84]. Potassium supplementation up to the required level was described as involved in the tolerance to a wide variety of stresses [84,276] and, recently, it was demonstrated that S. cerevisiae cells react to potassium concentration by a rapid, continuous, and precise adjustment of both the affinity and maximum velocity of their Trk1 protein [277]. However, the role of Trk1 in tolerance to different stresses is not completely clear: under formic acid (C1) stress, the deletion of TRK1 led to increased tolerance to formic acid, contrasting with what was described for acetic acid and the demonstrated role that K+ concentration has in tolerance [278]. The relevance of plasma membrane efflux pumps in the development of superior yeasts was recently reviewed [135]. Furthermore, the genetic alterations (gene deletion or overexpression) with a direct effect on the tolerance of S. cerevisiae towards individual inhibitors or combinations of inhibitors found in lignocellulosic hydrolysates or other feedstocks of interest in the context of a circular bio-economy were compiled in several works [81,89,262,279,280,281]. For example, the overexpression of Ace2, a transcription factor required for septum destruction after cytokinesis and Sfp1, a transcription factor that regulates ribosomal protein and biogenesis genes in S. cerevisiae, was beneficial to increase tolerance to acetic acid, furfural, and a mixture of acetic acid and furfural [282]. Regarding another example of transcription factor engineering, a single amino acid exchange at position 135 (serine to phenylalanine) in Haa1, a major TF involved in adaptation and tolerance to acetic and formic acids stresses [85,283], contributed to an increase in acetic acid tolerance [284].
7.5. Adaptive Laboratory Evolution (ALE) to Improve Yeast Robustness and Substrate Utilisation
Adaptive laboratory evolution (ALE) techniques [285,286] are a suitable alternative to the use of genetic engineering when the necessary efficient tools are not available as is the case for most of the non-Saccharomyces yeasts and they also constitute a highly convenient strategy to complement genetic manipulation experiments in Y. lipolytica [256,287,288]. The major objectives for such adaptation are to increase in tolerance of the producing yeasts to the inhibitors present in lignocellulosic hydrolysates or other residual feedstocks and the efficiency of substrate utilisation [285,286]. Of course, in the specific context of this review paper, this is an essential objective to assure yeast robustness [136] under the above-referred challenging conditions or, in other words, to maintain the high levels of oil production in the evolved strains. For example, a genetically manipulated strain of R. toruloides was adapted to undetoxified wheat straw hydrolysates leading to a higher rate of xylose consumption [289]. The genes DGAT1 (diacylglycerol acyl-CoA transferase type 2) and SCD1 (stearoyl-CoA desaturase), under the control of the xylose reductase (XYL1) promoter, previously reported to enhance lipid production in oleaginous yeasts [230,289] were overexpressed in this improved strain, leading to a lipid concentration of 39.5 g/L and lipid productivity of 0.334 g/Lh, representing the highest values described in the literature [289]. Considering the tolerance to the inhibitors present in lignocellulosic hydrolysate, an ALE attempt also using R. toruloides was performed to increase tolerance to HMF, furfural, acetic acid, and better performance for the accumulation of lipids and carotenoids was obtained [290]. Increased tolerance to inhibitors present in lignocellulosic biomass, either alone (in the case of formic acid) or in a mixture of inhibitors (formic acid, acetic acid, furfural and HMF) in Metshnikowia pulcherrima was also obtained using ALE, leading to a decreased duration of the latency period and an increased specific growth rate after growth resumption [291]. The lipid content was also increased by 50% in the evolved strain compared to the parental strain [291].
8. Conclusions and Future Perspectives
The efficient use of yeasts as cell factories for the production of lipids from lignocellulosic biomasses or other residual feedstocks is a major challenge in the transition towards a sustainable and low-carbon bio-economy. The genetic and physiological diversity of oleaginous yeasts is an advantage for the transition to greener biofuels given that some of them are able to efficiently consume all the carbon sources present in those feedstocks and tolerate the growth and metabolism inhibitors that are present or that result from their pre-treatment. In order to increase the performance of lipid production, it is crucial to match the oleaginous yeast species/strain to be used in the chosen feedstock. Although Rhodotorula toruloides and Yarrowia lipolytica stand out in the scientific literature as the most studied and promising yeast species, strain performance is highly variable and other yeast species/strains are also emerging as highly promising. However, while R. toruloides is characterised to efficiently utilise most of the sugars and other carbon sources present in biomass hydrolysates, has a high lipid production yield and is robust, Y. lipolytica allows easy genetic manipulation since several efficient genetic tools are already available. Multidisciplinary approaches, combining and integrating data from genome-wide analyses, the exploration of metabolic models and a holistic understanding of the physiology of these yeasts are expected to guide the rational construction of yeasts with superior characteristics and their use under appropriate conditions. This will contribute to rendering current biodiesel production greener and making the bioprocess more economically sustainable.
It is worth mentioning that the fatty acids from the TAGs, produced by native and engineered oleaginous yeast strains by modifying the lipid profile toward other chain lengths and saturation types, can also be used to produce other products than biodiesel for higher value markets (pharmaceuticals, nutraceuticals, cosmetics, food) [231,292,293,294].
Author Contributions
Conceptualisation, I.S.-C.; methodology, I.S.-C. and M.N.M.; validation, I.S.-C., investigation, I.S.-C., M.N.M. and P.M.; writing—original draft preparation, M.N.M.; writing—review and editing, I.S.-C. and M.N.M.; supervision, I.S.-C.; project administration, I.S.-C.; funding acquisition, I.S.-C. All authors have read and agreed to the published version of the manuscript.
Funding
Research work in the laboratory of I.S.-C. on oleaginous yeasts is funded by Move2LowC project (POCI-01-0247-FEDER-046117), cofinanced by Programa Operacional Competitividade e Internacionalização (POCI), Programa Operacional Regional de Lisboa, Portugal 2020 and the European Union, through the European Regional Development Fund (ERDF). It was also partially funded by ‘Fundação para a Ciência e a Tecnologia’ (FCT) project ERA-IB-2/0003/2015 and a Ph.D. fellowship to MNM (FCT-PhD Programme AEM—Applied and Environmental Microbiology PD/BD/146167/2019). Funding received from FCT by iBB—Institute for Bioengineering and Biosciences (UIDB/04565/2020 and UIDP/04565/2020) and by i4HB (LA/P/0140/2020), is also acknowledged. PM thanks to North Portugal Regional Operational Program (Norte2020) under the Portugal 2020 (Operational Program for Competitiveness and Internationalisation) for the Grant (NORTE-06-3559-FSE-000070) and to FCT.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Leong, H.Y.; Chang, C.-K.; Khoo, K.S.; Chew, K.W.; Chia, S.R.; Lim, J.W.; Chang, J.-S.; Show, P.L. Waste biorefinery towards a sustainable circular bioeconomy: A solution to global issues. Biotechnol. Biofuels 2021, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chandel, A.K.; Garlapati, V.K.; Kumar, S.P.J.; Hans, M.; Singh, A.K. The role of renewable chemicals and biofuels in building a bioeconomy. Biofuels Bioprod. Biorefin. 2020, 14, 830–844. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Sartaj, K.; Pruthi, V.; Pruthi, P.A. Sustainable biodiesel production from oleaginous yeasts utilizing hydrolysates of various non-edible lignocellulosic biomasses. Renew. Sustain. Energy Rev. 2016, 62, 836–855. [Google Scholar] [CrossRef]
- Ahmed, M.; Abdullah, A.; Patle, D.S.; Shahadat, M.; Ahmad, Z.; Athar, M.; Aslam, M.; Vo, D.-V.N. Feedstocks, catalysts, process variables and techniques for biodiesel production by one-pot extraction-transesterification: A review. Environ. Chem. Lett. 2021, 20, 335–378. [Google Scholar] [CrossRef]
- Chatzifragkou, A.; Papanikolaou, S. Effect of impurities in biodiesel-derived waste glycerol on the performance and feasibility of biotechnological processes. Appl. Microbiol. Biotechnol. 2012, 95, 13–27. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Sakinah, A.M.M.; Zularisam, A.W.; Sirohi, R.; Khilji, I.A.; Reddy, V.J.; Pandey, A. Bioconversion of Glycerol into Biofuels—Opportunities and Challenges. BioEnergy Res. 2021, 1–16. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Sakinah, A.M.; Zularisam, A. Opportunities of biodiesel industry waste conversion into value-added products. Mater. Today Proc. 2021, 57, 1014–1020. [Google Scholar] [CrossRef]
- Kamineni, A.; Consiglio, A.L.; MacEwen, K.; Chen, S.; Chifamba, G.; Shaw, A.J.; Tsakraklides, V. Increasing lipid yield in Yarrowia lipolytica through phosphoketolase and phosphotransacetylase expression in a phosphofructokinase deletion strain. Biotechnol. Biofuels 2021, 14, 1–14. [Google Scholar] [CrossRef]
- Beopoulos, A.; Nicaud, J.-M.; Gaillardin, C. An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl. Microbiol. Biotechnol. 2011, 90, 1193–1206. [Google Scholar] [CrossRef]
- Saha, R.; Mukhopadhyay, M. Prospect of metabolic engineering in enhanced microbial lipid production: Review. Biomass Convers. Biorefin. 2021, 1–22. [Google Scholar] [CrossRef]
- Caporusso, A.; Capece, A.; De Bari, I. Oleaginous Yeasts as Cell Factories for the Sustainable Production of Microbial Lipids by the Valorization of Agri-Food Wastes. Fermentation 2021, 7, 50. [Google Scholar] [CrossRef]
- Ratledge, C. Resources conservation by novel biological processes. I Grow fats from wastes. Chem. Soc. Rev. 1979, 8, 283–296. [Google Scholar] [CrossRef]
- Rebello, S.; Abraham, A.; Madhavan, A.; Sindhu, R.; Binod, P.; Babu, A.K.; Aneesh, E.M.; Pandey, A. Non-conventional Yeast cell factories for sustainable bioprocesses. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Sun, L.; Alper, H.S. Non-conventional hosts for the production of fuels and chemicals. Curr. Opin. Chem. Biol. 2020, 59, 15–22. [Google Scholar] [CrossRef]
- Spencer, A.R.D.S.J.; Spencer, J.; Laluce, C. Non-conventional yeasts. Appl. Microbiol. Biotechnol. 2002, 58, 147–156. [Google Scholar] [CrossRef]
- Patra, P.; Das, M.; Kundu, P.; Ghosh, A. Recent advances in systems and synthetic biology approaches for developing novel cell-factories in non-conventional yeasts. Biotechnol. Adv. 2021, 47, 107695. [Google Scholar] [CrossRef]
- Geijer, C.; Ledesma-Amaro, R.; Tomás-Pejó, E. Unraveling the potential of non-conventional yeasts in biotechnology. FEMS Yeast Res. 2022, 22. [Google Scholar] [CrossRef]
- Abeln, F.; Chuck, C.J. The history, state of the art and future prospects for oleaginous yeast research. Microb. Cell Fact. 2021, 20, 1–31. [Google Scholar] [CrossRef]
- Llamas, M.; Magdalena, J.A.; González-Fernández, C.; Tomás-Pejó, E. Volatile fatty acids as novel building blocks for oil-based chemistry via oleaginous yeast fermentation. Biotechnol. Bioeng. 2019, 117, 238–250. [Google Scholar] [CrossRef]
- Valdés, G.; Mendonça, R.; Aggelis, G. Lignocellulosic Biomass as a Substrate for Oleaginous Microorganisms: A Review. Appl. Sci. 2020, 10, 7698. [Google Scholar] [CrossRef]
- Liu, H.; Song, Y.; Fan, X.; Wang, C.; Lu, X.; Tian, Y. Yarrowia lipolytica as an Oleaginous Platform for the Production of Value-Added Fatty Acid-Based Bioproducts. Front. Microbiol. 2021, 11, 608662. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Maiti, M.K. Lipid production by oleaginous yeasts. Adv. Appl. Microbiol. 2021, 116, 1–98. [Google Scholar] [CrossRef] [PubMed]
- Yurkov, A.M. Yeasts of the soil—Obscure but precious. Yeast 2018, 35, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasconcelos, B.; Teixeira, J.C.; Dragone, G. Oleaginous yeasts for sustainable lipid production—From biodiesel to surf boards, a wide range of “green” applications. Appl. Microbiol. Biotechnol. 2019, 103, 3651–3667. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, F.A.G.; Colen, G.; Takahashi, J.A. Yarrowia lipolytica and Its Multiple Applications in the Biotechnological Industry. Sci. World J. 2014, 2014, 476207. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Alper, H.S. Draft Genome Sequence of the Oleaginous Yeast Yarrowia lipolytica PO1f, a Commonly Used Metabolic Engineering Host. Genome Announc. 2014, 2, e00652-14. [Google Scholar] [CrossRef] [Green Version]
- Larroude, M.; Rossignol, T.; Nicaud, J.-M.; Ledesma-Amaro, R. Synthetic biology tools for engineering Yarrowia lipolytica. Biotechnol. Adv. 2018, 36, 2150–2164. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Mitra, M.; Maiti, M.K. Recent advances in lipid metabolic engineering of oleaginous yeasts. Biotechnol. Adv. 2021, 53, 107722. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts. A Taxonomic Study, 5th ed.; Elsevier: New York, NY, USA, 2011. [Google Scholar]
- Yeeh, Y. Rhodotorula. Encycl. Food Microbiol. 1999, 1900–1905. [Google Scholar]
- Park, Y.-K.; Nicaud, J.-M.; Ledesma-Amaro, R. The Engineering Potential of Rhodosporidium toruloides as a Workhorse for Biotechnological Applications. Trends Biotechnol. 2018, 36, 304–317. [Google Scholar] [CrossRef]
- Gong, Z.; Shen, H.; Zhou, W.; Wang, Y.; Yang, X.; Zhao, Z.K. Efficient conversion of acetate into lipids by the oleaginous yeast Cryptococcus curvatus. Biotechnol. Biofuels 2015, 8, 189. [Google Scholar] [CrossRef]
- Chen, X.; Li, Z.; Zhang, X.; Hu, F.; Ryu, D.D.Y.; Bao, J. Screening of Oleaginous Yeast Strains Tolerant to Lignocellulose Degradation Compounds. Appl. Biochem. Biotechnol. 2009, 159, 591–604. [Google Scholar] [CrossRef]
- Huang, C.; Wu, H.; Smith, T.J.; Liu, Z.-J.; Lou, W.-Y.; Zong, M.-H. In vivo detoxification of furfural during lipid production by the oleaginous yeast Trichosporon fermentans. Biotechnol. Lett. 2012, 34, 1637–1642. [Google Scholar] [CrossRef]
- Yu, X.; Zheng, Y.; Dorgan, K.M.; Chen, S. Oil production by oleaginous yeasts using the hydrolysate from pretreatment of wheat straw with dilute sulfuric acid. Bioresour. Technol. 2011, 102, 6134–6140. [Google Scholar] [CrossRef]
- Wang, J.; Gao, Q.; Zhang, H.; Bao, J. Inhibitor degradation and lipid accumulation potentials of oleaginous yeast Trichosporon cutaneum using lignocellulose feedstock. Bioresour. Technol. 2016, 218, 892–901. [Google Scholar] [CrossRef]
- Huang, C.; Wu, H.; Li, R.-F.; Zong, M.-H. Improving lipid production from bagasse hydrolysate with Trichosporon fermentans by response surface methodology. New Biotechnol. 2011, 29, 372–378. [Google Scholar] [CrossRef]
- Martins, L.; Palma, M.; Angelov, A.; Nevoigt, E.; Liebl, W.; Sá-Correia, I. Complete Utilization of the Major Carbon Sources Present in Sugar Beet Pulp Hydrolysates by the Oleaginous Red Yeasts Rhodotorula toruloides and R. mucilaginosa. J. Fungi 2021, 7, 215. [Google Scholar] [CrossRef]
- Martins, L.M.; Monteiro, C.C.; Semedo, P.M.; Sá-Correia, I. Valorisation of pectin-rich agro-industrial residues by yeasts: Potential and challenges. Appl. Microbiol. Biotechnol. 2020, 104, 6527–6547. [Google Scholar] [CrossRef]
- Quarterman, J.; Slininger, P.J.; Kurtzman, C.P.; Thompson, S.R.; Dien, B.S. A survey of yeast from the Yarrowia clade for lipid production in dilute acid pretreated lignocellulosic biomass hydrolysate. Appl. Microbiol. Biotechnol. 2016, 101, 3319–3334. [Google Scholar] [CrossRef]
- Wang, J.; Ledesma-Amaro, R.; Wei, Y.; Ji, B.; Ji, X.-J. Metabolic engineering for increased lipid accumulation in Yarrowia lipolytica—A Review. Bioresour. Technol. 2020, 313, 123707. [Google Scholar] [CrossRef]
- Chmielarz, M.; Blomqvist, J.; Sampels, S.; Sandgren, M.; Passoth, V. Microbial lipid production from crude glycerol and hemicellulosic hydrolysate with oleaginous yeasts. Biotechnol. Biofuels 2021, 14, 1–11. [Google Scholar] [CrossRef]
- Klein, M.; Swinnen, S.; Thevelein, J.M.; Nevoigt, E. Glycerol metabolism and transport in yeast and fungi: Established knowledge and ambiguities. Environ. Microbiol. 2016, 19, 878–893. [Google Scholar] [CrossRef] [Green Version]
- Kumar, L.R.; Yellapu, S.K.; Tyagi, R.; Zhang, X. A review on variation in crude glycerol composition, bio-valorization of crude and purified glycerol as carbon source for lipid production. Bioresour. Technol. 2019, 293, 122155. [Google Scholar] [CrossRef]
- Leung, D.Y.; Wu, X.; Leung, M. A review on biodiesel production using catalyzed transesterification. Appl. Energy 2010, 87, 1083–1095. [Google Scholar] [CrossRef]
- Fei, Q.; O’Brien, M.; Nelson, R.; Chen, X.; Lowell, A.; Dowe, N. Enhanced lipid production by Rhodosporidium toruloides using different fed-batch feeding strategies with lignocellulosic hydrolysate as the sole carbon source. Biotechnol. Biofuels 2016, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tanimura, A.; Takashima, M.; Sugita, T.; Endoh, R.; Kikukawa, M.; Yamaguchi, S.; Sakuradani, E.; Ogawa, J.; Shima, J. Selection of oleaginous yeasts with high lipid productivity for practical biodiesel production. Bioresour. Technol. 2013, 153, 230–235. [Google Scholar] [CrossRef]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process. Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]
- Chen, Y.; Nie, X.; Ye, J.; Wang, Y.; Chen, J.; Xu, J. Biodiesel from Microorganisms: A Review. Energy Technol. 2021, 9. [Google Scholar] [CrossRef]
- Evans, C.T.; Scragg, A.H.; Ratledge, C. Regulation of Citrate Efflux from Mitochondria of Oleaginous and Non-Oleaginous Yeasts by Long-Chain Fatty Acyl-CoA Esters. Eur. J. Biochem. 1983, 132, 617–622. [Google Scholar] [CrossRef]
- Botham, P.A.; Ratledge, C. A Biochemical Explanation for Lipid Accumulation in Candida 107 and Other Oleaginous Micro-organisms. J. Gen. Microbiol. 1979, 114, 361–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, L.; Palmieri, F.; Runswick, M.J.; Walker, J.E. Identification by bacterial expression and functional reconstitution of the yeast genomic sequence encoding the mitochondrial dicarboxylate carrier protein. FEBS Lett. 1996, 399, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Ledesma-Amaro, R.; Lazar, Z.; Rakicka, M.; Guo, Z.; Fouchard, F.; Crutz-Le Coq, A.-M.; Nicaud, J.-M. Metabolic engineering of Yarrowia lipolytica to produce chemicals and fuels from xylose. Metab. Eng. 2016, 38, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Modeling Lipid Accumulation and Degradation in Yarrowia lipolytica Cultivated on Industrial Fats. Curr. Microbiol. 2003, 46, 398–402. [Google Scholar] [CrossRef]
- Beopoulos, A.; Mrozova, Z.; Thevenieau, F.; Le Dall, M.-T.; Hapala, I.; Papanikolaou, S.; Chardot, T.; Nicaud, J.-M. Control of Lipid Accumulation in the Yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 7779–7789. [Google Scholar] [CrossRef] [Green Version]
- Silverman, A.M.; Qiao, K.; Xu, P.; Stephanopoulos, G. Functional overexpression and characterization of lipogenesis-related genes in the oleaginous yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2016, 100, 3781–3798. [Google Scholar] [CrossRef] [Green Version]
- Chaiyaso, T.; Manowattana, A.; Techapun, C.; Watanabe, M. Efficient bioconversion of enzymatic corncob hydrolysate into biomass and lipids by oleaginous yeast Rhodosporidium paludigenum KM281510. Prep. Biochem. Biotechnol. 2019, 49, 545–556. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, S.; Hu, C.; Wang, Q.; Hua, Y.; Zhao, Z.K. Lipid production from Jerusalem artichoke by Rhodosporidium toruloides Y4. J. Ind. Microbiol. Biotechnol. 2010, 37, 581–585. [Google Scholar] [CrossRef]
- Slininger, P.J.; Dien, B.; Kurtzman, C.P.; Moser, B.; Bakota, E.L.; Thompson, S.R.; O’Bryan, P.J.; Cotta, M.A.; Balan, V.; Jin, M.; et al. Comparative lipid production by oleaginous yeasts in hydrolyzates of lignocellulosic biomass and process strategy for high titers. Biotechnol. Bioeng. 2016, 113, 1676–1690. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-E.; Vadlani, P.V.; Min, D. Sustainable Production of Microbial Lipids from Lignocellulosic Biomass Using Oleaginous Yeast Cultures. J. Sustain. Bioenergy Syst. 2017, 07, 36–50. [Google Scholar] [CrossRef] [Green Version]
- i Nogué, V.S.; Black, B.A.; Kruger, J.S.; Singer, C.A.; Ramirez, K.J.; Reed, M.L.; Cleveland, N.S.; Singer, E.R.; Yi, X.; Yeap, R.Y.; et al. Integrated diesel production from lignocellulosic sugars via oleaginous yeast. Green Chem. 2018, 20, 4349–4365. [Google Scholar] [CrossRef]
- Sitepu, I.; Jin, M.; Fernandez, J.E.; Sousa, L.D.C.; Balan, V.; Boundy-Mills, K.L. Identification of oleaginous yeast strains able to accumulate high intracellular lipids when cultivated in alkaline pretreated corn stover. Appl. Microbiol. Biotechnol. 2014, 98, 7645–7657. [Google Scholar] [CrossRef]
- Xavier, M.; Coradini, A.; Deckmann, A.; Franco, T. Lipid production from hemicellulose hydrolysate and acetic acid by Lipomyces starkeyi and the ability of yeast to metabolize inhibitors. Biochem. Eng. J. 2017, 118, 11–19. [Google Scholar] [CrossRef]
- Xavier, M.D.C.A.; Franco, T.T. Obtaining hemicellulosic hydrolysate from sugarcane bagasse for microbial oil production by Lipomyces starkeyi. Biotechnol. Lett. 2021, 43, 967–979. [Google Scholar] [CrossRef]
- Azad, A.K. Production of Microbial Lipids from Rice Straw Hydrolysates by Lipomyces starkeyi for Biodiesel Synthesis. J. Microb. Biochem. Technol. 2014, s8. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, Y.; Liu, H.; Zhang, J. Enhanced lipid production with undetoxified corncob hydrolysate by Rhodotorula glutinis using a high cell density culture strategy. Bioresour. Technol. 2015, 180, 32–39. [Google Scholar] [CrossRef]
- Galafassi, S.; Cucchetti, D.; Pizza, F.; Franzosi, G.; Bianchi, D.; Compagno, C. Lipid production for second generation biodiesel by the oleaginous yeast Rhodotorula graminis. Bioresour. Technol. 2012, 111, 398–403. [Google Scholar] [CrossRef]
- Miao, Z.; Tian, X.; Liang, W.; He, Y.; Wang, G. Bioconversion of corncob hydrolysate into microbial lipid by an oleaginous yeast Rhodotorula taiwanensis AM2352 for biodiesel production. Renew. Energy 2020, 161, 91–97. [Google Scholar] [CrossRef]
- de Oliveira, P.M.; Aborneva, D.; Bonturi, N.; Lahtvee, P.-J. Screening and Growth Characterization of Non-conventional Yeasts in a Hemicellulosic Hydrolysate. Front. Bioeng. Biotechnol. 2021, 9, 1–13. [Google Scholar] [CrossRef]
- Poontawee, R.; Yongmanitchai, W.; Limtong, S. Lipid production from a mixture of sugarcane top hydrolysate and biodiesel-derived crude glycerol by the oleaginous red yeast, Rhodosporidiobolus fluvialis. Process Biochem. 2018, 66, 150–161. [Google Scholar] [CrossRef]
- Gao, Q.; Cui, Z.; Zhang, J.; Bao, J. Lipid fermentation of corncob residues hydrolysate by oleaginous yeast Trichosporon cutaneum. Bioresour. Technol. 2014, 152, 552–556. [Google Scholar] [CrossRef]
- Chen, X.-F.; Huang, C.; Xiong, L.; Chen, X.-D.; Ma, L.-L. Microbial oil production from corncob acid hydrolysate by Trichosporon cutaneum. Biotechnol. Lett. 2012, 34, 1025–1028. [Google Scholar] [CrossRef]
- Hu, C.; Wu, S.; Wang, Q.; Jin, G.; Shen, H.; Zhao, Z.K. Simultaneous utilization of glucose and xylose for lipid production by Trichosporon cutaneum. Biotechnol. Biofuels 2011, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Xu, Z.; Chen, S.; Jin, M. Microbial lipid production from dilute acid and dilute alkali pretreated corn stover via Trichosporon dermatis. Bioresour. Technol. 2019, 295, 122253. [Google Scholar] [CrossRef]
- Huang, C.; Chen, X.-F.; Xiong, L.; Chen, X.-D.; Ma, L.-L. Oil production by the yeast Trichosporon dermatis cultured in enzymatic hydrolysates of corncobs. Bioresour. Technol. 2012, 110, 711–714. [Google Scholar] [CrossRef]
- Tsigie, Y.A.; Wang, C.-Y.; Truong, C.-T.; Ju, Y.-H. Lipid production from Yarrowia lipolytica Po1g grown in sugarcane bagasse hydrolysate. Bioresour. Technol. 2011, 102, 9216–9222. [Google Scholar] [CrossRef]
- Santek, M.I.; Beluhan, S.; Santek, B. Production of Microbial Lipids from Lignocellulosic Biomass. Adv. Biofuels Bioenergy 2018, 137–164. [Google Scholar] [CrossRef] [Green Version]
- Beig, B.; Riaz, M.; Naqvi, S.R.; Hassan, M.; Zheng, Z.; Karimi, K.; Pugazhendhi, A.; Atabani, A.E.; Chi, N.T.L. Current challenges and innovative developments in pretreatment of lignocellulosic residues for biofuel production: A review. Fuel 2020, 287, 119670. [Google Scholar] [CrossRef]
- Zha, Y.; Westerhuis, J.A.; Muilwijk, B.; Overkamp, K.M.; Nijmeijer, B.M.; Coulier, L.; Smilde, A.K.; Punt, P.J. Identifying inhibitory compounds in lignocellulosic biomass hydrolysates using an exometabolomics approach. BMC Biotechnol. 2014, 14, 22. [Google Scholar] [CrossRef] [Green Version]
- Vanmarcke, G.; Demeke, M.M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Identification of the major fermentation inhibitors of recombinant 2G yeasts in diverse lignocellulose hydrolysates. Biotechnol. Biofuels 2021, 14, 1–15. [Google Scholar] [CrossRef]
- Klinke, H.B.; Thomsen, A.B.; Ahring, B.K. Inhibition of ethanol-producing yeast and bacteria by degradation products produced during pre-treatment of biomass. Appl. Microbiol. Biotechnol. 2004, 66, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Kawahata, M.; Masaki, K.; Fujii, T.; Iefuji, H. Yeast genes involved in response to lactic acid and acetic acid: Acidic conditions caused by the organic acids in Saccharomyces cerevisiae cultures induce expression of intracellular metal metabolism genes regulated by Aft1p. FEMS Yeast Res. 2006, 6, 924–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mira, N.P.; Palma, M.; Guerreiro, J.F.; Sá-Correia, I. Genome-wide identification of Saccharomyces cerevisiae genes required for tolerance to acetic acid. Microb. Cell Fact. 2010, 9, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, M.; Guerreiro, J.F.; Sá-Correia, I. Adaptive Response and Tolerance to Acetic Acid in Saccharomyces cerevisiae and Zygosaccharomyces bailii: A Physiological Genomics Perspective. Front. Microbiol. 2018, 9, 274. [Google Scholar] [CrossRef]
- Palma, M.; Sá-Correia, I. Physiological Genomics of the Highly Weak-Acid-Tolerant Food Spoilage Yeasts of Zygosaccharomyces bailii sensu lato. Yeasts Biotechnol. Hum. Health 2019, 58, 85–109. [Google Scholar] [CrossRef]
- Xu, J.; Liu, N.; Qiao, K.; Vogg, S.; Stephanopoulos, G. Application of metabolic controls for the maximization of lipid production in semicontinuous fermentation. Proc. Natl. Acad. Sci. USA 2017, 114, E5308–E5316. [Google Scholar] [CrossRef] [Green Version]
- Cordova, L.T.; Lad, B.C.; Ali, S.A.; Schmidt, A.J.; Billing, J.; Pomraning, K.; Hofstad, B.; Swita, M.S.; Collett, J.R.; Alper, H.S. Valorizing a hydrothermal liquefaction aqueous phase through co-production of chemicals and lipids using the oleaginous yeast Yarrowia lipolytica. Bioresour. Technol. 2020, 313, 123639. [Google Scholar] [CrossRef]
- Cunha, J.T.; Romaní, A.; Costa, C.E.; Sá-Correia, I.; Domingues, L. Molecular and physiological basis of Saccharomyces cerevisiae tolerance to adverse lignocellulose-based process conditions. Appl. Microbiol. Biotechnol. 2018, 103, 159–175. [Google Scholar] [CrossRef] [Green Version]
- Sjulander, N.; Kikas, T. Origin, Impact and Control of Lignocellulosic Inhibitors in Bioethanol Production—A Review. Energies 2020, 13, 4751. [Google Scholar] [CrossRef]
- Sitepu, I.; Selby, T.; Lin, T.; Zhu, S.; Boundy-Mills, K. Carbon source utilization and inhibitor tolerance of 45 oleaginous yeast species. J. Ind. Microbiol. Biotechnol. 2014, 41, 1061–1070. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.R.M.; Bertilsson, M.; Gorwa-Grauslund, M.F.; Gorsich, S.; Lidén, G. Metabolic effects of furaldehydes and impacts on biotechnological processes. Appl. Microbiol. Biotechnol. 2009, 82, 625–638. [Google Scholar] [CrossRef]
- Boyer, L.; Vega, J.; Klasson, K.T.; Clausen, E.; Gaddy, J. The effects of furfural on ethanol production by saccharomyces cereyisiae in batch culture. Biomass Bioenergy 1992, 3, 41–48. [Google Scholar] [CrossRef]
- Konzock, O.; Zaghen, S.; Norbeck, J. Tolerance of Yarrowia lipolytica to inhibitors commonly found in lignocellulosic hydrolysates. BMC Microbiol. 2021, 21, 1–10. [Google Scholar] [CrossRef]
- Kumar, K.K.; Deeba, F.; Sauraj; Negi, Y.S.; Gaur, N.A. Harnessing pongamia shell hydrolysate for triacylglycerol agglomeration by novel oleaginous yeast Rhodotorula pacifica INDKK. Biotechnol. Biofuels 2020, 13, 1–14. [Google Scholar] [CrossRef]
- Nakagame, S.; Shimizu, Y.; Saddler, J.N. The Production of Lipids Using 5-Hydorxymethy Furfural Tolerant Rhodotorula graminis Grown on the Hydrolyzates of Steam Pretreated Softwoods. Sustainability 2020, 12, 755. [Google Scholar] [CrossRef] [Green Version]
- Ruyters, S.; Mukherjee, V.; Verstrepen, K.; Thevelein, J.M.; Willems, K.A.; Lievens, B. Assessing the potential of wild yeasts for bioethanol production. J. Ind. Microbiol. Biotechnol. 2014, 42, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Radecka, D.; Mukherjee, V.; Mateo, R.Q.; Stojiljkovic, M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Looking beyond Saccharomyces: The potential of non-conventional yeast species for desirable traits in bioethanol fermentation. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Lam, F.H.; Turanlı-Yıldız, B.; Liu, D.; Resch, M.G.; Fink, G.R.; Stephanopoulos, G. Engineered yeast tolerance enables efficient production from toxified lignocellulosic feedstocks. Sci. Adv. 2021, 7, eabf7613. [Google Scholar] [CrossRef]
- Pomraning, K.R.; Collett, J.R.; Kim, J.; Panisko, E.A.; Culley, D.E.; Dai, Z.; Deng, S.; Hofstad, B.A.; Butcher, M.G.; Magnuson, J.K. Transcriptomic analysis of the oleaginous yeast Lipomyces starkeyi during lipid accumulation on enzymatically treated corn stover hydrolysate. Biotechnol. Biofuels 2019, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Matsushika, A.; Inoue, H.; Kodaki, T.; Sawayama, S. Ethanol production from xylose in engineered Saccharomyces cerevisiae strains: Current state and perspectives. Appl. Microbiol. Biotechnol. 2009, 84, 37–53. [Google Scholar] [CrossRef]
- Lee, J.W.; Yook, S.; Koh, H.; Rao, C.V.; Jin, Y.-S. Engineering xylose metabolism in yeasts to produce biofuels and chemicals. Curr. Opin. Biotechnol. 2020, 67, 15–25. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Rao, C.V. Microbial conversion of xylose into useful bioproducts. Appl. Microbiol. Biotechnol. 2018, 102, 9015–9036. [Google Scholar] [CrossRef]
- Bommareddy, R.R.; Sabra, W.; Maheshwari, G.; Zeng, A.-P. Metabolic network analysis and experimental study of lipid production in Rhodosporidium toruloides grown on single and mixed substrates. Microb. Cell Fact. 2015, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayikci, Ö.; Nielsen, J. Glucose repression in Saccharomyces cerevisiae. FEMS Yeast Res. 2015, 15, fov068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinuselvi, P.; Kim, M.-K.; Lee, S.-K.; Ghim, C.-M. Rewiring carbon catabolite repression for microbial cell factory. BMB Rep. 2012, 45, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Brandenburg, J.; Blomqvist, J.; Shapaval, V.; Kohler, A.; Sampels, S.; Sandgren, M.; Passoth, V. Oleaginous yeasts respond differently to carbon sources present in lignocellulose hydrolysate. Biotechnol. Biofuels 2021, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tiukova, I.A.; Brandenburg, J.; Blomqvist, J.; Sampels, S.; Mikkelsen, N.; Skaugen, M.; Arntzen, M.; Nielsen, J.; Sandgren, M.; Kerkhoven, E.J. Proteome analysis of xylose metabolism in Rhodotorula toruloides during lipid production. Biotechnol. Biofuels 2019, 12, 137. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.K.; Behera, S.; Arora, R.; Kumar, S.; Sani, R.K. Xylose transport in yeast for lignocellulosic ethanol production: Current status. J. Biosci. Bioeng. 2018, 125, 259–267. [Google Scholar] [CrossRef]
- Kuanyshev, N.; Deewan, A.; Jagtap, S.S.; Liu, J.; Selvam, B.; Chen, L.; Shukla, D.; Rao, C.V.; Jin, Y. Identification and analysis of sugar transporters capable of co-transporting glucose and xylose simultaneously. Biotechnol. J. 2021, 16, 2100238. [Google Scholar] [CrossRef]
- Ryu, S.; Hipp, J.; Trinh, C.T. Activating and Elucidating Metabolism of Complex Sugars in Yarrowia lipolytica. Appl. Environ. Microbiol. 2016, 82, 1334–1345. [Google Scholar] [CrossRef] [Green Version]
- Ryu, S.; Trinh, C.T. Understanding Functional Roles of Native Pentose-Specific Transporters for Activating Dormant Pentose Metabolism in Yarrowia lipolytica. Appl. Environ. Microbiol. 2018, 84, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, A.; Singh, R.; Das, A.K.; Maiti, M.K. Characterization of two sugar transporters responsible for efficient xylose uptake in an oleaginous yeast Candida tropicalis SY005. Arch. Biochem. Biophys. 2020, 695, 108645. [Google Scholar] [CrossRef]
- Pinheiro, M.J.; Bonturi, N.; Belouah, I.; Miranda, E.A.; Lahtvee, P.-J. Xylose Metabolism and the Effect of Oxidative Stress on Lipid and Carotenoid Production in Rhodotorula toruloides: Insights for Future Biorefinery. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Sun, T.; Yu, Y.; Wang, K.; Ledesma-Amaro, R.; Ji, X.-J. Engineering Yarrowia lipolytica to produce fuels and chemicals from xylose: A review. Bioresour. Technol. 2021, 337, 125484. [Google Scholar] [CrossRef]
- Lee, H. Review: The structure and function of yeast xylose (aldose) reductases. Yeast 1998, 14, 977–984. [Google Scholar] [CrossRef]
- Peng, B.; Shen, Y.; Li, X.; Chen, X.; Hou, J.; Bao, X. Improvement of xylose fermentation in respiratory-deficient xylose-fermenting Saccharomyces cerevisiae. Metab. Eng. 2012, 14, 9–18. [Google Scholar] [CrossRef]
- Jin, Y.-S.; Jones, S.; Shi, N.-Q.; Jeffries, T.W. Molecular Cloning of XYL3 (d -Xylulokinase) from Pichia stipitis and Characterization of Its Physiological Function. Appl. Environ. Microbiol. 2002, 68, 1232–1239. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Chen, X.-F.; Yang, X.-Y.; Xiong, L.; Lin, X.-Q.; Yang, J.; Wang, B.; Chen, X.-D. Bioconversion of Corncob Acid Hydrolysate into Microbial Oil by the Oleaginous Yeast Lipomyces starkeyi. Appl. Biochem. Biotechnol. 2013, 172, 2197–2204. [Google Scholar] [CrossRef]
- Freer, S.; Skory, C.; Bothast, R. D-Xylose metabolism in Rhodosporidium toruloides. Biotechnol. Lett. 1997, 19, 1119–1122. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Rao, C.V. Production of d-arabitol from d-xylose by the oleaginous yeast Rhodosporidium toruloides IFO0880. Appl. Microbiol. Biotechnol. 2017, 102, 143–151. [Google Scholar] [CrossRef]
- Whitworth, D.A.; Ratledge, C. Phosphoketolase in Rhodotorula graminis and Other Yeasts. J. Gen. Microbiol. 1977, 102, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.T.; Rafledge, C. Induction of Xylulose-5-Phosphate Phosphoketolase in a Variety of Yeasts Grown on D-Xylose: The Key to Efficient Xylose Metabolism. Arch. Microbiol. 1984, 139, 48–52. [Google Scholar] [CrossRef]
- Calvey, C.H.; Su, Y.-K.; Willis, L.B.; McGee, M.; Jeffries, T.W. Nitrogen limitation, oxygen limitation, and lipid accumulation in Lipomyces starkeyi. Bioresour. Technol. 2016, 200, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, G.M.; Hussain, M.S.; Gambill, L.; Gao, D.; Yaguchi, A.; Blenner, M. Engineering xylose utilization in Yarrowia lipolytica by understanding its cryptic xylose pathway. Biotechnol. Biofuels 2016, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wei, N.; Quarterman, J.; Kim, S.R.; Cate, J.H.; Jin, Y.-S. Enhanced biofuel production through coupled acetic acid and xylose consumption by engineered yeast. Nat. Commun. 2013, 4, 2580. [Google Scholar] [CrossRef] [Green Version]
- Abbott, D.A.; Knijnenburg, T.A.; De Poorter, L.M.; Reinders, M.J.; Pronk, J.; Van Maris, A.J. Generic and specific transcriptional responses to different weak organic acids in anaerobic chemostat cultures of Saccharomyces cerevisiae. FEMS Yeast Res. 2007, 7, 819–833. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Hu, J.; Fan, L.; Chen, Q. RNA-Seq-based transcriptomic and metabolomic analysis reveal stress responses and programmed cell death induced by acetic acid in Saccharomyces cerevisiae. Sci. Rep. 2017, 7, 42659. [Google Scholar] [CrossRef]
- Mira, N.P.; Becker, J.D.; Sá-Correia, I. Genomic Expression Program Involving the Haa1p-Regulon in Saccharomyces cerevisiae Response to Acetic Acid. OMICS A J. Integr. Biol. 2010, 14, 587–601. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, R.A.; Vitorino, M.V.; Godinho, C.P.; Bourbon-Melo, N.; Robalo, T.T.; Fernandes, F.; Rodrigues, M.S.; Sá-Correia, I. Yeast adaptive response to acetic acid stress involves structural alterations and increased stiffness of the cell wall. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Godinho, C.P.; Vitorino, M.V.; Robalo, T.T.; Fernandes, F.; Rodrigues, M.S.; Sá-Correia, I. Crosstalk between Yeast Cell Plasma Membrane Ergosterol Content and Cell Wall Stiffness under Acetic Acid Stress Involving Pdr18. J. Fungi 2022, 8, 103. [Google Scholar] [CrossRef]
- Lindberg, L.; Santos, A.X.; Riezman, H.; Olsson, L.; Bettiga, M. Lipidomic Profiling of Saccharomyces cerevisiae and Zygosaccharomyces bailii Reveals Critical Changes in Lipid Composition in Response to Acetic Acid Stress. PLoS ONE 2013, 8, e73936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godinho, C.; Prata, C.S.; Pinto, S.; Cardoso, C.; Bandarra, N.M.; Fernandes, F.; Sá-Correia, I. Pdr18 is involved in yeast response to acetic acid stress counteracting the decrease of plasma membrane ergosterol content and order. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Godinho, C.P.; Sá-Correia, I. Physiological Genomics of Multistress Resistance in the Yeast Cell Model and Factory: Focus on MDR/MXR Transporters. Yeasts Biotechnol. Hum. Health 2019, 58. [Google Scholar] [CrossRef]
- Sá-Correia, I.; Godinho, C.P. Exploring the biological function of efflux pumps for the development of superior industrial yeasts. Curr. Opin. Biotechnol. 2021, 74, 32–41. [Google Scholar] [CrossRef]
- Olsson, L.; Rugbjerg, P.; Pianale, L.T.; Trivellin, C. Robustness: Linking strain design to viable bioprocesses. Trends Biotechnol. 2022, 1–14. [Google Scholar] [CrossRef]
- Gao, R.; Li, Z.; Zhou, X.; Bao, W.; Cheng, S.; Zheng, L. Enhanced lipid production by Yarrowia lipolytica cultured with synthetic and waste-derived high-content volatile fatty acids under alkaline conditions. Biotechnol. Biofuels 2020, 13, 3–16. [Google Scholar] [CrossRef]
- Gao, R.; Li, Z.; Zhou, X.; Cheng, S.; Zheng, L. Oleaginous yeast Yarrowia lipolytica culture with synthetic and food waste-derived volatile fatty acids for lipid production. Biotechnol. Biofuels 2017, 10, 247. [Google Scholar] [CrossRef]
- Ruan, Z.; Hollinshead, W.; Isaguirre, C.; Tang, Y.J.; Liao, W.; Liu, Y. Effects of inhibitory compounds in lignocellulosic hydrolysates on Mortierella isabellina growth and carbon utilization. Bioresour. Technol. 2015, 183, 18–24. [Google Scholar] [CrossRef]
- Rodrigues, F.; Sousa, M.J.; Ludovico, P.; Santos, H.; Côrte-Real, M.; Leão, C. The Fate of Acetic Acid during Glucose Co-Metabolism by the Spoilage Yeast Zygosaccharomyces bailii. PLoS ONE 2012, 7, e52402. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.-F.; Liu, J.-N.; Lu, L.-J.; Peng, K.-M.; Yang, G.-X.; Liu, J. Culture strategies for lipid production using acetic acid as sole carbon source by Rhodosporidium toruloides. Bioresour. Technol. 2016, 206, 141–149. [Google Scholar] [CrossRef]
- Gong, Z.; Zhou, W.; Shen, H.; Yang, Z.; Wang, G.; Zuo, Z.; Hou, Y.; Zhao, Z.K. Co-fermentation of acetate and sugars facilitating microbial lipid production on acetate-rich biomass hydrolysates. Bioresour. Technol. 2016, 207, 102–108. [Google Scholar] [CrossRef]
- Hu, C.; Zhao, X.; Zhao, J.; Wu, S.; Zhao, Z. Effects of biomass hydrolysis by-products on oleaginous yeast Rhodosporidium toruloides. Bioresour. Technol. 2009, 100, 4843–4847. [Google Scholar] [CrossRef]
- Béligon, V.; Poughon, L.; Christophe, G.; Lebert, A.; Larroche, C.; Fontanille, P. Improvement and modeling of culture parameters to enhance biomass and lipid production by the oleaginous yeast Cryptococcus curvatus grown on acetate. Bioresour. Technol. 2015, 192, 582–591. [Google Scholar] [CrossRef]
- Fontanille, P.; Kumar, V.; Christophe, G.; Nouaille, R.; Larroche, C. Bioconversion of volatile fatty acids into lipids by the oleaginous yeast Yarrowia lipolytica. Bioresour. Technol. 2012, 114, 443–449. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Sakinah, A.M.M.; Zularisam, A.W.; Pandey, A. Glycerol waste to value added products and its potential applications. Syst. Microbiol. Biomanuf. 2021, 1, 378–396. [Google Scholar] [CrossRef]
- Do, D.T.H.; Theron, C.W.; Fickers, P. Organic Wastes as Feedstocks for Non-Conventional Yeast-Based Bioprocesses. Microorganisms 2019, 7, 229. [Google Scholar] [CrossRef] [Green Version]
- Kumar, L.R.; Yellapu, S.K.; Tyagi, R.; Drogui, P. Microbial lipid and biodiesel production from municipal sludge fortified with crude glycerol medium using pH-based fed-batch strategy. J. Environ. Chem. Eng. 2021, 9, 105032. [Google Scholar] [CrossRef]
- Filippousi, R.; Antoniou, D.; Tryfinopoulou, P.; Nisiotou, A.A.; Nychas, G.; Koutinas, A.A.; Papanikolaou, S. Isolation, identification and screening of yeasts towards their ability to assimilate biodiesel-derived crude glycerol: Microbial production of polyols, endopolysaccharides and lipid. J. Appl. Microbiol. 2019, 127, 1080–1100. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Tyagi, R.D.; Drogui, P. Utilization of methanol in crude glycerol to assist lipid production in non-sterilized fermentation from Trichosporon oleaginosus. Bioresour. Technol. 2018, 253, 8–15. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Drogui, P.; Tyagi, R.D. The pH-based fed-batch for lipid production from Trichosporon oleaginosus with crude glycerol. Bioresour. Technol. 2018, 259, 237–243. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Yan, S.; Tyagi, R.D.; Drogui, P. Lipid production from fed-batch fermentation of crude glycerol directed by the kinetic study of batch fermentations. Fuel 2017, 209, 1–9. [Google Scholar] [CrossRef]
- Liu, L.-P.; Hu, Y.; Lou, W.-Y.; Li, N.; Wu, H.; Zong, M.-H. Use of Crude Glycerol as Sole Carbon Source for Microbial Lipid Production by Oleaginous Yeasts. Appl. Biochem. Biotechnol. 2016, 182, 495–510. [Google Scholar] [CrossRef]
- Dobrowolski, A.; Mituła, P.; Rymowicz, W.; Mirończuk, A.M. Efficient conversion of crude glycerol from various industrial wastes into single cell oil by yeast Yarrowia lipolytica. Bioresour. Technol. 2016, 207, 237–243. [Google Scholar] [CrossRef]
- Gao, Z.; Ma, Y.; Wang, Q.; Zhang, M.; Wang, J.; Liu, Y. Effect of crude glycerol impurities on lipid preparation by Rhodosporidium toruloides yeast 32489. Bioresour. Technol. 2016, 218, 373–379. [Google Scholar] [CrossRef]
- Saenge, C.; Cheirsilp, B.; Suksaroge, T.T.; Bourtoom, T. Potential use of oleaginous red yeast Rhodotorula glutinis for the bioconversion of crude glycerol from biodiesel plant to lipids and carotenoids. Process Biochem. 2011, 46, 210–218. [Google Scholar] [CrossRef]
- Leiva-Candia, D.; Tsakona, S.; Kopsahelis, N.; García, I.; Papanikolaou, S.; Dorado, M.; Koutinas, A. Biorefining of by-product streams from sunflower-based biodiesel production plants for integrated synthesis of microbial oil and value-added co-products. Bioresour. Technol. 2015, 190, 57–65. [Google Scholar] [CrossRef]
- Sestric, R.; Munch, G.; Cicek, N.; Sparling, R.; Levin, D.B. Growth and neutral lipid synthesis by Yarrowia lipolytica on various carbon substrates under nutrient-sufficient and nutrient-limited conditions. Bioresour. Technol. 2014, 164, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Jin, G.; Gong, Z.; Shen, H.; Bai, F.; Zhao, Z.K. Recycling biodiesel-derived glycerol by the oleaginous yeast Rhodosporidium toruloides Y4 through the two-stage lipid production process. Biochem. Eng. J. 2014, 91, 86–91. [Google Scholar] [CrossRef]
- Liang, Y.; Cui, Y.; Trushenski, J.; Blackburn, J.W. Converting crude glycerol derived from yellow grease to lipids through yeast fermentation. Bioresour. Technol. 2010, 101, 7581–7586. [Google Scholar] [CrossRef] [PubMed]
- Fabarius, J.T.; Wegat, V.; Roth, A.; Sieber, V. Synthetic Methylotrophy in Yeasts: Towards a Circular Bioeconomy. Trends Biotechnol. 2021, 39, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Cotton, C.A.; Claassens, N.J.; Vaquerizo, S.B.; Bar-Even, A. Renewable methanol and formate as microbial feedstocks. Curr. Opin. Biotechnol. 2019, 62, 168–180. [Google Scholar] [CrossRef]
- Espinosa, M.I.; Gonzalez-Garcia, R.A.; Valgepea, K.; Plan, M.R.; Scott, C.; Pretorius, I.S.; Marcellin, E.; Paulsen, I.T.; Williams, T.C. Adaptive laboratory evolution of native methanol assimilation in Saccharomyces cerevisiae. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Cai, H.; Doi, R.; Shimada, M.; Hayakawa, T.; Nakagawa, T. Metabolic regulation adapting to high methanol environment in the methylotrophic yeast Ogataea methanolica. Microb. Biotechnol. 2021, 14, 1512–1524. [Google Scholar] [CrossRef]
- Wainaina, S.; Lukitawesa; Awasthi, M.K.; Taherzadeh, M. Bioengineering of anaerobic digestion for volatile fatty acids, hydrogen or methane production: A critical review. Bioengineered 2019, 10, 437–458. [Google Scholar] [CrossRef] [Green Version]
- Lukitawesa; Patinvoh, R.J.; Millati, R.; Sárvári-Horváth, I.; Taherzadeh, M.J. Factors influencing volatile fatty acids production from food wastes via anaerobic digestion. Bioengineered 2019, 11, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Kolouchová, I.; Schreiberová, O.; Sigler, K.; Masák, J.; Řezanka, T. Biotransformation of volatile fatty acids by oleaginous and non-oleaginous yeast species. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Llamas, M.; Dourou, M.; González-Fernández, C.; Aggelis, G.; Tomás-Pejó, E. Screening of oleaginous yeasts for lipid production using volatile fatty acids as substrate. Biomass Bioenergy 2020, 138, 105553. [Google Scholar] [CrossRef]
- Llamas, M.; Tomás-Pejó, E.; González-Fernández, C. Volatile fatty acids from organic wastes as novel low-cost carbon source for Yarrowia lipolytica. New Biotechnol. 2020, 56, 123–129. [Google Scholar] [CrossRef]
- Fei, Q.; Chang, H.N.; Shang, L.; Choi, J.-D.; Kim, N.; Kang, J. The effect of volatile fatty acids as a sole carbon source on lipid accumulation by Cryptococcus albidus for biodiesel production. Bioresour. Technol. 2011, 102, 2695–2701. [Google Scholar] [CrossRef]
- Béligon, V.; Noblecourt, A.; Christophe, G.; Lebert, A.; Larroche, C.; Fontanille, P. Proof of concept for biorefinery approach aiming at two bioenergy production compartments, hydrogen and biodiesel, coupled by an external membrane. Biofuels 2016, 9, 163–174. [Google Scholar] [CrossRef]
- Liu, J.; Yuan, M.; Shen, Z.-H.; Peng, K.-M.; Lu, L.-J.; Huang, X.-F. Bioconversion of volatile fatty acids derived from waste activated sludge into lipids by Cryptococcus curvatus. Bioresour. Technol. 2016, 211, 548–555. [Google Scholar] [CrossRef]
- Vajpeyi, S.; Chandran, K. Microbial conversion of synthetic and food waste-derived volatile fatty acids to lipids. Bioresour. Technol. 2015, 188, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Kim, J.Y.; Cho, H.U.; Park, H.R.; Park, J.M. Bioconversion of volatile fatty acids from macroalgae fermentation into microbial lipids by oleaginous yeast. Chem. Eng. J. 2014, 264, 735–743. [Google Scholar] [CrossRef]
- Boonyarit, J.; Polburee, P.; Khaenda, B.; Zhao, Z.K.; Limtong, S. Lipid Production from Sugarcane Top Hydrolysate and Crude Glycerol with Rhodosporidiobolus fluvialis Using a Two-Stage Batch-Cultivation Strategy with Separate Optimization of Each Stage. Microorganisms 2020, 8, 453. [Google Scholar] [CrossRef] [Green Version]
- Rakicka, M.; Lazar, Z.; Dulermo, T.; Fickers, P.; Nicaud, J.M. Lipid production by the oleaginous yeast Yarrowia lipolytica using industrial by-products under different culture conditions. Biotechnol. Biofuels 2015, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Poontawee, R.; Limtong, S. Feeding Strategies of Two-Stage Fed-Batch Cultivation Processes for Microbial Lipid Production from Sugarcane Top Hydrolysate and Crude Glycerol by the Oleaginous Red Yeast Rhodosporidiobolus fluvialis. Microorganisms 2020, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y.; Valéro, J.R. Lipid production from Trichosporon oleaginosus cultivated with pre-treated secondary wastewater sludge. Fuel 2014, 134, 274–282. [Google Scholar] [CrossRef]
- Donzella, S.; Serra, I.; Fumagalli, A.; Pellegrino, L.; Mosconi, G.; Scalzo, R.L.; Compagno, C. Recycling industrial food wastes for lipid production by oleaginous yeasts Rhodosporidiobolus azoricus and Cutaneotrichosporon oleaginosum. Biotechnol. Biofuels Bioprod. 2022, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Thiru, M.; Sankh, S.; Rangaswamy, V. Process for biodiesel production from Cryptococcus curvatus. Bioresour. Technol. 2011, 102, 10436–10440. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.-W.; Yang, Y.-C.; Yu, Y.-H. Using crude glycerol and thin stillage for the production of microbial lipids through the cultivation of Rhodotorula glutinis. J. Biosci. Bioeng. 2012, 114, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zong, M.; Wu, H. Efficient lipid production with Trichosporonfermentans and its use for biodiesel preparation. Bioresour. Technol. 2008, 99, 7881–7885. [Google Scholar] [CrossRef]
- Awad, D.; Bohnen, F.; Mehlmer, N.; Brueck, T. Multi-Factorial-Guided Media Optimization for Enhanced Biomass and Lipid Formation by the Oleaginous Yeast Cutaneotrichosporon oleaginosus. Front. Bioeng. Biotechnol. 2019, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Chopra, J.; Sen, R. Process optimization involving critical evaluation of oxygen transfer, oxygen uptake and nitrogen limitation for enhanced biomass and lipid production by oleaginous yeast for biofuel application. Bioprocess Biosyst. Eng. 2018, 41, 1103–1113. [Google Scholar] [CrossRef]
- Evans, C.T.; Ratledge, C. Effect of Nitrogen Source on Lipid Accumulation in Oleaginous Yeasts. Microbiology 1984, 130, 1693–1704. [Google Scholar] [CrossRef] [Green Version]
- Kitcha, S.; Cheirsilp, B. Enhancing Lipid Production from Crude Glycerol by Newly Isolated Oleaginous Yeasts: Strain Selection, Process Optimization, and Fed-Batch Strategy. BioEnergy Res. 2012, 6, 300–310. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Vallejo, J.; Veiga-Crespo, P.; Villa, T.G. Oily yeasts as oleaginous cell factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part II: Technology and potential applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Probst, K.V.; Schulte, L.R.; Durrett, T.P.; Rezac, M.E.; Vadlani, P.V. Oleaginous yeast: A value-added platform for renewable oils. Crit. Rev. Biotechnol. 2015, 36, 942–955. [Google Scholar] [CrossRef]
- Boulton, C.; Ratledge, C. Cryptococcus terricolus, an oleaginous yeast re-appraised. Appl. Microbiol. Biotechnol. 1984, 20. [Google Scholar] [CrossRef]
- Lopes, H.J.S.; Bonturi, N.; Kerkhoven, E.J.; Miranda, E.A.; Lahtvee, P.-J. C/N ratio and carbon source-dependent lipid production profiling in Rhodotorula toruloides. Appl. Microbiol. Biotechnol. 2020, 104, 2639–2649. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lim, E.Y.; Loh, K.-C.; Dai, Y.; Tong, Y.W. Two-Stage Fermentation of Lipomyces starkeyi for Production of Microbial Lipids and Biodiesel. Microorganisms 2021, 9, 1724. [Google Scholar] [CrossRef] [PubMed]
- Christophe, G.; Deo, J.L.; Kumar, V.; Nouaille, R.; Fontanille, P.; Larroche, C. Production of Oils from Acetic Acid by the Oleaginous Yeast Cryptococcus curvatus. Appl. Biochem. Biotechnol. 2011, 167, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, I.C.S.; Chevalot, I.; Komaitis, M.; Marc, I.; Aggelis, G. Single cell oil production by Yarrowia lipolytica growing on an industrial derivative of animal fat in batch cultures. Appl. Microbiol. Biotechnol. 2002, 58, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Sabra, W.; Bommareddy, R.R.; Maheshwari, G.; Papanikolaou, S.; Zeng, A.-P. Substrates and oxygen dependent citric acid production by Yarrowia lipolytica: Insights through transcriptome and fluxome analyses. Microb. Cell Fact. 2017, 16, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.-W.; Zhang, Z. Effects of dissolved oxygen level on cell growth and total lipid accumulation in the cultivation of Rhodotorula glutinis. J. Biosci. Bioeng. 2011, 112, 71–74. [Google Scholar] [CrossRef]
- Capusoni, C.; Rodighiero, V.; Cucchetti, D.; Galafassi, S.; Bianchi, D.; Franzosi, G.; Compagno, C. Characterization of lipid accumulation and lipidome analysis in the oleaginous yeasts Rhodosporidium azoricum and Trichosporon oleaginosus. Bioresour. Technol. 2017, 238, 281–289. [Google Scholar] [CrossRef]
- Davies, R.; Holdsworth, J.; Reader, S. The effect of low oxygen uptake rate on the fatty acid profile of the oleaginous yeast Apiotrichum curvatum. Appl. Microbiol. Biotechnol. 1990, 33, 569–573. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Tan, T. Lipid and carotenoid production by Rhodotorula glutinis under irradiation/high-temperature and dark/low-temperature cultivation. Bioresour. Technol. 2014, 157, 149–153. [Google Scholar] [CrossRef]
- Tezaki, S.; Iwama, R.; Kobayashi, S.; Shiwa, Y.; Yoshikawa, H.; Ohta, A.; Horiuchi, H.; Fukuda, R. Δ12-fatty acid desaturase is involved in growth at low temperature in yeast Yarrowia lipolytica. Biochem. Biophys. Res. Commun. 2017, 488, 165–170. [Google Scholar] [CrossRef]
- Granger, L.-M.; Perlot, P.; Goma, G.; Pareilleux, A. Effect of various nutrient limitations on fatty acid production by Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 1993, 38, 784–789. [Google Scholar] [CrossRef]
- Juanssilfero, A.B.; Kahar, P.; Amza, R.L.; Miyamoto, N.; Otsuka, H.; Matsumoto, H.; Kihira, C.; Thontowi, A.; Yopi; Ogino, C.; et al. Effect of inoculum size on single-cell oil production from glucose and xylose using oleaginous yeast Lipomyces starkeyi. J. Biosci. Bioeng. 2018, 125, 695–702. [Google Scholar] [CrossRef]
- Amaretti, A.; Raimondi, S.; Sala, M.; Roncaglia, L.; De Lucia, M.; Leonardi, A.; Rossi, M. Single cell oils of the cold-adapted oleaginous yeast Rhodotorula glacialis DBVPG 4785. Microb. Cell Fact. 2010, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Jiru, T.M.; Groenewald, M.; Pohl, C.; Steyn, L.; Kiggundu, N.; Abate, D. Optimization of cultivation conditions for biotechnological production of lipid by Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 for biodiesel preparation. 3 Biotech 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Osman, M.E.; Abdel-Razik, A.B.; Zaki, K.I.; Mamdouh, N.; El-Sayed, H. Isolation, molecular identification of lipid-producing Rhodotorula diobovata: Optimization of lipid accumulation for biodiesel production. J. Genet. Eng. Biotechnol. 2022, 20, 1–15. [Google Scholar] [CrossRef]
- Weiss, S.; Samson, F.; Navarro, D.; Casaregola, S. YeastIP: A database for identification and phylogeny of Saccharomycotinayeasts. FEMS Yeast Res. 2013, 13, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, P.; Oliveira, J.D.S.; Pais, P.; Antunes, M.; Palma, M.; Cavalheiro, M.; Galocha, M.; Godinho, C.; Martins, L.M.; Bourbon, N.; et al. YEASTRACT+: A portal for cross-species comparative genomics of transcription regulation in yeasts. Nucleic Acids Res. 2019, 48, D642–D649. [Google Scholar] [CrossRef]
- Godinho, C.P.; Palma, M.; Oliveira, J.; Mota, M.N.; Antunes, M.; Teixeira, M.C.; Monteiro, P.T.; Sá-Correia, I. The NC Yeastract and CommunityYeastract databases to study gene and genomic transcription regulation in non-conventional yeasts. FEMS Yeast Res. 2021, 21, 1–10. [Google Scholar] [CrossRef]
- Oliveira, J.; Antunes, M.; Godinho, C.P.; Teixeira, M.C.; Sá-Correia, I.; Monteiro, P.T. From a genome assembly to full regulatory network prediction: The case study of Rhodotorula toruloides putative Haa1-regulon. BMC Bioinform. 2021, 22, 1–17. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, Y.; Zhang, J.; Zhao, M.; Tang, M.; Zhou, W.; Gong, Z. A metabolic model of Lipomyces starkeyi for predicting lipogenesis potential from diverse low-cost substrates. Biotechnol. Biofuels 2021, 14, 148. [Google Scholar] [CrossRef]
- Aburatani, S.; Ishiya, K.; Itoh, T.; Hayashi, T.; Taniguchi, T.; Takaku, H. Inference of Regulatory System for TAG Biosynthesis in Lipomyces starkeyi. Bioengineering 2020, 7, 148. [Google Scholar] [CrossRef]
- Tiukova, I.A.; Prigent, S.; Nielsen, J.; Sandgren, M.; Kerkhoven, E.J. Genome-scale model of Rhodotorula toruloides metabolism. Biotechnol. Bioeng. 2019, 116, 3396–3408. [Google Scholar] [CrossRef]
- Pham, N.; Reijnders, M.; Suarez-Diez, M.; Nijsse, B.; Springer, J.; Eggink, G.; Schaap, P.J. Genome-scale metabolic modeling underscores the potential of Cutaneotrichosporon oleaginosus ATCC 20509 as a cell factory for biofuel production. Biotechnol. Biofuels 2021, 14, 1–17. [Google Scholar] [CrossRef]
- Poorinmohammad, N.; Kerkhoven, E.J. Systems-level approaches for understanding and engineering of the oleaginous cell factory Yarrowia lipolytica. Biotechnol. Bioeng. 2021, 118, 3640–3654. [Google Scholar] [CrossRef]
- Chen, Y.; Li, F.; Nielsen, J. Genome-scale modeling of yeast metabolism: Retrospectives and perspectives. FEMS Yeast Res. 2022, 22, 1–9. [Google Scholar] [CrossRef]
- Lu, H.; Kerkhoven, E.J.; Nielsen, J. Multiscale models quantifying yeast physiology: Towards a whole-cell model. Trends Biotechnol. 2021, 40, 291–305. [Google Scholar] [CrossRef]
- Claus, S.; Jezierska, S.; Elbourne, L.D.H.; Van Bogaert, I. Exploring the transportome of the biosurfactant producing yeast Starmerella bombicola. BMC Genom. 2022, 23, 1–17. [Google Scholar] [CrossRef]
- Gajdoš, P.; Ledesma-Amaro, R.; Nicaud, J.-M.; Čertík, M.; Rossignol, T. Overexpression of diacylglycerol acyltransferase in Yarrowia lipolytica affects lipid body size, number and distribution. FEMS Yeast Res. 2016, 16, fow062. [Google Scholar] [CrossRef] [Green Version]
- Quarterman, J.C.; Slininger, P.J.; E Hector, R.; Dien, B.S. Engineering Candida phangngensis—An oleaginous yeast from the Yarrowia clade—For enhanced detoxification of lignocellulose-derived inhibitors and lipid overproduction. FEMS Yeast Res. 2018, 18, 102. [Google Scholar] [CrossRef]
- Polburee, P.; Ohashi, T.; Tsai, Y.-Y.; Sumyai, T.; Lertwattanasakul, N.; Limtong, S.; Fujiyama, K. Molecular cloning and overexpression of DGA1, an acyl-CoA-dependent diacylglycerol acyltransferase, in the oleaginous yeast Rhodosporidiobolus fluvialis DMKU-RK253. Microbiology 2018, 164, 1–10. [Google Scholar] [CrossRef]
- Zhang, S.; Skerker, J.M.; Rutter, C.D.; Maurer, M.J.; Arkin, A.P.; Rao, C.V. Engineering Rhodosporidium toruloides for increased lipid production. Biotechnol. Bioeng. 2015, 113, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, J.; Tsakraklides, V.; Kamineni, A.; Greenhagen, E.H.; Consiglio, A.L.; MacEwen, K.; Crabtree, D.V.; Afshar, J.; Nugent, R.L.; Hamilton, M.A.; et al. Engineering of a high lipid producing Yarrowia lipolytica strain. Biotechnol. Biofuels 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, K.; Abidi, S.H.I.; Liu, H.; Zhang, H.; Chakraborty, S.; Watson, N.; Ajikumar, P.K.; Stephanopoulos, G. Engineering lipid overproduction in the oleaginous yeast Yarrowia lipolytica. Metab. Eng. 2015, 29, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazeck, J.; Hill, A.; Liu, L.; Knight, R.; Miller, J.; Pan, A.; Otoupal, P.; Alper, H.S. Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat. Commun. 2014, 5, 3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Ito, M.; Skerker, J.M.; Arkin, A.; Rao, C.V. Metabolic engineering of the oleaginous yeast Rhodosporidium toruloides IFO0880 for lipid overproduction during high-density fermentation. Appl. Microbiol. Biotechnol. 2016, 100, 9393–9405. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, L.; Chen, H.; Chen, Y.Q.; Ratledge, C.; Song, Y.; Chen, W. Regulatory properties of malic enzyme in the oleaginous yeast, Yarrowia lipolytica, and its non-involvement in lipid accumulation. Biotechnol. Lett. 2013, 35, 2091–2098. [Google Scholar] [CrossRef]
- Tang, W.; Zhang, S.; Tan, H.; Zhao, Z.K. Molecular Cloning and Characterization of a Malic Enzyme Gene from the Oleaginous Yeast Lipomyces starkeyi. Mol. Biotechnol. 2010, 45, 121–128. [Google Scholar] [CrossRef]
- Wasylenko, T.M.; Ahn, W.S.; Stephanopoulos, G. The oxidative pentose phosphate pathway is the primary source of NADPH for lipid overproduction from glucose in Yarrowia lipolytica. Metab. Eng. 2015, 30, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Qiao, K.; Wasylenko, T.M.; Zhou, K.; Xu, P.; Stephanopoulos, G. Lipid production in Yarrowia lipolytica is maximized by engineering cytosolic redox metabolism. Nat. Biotechnol. 2017, 35, 173–177. [Google Scholar] [CrossRef]
- Tai, M.; Stephanopoulos, G. Engineering the push and pull of lipid biosynthesis in oleaginous yeast Yarrowia lipolytica for biofuel production. Metab. Eng. 2012, 15, 1–9. [Google Scholar] [CrossRef]
- Xue, S.-J.; Chi, Z.; Zhang, Y.; Li, Y.-F.; Liu, G.-L.; Jiang, H.; Hu, Z.; Chi, Z.-M. Fatty acids from oleaginous yeasts and yeast-like fungi and their potential applications. Crit. Rev. Biotechnol. 2018, 38, 1049–1060. [Google Scholar] [CrossRef]
- Xu, P.; Qiao, K.; Ahn, W.S.; Stephanopoulos, G. Engineering Yarrowia lipolytica as a platform for synthesis of drop-in transportation fuels and oleochemicals. Proc. Natl. Acad. Sci. USA 2016, 113, 10848–10853. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Sharpe, P.L.; Hong, S.-P.; Yadav, N.S.; Xie, D.; Short, D.R.; Damude, H.G.; A Rupert, R.; E Seip, J.; Wang, J.; et al. Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat. Biotechnol. 2013, 31, 734–740. [Google Scholar] [CrossRef]
- Kretschmer, M.; Klose, J.; Kronstad, J.W. Defects in Mitochondrial and Peroxisomal β-Oxidation Influence Virulence in the Maize Pathogen Ustilago maydis. Eukaryot. Cell 2012, 11, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Camões, F.; Islinger, M.; Guimarães, S.C.; Kilaru, S.; Schuster, M.; Godinho, L.F.; Steinberg, G.; Schrader, M. New insights into the peroxisomal protein inventory: Acyl-CoA oxidases and -dehydrogenases are an ancient feature of peroxisomes. Biochim. Biophys. Acta 2015, 1853, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Freitag, J.; Ast, J.; Bölker, M. Cryptic peroxisomal targeting via alternative splicing and stop codon read-through in fungi. Nature 2012, 485, 522–525. [Google Scholar] [CrossRef]
- Bhutada, G.; Kavšček, M.; Ledesma-Amaro, R.; Thomas, S.; Rechberger, G.N.; Nicaud, J.M.; Natter, K. Sugar versus Fat: Elimination of Glycogen Storage Improves Lipid Accumulation in Yarrowia lipolytica. FEMS Yeast Res. 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Yarrowia lipolytica as a potential producer of citric acid from raw glycerol. J. Appl. Microbiol. 2002, 92, 737–744. [Google Scholar] [CrossRef]
- Wang, Z.-P.; Xu, H.-M.; Wang, G.-Y.; Chi, Z.; Chi, Z.-M. Disruption of the MIG1 gene enhances lipid biosynthesis in the oleaginous yeast Yarrowia lipolytica ACA-DC 50109. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2013, 1831, 675–682. [Google Scholar] [CrossRef]
- Seip, J.; Jackson, R.; He, H.; Zhu, Q.; Hong, S.-P. Snf1 Is a Regulator of Lipid Accumulation in Yarrowia lipolytica. Appl. Environ. Microbiol. 2013, 79, 7360–7370. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Markham, K.; Blazeck, J.; Zhou, N.; Leon, D.; Otoupal, P.; Alper, H.S. Surveying the lipogenesis landscape in Yarrowia lipolytica through understanding the function of a Mga2p regulatory protein mutant. Metab. Eng. 2015, 31, 102–111. [Google Scholar] [CrossRef]
- Tsai, Y.-Y.; Ohashi, T.; Wu, C.-C.; Bataa, D.; Misaki, R.; Limtong, S.; Fujiyama, K. Delta-9 fatty acid desaturase overexpression enhanced lipid production and oleic acid content in Rhodosporidium toruloides for preferable yeast lipid production. J. Biosci. Bioeng. 2019, 127, 430–440. [Google Scholar] [CrossRef]
- Wu, C.-C.; Ohashi, T.; Kajiura, H.; Sato, Y.; Misaki, R.; Honda, K.; Limtong, S.; Fujiyama, K. Functional characterization and overexpression of Δ12-desaturase in the oleaginous yeast Rhodotorula toruloides for production of linoleic acid-rich lipids. J. Biosci. Bioeng. 2021, 131, 631–639. [Google Scholar] [CrossRef]
- Yan, F.X.; Dong, G.R.; Qiang, S.; Niu, Y.J.; Hu, C.Y.; Meng, Y.H. Overexpression of △12, △15-Desaturases for Enhanced Lipids Synthesis in Yarrowia lipolytica. Front. Microbiol. 2020, 11, 289. [Google Scholar] [CrossRef]
- Lazar, Z.; Dulermo, T.; Neuvéglise, C.; Coq, A.-M.C.-L.; Nicaud, J.-M. Hexokinase—A limiting factor in lipid production from fructose in Yarrowia lipolytica. Metab. Eng. 2014, 26, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kang, X.; Xiao, N.; Gao, M.; Zhao, Y.; Zhang, B.; Song, Y. Intracellular expression of Vitreoscilla haemoglobin improves lipid production in Yarrowia lipolytica. Lett. Appl. Microbiol. 2019, 68, 248–257. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, L.; Chen, H.; Chen, Y.Q.; Chen, W.; Song, Y.; Ratledge, C. Enhanced lipid accumulation in the yeast Yarrowia lipolytica by over-expression of ATP: Citrate lyase from Mus musculus. J. Biotechnol. 2014, 192, 78–84. [Google Scholar] [CrossRef]
- Xu, P.; Qiao, K.; Stephanopoulos, G. Engineering oxidative stress defense pathways to build a robust lipid production platform in Yarrowia lipolytica. Biotechnol. Bioeng. 2017, 114, 1521–1530. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Dulermo, T.; Nicaud, J.M. Engineering Yarrowia lipolytica to produce biodiesel from raw starch. Biotechnol. Biofuels 2015, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Sun, W.; Shen, H.; Zhang, S.; Jiao, X.; Zhao, Z.K. Expression of phosphotransacetylase in Rhodosporidium toruloides leading to improved cell growth and lipid production. RSC Adv. 2018, 8, 24673–24678. [Google Scholar] [CrossRef] [Green Version]
- Donzella, S.; Cucchetti, D.; Capusoni, C.; Rizzi, A.; Galafassi, S.; Chiara, G.; Compagno, C. Engineering cytoplasmic acetyl-CoA synthesis decouples lipid production from nitrogen starvation in the oleaginous yeast Rhodosporidium azoricum. Microb. Cell Fact. 2019, 18, 1–10. [Google Scholar] [CrossRef]
- Podolsky, I.A.; Seppälä, S.; Xu, H.; Jin, Y.-S.; O’Malley, M.A. A SWEET surprise: Anaerobic fungal sugar transporters and chimeras enhance sugar uptake in yeast. Metab. Eng. 2021, 66, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Perpelea, A.; Wijaya, A.W.; Martins, L.C.; Rippert, D.; Klein, M.; Angelov, A.; Peltonen, K.; Teleki, A.; Liebl, W.; Richard, P.; et al. Towards valorization of pectin-rich agro-industrial residues: Engineering of Saccharomyces cerevisiae for co-fermentation of d-galacturonic acid and glycerol. Metab. Eng. 2021, 69, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Niehus, X.; Coq, A.-M.C.-L.; Sandoval, G.; Nicaud, J.-M.; Ledesma-Amaro, R. Engineering Yarrowia lipolytica to enhance lipid production from lignocellulosic materials. Biotechnol. Biofuels 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Alper, H.S. Enabling xylose utilization in Yarrowia lipolytica for lipid production. Biotechnol. J. 2016, 11, 1230–1240. [Google Scholar] [CrossRef]
- Yook, S.D.; Kim, J.; Gong, G.; Ko, J.K.; Um, Y.; Han, S.O.; Lee, S. High-yield lipid production from lignocellulosic biomass using engineered xylose-utilizing Yarrowia lipolytica. GCB Bioenergy 2020, 12, 670–679. [Google Scholar] [CrossRef]
- Prabhu, A.; Ledesma-Amaro, R.; Lin, C.S.K.; Coulon, F.; Thakur, V.K.; Kumar, V. Bioproduction of succinic acid from xylose by engineered Yarrowia lipolytica without pH control. Biotechnol. Biofuels 2020, 13, 1–15. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, S.; Gao, X.; Li, M.; Li, D.; Lu, W. Enhanced Protopanaxadiol Production from Xylose by Engineered Yarrowia lipolytica. Microb. Cell Fact. 2019, 18, 1–12. [Google Scholar] [CrossRef]
- Yao, F.; Liu, S.-C.; Wang, D.-N.; Liu, Z.-J.; Hua, Q.; Wei, L.-J. Engineering oleaginous yeast Yarrowia lipolytica for enhanced limonene production from xylose and lignocellulosic hydrolysate. FEMS Yeast Res. 2020, 20, 46. [Google Scholar] [CrossRef]
- Wei, W.; Zhang, P.; Shang, Y.; Zhou, Y.; Ye, B.-C. Metabolically engineering of Yarrowia lipolytica for the biosynthesis of naringenin from a mixture of glucose and xylose. Bioresour. Technol. 2020, 314, 123726. [Google Scholar] [CrossRef]
- Brandt, B.A.; Jansen, T.; Volschenk, H.; Görgens, J.F.; Van Zyl, W.H.; Haan, R.D. Stress modulation as a means to improve yeasts for lignocellulose bioconversion. Appl. Microbiol. Biotechnol. 2021, 105, 4899–4918. [Google Scholar] [CrossRef]
- Deparis, Q.; Claes, A.; Foulquié-Moreno, M.R.; Thevelein, J.M. Engineering tolerance to industrially relevant stress factors in yeast cell factories. FEMS Yeast Res. 2017, 17, 1–17. [Google Scholar] [CrossRef]
- Lin, N.-X.; Xu, Y.; Yu, X.-W. Overview of yeast environmental stress response pathways and the development of tolerant yeasts. Syst. Microbiol. Biomanuf. 2021, 2, 232–245. [Google Scholar] [CrossRef]
- Mohedano, M.T.; Konzock, O.; Chen, Y. Strategies to increase tolerance and robustness of industrial microorganisms. Synth. Syst. Biotechnol. 2021, 7, 533–540. [Google Scholar] [CrossRef]
- Dai, L.; Korolev, K.S.; Gore, J. Relation between stability and resilience determines the performance of early warning signals under different environmental drivers. Proc. Natl. Acad. Sci. USA 2015, 112, 10056–10061. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Son, H.F.; Hwang, S.; Gong, G.; Ko, J.K.; Um, Y.; Han, S.O.; Lee, S.-M. Improving Lipid Production of Yarrowia lipolytica by the Aldehyde Dehydrogenase-Mediated Furfural Detoxification. Int. J. Mol. Sci. 2022, 23, 4761. [Google Scholar] [CrossRef]
- Wu, C.-C.; Tsai, Y.-Y.; Ohashi, T.; Misaki, R.; Limtong, S.; Fujiyama, K. Isolation of a thermotolerant Rhodosporidium toruloides DMKU3-TK16 mutant and its fatty acid profile at high temperature. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Wu, C.-C.; Ohashi, T.; Misaki, R.; Limtong, S.; Fujiyama, K. Ethanol and H2O2 stresses enhance lipid production in an oleaginous Rhodotorula toruloides thermotolerant mutant L1-1. FEMS Yeast Res. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, H.; Chen, X.; Liu, L. Engineering microbial membranes to increase stress tolerance of industrial strains. Metab. Eng. 2018, 53, 24–34. [Google Scholar] [CrossRef]
- Liu, Y.; Koh, C.M.J.; Yap, S.A.; Cai, L.; Ji, L. Understanding and exploiting the fatty acid desaturation system in Rhodotorula toruloides. Biotechnol. Biofuels 2021, 14, 1–17. [Google Scholar] [CrossRef]
- Matsuzawa, T.; Maehara, T.; Kamisaka, Y.; Ara, S.; Takaku, H.; Yaoi, K. Identification and characterization of Δ12 and Δ12/Δ15 bifunctional fatty acid desaturases in the oleaginous yeast Lipomyces starkeyi. Appl. Microbiol. Biotechnol. 2018, 102, 8817–8826. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, S.; Pötter, M.; Sun, W.; Li, L.; Yang, X.; Jiao, X.; Zhao, Z.K. Overexpression of Δ12-Fatty Acid Desaturase in the Oleaginous Yeast Rhodosporidium toruloides for Production of Linoleic Acid-Rich Lipids. Appl. Biochem. Biotechnol. 2016, 180, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Kamisaka, Y.; Kimura, K.; Uemura, H.; Ledesma-Amaro, R. Modulation of gluconeogenesis and lipid production in an engineered oleaginous Saccharomyces cerevisiae transformant. Appl. Microbiol. Biotechnol. 2016, 100, 8147–8157. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, T.; Kamisaka, Y.; Maehara, T.; Takaku, H.; Yaoi, K. Identification and characterization of two fatty acid elongases in Lipomyces starkeyi. Appl. Microbiol. Biotechnol. 2020, 104, 2537–2544. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Nicaud, J. Screening a genomic library for genes involved in propionate tolerance in Yarrowia lipolytica. Yeast 2019, 37, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Yang, H.; Brennan, C.S.; Coldea, T.E.; Zhao, H. Cellular mechanism for the improvement of multiple stress tolerance in brewer’s yeast by potassium ion supplementation. Int. J. Food Sci. Technol. 2019, 55, 2419–2427. [Google Scholar] [CrossRef]
- Masaryk, J.; Sychrová, H. Yeast Trk1 Potassium Transporter Gradually Changes Its Affinity in Response to Both External and Internal Signals. J. Fungi 2022, 8, 432. [Google Scholar] [CrossRef]
- Henriques, S.F.; Mira, N.P.; Sá-Correia, I. Genome-wide search for candidate genes for yeast robustness improvement against formic acid reveals novel susceptibility (Trk1 and positive regulators) and resistance (Haa1-regulon) determinants. Biotechnol. Biofuels 2017, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Cámara, E.; Olsson, L.; Zrimec, J.; Zelezniak, A.; Geijer, C.; Nygård, Y. Data mining of Saccharomyces cerevisiae mutants engineered for increased tolerance towards inhibitors in lignocellulosic hydrolysates. Biotechnol. Adv. 2022, 57, 107947. [Google Scholar] [CrossRef]
- Moreno, A.D.; González-Fernández, C.; Tomás-Pejó, E. Insights into cell robustness against lignocellulosic inhibitors and insoluble solids in bioethanol production processes. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef]
- Stojiljkovic, M.; Claes, A.; Deparis, Q.; Demeke, M.; Subotic, A.; Foulquié-Moreno, M.; Thevelein, J. Whole-Genome Transformation of Yeast Promotes Rare Host Mutations with a Single Causative SNP Enhancing Acetic Acid Tolerance. Mol. Cell. Biol. 2022, 42. [Google Scholar] [CrossRef]
- Chen, Y.; Sheng, J.; Jiang, T.; Stevens, J.; Feng, X.; Na Wei, N. Transcriptional profiling reveals molecular basis and novel genetic targets for improved resistance to multiple fermentation inhibitors in Saccharomyces cerevisiae. Biotechnol. Biofuels 2016, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.; Mira, N.; Vargas, R.; Canelhas, I.; Sá-Correia, I. Saccharomyces cerevisiae adaptation to weak acids involves the transcription factor Haa1p and Haa1p-regulated genes. Biochem. Biophys. Res. Commun. 2005, 337, 95–103. [Google Scholar] [CrossRef]
- Swinnen, S.; Henriques, S.F.; Shrestha, R.; Ho, P.-W.; Sá-Correia, I.; Nevoigt, E. Improvement of yeast tolerance to acetic acid through Haa1 transcription factor engineering: Towards the underlying mechanisms. Microb. Cell Fact. 2017, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.; Yen, H.-W.; Philippidis, G.P. Harnessing the Power of Mutagenesis and Adaptive Laboratory Evolution for High Lipid Production by Oleaginous Microalgae and Yeasts. Sustainability 2020, 12, 5125. [Google Scholar] [CrossRef]
- Sandberg, T.E.; Salazar, M.J.; Weng, L.L.; Palsson, B.O.; Feist, A.M. The emergence of adaptive laboratory evolution as an efficient tool for biological discovery and industrial biotechnology. Metab. Eng. 2019, 56, 1–16. [Google Scholar] [CrossRef]
- Daskalaki, A.; Perdikouli, N.; Aggeli, D.; Aggelis, G. Laboratory evolution strategies for improving lipid accumulation in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2019, 103, 8585–8596. [Google Scholar] [CrossRef]
- Zhou, L.; Xu, Z.; Wen, Z.; Lu, M.; Wang, Z.; Zhang, Y.; Zhou, H.; Jin, M. Combined adaptive evolution and transcriptomic profiles reveal aromatic aldehydes tolerance mechanisms in Yarrowia lipolytica. Bioresour. Technol. 2021, 329, 124910. [Google Scholar] [CrossRef]
- Díaz, T.; Fillet, S.; Campoy, S.; Vázquez, R.; Viña, J.; Murillo, J.; Adrio, J.L. Combining evolutionary and metabolic engineering in Rhodosporidium toruloides for lipid production with non-detoxified wheat straw hydrolysates. Appl. Microbiol. Biotechnol. 2018, 102, 3287–3300. [Google Scholar] [CrossRef]
- Liu, Z.; Radi, M.; Mohamed, E.T.; Feist, A.M.; Dragone, G.; Mussatto, S.I. Adaptive laboratory evolution of Rhodosporidium toruloides to inhibitors derived from lignocellulosic biomass and genetic variations behind evolution. Bioresour. Technol. 2021, 333, 125171. [Google Scholar] [CrossRef]
- Hicks, R.H.; Sze, Y.; Chuck, C.J.; Henk, D.A. Enhanced Inhibitor Tolerance and Increased Lipid Productivity through Adaptive Laboratory Evolution in the Oleaginous Yeast Metshnikowia Pulcherrima. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Banerjee, S.; Pandit, S.; Gupta, P.K.; Mathriya, A.S.; Kumar, S.; Lahiri, D.; Nag, M.; Ray, R.R.; Joshi, S. A comprehensive review on enhanced production of microbial lipids for high-value applications. Biomass Convers. Biorefin. 2021, 1–24. [Google Scholar] [CrossRef]
- Szczepańska, P.; Hapeta, P.; Lazar, Z. Advances in production of high-value lipids by oleaginous yeasts. Crit. Rev. Biotechnol. 2021, 42, 1–22. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Nicaud, J.-M. Yarrowia lipolytica as a biotechnological chassis to produce usual and unusual fatty acids. Prog. Lipid Res. 2016, 61, 40–50. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree of biotechnologically relevant fungi (yeasts and filamentous fungi), highlighting the diversity of oleaginous yeasts. The tree was constructed using the maximum-likelihood method based on the alignment of the small subunit (18S) ribosomal DNA sequence. The sequences used were obtained from NCBI database. The underlined species were described as oleaginous. Ascomycete yeasts are shown in blue, basidiomycetous yeasts are in red and filamentous fungi are in green.
Figure 1.
Phylogenetic tree of biotechnologically relevant fungi (yeasts and filamentous fungi), highlighting the diversity of oleaginous yeasts. The tree was constructed using the maximum-likelihood method based on the alignment of the small subunit (18S) ribosomal DNA sequence. The sequences used were obtained from NCBI database. The underlined species were described as oleaginous. Ascomycete yeasts are shown in blue, basidiomycetous yeasts are in red and filamentous fungi are in green.

Figure 2.
Lipid biosynthesis pathways in oleaginous yeasts. The pathways involved in lipid synthesis (de novo and ex novo synthesis) are summarised in this figure. The explanation of each step of the pathway is described in the main text. Abbreviations: ACC, Acetyl-CoA carboxylase; ACL, ATP-citrate lyase; ADH, alcohol dehydrogenase; 1,3-BPG, 1,3-bisphosphoglycerate; DAG, diacylglycerol; DHAP, dihydroxyacetone phosphate; ELO, Elongase; ER, endoplasmic reticulum; F6P, fructose-6-phosphate; F-1,6-BP, Fructose 1,6-bisphosphate; FAS, fatty acid synthase; FFA, free fatty acids; G6P, glucose-6-phosphate; G3P, glycerol-3-phosphate; GA3P, glyceraldehyde-3-phosphate; LPA, lysophosphatidic acid; OLE, desaturase; 2PG, 2-phosphoglycerate; 3PG, 3-phosphoglycerate; PA, phosphatidic acid; PEP, phosphoenolpyruvate; PPP, pentose phosphate pathway; Pyr, pyruvate; Pyr DH, pyruvate dehydrogenase; TAG, triacylglycerol.
Figure 2.
Lipid biosynthesis pathways in oleaginous yeasts. The pathways involved in lipid synthesis (de novo and ex novo synthesis) are summarised in this figure. The explanation of each step of the pathway is described in the main text. Abbreviations: ACC, Acetyl-CoA carboxylase; ACL, ATP-citrate lyase; ADH, alcohol dehydrogenase; 1,3-BPG, 1,3-bisphosphoglycerate; DAG, diacylglycerol; DHAP, dihydroxyacetone phosphate; ELO, Elongase; ER, endoplasmic reticulum; F6P, fructose-6-phosphate; F-1,6-BP, Fructose 1,6-bisphosphate; FAS, fatty acid synthase; FFA, free fatty acids; G6P, glucose-6-phosphate; G3P, glycerol-3-phosphate; GA3P, glyceraldehyde-3-phosphate; LPA, lysophosphatidic acid; OLE, desaturase; 2PG, 2-phosphoglycerate; 3PG, 3-phosphoglycerate; PA, phosphatidic acid; PEP, phosphoenolpyruvate; PPP, pentose phosphate pathway; Pyr, pyruvate; Pyr DH, pyruvate dehydrogenase; TAG, triacylglycerol.

Figure 3.
Metabolic pathways of xylose assimilation in yeast. The oxidative pentose phosphate pathway is highlighted in dark blue, and the non-oxidative pentose pathway is shown in light blue. Abbreviations: ACC, Acetyl-CoA carboxylase; ACL, ATP-citrate lyase; ADH, alcohol dehydrogenase; ADP, adenosine diphosphate; ATP, adenosine triphosphate;1,3-BPG, 1,3-bisphosphoglycerate; CO2, carbon dioxide; DAG, diacylglycerol; DHAP, dihydroxyacetone phosphate; F6P, fructose-6-phosphate; F-1,6-BP, Fructose 1,6-bisphosphate; FAS, fatty acid synthase; FFA, free fatty acids; G6P, glucose-6-phosphate; G3P, glycerol-3-phosphate; GA3P, glyceraldehyde-3-phosphate; G6PD, glucose 6-phosphate dehydrogenase; 6PGL, 6-phosphogluconolactonase; 6PGD, 6-phosphogluconate dehydrogenase; RPI, ribose-5-phosphate isomerase; R LPA, lysophosphatidic acid; NAD, nicotinamide adenine dinucleotide; 2PG, 2-phosphoglycerate; 3PG, 3-phosphoglycerate; PA, phosphatidic acid; PEP, phosphoenolpyruvate; PK, phosphoketolase; PPP, pentose phosphate pathway; PTA, phosphotransacetylase; Pyr, pyruvate; Pyr DH, pyruvate dehydrogenase; RPE, ribulose 5-phosphate 3-epimerase; RPI, ribose-5-phosphate isomerase; TAG, triacylglycerol; TAL, transaldolase; TCA, tricarboxylic acid cycle; TKL, transketolase; XD, xylonate dehydratase; XDH, xylitol dehydrogenase; XI, xylose isomerase; XK, xylulose kinase; XR, xylose reductase.
Figure 3.
Metabolic pathways of xylose assimilation in yeast. The oxidative pentose phosphate pathway is highlighted in dark blue, and the non-oxidative pentose pathway is shown in light blue. Abbreviations: ACC, Acetyl-CoA carboxylase; ACL, ATP-citrate lyase; ADH, alcohol dehydrogenase; ADP, adenosine diphosphate; ATP, adenosine triphosphate;1,3-BPG, 1,3-bisphosphoglycerate; CO2, carbon dioxide; DAG, diacylglycerol; DHAP, dihydroxyacetone phosphate; F6P, fructose-6-phosphate; F-1,6-BP, Fructose 1,6-bisphosphate; FAS, fatty acid synthase; FFA, free fatty acids; G6P, glucose-6-phosphate; G3P, glycerol-3-phosphate; GA3P, glyceraldehyde-3-phosphate; G6PD, glucose 6-phosphate dehydrogenase; 6PGL, 6-phosphogluconolactonase; 6PGD, 6-phosphogluconate dehydrogenase; RPI, ribose-5-phosphate isomerase; R LPA, lysophosphatidic acid; NAD, nicotinamide adenine dinucleotide; 2PG, 2-phosphoglycerate; 3PG, 3-phosphoglycerate; PA, phosphatidic acid; PEP, phosphoenolpyruvate; PK, phosphoketolase; PPP, pentose phosphate pathway; PTA, phosphotransacetylase; Pyr, pyruvate; Pyr DH, pyruvate dehydrogenase; RPE, ribulose 5-phosphate 3-epimerase; RPI, ribose-5-phosphate isomerase; TAG, triacylglycerol; TAL, transaldolase; TCA, tricarboxylic acid cycle; TKL, transketolase; XD, xylonate dehydratase; XDH, xylitol dehydrogenase; XI, xylose isomerase; XK, xylulose kinase; XR, xylose reductase.

Figure 4.
Metabolic pathways involved in glycerol catabolisation for the production of lipids. Abbreviations: ADP, adenosine diphosphate; ATP, adenosine triphosphate; DAG, diacylglycerol; LPA, lysophosphatidic acid; NAD, nicotinamide adenine dinucleotide; PA, phosphatidic acid: TAG, triacylglycerol.
Figure 4.
Metabolic pathways involved in glycerol catabolisation for the production of lipids. Abbreviations: ADP, adenosine diphosphate; ATP, adenosine triphosphate; DAG, diacylglycerol; LPA, lysophosphatidic acid; NAD, nicotinamide adenine dinucleotide; PA, phosphatidic acid: TAG, triacylglycerol.

Table 1.
Lipid production by various oleaginous yeasts based on hydrolysates of lignocellulosic biomasses. The species contemplated in this analysis were: Cryptococcus aerius, Cryptococcus albidus, Cryptococcus curvatus, Cryptococcus humicola, Lipomyces kononenkoae, Lipomyces starkeyi, Lipomyces tetrasporus, Rhodotorula glutinis, Rhodotorula graminis, Rhodotorula. paludigenum, Rhodotorula. taiwanensis, Rhodotorula toruloides, Rhodosporidiobolus fluvialis, Saitoella coloradoensis, Saitoella complicata, Trichosporon cutaneum, Trichosporon dermatis, Trichosporon guehoae, Trichosporon oleaginosus, and Yarrowia lipolytica.
Table 1.
Lipid production by various oleaginous yeasts based on hydrolysates of lignocellulosic biomasses. The species contemplated in this analysis were: Cryptococcus aerius, Cryptococcus albidus, Cryptococcus curvatus, Cryptococcus humicola, Lipomyces kononenkoae, Lipomyces starkeyi, Lipomyces tetrasporus, Rhodotorula glutinis, Rhodotorula graminis, Rhodotorula. paludigenum, Rhodotorula. taiwanensis, Rhodotorula toruloides, Rhodosporidiobolus fluvialis, Saitoella coloradoensis, Saitoella complicata, Trichosporon cutaneum, Trichosporon dermatis, Trichosporon guehoae, Trichosporon oleaginosus, and Yarrowia lipolytica.
| Species and Strain | Feedstock and Bioprocess Type | Lipid Content (%) Lipid Titer (g/L) Lipid Productivity (g/Lh) | Ref. |
|---|---|---|---|
| C. aerius Y-1399 | Corn stover hydrolysate. Using 96-well plate, 25 °C, 400 rpm, pH 6.0, C/N 62:1, 59.3 g/L Glu, 36.3 g/L Xyl, 5.6 g/L Ara, 0.6 mM HMF, 0.09 mM Fur, 2.4 g/L AcA. | - 9.8 0.089 | [60] |
| C. albidus ATCC 10672 | Sorghum stalk hydrolysate. Flask, 25 °C, 200 rpm, 51% Glu, 30% Xyl, 2.9% Ara. | 42.0 4.6 - | [61] |
| Switchgrass hydrolysate. Flask, 25 °C, 200 rpm, 58% Glu, 26% Xyl. | 44.0 4.7 - | ||
| C. curvatus ATCC 20509 | Corn stover hydrolysate. Bioreactor, 30 °C, pH 5.2, 1 vvm, 25% DO, 97.47 g/L Glu, 58.02 g/L Xyl, 3.51 g/L Gal, 8.56 g/L Ara, 0.24 g/L Fruc. Supplementation with 2.00 g/L YE, 4.00 g/L pep and 1.60 g/L YNB. | 63.1 21.4 0.220 | [62] |
| Wheat straw hydrolysate. Flask, 28 °C, 200 rpm, 3.2 g/L Glu, 14.0 g/L Xyl, 3.7 g/L Ara, 0.8 g/L Gal, 4.2 g/L AcA, 0.03 g/L Fur and 0.02 g/L HMF. | 27.1 4.2 - | [36] | |
| C. humicola UCDFST 10-1004 | Corn stover hydrolysate. Flask, 30 °C, 200 rpm, pH 5.5, 63.2 g/L Glu, 28.9 g/L Xyl. | 40.0 15.5 - | [63] |
| L. kononenkoae Y-7042 | Corn stover hydrolysate. Using 96-well plate, 25 °C, 400 rpm, pH 6.0, C/N 62:1, 59.3 g/L Glu, 36.3 g/L Xyl, 5.6 g/L Ara, 0.6 mM HMF, 0.09 mM Fur, 2.4 g/L AcA. | - 11.3 0.081 | [60] |
| L. starkeyi DSM 70296 | Sugarcane bagasse hydrolysate. Flask, 28 °C, 200rpm, pH 5.5, C/N 50, 13.1 g/L Xyl, 2.2 g/L Glu, 2.1 g/L AcA, 2.2 g/L Ara, 0.02 g/L Fur, 0.02 g/L HMF. | 26.9 - - | [64] |
| Sugarcane bagasse hydrolysate. Bioreactor, 28 °C, 400rpm, pH 5.5, 1 vvm, C/N 50, 13.1 g/L Xyl, 2.2 g/L Glu, 2.1 g/L AcA, 2.2 g/L Ara, 0.02 g/L Fur, 0.02 g/L HMF. | 26.1 - - | ||
| Sugarcane bagasse hydrolysate. Flask, 28 °C, 200 rpm, pH 5.5 <3 g/L Glu, 18.5 g/L Xyl, <3 g/L Ara, 3.6 g/L AcA, Furfural < 300 ppm, HMF < 200 ppm. | 27.8 3.5 0.040 | [65] | |
| L. starkeyi ATCC 56304 | Sorghum stalk hydrolysate. Flask, 25 °C, 200 rpm. 51% Glu, 30% Xyl, 2.9% Ara. | 44.0 7.9 - | [61] |
| L. starkeyi ATCC 56304 | Switchgrass hydrolysate. Flask, 25 °C, 200 rpm, 58% Glu, 26% Xyl. | 39.0 6.5 - | [61] |
| L. starkeyi - | Rice straw hydrolysate. Flask, 30 °C, 160 rpm. | 36.0 4.6 - | [66] |
| L. starkeyi ATCC 12659 | Wheat straw hydrolysate. Flask, 28 °C, 200 rpm, 3.2 g/L Glu, 14.0 g/L Xyl, 3.7 g/L Ara, 0.8 g/L Gal, 4.2 g/L AcA, 0.03 g/L Fur, 0.02 g/L HMF. | 29.1 3.7 - | [36] |
| L. tetrasporus Y-11562 | Corn stover hydrolysate. Using 96-well plate, 25 °C, pH 6.0, 400 rpm, C/N 62:1. 59.3 g/L Glu, 36.3 g/L Xyl, 5.6 g/L Ara, 0.6 mM HMF, 0.09 mM Fur, 2.4 g/L AcA. | - 11.9 0.100 | [60] |
| R. glutinis CGMCC 2.703 | Corn stover hydrolysate. Bioreactor, 30 °C, 2.27 g/L Glu, 40.1 g/L Xyl, 0.171 g/L Fur, 0.483 g/L HMF. | 36.4 5.5 - | [67] |
| R. glutinis ATCC 204091 | Wheat straw hydrolysate. Flask, 28 °C, 200 rpm, 3.2 g/L Glu, 14.0 g/L Xyl, 3.7 g/L Ara, 0.8 g/L Gal, 4.2 g/L AcA, 0.03 g/L Fur and 0.02 g/L HMF. | 20.7 2.4 - | [36] |
| R. graminis DBVPG 4620 | Corn stover hydrolysate. Bioreactor, 30 °C, 900 rpm, pH 6.0. 126 g/L Glu, 6.5 g/L Gal, 87.1 g/L Xyl, 7.5 g/L Ara, 2.9 g/L Man, 4.9 g/L AcA, 0.46 g/L Fur, 1.85 g/L HMF. Supplementation with CSS, YE and salts. | 34.0 - 0.210 | [68] |
| R. paludigenum KM281510 | Corncob hydrolysate. Flask, 25 °C, 200 rpm, pH 6.0, 54.98 g/L Glu, 19.32 g/L Xyl, 1.13 g/L Ara. | 58.4 3.3 - | [58] |
| Rhodotorula sp. KM281508 | 47.4 2.3 - | ||
| R. paludigenum KM281510 | Corncob hydrolysate. Bioreactor, 25 °C, 200 rpm, pH 6.5, 54.98 g/L Glu, 19.32 g/L Xyl, 1.13 g/L Ara. | 73.0 20.3 0.101 | |
| R. taiwanensis AM2353 | Corncob hydrolysate. Bioreactor, 26 °C, 200 rpm, pH 7.0, 1.75 L/min aeration, 7.22 g/L Glu, 36.79 g/L Xyl, 0.02 HMF. Supplementation with 0.5% YE. | 60.3 - - | [69] |
| R. toruloides ATCC 10788 | Wheat straw hydrolysate. Flask, 28 °C, 200 rpm, 3.2 g/L Glu, 14.0 g/L Xyl, 3.7 g/L Ara, 0.8 g/L Gal, 4.2 g/L AcA, 0.03 g/L Fur and 0.02 g/L HMF. | 24.6 2.4 - | [36] |
| R. toruloides CCT 7815 | Hemicellulosic hydrolysate from Birch. Bioreactor, 30 °C, 400–800 rpm, pH 6.0, DO > 25% 87.1 g/L Glu, 29.8 g/L Gal, 21.0 g/L Man, 298.1 g/L Xyl, 14.1 g/L Ara, 20.5 g/L AcA, 33 g/L phenols | 41.0 11.0 - | [70] |
| R. toruloides DSMZ 4444 | Corn stover hydrolysate. Bioreactor, 30 °C, pH 5.2, 1 vvm, 25% DO, 97.47 g/L Glu, 58.02 g/L Xyl, 3.51 g/L Gal, 8.56 g/L Ara, 0.24 g/L Fruc. Supplementation with 2 g/L YE, 4 g/L Pep and 1.6 g/L YNB. | 60.8 23.3 0.170 | [62] |
| R. toruloides Y-1091 | Corn stover hydrolysate. Using 96-well plate, 25 °C, 400 rpm, pH 6.0, C/N 62:1, 59.3 g/L Glu, 36.3 g/L Xyl, 5.6 g/L Ara, 0.6 mM HMF, 0.09 mM Fur, 2.4 g/L AcA. | - 8.8 0.095 | [60] |
| R. toruloides Y4 | Jerusalem artichoke extracts and hydrolysate. Bioreactor, 30 °C, 200–600 rpm, pH 6.0, 40–50% DO. | 56.5 39.6 - | [59] |
| R. fluvialis DMKU-SP314 | Sugar cane top hydrolysate. Flask, 28 °C, 150 rpm, pH 5.5, 21.4 g/L Glu, 7.1 g/L Xyl, 7.1 g/L unidentified sugars. | 43.7 3.4 0.016 | [71] |
| S. coloradoensis YB-2330 | Corn stover hydrolysate. Using 96-well plate, 25 °C, 400 rpm, pH 6.0, C/N 62:1, 59.3 g/L Glu, 36.3 g/L Xyl, 5.6 g/L Ara, 0.6 mM HMF, 0.09 mM Fur, 2.4 g/L AcA. | - 8.5 0.071 | [60] |
| S. complicata Y-17804 | - 7.4 0.067 | ||
| T. cutaneum ATCC 20271 | Corncob hydrolysate. Flask, 30 °C, 180 rpm, pH 5.0, 98.9 g/L Glu, 16.6 g/L Xyl. | 32.1 12.3 - | [72] |
| T. cutaneum CH002 | Corncob hydrolysate. Flask, 28 °C, 150 rpm, pH 7.0, 45.7 g/L sugars, 0.06 g/L Fur, 0.32 g/L HMF, 0.04 g/L AcA, 0.03 g/L ButA. | 36.0 7.9 - | [73] |
| T. cutaneum AS 2.571 | Corn stover hydrolysate. Flask, 30 °C, 200 rpm, pH 6.0, 60 g/L total sugars. | 39.2 7.6 0.078 | [74] |
| T. dermatis 32903 | Corn stover hydrolysate. Flask, 30 °C, 250 rpm, C/N 110, 43.41 g/L Glu, 22.69 g/L Xyl, 3.79 g/L Ara, 1.82 g/L Cel, 2.32 g/L AcA, 1.32 g/L Fur, 2.62 g/L HMF. | 24.2 7.5 0.104 | [75] |
| T. dermatis CH007 | Corn stover hydrolysate. Flask, 28 °C, 150 rpm, pH 7.0, 35.6 g/L Glu, 8 g/L Cel, 16.5 g/L Xyl. | 40.1 9.8 - | [76] |
| T. guehoae UCDFST 6059 | Corn stover hydrolysate. Bioreactor, 30 °C, pH 5.2, 1 vvm, 25% DO, 97.47 g/L Glu, 58.02 g/L Xyl, 3.51 g/L Gal, 8.56 g/L Ara, 0.24 g/L Fruc. Supplementation with 2 g/L YE, 4 g/L Pep and 1.6 g/L YNB. | 48.3 14.2 0.120 | [62] |
| T. oleaginosus ATCC 20509 | Sorghum stalk hydrolysate. Flask, 25 °C, 200 rpm, 51% Glu, 30% Xyl, 2.9% Ara. | 60.0 13.1 - | [61] |
| Switchgrass hydrolysate. Flask, 25 °C, 200 rpm, 58% Glu, 26% Xyl. | 58.0 12.3 - | ||
| Y. lipolytica YB-392 | Corn stover hydrolysate. Using 96-well plate, 25 °C, 400 rpm, pH 6.0, C/N 62:1, 59.3 g/L Glu, 36.3 g/L Xyl, 5.6 g/L Ara, 0.6 mM HMF, 0.09 mM Fur, 2.4 g/L AcA. | - 5.8 0.096 | [60] |
| Y. lipolytica YB-437 | Corn stover hydrolysate. Using 96-well plate, 25 °C, 400 rpm, pH 6.0, C/N 62:1, 59.3 g/L Glu, 36.3 g/L Xyl, 5.6 g/L Ara, 0.6 mM HMF, 0.09 mM Fur, 2.4 g/L AcA. | - 5.8 0.061 | |
| Y. lipolytica Po1g | Sugarcane bagasse hydrolysate. Flask, 26 °C, 160 rpm, pH 6.5, 3.98 g/L Glu, 13.59 g/L Xyl, 2.78 g/L Ara. | 58.5 6.7 0.070 | [77] |
Notes: The taxa displayed on the table refer to the original designation found in the corresponding articles. -, no data available. Lipid content = g of lipids/g of dry weight (%). Abbreviations: Acetic Acid (AcA); Arabinose (Ara); Butyric acid (ButA); Cellobiose (Cel); Corn Steep Solids (CSS); Dissolved oxygen (DO); Glucose (Glu); Fructose (Fruc); Furfural (Fur); Galactose (Gal); Mannose (Man); Peptone (Pep); References (Ref.); Xylose (Xyl); Yeast Extract (YE).
Table 2.
Effect of the supplementation of cultivation media with different concentrations of furfural/HMF in lipid production by Rhodotorula graminis, Lipomyces starkeyi, Rhodotorula glutinis, Rhodotorula toruloides and Trichosporon cutaneum.
Table 2.
Effect of the supplementation of cultivation media with different concentrations of furfural/HMF in lipid production by Rhodotorula graminis, Lipomyces starkeyi, Rhodotorula glutinis, Rhodotorula toruloides and Trichosporon cutaneum.
| Species and Strain | Experimental Condition | Lipid Content (%) Lipid Titer (g/L) Lipid Productivity (g/Lh) | Ref. |
|---|---|---|---|
| R. graminis DBVPG 4620 (adapted strain) | Flask, CSL medium with HMF (0.4%) | - 1.8 - | [96] |
| R. graminis DBVPG 4620 (parental strain) | ND ND ND | ||
| R. graminis DBVPG 4620 | Flask, Medium B, control condition | 43.0 5.52 0.061 | [68] |
| Flask, Medium B + Fur (1.5 g/L) | 28.0 3.03 0.034 | ||
| Flask, Medium B + HMF (1.5 g/L) | 49.0 7.73 0.086 | ||
| L. starkeyi 2.1390 | Flask, N-limited medium, control condition | 37.2 2.29 0.021 | [34] |
| Flask, N-limited medium + 0.5 g/L Fur | 30.3 1.64 - | ||
| Flask, N-limited medium + 0.5 g/L HMF | 31.3 2.22 - | ||
| L. starkeyi 2.1608 | Flask, N-limited medium, control condition | 21.8 2.04 0.097 | |
| Flask, N-limited medium + 0.5 g/L HMF | 23.8 2.00 - | ||
| L. starkeyi 2.1608 | Flask, N-limited medium + 1.0 g/L HMF | 24.6 2.08 - | [34] |
| R. glutinis 2.107 | Flask, N-limited medium, control condition | 13.0 0.52 0.042 | |
| Flask, N-limited medium + 0.5 g/L Fur | 5.51 0.20 - | ||
| Flask, N-limited medium + 0.5 g/L HMF | 11.0 0.56 - | ||
| Flask, N-limited medium + 1.0 g/L HMF | 6.56 0.20 - | ||
| Flask, N-limited medium + 2.0 g/L HMF | 8.19 0.24 - | ||
| R. glutinis 2.704 | Flask, N-limited medium, control condition | 16.7 0.92 0.057 | |
| Flask, N-limited medium + 0.5 g/L HMF | 6.43 0.34 0.023 | ||
| Flask, N-limited medium + 1.0 g/L HMF | 6.22 0.24 - | ||
| Flask, N-limited medium + 2.0 g/L HMF | 4.49 0.14 - | ||
| R. toruloides 2.1389 | Flask, N-limited medium, control condition | 39.3 1.67 0.044 | |
| Flask, N-limited medium + 0.5 g/L HMF | 22.4 0.76 - | ||
| Flask, N-limited medium + 1.0 g/L HMF | 16.5 0.40 - | ||
| Flask, N-limited medium + 2.0 g/L HMF | 14.7 0.28 - | ||
| T. cutaneum 2.1374 | Flask, N-limited medium, control condition | 39.8 1.09 0.011 | |
| Flask, N-limited medium+ 0.5 g/L Fur | 42.5 1.25 - | ||
| Flask, N-limited medium + 1.0 g/L Fur | 30.6 0.54 - | ||
| T. cutaneum 2.1374 | Flask, N-limited medium + 0.5 g/L HMF | 46.8 0.83 - | [34] |
| Flask, N-limited medium + 1.0 g/L HMF | 44.2 1.14 - | ||
| Flask, N-limited medium + 2.0 g/L HMF | 43.8 1.04 - |
Notes: The taxa displayed on the table refer to the original designation found in the corresponding articles. ND, not detected; -, no data available; Lipid content = g of produced lipids/g dry weight (%); Lipid titer = g of produced lipids/L of culture; Lipid productivity = g of produced lipids/L of culture per hour. Abbreviations: Corn steep liquor (CSL); Furfural (Fur); Nitrogen (N); References (Ref.).
Table 3.
Lipid production by oleaginous yeasts (Cryptococcus curvatus, Rhodotorula toruloides, Lipomyces starkeyi, Rhodotorula glutinis, Rhodotorula minuta, Rhodotorula mucilaginosa, Trichosporon cutaneum, Trichosporon fermentans and Yarrowia lipolytica) using acetic acid as carbon source. The experimental conditions column also contains information on the culture method used: flasks, two-stage batch, sequential batch, fed-batch and semicontinuous fermentation.
Table 3.
Lipid production by oleaginous yeasts (Cryptococcus curvatus, Rhodotorula toruloides, Lipomyces starkeyi, Rhodotorula glutinis, Rhodotorula minuta, Rhodotorula mucilaginosa, Trichosporon cutaneum, Trichosporon fermentans and Yarrowia lipolytica) using acetic acid as carbon source. The experimental conditions column also contains information on the culture method used: flasks, two-stage batch, sequential batch, fed-batch and semicontinuous fermentation.
| Species and Strain | Experimental Condition | Lipid Content (%) Lipid Titer (g/L) Lipid Productivity (g/Lh) | Ref. |
|---|---|---|---|
| R. toruloides AS 2.1389 | Flask, 20 g/L AcA, pH 6.0, C/N 200 | 48.2 - 0.025 | [141] |
| Flask, 4 g/L AcA, pH 6.0, C/N 230 | 15.2 - 0.009 | ||
| Flask (two-stage batch), 1st step, 40 g/L Glu; 2nd step, 20 g/L AcA, pH 6.0, C/N 200 | 50.1 - 0.011 | ||
| Flask (two-stage batch), 1st step, 40 g/L Glu; 2nd step, 5 g/L AcA, pH 6.0, C/N 200 | 13.7 - 0.002 | ||
| Flask (Sequencing batch), 4 g/L AcA, pH 6.0, C/N 100 | 38.6 - 0.024 | ||
| C. curvatus ATCC 20509 | Bioreactor (fed-batch), AcA (5 g/L), pH 7.0, C/N 300 | 53.0 25.0 - | [144] |
| Flask, acetate assimilation medium [30 g/L AcA], pH 7.0, C/N 50 | 73.4 4.2 - | [33] | |
| Bioreactor, N-rich acetate medium, containing 5 g/L AcA, pH 7.0, C/N 1.76 | 56.7 0.8 0.030 | ||
| Bioreactor, N-limited acetate medium containing 30 g/L AcA, pH 7.0, C/N 33.5 | 66.4 3.4 0.033 | ||
| L. starkeyi AS 2.1560 | Flask, acetate assimilation medium [30 g/L AcA], pH 7.0, C/N 50 | 17.1 0.6 - | |
| R. glutinis AS 2.107 | Flask, acetate assimilation medium [30 g/L AcA], pH 7.0, C/N 50 | 27.0 0.7 - | [33] |
| R. minuta AS 2.277 | 30.2 0.5 - | ||
| R. mucilaginosa AS 2.1515 | 21.8 0.6 - | ||
| R. toruloides ATCC 10788 | 33.0 0.4 - | ||
| R. toruloides Y4 | 54.9 1.5 - | ||
| T. cutaneum AS 2.571 | 58.5 4.4 - | ||
| T. fermentans CICC 1368 | 55.4 3.8 - | ||
| Y. lipolytica AS 2.1398 | 12.2 0.5 - | ||
| Y. lipolytica MUCL 28849 | Bioreactor (fed-batch), 3x4 g/L AcA, pH 5.6, C/N 50 | 30.8 1.8 - | [145] |
| Bioreactor (two-stage fed-batch), 1st step, 40 g/L Glu; 2nd step, 5 g C/L AcA, pH 5.6, C/N 50 | 40.7 12.4 0.160 | ||
| Bioreactor (two-stage fed-batch), 1st step, 40 g/L Gly; 2nd step, 5g C/L AcA, pH 5.6, C/N 50 | 38.4 15.7 0.330 | ||
| Y. lipolytica MTYL065 | Bioreactor (semicontinuous fermentation), 3% AcA feed, pH 7.0, C/N 32 | 52.6 10.0 0.070 | [87] |
| Bioreactor (semicontinuous fermentation), joint feed of 3% acetic acid and acetate, C/N 13.2 (first 72 h) C/N 102 afterward. | 56.9 33.0 0.230 | ||
| Bioreactor (semicontinuous fermentation), optimised carbon and nitrogen feed. | 59.2 115.0 0.800 | ||
| Bioreactor (semicontinuous fermentation), optimised carbon and nitrogen feed. | 59.2 115.0 0.800 |
Notes: The taxa displayed on the table refer to the original designation found in the corresponding articles. -, no data available. Lipid content: g lipids/g dry weight (%); Lipid concentration: g of produced lipid/L of culture; Lipid yield: g of produced lipids/L of culture per hour. Abbreviations: Acetic acid (AcA); Glucose (Glu); Glycerol (Gly); Nitrogen (N); References (Ref.).
Table 4.
Lipid production by Cryptococcus curvatus, Naganishia uzbekistanensis, Rhodotorula glutinis, Rhodotorula kratochvilovae, Rhodotorula toruloides, Trichosporon fermentans, Trichosporon oleaginosus and Yarrowia lipolytica using crude glycerol as C-source.
Table 4.
Lipid production by Cryptococcus curvatus, Naganishia uzbekistanensis, Rhodotorula glutinis, Rhodotorula kratochvilovae, Rhodotorula toruloides, Trichosporon fermentans, Trichosporon oleaginosus and Yarrowia lipolytica using crude glycerol as C-source.
| Species and Strain | Experimental Condition | Lipid Content (%) Lipid Titer (g/L) Lipid Productivity (g/Lh) | Ref. |
|---|---|---|---|
| Y. lipolytica SKY7 | Bioreactor (fed-batch), 20 g/L suspended solids of washed sludge fortified with crude Gly (10.08 g/L FFA with 5.78 g/L Gly), 28 °C, 400–600 rpm, pH 6.0. | - 31.4 - | [148] |
| Y.lipolytica FMCC Y73 | Flask, 40 g/L Gly (crude Gly purity = 90%), supplemented with 2.0 g/L Pep and 1.0 g/L YE. 29 °C, 190 rev. min−1, pH 6.0. | 16.9 1.2 - | [149] |
| Y.lipolytica FMCC Y74 | 10.7 0.8 - | ||
| Y.lipolytica FMCC Y75 | 19.1 1.3 - | ||
| Rhodotorula sp. FMCC Y78 | 11.3 1.0 - | ||
| Rhodotorula sp. FMCC Y76 | 18.7 1.7 - | ||
| R. glutinis NRRL YB-252 | 19.3 2.1 - | ||
| C. curvatus NRRL Y-1511 | 8.4 1.3 - | ||
| N. uzbekistanensis FMCC Y72 | 34.4 1.1 - | ||
| R. kratochvilovae FMCC Y70 | 19.8 1.7 - | ||
| R. kratochvilovae FMCC Y71 | 16.7 1.5 - | ||
| Debaryomyces sp. FMCC Y68 | 29.9 2.0 - | ||
| Debaryomyces sp. FMCC Y69 | Flask, 40 g/L Gly (crude Gly purity = 90%), supplemented with 2.0 g/L Pep and 1.0 g/L YE. 29 °C, 190 rev. min−1, pH 6.0. | 22.4 1.7 - | [149] |
| Debaryomyces sp. FMCC Y68 | Flask, 80 g/L Gly (crude Gly purity = 90%), supplemented with 2.0 g/L Pep and 1.0 g/L YE. 29 °C, 190 rev. min−1, pH 6.0. | 16.9 2.1 - | |
| Debaryomyces sp. FMCC Y69 | 38.9 2.3 - | ||
| R. glutinis NRRL YB-252 | 38.2 7.2 - | ||
| C. curvatus NRRL Y-1511 | 23.9 4.5 - | ||
| N. uzbekistanensis FMCC Y72 | Flask, 55 g/L Gly (crude Gly purity = 90%), supplemented with 2.0 g/L Pep and 1.0 g/L YE. 29 °C, 150–450 rev. min−1, pH 6.0, 2 vvm. | 31.1 3.3 - | |
| T. oleaginosus ATCC 20905 | Bioreactor (fed-batch), 6.07 g/L Gly (from a crude Gly with a purity of 15.05%), non-sterilised conditions, 30 °C, 300–500 rpm, pH 5.0, DO > 35% (v/v). | 48.1 20.8 - | [150] |
| Bioreactor (fed-batch), 6.22 g/L Gly (from a crude Gly with a purity of 15.05%) + 10.08 g/L FFA 30 °C, pH 5.0, DO > 30% (v/v). | 54.5 35.8 - | [151] | |
| Bioreactor (batch), 10.28 g C/L (crude Gly purity = 13.24%), C/N 20, 30 °C, 300–500 rpm, pH 5.0, DO > 35%. | 23.0 3.1 0.05 | [152] | |
| Bioreactor (batch), 15.30 g C/L (crude Gly purity = 13.24%), C/N 30, 30 °C, 300–500 rpm, pH 5.0, DO > 35%. | 47.5 11.3 0.21 | ||
| Bioreactor (batch), 22.84 g Carbon/L (crude Gly purity = 13.24%) C/N 45, 30 °C, 300–500 rpm, pH 5.0, DO > 35%. | 49.0 12.1 0.22 | ||
| Bioreactor (batch), 29.69 g Carbon/L (crude Gly purity = 13.24%) C/N 60, 30 °C, 300–500 rpm, pH 5.0, DO > 35%. | 52.0 10.0 0.18 | ||
| Bioreactor (fed-batch), ~46.26 g Carbon/L (crude Gly purity = 13.24%) C/N 45, 30 °C, 300–500 rpm, pH 5.0, DO > 35%. | 49.9 21.9 0.42 | ||
| T. cutaneum AS 2.0571 | Flask, 70 g/L crude Gly (75.1% purity) C/N 60, 30 °C, 200 rpm. | 32.2 5.6 - | [153] |
| T. fermentans CICC 1368 | Flask, 50 g/L crude Gly (75.1% purity) C/N 60, 25 °C, 160 rpm. | 32.4 5.2 - | [153] |
| Y. lipolytica A101 | Flask, 50 g/L crude Gly (purity of 80%, from soap production). Supplementation with YNB and (NH4)2SO4. C/N 100, 28 °C, 240 rpm, pH 6.0. | 24.9 1.7 - | [154] |
| Flask, 50 g/L crude Gly (purity of 80%, from biodiesel). Supplementation with YNB and (NH4)2SO4. C/N 100, 28 °C, 240 rpm, pH 6.0. | 24.3 0.9 - | ||
| Flask, 50 g/L crude Gly (purity of 42%, from stearin production). Supplementation with YNB and (NH4)2SO4. C/N 100, 28 °C, 240 rpm, pH 6.0. | 28.0 0.7 - | ||
| R. toruloides 32489 | Flask, crude Gly concentration equivalent to a carbon weight of 20 g/L Glu (crude Gly purity = 49%). Supplementation with 2 g/L Pep, 0.5 g/L (NH4)2SO4, 1 g/L K3PO4 and 0.5 g/L MgSO4. C/N 60, 30 °C, 200 rpm, pH 7.0. | 41.8 6.2 - | [155] |
| R. glutinis TISTR 5159 | Bioreactor (fed-batch), 9.5% crude Gly (50% purity) C/N 85, 30 °C, 100 rpm, pH 6.0, 2 vvm. | 60.7 6.05 - | [156] |
| C. curvatus ATCC 20509 | Bioreactor (fed-batch), 100 g/L Gly (crude Gly purity = 91%) + sunflower meal hydrolysate, 28 °C, 200–700 rpm, pH 6.0, 1vvm. | 47.1 17.9 0.09 | [157] |
| Bioreactor (fed-batch), 100 g/L Gly (crude Gly purity = 91%) + pretreated sunflower meal hydrolysate, 28 °C, 200–700 rpm, pH 6.0, 1vvm. | 52.9 18.3 0.11 | ||
| R. toruloides DSM 4444 | Bioreactor (fed-batch), 50 g/L Gly (crude Gly purity = 91%) + sunflower meal hydrolysate, 28 °C, 200–700 rpm, pH 6.0, 1vvm. | 37.8 18.1 0.14 | |
| Bioreactor (fed-batch), 50 g/L Gly (crude Gly purity = 91%) + pretreated sunflower meal hydrolysate, 28 °C, 200–700 rpm, pH 6.0, 1vvm. | 51.3 19.2 0.17 | ||
| Y. lipolytica ATCC 20460 | Flask, 343 mM Gly (from crude Gly purity between 78–86%) 30 °C, 125 rpm. | 11.6 - - | [158] |
| R. toruloides Y4 | Flask, 20 g/L Gly (from a crude Gly containing 50 g/L Gly) 30 °C, 200 rpm, pH 5.5. | 21.6 2.5 - | [159] |
| Flask, 50 g/L Gly (from a crude Gly containing 50 g/L Gly) 30 °C, 200 rpm, pH 5.5. | 35.4 6.9 - | ||
| Flask, 100 g/L Gly (from a crude Gly containing 50 g/L Gly) 30 °C, 200 rpm, pH 5.5. | 42.5 8.6 - | ||
| Flask, 150 g/L Gly (from a crude Gly containing 50 g/L Gly) 30 °C, 200 rpm, pH 5.5. | 40.4 6.6 - | ||
| Flask, 200 g/L Gly (from a crude Gly containing 50 g/L Gly) 30 °C, 200 rpm, pH 5.5. | 41.5 5.6 - | ||
| C. curvatus ATCC 20509 | Bioreactor (fed-batch), 25.8 g/L glycerol (crude Gly purity = 48.7%). Supplementation with NH4Cl. C/N 30, 28 °C, pH 5.5. | 44.6 13.92 0.05 | [160] |
| C. curvatus ATCC 20509 | Bioreactor (fed-batch), 32 g/L Gly (crude glycerol purity = 48.7%). Supplementation with NH4Cl. C/N 30, 28 °C, pH 5.5. | 52.9 17.40 0.06 | [160] |
Notes: The taxa displayed on the table refer to the original designation found in the corresponding articles. -, no data available; Lipid content: g lipids/g dry weight (%); Lipid concentration: g of produced lipid/L of culture; Lipid yield: g of produced lipids/L of culture per hour. Abbreviations: Glycerol (Gly); References (Ref).
Table 5.
Lipid production by oleaginous yeasts (Cryptococcus albidus, Cryptococcus curvatus, and Yarrowia lipolytica) using a mixture of VFAs as carbon source. The experimental conditions column also contains information on the culture method used: flasks, two-stage batch, sequential batch and fed-batch.
Table 5.
Lipid production by oleaginous yeasts (Cryptococcus albidus, Cryptococcus curvatus, and Yarrowia lipolytica) using a mixture of VFAs as carbon source. The experimental conditions column also contains information on the culture method used: flasks, two-stage batch, sequential batch and fed-batch.
| Species and Strain | Experimental Condition | Lipid Content (%) Lipid Titer (g/L) Lipid Productivity (g/Lh) | Ref. |
|---|---|---|---|
| C. curvatus ATCC 20509 | Bioreactor (fed-batch), VFAs, mainly acetate and butyrate (12 + 4 g/L), pH 7.0, C/N increased to 15 when [VFAs] reached 2 g/L. | 42.0 - - | [171] |
| C. curvatus MUCL 29819 | Flask (Sequencing batch), 3.35 g/L VFAs from activated sludge. | 39.6 - 0.03 | [172] |
| C. albidus ATCC 10672 | Bioreactor, AcA: ProA: ButA (5:1:4), pH 6.0 COD/N ratio 25:1. | 28.3 0.32 - | [173] |
| C. curvatus ATCC 20509 | Bioreactor (repeated batch), VFAs (9.27 g/L), pH 7.0. | 61.0 1.36 - | [174] |
| Y. lipolytica MUCL 28849 | Bioreactor (two-stage fed-batch), 1st step, 40 g/L Glucose; 2nd step, 5g C/L [AcA:ProA:ButA] (3:1:1), pH 5.6, C/N 50. | 40.2 16.50 0.33 | [145] |
| Bioreactor (two-stage fed-batch), 1st step, 40 g/L Glycerol; 2nd step, 5g C/L [AcA:ProA:ButA] (3:1:1), pH 5.6, C/N 50. | 34.6 14.19 0.28 | ||
| C. albidus var. albidus ATCC 10672 | Flask, 2 g/L [AcA:ProA:ButA] (4:3:3), pH 6.0. | 19.8 0.13 - | [170] |
| Flask, 2 g/L [AcA:ProA:ButA] (8:1:1), pH 6.0. | 27.8 0.33 - | ||
| Flask, 2 g/L [AcA:ProA:ButA] 7:2:1, pH 6.0. | 26.1 0.29 - | ||
| Flask, 2 g/L [AcA:ProA:ButA] 6:1:3, pH 6.0. | 27.0 0.31 - | ||
| Flask, 5 g/L [AcA:ProA:ButA] 6:1:3, pH 6.0. | 24.9 0.64 - | ||
| Flask, 8 g/L [AcA:ProA:ButA] 6:1:3, pH 6.0. | 11.9 0.09 - |
Notes: The taxa displayed on the table refer to the original designation found in the corresponding articles. -, no data available; Lipid content: g lipids/g dry weight (%); Lipid concentration: g of produced lipid/L of culture; Lipid yield: g of produced lipids/L of culture per hour. Abbreviations: Acetic acid:propionic acid:butyric acid (AcA:ProA:ButA).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mota, M.N.; Múgica, P.; Sá-Correia, I. Exploring Yeast Diversity to Produce Lipid-Based Biofuels from Agro-Forestry and Industrial Organic Residues. J. Fungi 2022, 8, 687. https://doi.org/10.3390/jof8070687
AMA Style
Mota MN, Múgica P, Sá-Correia I. Exploring Yeast Diversity to Produce Lipid-Based Biofuels from Agro-Forestry and Industrial Organic Residues. Journal of Fungi. 2022; 8(7):687. https://doi.org/10.3390/jof8070687
Chicago/Turabian StyleMota, Marta N., Paula Múgica, and Isabel Sá-Correia. 2022. "Exploring Yeast Diversity to Produce Lipid-Based Biofuels from Agro-Forestry and Industrial Organic Residues" Journal of Fungi 8, no. 7: 687. https://doi.org/10.3390/jof8070687
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.