
Taxonomic and Phylogenetic Reassessment of Pyrgidium (Mycocaliciales) and Investigation of Ascospore Morphology



 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Herbarium Examination and Morphological Studies
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
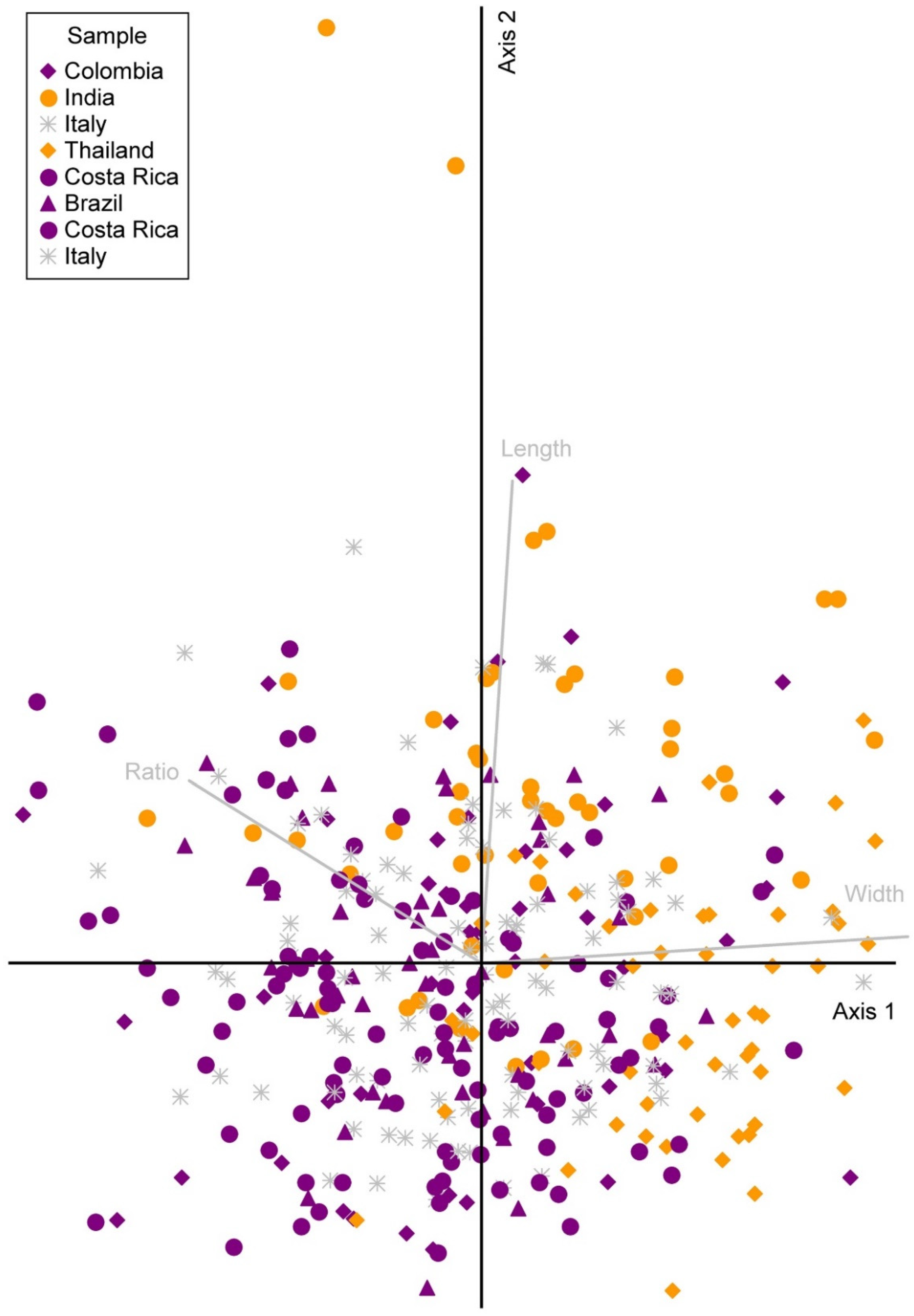
2.4. PCA
3. Results
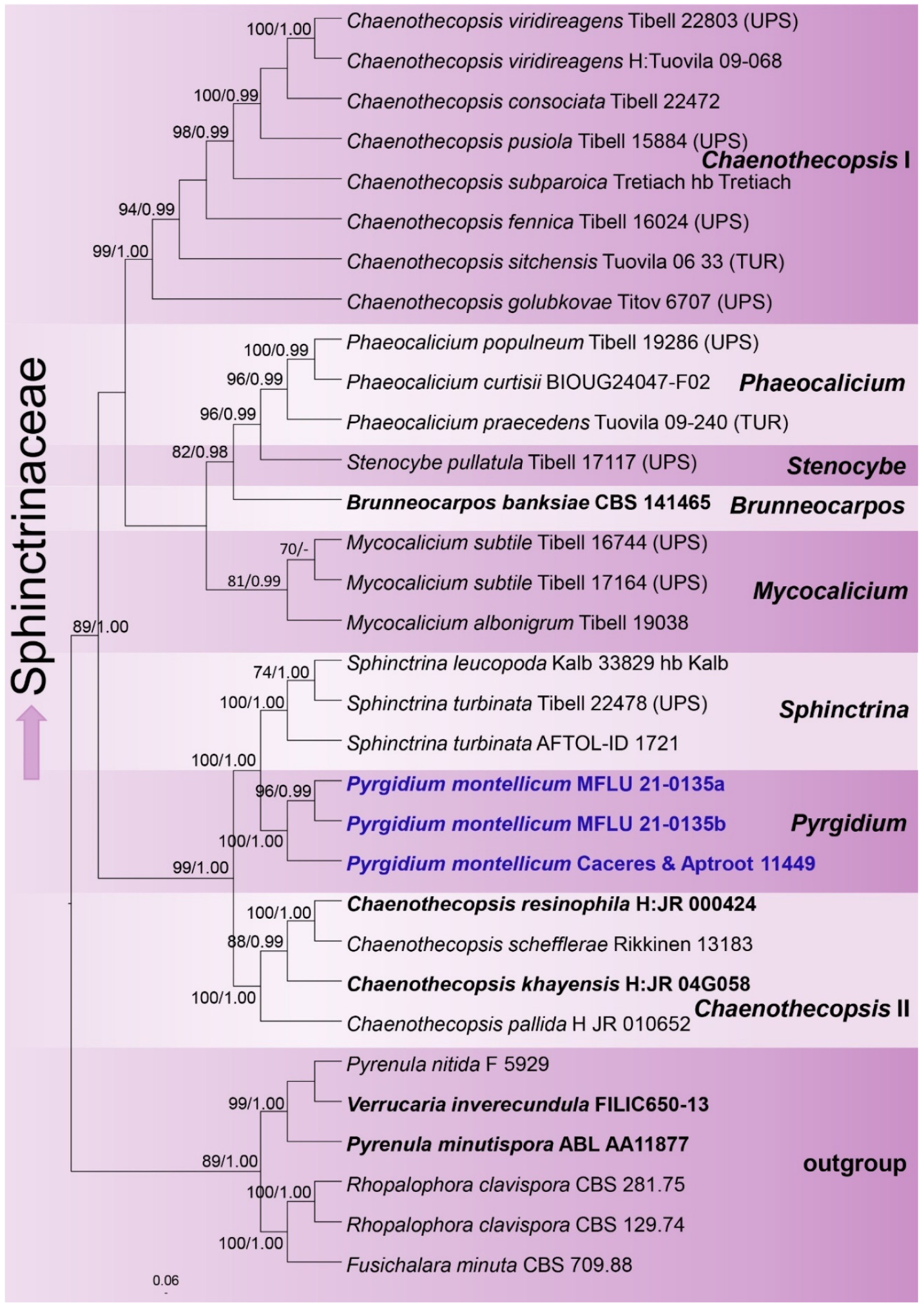
3.1. Phylogenetic Analyses
3.2. PCA
4. Taxonomy
4.1. Sphinctrinaceae M. Choisy, Bull. Mens. Soc. Linn. Soc. Bot. Lyon 19: 65 (1950)
4.2. Pyrgidium Nyl., Flora, Regensburg 50: 3 (1867)
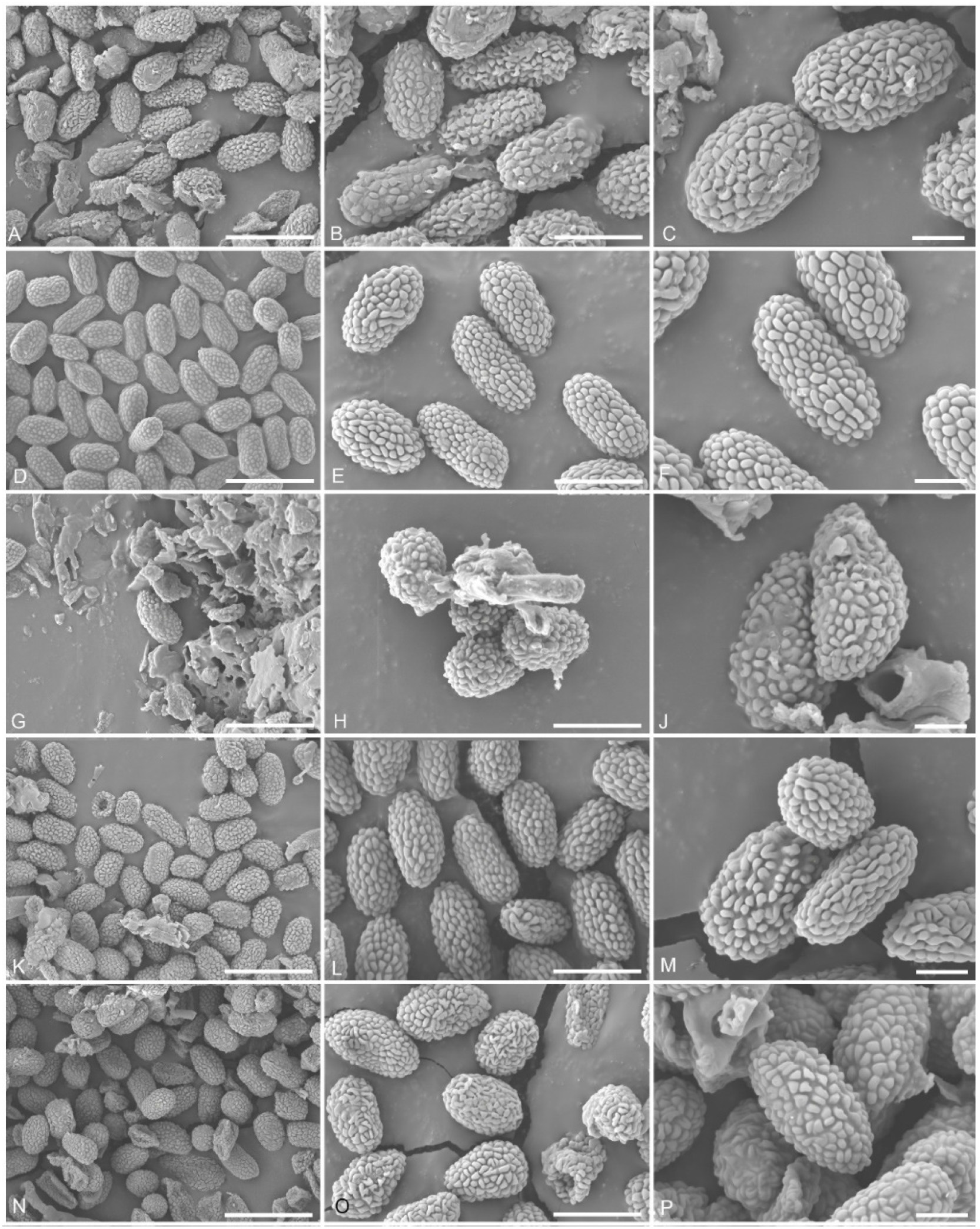
4.3. Pyrgidium montellicum (Beltr.) Tibell, Lichenologist 14(3): 239 (1982) (Figure 3 and Figure 4)
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prieto, M.; Baloch, E.; Tehler, A.; Wedin, M. Mazaedium evolution in the Ascomycota (Fungi) and the classification of mazaediate groups of formerly unclear relationship. Cladistics 2013, 29, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Rikkinen, J.; Meinke, S.K.L.; Grabenhorst, H.; Gröhn, C.; Kobbert, M.; Wunderlich, J.; Schmidt, A.R. Calicioid lichens and fungi in amber–Tracing extant lineages back to the Paleogene. Geobios 2018, 51, 469–479. [Google Scholar] [CrossRef]
- Fries, E. Lichenum dianome nova. Lund 1817, 107, 1403–1418. [Google Scholar]
- Wedin, M.; Tibell, L. Phylogeny and evolution of Caliciaceae, Mycocaliciaceae, and Sphinctrinaceae (Ascomycota), with notes on the evolution of the prototunicate ascus. Canad. J. Bot. 1997, 75, 1236–1242. [Google Scholar] [CrossRef]
- Tibell, L. Caliciales—Flora Neotropica, Monograph 69; New York Botanical Garden: New York, NY, USA, 1996. [Google Scholar]
- Tibell, L.; Wedin, M. Mycocaliciales, a new order for nonlichenized calicioid fungi. Mycologia 2000, 92, 577–581. [Google Scholar] [CrossRef]
- Temu, S.G.; Tibell, S.; Tibuhwa, D.D.; Tibell, L. Crustose Calicioid Lichens and Fungi in Mountain Cloud Forests of Tanzania. Microorganisms 2019, 7, 491. [Google Scholar] [CrossRef]
- Vainio, E.A. Lichenographia fennica III, Coniocarpeae. Acta Soc. Pro Fauna Flora Fenn. 1927, 57, 1–138. [Google Scholar]
- Schmidt, A. Anatomisch-taxonomische Untersu- chungen an Europaischen Arten der Flechtenfamilie Caliciaceae. Mitteilung zur Staatsinstitut fuXr Aligemeine Botanik, Hamburg. Hambg. Staatsinst. Allg. Bot. Mitt. 1970, 13, 111–166. [Google Scholar]
- Tibell, L.B. Distribution patterns and dispersal strategies of Caliciales. Bot. J. Lin. Soc. 1994, 116, 159–202. [Google Scholar] [CrossRef]
- Tibell, L. Phaeocalicium (Mycocaliciaceae, Ascomycetes) in Northern Europe. Annales Botanici Fennici 1996, 33, 205–221. [Google Scholar]
- Tibell, L.B. A reappraisal of the taxonomy of Caliciales. Beih. Nova Hedwig. 1984, 79, 597–713. [Google Scholar]
- Prieto, M.; Wedin, M. Phylogeny, taxonomy and diversification events in the Caliciaceae. Fung Divers. 2017, 82, 221–238. [Google Scholar] [CrossRef]
- Geiser, D.M.; Gueidan, C.; Miadlikowska, J.; Lutzoni, F.; Kauff, F.; Hofstetter, V.; Fraker, E.; Schoch, C.L.; Tibell, L.; Untereiner, W.A.; et al. Eurotiomycetes: Eurotiomycetidae and Chaetothyriomycetidae. Mycologia 2006, 98, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Lumbsch, H.T.; Huhndorf, S.M. Outline of Ascomycota–2007. Myconet 2007, 13, 1–58. [Google Scholar]
- Schmitt, I. 8 Fruiting Body Evolution in the Ascomycota: A Molecular perspective integrating lichenized and non-lichenized groups. In Evolution of Fungi and Fung. Like Organisms; Springer: Berlin, Germany, 2011; pp. 187–204. [Google Scholar]
- Sadowski, E.M.; Beimforde, C.; Gube, M.; Rikkinen, J.; Singh, H.; Seyfullah, L.J.; Heinrichs, J.; Nascimbene, P.C.; Reitner, J.; Schmidt, A.R. The anamorphic genus Monotosporella (Ascomycota) from Eocene amber and from modern Agathis resin. Fung Biol. 2012, 116, 1099–1110. [Google Scholar] [CrossRef]
- Gueidan, C.; Aptroot, A.; da Silva Cáceres, M.E.; Badali, H.; Stenroos, S. A reappraisal of orders and families within the subclass Chaetothyriomycetidae (Eurotiomycetes, Ascomycota). Mycol. Prog. 2014, 13, 1027–1039. [Google Scholar] [CrossRef]
- Geiser, D.M.; LoBuglio, K.F.; Gueidan, C. 5 Pezizomycotina: Eurotiomycetes. Syst. Evol. 2015, 7B, 121–141. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera–Ascomycota. Fung Divers. 2017, 86, 1–594. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Divakar, P.K.; Rajeshkumar, K.C.; Weerahewa, D.; Delgado, G.; Wang, Y.; Fu, L. Notes for genera update—Ascomycota: 6616-6821. Mycosphere 2018, 9, 115–140. [Google Scholar] [CrossRef]
- Wijayawardene, N.; Hyde, K.; Al-Ani, L.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.-L.; Aptroot, A.; Saxena, R.; Tokarev, Y.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1160–1456. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdogdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa-2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Titov, A.; Tibell, L. Chaenothecopsis in the Russian Far East. Nord. J. Bot. 1993, 13, 313–329. [Google Scholar] [CrossRef]
- Tibell, L.; Titov, A. Species of Chaenothecopsis and Mycocalicium (Caliciales) on exudate. Bryologist 1995, 98, 550–560. [Google Scholar] [CrossRef]
- Tuovila, H.; Davey, M.L.; Yan, L.; Huhtinen, S.; Rikkinen, J. New resinicolous Chaenothecopsis species from China. Mycologia 2014, 106, 989–1003. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R.; et al. Fungal diversity notes 111–252—Taxonomic and phylogenetic contributions to fungal taxa. Fung. Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M.; Le Roux, J.J.; Strasberg, D.; Edwards, J.; Roets, F.; Hubka, V.; Taylor, P.W.J.; Heykoop, M.; et al. Fungal Planet description sheets: 400–68. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef]
- Choisy, M. Catalogue des lichens de la region Lyonnaise. Bull. Mens. Soc. Linn. Lyon 1950, 19, 57–72. [Google Scholar]
- Muniz, D.; Llop, E.; Hladun, N.L. Sphinctrina paramerae, a new Mediterranean lichenicolous species with non-septate spores. Lichenologist 2013, 45, 137–143. [Google Scholar] [CrossRef]
- Jaklitsch, W.; Baral, H.-O.; Lücking, R.; Lumbsch, H.T.; Frey, W. Syllabus of Plant Families–Adolf Engler’s Syllabus der Pflanzenfamilien; Borntraeger: Stuttgart, Germany, 2016; p. 288. [Google Scholar]
- Lücking, R.; Hodkinson, B.P.; Leavitt, S.D. Corrections and amendments to the 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota. Bryologist 2017, 120, 58–69. [Google Scholar] [CrossRef]
- Falswal, A.; Bhandari, B.S. A New Lichenicolous Fungus from Garhwal Himalayan Region of Uttarakhand, India. Acta Bot. 2021, 63, 297–302. [Google Scholar] [CrossRef]
- Nylander, W. Addenda nova ad Lichenographiam Europaeam. Continuatio quarta. Flora 1867, 50, 177–180. [Google Scholar]
- Nádvorník, J. Systematische Übersicht der mitteleuropäischen Arten der Flechtenfamilie Caliciaceae. Stud. Bot. Cech. 1942, 5, 6–46. [Google Scholar]
- Tibell, L. Caliciales of Costa Rica. Lichenologist 1982, 14, 219–254. [Google Scholar] [CrossRef]
- Index Fungorum. 2022. Available online: http://www.indexfungorum.org/names/Names.asp (accessed on 15 May 2022).
- Mycobank. 2022. Available online: https://www.mycobank.org/page/Simple%20names%20search (accessed on 15 January 2022).
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fung. Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2652–2676. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Messuti, M.I.; Vidal-Russell, R.; Amico, G.C.; Lorenzo, L.E. Chaenothecopsis quintralis, a new species of calicioid fungus. Mycologia 2012, 104, 1222–1228. [Google Scholar] [CrossRef]
- Schoch, C.L.; Sung, G.H.; López-Giráldez, F.; Townsend, J.P.; Miadlikowska, J.; Hofstetter, V.; Robbertse, B.; Matheny, P.B.; Kauff, F.; Wang, Z.; et al. The Ascomycota tree of life: A phylum-wide phylogeny clarifies the origin and evolution of fundamental reproductive and ecological traits. Syst. Boil. 2009, 58, 224–239. [Google Scholar] [CrossRef]
- Zoller, S.; Scheidegger, C.; Sperisen, C. PCR primers for the amplification of mitochondrial small subunit ribosomal DNA of lichen-forming ascomycetes. Lichenologist 1999, 31, 511–516. [Google Scholar] [CrossRef]
- Swindell, S.R.; Plasterer, T.N. Seqman. In Sequence Data Analysis Guidebook; Springer: Berlin, Germany, 1991; pp. 75–89. [Google Scholar]
- Tibell, L.; Vinuesa, M. Chaenothecopsis in a molecular phylogeny based on nuclear rDNA ITS and LSU sequences. Taxon 2005, 54, 427–442. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinf. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Ac. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. ALTER: Program-oriented conversion of DNA and protein alignments. Nucl. Acids Res. 2010, 38, 14–18. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post–analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the SC10 Workshop on Gateway Computing Environments (GCE10), New Orleans, LA, USA, 5–28 September 2010. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformation 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest 2.0. Program Distributed by the Author; Evolutionary Biology Centre; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A. FigTree. Version 1.4. 2; University of Edinburgh: Edinburgh, UK, 2014. [Google Scholar]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for Fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef]
- Tuovila, H.; Schmidt, A.R.; Beimforde, C.; Dörfelt, H.; Grabenhorst, H.; Rikkinen, J. Stuck in time—A new Chaenothecopsis species with proliferating ascomata from Cunninghamia resin and its fossil ancestors in European amber. Fung. Divers. 2013, 58, 199–213. [Google Scholar] [CrossRef]
- Beimforde, C.; Tuovila, H.; Schmidt, A.R.; Lee, W.G.; Gube, M.; Rikkinen, J. Chaenothecopsis schefflerae (Ascomycota: Mycocaliciales): A widespread fungus on semi-hardened exudates of endemic New Zealand Araliaceae. N. Zeal. J. Bot. 2017, 55, 387–406. [Google Scholar] [CrossRef]
- Réblová, M.; Untereiner, W.A.; Štěpánek, V.; Gams, W. Disentangling Phialophora section Catenulatae: Disposition of taxa with pigmented conidiophores and recognition of a new subclass, Sclerococcomycetidae (Eurotiomycetes). Mycol. Progr. 2017, 16, 27–46. [Google Scholar] [CrossRef]
- Telfer, A.C.; Young, M.R.; Quinn, J.; Perez, K.; Sobel, C.N.; Sones, J.E.; Levesque-Beaudin, V.; Derbyshire, R.; Fernandez-Triana, J.; Rougerie, R.; et al. Biodiversity inventories in high gear: DNA barcoding facilitates a rapid biotic survey of a temperate nature reserve. Biodivers. Data J. 2015, 3, e6313. [Google Scholar] [CrossRef]
- Gueidan, C.; Aptroot, A.; da Silva Cáceres, M.E.; Binh, N.Q. Molecular phylogeny of the tropical lichen family Pyrenulaceae: Contribution from dried herbarium specimens and FTA card samples. Mycol. Prog. 2016, 15, 7. [Google Scholar] [CrossRef]
- Del Prado, R.; Schmitt, I.; Kautz, S.; Palice, Z.; Lücking, R.; Lumbsch, H.T. Molecular data place Trypetheliaceae in Dothideomycetes. Mycol. Res. 2006, 110, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, G.; Wolseley, P.A.; Arachchige, O.; da Silva Cáceres, M.E.; Jayalal, U.; Aptroot, A. Eight new lichen species and 88 new records from Sri Lanka. Bryologist 2016, 119, 131–142. [Google Scholar] [CrossRef]
- Pykälä, J.; Launis, A.; Myllys, L. Taxonomy of the Verrucaria kalenskyi-V. xyloxena species complex in Finland. Nova Hedw. 2019, 109, 489–511. [Google Scholar] [CrossRef]
- Ekanayaka, A.H.; Ariyawansa, H.A.; Hyde, K.D.; Jones, E.B.G.; Daranagama, D.A.; Phillips, A.J.L.; Hongsanan, S.; Jayasiri, S.C.; Zhao, Q. Discomycetes: The apothecial representatives of the phylum Ascomycota. Fung. Divers. 2017, 87, 237–298. [Google Scholar] [CrossRef]
- Titov, A. Further notes on calicioid lichens and fungi from the Gongga Mountains (Sichuan, China). Lichenologist 2001, 33, 303–314. [Google Scholar] [CrossRef]
- Nayaka, S.; Upreti, D.K. Status of lichen diversity in Western Ghats, India. Sahyadri E-News, Western Ghats BioDivers. Infor. Sys. 2005, 16, 1–28. [Google Scholar]
- Aptroot, A.; Saipunkaew, W.; Sipman, H.J.M.; Sparrius, L.B.; Wolseley, P.A. New lichens from Thailand, mainly microlichens from Chiang Mai. Fung. Divers. 2007, 24, 75–134. [Google Scholar]
- Seaward, M.R.D.; Sipman, H.J.M.; Sohrabi, M. A revised checklist of lichenized, lichenicolous and allied fungi for Iran. Sauteria 2008, 15, 459–520. [Google Scholar]
- Nascimbene, J.; Marini, L. Oak forest exploitation and black-locust invasion caused severe shifts in epiphytic lichen communities in Northern Italy. Sci. Tot. Env. 2010, 408, 5506–5512. [Google Scholar] [CrossRef] [PubMed]
- da Silva Cáceres, M.E.; de Lima, E.L.; Nascimento, A.A.; Lücking, R. Liquens brasileiros: Novas descobertas evidenciam a riqueza no Norte e Nordeste do país. Bol. Mus. Biol. Mello Leitão. 2014, 35, 101–119. [Google Scholar]
- Crespo, A.; Lumbsch, H.T. Cryptic species in lichen-forming fungi. IMA Fungus 2010, 1, 167–170. [Google Scholar] [CrossRef]
- Sato, H.; Ohta, R.; Murakami, N. Molecular prospecting for cryptic species of the Hypholoma fasciculare complex: Toward the effective and practical delimitation of cryptic macrofungal species. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Vinuesa, M.D.L.A.; Sanches-Puelles, J.M.; Tibell, L. Intraspecific variation in Mycocalicium subtile (Mycocaliciaceae) elucidated by morphology and the sequences of the ITS1-5.8 S-ITS2 region of rDNA. Mycol. Res. 2001, 105, 323–330. [Google Scholar]
- Nilsson, R.H.; Kristiansson, E.; Ryberg, M.; Hallenberg, N.; Larsson, K.H. Intraspecific ITS variability in the kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol. Bioinfor. 2008, 4, 193–201. [Google Scholar] [CrossRef]
- Ellis, J.B.; Everhart, B.M. New species of fungi from various localities. J. Myc. 1888, 4, 62–65. [Google Scholar] [CrossRef]
- Barr, M.E. Redisposition of some taxa described by J. B. Ellis. Mycotaxon 1993, 46, 45–76. [Google Scholar]
- Jaklitsch, W.M.; Gardiennet, A.; Voglmayr, H. Resolution of morphology-based taxonomic delusions: Acrocordiella, Basiseptospora, Blogiascospora, Clypeosphaeria, Hymenopleella, Lepteutypa, Pseudapiospora, Requienella, Seiridium and Strickeria. Pers. Mol. Phyl. Evol. Fungi 2016, 37, 82. [Google Scholar] [CrossRef]
- Voglmayr, H.; Aguirre-Hudson, M.B.; Wagner, H.G.; Tello, S.; Jaklitsch, W.M. Lichens or endophytes? The enigmatic genus Leptosillia in the Leptosilliaceae fam. nov. (Xylariales), and Furfurella gen. nov. (Delonicicolaceae). Pers. Mol. Phyl. Evol. Fungi 2019, 42, 228. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fung. Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Lücking, R.; Ertz, D.; Wanasinghe, D.N.; Karunarathna, S.C.; Camporesi, E.; Hyde, K.D. Evolution of non-lichenized, saprotrophic species of Arthonia (Ascomycota, Arthoniales) and resurrection of Naevia, with notes on Mycoporum. Fung. Divers. 2020, 102, 205–224. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Lücking, R.; Ertz, D.; Karunarathna, S.C.; Wanasinghe, D.N.; Lumyong, S.; Hyde, K.D. The evolution of life modes in Stictidaceae, with three novel taxa. J. Fung. 2021, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Kossowska, M.; Faltynowicz, W.; Dimos-Zych, M.; Faltynowicz, H.; Patejuk, K.; Piegdon, A.; Buksakowska, M.; Jarema, P. Additions to the lichen biota of the Sudety Mountains. I. Records from the Karkonosze Mountains. Acta Mycologica 2018, 53, 1–10. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The variety of fungal-algal symbioses, their evolutionary significance, and the nature of lichens. Bot. J. Lin. Soc. 1988, 96, 3–20. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Volkmann-Kohlmeyer, B. A new marine Xylomyces on Rhizophora from the Caribbean and Hawaii. Fung. Divers. 1998, 1, 159–164. [Google Scholar]
- Wirth, V. Die Flechten Baden-Württembergs. Teil 1; Ulmer: Stuttgart, Germany, 1995. [Google Scholar]
- Groner, U. The genus Chaenothecopsis (Mycocaliciaceae) in Switzerland, and a key to the European species. Lichenol. 2006, 38, 395–406. [Google Scholar] [CrossRef]
- Nimis, P.L.; Puntillo, D. Keys to the Lichens of Italy-08) Calicioid Species (Both Lichenized and Non-Lichenized). Available online: https://italic.units.it/flora/index.php?procedure=ext_key_home&key_id=1619 (accessed on 24 November 2021).
- Tuovila, H.; Huhtinen, S. New methods for mycocalicioid fungi. Lichenologist 2020, 52, 403–413. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Region | Primers | PCR Condition | References |
|---|---|---|---|
| ITS | ITS4 and ITS5 | 95 °C: 4 min, (94 °C: 1 min, 54 °C: 1 min, 72 °C: 45 s) × 35 cycles 72 °C: 5 min | [7,42] |
| LSU | LROR and LR5 | 94 °C: 5 min, (94 °C: 40 s, 52 °C: 40 s, 72 °C: 40 s) × 35 cycles 72 °C: 10 min | [43,44] |
| SSU | NS1 and NS4 | 95 °C: 15 min, (95 °C: 27 s, 54–56 °C: 30 s, 72 °C: 1 min) × 35 cycles 72 °C: 5 min | [42,45] |
| mtSSU | mtSSU1 and mtSSU3R | 94 °C: 3 min, (94 °C: 3 min, 52 °C: 1 min, 72 °C: 1 min) × 35 cycles 72 °C: 10 min | [46] |
| Taxa | Strain | GenBank Accessions | ||
|---|---|---|---|---|
| LSU | ITS | References | ||
| Brunneocarpos banksiae | CBS 141465 | NG_066277 | - | [29] |
| Chaenothecopsis consociata | Tibell 22472 | DQ008999 | AY795851 | [48] |
| Chaenothecopsis khayensis | H:JR 04G058 | - | NR_120165 | [57] |
| Chaenothecopsis resinophila | H:JR 000424 | JX122782 | JX122780 | [58] |
| Chaenothecopsis schefflerae | Rikkinen 13183 | KY499967 | KY499965 | [59] |
| Chaenothecopsis subparoica | Tretiach (hb. Tretiach) | - | AY795869 | [48] |
| Chaenothecopsis viridireagens | H:Tuovila 09-068 | JX119117 | JX119108 | [58] |
| Chaenothecopsis pallida | H:JR 010652 | JX122781 | JX122779 | [58] |
| Chaenothecopsis pusiola | Tibell 15884 (UPS) | - | AY795865 | [48] |
| Chaenothecopsis fennica | Tibell 16024 (UPS) | AY795995 | AY795857 | [48] |
| Chaenothecopsis sitchensis | Tuovila 06-33 (TUR) | KF157988 | - | [59] |
| Chaenothecopsis golubkovae | Titov 6707 (UPS) | AY795996 | AY795859 | [48] |
| Chaenothecopsis viridireagens | Tibell 22803 (UPS) | DQ013257 | AY795872 | [48] |
| Fusichalara minuta | CBS 709.88 | KX537758 | KX537754 | [60] |
| Mycocalicium subtile | Tibell 16744 (UPS) | AY796004 | - | [48] |
| Mycocalicium subtile | Tibell 17164 (UPS) | AY796005 | - | [48] |
| Mycocalicium albonigrum | Tibell 19038 | AY796001 | AF223966 | [48] |
| Phaeocalicium curtisii | BIOUG24047-F02 | - | KT695401 | [61] |
| Phaeocalicium populneum | Tibell 19286 (UPS) | AY796009 | AY795874 | [48] |
| Phaeocalicium praecedens | Tuovila 09-240 (TUR) | KC590486 | KC590481 | [27] |
| Pyrenula minutispora | ABL AA11877 | - | KT820119 | [62] |
| Pyrenula nitida | F 5929 | DQ329023 | JQ927458 | [63,64] |
| Pyrgidium montellicum | MFLU 21-0135a | ON979678 | ON979674 | This study |
| Pyrgidium montellicum | MFLU 21-0135b | - | OP094605 | This study |
| Pyrgidium montellicum | Cáceres and Aptroot 11449 | OP077215 | ON979667 | This study |
| Rhopalophora clavispora | CBS 129.74 | MH872573 | KX537751 | [60] |
| Rhopalophora clavispora | CBS 281.75 | KX537756 | KX537752 | [50] |
| Sphinctrina leucopoda | Kalb 33829 (hb. Kalb) | AY796006 | AY795875 | [48] |
| Sphinctrina turbinata | AFTOL-ID 1721 | EF413632 | - | [14] |
| Sphinctrina turbinata | Tibell 22478 (UPS) | - | AY795876 | [14] |
| Stenocybe pullatula | Tibell 17117 (UPS) | AY796008 | AY795878 | [48] |
| Verrucaria inverecundula | FILIC650-13 | - | MK138796 | [65] |
| P.bengaliense (Kurz 1866; Isotype UPS) | Trachylia leptoconia (Lindig 2865; Isotype UPS) | Acolium montellicum (L-008798; Syntype UPS) | Acolium montellicum (L-996762) | P. montellicum (Tibell 8306; Non-Type UPS) | P. montellicm Tibel (8232; Non-Type UPS) | P. montellicum (Cáceres and Aptroot 11449) | P. montellicum (MFLU 21-0135) | |
|---|---|---|---|---|---|---|---|---|
| Thallus | Absent | Farinose | Farinose | Farinose | Farinose | Absent | Absent | Farinose |
| Ascomata width (μm) | 190–205 | 220–240 | 215–225 | 185–190 | 180–195 | 190–205 | 225–235 | 180–330 |
| Ascomata height (μm) | 180–220 | 220–235 | 160–170 | 175–190 | 210–290 | 170–190 | 130–145 | 135–195 |
| Exciple (lateral) (μm) | 30–55 | 22–28 | 26–31 | 25–40 | 11–30 | 30–39 | 30–45 | 25–55 |
| Exciple (base) (μm) | 60–90 | 35–55 | 33–46 | 28–40 | 27–38 | 27–42 | 34–46 | 30–60 |
| Paraphyses width (μm) | 0.9–2 | 1–2 | 0.8–1.6 | 1–2 | 0.7–1.7 | 0.4–1.6 | – | 1–2 |
| Asci length (μm) | 18–33 | 34–56 | 31–36 | 29–39 | 28–32 | 25–40 | – | 25–40 |
| Asci width (μm) | 4–9 | 4–5 | 3.5–5 | 3–6 | 3–4 | 2–5 | – | 5–7 |
| Ascospore length (μm) | 5.5–10 | 4.5–9 | 4–5.5 | 5–7 | 4.5–7 | 4.5–7 | 5–8 | 6–9 |
| Ascospore width (μm) | 3–5 | 2–5 | 2.6–3.8 | 2–4 | 2.5–4 | 2–4 | 2.7–4 | 2.5–4.5 |
| Q value | 1.94 | 1.82 | 1.76 | 1.85 | 1.73 | 1.98 | 1.81 | 1.55 |
| No. of septates per ascospore | 0–1 | 0–1 | 0–1 | 0–1 | 0–1 | 0–1 | 0–1 | 0–1 |
| Geographical occurrence | India | Colombia | Italy | Italy | Costa Rica | Costa Rica | Brazil | Thailand |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thiyagaraja, V.; Ertz, D.; Lücking, R.; Wanasinghe, D.N.; Aptroot, A.; Cáceres, M.E.d.S.; Hyde, K.D.; Tapingkae, W.; Cheewangkoon, R. Taxonomic and Phylogenetic Reassessment of Pyrgidium (Mycocaliciales) and Investigation of Ascospore Morphology. J. Fungi 2022, 8, 966. https://doi.org/10.3390/jof8090966
Thiyagaraja V, Ertz D, Lücking R, Wanasinghe DN, Aptroot A, Cáceres MEdS, Hyde KD, Tapingkae W, Cheewangkoon R. Taxonomic and Phylogenetic Reassessment of Pyrgidium (Mycocaliciales) and Investigation of Ascospore Morphology. Journal of Fungi. 2022; 8(9):966. https://doi.org/10.3390/jof8090966
Chicago/Turabian StyleThiyagaraja, Vinodhini, Damien Ertz, Robert Lücking, Dhanushka N. Wanasinghe, André Aptroot, Marcela Eugenia da Silva Cáceres, Kevin D. Hyde, Wanaporn Tapingkae, and Ratchadawan Cheewangkoon. 2022. "Taxonomic and Phylogenetic Reassessment of Pyrgidium (Mycocaliciales) and Investigation of Ascospore Morphology" Journal of Fungi 8, no. 9: 966. https://doi.org/10.3390/jof8090966