
Morphological and Phylogenetic Characterization of Three Novel Thaxterogaster (Cortinariaceae) Species from China with an Emphasis on Their Subtropical Distribution
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens and Morphological Description
2.2. Molecular Phylogeny
3. Results
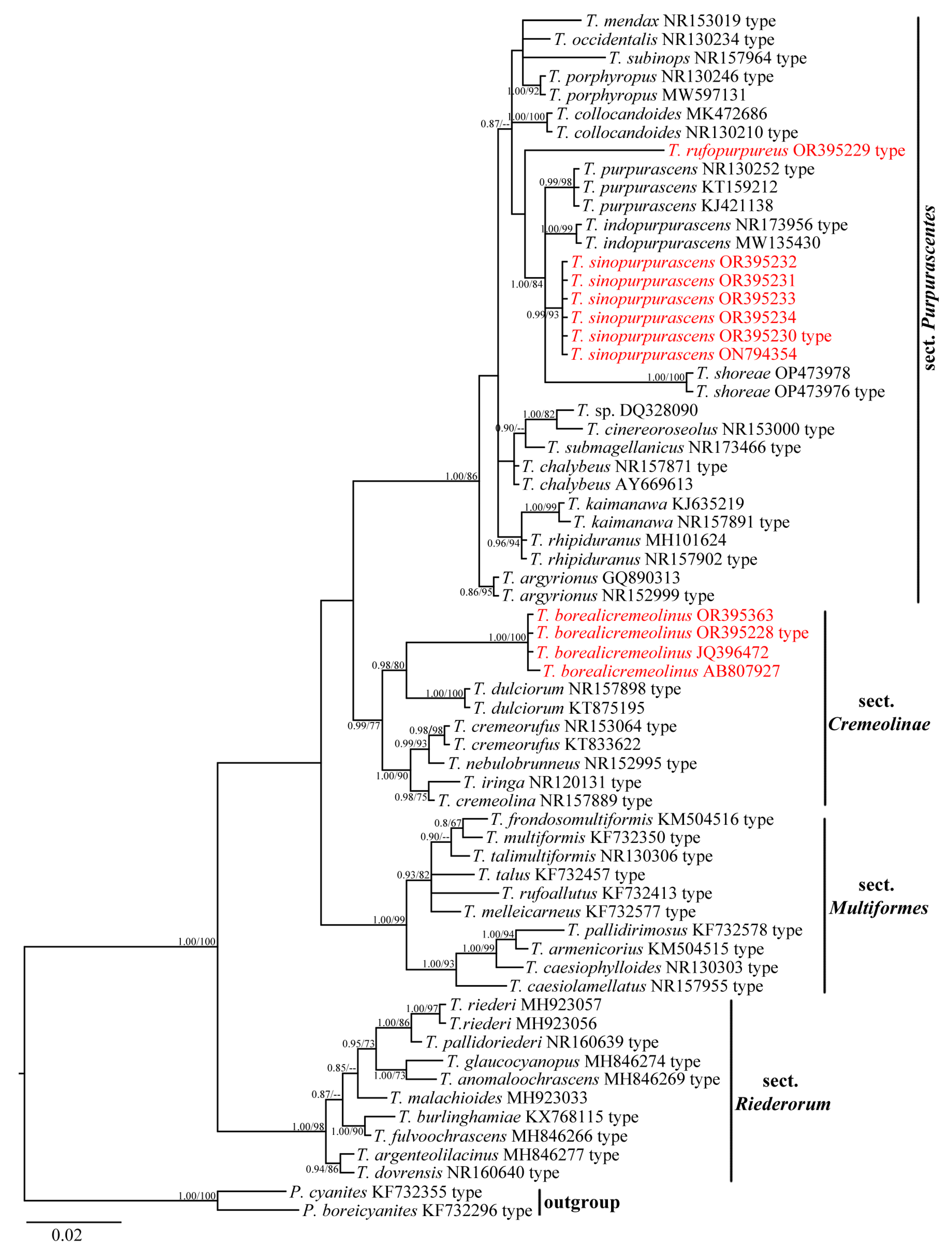
3.1. Molecular Phylogeny
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, R. Thaxterogaster—A new link between Gastromycetes and Agaricales. Mycologia 1951, 43, 215–228. [Google Scholar] [CrossRef]
- Beaton, G.; Pegler, D.N.; Young, T.W.K. Gasteroid Basidiomycota of Victoria State, Australia. 3. Cortinariales. Kew Bull. 1985, 40, 167–204. [Google Scholar] [CrossRef]
- Peintner, U.; Moser, M.; Vilgalys, R. Thaxterogaster is a taxonomic synonym of Cortinarius: New combinations and new names. Mycotaxon 2002, 81, 177–184. [Google Scholar]
- Liimatainen, K.; Kim, J.T.; Pokorny, L.; Kirk, P.M.; Dentinger, B.; Niskanen, T. Taming the beast: A revised classification of Cortinariaceae based on genomic data. Fungal Divers. 2022, 112, 89–170. [Google Scholar] [CrossRef]
- Soop, K. Dix genres pour remplacer Cortinarius. J. JEC 2022, 24, 22–24. [Google Scholar]
- Dai, Y.C.; Zhou, L.W.; Yang, Z.L.; Wen, H.A.; Bau, T.; Li, T.H. A revised checklist of edible fungi in China. Mycosystema 2010, 29, 1–21. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L. A revised checklist of medicinal fungi in China. Mycosystema 2008, 27, 801–824. [Google Scholar] [CrossRef]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963. [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979. [Google Scholar]
- Wang, K.; Xie, M.L.; Liu, T.Z.; Li, G.J.; Liu, D.M.; Wei, T.Z. Taxonomy of Cortinarius in China 1. Subgenus Phlegmacium. Mycosystema 2022, 41, 1370–1397. [Google Scholar] [CrossRef]
- Xie, M.L. Taxonomic, molecular phylogenetic and biogeographic studies of Cortinarius in China. Ph.D. Thesis, Northeast Normal University, Changchun, China, 2022. [Google Scholar]
- Xie, M.L.; Wei, T.Z.; Fu, Y.P.; Li, D.; Qi, L.L.; Xing, P.J.; Cheng, G.H.; Ji, R.Q.; Li, Y. Three new species of Cortinarius subgenus Telamonia (Cortinariaceae, Agaricales) from China. MycoKeys 2020, 69, 91–109. [Google Scholar] [CrossRef]
- Xie, M.L.; Chen, J.L.; Phukhamsakda, C.; Dima, B.; Fu, Y.P.; Ji, R.Q.; Wang, K.; Wei, T.Z.; Li, Y. Cortinarius subsalor and C. tibeticisalor spp. nov., two new species from the section Delibuti from China. PeerJ 2021, 9, e11982. [Google Scholar] [CrossRef]
- Xie, M.L.; Phukhamsakda, C.; Wei, T.Z.; Li, J.P.; Wang, K.; Wang, Y.; Ji, R.Q.; Li, Y. Morphological and phylogenetic evidence reveal five new telamonioid species of Cortinarius (Agaricales) from East Asia. J. Fungi 2022, 8, 257. [Google Scholar] [CrossRef]
- Xie, M.L.; Li, Y.; Wang, K.; Ji, R.Q.; Wei, T.Z. Cortinarius liyui sp. nov., a new species of Cortinarius sect. Liyuorum sect. nov. from China. Mycosystema 2023, 42, 244–251. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of total cellular DNA from plants, algae and fungi. In Plant Molecular Biology Manual; Gelvin, S.B., Schilperoort, R.A., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 1994; pp. 183–190. [Google Scholar]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biology Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program distributed by the author. Evolutionary Biology Centre, Uppsala University.ampignons de l’Équateur (Pugillus IV). Bull. L’Herb. Boissier 2004, 3, 53–74. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. RaxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Gasparini, B.; Soop, K. Contribution to the knowledge of Cortinarius (Agaricales, Cortinariaceae) of Tasmania (Australia) and New Zealand. Australas. Mycol. 2008, 27, 173–203. [Google Scholar]
- Soop, K. A contribution to the study of the cortinarioid mycoflora of New Zealand, VII. N. Zeal. J. Bot. 2016, 54, 344–365. [Google Scholar] [CrossRef]
- Danks, M.; Lebel, T.; Vernes, K. Cort short on a mountaintop—Eight new species of truffle Cortinarius from sub-alpine Australia and affinities to sections within the genus. Persoonia 2010, 24, 106–126. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; Dima, B.; Dovana, F.; et al. Fungal Planet description sheets: 1112–1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef] [PubMed]
- Saar, G.; Brandrud, T.E.; Dima, B.; Schmidt-Stohn, G.; Bellù, F.; Frøslev, T.; Oertel, B.; Soop, K. Cortinarius Untergattung Phlegmacium Sektion Purpurascentes in Europa. J. JEC 2014, 16, 140–161. [Google Scholar]
- Ghosh, A.; Chakraborty, D.; Hembrom, M.E.; Vizzini, A.; Das, K. Thaxterogaster shoreae, a new species of Thaxterogaster subg. Scauri sect. Purpurascentes from Sal Forest of India based on morphology and molecular phylogeny. Taiwania 2023, 68, 23–30. [Google Scholar] [CrossRef]
- Soop, K.; Dima, B.; Cooper, J.A.; Park, D.; Oertel, B. A phylogenetic approach to a global supraspecific taxonomy of Cortinarius (Agaricales) with an emphasis on the southern mycota. Persoonia 2019, 42, 261–290. [Google Scholar] [CrossRef]
- Shen, M.; Zhang, C.Q.; Ma, Y.P.; Stephane, W.; Pierre-Arthur, M.; Marc-André, S. Mycorrhizal features and fungal partners of four mycoheterotrophic Monotropoideae (Ericaceae) species from Yunnan, China. Symbiosis 2012, 57, 1–13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Specimen Status | GenBank Accession no. | Locality |
|---|---|---|---|---|
| Thaxterogaster anomalo-ochrascens | PC: RH2805 | Holotype | MH846269 | France |
| T. argenteolilacinus | M: MM48/752 | Holotype | MH846277 | Austria |
| T. argyrionus | MD162 | GQ890313 | Australia | |
| T. argyrionus | MEL: 2331642 | Holotype | NR152999 | Australia |
| T. armenicorius | KS-CO1865 | Holotype | KM504515 | France |
| T. borealicremeolinus | HMAS 287398 | Holotype | OR395228 | China |
| T. borealicremeolinus | LY418 | OR395363 | China | |
| T. borealicremeolinus | 38-1A | JQ396472 | China | |
| T. borealicremeolinus | Sa17-501 | AB807927 | Japan | |
| T. burlinghamiae | DBB37303 | Holotype | KX768115 | USA |
| T. caesiolamellatus | PC: PML4905 | Holotype | NR157955 | France |
| T. caesiophylloides | H: 6029792 | Holotype | NR130303 | Finland |
| T. chalybeus | PDD: 73146 | Holotype | NR157871 | New Zealand |
| T. chalybeus | CO1342 | AY669613 | Germany | |
| T. rufopurpureus | HMAS 287399 | Holotype | OR395229 | China |
| T. cinereoroseolus | MEL: 2331646 | Holotype | NR153000 | Australia |
| T. collocandoides | CR502 | MK472686 | Luxembourg | |
| T. collocandoides | PC: PML5087 | Holotype | NR130210 | France |
| T. cremeolina | PDD: 70506 | Holotype | NR157889 | New Zealand |
| T. cremeorufus | PDD: 94056 | Holotype | NR153064 | New Zealand |
| T. cremeorufus | PDD: 72649 | KT833622 | New Zealand | |
| T. dovrensis | O: TEB112-80 | Holotype | NR160640 | Norway |
| T. dulciorum | PDD: 78797 | Holotype | NR157898 | New Zealand |
| T. dulciorum | PDD: 107708 | KT875195 | New Zealand | |
| T. frondosomultiformis | TG2000-218 | Holotype | KM504516 | Italy |
| T. fulvo-ochrascens | PC: RH314 | Holotype | MH846266 | France |
| T. glaucocyanopus | G: 5034 | Holotype | MH846274 | France |
| T. indopurpurascens | KCS: 2442 | Holotype | NR173956 | India |
| T. indopurpurascens | KCS: 2529 | MW135430 | India | |
| T. iringa | PDD: 73135 | Holotype | NR120131 | New Zealand |
| T. kaimanawa | PDD: 101872 | KJ635219 | New Zealand | |
| T. kaimanawa | PDD: 73133 | Holotype | NR157891 | New Zealand |
| T. malachioides | TEB340-16 | MH923033 | Sweden | |
| T. melleicarneus | H: IK01-053 | Holotype | KF732577 | Estonia |
| T. mendax | PC: AB07-10-162 | Holotype | NR153019 | France |
| T. multiformis | S: F44806 | Neotype | KF732350 | Sweden |
| T. nebulobrunneus | MEL: 2331648 | Holotype | NR152995 | Australia |
| T. occidentalis | MICH: 10382 | Holotype | NR130234 | USA |
| T. pallidirimosus | H: 6035694 | Holotype | KF732578 | Finland |
| T. pallidoriederi | BOZ: Bellu 30-09-2011 | Holotype | NR160639 | Italy |
| T. porphyropus | S: F47381 | Neotype | NR130246 | Sweden |
| T. porphyropus | SAT-16-237-03 | MW597131 | USA | |
| T. purpurascens | H: IK98-2121 | Neotype | NR130252 | Sweden |
| T. purpurascens | M: 0275842 | KT159212 | Portugal | |
| T. purpurascens | TUB 019710 | KJ421138 | Austria | |
| T. rhipiduranus | PDD: 72617 | MH101624 | New Zealand | |
| T. rhipiduranus | PDD: 88269 | Holotype | NR157902 | New Zealand |
| T. riederi | BOZ: Bellu 12-08-2012 | MH923057 | Italy | |
| T. riederi | TEB141-10 | MH923056 | Sweden | |
| T. rufoallutus | PC: PML635 | Holotype | KF732413 | France |
| T. shoreae | AGJH-017 | OP473978 | India | |
| T. shoreae | AGDC_21-04 | Holotype | OP473976 | India |
| T. sinopurpurascens | HMAS 287400 | Holotype | OR395230 | China |
| T. sinopurpurascens | HMAS 287401 | OR395231 | China | |
| T. sinopurpurascens | HMAS 287402 | OR395232 | China | |
| T. sinopurpurascens | HMAS 287403 | OR395233 | China | |
| T. sinopurpurascens | HMAS 287404 | OR395234 | China | |
| T. sinopurpurascens | HKAS 122529 | ON794354 | China | |
| T. sp. | H0920 | DQ328090 | Australia | |
| T. subinops | PC: PML5119 | Holotype | NR157964 | France |
| T. submagellanicus | MEL: 2305432 | Isoparatype | NR173466 | Australia |
| T. talimultiformis | UPS: AT2004096 | Holotype | NR130306 | Sweden |
| T. talus | S: F44793 | Neotype | KF732457 | Sweden |
| Phlegmacium boreicyanites | S: CFP931 | Holotype | KF732296 | Sweden |
| P. cyanites | UPS: AT2005069 | Neotype | KF732355 | Sweden |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, M.-L.; Feng, N.; Lin, W.-F.; Su, W.-Y.; Li, Y.; Yang, Z.-Q. Morphological and Phylogenetic Characterization of Three Novel Thaxterogaster (Cortinariaceae) Species from China with an Emphasis on Their Subtropical Distribution. J. Fungi 2023, 9, 1058. https://doi.org/10.3390/jof9111058
Xie M-L, Feng N, Lin W-F, Su W-Y, Li Y, Yang Z-Q. Morphological and Phylogenetic Characterization of Three Novel Thaxterogaster (Cortinariaceae) Species from China with an Emphasis on Their Subtropical Distribution. Journal of Fungi. 2023; 9(11):1058. https://doi.org/10.3390/jof9111058
Chicago/Turabian StyleXie, Meng-Le, Na Feng, Wen-Fei Lin, Wen-Ying Su, Yi Li, and Zhen-Quan Yang. 2023. "Morphological and Phylogenetic Characterization of Three Novel Thaxterogaster (Cortinariaceae) Species from China with an Emphasis on Their Subtropical Distribution" Journal of Fungi 9, no. 11: 1058. https://doi.org/10.3390/jof9111058