
TtCel7A: A Native Thermophilic Bifunctional Cellulose/Xylanase Exogluclanase from the Thermophilic Biomass-Degrading Fungus Thielavia terrestris Co3Bag1, and Its Application in Enzymatic Hydrolysis of Agroindustrial Derivatives
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Microorganisms and Growth Conditions
2.3. Cellulase and Xylanase Activity Assays
2.4. Enzyme Purification
2.5. Protein Concentration
2.6. SDS-PAGE and Zymogram Analysis
2.7. Carbohydrate Content
2.8. Biochemical Properties of TtCel7A
2.8.1. Optimal pH and pH Stability
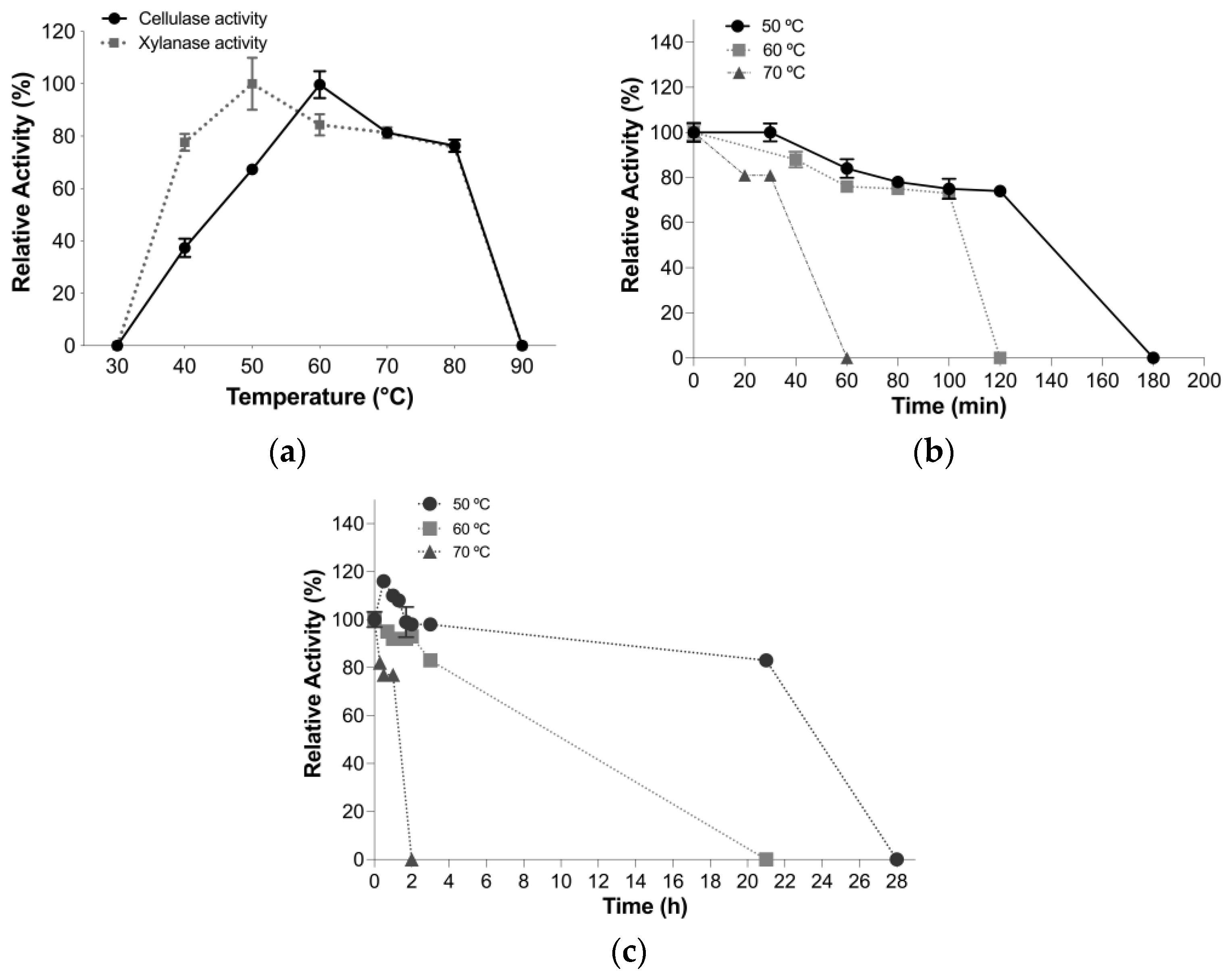
2.8.2. Optimal Temperature and Thermal Stability Assay
2.8.3. Substrate Specificity and Kinetic Parameters
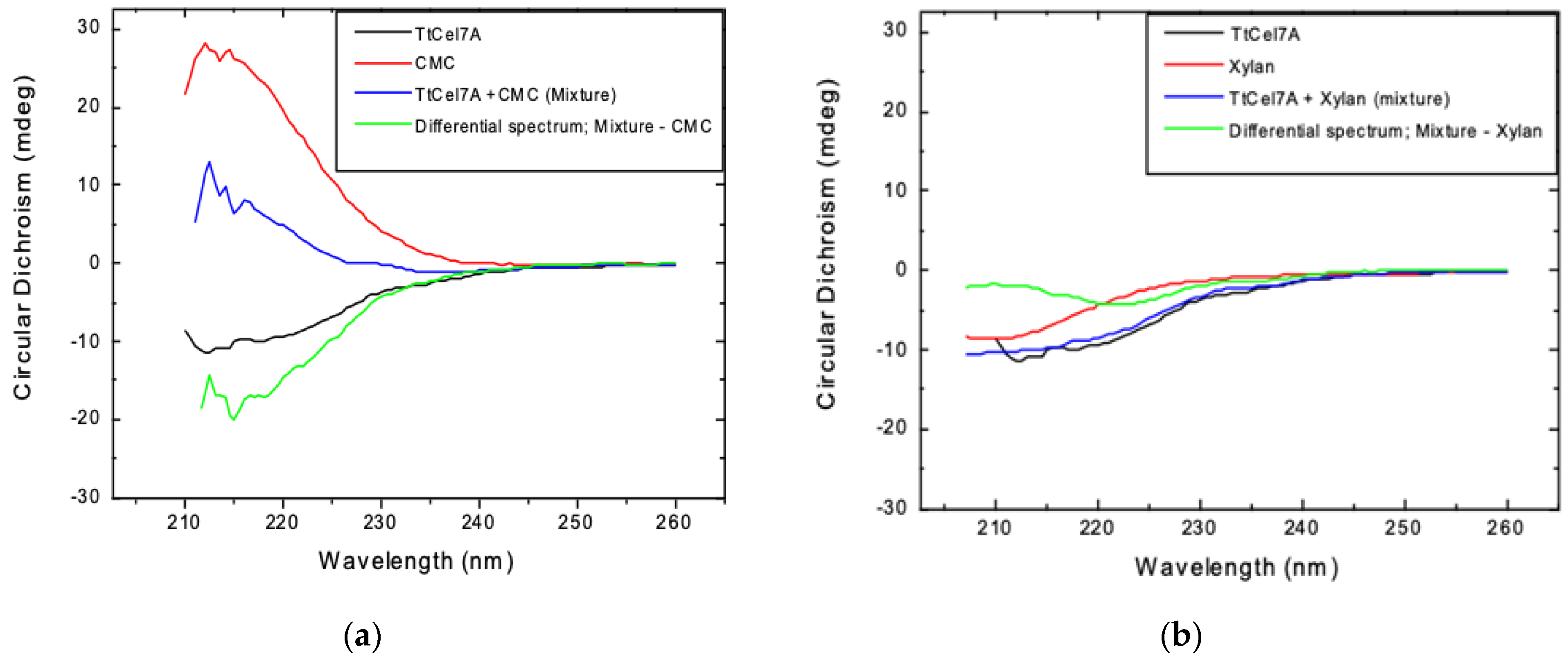
2.8.4. Circular Dichroism
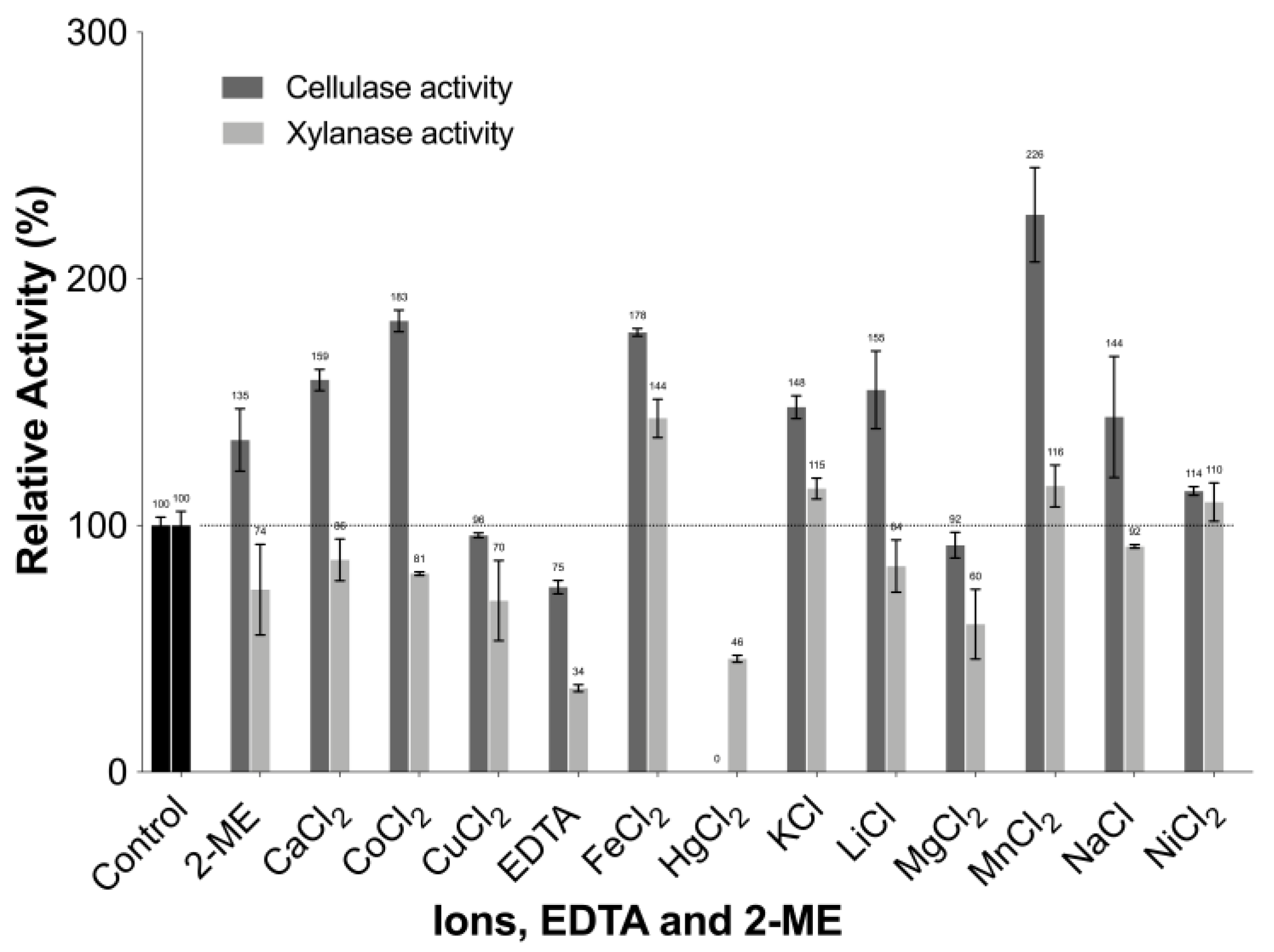
2.8.5. Effect of Ions, EDTA, and 2-mercaptoethanol (2-ME) on Enzyme Activity
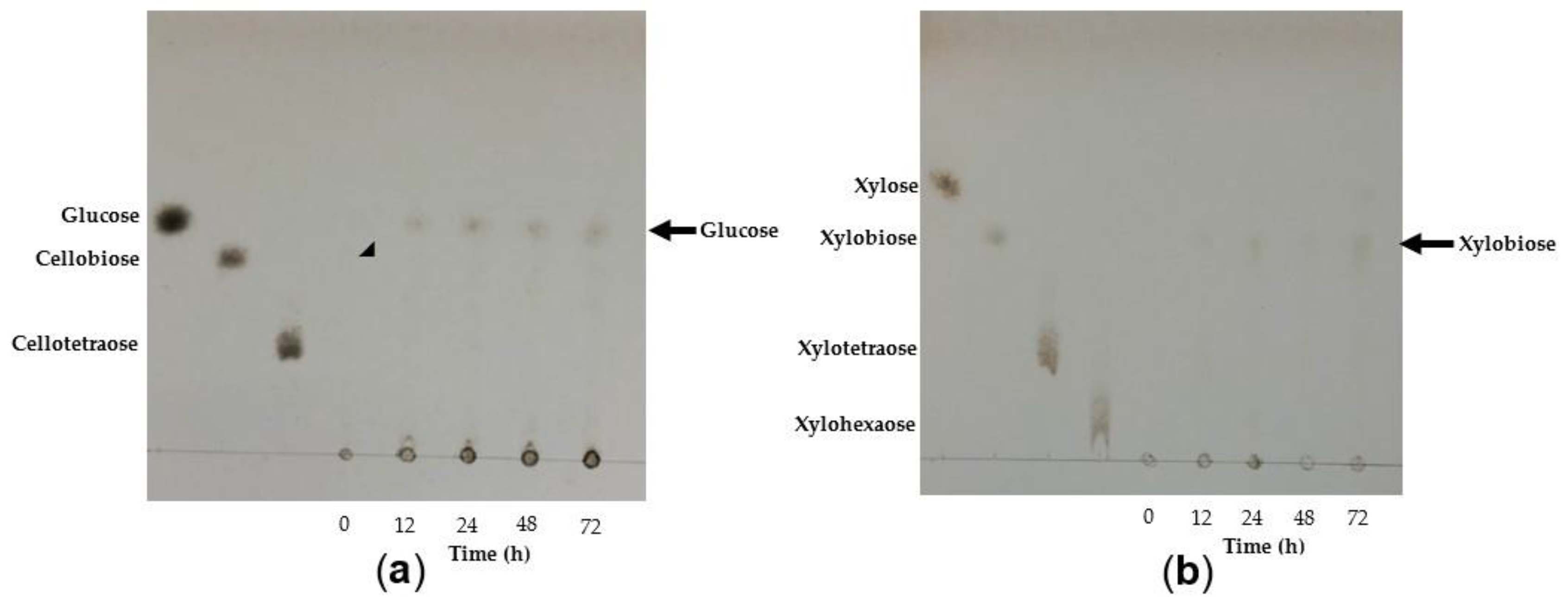
2.8.6. Analysis of TtCel7A Hydrolysis Products
2.9. Partial Amino Acid Sequencing of TtCel7A
2.10. Total RNA Extraction and Synthesis of cDNA from T. terrestris Co3Bag1
2.11. Analysis of TtCel7A Encoding Gene
2.12. Analysis of the 2D and 3D Structures of TtCelA
2.13. Phylogenetic Reconstruction
3. Results
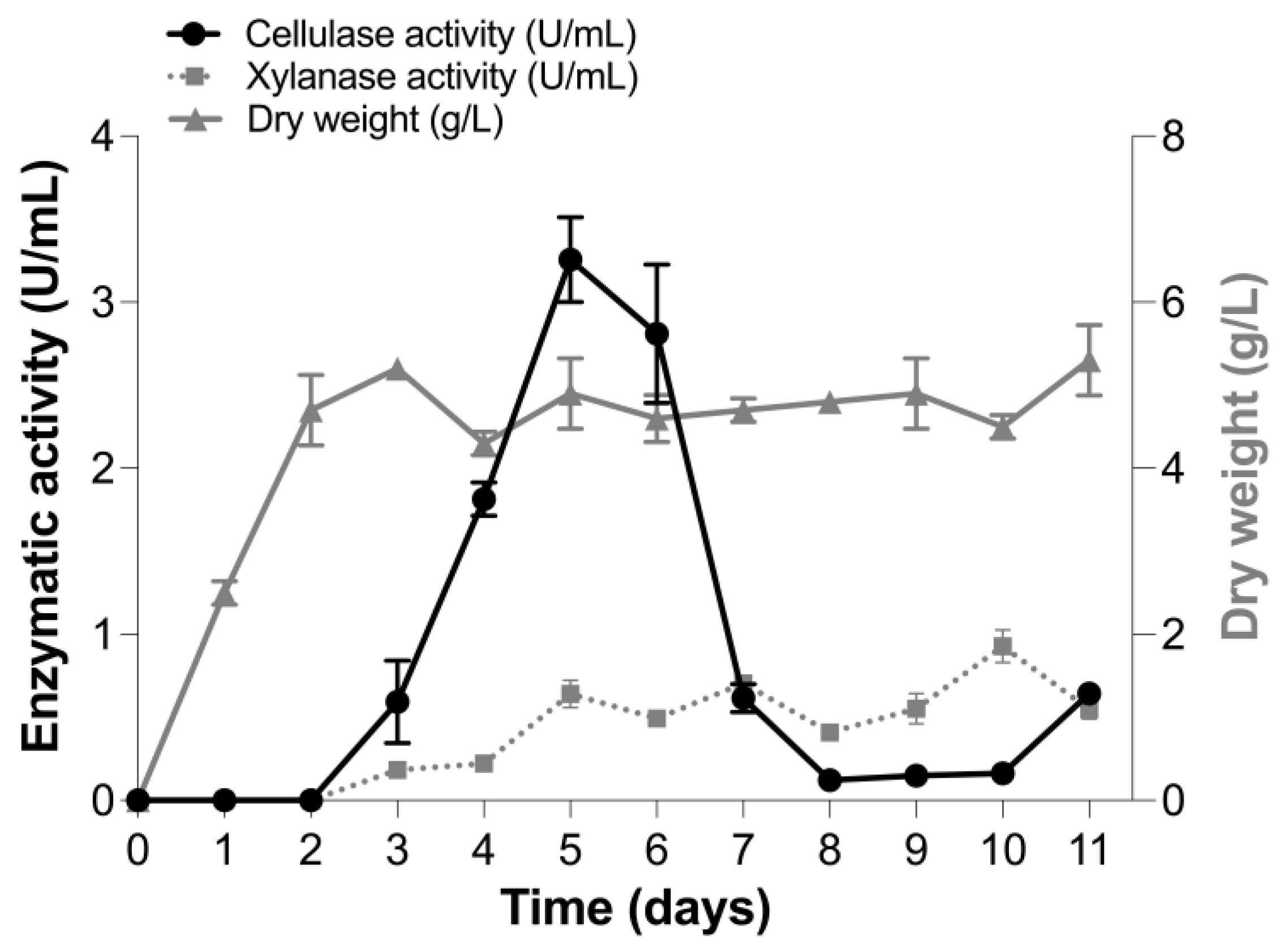
3.1. Production of Cellulase and Xylanase Activities
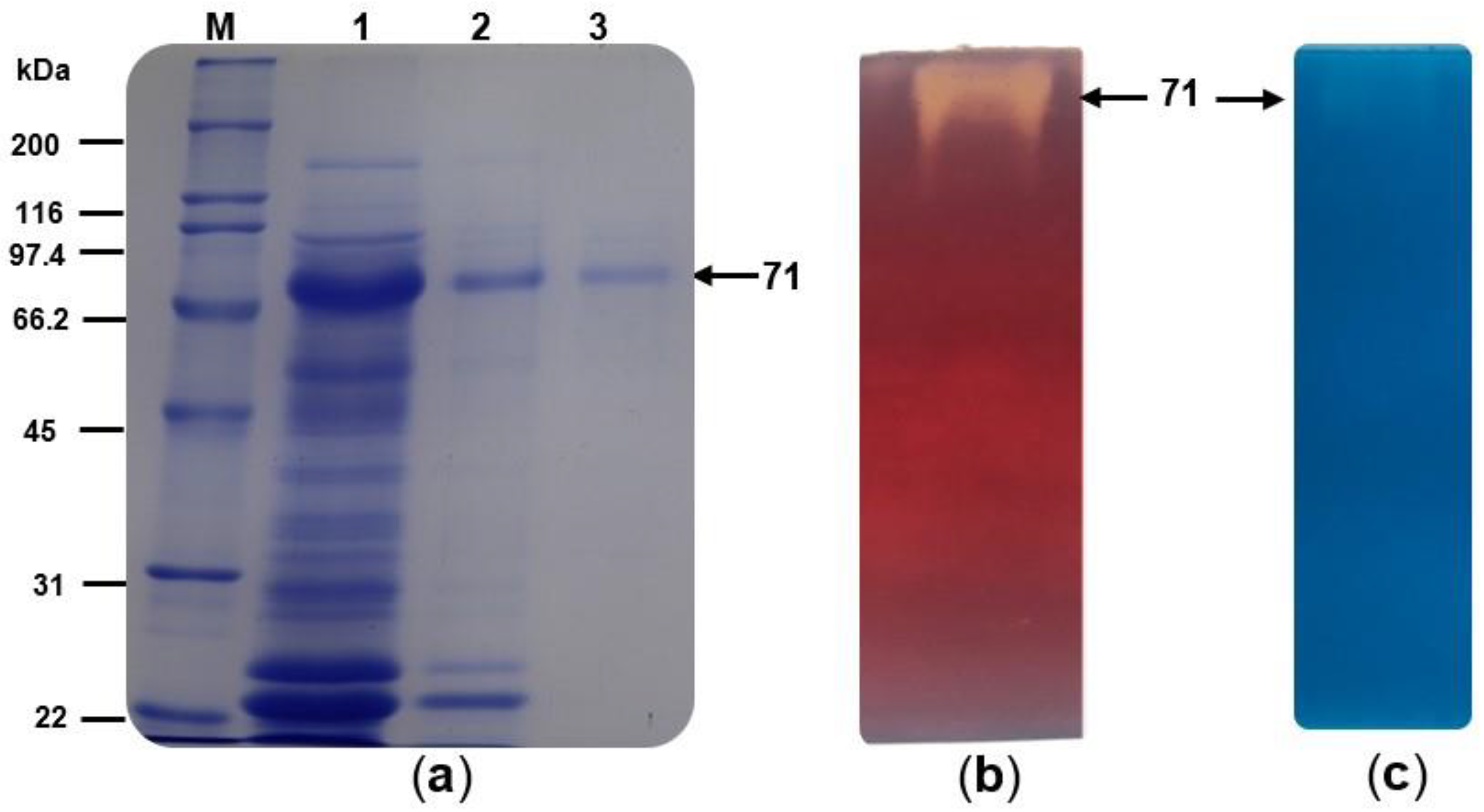
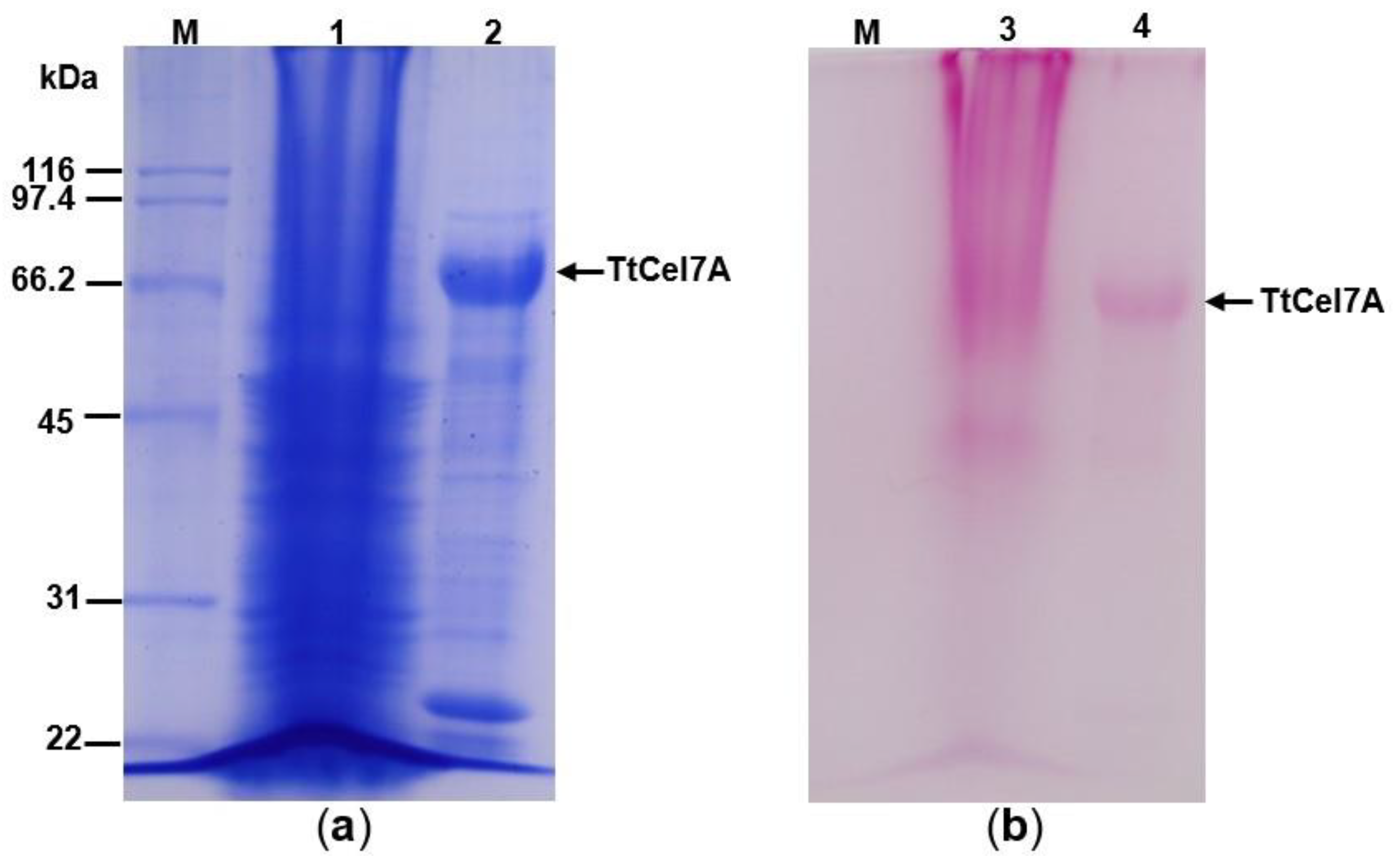
3.2. Enzyme Purification and Zymogram Analysis
3.3. Carbohydrate Content
3.4. Biochemical Properties of TtCel7A
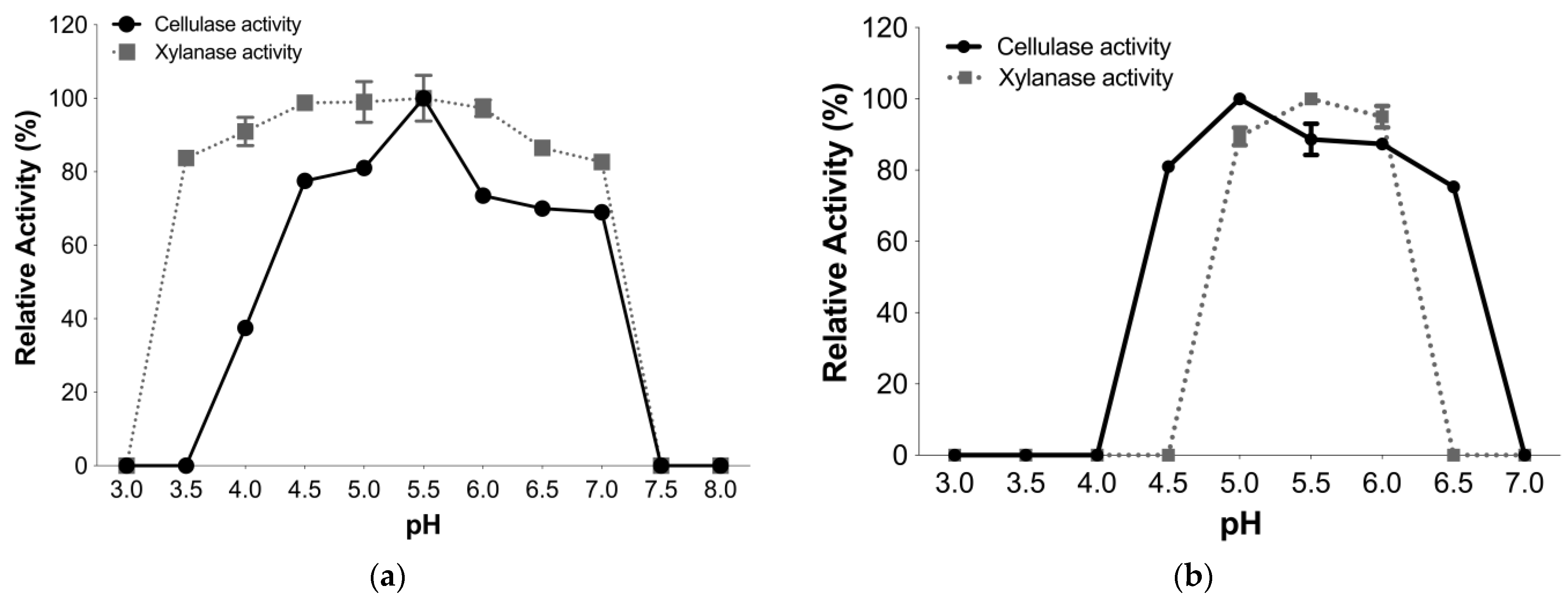
3.4.1. Optimal pH and pH Stability
3.4.2. Optimal Temperature and Thermal Stability Assays
3.4.3. Substrate Specificity and Kinetic Parameters
3.4.4. Circular Dichroism
3.4.5. Effect of Metal Ions, EDTA, and 2-mercaptoethanol (2-ME) on Enzyme Activity
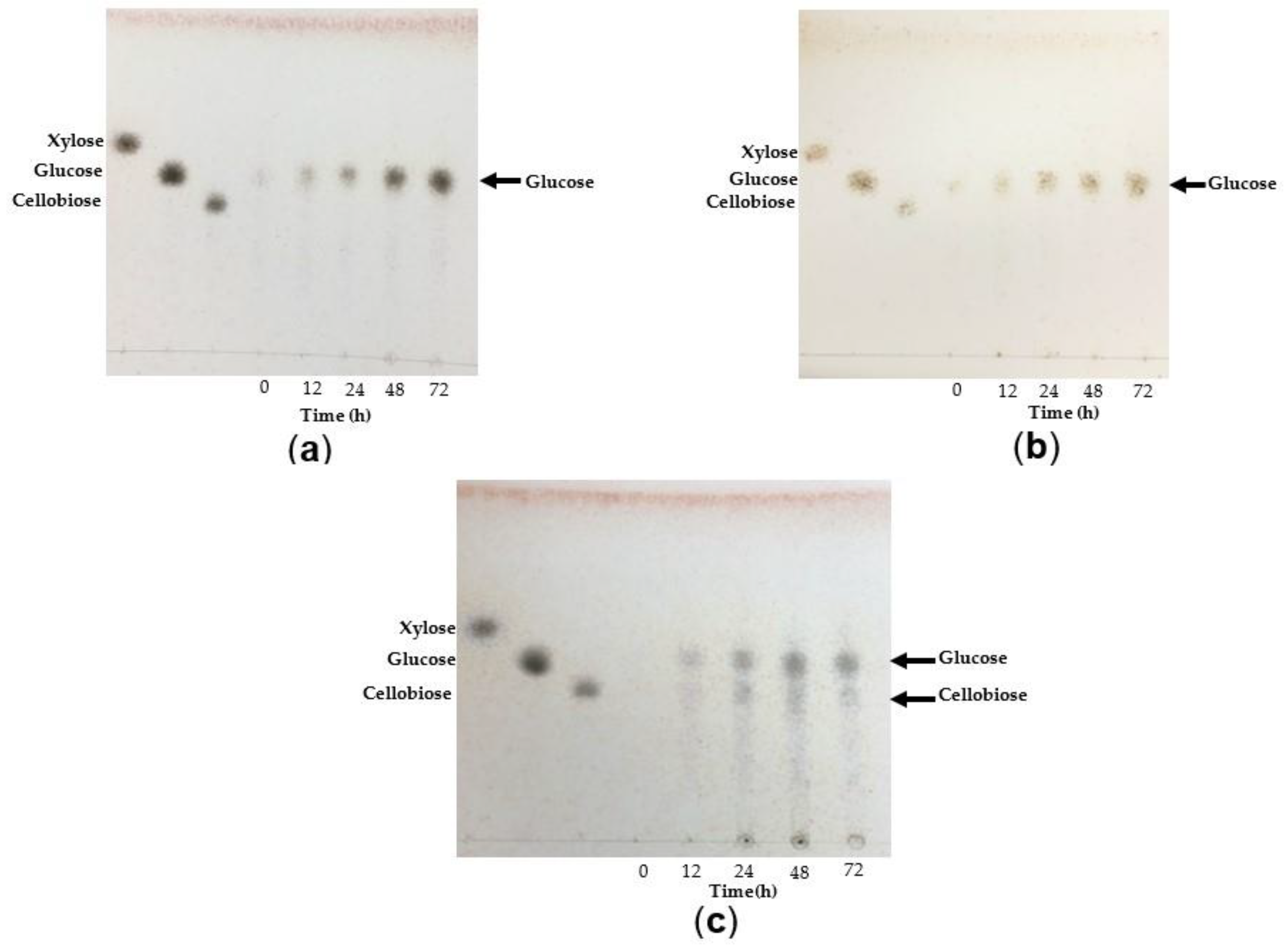
3.4.6. Analysis of TtCel7A Hydrolysis Products
3.5. Analysis of TtCel7A Encoding Gene
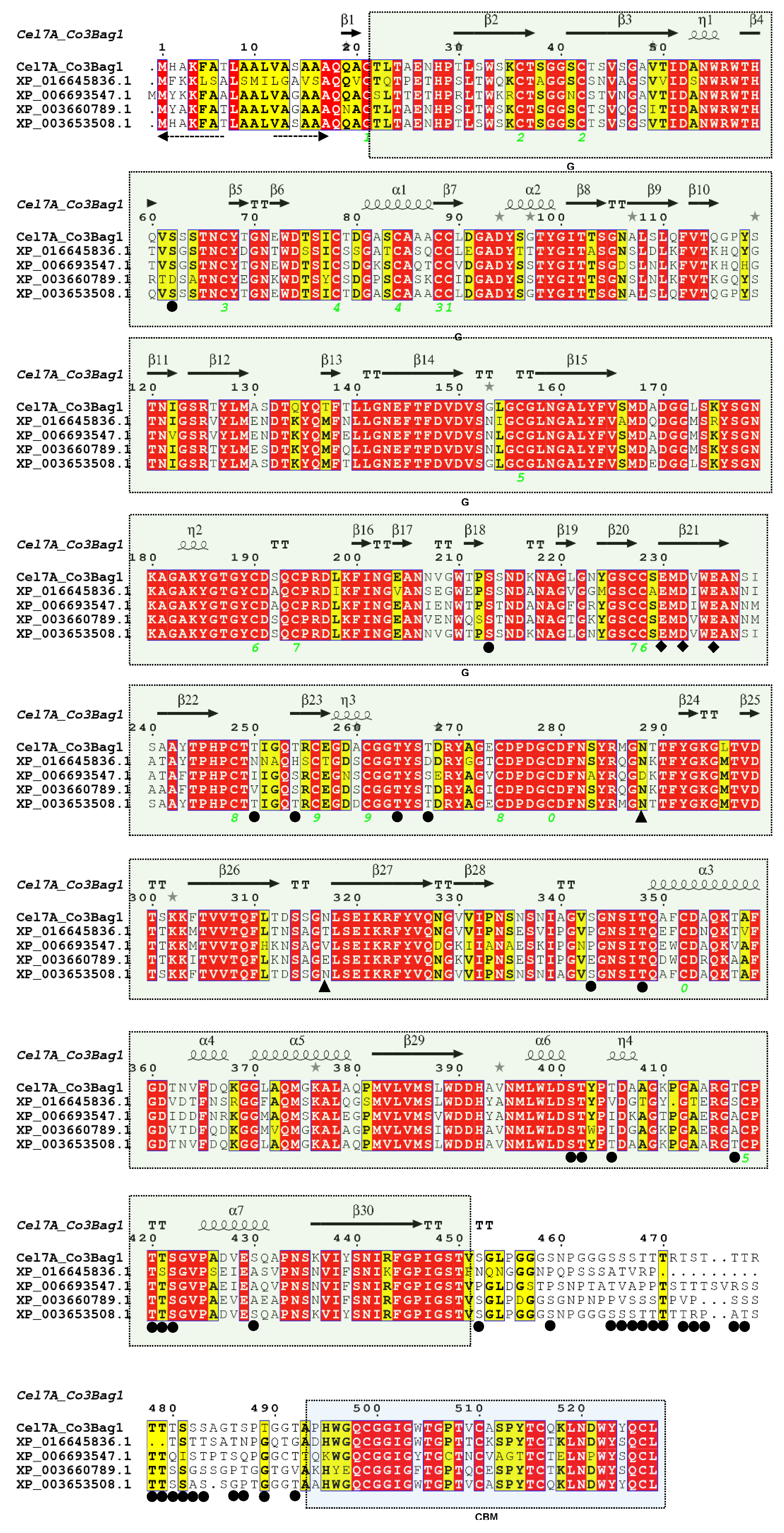
3.6. Identity of TtCel7A by Partial Amino Acid Sequencing
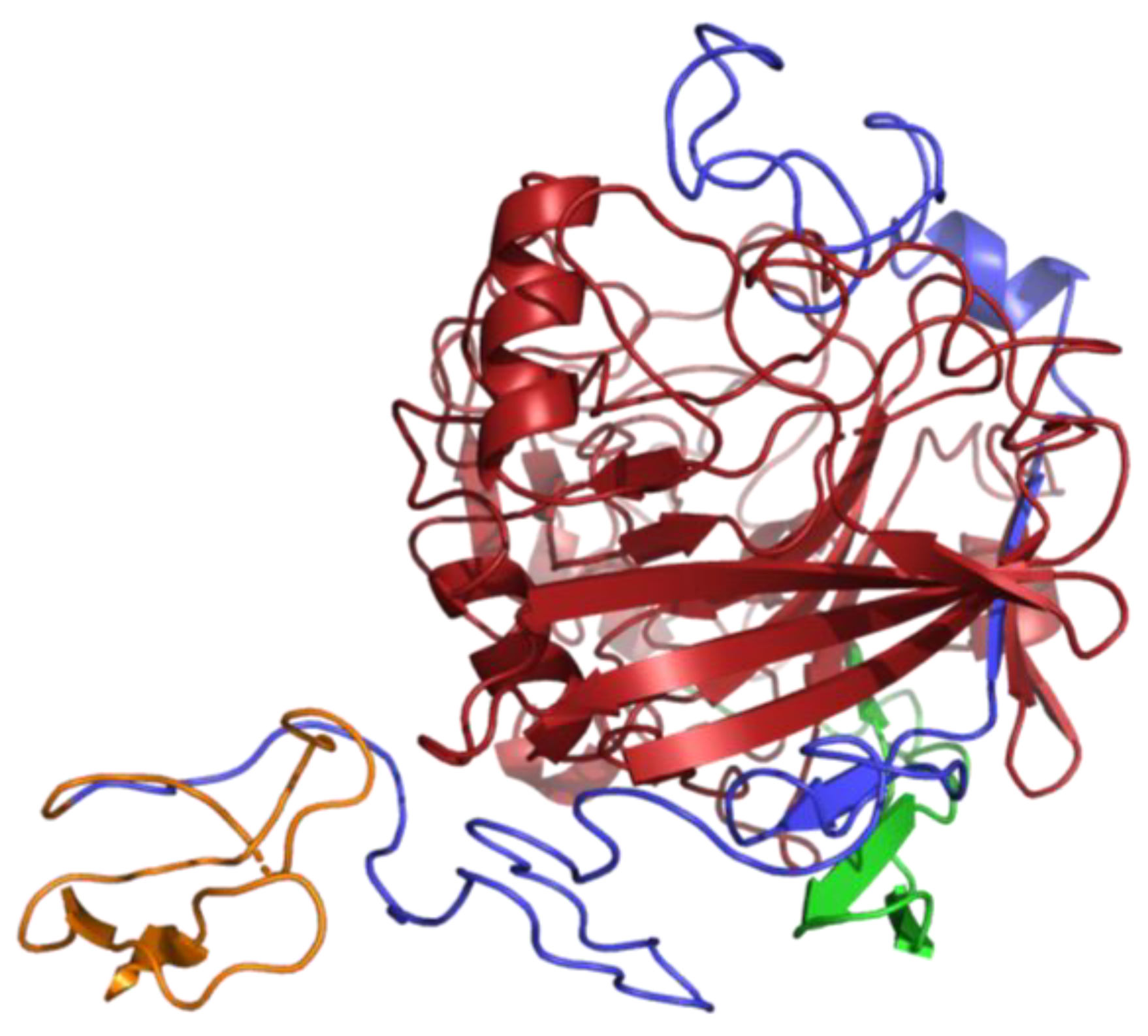
3.7. Analysis of 2D and 3D Structure of TtCel7A
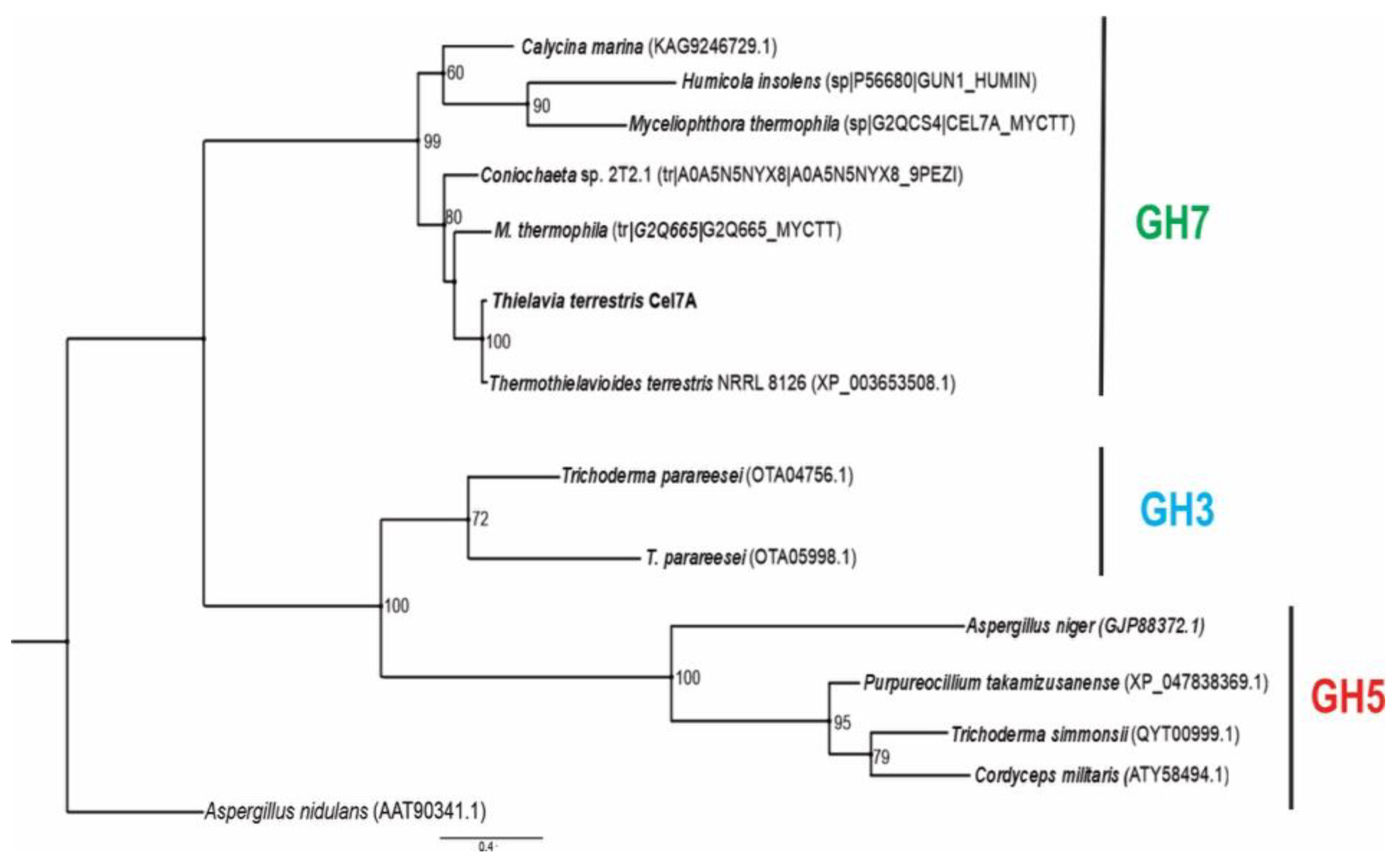
3.8. Phylogenetic Reconstruction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gama, A.R.; Brito-Cunha, C.C.Q.; Campos, I.T.N.; De Souza, G.R.L.; Carneiro, L.C.; Bataus, L.A.M. Streptomyces thermocerradoensis I3 secretes a novel bifunctional xylanase/endoglucanase under solid-state fermentation. Biotechnol. Prog. 2020, 36, e2934. [Google Scholar] [CrossRef] [PubMed]
- Pabbathi, N.P.P.; Velidandi, A.; Tavarna, T.; Gupta, S.; Raj, R.S.; Gandam, P.K.; Baadhe, R.R. Role of metagenomics in prospecting novel endoglucanases, accentuating functional metagenomics approach in second-generation biofuel production: A review. Biomass Conv. Bioref. 2021, 13, 1371–1398. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Ding, M.; Bao, L.; Chen, Y.; Zhou, J.; Lu, H. Characterization of a bifunctional xylanase/endoglucanase from yak rumen microorganisms. Appl. Microbiol. Biotechnol. 2011, 90, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Payne, C.M.; Knott, B.C.; Mayes, H.B.; Hansson, H.; Himmel, M.E.; Sandgren, M.; Ståhlberg, J.; Beckham, G.T. Fungal Cellulases. Chem. Rev. 2015, 115, 1308–1448. [Google Scholar] [CrossRef] [Green Version]
- Pamidipati, S.; Ahmed, A. Degradation of Lignin in Agricultural Residues by locally Isolated Fungus Neurospora discrete. Appl. Biochem. Biotechnol. 2017, 181, 1561–1572. [Google Scholar] [CrossRef]
- Taylor, L.E.; Knott, B.C.; Baker, J.O.; Alahuhta, P.M.; Hobdey, S.E.; Linger, J.G.; Lunin, V.V.; Amore, A.; Subramanian, V.; Podkaminer, K.; et al. Engineering enhanced cellobiohydrolase activity. Nat. Commun. 2018, 9, 1186. [Google Scholar] [CrossRef] [Green Version]
- Chai, S.; Zhang, X.; Jia, Z.; Xu, X.; Zhang, Y.; Wang, S.; Feng, Z. Identification and characterization of a novel bifunctional cellulase/hemicellulase from a soil metagenomic library. Appl. Microbiol. Biotechnol. 2020, 104, 7563–7572. [Google Scholar] [CrossRef]
- Cragg, S.M.; Beckham, G.T.; Bruce, N.C.; Bugg, T.D.H.; Distel, D.L.; Dupree, P.; Etxabe, A.G.; Goodell, B.S.; Jellison, J.; Mcgeehan, J.E.; et al. Lignocellulose degradation mechanisms across the Tree of Life. Curr. Opin. Chem. Biol. 2015, 29, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Gündüz Ergün, B.; Çalik, P. Lignocellulose degrading extremozymes produced by Pichia pastoris: Current status and future prospects. Bioprocess Biosyst. Eng. 2016, 39, 1–36. [Google Scholar] [CrossRef]
- Rattu, G.; Joshi, S.; Satyanarayana, T. Bifunctional recombinant cellulase-xylanase (rBhcell-xyl) from the polyextremophilic bacterium Bacillus halodurans TSLV1 and its utility in valorization of renewable agro-residues. Extremophiles 2016, 20, 831–842. [Google Scholar] [CrossRef]
- Soni, S.K.; Sharma, A.; Soni, R. Cellulases: Role in Lignocellulosic Biomass Utilization. In Cellulases, Methods in Molecular Biology; Lübeck, M., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1796, pp. 3–23. [Google Scholar] [CrossRef]
- Klippel, B.; Blank, S.; Janzer, V.-A.; Piascheck, H.; Moccand, C.; Bel-Rhlid, R.; Antranikian, G. Characterization of a thermoactive endoglucanase isolated from a biogas plant metagenome. Extremophiles 2019, 23, 479–486. [Google Scholar] [CrossRef]
- Duo-Chuan, L.; An-Na, L.; Anastassios, C.P. Cellulases from Thermophilic Fungi: Recent Insights and Biotechnological Potential. In Fungi in Extreme Environments: Ecological Role and Biotechnological Significance; Springer: Berlin/Heidelberg, Germany, 2011; pp. 395–417. [Google Scholar] [CrossRef] [Green Version]
- Rana, V.; Rana, D. Role of Microorganisms in Lignocellulosic Biodegradation. In Renewable Biofuels; Springer: Cham, Switzerland, 2017; pp. 19–67. [Google Scholar] [CrossRef]
- Escuder-Rodriguez, J.J.; Decastro, M.E.; Cerdan, M.E.; Rodriguez-Belmonte, E.; Becerra, M.; Gonzalez-Siso, M.I. Cellulases from Thermophiles Found by Metagenomics. Microorganisms 2018, 6, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, N.; Kumar, B.; Agarwal, K.; Chaturvedi, V.; Verma, P. Purification and characterization of a thermo-acid/alkali stable xylanases from Aspergillus oryzae LC1 and its application in Xylo-oligosaccharides production from lignocellulosic agricultural wastes. Int. J. Biol. Macromol. 2019, 122, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, R.C.; Gupta, R.; Singh, A. Microbial cellulases and their industrial applications. Enzyme Res. 2011, 2011, 280696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.K.; Singhania, R.R.; Sim, S.J.; Pandey, A. Thermostable cellulases: Current status and perspectives. Bioresour. Technol. 2019, 279, 385–392. [Google Scholar] [CrossRef]
- Basit, A.; Liu, J.; Rahim, K.; Jiang, W.; Lou, H. Thermophilic xylanases: From bench to bottle. Crit. Rev. Biotechnol. 2018, 38, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, K.; Sahoo, R.K.; Gaur, M.; Subudhi, E. Cellulolytic thermophilic microorganisms in white biotechnology: A review. Folia Microbiol. 2020, 65, 25–43. [Google Scholar] [CrossRef]
- Poças-Fonseca, M.J.; De Melo Matos, R.W.; Mello-De-Sousa, T.M. The Lignocellulolytic System of Thermophilic Fungi and Actinomycetes: Structure, Regulation and Biotechnological Applications. In Thermophilic Microbes in Environmental and Industrial Biotechnology; Satyanarayana, T., Littlechild, J., Kawarabayasi, Y., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 731–769. [Google Scholar] [CrossRef]
- Kadowaki, M.A.S.; Higasi, P.; De Godoy, M.O.; Prade, R.A.; Polikarpov, I. Biochemical and structural insights into a thermostable cellobiohydrolase from Myceliophthora thermophila. FEBS J. 2018, 285, 559–579. [Google Scholar] [CrossRef] [Green Version]
- Maheshwari, R.; Bharadwaj, G.; Bhat, M.K. Thermophilic Fungi: Their Physiology and Enzymes. Microbiol. Mol. Biol. Rev. 2000, 64, 461–488. [Google Scholar] [CrossRef] [Green Version]
- Pereira, J.C.; Marques, N.P.; Rodrigues, A.; Oliveira, T.B.D.; Boscolo, M.; Silva, R.; Gomes, E.; Martins, D.A.B. Thermophilic fungi as new sources for production of cellulases and xylanases with potential use in sugarcane bagasse saccharification. J. Appl. Microbiol. 2015, 118, 928–939. [Google Scholar] [CrossRef] [Green Version]
- Borisova, A.S.; Eneyskaya, E.V.; Jana, S.; Badino, S.F.; Kari, J.; Amore, A.; Karlsson, M.; Hansson, H.; Sandgren, M.; Himmel, M.E.; et al. Correlation of structure, function and protein dynamics in GH7 cellobiohydrolases from Trichoderma atroviride, T. reesei and T. harzianum. Biotechnol. Biofuels 2018, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnaouri, A.C.; Topakas, E.; Christakopoulos, P. Cloning, expression, and characterization of a thermostable GH7 endoglucanase from Myceliophthora thermophila capable of high-consistency enzymatic liquefaction. Appl. Microbiol. Biotechnol. 2014, 98, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, V.O.A.; Serpa, V.I.; Godoy, A.S.; Camilo, C.M.; Bernardes, A.; Rezende, C.A.; Junior, N.P.; Franco Cairo, J.P.L.; Squina, F.M.; Polikarpov, I. Recombinant Trichoderma harzianum endoglucanase I (Cel7B) is a highly acidic and promiscuous carbohydrate-active enzyme. Appl. Microbiol. Biotechnol. 2015, 99, 9591–9604. [Google Scholar] [CrossRef] [PubMed]
- Hua, C.; Li, W.; Han, W.; Wang, Q.; Bi, P.; Han, C.; Zhu, L. Characterization of a novel thermostable GH7 endoglucanase from Chaetomium thermophilum capable of xylan hydrolysis. Int. J. Biol. Macromol. 2018, 117, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Yang, R.; Sun, Y.; Liu, M.; Zhou, L.; Li, D. Identification and Characterization of a Novel Hyperthermostable Bifunctional Cellobiohydrolase-Xylanase Enzyme for Synergistic Effect with Commercial Cellulase on Pretreated Wheat Straw Degradation. Front. Bioeng. Biotechnol. 2020, 8, 296. [Google Scholar] [CrossRef] [PubMed]
- Berka, R.M.; Grigoriev, I.V.; Otillar, R.; Salamov, A.; Grimwood, J.; Reid, I.; Ishmael, N.; John, T.; Darmond, C.; Moisan, M.-C.; et al. Comparative genomic analysis of the thermophilic biomass-degrading fungi Myceliophthora thermophila and Thielavia terrestris. Nat. Biotechnol. 2011, 29, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Mendoza, J.; Santiago-Hernández, A.; Alvarez-Zúñiga, M.T.; Gutiérrez-Antón, M.; Aguilar-Osorio, G.; Hidalgo-Lara, M.E. Purification and biochemical characterization of a novel thermophilic exo-β-1,3-glucanase from the thermophile biomass-degrading fungus Thielavia terrestris Co3Bag1. Electron. J. Biotechnol. 2019, 41, 60–71. [Google Scholar] [CrossRef]
- García-Huante, Y.; Cayetano-Cruz, M.; Santiago-Hernández, A.; Cano-Ramírez, C.; Marsch-Moreno, R.; Campos, J.E.; Aguilar-Osorio, G.; Benítez-Cardoza, C.G.; Trejo-Estrada, S.; Hidalgo-Lara, M.E. The thermophilic biomass-degrading fungus Thielavia terrestris Co3Bag1 produces a hyperthermophilic and thermostable β-1,4-xylanase with exo- and endo-activity. Extremophiles 2017, 21, 175–186. [Google Scholar] [CrossRef]
- Woon, J.S.-K.; Mackeen, M.M.; Mahadi, N.M.; Illias, R.M.; Abdul Murad, A.M.; Abu Bakar, F.D. Expression and characterization of a cellobiohydrolase (CBH7B) from the thermophilic fungus Thielavia terrestris in Pichia pastoris. Biotechnol. Appl. Biochem. 2016, 63, 690–698. [Google Scholar] [CrossRef]
- Tien, M.; Kirk, T.K. Lignin peroxidase of Phanerochaete chrysosporium. Methods Enzymol. 1988, 161, 238–249. [Google Scholar] [CrossRef]
- Zouari-Mechichi, H.; Mechichi, T.; Dhouib, A.; Sayadi, S.; Martínez, A.T.; Martínez, M.J. Laccase purification and characterization from Trametes trogii isolated in Tunisia: Decolorization of textile dyes by the purified enzyme. Enzym. Microb. Technol. 2006, 39, 141–148. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Cano-Ramírez, C.; Santiago-Hernández, A.; Rivera-Orduña, F.N.; García-Huante, Y.; Zúñiga, G.; Hidalgo-Lara, M.E. Expression, purification and characterization of an endoglucanase from Serratia proteamaculans CDBB-1961, isolated from the gut of Dendroctonus adjunctus (Coleoptera: Scolytinae). AMB Express 2016, 6, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacharius, R.M.; Zell, T.E.; Morrison, J.H.; Woodlock, J.J. Glycoprotein staining following electrophoresis on acrylamide gels. Anal. Biochem. 1969, 30, 148–152. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Pavón-Orozco, P.; Santiago-Hernández, A.; Rosengren, A.; Hidalgo-Lara, M.E.; Stålbrand, H. The family II carbohydrate-binding module of xylanase CflXyn11A from Cellulomonas flavigena increases the synergy with cellulase TrCel7B from Trichoderma reesei during the hydrolysis of sugarcane bagasse. Bioresour. Technol. 2012, 104, 622–630. [Google Scholar] [CrossRef]
- Nakazawa, H.; Okada, K.; Kobayashi, R.; Kubota, T.; Onodera, T.; Ochiai, N.; Omata, N.; Ogasawara, W.; Okada, H.; Morikawa, Y. Characterization of the catalytic domains of Trichoderma reesei endoglucanase I, II and III, expressed in Escherichia coli. Appl. Microbiol. Biotechnol. 2008, 81, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Voutilainen, S.P.; Puranen, T.; Siika-Aho, M.; Lappalainen, A.; Alapuranen, M.; Kallio, J.; Hooman, S.; Viikari, L.; Vehmaanperä, J.; Koivula, A. Cloning, expression, and characterization of novel thermostable family 7 cellobiohydrolases. Biotechnol. Bioeng. 2008, 101, 515–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, N.; Qian, Y.; Wang, W.; Zhong, Y.; Dai, M. Heterologous expression of Talaromyces emersonii cellobiohydrolase Cel7A in Trichoderma reesei increases the efficiency of corncob residues saccharification. Biotechnol. Lett. 2018, 40, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Alokika; Singh, B. Production, characteristics, and biotechnological applications of microbial xylanases. Appl. Microbiol. Biotechnol. 2019, 103, 8763–8784. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Tian, H.; Cheng, Y.; Jiang, Z.; Yang, S. Purification and characterization of a thermostable cellulase-free xylanase from the newly isolated Paecilomyces themophila. Enzym. Microb. Technol. 2006, 38, 780–787. [Google Scholar] [CrossRef]
- Jiang, Z.; Cong, Q.; Yan, Q.; Kumar, N.; Du, X. Characterisation of a thermostable xylanase from Chaetomium sp. and its application in Chinese steamed bread. Food Chem. 2010, 120, 457–462. [Google Scholar] [CrossRef]
- Liu, Y.; Dun, B.; Shi, P.; Ma, R.; Luo, H.; Bai, Y.; Xie, X.; Yao, B. A Novel GH7 Endo- β -1,4-Glucanase from Neosartorya fischeri P1 with Good Thermostability, Broad Substrate Specificity and Potential Application in the Brewing Industry. PLoS ONE 2015, 10, e0137485. [Google Scholar] [CrossRef]
- Liao, H.; Zheng, H.; Li, S.; Wei, Z.; Mei, X.; Ma, H.; Shen, Q.; Xu, Y. Functional diversity and properties of multiple xylanases from Penicillium oxalicum GZ-2. Sci. Rep. 2015, 5, 12631. [Google Scholar] [CrossRef] [Green Version]
- Beckham, G.T.; Bomble, Y.J.; Matthews, J.F.; Taylor, C.B.; Resch, M.G.; Yarbrough, J.M.; Decker, S.R.; Bu, L.; Zhao, X.; Mccabe, C.; et al. The O-glycosylated linker from the Trichoderma reesei Family 7 cellulase is a flexible, disordered protein. Biophys. J. 2010, 99, 3773–3781. [Google Scholar] [CrossRef] [Green Version]
- Woon, J.S.-K.; Mackeen, M.M.; Sudin, A.H.B.; Mahadi, N.M.; Illias, R.M.; Murad, A.M.A.; Bakar, F.D.A. Production of an oligosaccharide-specific cellobiohydrolase from the thermophilic fungus Thielavia terrestris. Biotechnol. Lett. 2016, 38, 825–832. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|
| Crude extract | 124 | 239 | 0.52 | 100 | 1.0 |
| 80% (NH4)2SO4 precipitation | 26.00 | 45.61 | 0.57 | 20.96 | 1.10 |
| Anion exchange chromatography | 10.85 | 1.73 | 6.27 | 8.75 | 12.06 |
| Gel filtration chromatography | 7.62 | 0.23 | 33.13 | 6.15 | 63.71 |
| Substrates | KM (mg/mL) | Vmax (U/mg) | Catalytic Efficiency (mL/mg s) | kcat (s−1) |
|---|---|---|---|---|
| CMC a | 3.12 | 50 | 0.021 | 0.07 |
| Beechwood xylan b | 0.17 | 42.75 | 0.023 | 3.9 × 10−3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-López, A.; Santiago-Hernández, A.; Cayetano-Cruz, M.; García-Huante, Y.; Campos, J.E.; Bustos-Jaimes, I.; Marsch-Moreno, R.; Cano-Ramírez, C.; Benitez-Cardoza, C.G.; Hidalgo-Lara, M.E. TtCel7A: A Native Thermophilic Bifunctional Cellulose/Xylanase Exogluclanase from the Thermophilic Biomass-Degrading Fungus Thielavia terrestris Co3Bag1, and Its Application in Enzymatic Hydrolysis of Agroindustrial Derivatives. J. Fungi 2023, 9, 152. https://doi.org/10.3390/jof9020152
López-López A, Santiago-Hernández A, Cayetano-Cruz M, García-Huante Y, Campos JE, Bustos-Jaimes I, Marsch-Moreno R, Cano-Ramírez C, Benitez-Cardoza CG, Hidalgo-Lara ME. TtCel7A: A Native Thermophilic Bifunctional Cellulose/Xylanase Exogluclanase from the Thermophilic Biomass-Degrading Fungus Thielavia terrestris Co3Bag1, and Its Application in Enzymatic Hydrolysis of Agroindustrial Derivatives. Journal of Fungi. 2023; 9(2):152. https://doi.org/10.3390/jof9020152
Chicago/Turabian StyleLópez-López, Azucena, Alejandro Santiago-Hernández, Maribel Cayetano-Cruz, Yolanda García-Huante, Jorge E. Campos, Ismael Bustos-Jaimes, Rodolfo Marsch-Moreno, Claudia Cano-Ramírez, Claudia G. Benitez-Cardoza, and María Eugenia Hidalgo-Lara. 2023. "TtCel7A: A Native Thermophilic Bifunctional Cellulose/Xylanase Exogluclanase from the Thermophilic Biomass-Degrading Fungus Thielavia terrestris Co3Bag1, and Its Application in Enzymatic Hydrolysis of Agroindustrial Derivatives" Journal of Fungi 9, no. 2: 152. https://doi.org/10.3390/jof9020152