
The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems
by
 , ,
, ,
Ulla Kaasalainen
1,2,*,
Paul M. Kirika
3,
Neduvoto P. Mollel
4,
Andreas Hemp
5 and
Jouko Rikkinen
1,6 1
Finnish Museum of Natural History, University of Helsinki, P.O. Box 7, 00014 Helsinki, Finland
2
Department of Geobiology, University of Göttingen, Goldschmidtstraße 3, 37077 Göttingen, Germany
3
National Museums of Kenya, East African Herbarium, Museum Hill Road, P.O. Box 45166, Nairobi 00100, Kenya
4
National Herbarium, Tropical Pesticides Research Institute, P.O. Box 3024, Arusha 23201, Tanzania
5
Department of Plant Systematics, University of Bayreuth, Universitätsstraße 30, 95440 Bayreuth, Germany
6
Organismal and Evolutionary Biology Research Programme, Faculty of Biological and Environmental Sciences, University of Helsinki, P.O. Box 65, 00014 Helsinki, Finland
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(2), 246; https://doi.org/10.3390/jof9020246
Submission received: 30 December 2022
/
Revised: 3 February 2023
/
Accepted: 6 February 2023
/
Published: 12 February 2023
(This article belongs to the Special Issue Species Diversity, Ecology and Evolutionary History of Lichenized Fungi)
Abstract
:The lichen flora of Africa is still poorly known. In many parts of the tropics, recent studies utilizing DNA methods have revealed extraordinary diversity among various groups of lichenized fungi, including the genus Sticta. In this study, East African Sticta species and their ecology are reviewed using the genetic barcoding marker nuITS and morphological characters. The studied regions represent montane areas in Kenya and Tanzania, including the Taita Hills and Mt. Kilimanjaro, which belong to the Eastern Afromontane biodiversity hotspot. Altogether 14 Sticta species are confirmed from the study region, including the previously reported S. fuliginosa, S. sublimbata, S. tomentosa, and S. umbilicariiformis. Sticta andina, S. ciliata, S. duplolimbata, S. fuliginoides, and S. marginalis are reported as new to Kenya and/or Tanzania. Sticta afromontana, S. aspratilis, S. cellulosa, S. cyanocaperata, and S. munda, are described as new to science. The abundance of new diversity detected and the number of taxa represented by only few specimens show that more comprehensive sampling of the region may be needed to reveal the true diversity of Sticta in East Africa. More generally, our results highlight the need for further taxonomic studies of lichenized fungi in the region.
1. Introduction
Tropical mountains, and especially their forests, are hot spots of biodiversity and endemism [1,2,3,4]. In East Africa, montane regions, such as the ancient Eastern Arc Mountains which range from southern Tanzania to Kenya, and the much younger volcanic mountains, such as Mt. Kilimanjaro in Tanzania and Mt. Elgon at the border of Kenya and Uganda, are surrounded by vast stretches of arid woodlands and savannas [5,6]. Still, especially the humid upper windward slopes of these mountains have provided refuge for the montane rainforests already for millions of years [6,7]. During this time, climatic induced fluctuations in the areal extent and isolation of moist montane forests have generated remarkably high levels of diversity and local endemism [1,3,6,8,9] in what is now known as the Eastern Afromontane biodiversity hotspot [10,11].
The lichen flora of Africa is poorly known and is still largely based on information collected during the 20th century [12]. So far, only few groups of parmelioid and cyanolichens have been studied in any detail, and especially the more recent application of DNA methods has revealed high levels of previously unknown diversity [13,14,15,16,17,18]. Sticta (Schreb.) Ach. (Lobariaceae, Peltigerales) is a genus of foliose macrolichens characterized by well-differentiated pores called cyphellae on the lower surface. Recent molecular studies from different parts of the globe have demonstrated that the genus is very rich in species, with altogether 500 or more species [19,20,21,22,23]. Many traditionally delimited Sticta species, such as S. fuliginosa and S. weigelii, have been shown to represent complexes of taxa with a somewhat similar gross morphology, often called morphodemes [19,20,21,22,23]. High species diversity has been found from tropical montane areas of the Neotropics, and Madagascar and nearby islands [19,24,25]. In their classical guide to East African macrolichens, Swinscow and Krog [12] listed ten Sticta species from an East African region encompassing Ethiopia, Kenya, Tanzania, and Uganda. Since then, only one additional species has been reported from this region [26]. These reported taxa include S. ambavillaria (Bory) Ach., S. cyphellulata (Müll. Arg.) Hue, S. dichotoma Delise, S. fuliginosa (Hoffm.) Ach., S. kunthii Hook. f., S. limbata (Sm.) Ach., S. orbicularis (Braun) Hue, S. papyracea Delise, S. sublimbata (Steiner) Swinscow and Krog, S. tomentosa (Swartz) Ach., and S. weigelii var. weigelii (Ach.) Vainio and S. weigelii var. xanthotropa (Krempelh.) Hue [12,26,27,28,29,30,31], of which the latter is now known as S. xanthotropa (Kremp.) D. J. Galloway [32]. Sticta papyracea has been treated as a synonym of S. variabilis Ach. [33], and the reports of Sticta limbata from many parts of the world, including East Africa, probably represent S. umbilicariiformis Hochsc. Ex Flotow [20], a species originally described from Ethiopia. Additionally, S. duplolimbata (Hue) Vain. And S. ciliata Taylor have been reported from Rwanda [20].
In this study, we review Sticta species and their ecology in the montane regions of Kenya and Tanzania using the barcoding nuITS genetic marker and provide a wealth of new information on their diversity and ecology.
2. Materials and Methods
2.1. Study Locations and Sampling
Specimens were collected from four mountain regions in East Africa, including the dormant volcano Mt. Kilimanjaro in Tanzania, the Taita Hills, and Mt. Kasigau, which represent the northeastern end of the Eastern Arc Mountain Range in Kenya, and Mt. Elgon at the border of Kenya and Uganda. All the mountains are less than 400 km from the equator and separated from each other by wide semiarid plains with a tropical climate with two distinct rainy seasons.
The high Mt. Kilimanjaro (5895 m) supports a wide range of natural vegetation types from natural savanna to alpine Helichrysum heath, in addition to which human activity has produced a variety of additional habitat types. The sampling in the Kilimanjaro region was done in 2016–2017 along five replicate transects on the southern and southeastern slopes of the mountain. The 65 sampling plots represent the following 13 natural and disturbed ecosystems, with 5 replicate plots in each ecosystem type: natural savanna and maize fields (800–1100 masl), lower montane forests, traditional Chagga home gardens, commercial coffee farms, and grasslands (1100–2000 masl), montane Ocotea forest and selectively logged Ocotea forest (2100─2800 masl), upper montane Podocarpus forest and secondary forest dominated by Erica excelsa as a result of repeated forest fires (2800–3100 masl), subalpine Erica trimera forest and fire disturbed E. trimera forest/shrubbery (3500–4000 masl), and alpine Helichrysum heath (4000–4650 masl). For a more detailed description of the sampled environments on Mt. Kilimanjaro, see [13,34]. On each plot, lichen specimens were collected from a 5 × 20 m central plot and along two 50 m transects. Additionally, also a larger 20 × 50 m plot was sampled for branches that had dropped from the canopy.
The Taita Hills consists of three mountain massifs: Dabida, Mbololo, and Sagalla. While the potential natural vegetation on the moist upper slopes consists of evergreen Ocotea forest, long-lasting and intensive human influence has fragmented the indigenous forest into small and often heavily disturbed, isolated patches [35,36,37]. The sampling of lichens in the Taita Hills took place during several field trips mainly in 2009–2011, encompassing all the main remaining forest fragments: On the Dabida massif, fragments of indigenous montane forest mainly occur on the highest peaks and ridges, including Ngangao (120 ha), Chawia (86 ha), Yale (16 ha), Fururu (8 ha), Macha (3 ha), Mwachora (2 ha), Vuria (<1 ha), and Shomoto Hill (<0.2 ha) [36,38]. Mt. Sagalla in the southeast harbors a small indigenous forest patch of 2 ha surrounded by plantation forest, while Mbololo in the northeast has a relatively well-preserved 185 ha moist montane forest on top of a single ridge [39]. Additionally, few specimens were collected from Maktau Hill, an isolated peak with a small patch of dry woodland vegetation, lying west of the Dabida massif. All the collection localities in the Taita Hills were situated between 1300 masl (Maktau Hill) and 2208 masl (Vuria). For a more detailed description of the sampled forest fragments especially on the Dabida massif, see [13].
Mt. Kasigau is situated approximately 50 km southeast of the Taita Hills and, unlike the highly fragmented forests of the Taita Hills, the forest and woodland on Mt. Kasigau has remained relatively intact. The vegetation includes a transition from the Acacia-Commiphora bushland on the surrounding plains in 520 masl through a lower montane woodland to an evergreen forest, reaching the summit at 1641 masl [40,41]. On Mt. Kasigau, specimens were mainly collected in 2010, along four transects corresponding to the northern, eastern, southern, and western slopes of the mountain, as described in [42]. On Mt. Elgon, specimens were collected in 2016 from the ericaceous zone of the mountain.
Local species abundances in the Mt. Kilimanjaro sampling plots were defined based on species presence on the central plot and along the two transects, the value thus ranging from 0–3 for each sampling plot. Abundances in habitat types were defined as the sum of abundances in the five sample plots representing each habitat type. In the Taita Hills, abundance was defined as the number of different forest fragments from which a species was collected (0–11); abundance was not estimated for the individual forest fragments. On Mt. Kasigau, the abundance was defined as the number of elevational transects from which the species was collected (0–4).
2.2. Morphological Inspection
In sum, 373 Sticta specimens from Mt. Kilimanjaro, the Taita Hills, Mt. Kasigau, and Mt. Elgon (Table A1) were studied and identified based on morphological characters and the previous literature from the study region [12,26]. The morphological and anatomical characters were assessed using a Leica S8AP0 stereo and an Olympus BX51 compound light microscopes, the latter equipped with a Deltapix Invenio 12EIII camera. The description and naming of characters (branching, vegetative propagules, tomentum) follows Moncada et al. [43]. Spot reactions were checked from medulla with 10% KOH (K), sodium hypochlorite solution (liquid bleach; C), and 1,4-phenylenediamine in ethanol (Pd).
2.3. DNA Sequencing
The DNA extractions were made using the GeneJET Genomic DNA Purification Kit (Thermo Fisher Scientific, Waltham, MA, USA). For the extraction, a clean piece of lichen thallus (~0.1 cm2) was selected under a preparation microscope using a sterile needle or scalpel. Amplification and sequencing of the nuclear fungal internal transcribed spacer (ITS: ITS1-5.8S-ITS2) was performed as in [44], using primers ITS1 or ITS5, and ITS4 [45]. Sequencing was performed by Macrogen Europe (Amsterdam, the Netherlands) and LGC Genomics (Berlin, Germany). Sequences were edited with CodonCode Aligner [46]. The newly obtained ITS sequences were deposited in the NCBI GenBank database [47]. The specimen information, collection locations and the GenBank accession numbers are listed in Table A1.
2.4. Phylogenetic Analyses
The generated ITS sequence dataset was complemented with sequences downloaded from the GenBank [47]. The initial alignment of the ITS was done using MAFFT on the online server [48] and adjusted by hand using PhyDE v. 0.9771 [49]. Ambiguous regions were removed from the ITS alignment before the analysis of the complete dataset resulting an alignment of 346 sequences and 602 characters.
Bayesian analyses were performed using MrBayes v.3.2.7 [50,51] on CIPRES Science Gateway [52]. To allow possible deviating substitution models for the different regions the data sets were divided in a partition of three subsets (1: ITS1; 2: 5.8S; 3: ITS2). The best fitting nucleotide substitution models were selected by jModelTest [53] using AIC and BIC, and GTR + Γ was used for ITS1, SYM + Γ for 5.8S, and GTR + I + Γ for ITS2. Posterior probability distributions of trees were calculated using the Metropolis-coupled Markov chain Monte Carlo (MCMCMC) method and the search strategies suggested by Huelsenbeck et al. [54,55]. Three runs with four chains with 10 × 106 generations each were run simultaneously. First, 25% of the trees were discarded (burnin), and the convergence of the runs confirmed with Tracer v. 1.7.1 [56] before the calculations for the 50% majority consensus tree and clade posterior probabilities (PP) were made. The trees were visualized using TreeGraph2 v2.15.0 [57].
To further analyze the phylogenetic relationships in the Sticta umbilicariiformis—fuliginosa clade, further analyses were run for selected specimens using more of the ITS region, following the same practices as in the first analysis. The alignment included 37 sequences and 495 characters with S. duplolimbata (KT281696), S. andensis (KC732547), and S. pseudolimbata (KC732564) as outgroup sequences. SYM + Γ was selected for ITS1, JC for 5.8S, and HKY + I for ITS2 as substitution models, and the analysis was run for 5 × 106 generations. The sequence alignment files and the resulting tree files from the phylogenetic analyses are available in the Zenodo repository (https://zenodo.org/) with doi 10.5281/zenodo.7575780.
3. Results
Of the studied 373 Sticta specimens, a good quality ITS sequence was obtained from 233 specimens. ITS variant information for each sequenced lichen specimen is listed in Table A1.
3.1. Phylogenetic Analyses of the Specimens
The Bayesian analysis of the nuITS region of the genus Sticta revealed that several of the morphologically identified species included representatives of more than one phylogenetic lineage (Figure S1).
3.1.1. Specimens with Soredia or Pustules
Based on the previous literature from the region [12], the sorediate specimens were identified as either S. sublimbata or S. limbata. All specimens with soredia that were collected from the lower to middle montane forest zones formed a well-supported (PP = 1) clade together with S. sublimbata specimens from Réunion and Japan (Figure 1). However, the sorediate-pustular specimens from higher elevation habitats did not group together with S. limbata, but were closely related to a previously sequenced S. umbilicariiformis specimen from Rwanda (Figure 2).
3.1.2. Fertile Specimens without Symbiotic Propagules
Three frequently fertile Sticta species that lack symbiotic propagules had previously been reported from the region, i.e., S. ambavillaria, S. kunthii, and S. tomentosa [12,26], and specimens resembling the descriptions of all these taxa were also present in our material. All specimens matching the description of S. tomentosa fell into a well-supported (PP = 1) clade, which mainly consists of S. tomentosa specimens from Colombia and Hawaii (Figure 1). All other specimens belonged to the S. umbilicariiformis—fuliginosa group (Figure 2). In the additional analysis, six specimens formed a well-supported (PP = 0.992) clade (S. munda); however, many morphologically similar specimens, i.e., fertile with pubescent or nodulous apothecial margins and with smooth, scrobiculate, foveolate to pitted upper surface, were mainly placed in the poorly resolved S. umbilicariiformis group, with some specimens in the well-supported (PP = 1) S. aspratilis clade.
3.1.3. Specimens with Laminal Isidia (Sticta fuliginosa Morphodeme)
Our specimens identified as S. fuliginosa, based on the previous literature from the region [12], fell into six different clades (Figure S1). These include three previously described species of the Sticta fuliginosa morphodeme, i.e., S. ciliata, S. fuliginoides (Figure 3), and S. fuliginosa (Figure 2). Additionally, four specimens (Sticta sp. B) representing two different ITS variants formed a well-supported (PP = 0.995) clade with one sequence obtained from a specimen from Rwanda identified as S. ciliata, forming a sister clade to S. parvilobata, a recently described species [23] from Puerto Rico (Figure 3). Additionally, three specimens (Sticta sp. A) were closely related to S. catharinae, another recently described species [22] from Bolivia (Figure 3). Nineteen specimens (S. aspratilis), mostly representing the S. fuliginosa morphodeme, but also including some fertile specimens without isidia, formed a well-supported (PP = 1) clade within the S. umbilicariiformis—fuliginosa group (Figure 2).
3.1.4. Specimens of the Sticta weigelii Morphodeme
The specimens belonging to the Sticta weigelii morphodeme, i.e., with cylindrical or flattened marginal isidia and often also with elongate lobes, were split into several different clades (Figure S1). The specimens with cylindrical isidia fell into a clade with specimens identified as S. weigelii from different parts of the world; however, the smaller clade consisting of specimens identified as S. weigelii s. str. [58] only includes GenBank sequences from the Neotropics (Figure 4).
Specimens with at least some flattened isidia were divided into three different clades. The majority fell within the S. umbilicariiformis—fuliginosa group and formed a clade (PP = 0.665) closely related to S. munda, S. umbilicariiformis, and S. aspratilis (Figure 2). One specimen was placed into a well-supported (PP = 0.992) clade comprised mainly of S. andina specimens (Figure 4), and four specimens (S. cyanocaperata) grouped together with S. caperata from Réunion and Madagascar (Figure 5).
3.1.5. Specimens with Marginal Isidia
Previously, two additional species with marginal isidia have been reported from the region, including S. cyphellulata and S. orbicularis [12]. Of the remaining marginally isidiate specimens, 18 clearly stipitate specimens formed a well-supported group with S. marginalis specimens from Réunion and Madagascar (Figure 5), and 22 formed a well-supported (PP = 1) group with S. duplolimbata specimens from other parts of the world (Figure 4). Additionally, specimens with cylindrical, mainly marginal isidia, but with an otherwise unique appearance (S. cellulosa), formed their own clade (PP = 0.868) among several recently described species mainly from the Neotropics (Figure 4). Additionally, two small and poorly developed specimens (Sticta sp. D) grouped (PP = 0.765) together with S. isidioimpressula but with a relatively long branch (Figure 4).
3.1.6. Sticta with Green Algae
Previously, two Sticta species with a green algal photobiont have been reported from East Africa: Sticta dichotoma and S. papyracea/variabilis [12]. Only four such specimens were collected by us, all resembling the description of S. papyracea. However, in the phylogenetic analysis, the sequenced specimens did not group together with S. variabilis or S. dichotoma, but formed a clade (PP = 0.854) with some specimens from Madagascar (Figure 5), identified as “Sticta sp. 2” by Simon et al. [25].
3.2. Species of Sticta Identified
According to the results of the phylogenetic analysis, our Sticta specimens represent 19 distinct species (Table A1). These include nine previously established species, S. andina, S. ciliata, S. duplolimbata, S. fuliginoides, S. fuliginosa, S. marginalis, S. sublimbata, S. tomentosa, and S. umbilicariiformis; five newly described species, S. afromontana, S. aspratilis, S. cellulosa, S. cyanocaperata, S. munda, and five putative species, Sticta sp. A (fuliginoides agg.), Sticta sp. B (ciliata agg.), Sticta sp. C (weigelii agg.), Sticta sp. D, and Sticta sp. 2. Brief descriptions of the established species and full descriptions of the novel species are provided, including a summary of observations on their ecology and distribution. All the observed species are included in the key. Sticta dichotoma is included in the key based on previous published reports from the region [12].
3.2.1. Key to Sticta Species in East Africa
- 1a.
- Main photobiont green alga—2.
- 1b.
- Main photobiont cyanobacterium—3.
- 2a.
- Thallus thin and fragile, with marginal lobules—Sticta sp. 2 (Figure 6a)
- 2b.
- Thallus robust, without marginal lobules—Sticta dichotoma
- 3a.
- Thallus with marginal to submarginal soralia (may occasionally form coralloid, isidia-resembling structures) and/or pustules—4.
- 3b.
- Thallus without soredia or pustules—5.
- 4a.
- With true, mainly marginal soredia (found in sub/lower montane habitats, mainly <2000 masl)—Sticta sublimbata
- 4b.
- With pustules that may appear sorediate (found in upper montane and subalpine habitats, >3500 masl)—Sticta umbilicariiformis
- 5a.
- Thallus without symbiotic propagules, often fertile—6.
- 5b.
- Thallus with marginal or laminal isidia and/or phyllidia—9.
- 6a.
- Apothecial margin with abundant white hairs, not nodulous; lobe margins often with projecting tufts of hair/tomentum; cyphellae urceolate with a small pore; ascospores large (>40 × 8 µm in diam.)—Sticta tomentosa
- 6b.
- Apothecial margin slightly pubescent at most, most often nodulous; cyphellae not strongly urceolate with a small pore; ascospores shorter—7.
- 7a.
- Ascospores 3-septate; lobes usually <3 cm long and wide—Sticta munda sp. nov.
- 7b.
- Ascospores 1-septate; lobes often larger—8.
- 8a.
- Ascospores 33–40 µm long—Sticta umbilicariiformis
- 8b.
- Ascospores usually shorter—Sticta aspratilis sp. nov.
- 9a.
- Dark isidia marginal, submarginal, and on scrobiculate ridges present at least near the thallus margins of the brown thallus—Sticta cellulosa sp. nov.
- 9b.
- Thallus without scrobiculate isidiate ridges—10.
- 10a.
- Isidia scattered over thallus lamina; without apothecia—11.
- 10b.
- Isidia predominantly on thallus margins (in fertile specimens often also on lamina)—14.
- 11a.
- Lobes elongate-obovate, fan-shaped, ascending from one attachment point with a funnel-like base; older thalli commonly with stalked lobules; lower side with pronounced, often clearly dome-like cyphellae (in middle montane to subalpine habitats, >2500 masl)—Sticta fuliginoides agg. (Sticta fuliginoides and Sticta sp. A)
- 11b.
- Lobes rounded and/or palmate, without a funnel-shaped base, stalked lobules not present—12.
- 12a.
- Thallus small (up to 2 cm in diam.), lobes widely rounded (wider than long) and revolute (often complete lobes becoming convex); lower side marginally etomentose with widely different sized and often flat cyphellae (in lower and middle montane habitats, <2600 masl)—Sticta ciliata agg. (Sticta ciliata and Sticta sp. B, Figure 6b)
- 12b.
- Thallus usually larger, lower side fully tomentose, cyphellae cupuliform to slightly urceolate—13.
- 13a.
- Lower side usually with abundant, arachnoid, moniliform secondary tomentum—Sticta aspratilis sp. nov.
- 13b.
- Lower side usually without arachnoid, moniliform secondary tomentum—Sticta fuliginosa
- 14a.
- Lobes palmate, clearly stipitate and ascending from one attachment point; lower side largely etomentose, often with a yellow hue; marginal isidia developing into lobules especially in older thalli—Sticta marginalis
- 14b.
- Lobes not clearly stipitate, palmate, and ascending; lower side tomentose—15.
- 15a.
- Lobes elongate, with rounded apices, often with dark marginal cilia; cyphellae distinctly raised and urceolate with a small opening (dome-like)—Sticta duplolimbata
- 15b.
- Lobe margins without dark marginal cilia; cyphellae not distinctly dome-like—16.
- 16a.
- Isidia cylindrical to coralloid, in congested dark heaps, occasionally developing into stalked lobules—Sticta sp. C (weigelii agg.) (Figure 6c)
- 16b.
- Isidia flattened and extending horizontally from the margins, usually not in congested heaps—17.
- 17a.
- Lower surface color is usually cream to light brown with light to brown tomentum (or lower surface brown with white tomentum); upper surface grey to chocolate brown—18.
- 17b.
- Lower surface dark brown (at least centrally) with dark tomentum; upper surface color is usually fawn to yellow–brown (occasionally light grey or dark brown)—19.
- 18a.
- Thallus thick and large; with marginal flattened isidia; primary tomentum of agglutinated hyphae—Sticta afromontana sp. nov.
- 18b.
- Rounded lobes with marginal and submarginal isidia and stalked lobules; lower surface at least centrally dark brown and glossy, tomentum white-light, only weakly agglutinated and hair-like, often entangled—Sticta sp. D
- 19a.
- K+ yellow; with moniliform secondary tomentum—Sticta andina
- 19b.
- K−; without moniliform secondary tomentum—Sticta cyanocaperata sp. nov.
Figure 6.
Sticta sp. 2, Sticta sp. B (ciliata agg.), Sticta sp. C (weigelii agg.), and Sticta andina. (a) Sticta sp. 2, the only green algal species collected by us, photographed in situ on Mt. Kasigau and showing the characteristic abundant and delicate marginal phyllidia (JR10K302). (b) Sticta sp. B, closely resembling S. ciliata, photographed in situ on Shomoto Hill and displaying the small, dark brown, rounded lobes and laminal isidia (JR10060). (c) Sticta sp. C photographed in situ in Sagalla forest with the characteristic congested heaps of cylindrical-coralloid isidia in the margins of the elongate lobes (JR16295). (d,e) Sticta andina (JR10117). (d) Photographed in situ on Vuria Mountain. (e) When dry, the species has a yellowish–brown upper surface and dark brown lower surface. Scale 0.5 cm in (e).
Figure 6.
Sticta sp. 2, Sticta sp. B (ciliata agg.), Sticta sp. C (weigelii agg.), and Sticta andina. (a) Sticta sp. 2, the only green algal species collected by us, photographed in situ on Mt. Kasigau and showing the characteristic abundant and delicate marginal phyllidia (JR10K302). (b) Sticta sp. B, closely resembling S. ciliata, photographed in situ on Shomoto Hill and displaying the small, dark brown, rounded lobes and laminal isidia (JR10060). (c) Sticta sp. C photographed in situ in Sagalla forest with the characteristic congested heaps of cylindrical-coralloid isidia in the margins of the elongate lobes (JR16295). (d,e) Sticta andina (JR10117). (d) Photographed in situ on Vuria Mountain. (e) When dry, the species has a yellowish–brown upper surface and dark brown lower surface. Scale 0.5 cm in (e).

3.2.2. Sticta afromontana Kaasalainen and Rikkinen sp. nov. (Figure 7)
Mycobank # MB847043
Species of Sticta lichenized with a cyanobacterium and characterized by robust thallus, flattened marginal isidia, and cream to mid–brown lower surface with moniliform secondary tomentum.
Type: Tanzania, Mt. Kilimanjaro, near the Maua Route, upper montane secondary forest with Erica excelsa, −3.1864° N 37.4403° E, 2820 masl, 11 March 2017, on a fallen branch, U. Kaasalainen UK170826e (H 9237169—holotype).
ITS barcoding marker accession (GenBank): OP999496 (holotype).
Description: Thallus rosetteform to irregular, 200–350 µm thick and up to 5 cm in diam., attached to substrate from the center of the lower side. Lobes robust, loosely adnate, elongate, palmate, up to 3 cm long and 2.5 cm wide, branching polytomous; margins crenate and crisped with abundant, mostly flattened isidia. Upper surface light grey to brown, smooth to slightly wavy to foveolate centrally, often with tufts of submarginal white hairs. Upper cortex paraplectenchymatous, 25–40 μm and 3(4) cell layers thick, the first layer(s) composed of smaller slightly flattened cells (~5 × 7 um), the others of larger (6–12 um) isodiametric cells. Photobiont Nostoc. Photobiont layer 45–85 μm thick, with Nostoc cells 5–6 μm in diam. Medulla 70–240 μm thick, with hyphae 2–4 µm wide. Cilia not present. Isidia abundantly present, mainly marginal but on fertile specimens also on lamina, grey to dark brown, glossy, coralloid, and mostly flattened and horizontal. Lower surface cream colored to (more rarely) brown, smooth or ridged, with abundant tomentum. Lower cortex paraplectenchymatous, 20–40 μm and 2–3 cell layers thick, with isodiametric cells 7–10 μm in diam. Primary tomentum usually with a brown and agglutinated stem, becoming white and squarrose towards the end. Secondary tomentum white, arachnoid, composed of moniliform assemblages. Rhizines infrequent, dark, slender. Cyphellae 45–90/cm2, cupuliform to slightly urceolate, with raised margins, pore up to 1.3 mm in diam., often with a darker ring surrounding the opening; cyphellar membrane white, 15–20 µm thick, composed of rounded, epapillose cells ~7 µm in diam. Apothecia infrequent and mainly found on specimens collected from optimal habitats; submarginal and laminal, sessile, up to 1.3 mm in diam. and 0.6 mm high (from the lower cortex of the lobe invagination); disc orange–brown; margin light brown with dark brown nodules. Exciple margin 95–115 μm wide. Epithecium 10–15 μm thick, orange–brown. Hymenium 100–120 μm thick. Hypothecium 55–70 µm thick, orange–brown. Ascospores fusiform, colorless, (1)3(5)-septate, 27–40 × 6–8 µm. Pycnidia not seen. Chemistry: K−, C−, Pd−.
Figure 7.
Sticta afromontana. (a) Type specimen UK170826e collected from the upper montane forest on Mt. Kilimanjaro. The specimen has laminal isidia that are not present in specimens without apothecia. (b) Specimen JR16366 illustrating typical habit with marginal isidia. (c) Specimen JR10112 photographed in situ on Vuria Mountain. (d) Cross section of an apothecium from the type specimen. (e,f) Thallus part of the same collection as the type imaged from the upper and lower side. Scales 0.5 cm in (a), (b), (e), and (f), 300 µm in (d).
Figure 7.
Sticta afromontana. (a) Type specimen UK170826e collected from the upper montane forest on Mt. Kilimanjaro. The specimen has laminal isidia that are not present in specimens without apothecia. (b) Specimen JR16366 illustrating typical habit with marginal isidia. (c) Specimen JR10112 photographed in situ on Vuria Mountain. (d) Cross section of an apothecium from the type specimen. (e,f) Thallus part of the same collection as the type imaged from the upper and lower side. Scales 0.5 cm in (a), (b), (e), and (f), 300 µm in (d).

Etymology: Sticta afromontana is one of the most common Sticta species in the studied afromontane region.
Ecology and distribution: Sticta afromontana is common, abundant and often fertile in the upper montane forest zone and present from lower montane forests to the ericaceous zone (1800–3510 masl) but has not been collected outside forest habitats. Sticta afromontana is particularly common on Mt. Kilimanjaro, where often the most abundant Sticta species, but also found in some Taita Hills forests. Epiphytic on tree trunks, branches, shrubs, and climbers. So far, only known from Tanzania and Kenya.
Selected specimens examined: Tanzania, Mt. Kilimanjaro, near Machame Route, lower montane forest, −3.1675° N 37.2363° E, 1920 masl, 15 March 2017, on a fallen branch, U. Kaasalainen UK170930c (H). Middle montane Ocotea forest, −3.0812° N 37.1444° E, 2260 masl, 9 March 2017, on fallen branch, U. Kaasalainen UK170792d (H). Upper montane Podocarpus forest, −3.1035° N 37.2604° E, 2850 masl, 9 March 2017, on mossy liana U. Kaasalainen UK170775b, U. Kaasalainen UK170779c. On fallen branch, U. Kaasalainen UK170781b (H). Near Marangu Route, upper montane Podocarpus forest, −3.1044° N 37.3046° E, 2800 masl, 13 March 2017, on tree trunk, U. Kaasalainen UK170886a (H). Near Maua Route, disturbed middle montane Ocotea forest, 2270 masl, on fallen branch, U. Kaasalainen UK171180a (H). Upper montane Podocarpus forest, −3.1936° N 37.4421° E, 2270 masl, 11 March 2017, on tree trunk, U. Kaasalainen UK170857a (H). On trunk of fallen tree, U. Kaasalainen UK170858g (H). Upper montane secondary forest with Erica excelsa, −3.1898° N 37.4390° E, 2880 masl, 11 March 2017, on a shrub branch, U. Kaasalainen UK170846e (H). −3.1864° N 37.4403° E, 2720 masl, 11 March 2017, on a fallen branch, U. Kaasalainen UK170806j (H). On a tree trunk, U. Kaasalainen UK170832c (H). Near the Mweka route, upper montane Podocarpus forest, −3.1659° N 37.3626° E, 2940 masl, 24 June 2017, on a fallen branch, U. Kaasalainen UK171526a (H). On a tree trunk, U. Kaasalainen UK171528b (H). Upper montane secondary forest with Erica excelsa, −3.1640° N 37.3675° E, 2990 masl, 25 June 2017, on a tree trunk, U. Kaasalainen UK171577f (H). Near the Umbwe route, middle montane Ocotea forest, −3.0824° N 37.1811° E, 2540 masl, 20 June 2017, on a tree trunk, U. Kaasalainen UK171490e (H). Kenya, Taita Hills, Vuria Mountain, −3.24° N 38.17° E, 2200 masl, J. Rikkinen JR10112 (EA), J. Rikkinen JR10121B (EA), J. Rikkinen and P. M. Kirika JR16366 (EA).
Notes: Sticta afromontana can be distinguished from other Sticta species in the region by its flattened marginal isidia and lack of K reaction (strong and immediate yellow in S. andina), presence of moniliform secondary hyphae (not present in S. cyanocaperata), and usually cream to light brown lower surface and primary tomentum (usually dark brown in S. andina and S. cyanocaperata). Sticta xanthotropa, previously reported from East Africa, has a thin and fragile thallus (robust in S. afromontana), shorter ascospores (24–36 µm), and different substrate ecology (grows on rocks and soil) [32] than S. afromontana.
3.2.3. Sticta andina B. Moncada, Lücking and Sérus. (Figure 6d,e)
A detailed description of S. andina is provided in [21]. Only one specimen of S. andina was identified from our material. It has a cyanobacterial main photobiont (Nostoc) and is characterized by flattened marginal isidia, yellowish–brown upper surface and dark brown lower surface with dark, short primary tomentum, and moniliform secondary tomentum. Chemistry: K+ yellow, C−, Pd−.
Morphologically, S. andina most resembles S. cyanocaperata; however, it can be easily identified based on the immediate, bright yellow K+ reaction in the medulla (S. andina is the only K+ species in our region). Sticta cyanocaperata also lacks the moniliform secondary tomentum present in S. andina.
Ecology and distribution: In other parts of the world (Columbia, Hawaii, Azores), S. andina has been reported to grow epiphytically in humid montane forests and in montane heathlands [21]. Our single specimen was collected from a moist montane forest on Vuria Mountain in the Taita Hills, and it shared an identical ITS sequence with a specimen previously collected from Hawaii (MT132671).
3.2.4. Sticta aspratilis Kaasalainen and Rikkinen sp. nov. (Figure 8)
Mycobank # MB847044
Species of Sticta lichenized with a cyanobacterium and characterized by its large thallus, rough upper surface with laminal isidia, abundant moniliform secondary tomentum on the lower side, and short one-septate ascospores.
Type: Kenya, Mount Elgon National Park, ericaceous zone, 2016, on a tree trunk, J. Rikkinen and P. M. Kirika JR16107 (EA—holotype).
ITS barcoding marker accession (GenBank): OP999437 (holotype).
Description: Thallus rosetteform to irregular, attached to substrate from the center of the lower side. Lobes 130–250 µm thick, loosely adnate, rarely ascending, usually rounded palmate, usually 2–4 cm wide and 2–3 cm long, but sometimes more elongate and up to 7 cm long; branching polytomous, margins entire to sinuose, sometimes slightly crisped with isidia, occasionally revolute. Upper surface grey–brown or more rarely brown, usually at least slightly glossy, uneven, ridged (isidiate specimens) and/or foveolate, with occasional eroded patches surrounded by isidia. Upper cortex paraplectenchymatous, 25–60 μm and 3–6 cell layers thick, composed of tightly packed cells of ~7 µm in diam. Photobiont Nostoc. Photobiont layer 30–75 μm thick, with Nostoc cells 5–6 μm in diam. Medulla 35–145 μm thick, with hyphae 3–4 µm wide. Cilia not present. Isidia abundantly present, laminal, and sometimes also present on the lobe margins, brown or grey, darker than the upper surface, glossy, nodular to branched or coralloid. Lower surface cream to light brown, smooth to occasionally slightly uneven, abundantly tomentose. Lower cortex paraplectenchymatous, 20–40 μm and 3–4 cell layers thick, with cells 7–10 μm in diam. Primary tomentum white to brown, composed of agglutinated hyphae. Secondary tomentum abundant, pale, arachnoid, composed of moniliform assemblages. Rhizines sparse, in scattered groups, long, slender, concolorous with primary tomentum. Cyphellae 90–270/cm2, cupuliform to slightly urceolate, with a raised margin, very variable is size, pore up to 1(2.4) mm in diam.; cyphellar membrane white, ~20 µm thick, with rounded, epapillose cells ~8 µm in diam. Apothecia occasional, only seen on specimens without isidia; submarginal and laminal, up to 2.7 mm wide; margin beige to brown with darker brown stripes or nodules, sometimes slightly pubescent or tomentose; disc red–brown. Exciple margin 120–150 μm wide. Epithecium 10–20 µm thick, orange–brown. Hymenium 90–140 µm thick. Hypothecium 50−75 µm thick, orange–brown. Ascospores fusiform, 1-septate, colorless, (21)26–33(35) × 5.5–8 µm in diam. Pycnidia not seen. Chemistry: K−, C−, Pd−.
Figure 8.
Sticta aspratilis. (a–d) Type specimen JR16107. (a) Photographed in situ in the ericaceous zone on Mt. Elgon. (b) The uneven and isidiate upper surface. (c) Abundantly tomentose lower surface. (d) Thallus cross section with a cyphella, showing the cyphellar membrane with rounded cells without papillae. (e) Specimen JR10155 photographed in situ on Yale, with gray and distinctly reticulate upper surface. (f,g) Fertile specimen UK171478a. (f) Foveolate upper surface with apothecia. (g) Cross section of an apothecium. Scales 0.5 cm in (b) and (c), 100 µm in (d) and (g), 1 cm in (f).
Figure 8.
Sticta aspratilis. (a–d) Type specimen JR16107. (a) Photographed in situ in the ericaceous zone on Mt. Elgon. (b) The uneven and isidiate upper surface. (c) Abundantly tomentose lower surface. (d) Thallus cross section with a cyphella, showing the cyphellar membrane with rounded cells without papillae. (e) Specimen JR10155 photographed in situ on Yale, with gray and distinctly reticulate upper surface. (f,g) Fertile specimen UK171478a. (f) Foveolate upper surface with apothecia. (g) Cross section of an apothecium. Scales 0.5 cm in (b) and (c), 100 µm in (d) and (g), 1 cm in (f).

Etymology: The specific epithet refers to the characteristic uneven roughness of the upper thallus surface.
Ecology and distribution: Sticta aspratilis is relatively common, but not very abundant in any habitat type. It has a wide elevational range on Mt. Kilimanjaro, the Taita Hills, and Mt. Elgon, extending from lower montane forests to the subalpine zone (1450–3720 masl). In addition to primary forests, it has also been collected from disturbed habitats. Epiphytic, mainly on canopy branches, but also on tree trunks at more open sites, occasionally also on soil in the subalpine zone. So far, S. aspratilis is only known from Kenya and Tanzania.
Selected specimens examined: Tanzania, Mt. Kilimanjaro, coffee farm in Maua, −3.1630° N 37.2813° E, 1660 masl, 23 May 2017, on coffee bush, U. Kaasalainen UK170975f (H). Near Machame Route, upper montane Podocarpus forest, −3.1035° N 37.2604° E, 2970 masl, 9 March 2017, on fallen branch, U. Kaasalainen UK170781c (H). Near Marangu Route, upper montane Podocarpus forest, −3.1044° N 37.3046° E, 2800 masl, 13 March 2017, on tree trunk, U. Kaasalainen UK170892c (H). On fallen branch, U. Kaasalainen UK170896b (H). Near Mweka Route, middle montane Ocotea forest near Mweka Route, −3.1722° N 37.3583° E, 2850 masl, 24 June 2017, on a fallen branch, U. Kaasalainen UK171515b (H). Upper montane Podocarpus forest, −3.1616° N 37.3632° E, 2970 masl, 26 June 2017, on a fallen tree, U. Kaasalainen UK171587j (H). Disturbed subalpine Erica vegetation, −3.1339° N 37.3702° E, 3720 masl, 25 June 2017, on shrub base, U. Kaasalainen UK171562c (H). Near Umbwe Route, middle montane Ocotea forest, −3.0819° N 37.1819° E, 2650 masl, 20 June 2017, on fallen branch, U. Kaasalainen UK171478a (H). Kenya, Taita Hills, field edge near the Fururu Forest, −3.25° N 38.20° E, 1650 masl, 20 January 2011, on tree trunk, U. Kaasalainen UK110551f (EA), UK110551g (EA). Shomoto Hill, −3.395° N 38.360° E, 1500 masl, 2010, epiphytic, J. Rikkinen JR10044A (EA), JR10057 (EA). Yale, −03.24° N 38.20° E, 1850 masl, 2010, epiphytic, J. Rikkinen JR10155A (EA), JR10155B (EA), JR10155C (EA), JR10171 (EA).
Notes: Sticta aspratilis most closely resembles S. fuliginosa which, however, usually lacks the moniliform secondary tomentum that characteristically covers the lower surface of S. aspratilis. Fertile specimens may resemble S. kunthii, previously reported from East Africa, and fertile specimens of S. umbilicariiformis. However, the upper surface of S. kunthii is marbled with maculae and papillate [32], while S. umbilicariiformis usually has longer ascospores (33–40 µm) than S. aspratilis.
3.2.5. Sticta cellulosa Kaasalainen sp. nov. (Figure 9)
Mycobank # MB847045
Species of Sticta lichenized with a cyanobacterium and characterized by its thick, brown lobes and tomentum, and the scrobiculate, isidiate ridges on the upper surface present at least near the margins.
Type: Tanzania, Mt. Kilimanjaro, near Umbwe Route, subalpine Erica trimera forest, −3.1114° N 37.3183° E, 3500 masl, 18 June 2017, on fallen branch, U. Kaasalainen UK171406e (H9237170—holotype).
ITS barcoding marker accession (GenBank): OP999548 (holotype).
Description: Thallus irregular, 150–300 µm thick, attached to substrate from the lower side of thallus. Lobes loosely adnate to ascending, elongate or rarely palmate, up to 3 cm wide and 4 cm long, branching polytomous, margins entire to sinuose, usually abundantly isidiate. Upper surface middle to dark brown or occasionally lighter yellow–brown, moderately glossy, smooth to foveolate centrally, with a scrobiculate pattern of isidiate ridges and hollows, at least submarginally, but occasionally spreading over a large part of the upper surface. Upper cortex paraplectenchymatous, 30–50 μm and 3–5 cell layers thick, cells up to 15 µm in diam., the first layer of cells often smaller and brownish in color. Photobiont Nostoc. Photobiont layer 20–70 μm thick, with Nostoc cells approximately 9 μm in diam. Medulla 50–180 μm thick, with hyphae 2–4 µm wide. Cilia not present. Isidia abundantly present and congested, marginal, submarginal, and on the laminal scrobiculate ridges, dark brown to almost black, glossy, cylindrical to coralloid, occasionally present also on the lower surface where grey. Lower surface dark brown, occasionally lighter towards margins, smooth, thickly and densely tomentose throughout. Lower cortex paraplectenchymatous, brown, 25–70 μm and 3–5 cell layers thick, with cells 6–14(20) μm in diam. Primary tomentum dark brown to almost black, composed of agglutinated hyphae. Secondary tomentum arachnoid, pale, composed of moniliform assemblages. Rhizines often present in small, scattered groups, clearly longer than tomentum, fasciculate, squarrose, dark brown and often with white tips. Cyphellae 13–50/cm2, cupuliform with a round pore and raised margins, larger may be more irregular and slightly urceolate, pore size very variable, up to 2.3 mm in diam.; cyphellar membrane cream-colored to slightly brown or yellow, 15–25 µm thick, composed of rounded, epapillose cells 5–12 µm in diam. Apothecia or pycnidia not seen. Chemistry: K−, C−, Pd−.
Figure 9.
Sticta cellulosa. (a,b) Type specimen UK171406e imaged from the upper and lower side. (c) Thallus part of the same collection as type specimen with abundant characteristic isidiate ridges. (d) Thallus cross section with a cyphella on the right showing the epapillate cells of the cyphellar membrane, and the moniliform assemblages which form secondary tomentum on the lower surface (UK171340t). Scales 0.5 cm in (a–c), 100 µm in (d).
Figure 9.
Sticta cellulosa. (a,b) Type specimen UK171406e imaged from the upper and lower side. (c) Thallus part of the same collection as type specimen with abundant characteristic isidiate ridges. (d) Thallus cross section with a cyphella on the right showing the epapillate cells of the cyphellar membrane, and the moniliform assemblages which form secondary tomentum on the lower surface (UK171340t). Scales 0.5 cm in (a–c), 100 µm in (d).

Etymology: The specific epithet refers to the characteristic reticulate pattern of the upper thallus surface.
Ecology and distribution: Sticta cellulosa seems to be rare and is found mainly in the subalpine zone on Mt. Kilimanjaro (2990–3520 masl). It occurs in primary and secondary (fire disturbed) Erica trimera forests and in the secondary upper montane forest with Erica excelsa. Epiphytic especially on Erica. So far, only known from Mt. Kilimanjaro, Tanzania.
Selected specimens examined: Tanzania, Mt. Kilimanjaro, near Machame Route, fire disturbed subalpine Erica vegetation, −3.0854° N 37.2794° E, 3520 masl, 15 June 2017, on Erica branches, U. Kaasalainen UK171340t (H). Near Mweka Route, fire disturbed upper montane Podocarpus forest now dominated by Erica excelsa, −3.1640° N 37.3675° E, 2990 masl, 25 June 2017, on tree trunk, U. Kaasalainen UK171584k (H). Near Umbwe Route, subalpine Erica trimera forest, −3.1114° N 37.3183° E, 3500 masl, 18 June 2017, on fallen branch, U. Kaasalainen UK171407j (H). On Erica, U. Kaasalainen UK171458k (H).
Notes: Well-developed specimens are easy to distinguish from other Sticta species based on their robust, often dark brown appearance, the isidiate scrobiculation at least along lobe margins, and thick dark brown tomentum of the lower surface. Poorly developed specimens may resemble other isidiate species with yellowish–brown upper surface, but can be distinguished on the basis of submarginal, cylindrical isidia.
3.2.6. Sticta ciliata Tayl. (Figure 10a)
Sticta ciliata has a cyanobacterial main photobiont (Nostoc) and rounded lobes with laminal isidia. A detailed description of S. ciliata is provided in [20]. In our region, the two sequenced specimens of S. ciliata have small, approximately 1–2 cm wide and 0.5–1.5 cm long, loosely adnate, rounded, obovate lobes with abundant, laminal isidia on the grey upper surface. The lobes are revolute making them convex. The lower surface is pale with a tomentose base, but with the tomentum becoming scarce or absent towards the margins, and with flat, variably sized cyphellae. Lobe margins are often ciliate. Chemistry: K−, C−, Pd−.
In our region, four other species with laminal isidia are present: Sticta aspratilis, S. fuliginoides, S. fuliginosa, and Sticta sp. B. Our S. ciliata specimens are rather few and poorly developed, but Magain and Sérusiaux [20] describe their diagnostic characters: Fresh specimens of S. ciliata have delicate and usually ciliate thallus margins, especially in young thalli, and abundant tiny papillae over the cells of the cyphellar membrane, however, the regeneration lobules of all other isidiate species can also have marginal cilia, and the fine anatomy of cyphellae can only be studied from fresh and well-preserved material. In our material, S. fuliginosa and S. aspratilis usually have larger thalli with a tomentose lower surface and cupuliform to slightly urceolate cyphellae. Sticta fuliginoides has a funnel-shaped base and usually occurs on higher elevations than S. ciliata. Sticta sp. B closely resembles S. ciliata in morphology and occurs in similar habitats. However, the material presently available is too scarce to allow a detailed morphological analysis to compare the two species.
Ecology and distribution: In other parts of the world, S. ciliata is known from Europe, Macaronesia, and possibly Colombia, where it grows as an epiphyte on tree trunks and on bryophytes, especially in well-preserved, humid forests [20]. The two confirmed specimens in our material were collected from moist montane forest, one from the Sagalla Mountain in the Taita Hills and one from the lower montane forest of Mt. Kilimanjaro. Both specimens were growing epiphytically on tree trunks. Sticta sp. B seems to be widely distributed in East Africa and has been collected from Kenya, Tanzania, and Rwanda, from similar moist lower to middle montane forests as S. ciliata.
Figure 10.
Sticta ciliata, Sticta duplolimbata, Sticta fuliginoides, Sticta fuliginosa, and Sticta marginalis. (a) Sticta ciliata from Chawia Forest with small revolute lobes and ciliate margins (UK110570a). (b) Sticta duplolimbata photographed in situ on Vuria Mountain, showing the elongate lobes with rounded apices and marginal cilia and isidia, and abundantly tomentose lower surface. (c) Sticta fuliginoides from an upper montane forest on Mt. Kilimanjaro with palmate lobes and a funnel-like base (UK171468d). (d,e) Sticta fuliginosa photographed in situ in the Taita Hills. (d) Specimen JR16358 with rounded lobes and abundant laminal isidia. (e) Specimen JR09A46 showing the white and abundantly tomentose lower surface. (f–h) Sticta marginalis photographed in situ in the Taita Hills. (f) Specimen JR29502 with stipitate, ascending lobes and marginal isidia. (g) Note the characteristic etomentose lower surface with inconspicuous cyphellae. (h) Delicate marginal isidia and stalked lobules occur commonly especially in ageing thalli.
Figure 10.
Sticta ciliata, Sticta duplolimbata, Sticta fuliginoides, Sticta fuliginosa, and Sticta marginalis. (a) Sticta ciliata from Chawia Forest with small revolute lobes and ciliate margins (UK110570a). (b) Sticta duplolimbata photographed in situ on Vuria Mountain, showing the elongate lobes with rounded apices and marginal cilia and isidia, and abundantly tomentose lower surface. (c) Sticta fuliginoides from an upper montane forest on Mt. Kilimanjaro with palmate lobes and a funnel-like base (UK171468d). (d,e) Sticta fuliginosa photographed in situ in the Taita Hills. (d) Specimen JR16358 with rounded lobes and abundant laminal isidia. (e) Specimen JR09A46 showing the white and abundantly tomentose lower surface. (f–h) Sticta marginalis photographed in situ in the Taita Hills. (f) Specimen JR29502 with stipitate, ascending lobes and marginal isidia. (g) Note the characteristic etomentose lower surface with inconspicuous cyphellae. (h) Delicate marginal isidia and stalked lobules occur commonly especially in ageing thalli.

3.2.7. Sticta cyanocaperata Kaasalainen sp. nov. (Figure 11)
Mycobank # MB847046
Species of Sticta lichenized with a cyanobacterium and characterized by its light grey to fawn, wrinkled upper surface, flattened marginal isidia, and brown, ridged (visible in thallus cross-section) lower surface without moniliform secondary tomentum.
Type: Tanzania, Mt. Kilimanjaro, near the Umbwe Route, middle montane Ocotea forest −3.0819° N 37.1819° E, 2650 masl, 20 June 2017, on a fallen branch, U. Kaasalainen UK171480d (H 9237171—holotype).
ITS barcoding marker accession (GenBank): OP999563 (holotype).
Description: Thallus rosetteform, (140)200–470 µm thick with thickenings on the lower side clearly visible in the cross-section, closely adnate centrally, loosely adnate marginally. Lobes elongate, polytomously branching, up to 5.5 cm long and 2 cm wide; margins often crisped, with darker, mostly flattened isidia, occasionally also with phyllidia. Upper surface is usually fawn to yellowish brown, occasionally light grey wavy to slightly wrinkled at least centrally. Upper cortex paraplectenchymatous, 30–70 μm and 4–6 cell layers thick, cells 6–13 µm in diam., the first layer(s) more compact. Photobiont Nostoc. Photobiont layer 30–90 μm thick, with Nostoc cells 5–7 μm in diam. Medulla 75–300 μm thick, with hyphae 3–4 µm wide. Cilia not present. Isidia marginal, usually dark brown, glossy, coralloid, flattened. Lower surface dark brown at least centrally, may become lighter towards margins, with ridges (may not always be visible under a preparation microscope, but present at least as thickenings in a thallus cross-section), with brown tomentum, lighter margins may be etomentose. Lower cortex paraplectenchymatous, (10)30–45 μm and (2)3 cell layers thick, cells 7–12 μm in diam. Primary tomentum usually dark brown, composed of agglutinated hyphae, may become entangled and matted centrally, and/or resemble arachnoid secondary tomentum. Moniliform secondary tomentum not present. Rhizines scattered in groups, dark brown to black, glossy, often with long, tapering, white tip, often hirsute from the lower parts. Cyphellae urceolate with a wide opening (up to 1 mm in diam.), pore margin raised or flat, occasionally thickened; cyphellar membrane light yellow, ~20 μm thick, cells rounded, epapillose, 6–7 μm in diam. Apothecia or pycnidia not seen. Chemistry: K−, C−, Pd−.
Figure 11.
Sticta cyanocaperata. (a) A fragment of the type specimen UK171480d. (b,c) Additional thalli part of the same collection as the type: (b) Showing the brown tomentum and pale thallus margins. (c) Characteristic light yellowish–brown, wrinkled upper surface and dark, flattened, marginal isidia. (d) Thallus cross section showing the uneven lower surface (UK170912a) and the lack of moniliform secondary tomentum. Scales 0.5 cm in (a–c), 100 µm in (d).
Figure 11.
Sticta cyanocaperata. (a) A fragment of the type specimen UK171480d. (b,c) Additional thalli part of the same collection as the type: (b) Showing the brown tomentum and pale thallus margins. (c) Characteristic light yellowish–brown, wrinkled upper surface and dark, flattened, marginal isidia. (d) Thallus cross section showing the uneven lower surface (UK170912a) and the lack of moniliform secondary tomentum. Scales 0.5 cm in (a–c), 100 µm in (d).

Etymology: The specific epithet refers to the cyanobacterial primary photobiont and characteristic wrinkles on the upper and lower surfaces, and to the close phylogenetic affiliation to Sticta caperata.
Ecology and distribution: Sticta cyanocaperata is common in the middle montane forests on Mt. Kilimanjaro (2270–2650 masl). Epiphytic on canopy branches, tree trunks, and lianas. So far, S. cyanocaperata is only known from Mt. Kilimanjaro, Tanzania.
Selected specimens examined: Tanzania, Mt. Kilimanjaro, near Marangu Route, disturbed middle montane Ocotea forest, −3.1207° N 37.3057° E, 2370 masl, 14 March 2017, on fallen branch, U. Kaasalainen UK170912a (H). U. Kaasalainen UK170912e (H). Near Maua Route, disturbed middle montane Ocotea forest, −3.1319° N 37.2717° E, 2270 masl, 29 May 2017, on liana, U. Kaasalainen UK171182c (H). Near Umbwe Route, middle montane Ocotea forest, −3.0824° N 37.1811° E, 2540 masl, 20 June 2017, on tree trunk, U. Kaasalainen UK171495f (H).
Notes: In shady habitats, S. cyanocaperata may have a rather different appearance in having a light grey upper surface, thin thallus, few isidia, and poorly developed lower side tomentum. However, such specimens can still be identified on the basis of the characteristic lower side ridges and lack of moniliform secondary tomentum. The closely related S. caperata most commonly has a green algal primary photobiont. The cyanomorph of S. caperata reported from Réunion [25] differs from S. cyanocaperata in having a marbled upper surface. Sticta xanthotropa, previously reported from East Africa [12], is distinguished for example based on the pale cream-colored to yellowish lower surface [32]. The most closely resembling species in the region with similar flattened marginal isidia and yellow–brown upper surface is S. andina which can be recognized due to the instant and strong K+ yellow reaction of the medulla.
3.2.8. Sticta duplolimbata (Hue) Vain. (Figure 10b)
Sticta duplolimbata has a cyanobacterial main photobiont (Nostoc) and marginal isidia. A detailed description of the species is provided in [59]. In our region, the species is characterized by loosely adnate, elongate, and most often light grey lobes with rounded apices, dark, mainly marginal cylindrical to coralloid isidia, and dark marginal cilia which, however, are not always present. The lower side is abundantly tomentose, with tomentum reaching the lobe margins, and usually pale but sometimes even dark brown. Cyphellae are conspicuous, raised, and distinctly urceolate with a small opening. Chemistry: K−, C−, Pd−.
Sticta duplolimbata is easy to distinguish from the other Sticta species in our region, especially by the dark cilia usually present at the rounded lobe apices and distinct, urceolate cyphellae. Galloway [59] mentions that S. duplolimbata has “lobes arising from short stalk”; however, this is not visible in most of our specimens, which are usually attached to their substrate by the tomentum of the central parts of the lower surface.
Ecology and distribution: In other parts of the world, S. duplolimbata is mainly known from the Western Pacific region [59]. Abundant on tree trunks, climbers, and canopy branches in lower and middle montane forests on Mt. Kilimanjaro, with fewer specimens from upper montane forests, Chagga home gardens, and moist montane forests of the Taita Hills (1800–3060 masl).
3.2.9. Sticta fuliginoides Magain and Sérus. (Figure 10c)
Sticta fuliginoides has a cyanobacterial main photobiont (Nostoc) and obovate lobes with laminal isidia. A detailed description of the species is provided in [20]. In our region, S. fuliginoides is characterized by the obovate lobes, ascending from a single attachment point. The lobes are usually up to 2.5 cm in diam and have a funnel-like base. The upper surface is grey, brown or yellowish–brown, smooth to strongly reticulate especially in ageing thalli, and with dark, mainly laminal, cylindrical to coralloid isidia, often developing into stalked lobules. The lower surface is white or beige, with abundant tomentum near the attachment point, but often etomentose towards the margins. Cyphellae are variable in size, usually urceolate and dome-like and raised especially towards the thallus margin, and if cupuliform, with a distinctly raised margin. Chemistry: K−, C−, Pd−.
Poorly developed thalli and/or specimens collected from suboptimal habitats may often have only one obovate-palmate ascending lobe with laminal isidia, and a mostly etomentose lower surface with pronounced cyphellae. In well-developed thalli, the ascending lobes usually have a funnel-shaped base quite distinct from those of other Sticta species with laminal isidia. Based on our phylogenetic analysis, three specimens, closely resembling S. fuliginoides in overall morphology, represent an additional undescribed taxon Sticta sp. A (Figure 3). However, the material presently at hand is insufficient for properly assessing the morphological characteristics of that species.
Ecology and distribution: In other parts of the world (Europe, Macaronesia, North America, Colombia) S. fuliginoides grows on mossy trees and rocks in humid forests and parks [20]. In our region it is an abundant epiphyte on tree trunks, canopy branches, and climbers, especially in upper montane forests on Mt. Kilimanjaro, but occurring in middle montane to subalpine habitats as well (2470–3520 masl).
3.2.10. Sticta fuliginosa (Hoffm.) Ach. (Figure 10d,e)
Sticta fuliginosa has a cyanobacterial main photobiont (Nostoc) and laminal isidia. A detailed description of the species and a discussion of differences between S. fuliginosa, S. ciliata, and S. fuliginoides are provided in [20]. In our region, S. fuliginosa is characterized by rounded palmate-obovate lobes, usually up to 5 cm in diam., with an uneven, most often greyish–brown upper surface and darker laminal isidia. The lower surface is pale with cupuliform to slightly urceolate cyphellae, and with a pale primary tomentum, but usually without a moniliform secondary tomentum. Chemistry: K−, C−, Pd−.
Of the other species with laminal isidia in our region, S. fuliginosa mostly resembles S. aspratilis which, however, has a well-developed arachnoid secondary tomentum on the lower surface, often making also the primary tomentum appear “furry”. Small thalli of S. fuliginosa often have dark brown, rounded lobes with ciliate and occasionally revolute margins very similar to those of S. ciliata agg. Sticta ciliata is usually much smaller and according to Magain and Sérusiaux [20], fresh specimens of S. ciliata and S. fuliginoides both have abundant papillae on the cells of cyphellar membrane, which do not occur in S. fuliginosa.
Ecology and distribution: Sticta fuliginosa is believed to be widely distributed in both hemispheres [20,60]. In our region, it is common in the Taita Hills forests and occurs on Mt. Kasigau. On Mt. Kilimanjaro it is common especially on canopy branches in middle montane forests but is also found in the lower montane to upper montane zones, including the Chagga home gardens (1840–2880 masl).
3.2.11. Sticta marginalis Bory (Figure 10f–h)
Sticta marginalis has a cyanobacterial main photobiont (Nostoc) and marginal isidia that often develop into stalked lobules. Thallus lobes are clearly stipitate, palmate, and ascending from a single attachment point. The upper surface is smooth, usually light grey with a yellow tinge. Isidia are dark, mainly marginal, cylindrical to coralloid, and often develop into characteristic stalked lobules especially in ageing thalli. Lower surface is light to dark brown, often with some yellow coloring, usually without tomentum or with a limited amount of short velvety hair; cyphellae are small and flat. Apothecia are not present in our material, but in the description of S. marginalis from Réunion, the apothecia are described submarginal and the ascospores brown, 1–3-septate, 40–48 × 8 µm [61]. Chemistry: K−, C−, Pd−.
Sticta marginalis is very characteristic looking and easily distinguished from the other Sticta species in the region based on the stipitate, palmate, ascending lobes with isidiate-lobulate margins, mostly naked lower surface, and flat cyphellae.
Ecology and distribution: In other parts of the world, Sticta marginalis is known from its type location Réunion and from Madagascar [61,62,63]. In our region, S. marginalis often grows as an epiphyte on tree trunks, but occasionally also on decaying wood and rock. It is common in moist lower and middle montane forests of Mt. Kilimanjaro, the Taita Hills, and Mt. Kasigau (1450–2470 masl).
Note: Sticta marginalis was described from Réunion [61] and has not previously been reported from the African continent. Another stipitate species S. orbicularis, originally described from Java in Indonesia [64], has previously been reported to occur in East Africa [12,28,30]. Sticta marginalis has even been suggested to be synonymous to S. orbicularis [65], and also Swinscow and Krog [12] noted that the taxon needs critical study and that S. pusilla Meissner may be its correct name. No DNA data is available from S. orbicularis and a description of the type material mentions that the material consists of only two immature specimens [64].
3.2.12. Sticta munda Kaasalainen sp. nov. (Figure 12)
Mycobank# MB847047
Species of Sticta lichenized with a cyanobacterium and characterized by palmate lobes with light grey upper surface, lack of symbiotic propagules, and submarginal apothecia with nodular margins and 3-septate, 30–40 µm long ascospores.
Type: Tanzania, Mt. Kilimanjaro, near the Mweka Route, upper montane secondary forest with Erica excelsa, −3.1640° N 37.3675° E, 2990 masl, 25 June 2017, on a tree trunk, U. Kaasalainen UK171584u (H 9237172—holotype).
ITS barcoding marker accession (GenBank): OP999600 (holotype).
Description: Thallus loosely adnate, often rosetteform, (120)150–180(210) µm thick and up to 5(7) cm in diam., attached from the center of the lower side or, more often, consisting of a single ascending, palmate lobe attached to substrate from its base. Lobes relatively thin, palmate, 1.5–3(4) cm in diam., usually isodiametric or wider than long, with rounded margins. Upper surface light grey or occasionally brownish towards margins, smooth to slightly wavy or foveolate. Upper cortex paraplectenchymatous, (20)25–35(50) μm and 3–4(6) cell layers thick, composed of isodiametric cells 7–11 μm in diam., the cells in the first layer(s) occasionally slightly flattened and/or smaller. Photobiont Nostoc. Photobiont layer (30)35–50(70) μm thick, with Nostoc cells 5–8 μm in diam. Medulla (35)40–75 μm thick, with hyphae 2–4 µm wide. Cilia, isidia, soredia, and phyllidia absent. Lower surface cream colored to light brown, smooth, with abundant tomentum. Lower cortex paraplectenchymatous, 15–35(40) μm and 2–3(4) cell layers thick, with isodiametric to slightly oblong cells 6–13 μm in diam. Primary tomentum with brown and agglutinated stems, becoming lighter and squarrose towards the tips. Secondary tomentum white, arachnoid, composed of moniliform assemblages. Rhizines not present. Cyphellae (40)50–200/cm2, cupuliform and rounded with a clearly raised margin when small to medium sized, irregular and slightly urceolate when large; pore rarely >0.8 mm in diam., often surrounded by a darker ring; cyphellar membrane white, cells rounded, epapillose, 6–9 μm in diam. Apothecia common, submarginal to laminal, up to 2 mm in diam.; disc light reddish to dark brown; margin light brown with darker brown nodules. Exciple margin 120–140 μm wide. Epithecium 10–15(30) μm thick, orange–brown. Hymenium (80)90–110 μm thick. Hypothecium (45)80–95 μm thick, orange–brown. Ascospores fusiform, colorless, 3-septate, 30–38(43) × (5)6–9 µm. Pycnidia not seen. Chemistry: K−, C−, Pd−.
Figure 12.
Sticta munda. (a–c) Type specimen UK171584u. (a,b) Typical single lobed, palmate, and abundantly fertile specimen seen from the upper and lower side. (c) A cross section of apothecium. (d) Specimen UK171582i with a rosetteform thallus. Scales 0.5 cm in (a), (b), and (d), 100 µm in (c).
Figure 12.
Sticta munda. (a–c) Type specimen UK171584u. (a,b) Typical single lobed, palmate, and abundantly fertile specimen seen from the upper and lower side. (c) A cross section of apothecium. (d) Specimen UK171582i with a rosetteform thallus. Scales 0.5 cm in (a), (b), and (d), 100 µm in (c).

Etymology: The specific epithet refers to the simple and elegant form of the species.
Ecology and distribution: Sticta munda is quite rare in the middle and upper montane forests on Mt. Kilimanjaro (2220–2990 masl). Epiphytic on canopy branches and tree trunks. So far, only known from Mt. Kilimanjaro, Tanzania.
Selected specimens examined: Tanzania, Mt. Kilimanjaro, near the Marangu route, upper montane Podocarpus forest, −3.1044° N 37.3046° E, 2800 masl, 13 March 2017, on a fallen branch, U. Kaasalainen UK170888c (H). Near the Mweka route, −3.1101° N 37.2130° E, 2470 masl, 24 June 2017, on a fallen branch, U. Kaasalainen UK171508j (H), U. Kaasalainen UK171510k (H). Upper montane secondary forest with Erica excelsa, −3.1640° N 37.3675° E, 2990 masl, 25 June 2017, on a tree trunk, U. Kaasalainen UK171582i (H). Near the Umbwe route, disturbed middle montane Ocotea forest, −3.0903° N 37.1724° E, 2220 masl, 20 June 2017, on a fallen branch, U. Kaasalainen UK171497d (H).
Notes: The six specimens of S. munda sequenced represented three different ITS variants. Although all the specimens formed a well-supported monophyletic clade, there are some morphological differences between specimens representing the different ITS variants. Sticta munda resembles S. ambavillaria from Réunion, which, however, is distinguished by the phylogenetic analysis (Figure 2). Sticta ambavillaria also has longer ascospores ((42)44.5–50 μm [33]) than S. munda, which was apparently also noticed by Swinscow and Krog [12] who reported short ascospores (30–40 × 6–10 µm) for their S. ambavillaria specimens collected from East Africa. Additionally, fertile specimens of S. umbilicariiformis and S. aspratilis can resemble S. munda, but they have one-septate ascospores and thicker lobes, which are often brown instead of grey and more distinctly foveolate.
3.2.13. Sticta sublimbata (J. Steiner) Swinscow and Krog (Figure 13a,b)
Sticta sublimbata has a cyanobacterial main photobiont (Nostoc) and marginal soralia. A detailed description of East African material is provided in [12]. The rosetteform thallus has adnate or loosely adnate, elongate more or less linear and narrow (usually < 1 cm wide) lobes with rounded apices that are often also revolute when dry. The upper surface is usually light leather brown to grey but may also be dark brown. The lower surface is light to dark brown, and usually has scarce, short tomentum, but also densely tomentose forms are quite common. Cyphellae are white with open, raised margins. Marginal linear soralia are almost always present and produce farinose to granular soredia, and sometimes also form aggregates resembling isidia. Apothecia not seen. Chemistry: K−, C−, Pd−.
In our region, S. sublimbata is the only sorediate species in lower montane forests and woodland below 2500 masl. Sticta umbilicariiformis, which is common in the upper montane and subalpine zones, can occasionally appear sorediate, but usually has much thicker and wider (several centimeters wide) lobes and an abundantly tomentose lower surface.
Ecology and distribution: In addition to Africa, S. sublimbata is known from Australia, New Zealand, and southern South America [59]. In our region, S. sublimbata is an abundant and common epiphyte especially on tree trunks, but it also grows among bryophytes on cliffs and other rock surfaces. Sticta sublimbata is especially common in the lower montane forests of the Taita Hills, but also occurs on Mt. Kasigau and Mt. Kilimanjaro, mainly below 2000 masl. It may even benefit from human activity as it seems most abundant in moderately disturbed habitats.
Figure 13.
Sticta sublimbata, Sticta tomentosa, and Sticta umbilicariiformis. (a,b) Sticta sublimbata. (a) Large and adnate thallus of specimen JR16275 photographed in situ on Sagalla Mountain. (b) Specimen JRCA2 photographed in situ in Ngangao Forest displaying characteristic rounded lobes and marginal soralia. (c,d) Sticta tomentosa photographed in situ on Vuria Mountain. (c) Specimen JR16368 with palmate lobes and abundant apothecia. (d) Specimen JRA14357 showing the lobe margins with tufts of hair, apothecia with white hairs on the margin, and white lower surface with prominent cyphellae. (e–g) Sticta umbilicariiformis. (e) Specimen JR16102 photographed in situ in the ericaceous zone on Mt. Elgon with characteristic pustules on thallus margins and submarginal lamina. (f,g) Fertile specimen UK171411c with strongly foveolate upper surface and tomentose apothecial margins. Scales 0.5 mm in (f), 1 cm in (g).
Figure 13.
Sticta sublimbata, Sticta tomentosa, and Sticta umbilicariiformis. (a,b) Sticta sublimbata. (a) Large and adnate thallus of specimen JR16275 photographed in situ on Sagalla Mountain. (b) Specimen JRCA2 photographed in situ in Ngangao Forest displaying characteristic rounded lobes and marginal soralia. (c,d) Sticta tomentosa photographed in situ on Vuria Mountain. (c) Specimen JR16368 with palmate lobes and abundant apothecia. (d) Specimen JRA14357 showing the lobe margins with tufts of hair, apothecia with white hairs on the margin, and white lower surface with prominent cyphellae. (e–g) Sticta umbilicariiformis. (e) Specimen JR16102 photographed in situ in the ericaceous zone on Mt. Elgon with characteristic pustules on thallus margins and submarginal lamina. (f,g) Fertile specimen UK171411c with strongly foveolate upper surface and tomentose apothecial margins. Scales 0.5 mm in (f), 1 cm in (g).

3.2.14. Sticta tomentosa (Sw.) Ach. (Figure 13c,d)
Sticta tomentosa has a cyanobacterial main photobiont (Nostoc) and palmate lobes without symbiotic propagules, but usually with apothecia. A detailed description of S. tomentosa is provided in [66] and, based on East African material, in [12]. Our material of S. tomentosa have light grey, palmate lobes ascending clearly from one attachment point, and with tufts of hairs projecting from the lobe margins. The lower surface is white with abundant white tomentum and has prominent, dome-like cyphellae. Sticta tomentosa does not produce isidia, soredia nor phyllidia, but is almost always fertile. The apothecia are mainly submarginal with long white hairs on the margins. Ascospores are fusiform, colorless, 3-septate, 39–50 × 7–11 µm (30–50 × 6–10 µm in [12]). Chemistry: K−, C−, Pd−.
Sticta tomentosa is the only species in the studied lower montane forests that is commonly fertile and does not produce any type of symbiotic propagules. Easily distinguished from other fertile species in the region based on the thick white hairs on apothecial margin. The apothecial margins of other species are pubescent at most, and this feature is usually only seen in specimens collected from high-elevation habitats. Furthermore, the three-septate ascospores of S. tomentosa are larger than those of any other species in the region. The ascospores in the type specimen of S. tomentosa were measured to be 27.5–33.5(−36) × 5.5–8.5 µm [66], which is considerably less than in our material. Previously, also Swinscow and Krog [12] reported a relatively large ascospore size (30–40(50) × 6–10 um) from East Africa.
Ecology and distribution: Sticta tomentosa is a pantropical species found especially in undisturbed, middle to high elevation rainforest habitats [60]. In our region, S. tomentosa is not very common and grows as an epiphyte on tree trunks. It can occasionally be locally abundant in lower montane forests, and also occurs in middle and upper montane forests (1650–3060 masl) in the Taita Hills and on Mt. Kilimanjaro.
3.2.15. Sticta umbilicariiformis Hochst. ex Flot. (Figure 13e–g)
Sticta umbilicariiformis has a cyanobacterial main photobiont (Nostoc) and typically many marginal pustules which can sometimes make it appear sorediate. Thalli of S. umbilicariiformis are often quite large with thick (usually 200–350 µm), wavy to foveolate, loosely adnate to ascending lobes 3–5 cm long and 2–4 cm wide. The upper surface is greyish brown with congested marginal, and often also laminal pustules that may occasionally appear sorediate. The lower surface is cream colored or, more rarely, brown, and thickly tomentose. Primary tomentum has brown and agglutinated stems and squarrose, white tips. Secondary tomentum is white and arachnoid, composed of moniliform assemblages. Cyphellae are slightly urceolate with a relatively wide opening (up to 1 mm in diam.) and raised margins, the pore is often surrounded by a darker ring.
Also fertile specimens of S. umbilicariiformis are common and usually lack symbiotic propagules. The thallus lobes of palmate fertile specimens are often thinner than the lobes of pustulate specimens and vary from wavy to strongly foveolate. Apothecia are submarginal and laminal, up to 2(3) mm wide, with brown disks and, occasionally, pubescent margins patterned with brown nodules. Ascospores are colorless, fusiform, 1-septate, 31–40 × 6.5–8 µm. Chemistry: K−, C−, Pd−.
Pustular thalli of S. umbilicariiformis are easily distinguished from other species in the region merely based on their habit. The only vaguely similar species is S. sublimbata which, however, produces true marginal soralia, has adnate and narrow lobes, and mainly occurs below 2000 masl. The characters that help to distinguish fertile specimens of S. umbilicariiformis from fertile specimens of S. aspratilis and S. munda are described under those species.
Ecology and distribution: Sticta umbilicariiformis is presently confirmed only from East Africa, but might have a much wider distribution reaching North America and Australia [20]. In our region, S. umbilicariiformis is common and abundant in the upper montane and subalpine zones on Mt. Kilimanjaro, and also occurs in middle montane zone (between 2540–3800 masl). It is by far the most abundant Sticta species in the subalpine zone, and was also collected from the Erica zone on Mt. Elgon. In the upper montane forest, S. umbilicariiformis mainly grows epiphytically on tree trunks and branches, in the subalpine zone also on rock and soil among bryophytes.
Notes: The phylogeny within the S. umbilicariiformis clade remains poorly resolved even in the more detailed analysis (Figure 2). Almost all pustular specimens represent the same ITS variant (umbilicariiformis1), closely related to a sequence from Rwanda (KT281697). The other ITS variants (umbilicariiformis2–5) are mainly from specimens without pustules, but which often have apothecia. The type specimen of S. umbilicariiformis (H-Nyl 33835; originally described in [67], lectotype designated in [20]) has both pustules and apothecia on the same thallus and the only fertile specimen of S. umbilicariiformis in our material that also has pustules belongs to ITS variant group umbilicariiformis4.
3.3. Sticta Diversity in the Montane Ecosystems of East Africa
Altogether 16 species of Sticta were collected from Mt. Kilimanjaro, of which S. cyanocaperata, S. fuliginoides, S. munda, Sticta sp. A, and Sticta sp. D were not found from other locations. The Taita Hills had 12 species, of which S. andina and Sticta sp. C (weigelii agg.) were not collected from the other locations. Mt. Kasigau had four species, and two species were collected from Mt. Elgon (Figure 14). A clear majority of all Sticta specimens were collected from montane forests, with not a single observation from lowland savannas or alpine Helichrysum heaths, or from agricultural or grassland habitats. The species diversity was highest in middle montane forests (Figure 14a). Sticta sublimbata was by far the most common Sticta species in the relatively open low elevation habitats. Sticta afromontana, S. duplolimbata, S. marginalis, and S. fuliginosa were common in lower to middle elevation forests, while upper montane and subalpine habitats were dominated by S. umbilicariiformis, S. afromontana, and S. fuliginoides (Figure 14b).
4. Discussion
A total of 19 Sticta species, including five species new to science, were found from the studied mountains in Kenya and Tanzania, and at least five other new species remain to be described pending more material. Of these species, only S. fuliginosa, S. sublimbata, S. tomentosa, and S. umbilicariiformis were known to be present in Kenya and/or Tanzania based on previous reports [12,20,28,29,30,31]. Sticta ciliata and S. duplolimbata had been previously reported from Rwanda [20] and are now confirmed to also occur in Kenya and Tanzania. Additionally, S. marginalis and S. fuliginoides are here reported as new for Kenya and Tanzania, and S. andina as new for Kenya.
Five species were described as new to science: Sticta afromontana, S. aspratilis, S. cellulosa, S. cyanocaperata, and S. munda, the three last mentioned of which have so far only been collected from Mt. Kilimanjaro. Additionally, five putative species, Sticta sp. A in the Sticta fuliginoides aggregate, Sticta sp. B in the Sticta ciliata aggregate, Sticta sp. C in the Sticta weigelii aggregate, Sticta sp. D, and Sticta sp. 2 were well resolved in the phylogenetic tree, but are not yet described due to insufficient material. In contrast to previous reports, it seems unlikely that S. ambavillaria, S. cyphellulata, S. limbata, S. kunthii, S. orbicularis, S. papyracea/variabilis, S. weigelii, or S. xanthotropa would occur in East Africa, as specimens with similar thallus morphologies are here shown to represent other species. Our study confirms the presence of 14 Sticta species in Kenya and 17 in Tanzania. Additionally, S. dichotoma and Sticta sp. 2 are expected to occur in Tanzania based on previous observations [12,28,30], raising the current total number of Sticta species in Tanzania to 19. A short synopsis of all Sticta species reported from East Africa is provided in Table 1.
Several previous studies have demonstrated that many of the “traditional” Sticta species, such as S. fuliginosa and S. weigelii, include taxa that belong to several different evolutionary lineages [19,20,21,68]. The previously reported S. fuliginosa is accompanied by several other taxa with laminal isidia also in East Africa: Sticta ciliata, S. fuliginoides, S. aspratilis, Sticta sp. B, and Sticta sp. A. Of these, S. fuliginoides and Sticta sp. A belong to the same large clade (Figure 3), are morphologically quite similar, and occur in similar habitats. The same applies to S. ciliata and Sticta sp. B. This suggests that there may still be significant undetected diversity hiding under the name S. fuliginosa, both in East Africa and globally.
Several Sticta species belonging to the S. weigelii morphodeme, i.e., those with elongate lobes and marginal isidia [21], were found from our region: Sticta andina, S. afromontana, S. cyanocaperata, and Sticta sp. C. Most of these have flattened isidia, but the isidia of Sticta sp. C are cylindrical, resembling those of S. weigelii s. str. [58,69]. However, Sticta sp. C does not have a color reaction with K, in contrast to the K+ yellow of S. weigelii s. str. [69], giving further support to our phylogenetic analysis, which indicated that S. weigelii s. str. has not been collected from East Africa. This supports the notion that S. weigelii s. str. may be restricted to the Neotropics [21,58]. In East Africa, Sticta specimens with flattened marginal isidia have previously been assigned to S. xanthotropa [12], a species that also mainly occurs in the Americas [32]. However, our results indicate that none of the East African species with flattened isidia actually represent S. xanthotropa: Sticta andina has a strong K+ yellow color reaction, while S. xanthotropa is K− [32]; Sticta afromontana has a robust thallus, while S. xanthotropa is described as “papery thin”, and the spores of S. afromontana are larger than those of S. xanthotropa [32]; Sticta cyanocaperata has a very dark lower surface, while it is pale in S. xanthotropa [32]. Furthermore, all the aforementioned species are mainly epiphytic, while S. xanthotropa has been reported to mainly grow on rocks and soil [32,70].
The group of species that have previously been reported from East Africa, but the presence of which we were not able to confirm include S. cyphellulata, S. orbicularis, S. limbata, S. ambavillaria and S. kunthii [12,26,27,28,30]. Our specimens morphologically matching the previous reports of S. orbicularis and S. cyphellulata were assigned to S. marginalis and S. duplolimbata, while the previously reported S. limbata represents S. umbilicariiformis, as already suggested by Magain and Sérusiaux [20]. Sticta ambavillaria and S. kunthii are both fertile species devoid of symbiotic propagules, and the previous reports from East Africa likely refer to S. munda and/or fertile specimens of S. aspratilis or S. umbilicariiformis. Based on our phylogenetic analyses, a specimen of S. ambavillaria (JQ735978) from Réunion from where the species was described [63], does not group with any of the East African specimens. Furthermore, already Swinscow and Krog [12] reported that the ascospores of East African specimens were shorter than what has been described from the type of S. ambavillaria [33]. Sticta kunthii was described from Peru and appears to have a mainly neotropical distribution [32,33]. It has been reported only once from East Africa, from an upper montane forest on Mt. Kenya. The specimen(s) were described to have “a thallus surface with numerous depressions, sometimes appearing almost pitted, a pale lower tomentum with medium-sized cyphellae, and apothecia with short marginal hairs” [26], which corresponds well with some fertile specimens of S. aspratilis and S. umbilicariiformis. However, the apothecia of these species are submarginal or laminal while those of S. kunthii are mainly marginal [32,33]. Furthermore, the apothecial margins of S. kunthii are distinctly hairy with long bundles of silky, white hairs [33], while those of East African specimens have only velvety stubble. No sequences have as yet been published from S. kunthii, but Moncada et al. [19] placed the species within the S. kunthii clade based on morphological evidence; in our material the only taxon belonging to that clade is the isidiate Sticta sp. D.
Sticta dichotoma and S. papyracea/variabilis are the only two species with green algae as the main photobiont previously reported to occur in East Africa, both from Tanzania where they are said to be rare [12,28,30]. While we did not find any specimens matching the description of S. dichotoma, the species may well be present in montane forests of Tanzania [12,28,30]. Our four Sticta specimens with a green algal photobiont all have marginal phyllidia, and thus correspond morphologically with S. papyracea [12]. Sticta papyracea and the synonymous S. variabilis were both originally described from Réunion [33,71]. However, in the phylogenetic analysis our specimens did not group together with S. variabilis from Réunion (Figure 5), but instead formed a clade with specimens of “Sticta sp. 2” by Simon et al. [25] collected from Madagascar. This putative species was described to have a green algal photobiont, elongated and dichotomously branching lobes, and apothecia, but lack lobules and phyllidia [25]. Hence, it seems quite possible, that our specimens represent yet another undescribed species. However, more collections are needed before definite conclusions can be made.
In the regions examined, the diversity of Sticta species was highest in indigenous moist forests of the middle montane zone. Two other Peltigeralean lichen genera, Leptogium and Peltigera, exhibited slightly different diversity patterns, with the highest diversity of Leptogium species recorded from moist lower montane forests and that of Peltigera species from the upper montane zone [13,17]. In comparison to the approximately 20 species of Sticta present in the study region, the genus Leptogium is much more diverse with possibly over 70 species in the region [13], while only 8 species of Peltigera have so far been collected from Mt. Kilimanjaro [17]. When comparing the natural and disturbed habitat types on Mt. Kilimanjaro, the number of Sticta species was usually at least slightly lower in disturbed habitats. Similar patterns has previously been reported also for Leptogium and Peltigera [13,17]. Even while the effects and degree of disturbance varied considerably between different habitat types, all the disturbed habitats tended to be at least slightly more open, and often considerably so, than the unaltered habitat types [34], with probable effects to illumination conditions, temperature, and humidity. A similar trend of decreasing lichen species diversity with increasing habitat disturbance has been observed also in previous studies [72,73,74]. Especially shade-adapted cyanolichens are easily negatively affected by disturbance, and these effects on total species diversity are not necessarily compensated by a concurrent increase in the number of heliophytic species [72].
5. Conclusions
At least 20 species of Sticta have now been confirmed to occur in East Africa, which is almost double to that known before. Only four of the presently accepted species are identified under the same species names that have been used in previous studies from the region, highlighting the general need of taxonomic revisions of lichenized fungi in Africa. The overall diversity of Sticta in East Africa is substantially higher than previously known, but seems to be lower than what has recently been recorded from some mountain regions in the Neotropics. Interestingly, even though our collections originated in relatively few mountain regions of Kenya and Tanzania, we still managed to collect specimens corresponding to almost all morphological species previously reported from East Africa. Both the relatively high number of novel taxa detected, and the percentage of species represented by only a few specimens and/or found from single localities, indicates that more comprehensive sampling will undoubtedly reveal further diversity in the genus Sticta in East Africa.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9020246/s1, Figure S1: Bayesian tree of the genus Sticta based on the nuITS genetic marker region.
Author Contributions
Conceptualization, U.K. and J.R.; methodology, U.K. and A.H.; formal analysis, U.K.; investigation, U.K.; resources, U.K., J.R. and P.M.K.; data curation, U.K.; writing—original draft preparation, U.K.; writing—review and editing, U.K., J.R., P.M.K., N.P.M. and A.H.; visualization, U.K. and J.R.; project administration, U.K. and J.R.; funding acquisition, U.K. and J.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by German Research Foundation, DFG grant number 408295270 (U.K.), and the European Union’s Horizon 2020 Research and Innovation Program Under the Marie Skłodowska-Curie Grant Agreement No. 705777 (U.K. and J.R.). Open access funding was provided by University of Helsinki.
Data Availability Statement
The sequence data presented in this study are openly available in the NCBI GenBank (www.ncbi.nlm.nih.gov/genbank/ accessed on 29 December 2022) with accession numbers OP999379–OP999611. The sequence alignment files and the resulting tree files from the phylogenetic analyses are openly available in the Zenodo repository (https://zenodo.org/) with doi 10.5281/zenodo.7575780 accessed on 29 December 2022.
Acknowledgments
We are grateful for the research infrastructure and research plots of the DFG-funded research unit ‘Kilimanjaro under global change’ (KiLi) utilized for the study. We also thank the staff of the Taita Research Station of the University of Helsinki for the provision of research facilities and accommodation. The work in Kenya was conducted under the National Council for Science and Technology permits no. NCST/RCD/17/012/33 and KWS/BRM/5001, and the work in Tanzania under the Tanzania Commission for Science and Technology permit no. 2016-368-NA-96-44. The work has been done in compliance with the Nagoya protocol on access and benefit sharing.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

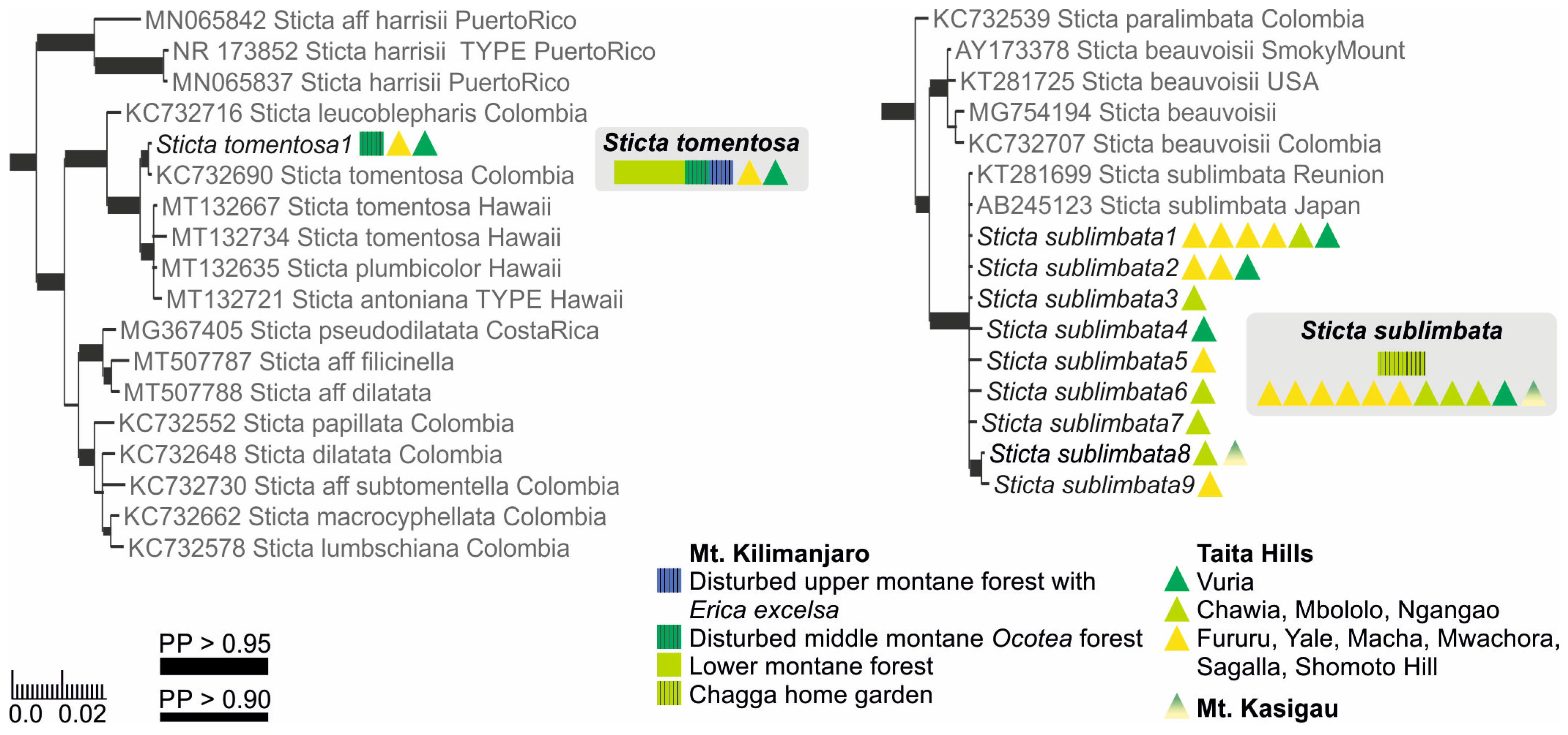{kind=link}
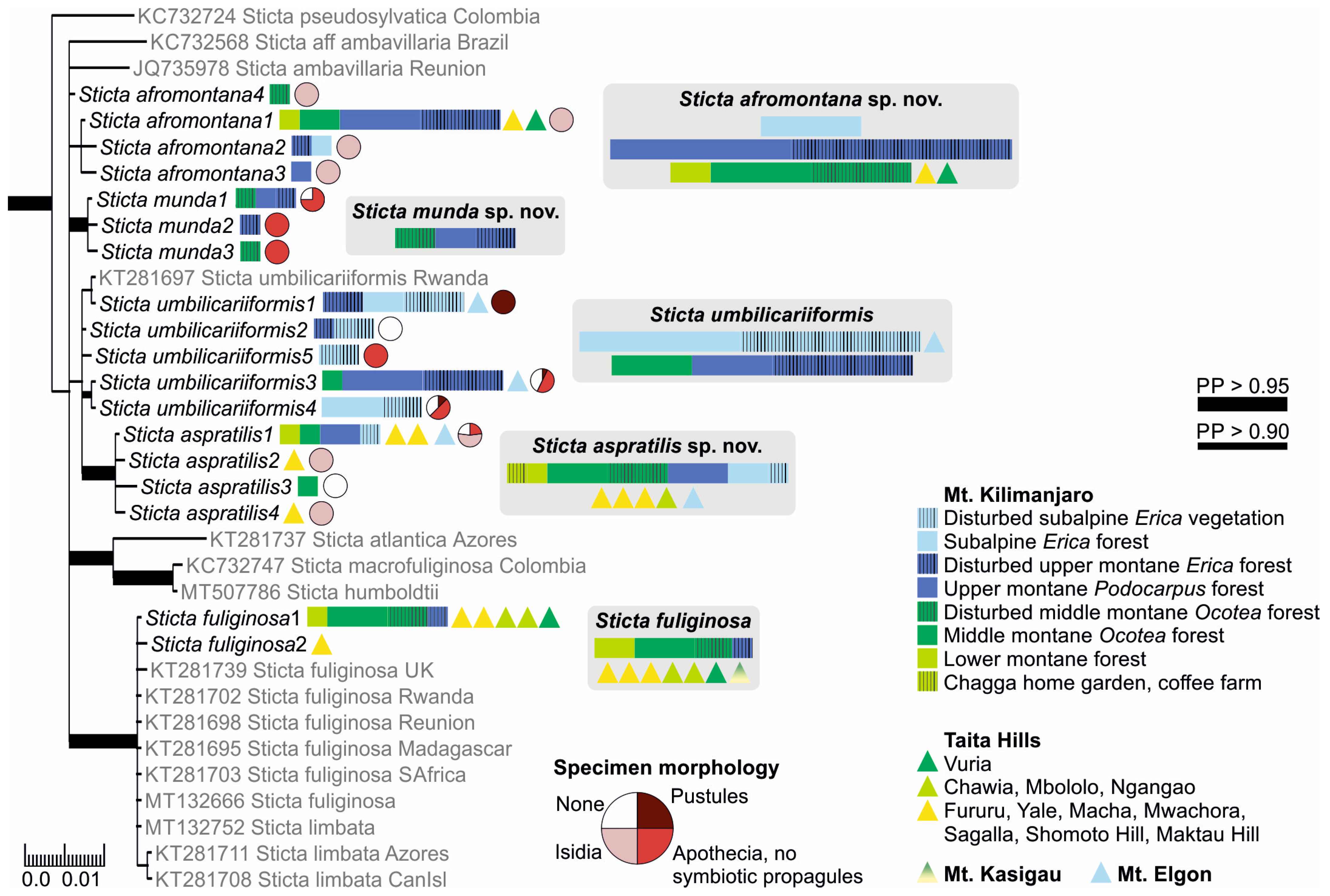{kind=link}
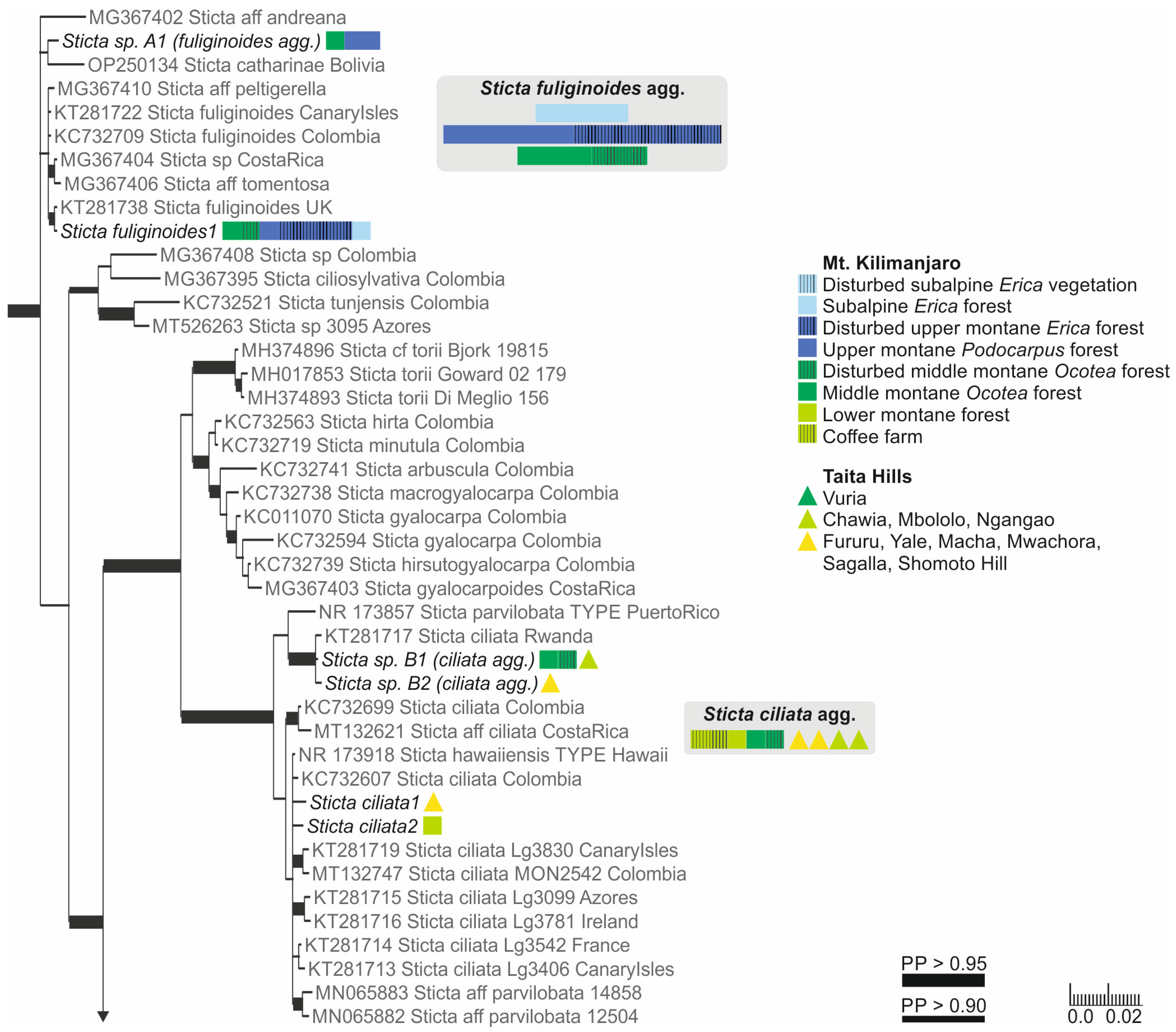{kind=link}
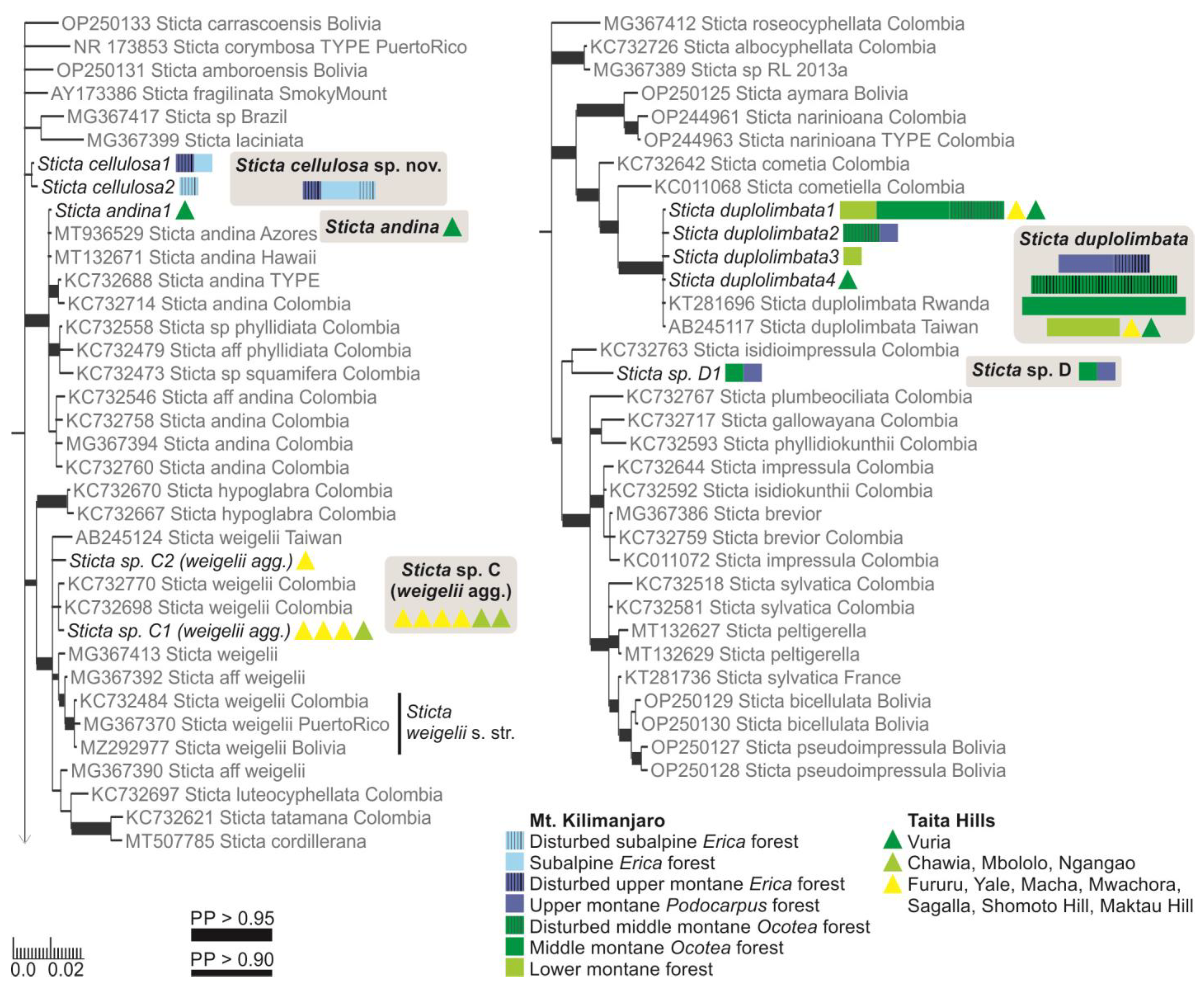{kind=link}
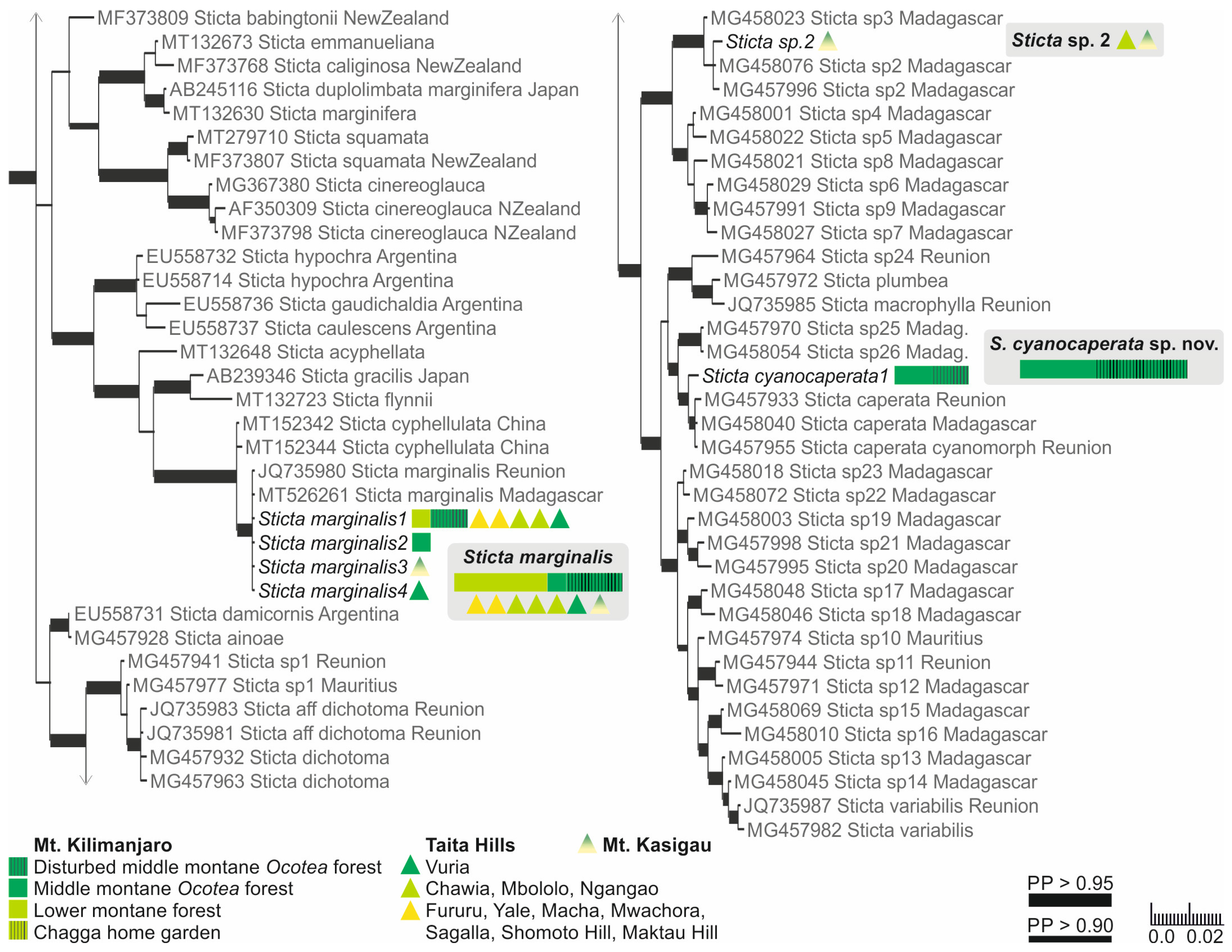{kind=link}
{kind=link}
{kind=link}
{kind=link}
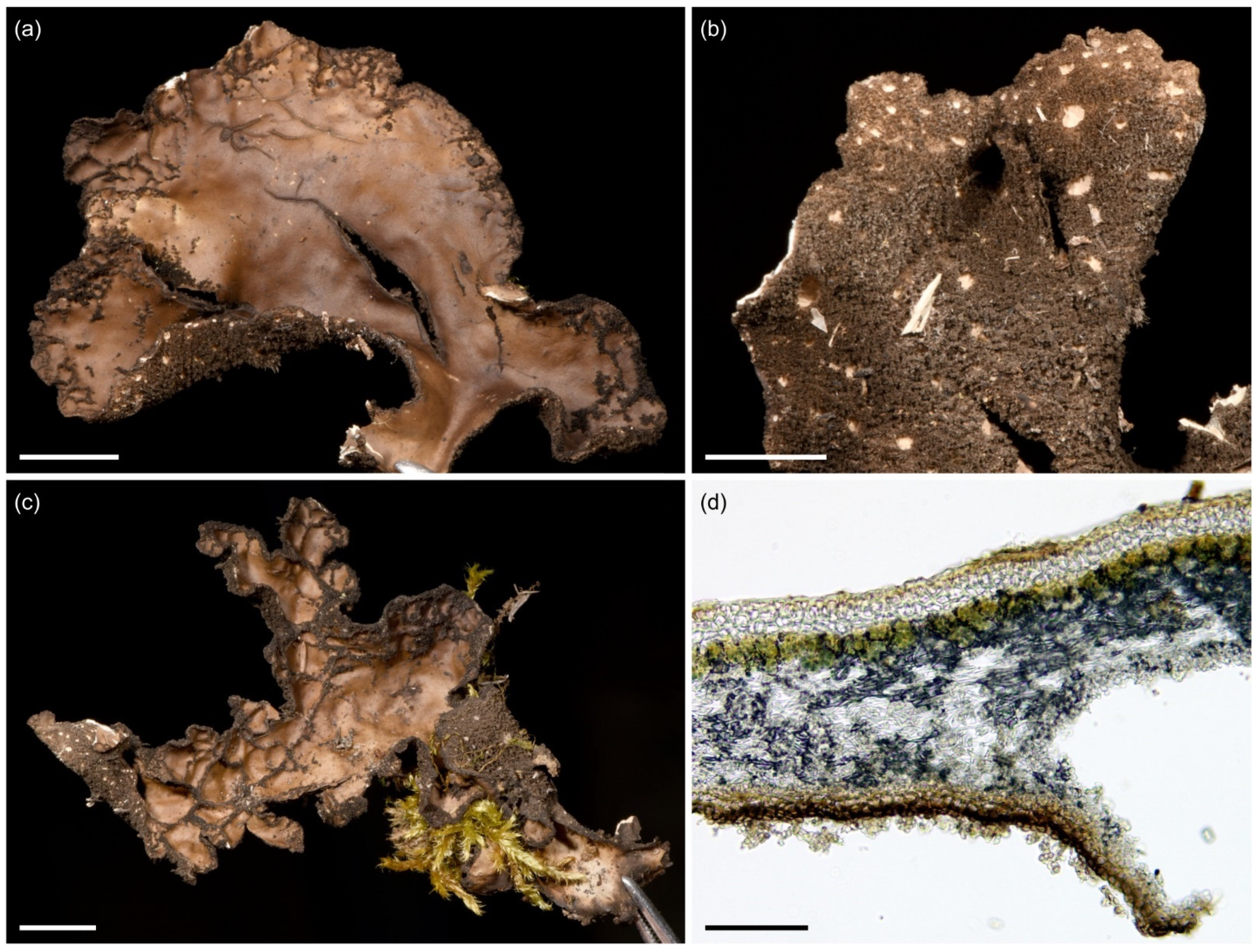{kind=link}
{kind=link}
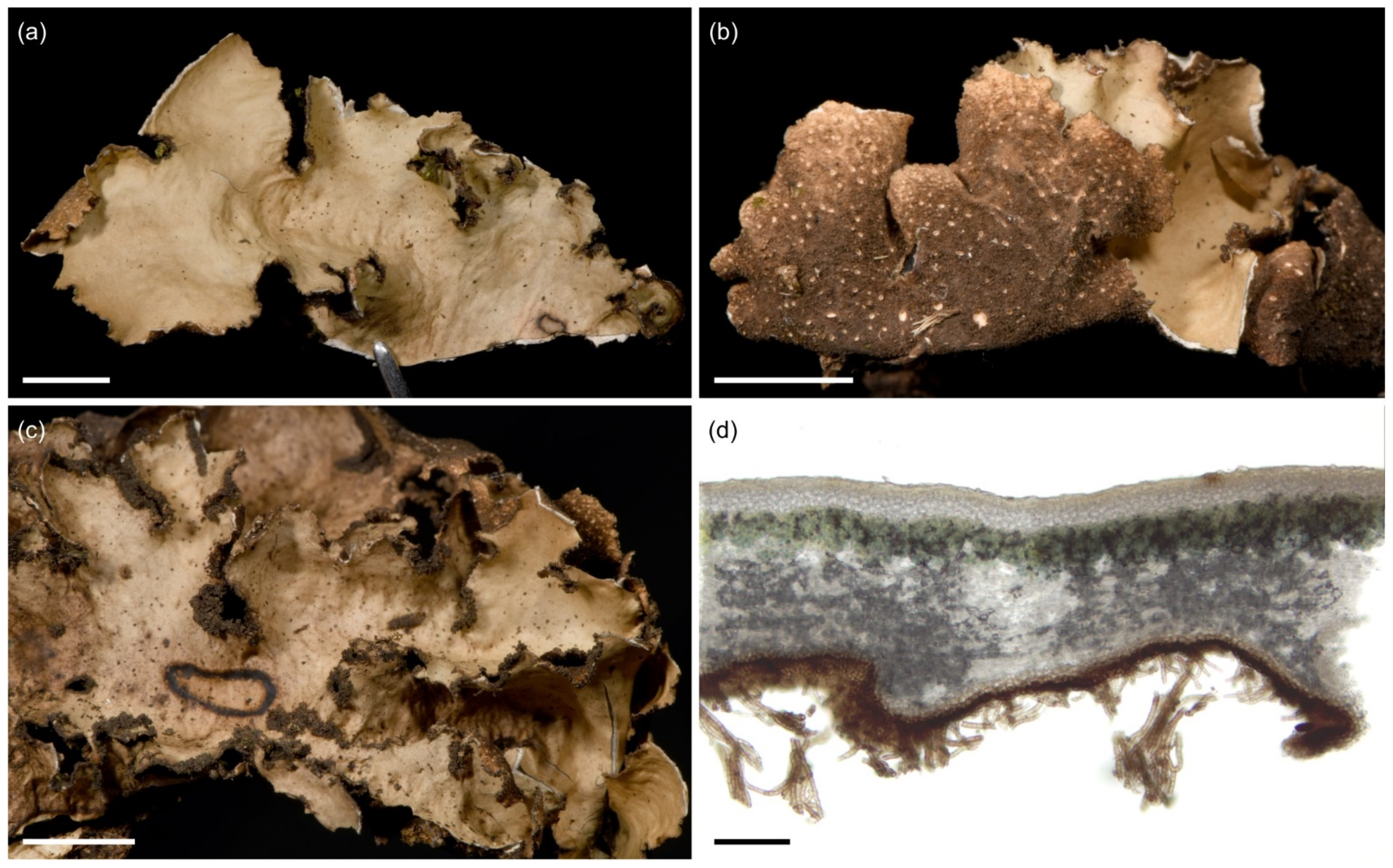{kind=link}
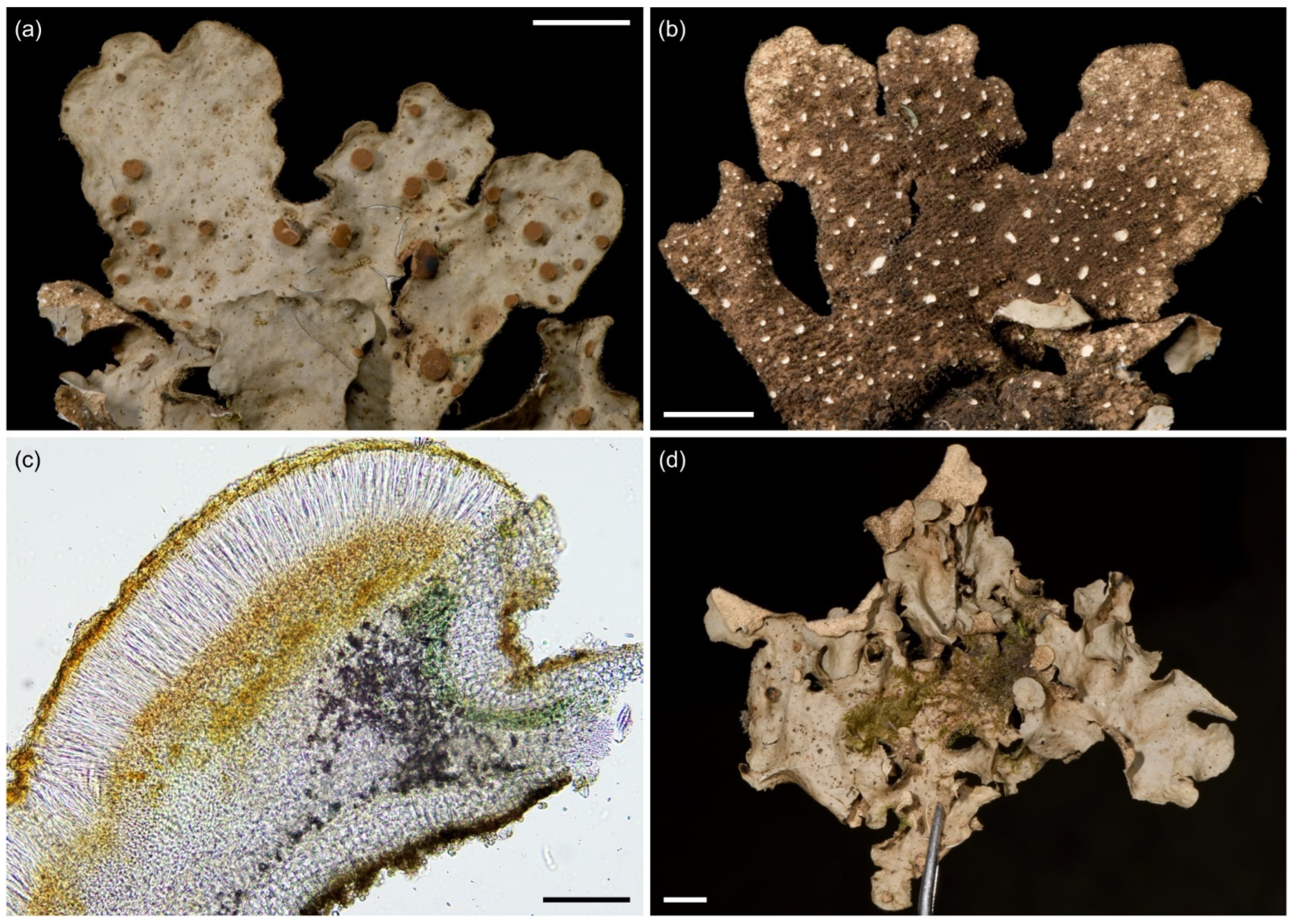{kind=link}
{kind=link}
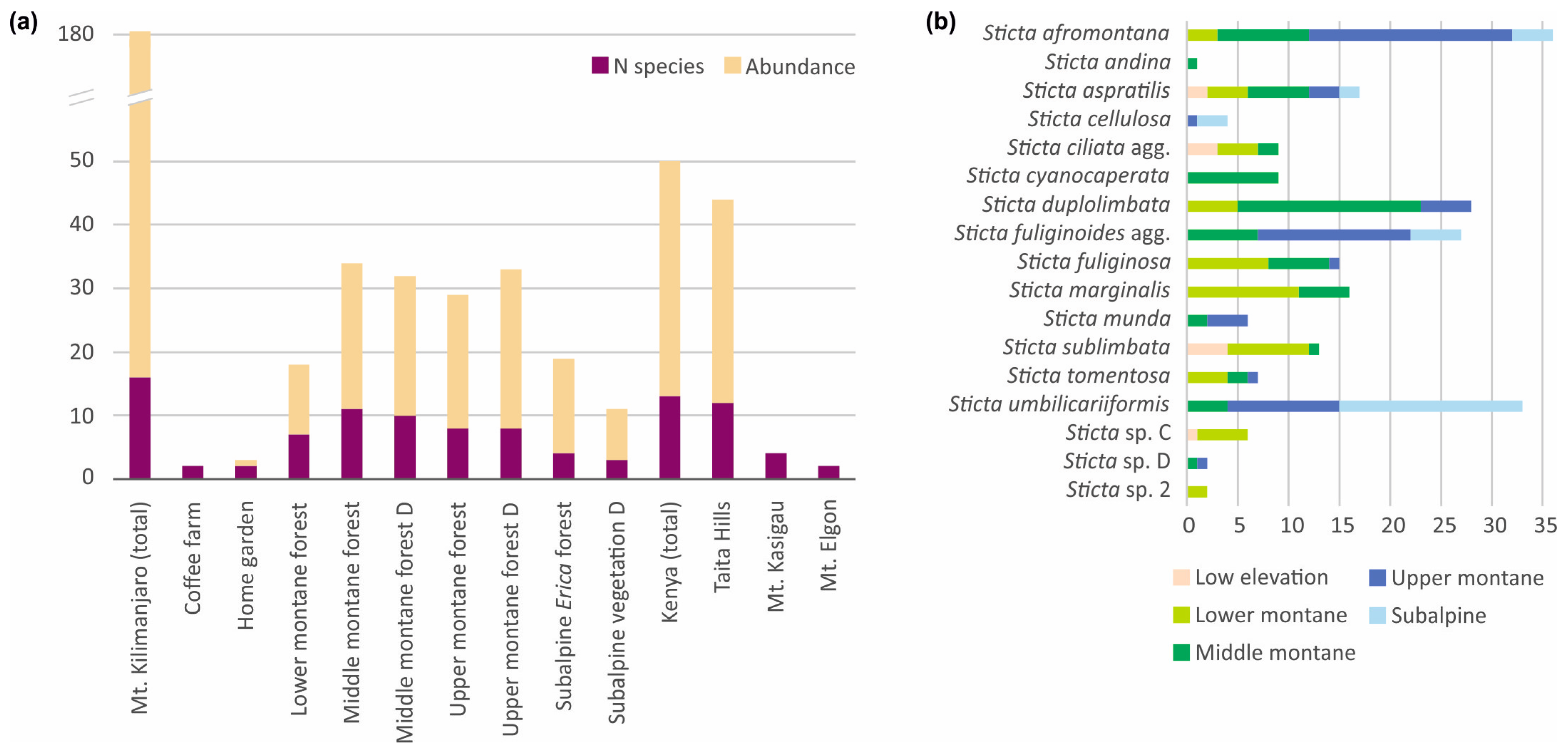{kind=link}
Table A1.
Specimen information. Specimens are collected from Kenya (Taita Hills, Mt. Kasigau, Mt. Elgon) and Tanzania (Kilimanjaro). Habitats on Mt. Kilimanjaro: Home = Chagga home garden, Cof = coffee farm, Flm = lower montane forest, FOc = middle montane Ocotea forest, FOD = disturbed Ocotea forest, FPo = upper montane Podocarpus forest, FPD = disturbed upper montane forest with Erica excelsa, FEr = subalpine Erica trimera forest, FED = disturbed subalpine Erica forest/shrubbery.
Table A1.
Specimen information. Specimens are collected from Kenya (Taita Hills, Mt. Kasigau, Mt. Elgon) and Tanzania (Kilimanjaro). Habitats on Mt. Kilimanjaro: Home = Chagga home garden, Cof = coffee farm, Flm = lower montane forest, FOc = middle montane Ocotea forest, FOD = disturbed Ocotea forest, FPo = upper montane Podocarpus forest, FPD = disturbed upper montane forest with Erica excelsa, FEr = subalpine Erica trimera forest, FED = disturbed subalpine Erica forest/shrubbery.
| Species | Collection ID | ITS Variant | Collection Location | Accession | |
|---|---|---|---|---|---|
| Sticta afromontana | JR10112 | afromontana1 | Taita Hills | Vuria | OP999402 |
| Sticta afromontana | JR10121B | afromontana1 | Taita Hills | Vuria | OP999404 |
| Sticta afromontana | JR10189 | afromontana1 | Taita Hills | Yale | OP999412 |
| Sticta afromontana | JR16366 | afromontana1 | Taita Hills | Vuria | OP999453 |
| Sticta afromontana | JR25 | - | Taita Hills | Yale | - |
| Sticta afromontana | JR31 | - | Taita Hills | Vuria | - |
| Sticta afromontana | UK110523c | - | Taita Hills | Vuria | - |
| Sticta afromontana | UK170771f | afromontana1 | Kilimanjaro | FPo1 | OP999477 |
| Sticta afromontana | UK170775b | afromontana3 | Kilimanjaro | FPo1 | OP999480 |
| Sticta afromontana | UK170779c | afromontana1 | Kilimanjaro | FPo1 | OP999481 |
| Sticta afromontana | UK170781b | afromontana1 | Kilimanjaro | FPo1 | OP999482 |
| Sticta afromontana | UK170792d | afromontana1 | Kilimanjaro | FOc2 | OP999484 |
| Sticta afromontana | UK170794d | - | Kilimanjaro | FOc2 | - |
| Sticta afromontana | UK170800b | - | Kilimanjaro | FOc1 | - |
| Sticta afromontana | UK170806j | afromontana1 | Kilimanjaro | FPD4 | OP999491 |
| Sticta afromontana | UK170807a | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170817h | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170822k | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170826e | afromontana1 | Kilimanjaro | FPD4 | OP999496 |
| Sticta afromontana | UK170830d | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170832c | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170834a | - | Kilimanjaro | FPD3 | - |
| Sticta afromontana | UK170836b | - | Kilimanjaro | FPD3 | - |
| Sticta afromontana | UK170841b | - | Kilimanjaro | FPD3 | - |
| Sticta afromontana | UK170842e | afromontana1 | Kilimanjaro | FPD3 | OP999498 |
| Sticta afromontana | UK170846e | afromontana1 | Kilimanjaro | FPD3 | OP999501 |
| Sticta afromontana | UK170856c | - | Kilimanjaro | FPo4 | - |
| Sticta afromontana | UK170857a | afromontana1 | Kilimanjaro | FPo4 | OP999503 |
| Sticta afromontana | UK170858g | afromontana1 | Kilimanjaro | FPo4 | OP999507 |
| Sticta afromontana | UK170880e | - | Kilimanjaro | FPo5 | - |
| Sticta afromontana | UK170886a | afromontana1 | Kilimanjaro | FPo5 | OP999509 |
| Sticta afromontana | UK170892b | - | Kilimanjaro | FPo5 | - |
| Sticta afromontana | UK170897ar | - | Kilimanjaro | FOD4 | - |
| Sticta afromontana | UK170911b | - | Kilimanjaro | FOD5 | - |
| Sticta afromontana | UK170915a | - | Kilimanjaro | FOD5 | - |
| Sticta afromontana | UK170930c | afromontana1 | Kilimanjaro | Flm1 | OP999523 |
| Sticta afromontana | UK170951g | - | Kilimanjaro | Flm1 | - |
| Sticta afromontana | UK171180a | afromontana4 | Kilimanjaro | FOD3 | OP999537 |
| Sticta afromontana | UK171185h | - | Kilimanjaro | FOD3 | - |
| Sticta afromontana | UK171405e | - | Kilimanjaro | FEr4 | - |
| Sticta afromontana | UK171413a | - | Kilimanjaro | FEr4 | - |
| Sticta afromontana | UK171438g | - | Kilimanjaro | FEr2 | - |
| Sticta afromontana | UK171439t | afromontana2 | Kilimanjaro | FEr2 | OP999554 |
| Sticta afromontana | UK171442c | - | Kilimanjaro | FEr2 | - |
| Sticta afromontana | UK171445j | - | Kilimanjaro | FEr2 | - |
| Sticta afromontana | UK171470v | - | Kilimanjaro | FPD1 | - |
| Sticta afromontana | UK171471c | - | Kilimanjaro | FPD1 | - |
| Sticta afromontana | UK171476e | afromontana1 | Kilimanjaro | FPD1 | OP999561 |
| Sticta afromontana | UK171490e | afromontana1 | Kilimanjaro | FOc3 | OP999566 |
| Sticta afromontana | UK171525a | - | Kilimanjaro | FPo2 | - |
| Sticta afromontana | UK171526a | afromontana1 | Kilimanjaro | FPo2 | OP999590 |
| Sticta afromontana | UK171528b | afromontana1_4 | Kilimanjaro | FPo2 | OP999591 |
| Sticta afromontana | UK171577f | afromontana2 | Kilimanjaro | FPD2 | OP999595 |
| Sticta afromontana | UK171578m | - | Kilimanjaro | FPD2 | - |
| Sticta afromontana | UK171581j | - | Kilimanjaro | FPD2 | - |
| Sticta afromontana | UK171582p | - | Kilimanjaro | FPD2 | - |
| Sticta afromontana | UK171584v | afromontana1 | Kilimanjaro | FPD2 | OP999601 |
| Sticta afromontana | UK171589b | - | Kilimanjaro | FPo3 | - |
| Sticta andina | JR10117 | andina1 | Taita Hills | Vuria | OP999403 |
| Sticta aspratilis | JR10044A | aspratilis4 | Taita Hills | Shomoto Hill | OP999394 |
| Sticta aspratilis | JR10057 | aspratilis1 | Taita Hills | Shomoto Hill | OP999395 |
| Sticta aspratilis | JR10155A | aspratilis1 | Taita Hills | Yale | OP999406 |
| Sticta aspratilis | JR10155B | aspratilis1 | Taita Hills | Yale | OP999407 |
| Sticta aspratilis | JR10155C | aspratilis1 | Taita Hills | Yale | OP999408 |
| Sticta aspratilis | JR10171 | aspratilis1 | Taita Hills | Yale | OP999409 |
| Sticta aspratilis | JR16103 | aspratilis1 | Mt. Elgon | Erica zone | OP999436 |
| Sticta aspratilis | JR16107 | aspratilis1 | Mt. Elgon | Erica zone | OP999437 |
| Sticta aspratilis | UK110551f | aspratilis2 | Taita Hills | Fururu | OP999475 |
| Sticta aspratilis | UK110551g | aspratilis2 | Taita Hills | Fururu | OP999476 |
| Sticta aspratilis | UK110566c | - | Taita Hills | Chawia | - |
| Sticta aspratilis | UK170781c | - | Kilimanjaro | FPo1 | - |
| Sticta aspratilis | UK170892c | aspratilis1 | Kilimanjaro | FPo5 | OP999510 |
| Sticta aspratilis | UK170896b | aspratilis1and2 | Kilimanjaro | FPo5 | OP999512 |
| Sticta aspratilis | UK170910b | - | Kilimanjaro | FOD5 | - |
| Sticta aspratilis | UK170912b | - | Kilimanjaro | FOD5 | - |
| Sticta aspratilis | UK170916c | aspratilis1 | Kilimanjaro | Flm6 | OP999520 |
| Sticta aspratilis | UK170975f | aspratilis1 | Kilimanjaro | Cof5 | OP999530 |
| Sticta aspratilis | UK171478a | aspratilis1 | Kilimanjaro | FOc4 | OP999562 |
| Sticta aspratilis | UK171487c | aspratilis1 | Kilimanjaro | FOc4 | OP999565 |
| Sticta aspratilis | UK171508k | - | Kilimanjaro | FOD2 | - |
| Sticta aspratilis | UK171515b | aspratilis3 | Kilimanjaro | FOc5 | OP999584 |
| Sticta aspratilis | UK171562c | aspratilis1 | Kilimanjaro | FED2 | OP999592 |
| Sticta aspratilis | UK171587j | aspratilis1 | Kilimanjaro | FPo3 | OP999607 |
| Sticta cellulosa | UK171340t | cellulosa2 | Kilimanjaro | FED1 | OP999545 |
| Sticta cellulosa | UK171406e | cellulosa1 | Kilimanjaro | FEr4 | OP999548 |
| Sticta cellulosa | UK171407j | cellulosa1 | Kilimanjaro | FEr4 | OP999549 |
| Sticta cellulosa | UK171458k | - | Kilimanjaro | FEr4 | - |
| Sticta cellulosa | UK171584k | cellulosa1 | Kilimanjaro | FPD2 | OP999599 |
| Sticta ciliata | JR16294b | ciliata1 | Taita Hills | Sagalla | OP999443 |
| Sticta ciliata | UK170944j | ciliata2 | Kilimanjaro | Flm1 | OP999528 |
| Sticta ciliata (agg.) | UK110570a | - | Taita Hills | Chawia | - |
| Sticta ciliata (agg.) | UK160552r | - | Kilimanjaro | Home3 | - |
| Sticta ciliata (agg.) | UK170984 | - | Kilimanjaro | Cof5 | - |
| Sticta cyanocaperata | UK170897f | - | Kilimanjaro | FOD4 | - |
| Sticta cyanocaperata | UK170912a | cyanocaperata1 | Kilimanjaro | FOD5 | OP999516 |
| Sticta cyanocaperata | UK170912e | cyanocaperata1 | Kilimanjaro | FOD5 | OP999518 |
| Sticta cyanocaperata | UK171182c | cyanocaperata1 | Kilimanjaro | FOD3 | OP999539 |
| Sticta cyanocaperata | UK171480d | cyanocaperata1 | Kilimanjaro | FOc4 | OP999563 |
| Sticta cyanocaperata | UK171483v | - | Kilimanjaro | FOc4 | - |
| Sticta cyanocaperata | UK171486b | - | Kilimanjaro | FOc4 | - |
| Sticta cyanocaperata | UK171495f | cyanocaperata1 | Kilimanjaro | FOc3 | OP999570 |
| Sticta cyanocaperata | UK171506g | - | Kilimanjaro | FOD2 | - |
| Sticta cyanocaperata | UK171510m | - | Kilimanjaro | FOD2 | - |
| Sticta duplolimbata | JR10194A | duplolimbata1 | Taita Hills | Yale | OP999416 |
| Sticta duplolimbata | JR10194B | - | Taita Hills | Yale | - |
| Sticta duplolimbata | JR16374b | duplolimbata4 | Taita Hills | Vuria | OP999460 |
| Sticta duplolimbata | JR16374d | duplolimbata1 | Taita Hills | Vuria | OP999462 |
| Sticta duplolimbata | JR68a | duplolimbata1_2_4 | Taita Hills | Vuria | OP999470 |
| Sticta duplolimbata | JR69 | duplolimbata1 | Taita Hills | Vuria | OP999471 |
| Sticta duplolimbata | UK170790 | duplolimbata1 | Kilimanjaro | FOc2 | OP999483 |
| Sticta duplolimbata | UK170799d | duplolimbata1 | Kilimanjaro | FOc1 | OP999489 |
| Sticta duplolimbata | UK170845o | - | Kilimanjaro | FPD3 | - |
| Sticta duplolimbata | UK170893a | duplolimbata1 | Kilimanjaro | FPo5 | OP999511 |
| Sticta duplolimbata | UK170914b | duplolimbata2 | Kilimanjaro | FOD5 | OP999519 |
| Sticta duplolimbata | UK170929c | duplolimbata1 | Kilimanjaro | Flm6 | OP999521 |
| Sticta duplolimbata | UK170931h | duplolimbata3 | Kilimanjaro | Flm1 | OP999524 |
| Sticta duplolimbata | UK170933g | - | Kilimanjaro | Flm1 | - |
| Sticta duplolimbata | UK170936g | - | Kilimanjaro | Flm1 | - |
| Sticta duplolimbata | UK170950c | - | Kilimanjaro | Flm1 | - |
| Sticta duplolimbata | UK170952e | duplolimbata1 | Kilimanjaro | Flm1 | OP999529 |
| Sticta duplolimbata | UK171178f | duplolimbata1 | Kilimanjaro | FOD3 | OP999536 |
| Sticta duplolimbata | UK171181e | duplolimbata2 | Kilimanjaro | FOD3 | OP999538 |
| Sticta duplolimbata | UK171182b | - | Kilimanjaro | FOD3 | - |
| Sticta duplolimbata | UK171188b | - | Kilimanjaro | FOD3 | - |
| Sticta duplolimbata | UK171472d | - | Kilimanjaro | FPD1 | - |
| Sticta duplolimbata | UK171483g | - | Kilimanjaro | FOc4 | - |
| Sticta duplolimbata | UK171490f | duplolimbata1 | Kilimanjaro | FOc3 | OP999567 |
| Sticta duplolimbata | UK171493b | - | Kilimanjaro | FOc3 | - |
| Sticta duplolimbata | UK171495e | duplolimbata1 | Kilimanjaro | FOc3 | OP999569 |
| Sticta duplolimbata | UK171497e | duplolimbata1 | Kilimanjaro | FOD1 | OP999572 |
| Sticta duplolimbata | UK171497f | duplolimbata1 | Kilimanjaro | FOD1 | OP999573 |
| Sticta duplolimbata | UK171502b | duplolimbata1 | Kilimanjaro | FOD1 | OP999574 |
| Sticta duplolimbata | UK171502c | duplolimbata1 | Kilimanjaro | FOD1 | OP999575 |
| Sticta duplolimbata | UK171503c | - | Kilimanjaro | FOD1 | - |
| Sticta duplolimbata | UK171504b | duplolimbata1 | Kilimanjaro | FOD2 | OP999576 |
| Sticta duplolimbata | UK171513f | duplolimbata1 | Kilimanjaro | FOc5 | OP999582 |
| Sticta duplolimbata | UK171519g | duplolimbata1 | Kilimanjaro | FOc5 | OP999585 |
| Sticta duplolimbata | UK171521h | - | Kilimanjaro | FOc5 | - |
| Sticta duplolimbata | UK171586p | duplolimbata2 | Kilimanjaro | FPo3 | OP999605 |
| Sticta duplolimbata | UK171590q | duplolimbata2 | Kilimanjaro | FPo3 | OP999609 |
| Sticta duplolimbata | UK171596e | duplolimbata1 | Kilimanjaro | Flm2 | OP999610 |
| Sticta fuliginoides | UK170808e | fuliginoides1 | Kilimanjaro | FPD4 | OP999494 |
| Sticta fuliginoides | UK170844c | fuliginoides1 | Kilimanjaro | FPD3 | OP999499 |
| Sticta fuliginoides | UK170858e | fuliginoides1 | Kilimanjaro | FPo4 | OP999505 |
| Sticta fuliginoides | UK171438f | fuliginoides1 | Kilimanjaro | FEr2 | OP999552 |
| Sticta fuliginoides | UK171468d | fuliginoides1 | Kilimanjaro | FPD1 | OP999558 |
| Sticta fuliginoides | UK171485c | fuliginoides1 | Kilimanjaro | FOc4 | OP999564 |
| Sticta fuliginoides | UK171504c | fuliginoides1 | Kilimanjaro | FOD2 | OP999577 |
| Sticta fuliginoides | UK171577i | fuliginoides1 | Kilimanjaro | FPD2 | OP999597 |
| Sticta fuliginoides (agg.) | UK170773c | - | Kilimanjaro | FPo1 | - |
| Sticta fuliginoides (agg.) | UK170775c | - | Kilimanjaro | FPo1 | - |
| Sticta fuliginoides (agg.) | UK170826g | - | Kilimanjaro | FPD4 | - |
| Sticta fuliginoides (agg.) | UK170845m | - | Kilimanjaro | FPD3 | - |
| Sticta fuliginoides (agg.) | UK170888g | - | Kilimanjaro | FPo5 | - |
| Sticta fuliginoides (agg.) | UK170889e | - | Kilimanjaro | FPo5 | - |
| Sticta fuliginoides (agg.) | UK170897g | - | Kilimanjaro | FOD4 | - |
| Sticta fuliginoides (agg.) | UK171439h | - | Kilimanjaro | FEr2 | - |
| Sticta fuliginoides (agg.) | UK171445h | - | Kilimanjaro | FEr2 | - |
| Sticta fuliginoides (agg.) | UK171456b | - | Kilimanjaro | FEr2 | - |
| Sticta fuliginoides (agg.) | UK171458m | - | Kilimanjaro | FEr4 | - |
| Sticta fuliginoides (agg.) | UK171467j | - | Kilimanjaro | FEr4 | - |
| Sticta fuliginoides (agg.) | UK171471d | - | Kilimanjaro | FPD1 | - |
| Sticta fuliginoides (agg.) | UK171473m | - | Kilimanjaro | FPD1 | - |
| Sticta fuliginoides (agg.) | UK171497ab | - | Kilimanjaro | FOD1 | - |
| Sticta fuliginoides (agg.) | UK171505d | - | Kilimanjaro | FOD2 | - |
| Sticta fuliginoides (agg.) | UK171514d | - | Kilimanjaro | FOc5 | - |
| Sticta fuliginoides (agg.) | UK171523m | - | Kilimanjaro | FPo2 | - |
| Sticta fuliginoides (agg.) | UK171582j | - | Kilimanjaro | FPD2 | - |
| Sticta fuliginoides (agg.) | UK171584l | - | Kilimanjaro | FPD2 | - |
| Sticta fuliginoides (agg.) | UK171586m | - | Kilimanjaro | FPo3 | - |
| Sticta fuliginoides (agg.) | UK171590r | - | Kilimanjaro | FPo3 | - |
| Sticta fuliginosa | JR09A46 | fuliginosa1 | Taita Hills | Ngangao | OP999381 |
| Sticta fuliginosa | JR09D21B | fuliginosa1 | Taita Hills | Ngangao | OP999384 |
| Sticta fuliginosa | JR10076 | fuliginosa1 | Taita Hills | Vuria | OP999399 |
| Sticta fuliginosa | JR10151 | fuliginosa1 | Taita Hills | Yale | OP999405 |
| Sticta fuliginosa | JR10190B | fuliginosa1 | Taita Hills | Yale | OP999413 |
| Sticta fuliginosa | JR10245 | fuliginosa2 | Taita Hills | Mwachora | OP999425 |
| Sticta fuliginosa | JR105A | fuliginosa1 | Taita Hills | Vuria | OP999428 |
| Sticta fuliginosa | JR105B | fuliginosa1 | Taita Hills | Vuria | OP999429 |
| Sticta fuliginosa | JR10K564 | - | Mt. Kasigau | Plot W12 | - |
| Sticta fuliginosa | JR16335 | fuliginosa1 | Taita Hills | Sagalla | OP999448 |
| Sticta fuliginosa | JR16354 | fuliginosa1 | Taita Hills | Vuria | OP999450 |
| Sticta fuliginosa | JR16358 | fuliginosa1 | Taita Hills | Vuria | OP999452 |
| Sticta fuliginosa | JR16457 | - | Taita Hills | Mbololo | |
| Sticta fuliginosa | JR16463 | fuliginosa1 | Taita Hills | Mbololo | OP999468 |
| Sticta fuliginosa | JR19 | - | Taita Hills | Yale | - |
| Sticta fuliginosa | JR3 | fuliginosa1 | Taita Hills | Yale | OP999469 |
| Sticta fuliginosa | UK110504b | - | Taita Hills | Ngangao | - |
| Sticta fuliginosa | UK110512a | - | Taita Hills | Vuria | - |
| Sticta fuliginosa | UK170794g | fuliginosa1 | Kilimanjaro | FOc2 | OP999486 |
| Sticta fuliginosa | UK170796a | fuliginosa1 | Kilimanjaro | FOc2 | OP999487 |
| Sticta fuliginosa | UK170797j | fuliginosa1 | Kilimanjaro | FOc1 | OP999488 |
| Sticta fuliginosa | UK170846f | fuliginosa1 | Kilimanjaro | FPD3 | OP999502 |
| Sticta fuliginosa | UK170900h | fuliginosa1 | Kilimanjaro | FOD4 | OP999513 |
| Sticta fuliginosa | UK170906b | fuliginosa1 | Kilimanjaro | FOD4 | OP999515 |
| Sticta fuliginosa | UK170930b | fuliginosa1 | Kilimanjaro | Flm1 | OP999522 |
| Sticta fuliginosa | UK171494n | fuliginosa1 | Kilimanjaro | FOc3 | OP999568 |
| Sticta fuliginosa | UK171504m | fuliginosa1 | Kilimanjaro | FOD2 | OP999579 |
| Sticta fuliginosa | UK171594e | - | Kilimanjaro | Flm2 | - |
| Sticta marginalis | JR_58_7 | marginalis3 | Mt. Kasigau | Plot W14 | OP999379 |
| Sticta marginalis | JR090X8a | - | Taita Hills | Chawia | - |
| Sticta marginalis | JR09D12A | marginalis1 | Taita Hills | Ngangao | OP999383 |
| Sticta marginalis | JR10028B | marginalis1 | Taita Hills | Vuria | OP999391 |
| Sticta marginalis | JR10176A | marginalis1 | Taita Hills | Yale | OP999410 |
| Sticta marginalis | JR10193A | marginalis1 | Taita Hills | Yale | OP999415 |
| Sticta marginalis | JR10202B | marginalis1 | Taita Hills | Yale | OP999417 |
| Sticta marginalis | JR10K532 | - | Mt. Kasigau | Plot W9 | - |
| Sticta marginalis | JR10K552A | - | Mt. Kasigau | Plot W11 | - |
| Sticta marginalis | JR10K552B | - | Mt. Kasigau | Plot W11 | - |
| Sticta marginalis | JR10K563 | - | Mt. Kasigau | Plot W12 | - |
| Sticta marginalis | JR110030A | marginalis4 | Taita Hills | Vuria | OP999432 |
| Sticta marginalis | JR110055 | marginalis1 | Taita Hills | Vuria | OP999433 |
| Sticta marginalis | JR16248 | marginalis1 | Taita Hills | Mbololo | OP999441 |
| Sticta marginalis | JR16294a | - | Taita Hills | Sagalla | - |
| Sticta marginalis | JR16312 | marginalis1 | Taita Hills | Sagalla | OP999446 |
| Sticta marginalis | JR16322a | marginalis1 | Taita Hills | Sagalla | OP999447 |
| Sticta marginalis | JR16420 | - | Taita Hills | Mbololo | - |
| Sticta marginalis | JR16440 | - | Taita Hills | Mbololo | - |
| Sticta marginalis | UK110501a | - | Taita Hills | Ngangao | - |
| Sticta marginalis | UK110535b | - | Taita Hills | Vuria | - |
| Sticta marginalis | UK170800m | marginalis2 | Kilimanjaro | FOc1 | OP999490 |
| Sticta marginalis | UK170927a | - | Kilimanjaro | Flm6 | - |
| Sticta marginalis | UK170932b | marginalis1 | Kilimanjaro | Flm1 | OP999525 |
| Sticta marginalis | UK170933h | marginalis1 | Kilimanjaro | Flm1 | OP999526 |
| Sticta marginalis | UK170934b | - | Kilimanjaro | Flm1 | - |
| Sticta marginalis | UK170937j | - | Kilimanjaro | Flm1 | - |
| Sticta marginalis | UK170942c | marginalis1 | Kilimanjaro | Flm1 | OP999527 |
| Sticta marginalis | UK170943d | - | Kilimanjaro | Flm1 | - |
| Sticta marginalis | UK170950d | - | Kilimanjaro | Flm1 | - |
| Sticta marginalis | UK171183a | marginalis1 | Kilimanjaro | FOD3 | OP999540 |
| Sticta marginalis | UK171183b | - | Kilimanjaro | FOD3 | - |
| Sticta marginalis | UK171186a | - | Kilimanjaro | FOD3 | - |
| Sticta marginalis | UK171504k | marginalis1 | Kilimanjaro | FOD2 | OP999578 |
| Sticta marginalis | UK171596k | marginalis1 | Kilimanjaro | Flm2 | OP999611 |
| Sticta munda | UK170888c | - | Kilimanjaro | FPo5 | - |
| Sticta munda | UK171497d | munda3 | Kilimanjaro | FOD1 | OP999571 |
| Sticta munda | UK171508j | munda1 | Kilimanjaro | FOD2 | OP999580 |
| Sticta munda | UK171510k | munda1 | Kilimanjaro | FOD2 | OP999581 |
| Sticta munda | UK171582i | munda2 | Kilimanjaro | FPD2 | OP999598 |
| Sticta munda | UK171584u | munda1 | Kilimanjaro | FPD2 | OP999600 |
| Sticta munda | UK171586k | munda1 | Kilimanjaro | FPo3 | OP999604 |
| Sticta munda | UK171586l | - | Kilimanjaro | FPo3 | - |
| Sticta sublimbata | JR09A60 | sublimbata1 | Taita Hills | Ngangao | OP999382 |
| Sticta sublimbata | JR09D6A | sublimbata1 | Taita Hills | Ngangao | OP999385 |
| Sticta sublimbata | JR09W12a | sublimbata8 | Mt. Kasigau | - | OP999387 |
| Sticta sublimbata | JR09Y12a | sublimbata6 | Taita Hills | Mbololo | OP999388 |
| Sticta sublimbata | JR09Y15b | sublimbata6 | Taita Hills | Mbololo | OP999389 |
| Sticta sublimbata | JR09Z2 | sublimbata1 | Taita Hills | Ngangao | OP999390 |
| Sticta sublimbata | JR10032A | sublimbata2 | Taita Hills | Shomoto Hill | OP999392 |
| Sticta sublimbata | JR10039B | sublimbata2 | Taita Hills | Shomoto Hill | OP999393 |
| Sticta sublimbata | JR10066A | sublimbata2 | Taita Hills | Shomoto Hill | OP999397 |
| Sticta sublimbata | JR10066B | sublimbata2 | Taita Hills | Shomoto Hill | OP999398 |
| Sticta sublimbata | JR10191 | sublimbata1 | Taita Hills | Yale | OP999414 |
| Sticta sublimbata | JR10208A | sublimbata5 | Taita Hills | Yale | OP999418 |
| Sticta sublimbata | JR10214B | sublimbata1 | Taita Hills | Yale | OP999420 |
| Sticta sublimbata | JR10220A | sublimbata2 | Taita Hills | Mwachora | OP999421 |
| Sticta sublimbata | JR10228D | sublimbata1 | Taita Hills | Mwachora | OP999422 |
| Sticta sublimbata | JR10230A | sublimbata1 | Taita Hills | Mwachora | OP999423 |
| Sticta sublimbata | JR10241 | sublimbata1 | Taita Hills | Mwachora | OP999424 |
| Sticta sublimbata | JR10282B | sublimbata1 | Taita Hills | Macha | OP999427 |
| Sticta sublimbata | JR10K494 | - | Mt. Kasigau | Plot W5 | - |
| Sticta sublimbata | JR10K495 | - | Mt. Kasigau | Plot W5 | - |
| Sticta sublimbata | JR16275 | sublimbata9 | Taita Hills | Sagalla | OP999442 |
| Sticta sublimbata | JR16355 | sublimbata2 | Taita Hills | Vuria | OP999451 |
| Sticta sublimbata | JR16367a | sublimbata2 | Taita Hills | Vuria | OP999454 |
| Sticta sublimbata | JR16367b | sublimbata2 | Taita Hills | Vuria | OP999455 |
| Sticta sublimbata | JR16373 | sublimbata4 | Taita Hills | Vuria | OP999458 |
| Sticta sublimbata | JR16374a | sublimbata1 | Taita Hills | Vuria | OP999459 |
| Sticta sublimbata | JR16374c | sublimbata2 | Taita Hills | Vuria | OP999461 |
| Sticta sublimbata | JR16417 | sublimbata3 | Taita Hills | Mbololo | OP999463 |
| Sticta sublimbata | JR16419 | sublimbata7 | Taita Hills | Mbololo | OP999464 |
| Sticta sublimbata | JR16443 | sublimbata8 | Taita Hills | Mbololo | OP999466 |
| Sticta sublimbata | UK110502 | - | Taita Hills | Ngangao | - |
| Sticta sublimbata | UK110510a | - | Taita Hills | Vuria | - |
| Sticta sublimbata | UK110520a | - | Taita Hills | Vuria | - |
| Sticta sublimbata | UK110539d | - | Taita Hills | Vuria | - |
| Sticta sublimbata | UK110547a | sublimbata1 | Taita Hills | Fururu | OP999473 |
| Sticta sublimbata | UK110547b | sublimbata1 | Taita Hills | Fururu | OP999474 |
| Sticta sublimbata | UK110554a | - | Taita Hills | Chawia | - |
| Sticta sublimbata | UK110556a | - | Taita Hills | Chawia | - |
| Sticta sublimbata | UK160482e | - | Kilimanjaro | Home1 | - |
| Sticta sublimbata | UK160552m | - | Kilimanjaro | Home3 | - |
| Sticta sublimbata | UK160557f | - | Kilimanjaro | Home3 | - |
| Sticta sublimbata | UK160559b | - | Kilimanjaro | Home3 | - |
| Sticta tomentosa | JR10086 | tomentosa1 | Taita Hills | Vuria | OP999400 |
| Sticta tomentosa | JR10095A | tomentosa1 | Taita Hills | Vuria | OP999401 |
| Sticta tomentosa | JR10260 | tomentosa1 | Taita Hills | Mwachora | OP999426 |
| Sticta tomentosa | JR16352 | tomentosa1 | Taita Hills | Vuria | OP999449 |
| Sticta tomentosa | JR16367c | tomentosa1 | Taita Hills | Vuria | OP999456 |
| Sticta tomentosa | JR16368 | tomentosa1 | Taita Hills | Vuria | OP999457 |
| Sticta tomentosa | UK110523b | tomentosa1 | Taita Hills | Vuria | OP999472 |
| Sticta tomentosa | UK170912c | tomentosa1 | Kilimanjaro | FOD5 | OP999517 |
| Sticta tomentosa | UK170934c | - | Kilimanjaro | Flm1 | - |
| Sticta tomentosa | UK170936f | - | Kilimanjaro | Flm1 | - |
| Sticta tomentosa | UK170943c | - | Kilimanjaro | Flm1 | - |
| Sticta tomentosa | UK170950b | - | Kilimanjaro | Flm1 | - |
| Sticta tomentosa | UK171473j | - | Kilimanjaro | FPD1 | - |
| Sticta umbilicariiformis | JR16098 | umbilicariiformis1 | Mt. Elgon | Erica zone | OP999434 |
| Sticta umbilicariiformis | JR16102 | umbilicariiformis3 | Mt. Elgon | Erica zone | OP999435 |
| Sticta umbilicariiformis | JR16108 | umbilicariiformis1 | Mt. Elgon | Erica zone | OP999438 |
| Sticta umbilicariiformis | UK170774k | umbilicariiformis3 | Kilimanjaro | FPo1 | OP999478 |
| Sticta umbilicariiformis | UK170775a | umbilicariiformis3 | Kilimanjaro | FPo1 | OP999479 |
| Sticta umbilicariiformis | UK170794c | - | Kilimanjaro | FOc2 | - |
| Sticta umbilicariiformis | UK170806m | umbilicariiformis3 | Kilimanjaro | FPD4 | OP999492 |
| Sticta umbilicariiformis | UK170808c | umbilicariiformis3 | Kilimanjaro | FPD4 | OP999493 |
| Sticta umbilicariiformis | UK170821d | - | Kilimanjaro | FPD4 | - |
| Sticta umbilicariiformis | UK170821h | umbilicariiformis1 | Kilimanjaro | FPD4 | OP999495 |
| Sticta umbilicariiformis | UK170826f | - | Kilimanjaro | FPD4 | - |
| Sticta umbilicariiformis | UK170842d | umbilicariiformis3 | Kilimanjaro | FPD3 | OP999497 |
| Sticta umbilicariiformis | UK170846d | umbilicariiformis3 | Kilimanjaro | FPD3 | OP999500 |
| Sticta umbilicariiformis | UK170858d | umbilicariiformis3 | Kilimanjaro | FPo4 | OP999504 |
| Sticta umbilicariiformis | UK171102b | umbilicariiformis1 | Kilimanjaro | FED5 | OP999531 |
| Sticta umbilicariiformis | UK171106b | umbilicariiformis2 | Kilimanjaro | FED5 | OP999532 |
| Sticta umbilicariiformis | UK171119b | umbilicariiformis5 | Kilimanjaro | FED5 | OP999533 |
| Sticta umbilicariiformis | UK171131a | umbilicariiformis5 | Kilimanjaro | FED4 | OP999534 |
| Sticta umbilicariiformis | UK171131b | umbilicariiformis1 | Kilimanjaro | FED4 | OP999535 |
| Sticta umbilicariiformis | UK171320e | umbilicariiformis4 | Kilimanjaro | FED1 | OP999541 |
| Sticta umbilicariiformis | UK171320f | umbilicariiformis4 | Kilimanjaro | FED1 | OP999542 |
| Sticta umbilicariiformis | UK171320g | umbilicariiformis4 | Kilimanjaro | FED1 | OP999543 |
| Sticta umbilicariiformis | UK171337 | umbilicariiformis1 | Kilimanjaro | FED1 | OP999544 |
| Sticta umbilicariiformis | UK171340s | - | Kilimanjaro | FED1 | - |
| Sticta umbilicariiformis | UK171404e | umbilicariiformis1 | Kilimanjaro | FEr4 | OP999546 |
| Sticta umbilicariiformis | UK171405f | umbilicariiformis1 | Kilimanjaro | FEr4 | OP999547 |
| Sticta umbilicariiformis | UK171411c | umbilicariiformis4 | Kilimanjaro | FEr4 | OP999550 |
| Sticta umbilicariiformis | UK171429 | umbilicariiformis4 | Kilimanjaro | FEr3 | OP999551 |
| Sticta umbilicariiformis | UK171433a | - | Kilimanjaro | FEr3 | - |
| Sticta umbilicariiformis | UK171435f | - | Kilimanjaro | FEr3 | - |
| Sticta umbilicariiformis | UK171438h | umbilicariiformis4 | Kilimanjaro | FEr2 | OP999553 |
| Sticta umbilicariiformis | UK171439u | umbilicariiformis1 | Kilimanjaro | FEr2 | OP999555 |
| Sticta umbilicariiformis | UK171442b | - | Kilimanjaro | FEr2 | - |
| Sticta umbilicariiformis | UK171449i | umbilicariiformis4 | Kilimanjaro | FEr2 | OP999556 |
| Sticta umbilicariiformis | UK171455h | umbilicariiformis1 | Kilimanjaro | FEr2 | OP999557 |
| Sticta umbilicariiformis | UK171458h | - | Kilimanjaro | FEr4 | - |
| Sticta umbilicariiformis | UK171467k | - | Kilimanjaro | FEr4 | - |
| Sticta umbilicariiformis | UK171468e | umbilicariiformis3 | Kilimanjaro | FPD1 | OP999559 |
| Sticta umbilicariiformis | UK171468f | umbilicariiformis3 | Kilimanjaro | FPD1 | OP999560 |
| Sticta umbilicariiformis | UK171494k | - | Kilimanjaro | FOc3 | - |
| Sticta umbilicariiformis | UK171514c | umbilicariiformis3 | Kilimanjaro | FOc5 | OP999583 |
| Sticta umbilicariiformis | UK171521f | umbilicariiformis3 | Kilimanjaro | FOc5 | OP999588 |
| Sticta umbilicariiformis | UK171524b | umbilicariiformis3 | Kilimanjaro | FPo2 | OP999589 |
| Sticta umbilicariiformis | UK171563a | umbilicariiformis2 | Kilimanjaro | FED2 | OP999593 |
| Sticta umbilicariiformis | UK171569d | umbilicariiformis4 | Kilimanjaro | FED2 | OP999594 |
| Sticta umbilicariiformis | UK171577h | umbilicariiformis1 | Kilimanjaro | FPD2 | OP999596 |
| Sticta umbilicariiformis | UK171578t | - | Kilimanjaro | FPD2 | - |
| Sticta umbilicariiformis | UK171584x | umbilicariiformis2 | Kilimanjaro | FPD2 | OP999602 |
| Sticta umbilicariiformis | UK171584y | umbilicariiformis1 | Kilimanjaro | FPD2 | OP999603 |
| Sticta umbilicariiformis | UK171590j | umbilicariiformis3 | Kilimanjaro | FPo3 | OP999608 |
| Sticta sp. A (fuliginoides agg.) | UK170858f | A1 | Kilimanjaro | FPo4 | OP999506 |
| Sticta sp. A (fuliginoides agg.) | UK170884b | A1 | Kilimanjaro | FPo5 | OP999508 |
| Sticta sp. A (fuliginoides agg.) | UK171519h | A1 | Kilimanjaro | FOc5 | OP999586 |
| Sticta sp. A (fuliginoides agg.) | UK171521g | - | Kilimanjaro | FOc5 | - |
| Sticta sp. B (ciliata agg.) | JR09D8A | B1 | Taita Hills | Ngangao | OP999386 |
| Sticta sp. B (ciliata agg.) | JR10060C | B2 | Taita Hills | Shomoto Hill | OP999396 |
| Sticta sp. B (ciliata agg.) | UK170792q | B1 | Kilimanjaro | FOc2 | OP999485 |
| Sticta sp. B (ciliata agg.) | UK170901b | B1 | Kilimanjaro | FOD4 | OP999514 |
| Sticta sp. C (weigelii agg.) | JR090X3 | C1 | Taita Hills | Chawia | OP999380 |
| Sticta sp. C (weigelii agg.) | JR10180B | C1 | Taita Hills | Yale | OP999411 |
| Sticta sp. C (weigelii agg.) | JR10180C | - | Taita Hills | Yale | - |
| Sticta sp. C (weigelii agg.) | JR10212B | C1 | Taita Hills | Yale | OP999419 |
| Sticta sp. C (weigelii agg.) | JR14634/082 | - | Taita Hills | Maktau Hill | - |
| Sticta sp. C (weigelii agg.) | JR16201a | C2 | Taita Hills | Fururu | OP999439 |
| Sticta sp. C (weigelii agg.) | JR16201b | C2 | Taita Hills | Fururu | OP999440 |
| Sticta sp. C (weigelii agg.) | JR16295 | C1 | Taita Hills | Sagalla | OP999444 |
| Sticta sp. C (weigelii agg.) | JR16310 | C1 | Taita Hills | Sagalla | OP999445 |
| Sticta sp. C (weigelii agg.) | JR16439 | C1 | Taita Hills | Mbololo | OP999465 |
| Sticta sp. C (weigelii agg.) | JR16462 | C1 | Taita Hills | Mbololo | OP999467 |
| Sticta sp. C (weigelii agg.) | UK110555e | - | Taita Hills | Chawia | - |
| Sticta sp. D | UK171519k | D1 | Kilimanjaro | FOc5 | OP999587 |
| Sticta sp. D | UK171586q | D1 | Kilimanjaro | FPo3 | OP999606 |
| Sticta sp. 2 | JR09Y12b | - | Taita Hills | Mbololo | - |
| Sticta sp. 2 | JR09Y13 | - | Taita Hills | Mbololo | - |
| Sticta sp. 2 | JR10K302 | sp. 2 | Mt. Kasigau | Plot E15 | OP999430 |
| Sticta sp. 2 | JR10K303 | sp. 2 | Mt. Kasigau | Plot E15 | OP999431 |
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Merckx, V.S.F.T.; Hendriks, K.P.; Beentjes, K.K.; Mennes, C.B.; Becking, L.E.; Peijnenburg, K.T.C.A.; Afendy, A.; Arumugam, N.; de Boer, H.; Biun, A.; et al. Evolution of endemism on a young tropical mountain. Nature 2015, 524, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Hemp, C.; Kehl, S.; Schultz, O.; Wägele, J.W.; Hemp, A. Climatic fluctuations and orogenesis as motors for speciation in East Africa: Case study on Parepistaurus Karsch, 1896 (Orthoptera). Syst. Entomol. 2015, 40, 17–34. [Google Scholar] [CrossRef]
- Schwery, O.; Onstein, R.E.; Bouchenak-Khelladi, Y.; Xing, Y.; Carter, R.J.; Linder, H.P. As old as the mountains: The radiations of the Ericaceae. New Phytol. 2015, 207, 355–367. [Google Scholar] [CrossRef]
- Griffiths, C.J. The ecological evolution of East Africa. In Biogeography and Ecology of the Rainforests of Eastern Africa; Lovett, J.C., Wasser, S.K., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 9–21. ISBN 0-521-43083-6. [Google Scholar]
- Wasser, S.K.; Lovett, J.C. (Eds.) Introduction to the biography and ecology of the rain forests of eastern Africa. In Biogeography and Ecology of the Rainforests of Eastern Africa; Cambridge University Press: Cambridge, UK, 1993; pp. 3–8. ISBN 0-521-43083-6. [Google Scholar]
- Lovett, J.C. Eastern Arc moist forest flora. In Biogeography and Ecology of the Rainforests of Eastern Africa; Lovett, J.C., Wasser, S.K., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 33–55. ISBN 0-521-43083-6. [Google Scholar]
- Hemp, A.; Hemp, C. Broken bridges: The isolation of Kilimanjaro’s ecosystem. Glob. Chang. Biol. 2018, 24, 3499–3507. [Google Scholar] [CrossRef]
- Hemp, C.; Grzywacz, B.; Warchałowska-Śliwa, E.; Hemp, A. Topography and climatic fluctuations boosting speciation: Biogeography and a molecular phylogeny of the East African genera Afroanthracites Hemp & Ingrisch and Afroagraecia Ingrisch & Hemp (Orthoptera, Tettigoniidae, Conocephalinae, Agraeciini). Org. Divers. Evol. 2016, 16, 211–223. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Robles, G.P.; Hoffmann, M.; Pilgrim, J.; Brooks, T.; Mittermeier, C.G.; Lamoreux, J.; Da Fonseca, G.A.B. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Ecoregions; CEMEX: Mexico City, Mexico, 2004; ISBN 968-6397-77-9. [Google Scholar]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global Biodiversity Conservation: The Critical Role of Hotspots. In Biodiversity Hotspots; Zachos, F.E., Habel, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar] [CrossRef]
- Swinscow, T.D.V.; Krog, H. Macrolichens of East Africa; Natural History Museum Publications: London, UK, 1988; ISBN 0565010395. [Google Scholar]
- Kaasalainen, U.; Tuovinen, V.; Kirika, P.; Mollel, N.; Hemp, A.; Rikkinen, J. Diversity of Leptogium (Collemataceae, Ascomycota) in East African Montane Ecosystems. Microorganisms 2021, 9, 314. [Google Scholar] [CrossRef]
- Kirika, P.M.; Divakar, P.K.; Buaruang, K.; Leavitt, S.D.; Crespo, A.; Gatheri, G.W.; Mugambi, G.; Benatti, M.N.; Lumbsch, H.T. Molecular phylogenetic studies unmask overlooked diversity in the tropical lichenized fungal genus Bulbothrix s.l. (Parmeliaceae, Ascomycota). Bot. J. Linn. Soc. 2017, 184, 387–399. [Google Scholar] [CrossRef]
- Kirika, P.M.; Divakar, P.K.; Crespo, A.; Leavitt, S.D.; Mugambi, G.; Gatheri, G.W.; Lumbsch, H.T. Polyphyly of the genus Canoparmelia—Uncovering incongruences between phenotype-based classification and molecular phylogeny within lichenized Ascomycota (Parmeliaceae). Phytotaxa 2016, 289, 36–48. [Google Scholar] [CrossRef]
- Kirika, P.M.; Divakar, P.K.; Crespo, A.; Lumbsch, H.T. Molecular and phenotypical studies on species diversity of Hypotrachyna (Parmeliaceae, Ascomycota) in Kenya, East Africa. Bryologist 2019, 122, 140–150. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Biermann, L.; Mollel, N.P.; Schmidt, A.R.; Hemp, A. Peltigera (Lecanoromycetes) on Mt Kilimanjaro, East Africa. Lichenologist 2022, 54, 231–243. [Google Scholar] [CrossRef]
- Suija, A.; Kaasalainen, U.; Kirika, P.M.; Rikkinen, J. Taitaia, a novel lichenicolous fungus in tropical montane forests in Kenya (East Africa). Lichenologist 2018, 50, 173–184. [Google Scholar] [CrossRef]
- Moncada, B.; Lücking, R.; Suárez, A. Molecular phylogeny of the genus Sticta (lichenized Ascomycota: Lobariaceae) in Colombia. Fungal Divers. 2014, 64, 205–231. [Google Scholar] [CrossRef]
- Magain, N.; Sérusiaux, E. Dismantling the treasured flagship lichen Sticta fuliginosa (Peltigerales) into four species in Western Europe. Mycol. Prog. 2015, 14, 125. [Google Scholar] [CrossRef]
- Moncada, B.; Mercado-Díaz, J.A.; Smith, C.W.; Bungartz, F.; Sérusiaux, E.; Lumbsch, H.T.; Lücking, R. Two new common, previously unrecognized species in the Sticta weigelii morphodeme (Ascomycota: Peltigeraceae). Willdenowia 2021, 51, 35–45. [Google Scholar] [CrossRef]
- Ossowska, E.A.; Moncada, B.; Kukwa, M.; Flakus, A.; Rodriguez-Flakus, P.; Olszewska, S.; Lücking, R. New species of Sticta (lichenised Ascomycota, lobarioid Peltigeraceae) from Bolivia suggest a high level of endemism in the Central Andes. Mycokeys 2022, 92, 131–160. [Google Scholar] [CrossRef]
- Mercado-Díaz, J.A.; Lücking, R.; Moncada, B.; Widhelm, T.J.; Lumbsch, H.T. Elucidating species richness in lichen fungi: The genus Sticta (Ascomycota: Peltigeraceae) in Puerto Rico. Taxon 2020, 69, 851–891. [Google Scholar] [CrossRef]
- Widhelm, T.J.; Bertoletti, F.R.; Asztalos, M.J.; Mercado-Díaz, J.A.; Huang, J.-P.; Moncada, B.; Lücking, R.; Magain, N.; Sérusiaux, E.; Goffinet, B.; et al. Oligocene origin and drivers of diversification in the genus Sticta (Lobariaceae, Ascomycota). Mol. Phylogenetics Evol. 2018, 126, 58–73. [Google Scholar] [CrossRef]
- Simon, A.; Goffinet, B.; Magain, N.; Sérusiaux, E. High diversity, high insular endemism and recent origin in the lichen genus Sticta (lichenized Ascomycota, Peltigerales) in Madagascar and the Mascarenes. Mol. Phylogenetics Evol. 2018, 122, 15–28. [Google Scholar] [CrossRef]
- Kirika, P.; Mugambi, G.; Lücking, R.; Lumbsch, H.T. New Records of Lichen-Forming Fungi from Kenya. J. East Afr. Nat. Hist. 2012, 101, 73–98. [Google Scholar] [CrossRef]
- Frisch, A.; Hertel, H. Flora of macrolichens in the alpine and subalpine zones of Mount Kenya (Kenya). Sauteria 1998, 9, 363–370. [Google Scholar]
- Alstrup, V.; Christensen, S.N. New records of lichens with cyanobacteria from Tanzania and Kenya. Cryptogam. Mycol. 2006, 27, 57–68. [Google Scholar]
- Kirika, P.M.; Ndiritu, G.G.; Mugambi, G.K.; Newton, L.E.; Lumbsch, H.T. Diversity and Altitudinal Distribution of Understorey Corticolous lichens in a tropical montane forest in Kenya (East Africa). Crypt. Biodivers. Assess. 2018, 2018, 47–70. [Google Scholar] [CrossRef]
- Krog, H. Corticolous macrolichens of low montane rainforests and moist woodlands of eastern Tanzania. Sommerfeltia 2000, 28, 1. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Tuovinen, V.; Mwachala, G.; Pellikka, P.; Rikkinen, J. Complex Interaction Networks Among Cyanolichens of a Tropical Biodiversity Hotspot. Front. Microbiol. 2021, 12, 672333. [Google Scholar] [CrossRef]
- Galloway, D.J.; Thomas, M.A. Sticta. In Lichen Flora of the Greater Sonoran Desert Region; Nash, T.H., III, Ryan, B.D., Diederich, P., Gries, C., Bungartz, F., Eds.; Lichens Unlimited, Arizona State University: Tempe, AZ, USA, 2004; Volume 2, pp. 513–524. ISBN 0971675910. [Google Scholar]
- Galloway, D.J. Studies on lichen genus Sticta (Schreber) Ach.: III*. Notes on species described by Bory de St-Vincent, William Hooker, and Delise, between 1804 and 1825. Nova Hedwig. 1995, 61, 147–188. [Google Scholar]
- Rutten, G.; Ensslin, A.; Hemp, A.; Fischer, M. Vertical and Horizontal Vegetation Structure across Natural and Modified Habitat Types at Mount Kilimanjaro. PLoS ONE 2015, 10, e0138822. [Google Scholar] [CrossRef]
- Bytebier, B. Taita Hills Biodiversity Project Report; National Museums of Kenya: Nairobi, Kenya, 2001. [Google Scholar]
- Pellikka, P.K.E.; Lötjönen, M.; Siljander, M.; Lens, L. Airborne remote sensing of spatiotemporal change (1955–2004) in indigenous and exotic forest cover in the Taita Hills, Kenya. Int. J. Appl. Earth Obs. Geoinf. 2009, 11, 221–232. [Google Scholar] [CrossRef]
- Beentje, H.J. An ecological and floristical study of the forests of the Taita Hills, Kenya. Utafiti 1988, 1, 23–66. [Google Scholar]
- Aerts, R.; Matthijsen, E.; Lehouck, V.; Beentje, H.; Bytebier, B.; Matthysen, E.; Gulinck, H.; Lens, L.; Muys, B. Woody plant communities of isolated Afromontane cloud forests in Taita Hills, Kenya. Plant Ecol. 2011, 212, 639–649. [Google Scholar] [CrossRef]
- Thijs, K.W.; Roelen, I.; Musila, W.M. Field Guide to the Woody Plants of Taita Hills, Kenya. J. East Afr. Nat. Hist. 2014, 102, 1–272. [Google Scholar] [CrossRef]
- Medley, K.E.; Maingi, J.K. Biogeographic Patterns of Forest Diversity at Mount Kasigau, Kenya. J. East Afr. Nat. Hist. 2015, 103, 1–24. [Google Scholar] [CrossRef]
- Henkin, M.A.; Medley, K.E.; Maingi, J.K. Biophysical analysis of afromontane forest community types at Mount Kasigau, Kenya. Afr. J. Ecol. 2015, 53, 454–464. [Google Scholar] [CrossRef]
- Enroth, J.; Nyqvist, P.; Malombe, I.; Pellikka, P.; Rikkinen, J. Additions to the moss flora of the Taita Hills and Mount Kasigau, Kenya. Pol. Bot. J. 2013, 58, 495–510. [Google Scholar] [CrossRef]
- Moncada, B.; Lücking, R. Ten new species of Sticta and counting: Colombia as a hot spot for unrecognized diversification in a conspicuous macrolichen genus. Phytotaxa 2012, 74, 1–29. [Google Scholar] [CrossRef]
- Fedrowitz, K.; Kaasalainen, U.; Rikkinen, J. Genotype variability of Nostoc symbionts associated with three epiphytic Nephroma species in a boreal forest landscape. Bryologist 2011, 114, 220–230. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PRC Protcols: A guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: London, UK, 1990; pp. 315–322. ISBN 0-12-372181-4. [Google Scholar]
- CodonCode Corporation. CodonCode Aligner v8; CodonCode Corporation: Centerville, UT, USA, 2018. [Google Scholar]
- National Center for Biotechnology Information. NCBI GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 9 October 2022).
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Müller, K.; Müller, J.; Quandt, D. PhyDE. Available online: http://www.phyde.de/ (accessed on 23 November 2022).
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian Inference of Phylogeny and Its Impact on Evolutionary Biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Larget, B.; Miller, R.E.; Ronquist, F. Potential Applications and Pitfalls of Bayesian Inference of Phylogeny. Syst. Biol. 2002, 51, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Ossowska, E.A. First records of Sticta weigelii s.str. from Bolivia confirmed by molecular data. Folia Cryptogam. Est. 2021, 58, 65–72. [Google Scholar] [CrossRef]
- Galloway, D.J. Studies on the lichen genus Sticta (Schreber) Ach.: V*. Australian species. Bryophyt. Divers. Evol. 1998, 15, 117–160. [Google Scholar] [CrossRef]
- Moncada, B.; Smith, C.W.; Lücking, R. A taxonomic reassessment of the genus Sticta (lichenized Ascomycota: Peltigeraceae) in the Hawaiian archipelago. Lichenologist 2021, 53, 117–133. [Google Scholar] [CrossRef]
- Nylander, W. Lichenes in Regionibus Exoticis Quibusnam Vigentes. Ann. Sci. Nat. Bot. 1859, 11, 205–264. [Google Scholar]
- Lindgren, H.; Moncada, B.; Lücking, R.; Magain, N.; Simon, A.; Goffinet, B.; Sérusiaux, E.; Nelsen, M.P.; Mercado-Díaz, J.A.; Widhelm, T.J.; et al. Cophylogenetic patterns in algal symbionts correlate with repeated symbiont switches during diversification and geographic expansion of lichen-forming fungi in the genus Sticta (Ascomycota, Peltigeraceae). Mol. Phylogenetics Evol. 2020, 150, 106860. [Google Scholar] [CrossRef]
- Magain, N.; Goffinet, B.; Sérusiaux, E. Further photomorphs in the lichen family Lobariaceae from Reunion (Mascarene archipelago) with notes on the phylogeny of Dendriscocaulon cyanomorphs. Bryol. 2012, 115, 243–254. [Google Scholar] [CrossRef]
- Hue, A.M. Lichens récoltés á Java en 1894-1895 par M. Jean Massart. I. Ann. Du Jard. Bot. De Buitenzorg 1901, 11, 171–194. [Google Scholar]
- Nylander, W. Synopsis Methodica Lichenum; Martinet: Paris, France, 1858. [Google Scholar]
- Galloway, D.J. Studies on the lichen genus Sticta (Schreber) Ach.: II. Typification of taxa from Swartz’s Prodromus of 1788. Bull. Nat. Hist. Mus. Lond. 1994, 24, 35–48. [Google Scholar]
- Flotow, J.V. Lichenen des Reisevereins und des Hochstetterschen Herbarii. Linnaea: Ein Journal für die Botanik in ihrem ganzen Umfange 1843, 17, 15–30. [Google Scholar]
- Moncada, B.; Luecking, R.K.; Lumbsch, H.T. Rewriting the evolutionary history of the lichen genus Sticta (Ascomycota: Peltigeraceae subfam. Lobarioideae) in the Hawaiian islands. Plant Fungal Syst. 2020, 65, 95–119. [Google Scholar] [CrossRef]
- Galloway, D.J. Notes on the holotype of Sticta damaecornis β weigelii Ach. (=Sticta weigelii). Lichenologist 2006, 38, 89–92. [Google Scholar] [CrossRef]
- Aptroot, A.; Stech, M. An updated checklist of the lichens of St. Eustatius, Netherlands Antilles. Mycokeys 2018, 33, 69–84. [Google Scholar] [CrossRef]
- Delise, D.F. Histoire des Lichens, Genre Sticta; Harvard University: Cambridge, MA, USA, 1825; Volume 1—2, 167. [Google Scholar]
- Benítez, Á.; Prieto, M.; Gonzáles, Y.; Aragón, G. Effects of tropical montane forest disturbance on epiphytic macrolichens. Sci. Total Env. 2012, 441, 169–175. [Google Scholar] [CrossRef]
- Tripp, E.A.; Lendemer, J.C.; McCain, C.M. Habitat quality and disturbance drive lichen species richness in a temperate biodiversity hotspot. Oecologia 2019, 190, 445–457. [Google Scholar] [CrossRef]
- Rikkinen, J. Cyanolichens. Biodivers. Conserv. 2015, 24, 973–993. [Google Scholar] [CrossRef]
Figure 1.
Clades with Sticta tomentosa and S. sublimbata of the Bayesian tree of the genus Sticta based on the nuITS marker region (Figure S1A,B). The colored polygons (rectangle, triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment and on Mt. Kasigau in one sampling transect. The grey boxes show the total abundance and distribution of the species, also including the data from unsequenced specimens. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.
Figure 1.
Clades with Sticta tomentosa and S. sublimbata of the Bayesian tree of the genus Sticta based on the nuITS marker region (Figure S1A,B). The colored polygons (rectangle, triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment and on Mt. Kasigau in one sampling transect. The grey boxes show the total abundance and distribution of the species, also including the data from unsequenced specimens. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.

Figure 2.
Phylogeny of the Sticta umbilicariiformis—fuliginosa group based on the nuITS region. The colored polygons (rectangle or triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment and on Mt. Kasigau in one sampling transect. The grey boxes show the total abundance and distribution of the species, also including the data from unsequenced specimens. The proportions of different structures (pustules, isidia, or just apothecia) among the specimens with specific ITS variants are indicated with pie charts for the new species and S. umbilicariiformis. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.
Figure 2.
Phylogeny of the Sticta umbilicariiformis—fuliginosa group based on the nuITS region. The colored polygons (rectangle or triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment and on Mt. Kasigau in one sampling transect. The grey boxes show the total abundance and distribution of the species, also including the data from unsequenced specimens. The proportions of different structures (pustules, isidia, or just apothecia) among the specimens with specific ITS variants are indicated with pie charts for the new species and S. umbilicariiformis. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.

Figure 3.
Clades with Sticta fuliginoides, S. ciliata, Sticta sp. A, and Sticta sp. B of the Bayesian tree of the genus Sticta based on the nuITS marker region (Figure S1C). The colored polygons (rectangle, triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment. The grey boxes show the total abundance and distribution of the species, also including the data from unsequenced specimens. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.
Figure 3.
Clades with Sticta fuliginoides, S. ciliata, Sticta sp. A, and Sticta sp. B of the Bayesian tree of the genus Sticta based on the nuITS marker region (Figure S1C). The colored polygons (rectangle, triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment. The grey boxes show the total abundance and distribution of the species, also including the data from unsequenced specimens. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.

Figure 4.
Clades with Sticta cellulosa, S. andina, Sticta sp. C (weigelii agg.), S. duplolimbata, and Sticta sp. D of the Bayesian tree of the genus Sticta based on the nuITS marker region (Figure S1D,E). The colored polygons (rectangle, triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment. The grey boxes show the total abundance and distribution of the species, including also the data from unsequenced specimens. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.
Figure 4.
Clades with Sticta cellulosa, S. andina, Sticta sp. C (weigelii agg.), S. duplolimbata, and Sticta sp. D of the Bayesian tree of the genus Sticta based on the nuITS marker region (Figure S1D,E). The colored polygons (rectangle, triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment. The grey boxes show the total abundance and distribution of the species, including also the data from unsequenced specimens. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.

Figure 5.
Clades with Sticta marginalis, Sticta sp. 2, and Sticta cyanocaperata in the Bayesian tree of the genus Sticta based on the nuITS marker region (Figure S1F). The colored polygons (rectangle, triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment and on Mt. Kasigau in one sampling transect. The grey boxes show the total abundance and distribution of the species, also including the data from unsequenced specimens. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.
Figure 5.
Clades with Sticta marginalis, Sticta sp. 2, and Sticta cyanocaperata in the Bayesian tree of the genus Sticta based on the nuITS marker region (Figure S1F). The colored polygons (rectangle, triangle) in the tree show the distribution of the ITS variants in the studied regions and ecosystem types: On Mt. Kilimanjaro, the different habitats are indicated by color and grid; the width of the rectangle indicate the number of sample plots in which the taxon was present in each ecosystem type (square = 1). In the Taita Hills, each triangle indicates presence in one forest fragment and on Mt. Kasigau in one sampling transect. The grey boxes show the total abundance and distribution of the species, also including the data from unsequenced specimens. Stronger support (PP > 0.9) for a clade is indicated with a thicker branch. The scale refers to nucleotide substitutions per site.

Figure 14.
Diversity and abundance of the genus Sticta in the study region. (a) Number of species (dark red) and abundance (light orange) in the different habitats and locations. D = disturbed. (b) The abundance of the Sticta species in different elevation zones and habitat types. Low elevation habitats: dry woodland on Mt. Kasigau, the Maktau Hill, Shomoto Hill in the Taita Hills, and coffee farm and home garden habitats on Mt. Kilimanjaro; Lower montane forests: most evergreen forests in the Taita Hills and on Mt. Kasigau, and lower montane forest on Mt. Kilimanjaro. Middle montane forests: middle montane forests on Mt. Kilimanjaro and on Vuria Mountain in the Taita Hills. Upper montane forest: such forests on Mt. Kilimanjaro. Subalpine: subalpine habitats on Mt. Kilimanjaro and Mt. Elgon. Sticta ciliata agg. includes specimens of S. ciliata and Sticta sp. B; Sticta fuliginoides agg. includes specimens of S. fuliginoides and Sticta sp. A.
Figure 14.
Diversity and abundance of the genus Sticta in the study region. (a) Number of species (dark red) and abundance (light orange) in the different habitats and locations. D = disturbed. (b) The abundance of the Sticta species in different elevation zones and habitat types. Low elevation habitats: dry woodland on Mt. Kasigau, the Maktau Hill, Shomoto Hill in the Taita Hills, and coffee farm and home garden habitats on Mt. Kilimanjaro; Lower montane forests: most evergreen forests in the Taita Hills and on Mt. Kasigau, and lower montane forest on Mt. Kilimanjaro. Middle montane forests: middle montane forests on Mt. Kilimanjaro and on Vuria Mountain in the Taita Hills. Upper montane forest: such forests on Mt. Kilimanjaro. Subalpine: subalpine habitats on Mt. Kilimanjaro and Mt. Elgon. Sticta ciliata agg. includes specimens of S. ciliata and Sticta sp. B; Sticta fuliginoides agg. includes specimens of S. fuliginoides and Sticta sp. A.

Table 1.
Summary of the genus Sticta in East Africa, including Ethiopia (E), Kenya (K), Rwanda (R), Tanzania (T), and Uganda (U). Distributions are reported according to our observations and previous studies listed in References. The ecological notes are based on our observations.
Table 1.
Summary of the genus Sticta in East Africa, including Ethiopia (E), Kenya (K), Rwanda (R), Tanzania (T), and Uganda (U). Distributions are reported according to our observations and previous studies listed in References. The ecological notes are based on our observations.
| Species | Distribution | Ecology | Comments | References |
|---|---|---|---|---|
| Sticta afromontana sp. nov. | K*, T* | Common especially in middle and upper montane forests. | ||
| Sticta ambavillaria | - | Not found in this study. | Previous reports may represent S. munda. | [12,27,28,29,30] |
| Sticta andina | K* | Middle montane forest (single observation). | ||
| Sticta aspratilis sp. nov. | K*, T* | Lower montane to subalpine zones. | ||
| Sticta cellulosa sp. nov. | T* | Rare in upper montane and subalpine zones. | ||
| Sticta ciliata | K*, T* | Rare in lower montane forest zone. | Previous report from Rwanda [20] represents Sticta sp. B. | |
| Sticta cyanocaperata sp. nov. | T* | Uncommon in middle montane forest. | ||
| Sticta cyphellulata | - | Not found in this study. | Previous reports may represent S. duplolimbata and/or S. marginalis. | [12,28,30] |
| Sticta dichotoma | T | Not found in this study. | [12,28,30] | |
| Sticta duplolimbata | K*, T*, R | Common in lower and middle montane forests. | [20] | |
| Sticta fuliginoides | T* | Common especially in upper montane forests. | ||
| Sticta fuliginosa | K, T, R | Common in lower and middle montane forests. | Some previous reports may represent other species of S. fuliginosa morphodeme. | [12,20,28,29,30,31] |
| Sticta kunthii | - | Not found in this study. | Previous report may represent S. umbilicariiformis and/or S. aspratilis. | [26] |
| Sticta limbata | - | Not found in this study. | Previous reports represent S. umbilicariiformis [20]. | [12,27] |
| Sticta marginalis | K*, T*, U* | Common in lower and middle montane forests. | Presence in Uganda based on previous reports of S. orbicularis [12,28,30]. | |
| Sticta munda sp. nov. | T* | Rare in middle and upper montane forests. | ||
| Sticta orbicularis | - | Not found in this study. | Previous reports may represent S. marginalis. | [12,28,30] |
| Sticta papyracea/variabilis | - | Not found in this study. | Sticta papyracea and S. variabilis are synonymous [33]. The previous reports may represent Sticta sp. 2. | [12,30] |
| Sticta sublimbata | E, K, T, U | Common in low elevation and lower montane habitats. | [12,30,31] | |
| Sticta tomentosa | K, T, U | Lower and middle montane forests. | [12,28,30,31] | |
| Sticta umbilicariiformis | E, K, T, R, U | Abundant in the upper montane and subalpine zones. | Presence in Uganda based on previous reports of S. limbata [12]. | [20] |
| Sticta weigelii | - | Not found in this study. | Previous reports may represent Sticta sp. C. | [12,27,28,29,30,31] |
| Sticta xanthotropa | - | Not found in this study. | Previously reported as S. weigelii var. xanthotropa. The reports may represent other species with marginal flattened isidia. | [12] |
| Sticta sp. A | T* | Rare in middle and upper montane forests. | Sticta fuliginoides agg. | |
| Sticta sp. B | K*, T*, R | Rare in lower and middle montane forests. | Sticta ciliata agg. Previously reported from Rwanda as S. ciliata [20]. | |
| Sticta sp. C | K* | Low elevation and lower montane habitats. | Sticta weigelii agg. May include two distinct taxa. | |
| Sticta sp. D | T* | Rare in middle and upper montane forests. | ||
| Sticta sp. 2 | K*, T | Rare in lower montane forests. | Presence in Tanzania based on previous reports of S. papyracea [12,30]. |
* New record for the region.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kaasalainen, U.; Kirika, P.M.; Mollel, N.P.; Hemp, A.; Rikkinen, J. The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems. J. Fungi 2023, 9, 246. https://doi.org/10.3390/jof9020246
AMA Style
Kaasalainen U, Kirika PM, Mollel NP, Hemp A, Rikkinen J. The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems. Journal of Fungi. 2023; 9(2):246. https://doi.org/10.3390/jof9020246
Chicago/Turabian StyleKaasalainen, Ulla, Paul M. Kirika, Neduvoto P. Mollel, Andreas Hemp, and Jouko Rikkinen. 2023. "The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems" Journal of Fungi 9, no. 2: 246. https://doi.org/10.3390/jof9020246
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.