
New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam

1
Komarov Botanical Institute of the Russian Academy of Sciences, 2 Prof. Popov Str., 197022 Saint Petersburg, Russia
2
Joint Vietnam-Russia Tropical Science and Technology Research Centre, Nguyen Van Huyen, Nghia Do, Cau Giay, Hanoi 122100, Vietnam
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(6), 621; https://doi.org/10.3390/jof9060621
Submission received: 25 April 2023
/
Revised: 24 May 2023
/
Accepted: 25 May 2023
/
Published: 27 May 2023
(This article belongs to the Special Issue Macromycetes of the Northern Hemisphere: Phylogeny, Taxonomy, Diversity, Distribution, Ecology, Conservation)
Abstract
:Four new species of Entoloma from Kon Chu Rang Nature Reserve and Ta Dung National Park were discovered during an investigation of the diversity of the mycobiota of Central Vietnam and are described here on the base of the molecular and morphological data. Phylogenetic analysis was based on nrITS1-5.8S-ITS2, nrLSU and tef1α regions. Illustrated descriptions of their macro- and microscopic features and discussion on similar taxa are given. Entoloma cycneum and E. peristerinum belong to the subgenus Cubospora. They are morphologically similar species and are characterized by white or whitish basidiomata with yellowish or beige tinges and with mainly smooth, glabrous, and hygrophanous pileus, longitudinally fibrillose or fibrillose-scaly white stipe, cuboid spores, and more or less cylindrical cheilocystidia, arising from hymenophoral trama. Entoloma peristerinum posseses initially more coloured beige conical pileus, discolouring to white with age and drying. The pileus of E. cycneum is initially white, hemisphaerical to convex, usually with thin pubescence near the margin. The species can be recognized also by the cheilocystidia form: serrulatum-type in E. cycneum vs. porphyrogriseum-type in E. peristerinum. Another two species belong to the subgenus Leptonia. Entoloma tadungense is close to E. percoelestinum from which it differs by smaller spores with pronounced angles, presence of the cheilocystidia, and the lilac discolouration of the stipe. E. dichroides is named after its similarity to E. dichroum, a dark blue coloured species with pronouncedly angled basidiospores. It is distinguished by the basidiospores form—irregularly 5(–6) angled with elongated apiculus, as well as by absence of the cheilocystidia and darker basidiomata with conical pileus. The article also describes the history of the study of the genus Entoloma in Vietnam with a list of 29 species mentioned in the publications for this country.
1. Introduction
According to estimates for 2018 [1], 21 species of the genus Entoloma (Fr.) P. Kumm. were known from the Central Vietnam, including unpublished data of the authors. In subsequent years, five more new species were described from this territory [2,3,4]. The full history of study of the genus Entoloma in Vietnam with a list of 29 species mentioned in the publications for this country is described in the discussion part. However, the real diversity of the genus is much higher, and many species are still waiting to be described.
As proven by recent molecular genetic studies, the shape of the basidiospores is a key feature in understanding evolution and relatedness in Entolomataceae [5,6,7].
The cuboid and cuboid-like form of spores is remarkable and interesting from a phylogenetic point of view in terms of the multiplicity of its occurrence in the course of evolution. The molecular genetic data [7] supports a clear segregation of clades with cuboid spores from cuboid-like ones (pentagonal, prismatic, pseudocuboid, squamiferum). The species with rhomboid spores also cluster separately in a well-supported clade but they are mixed with species with five or six angled spores [8,9].
Species with “true” cuboid spores (with six, more or less equal, quadrangular faces) traditionally were included in the subgenus/genus Inocephalus [Entoloma subgen. Inocephala Noordel., Inocephalus (Noordel.) P. D. Orton.]. However, the type of Inocephalus, Entoloma inocephalum (Romagnesi) Dennis, described from Madagascar, has 5–7 angled basidiospores in profile view [10]. The material identified as Entoloma inocephalum from Vietnam [11] has the same morphology and is used as a reference collection for the phylogenetic studies while authentic or Madagascar-collected material is not available [7]. It turns out that the /Inocephalus clade occupies an isolated position from the clades with cuboid spores. Therefore, two new subgenera were described to arrange species with such spores—Entoloma subgenus Cubospora Karstedt, Capelari, Largent, T. J. Baroni & Bergemann with E. luteolamellatum (Largent & Aime) Blanco-Dios as a type species (with a conical pileus) and Entoloma subgenus Cuboeccilia Karstedt, Capelari & Largent with E. omphalinoides (Largent) Blanco-Dios as a type species (with depressed pileus) [7]. The number of species with cuboid and cuboid-like spores described world-wide so far is estimated to be 165 [12] or 120 [7].
The first monographic contributions devoted to groups with cuboid spores were made by Horak [13,14,15,16,17], and refer to SE-Asia and South America. The following publications deal with the diversity of cuboid-spored species in different regions, e.g., Asia [11,12,18,19,20,21], Africa/Madagascar [10,22,23,24], Australia/Tasmania/New Zealand [25,26], South and Central America [27,28,29,30], and North America [31,32]. Generalization of accumulated data based on phylogenetic analysis was conducted by F. Karstedt et al. [7]. The analysis of records shows that the centre of diversity of the group with “true” cuboid spores is situated in tropical and subtropical regions of the Old and New Worlds. Two more species of the subgenus Cubospora are described from Vietnam here.
Another two new species belong to the subgenus Leptonia in the modern sense [8]. The multigene phylogeny based on nrITS, nrLSU, and mtSSU [5] showed that subgenus Leptonia in the previously considered volume [32,33,34,35,36] is polyphyletic. Sect. Leptonia of the subgenus belongs to the /Nolanea-Claudopus clade, and Cyanula and Griseorubida to the /Inocephalus-Cyanula clade. Based on these data, Cyanula has been raised to the subgenus level [26]. Recent studies [37,38] have shown that a clade of species around Entoloma ameides (sect. Ameides (Largent) Reschke et al.) previously classified under the subgenus Nolanea now is grouped with the subgenus Leptonia as a sister to the /Leptonia clade with high support. For that reason, the /Ameides clade is treated as a new section of subgenus Leptonia [8]. The subgenus Leptonia now combines species with more or less coloured (predominantly with blue, brown, but also yellow or green tinges) basidiomata, characterized also by the presence of clamp-connections, absence of brilliant granules, and more or less fibrillose to squamulose stipe, and includes around 50 species world-wide [8,26,32,39]; Gasteroid forms also occur [37]. The highest diversity of this group was found in the temperate zone of the Holarctic. In the tropics, representatives of the subgenus are rare, and in Vietnam they are recorded for the first time.
2. Materials and Methods
2.1. Collecting and Site Description
The material for this study was collected during the expeditions of the Joint Vietnam-Russia Tropical Science and Technology Research Centre (VRTC) to the Central Highlands of Vietnam.
The Kon Chu Rang Nature Reserve is located in the north-eastern part of Gia Lai Province of Vietnam (Son Lang Commune, K’Bang District), between 14.50° N–14.58° N and 108.5° E–108.65° E. The area of the reserve is 15,446 hectares, of which 99% are primary and intact forests, which is the highest forest coverage in the country. The relief is hilly to mountainous in the northern part, with heights from 800 to 1452 m a. s. l. (Kon Chu Rang Mt). The average annual temperature is about 21 °C (from 28 °C in May to 12 °C in January). The average annual precipitation is about 1900–2000 mm with the peak in September (340 mm). The dry season is from January to April [40,41]. The main forest type in the reserve is middle-mountain evergreen broad-leaved and mixed forest dominated by Fagaceae (Lithocarpus, Quercus, Castanopsis), Lauraceae, Fabaceae, Clusiaceae, Myrtaceae, Ericaceae, Burseraceae, and Magnoliaceae, mixed with gymnosperms (Dacrycarpus imbricatus, Dacrydium elatum), distributed at elevations between 900 and 1500 m in the north-west of the nature reserve. The first data on the mycobiota of Kon Chu Rang Nature Reserve were published only recently, including those on several subgenera of Entoloma, and on the Boletaceae [3,4,42,43].
Ta Dung National Park is located in the Central Highlands of Vietnam within the administrative boundary of Dak Som commune, Dak G’long district, Dak Nong province, 50 km northeast of Gia Nghia commune’s exam centre. It extends between 11.79° N–11.99° N and 107.89° E–108.11° E, occupying a total area of 20,973.7 hectares. The National Park is located on the Dak Nong Plateau and part of the Di Linh Plateau. Dak Nong Plateau is an arched elevation with an average height of 600–1200 m and has many mountain ranges with an average height of 1200–1500 m, with Ta Dung peak being 1982 m high, the lowest elevation being the land. Ta Dung National Park is situated in an area with a tropical highland monsoon climate regime with two distinct seasons, the rainy season lasts from April to October, the dry season—from November to March. The average annual temperature is 22.0 °C. The total average annual rainfall is 2339 mm; precipitation falls mainly from May to October. Ta Dung National Park is characterized by subtropical humid evergreen closed forests and mixed broadleaf and coniferous closed forests [44]. The mycobiota of the National Park previously was not studied.
Specimens were photographed in the field, and their macromorphological characters, such as size, colour, shape, and surface of all parts of the basidiomata as well as odour, were documented before drying. Colour codes refer to Kornerup & Wanscher [45]. GPS coordinates of collection site, habitat, and substrate type were also documented for each collection. Specimens were then dried either in airtight plastic containers with silica gel, or with an electric dryer at a temperature ca. 50 °C, placed on a piece of absorbent paper and packed in plastic Ziploc bags with small amounts of silica gel.
2.2. Morphological Study
Microscopic measurements and drawings were made with an AxioScope A1 light microscope equipped with Zeiss AxioCam 1Cc3 digital camera with AxioVisionRel.4.6 software (CarlZeiss, Jena, Saxe-Weimar-Eisenach, Germany). Spores, basidia, and cystidia were observed in squash preparations of small parts of the lamellae in 5% KOH or 1% Congo Red in concentrated NH4OH. The pileipellis was examined from a radial section of the pileus in 5% KOH. Basidiospore dimensions were based on 20 spores; cystidia and basidia dimensions on at least 10 structures per collection. Basidia were measured without sterigmata, and the spores without apiculus. Spore length to width ratios were reported as Q. When studying the specimens and compiling morphological descriptions, we followed the recommendations and terminology of [8]. The dried specimens were deposited in the Mycological Herbarium of the Komarov Botanical Institute RAS (LE) and in the Herbarium of the Joint Vietnam–Russia Tropical Science and Technology Research Centre, Hanoi (VRTC).
2.3. DNA Extraction, Amplification, and Sequencing
PCR products were obtained without DNA purification step using the Thermo Scientific Phire Tissue Direct PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA) standard protocol. The ribosomal ITS1–5.8S–ITS2 region was amplified with the fungal specific primers ITS1F and ITS4B [46]; http://www.biology.duke.edu/fungi/mycolab/primers.htm, accessed on 1 March 2017. Sequences of nrLSU-rDNA were generated using primers LR0R and LR5 [47]. Primers EF1-983F and EF1-1567R were used to amplify approximately 500 bp of tef1 [48]. For ITS, PCR was carried out under the following cycling parameters: initial denaturation: 98 °C for 4 min; followed by 35 cycles: 98 °C for 1 min, 52 °C for 1 min, and 72 °C for 1 min, and final extension at 72 °C for 3 min. For nrLSU: initial denaturing at 98 °C for 5 min; then 12 cycles of denaturing at 98 °C for 5 s, annealing at 67 °C for 1 min, extension at 72 °C for 1.5 min; then 35 cycles of denaturing at 98 °C for 5 s, annealing at 56 °C for 1 min, extension at 72 °C for 1.5 min; and a final extension step of 72 °C for 10 min. For tef1-a: initial denaturing at 98 °C for 5 min; then 8 cycles of denaturing at 98 °C for 5 s, annealing at 60 °C for 40 s, extension at 72 °C for 2 min; then 36 cycles of denaturing at 98 °C for 5 s, annealing at 53 °C for 1.5 min, extension at 72 °C for 2 min; and a final extension step of 72 °C for 10 min.
PCR products were visualized using agarose gel electrophoresis and Gel Red staining, and subsequently purified with the Fermentas Genomic DNA Purification Kit (Thermo Fisher Scientific Inc., Waltham, MA, USA). Sequencing was performed with an ABI model 3500 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA).
This work was carried out using equipment of the Core Facility Centre ‘Cell and Molecular Technologies in Plant Science’ of the Komarov Botanical Institute. Raw data were edited and assembled in MEGA X [49]. Newly generated sequences have been deposited in the GenBank.
2.4. Alignment and Phylogenetic Analyses
For this study, 12 nrITS and 9 tef1α, and 10 nrLSU sequences were newly generated. In addition, 38 nrITS and 20 tef1α, and 43 nrLSU sequences, including outgroups, were retrieved from the GenBank database (www.ncbi.nlm.nih.gov/genbank, accessed on 15 March 2023), using the BLASTn application (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 15 March 2023). The information on all these sequences is presented the Table 1.
Three datasets were analysed: nrITS, tef1α, and nrLSU. DNA sequences were aligned with the MAFFT v.7.110 web tool [56] using the G-INS-i option, and then manually modified where necessary in MEGA X [49]. To determine the phylogenetic positions of the studied collections, both datasets were analysed using Bayesian Analysis (BA). BA was performed using MrBayes 3.2.1 [57], under a GTR model. The analyses were run with two parallel searches: four chains for 5 million generations for ITS and LSU and for 1 million generations for tef1α, starting with a random tree. The trees were sampled every 100 generations. To check for convergence of MCMC analyses and to obtain estimates of the posterior distribution of parameter values, Tracer v1.7.2 was used [58]. The phylogenetic trees were edited in Adobe Illustrator CS4. Posterior probability (PP) values ≥ 0.95 are considered significant.
3. Results
3.1. Phylogenetic Analysis
The full nrITS dataset contained 50 sequences with 1059 characters (gaps included). The/Entocybe clade was selected as outgroup due to its basal position in the Entoloma phylogeny [6]. Besides our specimens, the tree includes 7 more sequences of the subgenus Cubospora retrieved from the GenBank NCBI data base, 16 representatives of the subgenus Leptonia, and 1–3 representatives of the other main subdivisions of the Entoloma s.l.
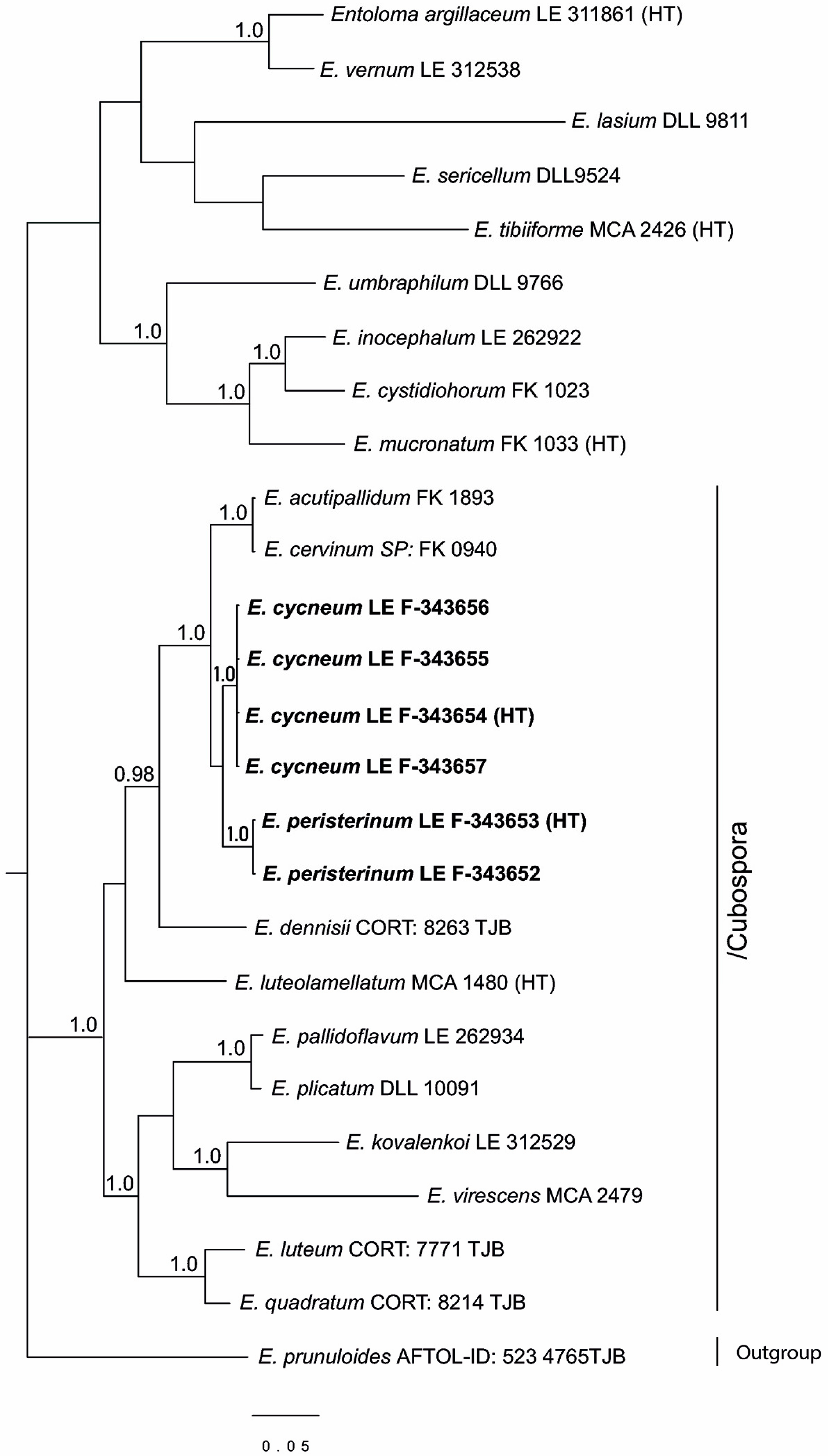
Since in the GenBank the ITS data are absent for many species of the subgenus Cubospora, a tree was constructed for this subgenus based on the tef1α as well. For the subgenus Leptonia, such information is insufficient for analysis. The full tef1α dataset contained 26 sequences with 535 characters (gaps included). It included Entoloma prunuloides as an outgroup due to its basal position in the Entoloma phylogeny [53,54], and E. luteolamellatum, the type-species of the subgenus Cubospora. In addition to our specimens, 11 more representatives of this subgenus and 1–3 representatives of the other main subgenera of the Entoloma s.l. were included in the analysis.
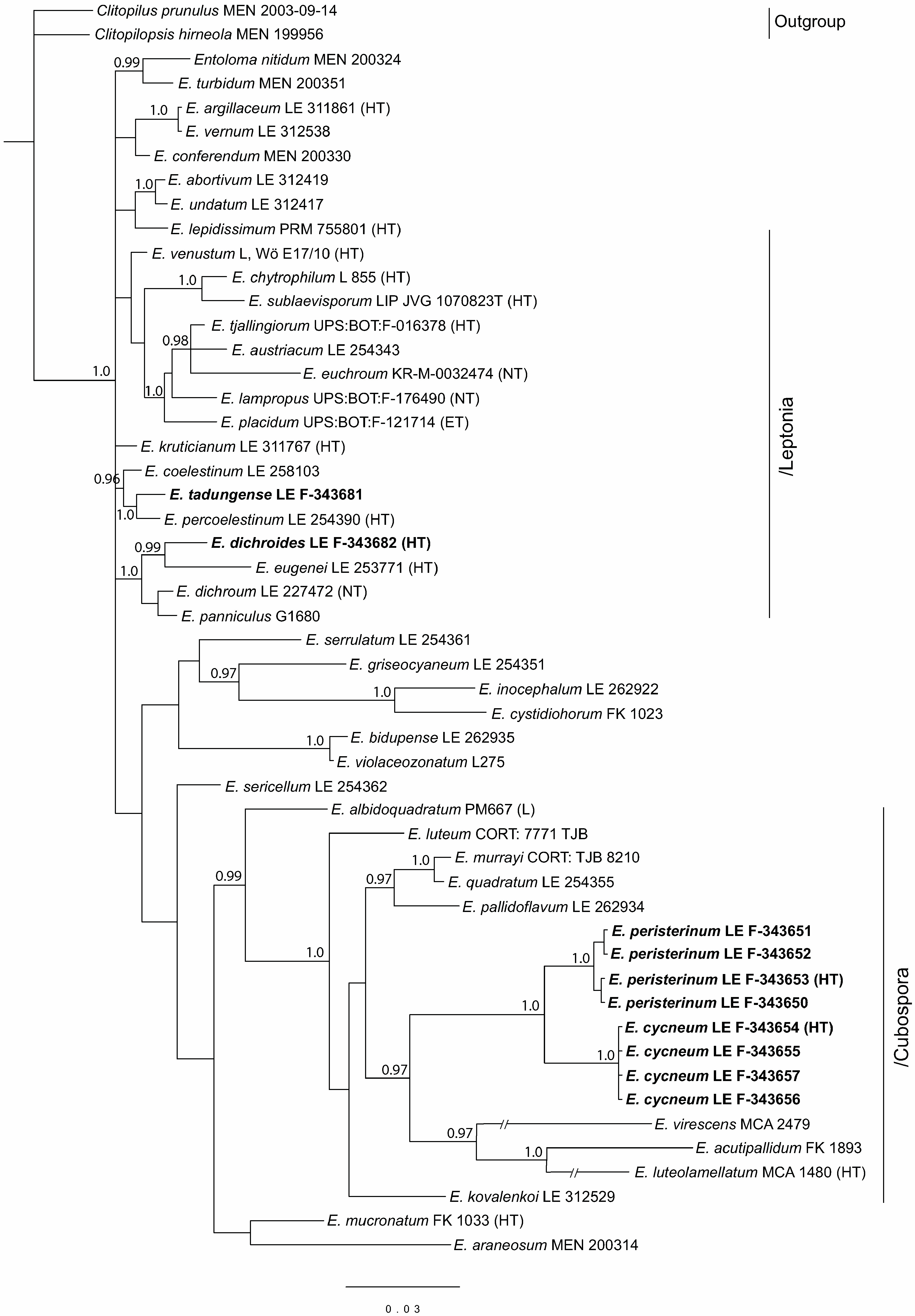
The full nrLSU dataset contained 52 sequences with 781 characters (gaps included). Clitopilus prunulus and Clitopilopsis hirneola were chosen as an outgroup because of their basal position in the Entolomataceae phylogeny [5]. In addition to new species specimens, 8 representatives of the subgenus Cubospora and 15 sequences of the subgenus Leptonia, as well as 1–3 sequences of the other main subgenera of the Entoloma s.l., were added to the analysis.
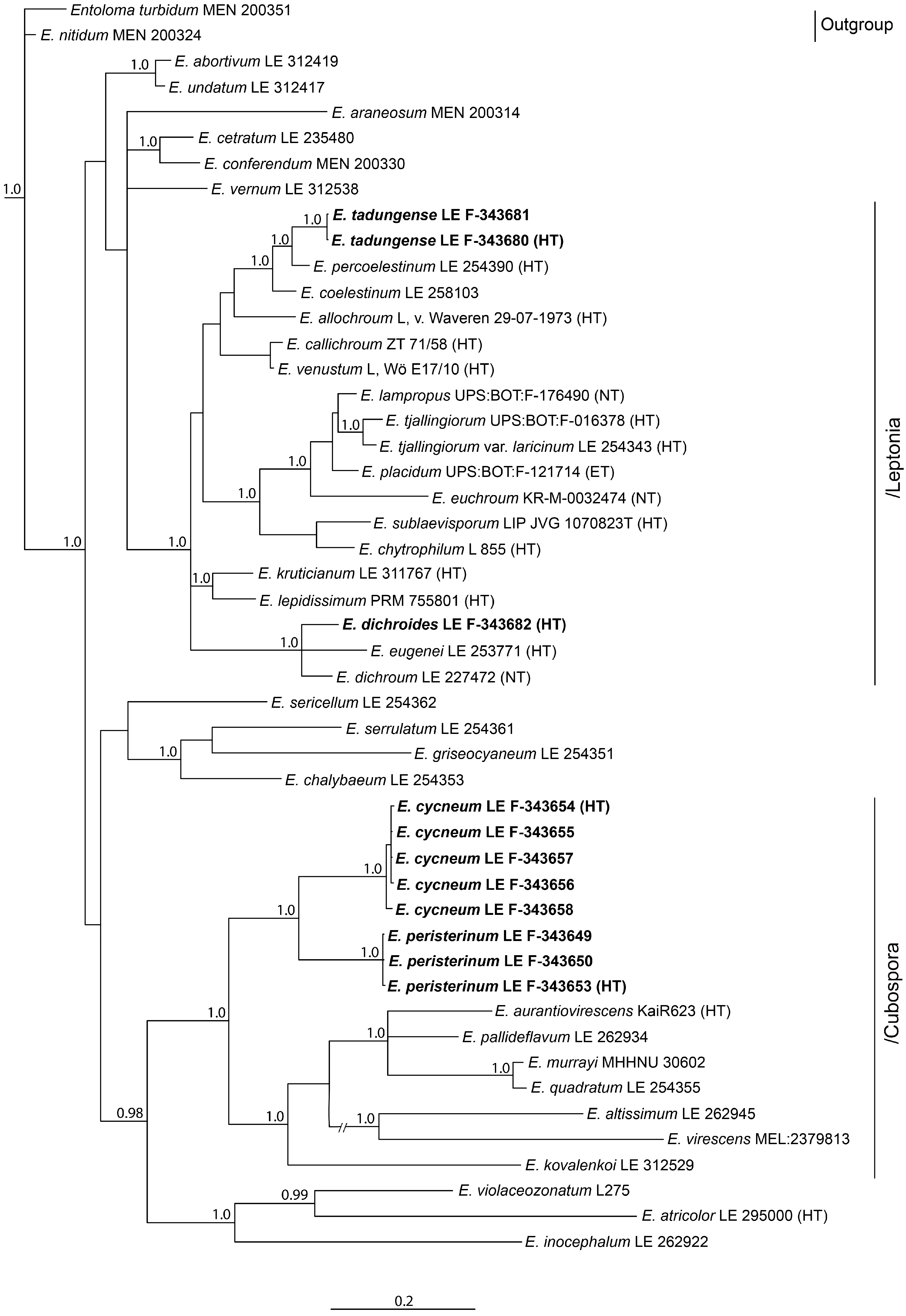
The results of the phylogenetic analysis are presented in the Figure 1 (nrITS), Figure 2 (tef1α), and Figure 3 (nrLSU). Entoloma cycneum and E. peristerinum form highly supported branches within the/Cubospora clade in all trees. E. tadungense and E. dichroides clearly nest within the/Leptonia clade in the ITS tree. They are not represented on the tef1α tree. Generally, in the LSU tree, the topology mostly is not or hardly resolved due to small differences between the sequences in the subgenera. At the same time, the subgenus Cubospora forms a highly supported clade in which two new species (E. cycneum and E. peristerinum) nest.
Subgenus Leptonia does not form a separate clade in the LSU tree, but it is represented by several small clades and singletons. However, it is clearly seen here that Entoloma tadungense clusters together with E. percoelestinum and E. coelestinum with high support. Entoloma dichroides distinctly nests in the/dichroum clade, but it differs from the known Leptonia species.
3.2. Taxonomy
Entoloma subgenus Cubospora Karstedt, Capelari, Largent, T. J. Baroni & Bergemann, in Phytotaxa 391(1): 20 (2019).
Entoloma cycneum O.V. Morozova & T.H.G. Pham, sp. nov. (Figure 4).
MycoBank: MB848527.
Etymology. From “κύκνος” (Greek)—swan, due to colour similarity with white swan.
Holotype. Vietnam, Gia Lai Province, K’Bang District, Son Lang Commune, Kon Chu Rang Nature Reserve, N 14.49436°, E 108.54428°, 1030 m a. s. l., on soil in middle-mountain evergreen mixed forest with a predominance of Podocarpaceae (Dacrydium elatum, Dacrycarpus imbricatus), Magnoliaceae, Burseraceae (Canarium), Myrtaceae (Syzygium), 30 May 2016, O.V. Morozova (LE F-343654). Isotype in VRTC (299VN16).
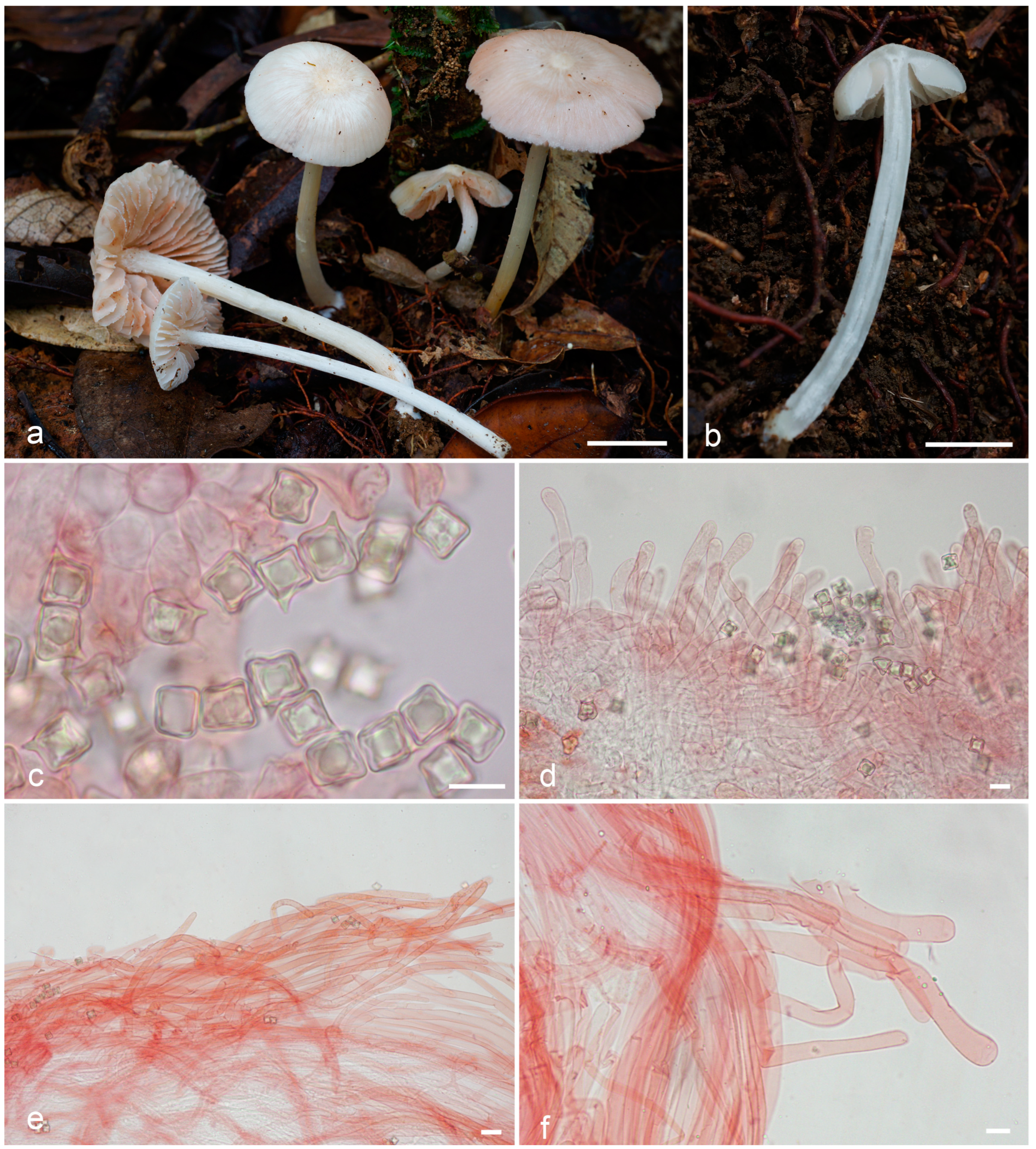
Diagnosis. Entoloma cycneum is characterized by initially white or whitish basidiomata with mainly smooth glabrous with thin pubescence near the margin, hygrophanous pileus, longitudinally fibrillose or fibrillose-scaly stipe, cuboid spores, and long, more or less cylindrical cheilocystidia of serrulatum-type.
Basidiomata small to medium-sized. Pileus 10–25 mm diam., firstly hemispherical, then convex, plano-convex with small papilla, with a slightly involute then straight crenulate margin, hygrophanous, smooth, mostly glabrous, but covered with thin fibrils on the pileus margin, in wet conditions translucently striate almost up to the centre, white to cream (3A1–2; 4A1–2), yellowish in places of damage, lighter towards margin, slightly darker in radial hygrophanous stripes, then dries up to white. Lamellae moderately distant, adnexed, adnate-emarginate, ventricose, initially white, then cream, pale pink, with white denticulate edge. Stipe 30–60 × 1.5–3 mm, cylindrical, fistulous, white, longitudinally fibrillose or fibrillose-scaly, at least at the top, white tomentose at base. Context white or hyaline. Smell indistinct, taste not reported.
Basidiospores (7.2–)8.5–9.0(–9.7) × (7.2–)8.0–8.5(–8.7) μm, Q = 1.0–1.2, isodiametrical or subisodiametrical, cuboid, rarely with 5 angles in side-view. Basidia 37–50 × 11–12.5 μm, 4-spored, clavate, constricted in the middle part, clamped. Cheilocystidia 95–160 × 7.5–9 μm, of serrulatum-type, flexuous-cylindrical, sometimes capitate or narrowly clavate, septate, not pigmented, with granular content, forming sterile lamellae edge. Hymenophoral trama regular, made up of cylindrical or inflated hyphae 4–10 μm wide. Pileipellis a cutis of cylindrical hyphae up to 10 μm broad, some hyphae are rising. Pileitrama consists of cylindrical hyphae 4–6 μm broad, intertwined by oleiferous hyphae with brilliant content. Stipitipellis of cylindrical hyphae up to 7 μm broad. Caulocystidia in bundles of rising hairs, cylindrical or slightly broadened towards apex, up to 250 μm length and 12 μm broad. Clamp-connections present in all tissue. Brilliant granules abundant.
Habitat and distribution—In small groups on soil in middle-mountain evergreen mixed forest. Known from Vietnam.
Additional specimens examined. Vietnam, Gia Lai Province, K’Bang District, Son Lang Commune, Kon Chu Rang Nature Reserve, N 14.49439°, E 108.54591°, 990 m a. s. l., on soil in middle-mountain evergreen mixed forest with a predominance of Podocarpaceae (Dacrydium elatum, Dacrycarpus imbricatus), Magnoliaceae, Burseraceae (Canarium), Myrtaceae (Syzygium), 27 May 2016, O.V. Morozova (LE F-343656 (1687_243VN16); ibid., 28 May 2016, O.V. Morozova (LE F-343655 (1689_255VN16); LE F-343657 (1690_256VN16); Dak Nong Province, Dak Plao District, south-east macroslope of Mt. Ta Dung, trail along the stream valley in the saddle between M’neun Tchirke and the eastern spur of Ta Dung, N 11.87172°, E 108.08253°, 1310 m a. s. l., on soil in middle-mountain evergreen mixed forest with a participation of Fagaceae and Lauraceae (Litsea glutinosa), 3 June 2022, T.H.G. Pham (LE F-343658).
Entoloma peristerinum O.V. Morozova & T.H.G. Pham, sp. nov. (Figure 5).
MycoBank: MB848528.
Etymology. From “περιστερά” (Greek)—dove, due to colour similarity to white dove.
Holotype. Vietnam, Gia Lai Province, K’Bang District, Son Lang Commune, Kon Chu Rang Nature Reserve, N 14.49667°, E 108.56106°, 980 m a. s. l., on soil in middle-mountain evergreen mixed forest with a predominance of Podocarpaceae (Dacrydium elatum, Dacrycarpus imbricatus), Magnoliaceae, Burseraceae (Canarium), Myrtaceae (Syzygium), 28 May 2016, O.V. Morozova (LE F-343653). Isotype in VRTC (276VN16).
Diagnosis. Entoloma peristerinum is characterized by initially cream, pale beige to beige or greyish-beige becoming white basidiomata, with smooth, glabrous, hygrophanous pileus, longitudinally fibrillose or fibrillose-scaly stipe, cuboid spores and cheilocystidia mostly narrowly clavate or tapering towards the apex—of porphyrogriseum-type.
Basidiomata small to medium-sized. Pileus 10–30 mm diam., firstly conical, then broadly conical with distinct acute papilla, with deflexed then straight or undulating, slightly crenulate margin, hygrophanous, smooth, glabrous, in wet condition translucently striate almost up to the centre, cream, pale beige to beige or greyish-beige (4A2; 4B2–3; 5B2–3), lighter towards margin, with dark radial hygrophanous stripes and yellowish papilla, then dries up by white radial strokes, finally becomes completely white, silky. Lamellae moderately distant, adnate-emarginate, ventricose, cream, pale pink, with minutely serrate whitish edge. Stipe 30–70 × 1.5–3 mm, cylindrical, fistulous, white, longitudinally fibrillose or fibrillose-scaly, at least at the top, white tomentose at base. Context white. Smell indistinct, taste not reported.
Basidiospores (7.4–)8–9.5(–9.8) × (6.5–)7–8(–9.2) μm, Q = 1.0–1.2(–1.3), isodiametrical or subisidiametrical, cuboid, rarely with 5 angles in side-view. Basidia 47–60 × 11–14.5 μm, 4-spored, clavate, constricted in the middle part, clamped. Cheilocystidia 75–215 × 12–15 μm, of porphyrogriseum-type [8], cylindrical, narrowly clavate or tapering towards the apex, with granulose content, usually forming sterile lamellae edge; in young basidiomata lamellae edge can be heterogeneous with rare cheilocystidia. Hymenophoral trama regular, made up of cylindrical or inflated hyphae 3–8 μm wide. Pileipellis a cutis of cylindrical hyphae up to 10 μm broad. Pileitrama consists of cylindrical hyphae 4–6 μm broad, intertwined by oleiferous hyphae with brilliant content. Clamp-connections present in all tissue. Brilliant granules abundant.
Habitat and distribution—In small groups on soil in middle-mountain evergreen mixed forest. Known from Vietnam.
Additional specimens examined. Vietnam, Gia Lai Province, K’Bang District, Son Lang Commune, Kon Chu Rang Nature Reserve, N 14.49439°, E 108.54591°, 990 m a. s. l., on soil in middle-mountain evergreen mixed forest with a predominance of Podocarpaceae (Dacrydium elatum, Dacrycarpus imbricatus), Magnoliaceae, Burseraceae (Canarium), Myrtaceae (Syzygium), 28 May 2016, O.V. Morozova (LE F-343651 (1688_254VN16)); ibid., N 14.49667°, E 108.56106°, 980 m a. s. l., 28 May 2016, O.V. Morozova (LE F-343650 (1691_276bVN16)); ibid., N 14.49436°, E 108.54428°, 1030 m a. s. l., 30 May 2016, O.V. Morozova, LE F-343652 (1692_300VN16), LE F-343649 (1696_312VN16)).
Notes—Entoloma cycneum and E. peristerinum are morphologically very similar species characterized by white or whitish with yellowish or beige tinged basidiomata with mainly smooth, glabrous, and hygrophanous pileus, longitudinally fibrillose or fibrillose-scaly white stipe, cuboid spores and more or less cylindrical cheilocystidia, arising from hymenophoral trama. Entoloma peristerinum posseses initially more coloured beige conical pileus, discolouring to white with age and drying. The pileus of E. cycneum is initially white, hemisphaerical to convex, usually with thin pubescence near the margin. E. peristerinum differs from E. cycneum also by more differentiated porphyrogriseum-type [8] cheilocystidia, vs. serrulatum-type in E. cycneum. Yellow tinge can present in old or damaged basidiomata of both species.
Phylogenetically both new species are also close to each other but rather distant from other known species in the subgenus Cubospora [7]. Entoloma cervinum (Karstedt & Capelari) Blanco-Dios and E. acutipallidum E. Horak & Cheype from the South America are the closest species according to the phylogenetical analysis. Morphologically they differ from our species by darker, distinctly coloured pileus [28].
Several species with cuboid spores and predominantly white or whitish basidiomata have been described at different times from different regions of the Earth.
The pileus of Entoloma alboumbonatum Hesler from North America is darker coloured, and only umbo is white. This species is characterized by the clavate or capitate cheilocystidia, presence of the pleurocystidia, and absence of the clamp connections [31].
E. albidoquadratum Manim. & Noordel., described from India, is characterized by non hygrophaneous, non-translucently striate pileus, presence of pleurocystidia, and large spores [20]. E. minutoalbum E. Horak is a species of Southern Hemisphere, a common fungus of the subantarctic Nothofagus forests of Tierra del Fuego and New Zealand. Morphologically it differs from new species by small basidiomata (with the pileus less than 10 mm diam.), smaller spores, and absence of the cheilocystidia [13]. E. albogracile E. Horak is also species from the Southern Hemisphere—Papua New Guinea. It is characterized by the small size, pileus covered with minute scales and fibrils, and by the absence of clamp-connections [13]. E. laccaroides T.H. Li, E. Horak & Xiao Lan He is recognized by the umbilicate pileus, and numerous conspicuous broadly fusoid to utriform pleurocystidia [12].
Due to the discolouration new species would be compared with cuboid-spored light-yellow species. Entoloma dennisii from Trinidad is a rather robust species with pileus up to 5 cm broad, deep to pale yellow colour, and small spores. E. pallidoflavum differs by the predominance of light-yellow colour in the pileus, yellow content of the oleiferous hyphae, and smaller spores [13].
Entoloma subgen. Leptonia (Fr.) Noordel., in Persoonia 11: 146 (1981), emend. O. V. Morozova, Noordel. & Vila (2014); emend. Noordeloos et al. (2022).
Entoloma tadungense O.V. Morozova & T.H.G. Pham, sp. nov. (Figure 6).
MycoBank: MB848530.
Etymology. Named after the type-locality—Ta Dung National Park.
Holotype. Dak Nong Province, Dak Glong District, Ta Dung National Park, south-eastern macroslope of the ridge of the Ta Dung Mt, south-eastern slope of the Ta Dung Mt, TK 1805, N 11.86780°, E 108.11692°, 1240 m a.s.l., on soil in evergreen broad-leaved forest with Lithocarpus spp., Quercus sp., Schima sp., Acer flabellatum, Dilleniaceae, Myristicaceae, 11 October 2022, T.H.G. Pham, O.V. Morozova (LE F-343680). Isotype in VRTC (87VN22).
Diagnosis. Entoloma tadungense is distinguished among the other Leptonia species by the tiny dark blue basidiomata with discolouring to lilac of the stipe, and by the small spores with rather pronounced angles.
Basidiomata small-sized, mycenoid. Pileus 3–10 mm broad, conical, or hemispherical with umbo, becoming almost applanate, not hygrophanous, not translucently striate, with deflexed then straight margin, radially fibrillose, squamulose at centre, firstly uniformly dark blue, blackish blue (19F7–8; 20E6–8; 21F7–8), glabrescent, radially cracking showing the white background, and pallescent (up to 20C5–6; D5–6) with edge. Lamellae moderately distant, adnate emarginate, ventricose, white, becoming pinkish, with entire concolourous edge. Stipe 1.5–30 × 0.5–2 mm, cylindrical or broadened towards to base, fistulous, initially distinctly longitudinally fibrillose, glabrescent with age up to almost smooth, especially in the apex, firstly concolourous with pileus—dark blue, blackish blue (19F7–8; 20E6–8), then discoloured up to lilac, whitely tomentose at base. Context thin, concolourous with the surface. Smell indistinct, taste not reported.
Basidiospores (6.0–)6.8–6.9(–8.3) × (4.2–)4.8–4.9(–5.7) μm, Q = 1.3–1.4(–1.6), heterodiametrical, with 6–7 relatively sharp angles in side-view. Basidia 19.5–24 × 6.6–8.3 μm, 4-spored, narrowly clavate to subcylindrical, clamped. Lamellae edge fertile or heterogeneous. Cheilocystidia 24–67 × 9–17 μm, of poliopus-type [8], clavate or lageniform, more frequent near the pileus margin. Hymenophoral trama regular, made up of narrow, cylindrical hyphae up to 3 μm wide. Pileipellis a cutis with a transition to a plagiotrichoderm and a trichoderm of cylindrical to slightly inflated hyphae 10–20 μm wide with blue intracellular pigment. Stipitipellis a cutis with a transition to a plagiotrichoderm with rising hyphae forming hairs up to 5 μm wide. Clamp-connections present, but rare. Brilliant granules absent.
Habitat and distribution—In small groups on soil in middle-mountain evergreen mixed forest. Known from Vietnam.
Additional specimens examined. Dak Nong Province, Dak Glong District, Ta Dung National Park, south-eastern macroslope of the ridge of the Ta Dung Mt, south-eastern slope of the Ta Dung Mt, TK 1805, N11.86780, E 108.11692, 1240 m a.s.l., on soil in evergreen broad-leaved forest with Lithocarpus spp., Quercus sp., Schima sp., Dilleniaceae, Myristicaceae, Acer flabellatum, 11 Oct. 2022, T.H.G. Pham, O.V. Morozova, LE F-343681, duplicate in VRTC (88VN22); ibid., 15 October 2022, LE F-343683, duplicate in VRTC (138VN22), LE F-343684, duplicate in VRTC (139VN22).
Notes—Entoloma tadungense is a species of the subgenus Leptonia due to presence of clamp connections, absence of brilliant granules, longitudinally fibrillose stipe surface and plagiotrichoderm to trichoderm pileipellis. It resembles Entoloma percoelestinum O.V. Morozova, Noordel., Vila & Bulyonk. by the small-sized dark blue mycenoid basidiomata [39]. Microscopically the absence of cheilocystidia and small spores also make them similar. However, E. tadungense can be recognized by smaller spores with pronounced angles, as well as the lilac discolouration of the stipe. Molecular data support their differences (p-distance from the closest species E. percoelestinum (ITS1-5.8S-ITS2 region)—4.9%). The similar non tropical species with dark blue colour and small size of the basidiomata distinguish from the new species in the following: E. coelestinum (Fr.) Hesler—by the smooth stipe and slightly larger spores, E. lepidissimum (Svrček) Noordel. and E. venustum—by the coloured lamellae, distinctly larger spores, and presence of the cheilocystidia, E. chytrophilum possesses large nodulose spores [39]. The American species E. subcoelestinum (Largent) Blanco-Dios is characterized by the initially coloured lamellae, moniliform cells in the pileipellis, and larger indistinctly angular spores [32].
Entoloma dichroides O.V. Morozova & T.H.G. Pham, sp. nov. (Figure 7).
MycoBank MB 848531.
Etymology. Named after its similarity to Entoloma dichroum.
Holotype. Dak Nong Province, Dak Glong District, Ta Dung National Park, northwest of the Ta Dung Mt, TK 1781, N11.923056°, E 108.00194°, 1000 m a.s.l., on soil in evergreen broad-leaved forest with Parashorea chinensis, Rhodoleia championii, Fagaceae, Lauraceae, Hypericaceae, 1 June 2022, Pham T.H.G., LE F-343682. Isotype in VRTC (HG09).
Diagnosis. Entoloma dichroides is a species of subgenus Leptonia, characterized by the dark blue basidiomata with squamulose surface of the pileus and stipe, initially white lamellae, absence of the cheilocystidia, and spores with 5(–6) pronounced angles and elongated apiculus.
Basidiomata medium-sized, mycenoid. Pileus 40 mm diam, conical to conico-convex, with deflexed then straight margin, not hygrophanous, not translucently striate, dark blue (20F6–8), entirely fibrillose-squamulose with prominent dense squamules on whitish background. Lamellae narrowly adnexed to almost free, ventricose, moderately distant, initially white becoming yellowish-pink, brownish-pink when, with an entire concolourous edge. Stipe 60 × 3–4 mm, cylindrical, slightly broadened at base, hollow, dark blue, concolourous with the pileus, entirely fibrillose-squamulose with dark blue fibrils (20F6–8) on whitish background. Context white. Smell weak, taste not reported. Basidiospores 9.5–12.5 × 6.5–8.5 μm, on average 10.8 × 7.2 μm, Q = 1.4–1.6(1.7), Qav = 1.5, 5(–6)-angled, heterodiametric with pronounced angles and elongated apiculus. Basidia 31–41 × 10–13 μm, 4-spored, clamped. Lamella edge fertile. Cheilocystidia absent. Hymenophoral trama regular, made up of narrow cylindrical hyphae 2–6 μm wide. Pileipellis a trichoderm of cylindrical entangled 6–10 m wide hyphae with fusiform or irregularly shaped terminal elements, 90–200 × 13–19 μm, with dark blue intracellular pigment. Stipitipellis an entangled trichoderm of cylindrical hyphae, 6–12 m wide with cylindrical to lageniform, terminal elements, forming squamules, 4–12 μm wide and up to the 200 long, with dark blue intracellular pigment. Clamp-connections abundant. Brilliant granules absent.
Habitat and distribution—Solitary in middle-mountain evergreen mixed forest. Known from Vietnam.
Notes—Entoloma dichroides is similar to European E. dichroum and Australian E. panniculus due to dark blue basidiomata with squamulose surface of the pileus and stipe, initially white lamellae, and spores with pronounced angles [26]. New species differs from them by darker basidiomata with conical pileus, basidiospores with elongated apiculus, and absence of the cheilocystidia. The Eastern species E. eugenei Noordel. & O.V. Morozova is also close. It is recognized by the more robust basidiomata and presence of the cheilocystidia [21]. The p-distance (ITS1-5.8S-ITS2 region) of the new species from E. dichroum—6.9%, from E. eugenei—10.4%.
4. Discussion
The genus Entoloma is the second largest genus in the order Agaricales with over 1000 species worldwide [59]. However, studies devoted to it in Vietnam have not yet been carried out enough.
The first most important contribution to the study of Vietnamese mycobiota was made by N. Patouillard, who described many new species from Northern and Central Vietnam based on collections by V. Demange, L. Duport, P. A. Eberhardt, E. Poilane, and others. First records of the genus Entoloma (Rhodophyllus clypeatus, Rh. sericeus) were also published by him [60,61], including originally described from Vietnam Rh. submurinus [61]. Heim and Malençon [62] published information on E. madidum. The first checklist summarizing all the data on the species composition of fungi and slime-molds in Vietnam was published in 1998 and included 829 species, of which only 6 belonged to the genus Entoloma [63]. The same species are presented in the list of plant species of Vietnam [64]. In 2003, Le Ba Dung listed 300 species of macromycetes for the Central Highlands, with only two Entoloma species: E. lividum and E. madidum [65]. Later, Ngô Anh and Nguyễn Thị Kim Cúc recorded two more species of the genus in Thua Thien Hue Province—E. prunuloides in Bach Ma National Park [66] and E. abortivum in Phong Dien Nature Reserve [67]. The data on Vietnamese mycobiota have been summarized in a monography published in 2011–2012, which provides information on the ecology and distribution of more than 900 species of macromycetes [68,69] with only 3 Entoloma species.
More intensive studies of the genus Entoloma in Vietnam began in the frame of the work of the Joint Vietnam–Russia Tropical Science and Technology Research Centre in collaboration with the Komarov Botanical Institute RAS. In 2012, Morozova et al. [11] reported 12 species of the genus Entoloma (Agaricales, Basidiomycota) for the Bidoup—Nui Ba (Lam Dong Province) and Cat Tien (Dong Nai Province) National Parks, of which 11 species were recorded for Vietnam for the first time, and a new species for science, E. myriadophyllum O.V. Morozova, was described.
The above information was based only on morphological data. In connection with the revision of the system of the genus Entoloma [8], these data are not entirely credible and require confirmation. In cases where herbarium material is absent or old and destroyed, this is not possible. However, sequences were later obtained for some of these records. Some of the identifications were revised, and some of these findings were described as species new to science—E. daphnis and E. bidupense [4,42]. Our specimens with cuboid spores were used in the work of Karstedt and colleagues [7], where additional data on tef1α, mtSSU, rpb2, and LSU markers were obtained for them.
Further descriptions of new species have already been confirmed by molecular data. Thus, Entoloma flavovelutinum and E. porphyroleucum from the Bu Gia Map National Park (Binh Phuoc Province) [2,70], E. nigrovelutinum from Chu Yang Sin National Park (Dak Lak Province) [71], and E. atricolor [72], E. arion, E. argus, E. daphnis, E. icarus [4], E. kovalenkoi [3], from the Kon Chu Rang Nature Reserve were described.
As a result, the data on 29 species of Entoloma have been published for Vietnam so far. Only for 14 of them the molecular data have been published and submitted into the GenBank. The resulting list is presented here.
List of the species of the genus Entoloma mentioned in the publications for Vietnam, 1910–2022 (species, supported for the molecular data are marked by the asterisk (*)):
Entoloma abortivum (Berk. & M.A. Curtis) Donk [67]
*E. altissimum (Massee) E. Horak [11]
*E. argus O.V. Morozova, E.S. Popov, A.V. Alexandrova & Noordel. [4]
*E. arion O.V. Morozova, E.S. Popov, T.H.G. Pham & Noordel. [4]
*E. daphnis O.V. Morozova, E.S. Popov, T.H.G. Pham & Noordel. [4,11] (as E. nubilum Manim., Leelav. et Noordel.)
*E. flavovelutinum O.V. Morozova, E.S. Popov, A.V. Alexandrova & Xiao L. He [70]
*E. icarus O.V. Morozova, E.S. Popov & Noordel. [4]
*E. kovalenkoi O.V. Morozova, E.S. Popov & A.V. Alexandrova [3]
E. longistriatum (Peck) Noordel. [11]
*E. myriadophyllum O.V. Morozova [11]
*E. nigrovelutinum O.V. Morozova & A.V. Alexandrova [71]
*E. porphyroleucum O.V. Morozova, Noordel. & Dima [2]
*E. quadratum (Berk. et M. A. Curtis) E. Horak [11]
E. sulcatum (T.J. Baroni et Lodge) Noordel. et Co-David [11]
Four more species are described in the presented article. For E. pallidoflavum, the information on ITS sequence is published here.
In conclusion, it is important to note that the area of tropical forests has been drastically reduced in recent years, including in Vietnam. Along with them, the species confined to them disappear, often without even being assigned a name. Nature reserves and national parks serve to save biologically valuable forest areas. However, even here the species are vulnerable. It sometimes happens that type localities are destroyed during road reconstructions (e.g., E. atricolor, E. arion). It remains to be hoped that they will continue to be found in the adjacent forests. In central Vietnam, the greatest diversity of fungi of the genus Entoloma was observed in middle-mountain evergreen broad-leaved forests. They require special attention, study, and careful treatment.
The study of the fungal diversity of typical tropical forests complements the information on the biota of macromycetes in Vietnam and may be useful in the development of measures for the conservation of these valuable nature areas and the species inhabiting them.
Author Contributions
Conceptualization, collecting, formal analysis, investigation, writing—original draft preparation—O.M. and T.H.G.P. O.M. and T.H.G.P. contributed equally and should be considered as co-first authors. All authors have read and agreed to the published version of the manuscript.
Funding
The field work was funded by Joint Vietnam-Russia Tropical Science and Technology Research Centre (project E1.5 No3). The work with collections was provided in the framework of an institutional research project of the Komarov Botanical Institute RAS (122011900032-7).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The DNA sequence data obtained from this study have been deposited in GenBank NCBI (https://www.ncbi.nlm.nih.gov/genbank/, accessed on 13 April 2023).
Acknowledgments
The authors are grateful to the management and staff of the Kon Chu Rang Nature Reserve and Ta Dung National Park for their great help in the field work and for permission to collect in the forests. Many thanks to E.S. Popov (Komarov Botanical Institute RAS) for the help in the field work and with the literature.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pham, T.H.G.; Popov, E.S.; Morozova, O.V.; Nguyen, D.H. Basidiomycetous Macromycetes of the Tay Nguyen Plateau in Vietnam: The History of Investigation. Mikol. Fitopatol. 2018, 52, 306–318. [Google Scholar]
- Haelewaters, D.; Dima, B.; Abdel-Hafiz, B.I.I.; Abdel-Wahab, M.A.; Abul-Ezz, S.R.; Acar, I.; Aguirre-Acosta, E.; Aime, M.C.; Aldemir, S.; Ali, M.; et al. Fungal Systematics and Evolution 6. Sydowia 2020, 72, 231–356. [Google Scholar]
- Crous, P.W.; Osieck, E.R.; Jurjevi, Ž.; Boers, J.; Van Iperen, A.L.; Starink-Willemse, M.; Dima, B.; Balashov, S.; Bulgakov, T.S.; Johnston, P.R.; et al. Fungal Planet description sheets: 1284–1382. Persoonia 2021, 47, 178–374. [Google Scholar] [CrossRef]
- Morozova, O.; Popov, E.; Alexandrova, A.; Pham, T.H.G.; Noordeloos, M.E. Four new species of Entoloma (Entolomataceae, Agaricomycetes) subgenera Cyanula and Claudopus from Vietnam and their phylogenetic position. Phytotaxa 2022, 549, 1–21. [Google Scholar] [CrossRef]
- Co-David, D.; Langeveld, D.; Noordeloos, M.E. Molecular phylogeny and spore evolution of Entolomataceae. Persoonia 2009, 23, 147–176. [Google Scholar] [CrossRef]
- Baroni, T.J.; Matheny, P.B. A re-evaluation of gasteroid and cyphelloid species of Entolomataceae from eastern North America. Harv. Pap. Bot. 2011, 16, 293–310. [Google Scholar] [CrossRef]
- Karstedt, F.; Baroni, T.J.; Largent, D.L.; Capelari, M.; Bergemann, S.E. Phylogenetic and morphological analyses of species of the Entolomataceae (Agaricales, Basidiomycota) with cuboid basidiospores. Phytotaxa 2019, 391, 1–27. [Google Scholar] [CrossRef]
- Noordeloos, M.E.; Morozova, O.; Dima, B.; Reschke, K.; Jansen, G.M.; Brandrud, T.E.; Jordal, J.B.; Bendiksen, E.; Vila, J. Entoloma Sensu Lato. Subgenera Cyanula, Leptonia, Nolanea, Trichopilus, and the/Rhombisporum Clade. Fungi Europaei; Edizione Candusso: Alassio, Italy, 2022; Volume 5b, pp. 1–968. [Google Scholar]
- Noordeloos, M.E.; Jansen, G.M.; Jordal, J.B.; Morozova, O.; Weholt, Ø.; Brandrud, T.E.; Vila, J.; Karich, A.; Reschke, K.; Lorås, J.; et al. Molecular and morphological diversity in the /Rhombisporum clade of the genus Entoloma with a note on E. cocles. Mycol. Prog. 2022, 21, 48. [Google Scholar] [CrossRef]
- Romagnesi, H. Les Rhodophylles de Madagascar (Entoloma, Nolanea, Leptonia, Eccilia, Claudopus). Prodrome Flore Mycol. Madag. 1941, 2, 1–164. [Google Scholar]
- Morozova, O.V.; Popov, E.S.; Kovalenko, A.E. Studies on mycobiota of Vietnam. I. Genus Entoloma: New records and new species. Mikol. Fitopatol. 2012, 46, 184–200. [Google Scholar]
- He, X.-L.; Horak, E.; Li, T.-H.; Peng, W.-H.; Gan, B.-C. Two new cuboid-spored species of Entoloma s.l. (Agaricales, Entolomataceae) from Southern China. Cryptogam. Mycol. 2015, 36, 237–249. [Google Scholar] [CrossRef]
- Horak, E. On cuboid spored species of Entoloma. Sydowia 1976, 28, 171–236. [Google Scholar]
- Horak, E. Additions to ‘On cuboid spored species of Entoloma’. Sydowia 1977, 29, 289–299. [Google Scholar]
- Horak, E. Entoloma in South America. I. Sydowia 1978, 30, 40–110. [Google Scholar]
- Horak, E. Entoloma (Agaricales) in Indomalaya and Australasia. Nova Hedwig. Beih. 1980, 65, 1–352. [Google Scholar]
- Horak, E. Entoloma in South America. II. Sydowia 1982, 35, 75–99. [Google Scholar]
- Pegler, D.N. Agaric Flora of Sri Lanka. Kew Bull. Addit. Ser. 1986, 12, 1–519. [Google Scholar]
- Manimohan, P.; Joseph, A.V.; Leelavathy, K.M. The genus Entoloma in Kerala State, India. Mycol. Res. 1995, 99, 1083–1097. [Google Scholar] [CrossRef]
- Manimohan, P.; Noordeloos, M.E.; Dhanya, A.M. Studies on the genus Entoloma (Basidiomycetes, Agaricales) in Kerala State, India. Persoonia 2006, 19, 45–93. [Google Scholar]
- Noordeloos, M.E.; Morozova, O.V. New and noteworthy Entoloma species from the Primorsky Territory, Russian Far East. Mycotaxon 2010, 112, 231–255. [Google Scholar] [CrossRef]
- Romagnesi, H.; Gilles, G. Les Rhodophylles des forêts côtières du Gabon et de la Côte d’Ivoire. Nova Hedwig. Beih. 1979, 59, 1–649. [Google Scholar]
- Eyssartier, G.; Buyck, B.; Courtecuisse, R. New species and combinations in cuboid-spored Entoloma species from Madagascar. Mycol. Res. 2001, 105, 1144–1148. [Google Scholar] [CrossRef]
- Noordeloos, M.E.; Hausknecht, A. The genus Entoloma (Basidiomycetes, Agaricales) of the Mascarenes and Seychelles. Fungal Divers. 2007, 27, 111–144. [Google Scholar]
- Horak, E. Agaricales of New Zealand 1: Pluteaceae (Pluteus, Volvariella); Entolomataceae (Claudopus, Clitopilus, Entoloma, Pouzarella, Rhodocybe, Richoniella). Fungal Divers. Res. Ser. 2008, 19, 1–305. [Google Scholar]
- Noordeloos, M.E.; Gates, G.M. The Entolomataceae of Tasmania. Fungal Divers. Res. Ser. 2012, 22, 1–400. [Google Scholar] [CrossRef]
- Horak, E.; Cheype, J.-L. Entoloma acutopallidum et Entoloma pileofribrosum deux espèces nouvelles de la Guyane française. Bull. Trimest. Soc. Mycol. Fr. 2008, 124, 287–298. [Google Scholar]
- Karstedt, F.; Capelari, M. Inocephalus (Entolomataceae, Agaricales) from São Paulo State, Brazil. Nova Hedwig. 2013, 96, 279–308. [Google Scholar] [CrossRef]
- Karstedt, F.; Capelari, M. A new species of Entolomataceae with cuboidal basidiospores from the São Paulo Metropolitan Region, Brazil. Mycosphere 2015, 6, 69–73. [Google Scholar] [CrossRef]
- Pegler, D.N. Agaric flora of the Lesser Antilles. Kew Bull. Addit. Ser. 1983, 9, 1–668. [Google Scholar]
- Hesler, L.R. Entoloma in Southeastern North America. Nova Hedwig. Beih. 1967, 23, 1–196. [Google Scholar]
- Largent, D.L. Entolomatoid Fungi of the Western United States and Alaska; Mad River Press: Eureka, CA, USA, 1994; pp. 1–495. [Google Scholar]
- Largent, D.L. The genus Leptonia on the Pacific Coast of the United States including a study of North American types. Bibl. Mycol. 1977, 55, 1–286. [Google Scholar]
- Noordeloos, M.E. Entoloma subgenus Leptonia in northwestern Europe–I. Introduction and a revision of its section Leptonia. Persoonia 1982, 11, 451–471. [Google Scholar]
- Noordeloos, M.E. Entoloma Sensu Lato. Fungi Europaei; Giovanna Biella: Saronno, Italy, 1992; Volume 5, pp. 1–760. [Google Scholar]
- Noordeloos, M.E. Entoloma Sensu Lato. Fungi Europaei; Edizione Candusso: Alassio, Italy, 2004; Volume 5a, pp. 761–1378. [Google Scholar]
- Vidal, J.M.; Bellanger, J.-M.; Moreau, P.-A. Three new gasteroid species of the genus Entoloma found in Spain. Bol. Micol. FAMCAL 2016, 11, 53–78. [Google Scholar]
- Reschke, K.; Morozova, O.V.; Dima, B.; Dima, B.; Cooper, J.A.; Corriol, G.; Biketova, A.Y.; Piepenbring, M.; Noordeloos, M.E. Phylogeny, taxonomy, and character evolution in Entoloma subgenus Nolanea. Persoonia 2022, 49, 136–170. [Google Scholar] [CrossRef]
- Morozova, O.V.; Noordeloos, M.E.; Vila, J. Entoloma subgenus Leptonia in boreal-temperate Eurasia: Towards a phylogenetic species concept. Persoonia 2014, 32, 141–169. [Google Scholar] [CrossRef]
- New, M.; Lister, D.; Hulme, M.; Makin, I. A high-resolution data set of surface climate over global land areas. Clim. Res. 2002, 21, 1–25. [Google Scholar] [CrossRef]
- Tordoff, A.W.; Tran, Q.B.; Nguyen, D.T.; Le, M.H. (Eds.) Sourcebook of Existing and Proposed Protected Areas in Vietnam, 2nd ed.; CD-ROOM; Birdlife International in Indochina and Ministry of Agriculture and Rural Development: Hanoi, Vietnam, 2004; Available online: https://thiennhienviet.org.vn/sourcebook/source_book/index_EN.html (accessed on 15 December 2022).
- Morozova, O.V.; Noordeloos, M.E.; Popov, E.S.; Alexandrova, A.V. Three new species within the genus Entoloma (Basidiomycota, Agaricales) with clamped basidia and a serrulatum-type lamellae edge, and their phylogenetic position. Mycol. Prog. 2018, 17, 381–392. [Google Scholar] [CrossRef]
- Pham, T.H.G.; Morozova, O.V.; Alexandrova, A.V. Boletoid fungi (Boletaceae, Basidiomycota) of protected areas of Kon Tum Plateau (Central Highlands of Vietnam). Turczaninowia 2021, 24, 65–76. Available online: http://turczaninowia.asu.ru/article/view/10442 (accessed on 15 December 2022). [CrossRef]
- Nguyen, T.T. Study on structural characteristics, regeneration and determination of natural forest resilience in Ta Dung National Park, Dak Nong province. Summ. Rep. Sci. Technol. Top. 2021, 22–36. (In Vietnamese) [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978; pp. 1–252. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-a sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Reschke, K.; Noordeloos, M.E.; Manz, C.; Hofmann, T.A.; Rodriguez-Cedeno, J.; Dima, B.; Piepenbring, M. Fungal diversity in the tropics: Entoloma spp. in Panama. Mycol. Progress 2022, 21, 93–145. [Google Scholar] [CrossRef]
- Morgado, L.N.; Noordeloos, M.E.; Lamoureux, Y.; Geml, J. Multi-gene phylogenetic analyses reveal species limits, phylogeographic patterns, and evolutionary histories of key morphological traits in Entoloma (Agaricales, Basidiomycota). Persoonia 2013, 31, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M.; Leroux, J.J.; Strasberg, D.; Edwards, J.; Roets, F.; Hubka, V.; Taylor, P.W.J.; Heykoop, M.; et al. Fungal Planet description sheets: 400–468. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef]
- Chen, Z.H.; Zhang, P. Atlas of Macrofungi in Hunan; Hunan Normal University Press: Changsha, China, 2019; pp. 1–426. (In Chinese) [Google Scholar]
- Varga, T.; Krizsán, K.; Földi, C.; Dima, B.; Sánchez-García, M.; Sánchez-Ramírez, S.; Szöllősi, G.J.; Szarkándi, J.G.; Papp, V.; Albert, L.; et al. Megaphylogeny resolves global patterns of mushroom evolution. Nat. Ecol. Evol. 2019, 3, 668–678. [Google Scholar] [CrossRef]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.; Aime, M.C.; Moncalvo, J.M.; Ge, Z.W.; Yang, Z.L.; Slot, J.C.; Ammirati, J.F.; Baroni, T.J.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 984–997. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008; p. 771. [Google Scholar] [CrossRef]
- Patouillard, N.; Demange, V. Nouvelles contributions’a la flore mycologique du Tonkin. Bull. Soc. Mycol. Fr. 1910, 26, 31–48. [Google Scholar]
- Patouillard, N. Quelques champignons du Tonkin. Bull. Soc. Mycol. Fr. 1913, 29, 206–228. [Google Scholar]
- Heim, R.; Malençon, G. Champignons du Tonkin recueillies par M. V. Demange. Ann. Cryptogam. Exot. 1928, 1, 58–74. [Google Scholar]
- Kiet, T.T. Preliminary checklist of macrofungi of Vietnam. Feddes Reper. 1998, 109, 257–277. [Google Scholar] [CrossRef]
- Le, T.C. (Ed.) Danh lục các loài thực vật Việt Nam: Tập 1 [List of Plant Species in Vietnam: Volume 1]; Agriculture Publishing House: Hanoi, Vietnam, 2001; pp. 1–1183. (In Vietnamese) [Google Scholar]
- Le, B.D. Nấm lớn ở Tây Nguyên [Macrofungi of the Central Highlands]; Science and Technics Publishing House: Hanoi, Vietnam, 2003; pp. 1–261. (In Vietnamese) [Google Scholar]
- Ngo, A. Sự đa dạng của khu hệ nấm lớn ở VQG Bạch Mã, tỉnh Thừa Thiên Huế [Diversity of macrofungi in Bach Ma National Park, Thua Thien Hue province. J. Res. Dev. 2006, 1, 46–52. [Google Scholar]
- Ngo, A.; Nguyen, T.K.C. Sự đa dạng nấm lớn khu bảo tồn thiên nhiên Phong Điền tỉnh Thừa Thiên Huế. Hội nghị Khoa học toàn quốc về Sinh thái và Tài nguyên sinh vật lần thứ 5. [Diversity of macrofungi in Phong Dien Nature Reserve, Thua Thien Hue Province]. In Proceedings of the 5th National Science Conference on Ecology and Biological Resources, Hanoi, Vietnam, 18 October 2013; pp. 346–370. (In Vietnamese). [Google Scholar]
- Kiet, T.T. Macrofungi of Vietnam. V. 1; Science and Technics Publishing House: Hanoi, Vietnam, 2011; pp. 1–314. (In Vietnamese) [Google Scholar]
- Kiet, T.T. Macrofungi of Vietnam. V. 2; Science and Technics Publishing House: Hanoi, Vietnam, 2012; pp. 1–413. (In Vietnamese) [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Hernández-Restrepo, M.; Sutton, D.A.; Acharya, K.; Groenewald, J.Z. Fungal Planet description sheets: 320–370. Persoonia 2015, 34, 167–266. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy GS, J.; Smith, D.; Groenewald, J.Z. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef]
- Morozova, O.V.; Noordeloos, M.E.; Popov, E.S.; Alexandrova, A.V. Nomenclatural novelties. Index Fungorum 2020, 459, 1. [Google Scholar]
- Dörfelt, H.; Kiet, T.T.; Berg, A. Neue Makromyceten-Kollektionen von Vietnam und deren systematische und okogeographische Bedeutung. Feddes Repert. 2004, 115, 164–177. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree derived from Bayesian analysis, based on nrITS1-5.8S-ITS2 region data. Posterior probability (PP > 0.95) values from the Bayesian analysis are added at the nodes. The scale bar represents the number of nucleotide changes per site. The new species are in bold. HT—holotype.
Figure 1.
Phylogenetic tree derived from Bayesian analysis, based on nrITS1-5.8S-ITS2 region data. Posterior probability (PP > 0.95) values from the Bayesian analysis are added at the nodes. The scale bar represents the number of nucleotide changes per site. The new species are in bold. HT—holotype.

Figure 2.
Phylogenetic tree derived from Bayesian analysis, based on tef1α data. Posterior probability (PP > 0.95) values from the Bayesian analysis are added at the nodes. The scale bar represents the number of nucleotide changes per site. The new species are in bold. HT—holotype.
Figure 2.
Phylogenetic tree derived from Bayesian analysis, based on tef1α data. Posterior probability (PP > 0.95) values from the Bayesian analysis are added at the nodes. The scale bar represents the number of nucleotide changes per site. The new species are in bold. HT—holotype.

Figure 3.
Phylogenetic tree derived from Bayesian analysis, based on nrLSU data. Posterior probability (PP > 0.95) values from the Bayesian analysis are added at the nodes. The scale bar represents the number of nucleotide changes per site. The new species are in bold. HT—holotype.
Figure 3.
Phylogenetic tree derived from Bayesian analysis, based on nrLSU data. Posterior probability (PP > 0.95) values from the Bayesian analysis are added at the nodes. The scale bar represents the number of nucleotide changes per site. The new species are in bold. HT—holotype.

Figure 4.
Entoloma cycneum: (a,b). basidiocarps; (c). basidiospores; (d). cheilocystidia; (e). pileipellis; (f). caulocystidia (a,d–f, from LE F-343654, holotype; b—from LE F-343655). Scale bars (a,b) 1 cm, (c–f) 10 μm.
Figure 4.
Entoloma cycneum: (a,b). basidiocarps; (c). basidiospores; (d). cheilocystidia; (e). pileipellis; (f). caulocystidia (a,d–f, from LE F-343654, holotype; b—from LE F-343655). Scale bars (a,b) 1 cm, (c–f) 10 μm.

Figure 5.
Entoloma peristerinum: (a,b). basidiocarps; (c). basidiospores; (d). cheilocystidia; (e). basidium; (f). pileipellis; (g). caulocystidia (a,c–g, from LE F-343653, holotype; b—from LE F-343652). Scale bars (a,b) 1 cm, (c–e) 10 μm, (f,g) 20 μm.
Figure 5.
Entoloma peristerinum: (a,b). basidiocarps; (c). basidiospores; (d). cheilocystidia; (e). basidium; (f). pileipellis; (g). caulocystidia (a,c–g, from LE F-343653, holotype; b—from LE F-343652). Scale bars (a,b) 1 cm, (c–e) 10 μm, (f,g) 20 μm.

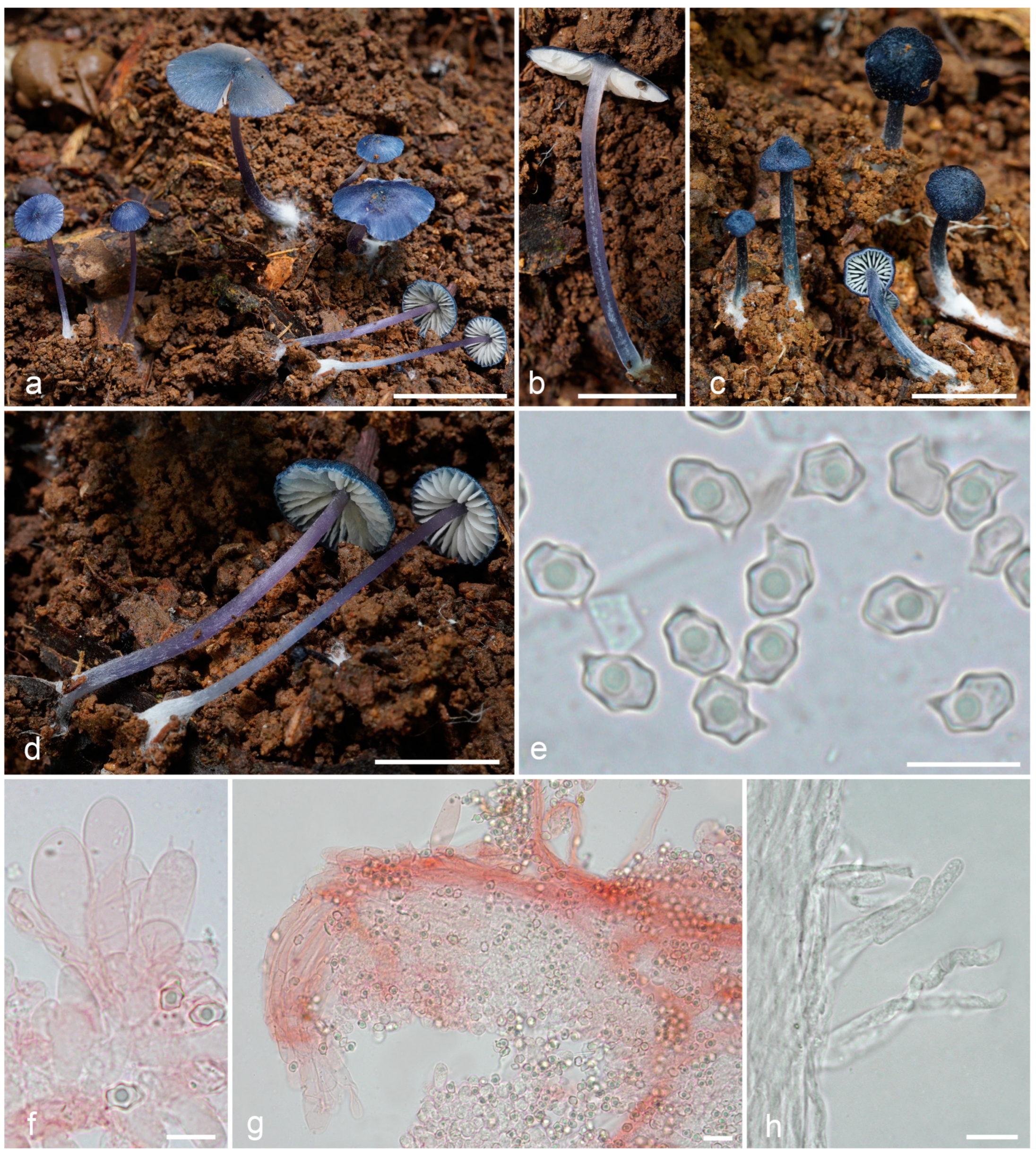
Figure 6.
Entoloma tadungense: (a–d). basidiocarps; (e). basidiospores; (f). cheilocystidia and basidium; (g). pileipellis; (h). caulocystidia (a–f,h from LE F-343680, holotype; g—from LE F-343681). Scale bars (a–d)—1 cm; (e,f,h)—10 μm; (g)—20 μm.
Figure 6.
Entoloma tadungense: (a–d). basidiocarps; (e). basidiospores; (f). cheilocystidia and basidium; (g). pileipellis; (h). caulocystidia (a–f,h from LE F-343680, holotype; g—from LE F-343681). Scale bars (a–d)—1 cm; (e,f,h)—10 μm; (g)—20 μm.

Figure 7.
Entoloma dichroides: (a–d). basidiocarps; (e,f). basidiospores; (g). basidiola and basidium; (h). stipitipellis; (i). pileipellis (all from LE F-343682). Scale bars (a–d)—1 cm; (e–i)—10 μm.
Figure 7.
Entoloma dichroides: (a–d). basidiocarps; (e,f). basidiospores; (g). basidiola and basidium; (h). stipitipellis; (i). pileipellis (all from LE F-343682). Scale bars (a–d)—1 cm; (e–i)—10 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Specimens and GenBank accession numbers of DNA sequences used in the molecular analyses (newly generated sequences are in bold).
Table 1.
Specimens and GenBank accession numbers of DNA sequences used in the molecular analyses (newly generated sequences are in bold).
| Species | Location | Voucher Number | Genbank Accession No. | References | ||
|---|---|---|---|---|---|---|
| ITS | LSU | tef | ||||
| Clitopilopsis hirneola (Fr.) Kühner (Clitopilus hirneolus (Fr.) Kühner & Romagn.) | Italy | MEN 199956 | — | GQ289211 | — | [5] |
| Clitopilus prunulus (Scop.) P. Kumm. | Belgium | MEN 2003-09-14 | — | GQ289149 | — | [5] |
| Entoloma abortivum (Berk. & M.A. Curtis) Donk | Russia: Far East | LE 312419 | MF476905 | MF487792 | — | [42] |
| E. acutipallidum E. Horak & Cheype | Brazil: Pará | SP: FK1893 | — | MG018325 | MH190147 | [7] |
| E. albidoquadratum Manim. & Noordel. | India: Kerala | PM667 (L) | — | GQ289151 | — | [5] |
| E. allochroum Noordel. | The Netherlands | v. Waveren, 29-07-1973, holotype (L) | KC898372 | — | — | [39] |
| E. altissimum (Massee) E. Horak | Vietnam | LE 262945 | MF476912 | — | — | [42] |
| E. argillaceum O.V. Morozova et al. | Russia: Caucasus | LE 311861, holotype | — | OL338531 | OL405537 | [38] |
| E. araneosum (Quél.) M.M. Moser | Belgium | MEN 200314 | KC710056 | GQ289153 | — | [5] |
| E. atricolor O.V. Morozova et al. | Vietnam | LE 295000, holotype | KY777496 | — | — | [42] |
| E. aurantiovirescens Reschke, Lotz-Winter & Noordel. | Panama | KaiR623, holotype | MZ611665 | — | — | [50] |
| E. austriacum Courtec. [E. tjallingiorum var. laricinum O.V. Morozova et al.] | Russia: Far East | LE 254343, holotype | KC898413 | KC898513 | — | [39] |
| E. bidupense O.V. Morozova & E.S. Popov | Vietnam | LE 262935 | — | NG_059265 | — | [42] |
| E. callichroum E. Horak & Noordel. | Switzerland | ZT 71/58, holotype | KC898350 | — | — | [39] |
| E. cervinum (Karstedt & Capelari) Blanco-Dios [Inocephalus cervinus Karstedt & Capelari] | Brazil | SP: FK 0940 | — | — | MH190138 | [7] |
| E. cetratum (Fr.) M.M. Moser | Russia: European part | LE 235480 | KC898450 | — | — | [39] |
| E. chalybaeum (Pers.) Noordel. | Russia: European part | LE 254353 | KC898445 | KC898500 | — | [39] |
| E. chytrophilum Wölfel, Noordel. & Dähncke | Spain: Canary Islands | L 855, holotype | KC898434 | KC898519 | — | [39] |
| E. coelestinum (Fr.) Hesler | Russia: Ural | LE 258103 | KC898362 | KC898524 | — | [39] |
| E. conferendum (Britzelm.) Noordel. | MEN 200330 | KC710055 | KC710133 | — | [51] | |
| E. cycneum O.V. Morozova et T.H.G. Pham | Vietnam | LE F-343654, holotype | OQ779461 | OQ804518 | OQ779183 | this work |
| E. cycneum | Vietnam | LE F-343655 | OQ779463 | OQ804519 | OQ779182 | this work |
| E. cycneum | Vietnam | LE F-343656 | OQ779462 | OQ804521 | OQ779181 | this work |
| E. cycneum | Vietnam | LE F-343657 | OQ779464 | OQ804520 | OQ779184 | this work |
| E. cycneum | Vietnam | LE F-343658 | OQ779465 | — | OQ779185 | this work |
| E. cystidiophorum Dennis [Inocephalus cystidiophorus (Dennis) Karstedt & Capelari] | Brazil: São Paulo | SP: FK1023 | — | MG018319 | MH190140 | [7] |
| E. dennisii E. Horak [Inocephalus dennisii (E. Horak) Karstedt & Capelari] | Puerto Rico | CORT: 8263 TJB | — | — | MH190164 | [7] |
| E. dichroides O.V. Morozova et T.H.G. Pham | Vietnam | LE F-343682 | OQ779472 | OQ804527 | — | this work |
| E. dichroum (Pers.) P. Kumm. | Russia: European part | LE 227472, neotype | KC898440 | — | — | [39] |
| E. dichroum | Russia: European part | LE 234260 | — | KC898527 | — | [39] |
| E. euchroum (Pers.) Donk | Russia: Caucasus | LE 262995 | KC898417 | KC898516 | — | [39] |
| E. eugenei Noordel. & O.V. Morozova | Russia: Primorsky Territory | LE 253771, holotype | KC898438 | KC898529 | — | [39] |
| E. griseocyaneum (Fr.) P. Kumm. | Russia: Caucasus | LE 254351 | KC898444 | KC898498 | — | [39] |
| E. inocephalum (Romagn.) Dennis | Vietnam | LE 262922 | KC898449 | MH259311 | MH190154 | [7,39] |
| E. kovalenkoi O.V. Morozova, E.S. Popov & A.V. Alexandrova | Vietnam | LE 312529 | OK257210 | OK257207 | OK256169 | [3] |
| E. kruticianum O.V. Morozova, M.Y. Dyakov, E.S. Popov & A.V. Alexandrova | Russia: European part | LE 311767, holotype | KU666558 | KU710222 | — | [52] |
| E. lampropus (Fr.) Hesler | Sweden | UPS:BOT:F-176490, neotype | KC898377 | KC898506 | — | [39] |
| E. lasium (Berk. & Broome) Noordel. & Co-David [Pouzarella lasia (Berk. & Broome) Largent & Abell-Davis] | Australia: Queensland | DLL9811 (BRI, CNS) | — | — | MG702641 | [7] |
| E. lepidissimum (Svrček) Noordel. | Czech Republic | PRM 755801, holotype | KC898364 | KC898532 | — | [39] |
| E. luteolamellatum (Largent & Aime) Blanco-Dios [Trichopilus luteolamellatus Largent & Aime] | Guyana | MCA 1480, holotype | — | MH190213 | MG702644 | [7] |
| E. luteum Peck [Inocephalus luteus (Peck) T.J. Baroni] | USA: New York | CORT: 7771 TJB | — | MH190212 | MH190161 | [7] |
| E. mucronatum (Karstedt & Capelari) Blanco-Dios [Inocephalus mucronatus Karstedt & Capelari] | Brazil: São Paulo | SP: FK1033, holotype | — | MH190174 | MH190141 | [7] |
| E. murrayi (Berk. & M.A. Curtis) Sacc. & P. Syd. | China | MHHNU 30602 | MK250917 | — | — | [53] |
| E. murrayi | USA | CORT: TJB 8210 | — | MH190193 | — | [7] |
| E. nitidum Quél. | Slovakia | MEN 200324 | KC710122 | GQ289175 | — | [5] |
| E. pallidoflavum(Henn. & E. Nyman) E. Horak | Vietnam | LE 262934 | OQ779469 | MH190183 | MH190155 | [7], this work |
| E. panniculus (Berk.) Sacc. | Australia | G1680 | — | MK278012 | — | [54] |
| E. percoelestinum O.V. Morozova, Noordel., Vila & Bulyonk. | Spain | LE 254390, holotype | KF745927 | KF745928 | — | [39] |
| E. peristerinum O.V. Morozova et T.H.G. Pham | Vietnam | LE F-343653, holotype | OQ779466 | OQ804522 | OQ779188 | this work |
| E. peristerinum | Vietnam | LE F-343649 | OQ779468 | — | — | this work |
| E. peristerinum | Vietnam | LE F-343650 | OQ779467 | OQ804524 | OQ779186 | this work |
| E. peristerinum | Vietnam | LE F-343652 | — | OQ804525 | OQ779187 | this work |
| E. peristerinum | Vietnam | LE F-343651 | — | OQ804523 | OQ779189 | this work |
| E. placidum (Fr.) Noordel. | Sweden | UPS:BOT:F-121714, epitype, | KC898394 | KC898514 | — | [39] |
| E. plicatum (Largent) Blanco-Dios | USA | DAR: DLL10091 | — | — | MG702627 | [7] |
| E. prunuloides (Fr.) Quél. | USA | AFTOL-ID: 523 4765 TJB | — | — | DQ457633 | [55] |
| E. quadratum (Berk. & M.A. Curtis) E. Horak [Inocephalus quadratus (Berk. & M.A. Curtis) T.J. Baroni] | Russia: Far East | LE254355 | KC898452 | KC898504 | — | [39] |
| E. quadratum | USA: New York | CORT: TJB 8214 | — | — | MH190162 | [7] |
| E. sericellum (Fr.) P. Kumm. [Alboleptonia sericella (Fr.) Largent & R.G. Benedict] | Russia: Caucasus | LE 254362 | KC898453 | — | — | [39] |
| E. sericellum | USA: California | HSU: DLL9524 | — | — | MG702617 | [7] |
| E. sericellum | Belgium | MEN 200315 | — | GQ289190 | — | [5] |
| E. serrulatum (Fr.) Hesler | Russia: Caucasus | LE 254361 | KC898447 | KC898501 | — | [39] |
| E. sublaevisporum Vila, Noordel. & O.V. Morozova | Spain | LIP JVG 1070823T, holotype | KC898436 | KC898518 | — | [39] |
| E. tadungense O.V. Morozova et T.H.G. Pham | Vietnam | LE F-343680, holotype | OQ779470 | — | — | this work |
| E. tadungense | Vietnam | LE F-343681 | OQ779471 | OQ804526 | — | this work |
| E. tibiiforme (Largent & Aime) Blanco-Dios [Trichopilus tibiiformis Largent & Aime] | Guyana | MCA2426, holotypus (BRG, LSUM) | — | — | MG702645 | [7] |
| E. tjallingiorum Noordel. var. tjallingiorum | Sweden | UPS:BOT:F-016378, holotype | KC898412 | KC898509 | — | [39] |
| E. turbidum (Fr.) Quél. | Slovakia | MEN 200351 | KC710060 | GQ289201 | — | [5] |
| E. umbrophilum Noordel. & Hauskn. [Leptonia umbrophila (Noordel. & Hauskn.) Largent] | Australia: Queensland | DLL9766 (BRI, CNS) | — | — | MG702638 | [7] |
| E. undatum (Gillet) M.M. Moser | Russia: European part | LE 312417 | MF476910 | MF487801 | — | [42] |
| E. venustum Wölfel & F. Hampe | Germany | L, Wö E17/10, holotype | KC898355 | KC898523 | — | [39] |
| E. vernum S. Lundell | Russia: European part | LE 312538 | OL338282 | OL338537 | OL405539 | [38] |
| E. violaceozonatum Noordel. & Liiv | Estonia | L 275, holotype | KC898448 | KC898502 | — | [39] |
| E. virescens (Sacc.) E. Horak ex Courtec. [Inocephalus virescens (Berk. & M.A. Curtis) Largent & Abell-Davis] | MEL:2379813 | MF977981 | — | — | Genbank | |
| E. virescens | Guyana | MCA 2479 | — | GU384622 | MG702629 | [7] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Morozova, O.; Pham, T.H.G. New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam. J. Fungi 2023, 9, 621. https://doi.org/10.3390/jof9060621
AMA Style
Morozova O, Pham THG. New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam. Journal of Fungi. 2023; 9(6):621. https://doi.org/10.3390/jof9060621
Chicago/Turabian StyleMorozova, Olga, and Thi Ha Giang Pham. 2023. "New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam" Journal of Fungi 9, no. 6: 621. https://doi.org/10.3390/jof9060621
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.