
Re-Valuation of the Taxonomic Status of Species within the Inocybe similis Complex
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
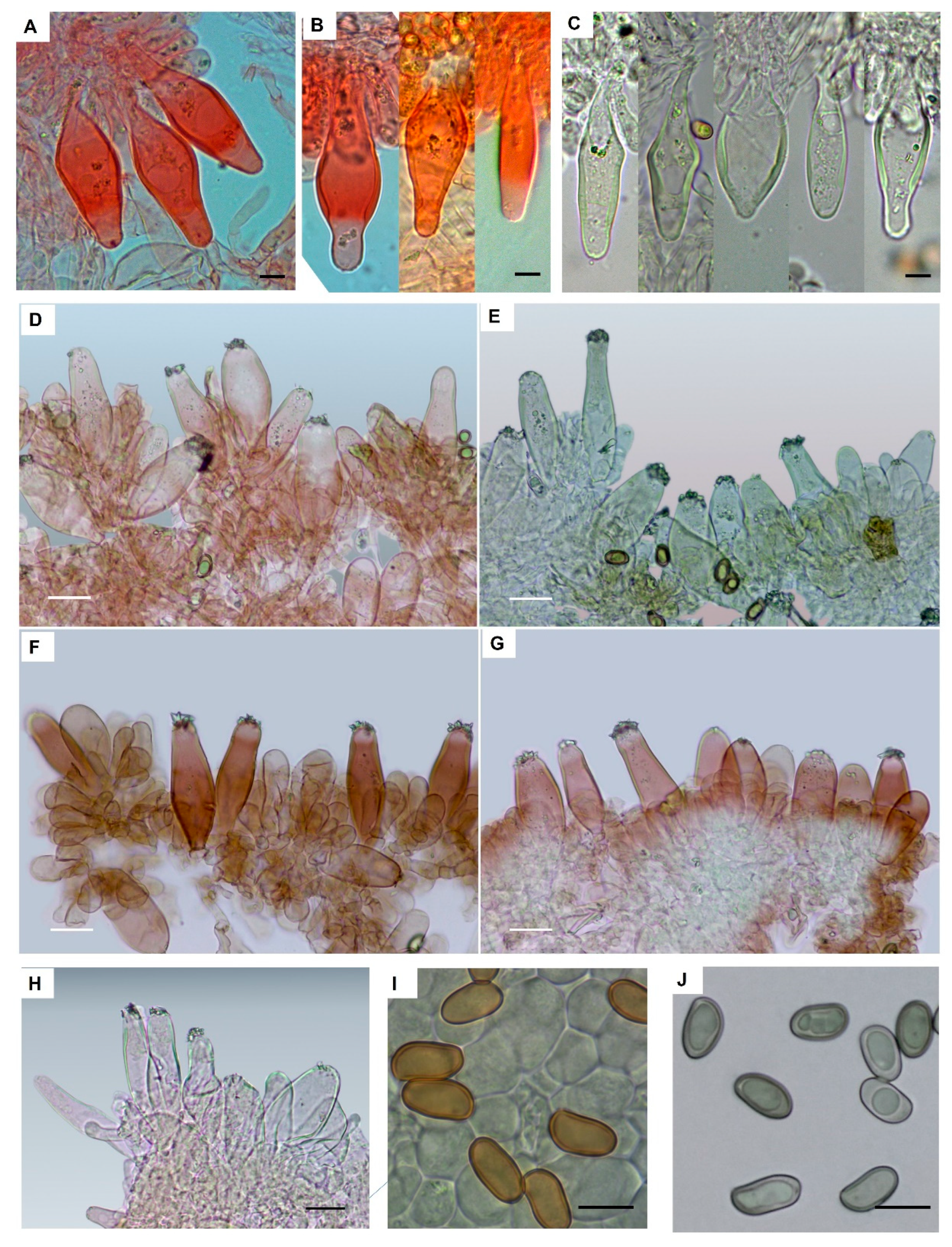
2.1. Morphology
2.2. Molecular Phylogeny
2.3. Statistical Analyses
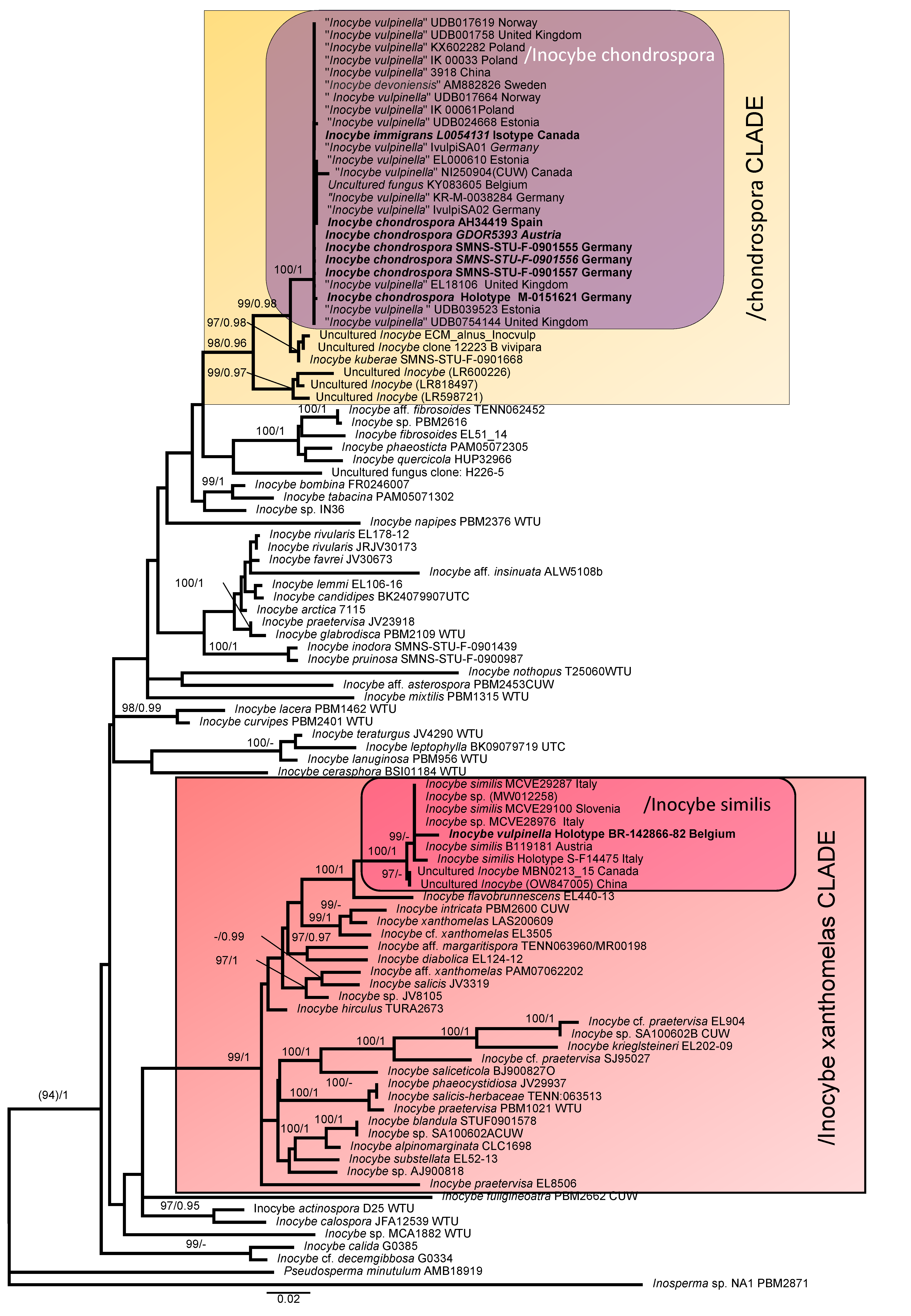
3. Results
3.1. Molecular Phylogeny
3.2. Statistical Analysis
3.3. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matheny, P.B.; Hobbs, A.M.; Esteve-Raventós, F. Genera of Inocybaceae: New skin for the old ceremony. Mycologia 2019, 112, 83–120. [Google Scholar] [CrossRef]
- Lurie, Y.; Wasser, S.P.; Taha, M.; Shehade, H.; Nijim, J.; Hoffmann, Y.; Basis, F.; Vardi, M.; Lavon, O.; Suaed, S.; et al. Mushroom poisoning from species of genus Inocybe (fiber head mushroom): A case series with exact species identification. Clin. Toxicol. 2009, 47, 562–565. [Google Scholar] [CrossRef]
- Patocka, J.; Wu, R.; Nepovimova, E.; Valis, M.; Wu, W.; Kuca, K. Chemistry and Toxicology of Major Bioactive Substances in Inocybe Mushrooms. Int. J. Mol. Sci. 2021, 22, 2218. [Google Scholar] [CrossRef]
- Kosentka, P.; Sprague, S.L.; Ryberg, M.; Gartz, J.; May, A.L.; Campagna, S.R.; Matheny, P.B. Evolution of the toxins muscarine and psilocybin in a family of mushroom-forming fungi. PLoS ONE 2013, 8, e64646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizzini, A.; Della Maggiora, M.; Tolaini, F.; Ercole, E. A new cryptic species in the genus Tubariomyces (Inocybaceae, Agaricales). Mycol. Prog. 2013, 12, 375–381. [Google Scholar] [CrossRef]
- Matheny, P.B.; Bougher, N.L. Fungi of Australia Inocybaceae; ABRS & CSIRO Publishing: Canberra/Melbourne, Australia, 2017; pp. 1–592. [Google Scholar]
- Cripps, C.L.; Larsson, E.; Vauras, J. Nodulose-spored Inocybe from the Rocky Mountain alpine zone molecularly linked to European and type specimens. Mycologia 2019, 112, 133–153. [Google Scholar] [CrossRef]
- Bandini, D.; Oertel, B.; Eberhardt, U. A fresh outlook on the smooth-spored species of Inocybe: Type studies and 18 new species. Mycol. Prog. 2021, 20, 1019–1114. [Google Scholar] [CrossRef]
- Einhellinger, A. Problems of Inocybe similis Bres., vulpinella Bruylants (incl. Inocybe chondrospora Einh. et Stangl) and its variety fuscolamellata Bon. Z. Mykol. 1994, 60, 365–372. [Google Scholar]
- Bresadola, J. Hymenomycetes novi vel minus cogniti. Ann. Mycol. 1905, 3, 159–164. [Google Scholar]
- Dovana, F.; Ferisin, G.; Bizio, E.; Bandini, D.; Olariaga, I.; Esteve-Raventós, F. A morphological and phylogenetic characterisation of Inocybe similis (Agaricales, Inocybaceae), a rare species described by Bresadola in 1905. Phytotaxa 2020, 474, 71–80. [Google Scholar] [CrossRef]
- Kuyper, T.W. A revision of the genus Inocybe in Europe 1. Subgenus Inosperma and the smooth-spored species of subgenus Inocybe. Pers. Suppl. 1986, 3, 1–247. [Google Scholar]
- Bruylants, J. Inocybe vulpinella sp. nov. Bull. Soc. Mycol. Fr. 1979, 85, 340–344. [Google Scholar]
- Einhellinger, A.; Stangl, J. Inocybe chondrospora Einhellinger & Stangl spec. nov. Z Mykol. 1979, 45, 163–165. [Google Scholar]
- Malloch, D. An undescribed species of Inocybe from mine wastes in Ontario. Can. J. Bot. 1982, 60, 40–45. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf material. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Dovana, F.; Contu, M.; Angeli, P.; Brandi, A.; Mucciarelli, M. Leucoagaricus ariminensis sp. nov., a lilac species from Italy. Mycotaxon 2017, 132, 205–216. [Google Scholar] [CrossRef]
- Stielow, B.; Hensel, G.; Strobelt, D.; Makonde, H.M.; Rohde, M.; Dijksterhuis, J.; Klenk, H.P.; Göker, M. Hoffmannoscypha, a novel genus of brightly coloured, cupulate Pyronemataceae closely related to Tricharina and Geopora. Mycol. Prog. 2013, 12, 675–686. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Tedersoo, L.; Mett, M.; Ishida, T.A.; Bahram, M. Phylogenetic relationships among host plants explain differences in fungal species richness and community composition in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4239–4246. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ryberg, M.; Nilsson, R.H.; Kristiansson, E.; Töpel, M.; Jacobsson, S.; Larsson, E. Mining metadata from unidentified ITS sequences in GenBank: A case study in Inocybe (Basidiomycota). BMC Evol. Biol. 2008, 8, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vauras, J.; Kokkonen, K. Finnish records on the genus Inocybe. The new species Inocybe saliceticola. Karstenia 2009, 48, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Osmundson, T.W.; Robert, V.A.; Schoch, C.L.; Baker, L.J.; Smith, A.; Robich, G.; Mizzan, L.; Garbelotto, M.M. Filling gaps in biodiversity knowledge for macrofungi: Contributions and assessment of an herbarium collection DNA barcode sequencing project. PLoS ONE 2013, 8, E62419. [Google Scholar] [CrossRef] [Green Version]
- Esteve-Raventós, F.; Moreno, G.; Bizio, E.; Alvarado, P. Inocybe flavobrunnescens, a new species in section Marginatae. Mycol. Prog. 2015, 14, 14. [Google Scholar] [CrossRef]
- Vauras, J.; Larsson, E. Inocybe caprimulgi and I. lacunarum, two new nodulose-spored species from Fennoscandia. Karstenia 2015, 55, 14. [Google Scholar] [CrossRef] [Green Version]
- Esteve-Raventós, F.; Alvarado, P.; Olariaga, I. Unraveling the Inocybe praetervisa group through type studies and ITS data: Inocybe praetervisoides sp. nov. from the Mediterranean region. Mycologia 2016, 108, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Bandini, D.; Oertel, B.; Moreau, P.-A.; Thines, M.; Ploch, S. Three new hygrophilous species of Inocybe, subgenus Inocybe. Mycol. Prog. 2019, 18, 1101–1119. [Google Scholar] [CrossRef]
- Kropp, B.R.; Matheny, P.B.; Nanagyulyan, S.G. Phylogenetic taxonomy of the Inocybe splendens group and evolution of supersection “Marginatae”. Mycologia 2010, 102, 560–573. Available online: http://www.jstor.org/stable/27811069 (accessed on 20 January 2017). [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformation 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, R.; Serra, F.; Tárraga, J.; Medina, I.; Carbonell, J.; Pulido, L.; de María, A.; Capella-Gutierrez, S.; Huerta-Cepas, J.; Gabaldón, T.; et al. Phylemon 2.0: A suite of web-tools for molecular evolution, phylogenetics, phylogenomics and hypotheses testing. Nucl. Acids Res. 2011, 39, W470–W474. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.F.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2015, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Le Vinh, S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P.; Teslenko, M. MrBayes Version 3.2 Manual: Tutorials and Model Summaries. 2011. Available online: Mrbayes.sourceforge.net/mb3.2_manual.pdf (accessed on 19 October 2021).
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendiburu, F.D.; Yaseen, M. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.4.0. Available online: https://myaseen208.github.io/agricolae/https://cran.r-project.org/package=agricolae (accessed on 10 December 2020).
- Esteve-Raventós, F. Inocybe vulpinella Bruyl. (Inocybaceae, Agaricomycetes), primer registro para la Península Ibérica. Zizak 2008, 5, 27–33. [Google Scholar]
- Stangl, J. Die Gattung Inocybe in Bayern. Hoppea 1989, 46, 1–409. [Google Scholar]
- Orton, P.D. New check list of British agarics and boleti. Part III. Notes on genera and species in the list. Trans. Br. Mycol. Soc. 1960, 43, 274. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Collection | Country | GenBank/UNITE | |
|---|---|---|---|---|
| ITS | nLSU | |||
| Inocybe actinospora | D25 WTU | Argentina | X | AY380363 |
| Inocybe alpinomarginata | CLC1698 | USA: Colorado | MK153647 | MK153647 |
| Inocybe arctica | 7115 | Sweden | KY033839 | KY033839 |
| Inocybe aff. asterospora | PBM2453 CUW | Unknown | X | AY702015 |
| Inocybe blandula | SMNS-STU-F-0901578 | Austria | MZ144124 | MZ144124 |
| Inocybe bombina | FR 0246007 | Germany | NR_173842 | X |
| Inocybe calida | G0385 | Hungary | MK278241 | MK278241 |
| Inocybe calospora | JFA12539WTU | Sweden | X | AY038313 |
| Inocybe candidipes | BK24079907 UTC | USA: Arizona | X | AY239019 |
| Inocybe cerasphora | BSI01184 WTU | Chile | X | AY380370 |
| /Inocybe chondrospora ** | ||||
| Inocybe vulpinella ** | 3918 | China | KR082885 | X |
| Inocybe chondrospora ** | AH34419 | Spain | OR088565 * | X |
| Inocybe vulpinella ** | EL000610 | Estonia | AM882825 | X |
| Inocybe vulpinella ** | EL18106 | United Kingdom | FN550898 | FN550898 |
| Inocybe chondrospora ** | GDOR5393 | Austria | OR088564 * | X |
| Inocybe vulpinella ** | IK-00033 | Poland | KX602283 | X |
| Inocybe vulpinella ** | IK-00034 | Poland | KX602282 | X |
| Inocybe vulpinella ** | IK-00061 | Poland | KX602284 | X |
| Inocybe vulpinella ** | IvulpiSA01 | Germany | MH389771 | X |
| Inocybe vulpinella ** | IvulpiSA02 | Germany | MH389774 | X |
| Inocybe vulpinella ** | KR-M-0038284 | Germany | MK929256 | X |
| Inocybe immigrans ** | L0054131 (Isotype) | Canada | MW539061 * | |
| Inocybe chondrospora ** | M-0151621 (Holotype) | Germany | OR098441 * | OR098446 * |
| Inocybe vulpinella ** | NI250904 (CUW) | Canada: Ontario | EU307834 | |
| Uncultured fungus ** | OTU_127 | Belgium | KY083605 | X |
| Uncultured fungus ** | saf_F1979 | Italy: Aosta Valley | MW164608 | X |
| Inocybe chondrospora ** | SMNS-STU-F-0901555 | Germany | MW539060 * | MW539060 * |
| Inocybe chondrospora ** | SMNS-STU-F-0901556 | Netherlands | MW539058 * | MW539058 * |
| Inocybe chondrospora ** | SMNS-STU-F-0901557 | Germany | MW539059 * | MW539059 * |
| Inocybe devoniensis ** | TAA17205 | Sweden | AM882826 | X |
| Inocybe vulpinella ** | UDB001758 | United Kingdom | UDB001758 | X |
| Inocybe vulpinella ** | UDB017619 | Norway | UDB017619 | X |
| Inocybe vulpinella ** | UDB024668 | Estonia | UDB024668 | X |
| Inocybe vulpinella ** | UDB039523 | Estonia | UDB039523 | X |
| Inocybe vulpinella ** | UDB0754144 | United Kingdom | UDB0754144 | X |
| Inocybe curvipes | PBM2401 WTU | USA: Washington | X | AY239022 |
| Inocybe cf. decemgibbosa | G0334 | Hungary | MK278224 | MK278224 |
| Inocybe diabolica | EL124-12 | Norway | KT958913 | KT958913 |
| Inocybe favrei | JV30673 | Norway | KY033798 | KY033798 |
| Inocybe aff. fibrosoides | TENN062452 | USA: Tennessee | MT237492 | MT228850 |
| Inocybe fibrosoides | EL51_14 | Sweden | KY033846 | KY033846 |
| Inocybe flavobrunnescens | EL440-13 | Spain | MK153641 | MK153641 |
| Inocybe fuligineoatra | PBM2662 CUW | USA: Tennessee | EU523589 | EU307831 |
| Inocybe glabrodisca | PBM 2109 (WTU) | USA: Washington | AY239023 | AY239023 |
| Inocybe glabrodisca | PBM2109 WTU | USA: Washington | X | AY239023 |
| Inocybe hirculus | TURA2673 | Finland | MT241840 | MT241840 |
| Inocybe inodora | SMNS-STU-F-0901439 | Austria | MT101875 | MT101875 |
| Inocybe aff. insinuata | ALW5108b (WTU) | USA: Louisiana | X | JN975030 |
| Inocybe intricata | PBM2600 CUW | USA: Tennessee | EU523561 | EU307835 |
| Inocybe krieglsteineri | EL202-09 | Sweden | KT958916 | KT958916 |
| Inocybe kuberae | SMNS-STU-F-0901668 | Germany | ON003427 | ON003427 |
| Inocybe lacera | PBM1462 WTU | USA: Washington | X | AY038318 |
| Inocybe lanuginosa | PBM956 WTU | USA: Washington | X | AY038319 |
| Inocybe lemmi | EL106-16 | Sweden | MG574395 | MG574395 |
| Inocybe leptophylla | BK09079719 UTC | USA: Utah | X | AY038320 |
| Inocybe aff. margaritispora | TENN:063960/MR00198 | USA: Tennessee | KP308775 | JN974998 |
| Inocybe mixtilis ceskae | PBM1315 WTU | USA: Washington | X | AY380387 |
| Inocybe napipes | PBM2376 WTU | Norway | X | AY239024 |
| Inocybe nothopus | Trappe 25060 (WTU) | Australia: New South Wales | X | AY380388 |
| Inocybe phaeocystidiosa | JV29937 | Sweden | MK153638 | MK153638 |
| Inocybe phaeosticta | PAM05072305 | France | HQ586873 | HQ641110 |
| Inocybe cf. praetervisa | EL904 | Sweden | AM882718 | AM882718 |
| Inocybe cf. praetervisa | SJ95027 | Sweden | AM882721 | AM882721 |
| Inocybe praetervisa | JV23918 | Italy | KY033785 | KY033785 |
| Inocybe praetervisa | EL8506 | Sweden | FN550892 | FN550892 |
| Inocybe praetervisa | PBM1021 WTU | USA: Washington | X | AY038322 |
| Inocybe pruinosa | SMNS-STU-F-0900987 | Germany | MT101877 | MT101877 |
| Inocybe quercicola | HUP32966 | Pakistan | MK368637 | MN812170 |
| Inocybe rivularis | JRJV30173 | Finland | KY033813 | KY033813 |
| Inocybe rivularis | EL178-12 | Finland | KY033823 | KY033823 |
| Inocybe saliceticola | BJ900827O | Sweden | AM882717 | AM882717 |
| Inocybe salicis | JV3319 | KT958906 | KT958906 | |
| Inocybe salicis-herbaceae | TENN:063513 | X | KP170999 | |
| /Inocybe similis | ||||
| Inocybe similis *** | B11-9-18-1 | Austria: Forchach | MT504413 | X |
| Inocybe vulpinella *** | BR-142866-82 (Holotype) | OR098440 * | X | |
| Uncultured Inocybe *** | MBN0213_15 | Canada | KC840624 | X |
| Inocybe similis *** | MCVE28976 | Italy: Grado | KY848219 | X |
| Inocybe similis *** | MCVE29100 | Slovenia | KY848218 | X |
| Inocybe similis *** | MCVE29287 | Italy | KY848217 | KY848221 |
| Inocybe similis *** | S-F14475 (Holotype) | Italy: Trentino Alto Adige | MT704951 | X |
| Inocybe sp. *** | Unknown | France | MW012258 | X |
| Uncultured Inocybe *** | Unknown | China | OW847005 | X |
| Inocybe sp. | IN36 | USA: Indiana | OM473594 | X |
| Inocybe sp. | JV8105 | KT958910 | KT958910 | |
| Inocybe sp. | SA100602B CUW | Slovakia | X | EU307838 |
| Inocybe sp. | AJ900818 | KT958923 | KT958923 | |
| Inocybe sp. | SA100602A CUW | Slovakia | KP636858 | EU307837 |
| Inocybe sp. | MCA1882 WTU | Guyana | X | AY509115 |
| Inocybe sp. PBM 2617 | PBM 2616 | USA: Tennessee | EU523567 | X |
| Inocybe substellata | EL52-13 | Sweden | KT958927 | KT958927 |
| Inocybe tabacina | PAM05071302 | France | HQ586865 | HQ641106 |
| Inocybe teraturgus | JV4290 WTU | ? | X | AY239027 |
| Inocybe aff. xanthomelas | PAM07062202 | France | HQ586861 | HQ641104 |
| Inocybe cf. xanthomelas | EL3505 | Norway | AM882989 | AM882989 |
| Inocybe xanthomelas | LAS200609 | Sweden | FN550895 | FN550895 |
| Uncultured fungus | clone: H226-5 | China: Huayuan, Hunan | AB636455 | X |
| Uncultured Inocybe | ECM_alnus_Inocvulp | France | JQ890277 | X |
| Uncultured Inocybe | clone 12223_B_vivipara | Germany | KF000619 | X |
| Uncultured Inocybe | China | LR598721 | X | |
| Uncultured Inocybe | China | LR600226 | X | |
| Uncultured Inocybe | China | LR818497 | X | |
| OUTGROUP | ||||
| Inosperma sp. NA2 | PBM2871 | USA: Connecticut | HQ201348 | HQ201348 |
| Pseudosperma minutulum | AMB 18919 | Italy | ON202637 | ON202637 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dovana, F.; Bandini, D.; Eberhardt, U.; Olariaga, I.; Bizio, E.; Ferisin, G.; Esteve-Raventós, F. Re-Valuation of the Taxonomic Status of Species within the Inocybe similis Complex. J. Fungi 2023, 9, 679. https://doi.org/10.3390/jof9060679
Dovana F, Bandini D, Eberhardt U, Olariaga I, Bizio E, Ferisin G, Esteve-Raventós F. Re-Valuation of the Taxonomic Status of Species within the Inocybe similis Complex. Journal of Fungi. 2023; 9(6):679. https://doi.org/10.3390/jof9060679
Chicago/Turabian StyleDovana, Francesco, Ditte Bandini, Ursula Eberhardt, Ibai Olariaga, Enrico Bizio, Giuliano Ferisin, and Fernando Esteve-Raventós. 2023. "Re-Valuation of the Taxonomic Status of Species within the Inocybe similis Complex" Journal of Fungi 9, no. 6: 679. https://doi.org/10.3390/jof9060679