
Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata)

1
M.L. Bean Life Science Museum and Department of Biology, Brigham Young University, Provo, UT 84602, USA
2
Department of Biology, Boise State University, Boise, ID 83725, USA
3
Plant and Wildlife Sciences, Brigham Young University, Provo, UT 84602, USA
4
Department of Biology, Eastern Washington University, Cheney, WA 99004, USA
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(7), 698; https://doi.org/10.3390/jof9070698
Submission received: 10 May 2023
/
Revised: 18 June 2023
/
Accepted: 19 June 2023
/
Published: 24 June 2023
(This article belongs to the Special Issue Lichens as Bioindicators of Global Change Drivers)
Abstract
:Thirty years after its designation as a federally endangered species, the Florida Perforate Cladonia (FPC) remains imperiled in isolated populations in the Florida scrub in the southeastern USA. For threatened and endangered species, such as FPC, reference genomes provide critical insight into genomic diversity, local adaptations, landscape-level genetics, and phylogenomics. Using high-throughput sequencing, we assemble the first draft nuclear and mitochondrial genomes for the FPC mycobiont—Cladonia perforata. We also assess genetic diversity within and among populations in southeastern Florida using genome-scale data and investigate diversity across the entire nuclear ribosomal cistron, including the standard DNA barcoding marker for fungi. The draft nuclear genome spanned 33.6 Mb, and the complete, circular mitochondrial genome was 59 Kb. We also generated the first chloroplast genome, to our knowledge, for the photobiont genus associated with FPC, an undescribed Asterochloris species. We inferred the presence of multiple, distinct mycobiont parental genotypes (genets) occurring at local scales in southeastern Florida, and strikingly, no genets were shared among even the closest sample sites. All sampled thalli shared identical mitochondrial genomes, while the nuclear ribosomal cistron showed limited variability—highlighting the genetic resolution provided by nuclear genome-scale datasets. The genomic resources generated here provide critical resources for informed conservation efforts for the FPC.
Keywords:
conservation; Florida scrub; genet; Illumina; MinION; Oxford Nanopore Technologies; ramet; reindeer lichens; transplant1. Introduction
Florida scrub habitat in the southeastern United States is characterized by well-drained, nutrient-poor sandy soils and supports a diverse assemblage of rare and endemic plant and animal species. In the Florida scrub, unique plant communities are associated with coastal dunes that formed during the Pleistocene, with the scrub vegetation likely remaining intact since the ancient formation of the dunes [1,2,3]. The Florida scrub also supports some of the highest densities of rare, endemic species in the United States, including over two-thirds of Florida’s reptiles and amphibians [4], a number of plants [5,6], arthropods [7], and lichens [8]. This pyrogenic ecosystem is a global biodiversity hotspot where many members of the community are dependent on periodic fire for habitat maintenance [9]. Varying fire return intervals create and maintain open sand gaps where many specialized species occur, including herbaceous plants and lichens [10,11].
Florida scrub habitats are imperiled and in rapid decline, primarily due to anthropogenic activities such as urbanization, agriculture, and land development [12,13], in addition to catastrophic fires and hurricanes [11,14,15]. Furthermore, the Florida scrub is vulnerable to climate change, which is expected to exacerbate the negative impacts of current threats, such as increased frequency and intensity of wildfires, droughts, and hurricanes, further reducing the available habitat for many of the specialized species that rely on this ecosystem [12,16].
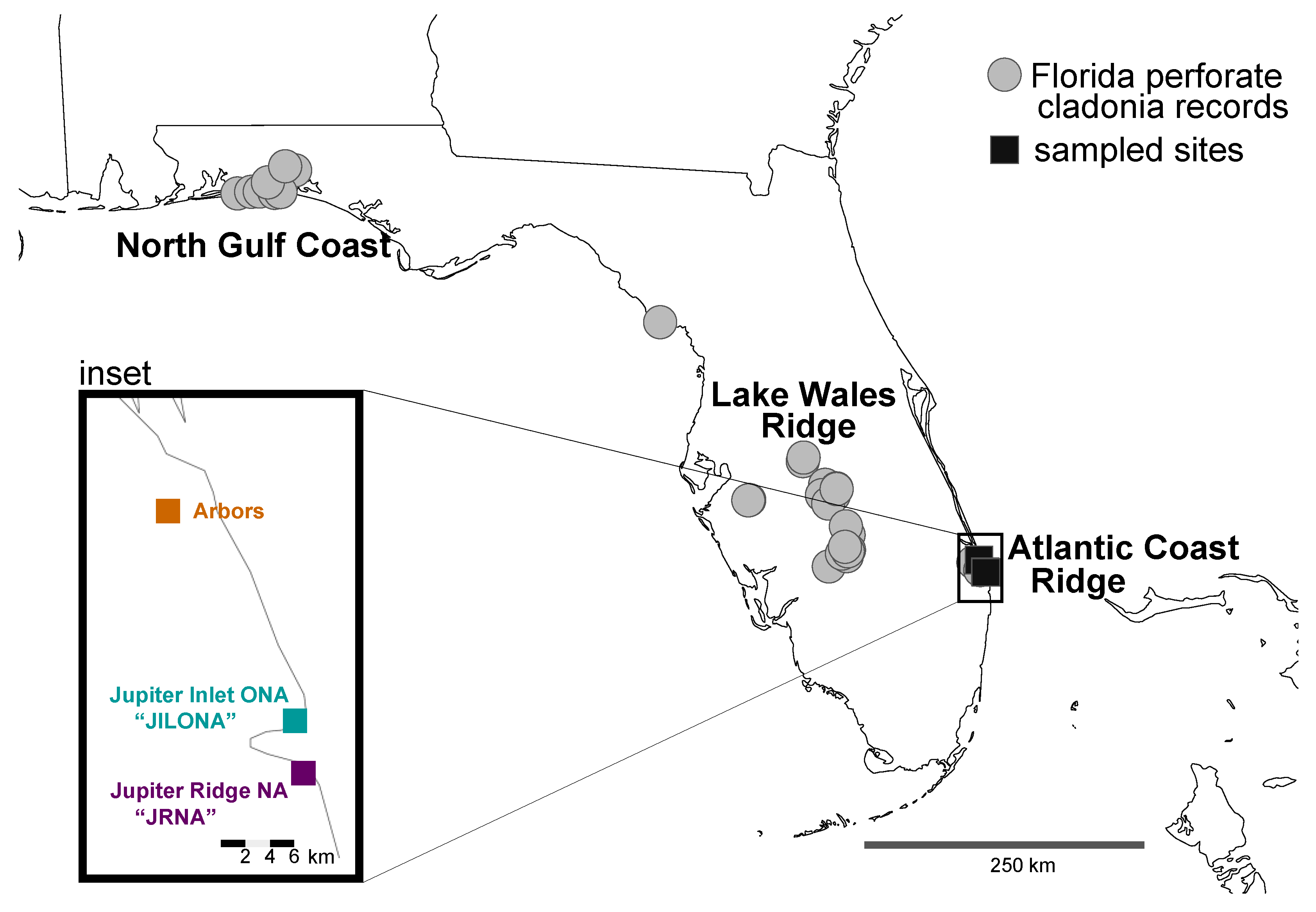
The Florida Perforate Cladonia (FPC; mycobiont = Cladonia perforata and photobiont = Asterochloris spp. [Chlorophyta, Trebouxiophyceae]) is a vagrant lichen occurring on white sandy soils in open areas in sand pine scrub, oak scrub and rosemary bald habitats in the Florida scrub [11,17]. FPC occurs in sixteen disjunct, isolated populations (at ca. 41 sites) from the panhandle area of Florida (North Gulf Coast) to the Lake Wales Ridge in central Florida to Martin and Palm Beach counties in South Florida (Atlantic Coast Ridge) [18] (Figure 1). This lichen likely has a naturally disjunct, patchy distribution, with ongoing local extirpation and recolonization events occurring throughout its long-term persistence in fire-maintained landscapes [11]. Whether these patch-level dynamics generated distinct, locally adapted variants is not known. Presently, habitat patches tend to be small, limited to a few square meters to several hectares, and these sites are exposed to disruptions by hurricanes and fires on very short time scales [11]. Due to significant loss of shrub habitat and limited and patchy occurrences [8,19], in 1993, the FPC was the first lichen to be federally listed as endangered in the U.S.A. and was one of the first lichens to be added to the IUCN Red List [20]. While some populations are relatively large and robust, others are in decline or under threat [21]. The main threats include habitat loss in high-value real estate areas, improper fire management strategies, and impacts from hurricanes.
FPC has a life history suggesting that this lichen reproduces exclusively via thallus fragmentation and clonal growth [21]. Spore-producing structures—apothecia—have not been observed in FPC, and the dispersal of fragments from source populations into open sand gaps in Florida scrub, including burned areas, is likely the primary method of short-term population recovery [11]. Populations grow via vegetative propagation, or clonal growth, in which parental genotypes or thalli (genets) produce individuals (ramets) that are capable of independent growth and dispersal [22]. However, recolonization from disjunct sites is unlikely and/or infrequent, as source populations are often kilometers away. Despite the apparently strict clonal propagation in FPC, genetic variation was found to be highly structured across populations spanning their entire distribution. Based on five microsatellite loci, Yahr [21] found low genetic diversity within geographically restricted sites but unique variations in the three major ridge systems (Figure 1).
Previous translocation success of FPC [18], in addition to successful experimental translocations of the common but similar and co-occurring Jester lichen (Cladonia leporina) and Powder-puff lichen (Cladonia evansii), highlight one avenue to offset potential losses from catastrophic events and reintroduce populations. However, genetic diversity within and among FPC populations is not well known, raising questions about how these populations are related and how to protect genetic diversity and potential locally adapted genets. This lack of information limits recovery efforts for this species, e.g., can individuals from healthy, robust populations be transplanted to help restore declining/extirpated populations?
‘Big data’ is critical for accelerating lichen conservation by informing the monitoring of rare and endangered species [23]. For threatened and endangered species, reference genomes provide critical insight into genomic diversity and architecture, local adaptations and genetic rescue, inbreeding and deleterious mutations, outbreeding and introgression, phylogenomics, and the structure and function of communities [24]. Here, we provide a genome-level perspective into the mycobiont (the fungal partner) of the FPC, Cladonia perforata. In this study, we aimed to characterize genetic diversity at three distinct sites in southeastern Florida using whole genome sequencing. Are individual thalli in these populations genetically homogenous, or does variation occur within and among the sites? Using high-throughput sequencing, we (i) provide the first draft genomes for C. perforata, including both the nuclear and mitochondrial genomes, (ii) assess genetic diversity within and among populations at the level of the genome (nuclear and mitochondrial), and (iii) investigate diversity across the entire nuclear ribosomal cistron, which includes the standard DNA barcoding marker for fungi. Our study provides the first genome-scale insight into this imperiled lichen.
2. Materials and Methods
2.1. Field Sampling
As part of ongoing research into the conservation of FPC in the Atlantic Coast Ridge in southeastern Florida, U.S.A., we were interested in genetic diversity occurring in sites that might be potentially impacted by transplanting to help restore declining/extirpated populations. Collections of Florida Perforate Cladonia (FPC) were made from three distinct sites in southeastern Florida on the Atlantic Coast Ridge in January 2022: (i) Jupiter Inlet Lighthouse Outstanding Natural Area (“JILONA”), (ii) Jupiter Ridge Natural Area (“JRNA”), and (iii) The Arbors Preserve (“Arbors”) (Figure 1 and Figure 2). These sites represent only the extreme southeastern populations of FPC, and populations in the North Gulf Coast and Lake Wales Ridge were not sampled for this study.
JILONA is part of the Bureau of Land Management’s National Conservation Lands system, covering 120 acres of sand pine scrub and oak scrub habitat in Jupiter, Palm Beach County (ca. 6 mASL). Associated species include sand selaginella (Selaginella arenicola), scrub frostweed (Crocanthemum nashii), gopher apple (Geobalanus oblongifolius), nodding pinweed (Lechea cernua), partridge pea (Chamaecrista fasciculata), scrub palmetto (Sabal etonia), and rosemary (Ceratiola ericoides). JRNA covers 271 acres of sand pine scrub and oak scrub habitat (10–18 mASL) in Juno Beach, Palm Beach County. JRNA is part of a state-owned Conservation and Recreation Lands project managed by Palm Beach County. Associated species include rosemary (Ceratiola ericoides), extensive Cladonia beds (Cladonia spp.), huckleberry (Vaccinium membranaceum), gopher apple (Geobalanus oblongifolius), nodding pinweed (Lechea cernua), saw palmetto (Serenoa repens). The final site, the Arbors, covers 72 acres of sand pine scrub and oak scrub habitat (10 mASL) on private land and is located behind the Arbors Subdivision off Ralph Fair Road in Hobe Sound, Martin County. Associated species include rosemary (Ceratiola ericoides), saw palmetto (Serenoa repens), gopher apple (Geobalanus oblongifolius), and huckleberry (Vaccinium membranaceum). The two southernmost sites, JILONA and JRNA, are separated by the Loxahatchee River and 4.3 km. The Arbors is ca. 20 km north of JILONA and JRNA.
To maximize the probability of sampling distinct genets, 10 FPC thalli were collected to maximize the spatial distance (tens to hundreds of meters) separating sampled thalli within each of the three natural area preserves. Each thallus was immediately placed into a bond paper lichen packet and allowed to air dry for several days before shipping. A GPS point was taken at the approximate midpoint within the preserve’s population, and photos were taken of the general area and FPC on site (Figure 2).
Letters of permission and authorization permits for collecting FPC at these sites include: permit #2996 from the Florida Dept. of Agriculture and Consumer Services Division of Plant Industry—for Permission to Harvest Endangered or Commercially Exploited Plant(s) or Plant Part(s). Within this permit is The Arbors authorization letter to collect FPC from their private preserve; permit #2997 from the Florida Dept. of Agriculture and Consumer Services Division of Plant Industry—for Permission to Harvest Endangered or Commercially Exploited Plant(s) or Plant Part(s). Within this permit is Palm Beach County’s authorization letter to collect FPC at the Jupiter Ridge Natural Area.
2.2. High-Throughput Sequencing and Genome Assembly
Limited genetic sequence data are currently available for FPC: two sequences representing the standard barcoding marker for fungi, the ITS (GenBank accession numbers AY753584 and AF457903), and a single sequence representing a fragment of the nuclear beta-tubulin gene (GenBank accession No. AF458570). To generate genome-scale resources for this study and future research, a single specimen, ‘Arbors7’ (Brigham Young University Herbarium of Non-Vascular Cryptogams, Provo, UT, USA), from the Arbors, was selected for long-read high throughput shotgun sequencing using Oxford Nanopore Technologies (ONT) MinION platform. For this specimen, DNA was extracted from three different portions of the same thallus using the E.Z.N.A. Plant DNA DS Mini Kit (Omega Bio-Tek, Inc., Norcross, GA, USA) following the manufacturer’s protocol. The ONT sequencing libraries were prepared using the ligation sequencing kit SQK-LSK114 with the Native Barcoding Kit 24 V14 (SQK-NBD114.24). The three barcoded libraries were sequenced on the MinION Mk1C system with an R10.4.1 flow cell (ONT).
Raw sequence FAST5 signals were called with Guppy v6.4.6 (ONT) using the dna_r10.3_450bps_hac.cfg model. The ONT sequencing generated 12.6 Gb from 3,735,355 reads, with an average length of 2407 bp. Nanopore reads were assembled using Flye v.2.9 [25]. The Flye assembly was further polished with the filtered Illumina reads (see below) following the Trycycler user manual using Polypolish v. 0.5.0 [26] and POLCA v. 4.0.5 [27].
The final nuclear genome sequence assembly was annotated using funannotate v1.8.7 (https://github.com/nextgenusfs/funannotate, accessed on 3 April 2023). Contigs were sorted by size, and simple repeats were identified, resulting in 1,263,743 (3.76%) masked nucleotides. Ab initio gene predication was conducted with annotations from Cladonia grayi Cgr/DA2myc/ss v2.0, Lobaria pulmonaria Scotland reference genome v1.0, Usnea florida ATCC 18,376 v1.0, and Xanthoria parietina 46-1-SA22 v1.1 and the Benchmarking Universal Single-Copy Orthologs (BUSCO) with the seed species set to Aspergillus fumigatus as training datasets. Biosynthetic gene clusters were annotated using antiSmash v6.0 for fungi, with all options enabled for the most thorough search possible [28].
To characterize diversity in FPC at multiple genetic scales, sequence fragments from the polished assembly were investigated to identify sequences representing (i) the lichen-forming fungus (LFF) nuclear genome, including BUSCO loci, (ii) the LFF nuclear ribosomal cistron, including the internal transcribed spacer region (ITS), and (iii) the LFF mitochondrial genome. To remove sequences from the assembly that did not represent the targeted LFF, we used a Diamond v0.9.32 blastx search against the NCBI nonredundant protein database to infer the taxonomic identity of each sequence [29]. We also compared the coverage of Illumina short reads (see below) mapped to the polished sequences to identify sequences with similar coverage and corroborate the taxonomic assignments. The LFF mitochondrial genome was identified from the assembly using BLAST comparisons [30] to known Cladonia spp. mitochondrial genomes. The mitochondrial genome was annotated using GeSeq as implemented in the CHLOROBOX platform using currently available NCBI RefSeq Lecanoromycetes mitochondrial genomes for BLAT reference sequences (https://chlorobox.mpimp-golm.mpg.de/geseq.html, accessed on 3 April 2023; [31]). LFF sequences from the polished ONT flye assembly were assessed to identify single-copy nuclear genes using the BUSCO v5.2.2 [32] and the “ascomycota_odb10” dataset for comparison.
As a byproduct of the assembly, we also identified the photobiont’s (Asterochloris sp.) chloroplast genome from the complete ONT flye assembly using BLAST comparisons [30] to known Trebouxiophyceae chloroplast genomes. The Asterochloris sp. chloroplast genome was also annotated using GeSeq implemented in the CHLOROBOX platform using currently available Trebouxiophyceae chloroplast genomes for BLAT reference sequences (https://chlorobox.mpimp-golm.mpg.de/geseq.html, accessed on 3 April 2023; [31]).
To assess genetic variation in the mycobiont partner (Cladonia perforata) of the FPC lichen, seven thalli from each area—JILONA, JRNA, and Arbors—were selected for Illumina shotgun sequencing. For these specimens, total genomic DNA was extracted using the E.Z.N.A. Plant DNA DS Mini Kit (Omega Bio-Tek, Inc., Norcross, GA, USA) and following the manufacturer’s protocol. We prepared total genomic DNA following the standard Illumina whole genome sequencing (WGS) library preparation process with Adaptive Focused Acoustics for shearing (Covaris), followed by an AMPure cleanup process. The DNA was then processed with the NEBNext Ultra™ II End Repair/dA-Tailing Module end-repair, together with the NEBNext Ultra™ II Ligation Module (New England Biolabs, Ipswich, MA, USA), while using standard Illumina index primers. Libraries were pooled and sequenced using the HiSeq 2500 sequencer in high output mode, by the DNA Sequencing Center at Brigham Young University, Provo, UT, USA, with 125 cycle paired-end (PE) reads.
2.3. Phylogenetic Datasets and Tree Inference
Three phylogenomic data matrices were created from the nuclear genome data: (i) one based on the 41 largest sequences (each fragment > 100 Kb and spanning over 32 Mb) in the polished Flye assembly, (ii) an alignment based on coding regions from the annotated nuclear genome (spanning ca. 13 Mb), and (iii) the final based on the 1250 complete, single-copy BUSCO markers. For the nuclear data matrices, we used RealPhy v1.12 [33] to align Illumina short reads from all samples to the targets implementing the following parameters: -readLength 100 -perBaseCov 5 -gapThreshold 0.5.
For the mitochondrial phylogenomic matrix, we constructed a multiple sequence alignment (MSA) from the complete mitochondrial genome. Illumina short reads from each specimen were mapped to the mitochondrial genome assembly (specimen ‘Arbors7’) using the Geneious Prime Read Mapper, with the “medium-low sensitivity/fast” settings, iterated five times [34]. The resulting consensus sequences were aligned using the program MAFFT v7 [35,36], implementing the G-INS-i alignment algorithm, and the remaining parameters were set to default values.
We also generated alignments of sequences representing the nuclear ribosomal cistron (nrDNA), which includes the standard barcoding marker for fungi, the ITS [37], the nuclear ribosomal small and large subunits, and the intergenic spacer region [38]. We identified the nrDNA cistron from the complete ONT Flye assembly using BLAST comparisons [30]. Illumina short reads were then mapped back to the reference nrDNA sequence using the Geneious Prime Read Mapper, with the “medium-low sensitivity/fast” settings, iterated five times [34]. Considering relative coverage, we selected the longest gap-free nrDNA region shared across all samples. The resulting consensus sequences were aligned using the program MAFFT v7 [35,36], implementing the G-INS-i alignment algorithm, and the remaining parameters were set to default values. Subsequently, the ITS region was extracted from the nrDNA alignment and re-aligned along with the two available ITS sequences from C. perforata on GenBank (accession numbers AY753584 and AF457903) using MAFFT as described above.
3. Results
3.1. Genomic Data
ONT long read and Illumina short read data are available under the NCBI BioProject ID PRJNA966217.
The initial ONT Flye assembly comprised 116 fragments spanning 34.4 Mb and had a mean coverage of 211. After excluding non-target sequences, the polished Cladonia perforata assembly (specimen ‘Arbors7’) comprised 72 fragments. It spanned 33.61 Mb with an N50 score of 1.1 Mb (describes the length of the shortest contig in the group of the longest contigs, which together represent 50% of the assembled genome) and an L50 of 12 (the smallest number of contigs that includes 50% of the total sequence assembly length) (Table 1; Supplementary File S1). The annotated nuclear alignment is available under the NCBI BioProject ID PRJNA982265.
BUSCO statistics suggest a relatively complete assembly, with 95.1% of the BUSCO markers recovered as a complete, single copy (1250 orthologs; Supplementary File S2), plus an additional 3% of the total BUSCO loci when including the fragmented, single copy regions. A total of 13,980 genes were predicted for the nuclear genome of C. perforata (Table 1). From the polished nuclear assembly, we extracted a 9.7 Kb fragment of the nuclear ribosomal cistron (GenBank Accession Nos. OQ890691-OQ890711) bounded by difficult-to-align/map intronic regions. The complete LFF mitochondrial genome was assembled, spanning 59,014 bp (GenBank accession No. OQ934048; Supplementary Files S3 and S4). We extracted and assembled ITS reads from the Asterochloris photobiont from the short read data, and the ITS was highly similar (99.5%) to a recently inferred, undescribed lineage ‘OUT11’ sensu [41] (results not shown). The assembled Asterochloris sp. (“OTU11” sensu [41]) chloroplast genome was identified by blast comparisons and spanned 218,874 bp (GenBank accession No. OQ934049; Supplementary Files S5 and S6).
3.2. Phylogenomic Inferences
A summary of phylogenomic data matrices is provided in Table 2.
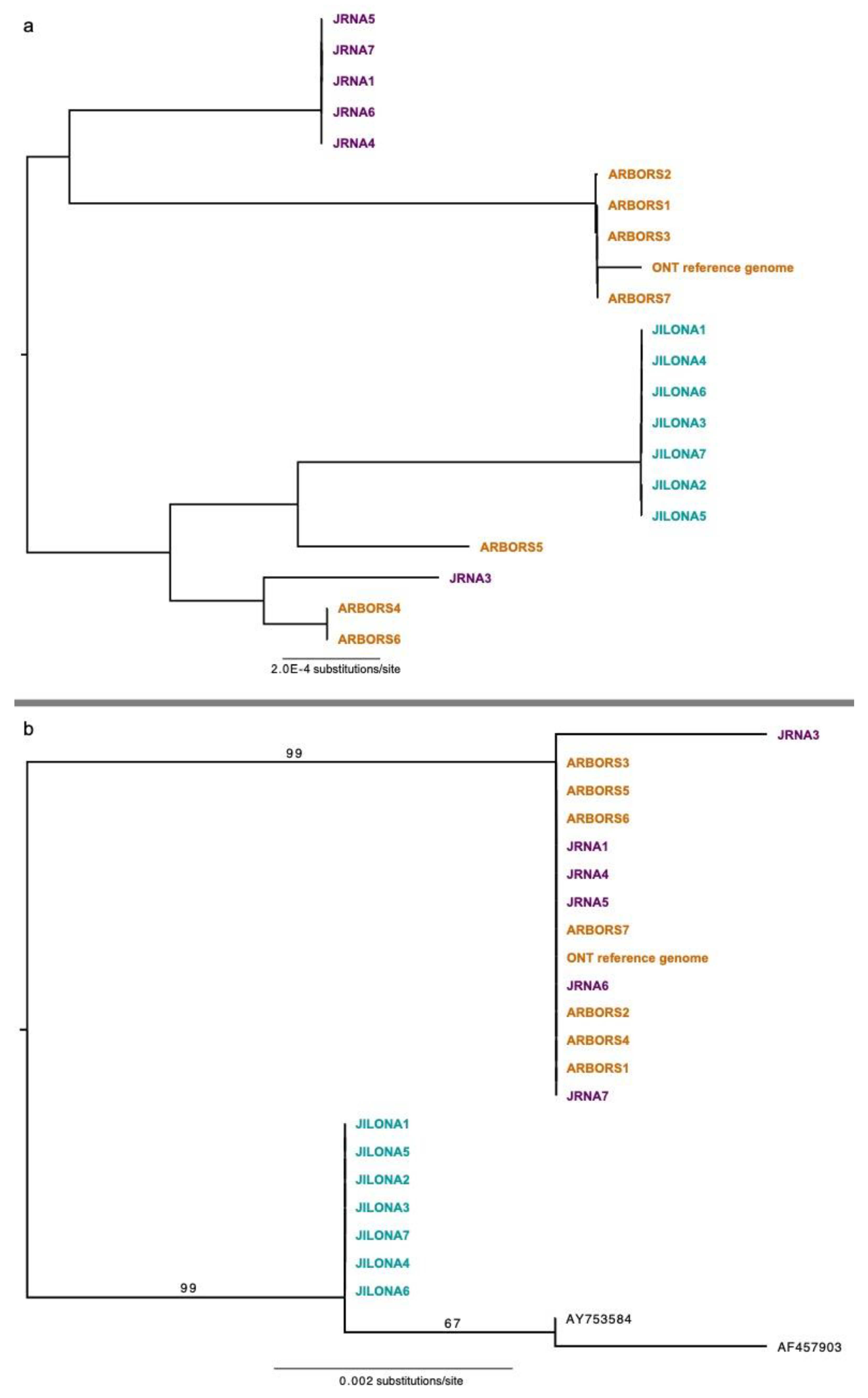
For the nuclear genome, coverage of Illumina short read data ranged from 4× to 15× per sample (Table 3). The phylogenomic data matrix based on the largest fragments (>100 Kb) comprised 32.1 Mb aligned nucleotide position characters (Supplementary File S7). In the nuclear REALPHY alignment, specimens Arbors5 and JILONA7 had 53.69% and 68.86% missing data, respectively, due to low coverage. All remaining specimens had less than 20% missing data, with an average of 6.82% missing data. The resulting ML topology revealed six distinct, closely related genets, with no genets shared among the sampling sites (Figure 3a), e.g., all sampled sites harbored unique genets. Phylogenies inferred from the alignment of the annotated coding regions (12.1 Mb) and the alignment of the concatenated 1250 BUSCO markers (1.9 Mb) recovered the same six genets, although relationships differed among the topologies inferred form different nuclear data matrices (Supplementary File S7).
Three genets were sampled in the Arbors (the smallest protected area sampled), two genets were sampled in JRNA (the largest protected area sampled), and a single genet was sampled in JILONA. Two of the six genets—one from the Arbors and the second from JRNA—were represented by a single sampled thallus.
The mitochondrial genomes of all sampled thalli were 100% identical. Coverage of the mitochondrial genome with Illumina short read data ranged from 61× to 287× per sample (Table 2). Phylogenies inferred from portions of the nuclear ribosomal cistron failed to recover the same six genets inferred from the nuclear genomic data matrices but did reveal two distinct genetic groups. The topology inferred from the ITS data revealed two well-supported clades: one comprising all specimens collected from JRNA and the Arbors, and the second contained the specimens collected from JILONA, plus the only two presently available sequences from GenBank (Figure 3b). ITS sequences generated from specimens collected at JILONA were 100% identical but differed from those on GenBank (Lake Wale Ridge, central Florida [Figure 1]) at a few nucleotide position characters. The topology inferred from a larger fragment of the nuclear ribosomal cistron also recovered all specimens collected from JRNA and the Arbors in a well-supported clade and specimens collected from JILONA in a separate, distinct clade. Average pairwise similarity across a 9.8 Kb fragment of the cistron and the ITS were 99.7% and 99.5%, respectively (Table 2).
4. Discussion
Thirty years after the initial designation of the Florida Perforate Cladonia lichen (FPC) as an endangered species, we provide critical genomic resources to aid in conservation efforts and understanding the biology of this imperiled lichen [23,24,42]. The draft nuclear genome generated by ONT long read data, and polished using Illumina data, spans over 33 Mb and is estimated to be over 95% complete (Table 1). The total genome size assembled here is slightly smaller than previously published Cladonia genomes, e.g., 33.6 Mb for C. perforata vs. 34.6 to 37.1 Mb in other Cladonia species [43,44]. While recent lichen-forming fungal genomes generated using ONT long read data have resulted in nearly complete, telomere-to-telomere chromosomal assemblies [43], our nuclear genome assembly of C. perforata was somewhat more fragmented (Table 1), while still within the range of high-quality draft genomes—see summary in [43]. The complete, circular mitochondrial genome for C. perforata was 59 Kb (Table 3), falling within the upper size range of other sequenced Cladonia spp. mitochondrial genomes, 45,312–60,062 bp [45]. Finally, we provide the first chloroplast genome, to our knowledge, for the photobiont genus associated with FPC, Asterochloris sp.
Using a genome-wide data matrix spanning over 32 Mb, we inferred the presence of multiple distinct mycobiont genets occurring at local scales in southeastern Florida (Figure 3a). Strikingly, no genets were shared among even the closest sample sites (separated by less than 5 km; Figure 1), with each sampled protected area harboring unique FPC genets (Figure 3a). The same six genets were inferred using a subset of the nuclear genome—1250 BUSCO loci, spanning 1.9 Mb. Our results are partly consistent with previous inferences from microsatellite data from C. perforata, revealing highly structured genetic variation among populations [21]. Yahr [21] found that C. perforata populations in the Atlantic Coast Ridge (area sampled for this study) were genetically more similar to each other than those occurring in the North Gulf Coast and the Lake Wales Ridge regions (Figure 1). While previous work documented low within-site variation, we found multiple distinct genets occurring in two of the three sites sampled (Table 3). This suggests that the range of genetic variation in FPC throughout its entire distribution is likely much higher than that sampled here. Highly genetically structured, isolated FPC populations would be consistent with the assumption that these are a part of unique Florida scrub communities that have likely remained intact since the formation of the dunes during the Pleistocene [1,2,3]. Interestingly, earlier works suggest that isolation by distance cannot explain the diversity within and among C. perforata populations across its distribution [21], and other factors potentially influencing dispersal merit additional attention. Restriction site-associated DNA sequencing data and other target enrichment approaches [46,47,48,49,50] will be essential for determining evolutionary relationships and diversity, landscape genetics, and the range of clonal ramets across the entire distribution of FPC.
Data from the nuclear ribosomal cistron (ca. 10 Kb) of the mycobiont nuclear genome, including the standard fungal DNA barcoding marker (the internal transcribed spacer region) [37], also provides evidence of genetic variation in the sampled specimens, recovering two distinct groups, rather than six inferred from nuclear genome-scale data (Figure 3). The ITS marker was insufficient to distinguish at least some genets, e.g., five putative genets from the “Arbors” and Jupiter Ridge Natural Area (JRNA) were indistinguishable using ITS data, while the single genet occurring in Jupiter Inlet Outstanding Natural Area (JILONA) was distinct from all the others based on a multiple sequence alignment of the ITS (Figure 3b). Additional genetic variation was found in the ITS region when including the two presently available ITS sequences from GenBank (Figure 3b) [21]. These results suggest that the standard fungal DNA barcode (ITS) may be a reasonable “first pass” marker for screening genetic diversity in the FPC mycobiont, given the relatively inexpensive cost compared to genome-scale sequencing. Specimen sampling for the present study was restricted to a narrow portion of the overall patchy distribution of FPC (Figure 1). We predict that with broader sampling spanning distinct populations throughout the entire distribution, additional mycobiont ITS haplotypes would be identified in FPC. This is supported by the differences observed among the ITS sequences inferred from our samples in southeastern Florida and the two sequences from GenBank from specimens collected in the Lake Wales Ridge region. However, nuclear genome-scale data is required to identify genetic variation on the scale of genets (Figure 3).
In contrast to the data from the nuclear genome, the entire mitochondrial genome was identical in all sampled specimens. In contrast to many animal lineages where mitochondrial genomes have high resolving power for population genetics and phylogeography [51], mitochondrial genomes may have more limited utility for identifying population structure in FPC [47]. Intraspecific variation in mitochondrial genomes has been observed in two widespread species in Cladonia [52]. Extended FPC sampling beyond the Atlantic Coast Ridge (Figure 1) will be critical to determining the full range of mitochondrial variation in this lichen.
FPC is a photobiont specialist, associating with multiple geographically structured and closely related Asterochloris genotypes [17]. These photobiont lineages appear to have limited geographic distributions, and restricted availability of compatible Asterochloris photobionts may further limit the successful establishment of FPC [17]. More recently, the geographical distribution of the different Asterochloris lineages associating with Cladonia species suggests that mycobiont identity and climate are the main predictors of patterns of Asterochloris genetic variation [41]. Conservation efforts to restore declining/extirpated populations should consider the availability of suitable Asterochloris partners.
An increasing number of published landscape genetic and genomic studies of LFF show that the genetic diversity of populations and rates of gene flow vary substantially between rare and common LFF. In the 21 individuals of FPC sampled from three sites, we found substantial genetic diversity within and between sites. Yahr [21] sampled throughout the distribution of FPC and inferred the population structure using five microsatellite markers. She found that all the populations were genetically distinct from each other, suggesting low rates of gene flow among sites. A population genomics study of the only other federally endangered lichen, Cetradonia linearis, similarly found evidence for strong isolation by distance and for high genetic diversity within populations [53]. These findings contrast strongly with studies of widespread LFF. Cladonia stellaris, for instance, was found to lack spatial genetic structure across a very large geographic area in Quebec, Canada [54], and Alors et al. [55] recovered a similar pattern in Parmelina carporrhizans. In a circum-Antarctic study of Pseudocyphellaria glabra, Widhelm et al. [46] found evidence for clades that largely correspond to continental-level regions with few instances of intercontinental gene flow, thus showing some spatial structure, though on a very large spatial scale. In Lobaria pulmonaria, a species with a circumboreal distribution that is rare and patchily distributed in some parts of its range, there is evidence for high levels of isolation by distance in some parts of its range [56] and evidence for gene pools with very large distributions in other parts of its range [57], which is perhaps driven by differing landscape-level processes occurring among sampled regions. The hypothesized relationship between rarity and migration rates in LFFs remains to be tested in a rigorous framework. If a similar pattern is found across many rare species, it would suggest that assisted migration may be an effective conservation action for rare LFF.
In the pyrogenic Florida scrub ecosystem, gap specialists, such as FPC, are sensitive to both alterations to fire regimes and microhabitat availability [58,59]. With increasingly fragmented habitat in the Florida scrub, the impact of disturbances on isolated FPC populations becomes increasingly problematic. In these fire-dependent communities, the dispersal of FPC from undisturbed habitats into recently burned areas is likely the primary method of short-term population recovery [11]. Post-fire and post-hurricane research and reintroduction observations suggest that FPC is not only limited by dispersal but also that populations may also be slow to increase in abundance [11].
From a conservation perspective, fine-scale genetic resolution is needed to resolve questions of dispersal and genetic diversity. Our results suggest that even at local scales, i.e., sites in close geographic proximity, such as those in the Atlantic Coast Ridge sampled here, transplants will likely introduce novel genets (Figure 3a). Whether distinct genets represent local adaptations remains unknown. Recent translocation studies resulted in significant population size increases at Jupiter Inlet Lighthouse Outstanding Natural Area [18], suggesting the likely novel genets transplanted to distinct sites were able to persist and grow. Considering dependency on fire for maintaining suitable habitat for FPC, infrequent dispersal events are likely fundamental to the persistence of FPC, although genetic variation cannot be accounted for strictly through isolation by distance [11,21].
Given the imperiled state of many FPC populations, we predict that the benefit of translocations from robust populations to restore disturbed populations outweighs the risk of introducing novel genetic variation. Pending additional genetic screening, restricting translocations to populations occurring within the same general region is likely advisable, e.g., populations within the North Gulf Coast, Lake Wales Ridge, or Atlantic Coast Ridge, respectively.
The genomic resources generated here—the nuclear and mitochondrial genomes of the mycobiont C. perforata and the photobiont’s chloroplast genome—provide critical resources for genome-informed conservation efforts for the FPC.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9070698/s1, File S1: Draft genome assembly for Cladonia perforata (mycobiont of the Florida Perforate Cladonia’ specimen ‘Arbors7’ [Herbarium BRY-C]), comprising 72 fragments and spanning 33.61 Mb, with an N50 score of 1.1 Mb (describes the length of the shortest contig in the group of the longest contigs which together represent 50% of the assembled genome) and an L50 of 12 (the smallest number of contigs that includes 50% of the total sequence assembly length); File S2: the 1250 complete, single-copy BUSCO nuclear markers (95.1% complete) extracted from the draft Cladonia perforata assembly (specimen ‘Arbors7’ [Herbarium BRY-C]); File S3: draft Cladonia perforata mitochondrial genome (specimen ‘Arbors7’ [Herbarium BRY-C]); File S4: Circular gene map of the mitochondrial genome of Cladonia perforata (mycobiont of the Florida Perforate Cladonia’, specimen ‘Arbors7’ [Herbarium BRY-C]). Genes drawn inside the circle are transcribed clockwise, and those outside the circle are transcribed counterclockwise. The darker grey in the inner circle represents the GC content; conversely, the lighter one represents the AT content. Gene functions or gene identifiers are displayed using colors indicated by the inner legend. The symbol ‘*’ indicates genes with introns; File S5: draft Asterochloris species chloroplast genome (photobiont of the Florida Perforate Cladonia’ specimen ‘Arbors7’ [Herbarium BRY-C]); File S6: Circular gene map of the chloroplast genome of Cladonia perforata (photobiont of the Florida Perforate Cladonia’, specimen ‘Arbors7’ [Herbarium BRY-C]). Genes drawn inside the circle are transcribed clockwise, and those outside the circle are transcribed counterclockwise. The darker grey in the inner circle represents the GC content; conversely, the lighter one represents the AT content. Gene functions or gene identifiers are displayed using colors indicated by the inner legend. The symbol ‘*’ indicates genes with introns; File S7: Multiple sequence alignments were used in this study. Nuclear phylogenomic data matrix based on all sequences larger than 100 Kb in the draft ONT Flye assembly—“cladonia_perforata_NUC_32mb_realphy_alignment.phy”; nuclear phylogenomic data matrix based on annotated coding regions from the draft ONT Flye assembly—“cladonia_perforata_NUC_cds_transcripts_realphy_alignment.phy”; matrix based on a fragment of the Cladonia perforata nuclear ribosomal cistron—“cladonia_perforata_NUC_cistron.fas”; matrix based on the ITS regions of the Cladonia perforata nuclear ribosomal cistron “cladonia_perforata_NUC_ITS.fas”; mitochondrial phylogenomic data matrix based on complete mitochondrial genomes—“cladonia_perforata_MT_alignment.fas”.
Author Contributions
Conceptualization, S.D.L. and A.D.; methodology, S.D.L., A.D., E.M. and J.L.A.; validation, S.D.L. and J.L.A.; formal analysis, S.D.L., E.M. and J.L.A.; investigation, S.D.L., A.D., E.M. and J.L.A.; resources, S.D.L. and A.D.; data curation, S.D.L.; writing—original draft preparation, S.D.L. and A.D.; writing—review and editing, S.D.L., A.D., E.M. and J.L.A.; visualization, S.D.L.; supervision, S.D.L.; project administration, S.D.L.; funding acquisition, S.D.L. and A.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Bureau of Land Management.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Oxford Nanopore Technologies long read and Illumina short read data are available under the NCBI BioProject ID PRJNA966217; the Cladonia perforata nuclear genome is available on GenBank under the NCBI BioProject ID PRJNA982265.; the Cladonia perforata mitochondrial genome is available on GenBank as accession No OQ934048; the chloroplast genome from the photobiont associated with Cladonia perforata specimen ‘Arbors 7’, an Asterochloris sp., is available on GenBank as accession No OQ934049; and all data matrices are available as supplemental files included in the published version of the manuscript.
Acknowledgments
We express sincere appreciation to Rebecca Yahr (Royal Botanic Garden Edinburgh) for providing invaluable insight based on her transformative research into the Florida Perforate Cladonia. We thank John Woodhouse for help with long-read sequencing and Ed Wilcox (DNA Sequencing Center, BYU) for help with the Illumina short-read sequencing. JLA was supported by NSF DEB #2115191.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Estill, J.C.; Cruzan, M.B. Phytogeography of rare plant species endemic to the Southeastern United States. Castanea 2001, 66, 3–23. [Google Scholar]
- Myers, R.L. Scrub and high pine. In Ecosystems of Florida; University of Central Florida Press: Orlando, FL, USA, 1990; Volume 765, pp. 150–193. [Google Scholar]
- Laessle, A.M. Relationship of sand pine scrub to former shore lines. Q. J. Fla. Acad. Sci. 1967, 30, 269–286. [Google Scholar]
- McCoy, E.D.; Britt, E.J.; Catenazzi, A.; Mushinsky, H.R. Fire and herpetofaunal diversity in the Florida scrub ecosystem. Nat. Areas J. 2013, 33, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Christman, S.P.; Judd, W.S. Notes on plants endemic to Florida scrub. Fla. Sci. 1990, 53, 52–73. [Google Scholar]
- Germain-Aubrey, C.C.; Soltis, P.S.; Neubig, K.M.; Thurston, T.; Soltis, D.E.; Gitzendanner, M.A. Using comparative biogeography to retrace the origins of an ecosystem: The case of four plants endemic to the Central Florida scrub. Int. J. Plant Sci. 2014, 175, 418–431. [Google Scholar] [CrossRef]
- Deyrup, M. Arthropods endemic to Florida scrub. Fla. Sci. 1989, 52, 254–270. [Google Scholar]
- Wilhelm, G.S.; Burkhalter, J.R. Cladonia Perforata, the Northwest Florida population. Bryologist 1990, 93, 66–68. [Google Scholar] [CrossRef]
- Noss, R.F.; Platt, W.J.; Sorrie, B.A.; Weakley, A.S.; Means, D.B.; Costanza, J.; Peet, R.K. How global biodiversity hotspots may go unrecognized: Lessons from the North American Coastal Plain. Divers. Distrib. 2015, 21, 236–244. [Google Scholar] [CrossRef]
- Menges, E.S.; Main, K.N.; Pickert, R.L.; Ewing, K. Evaluating a fire management plan for fire regime goals in a Florida landscape. Nat. Areas J. 2017, 37, 212–227. [Google Scholar] [CrossRef] [Green Version]
- Yahr, R. Ecology and post-fire recovery of Cladonia perforata, an endangered Florida-scrub lichen. For. Snow Landsc. Res. 2000, 75, 339–356. [Google Scholar]
- Lindsay, M.N.; Lewis, D.B.; Halstead, N.; Gainsbury, A.M. Fire severity effects on the herpetofaunal diversity of the Florida scrub, a biodiversity hotspot. Biodivers. Conserv. 2023, 32, 1857–1878. [Google Scholar] [CrossRef]
- McCoy, E.D.; Mushinsky, H.R. Effects of fragmentation on the richness of vertebrates in the Florida scrub habitat. Ecology 1994, 75, 446–457. [Google Scholar] [CrossRef]
- Li, J.; Powel, T.L.; Seiler, T.J.; Johnson, D.P.; Anderson, H.P.; Bracho, R.; Hungate, B.A.; Hinkle, C.R.; Drake, B.G. Impacts of urricane Frances on Florida scrub-oak ecosystem processes: Defoliation, net CO2 exchange and interactions with elevated CO2. Glob. Chang. Biol. 2007, 13, 1101–1113. [Google Scholar] [CrossRef]
- Menges, E.S.; Kohfeldt, N. Life history strategies of Florida scrub plants in relation to rire. Bull. Torrey Bot. Club 1995, 122, 282–297. [Google Scholar] [CrossRef]
- Rosentreter, R.; DeBolt, A. Lichens as an indicator of sea-level rise. Evansia 2023, 39, 162–168. [Google Scholar] [CrossRef]
- Yahr, R.; Vilgalys, R.; Depriest, P.T. Strong fungal specificity and selectivity for algal symbionts in Florida scrub Cladonia lichens. Mol. Ecol. 2004, 13, 3367–3378. [Google Scholar] [CrossRef]
- DeBolt, A. Transplant Success of Cladonia perforata (Florida Perforate Cladonia) at the Jupiter Inlet Lighthouse Outstanding Natural Area. Evansia 2021, 38, 32–42. [Google Scholar] [CrossRef]
- Buckley, A.; Hendrickson, T.O. The distribution of Cladonia perforata Evans on the southern Lake Wales Ridge in Highlands County, Florida. Bryologist 1988, 91, 354–356. [Google Scholar] [CrossRef]
- Yahr, R.L.S.G. Cladonia perforata. The IUCN Red List of Threatened Species 2003: E.T43994A10838980. Available online: file:///C:/Users/MDPI/Downloads/10.2305_IUCN.UK.2003.RLTS.T43994A10838980.en-1.pdf (accessed on 28 April 2023). [CrossRef]
- Yahr, R. Evolutionary and Ecological Pattern and Process in Cladonia Lichen Symbioses: A Population Genetic Approach. Ph.D. Thesis, Duke University, Durham, NC, USA, 2004. [Google Scholar]
- Barrett, S.C.H. Influences of clonality on plant sexual reproduction. Proc. Natl. Acad. Sci. USA 2015, 112, 8859–8866. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.L.; McMullin, R.T.; Tripp, E.A.; Lendemer, J.C. Lichen conservation in North America: A review of current practices and research in Canada and the United States. Biodivers. Conserv. 2019, 28, 3103–3138. [Google Scholar] [CrossRef]
- Formenti, G.; Theissinger, K.; Fernandes, C.; Bista, I.; Bombarely, A.; Bleidorn, C.; Ciofi, C.; Crottini, A.; Godoy, J.A.; Höglund, J.; et al. The era of reference genomes in conservation genomics. Trends Ecol. Evol. 2022, 37, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Holt, K.E. Polypolish: Short-read polishing of long-read bacterial genome assemblies. PLoS Comput. Biol. 2022, 18, e1009802. [Google Scholar] [CrossRef] [PubMed]
- Zimin, A.V.; Salzberg, S.L. The genome polishing tool POLCA makes fast and accurate corrections in genome assemblies. PLoS Comput. Biol. 2020, 16, e1007981. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Buchfink, B.; Reuter, K.; Drost, H.-G. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat. Methods 2021, 18, 366–368. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [Green Version]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Bertels, F.; Silander, O.K.; Pachkov, M.; Rainey, P.B.; van Nimwegen, E. Automated reconstruction of whole-genome phylogenies from short-sequence reads. Mol. Biol. Evol. 2014, 31, 1077–1088. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Rozewicki, J.; Yamada, K.D.; Katoh, K. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Consortium, F.B. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Naidoo, K.; Steenkamp, E.T.; Coetzee, M.P.A.; Wingfield, M.J.; Wingfield, B.D. Concerted evolution in the ribosomal RNA cistron. PLoS ONE 2013, 8, e59355. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree Version 1.4. 2008. Available online: http://tree.bio.ed.ac.uk/software/Wgtree/ (accessed on 9 January 2023).
- Pino-Bodas, R.; Stenroos, S. Global biodiversity patterns of the photobionts associated with the genus Cladonia (Lecanorales, Ascomycota). Microb. Ecol. 2021, 82, 173–187. [Google Scholar] [CrossRef]
- Blaxter, M.; Archibald, J.M.; Childers, A.K.; Coddington, J.A.; Crandall, K.A.; Di Palma, F.; Durbin, R.; Edwards, S.V.; Graves, J.A.M.; Hackett, K.J.; et al. Why sequence all eukaryotes? Proc. Natl. Acad. Sci. USA 2022, 119, e2115636118. [Google Scholar] [CrossRef]
- Pfeffer, B.; Lymbery, C.; Booth, B.; Allen, J.L. Chromosomal genome sequence assembly and mating-type (MAT) locus characterization of the leprose asexual lichenized fungus Lepraria neglecta (Nyl.) Erichsen. Lichenologist 2023, 55, 41–50. [Google Scholar] [CrossRef]
- Park, S.-Y.; Choi, J.; Lee, G.-W.; Kim, J.A.; Oh, S.-O.; Jeong, M.-H.; Yu, N.-H.; Kim, S.; Lee, Y.-H.; Hur, J.-S. Draft genome sequence of lichen-forming fungus Cladonia metacorallifera Strain KoLRI002260. Genome Announc. 2014, 2, e01065-13. [Google Scholar] [CrossRef] [Green Version]
- Brigham, L.M.; Allende, L.M.; Shipley, B.R.; Boyd, K.C.; Higgins, T.J.; Kelly, N.; Anderson Stewart, C.R.; Keepers, K.G.; Pogoda, C.S.; Lendemer, J.C.; et al. Genomic insights into the mitochondria of 11 eastern North American species of Cladonia. Mitochondrial DNA Part B 2018, 3, 508–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widhelm, T.J.; Grewe, F.; Huang, J.-P.; Ramanauskas, K.; Mason-Gamer, R.; Lumbsch, H.T. Using RADseq to understand the circum-Antarctic distribution of a lichenized fungus, Pseudocyphellaria glabra. J. Biogeogr. 2021, 48, 78–90. [Google Scholar] [CrossRef]
- Lücking, R.; Leavitt, S.D.; Hawksworth, D.L. Species in lichen-forming fungi: Balancing between conceptual and practical considerations, and between phenotype and phylogenomics. Fungal Divers. 2021, 109, 99–154. [Google Scholar] [CrossRef]
- Grewe, F.; Lagostina, E.; Wu, H.; Printzen, C.; Lumbsch, H.T. Population genomic analyses of RAD sequences resolves the phylogenetic relationship of the lichen-forming fungal species Usnea antarctica and Usnea aurantiacoatra. MycoKeys 2018, 43, 91–113. [Google Scholar] [CrossRef] [Green Version]
- Hale, H.; Gardner, E.M.; Viruel, J.; Pokorny, L.; Johnson, M.G. Strategies for reducing per-sample costs in target capture sequencing for phylogenomics and population genomics in plants. Appl. Plant Sci. 2020, 8, e11337. [Google Scholar] [CrossRef] [Green Version]
- Widhelm, T.J.; Grewe, F.; Huang, J.-P.; Mercado-Díaz, J.A.; Goffinet, B.; Lücking, R.; Moncada, B.; Mason-Gamer, R.; Lumbsch, H.T. Multiple historical processes obscure phylogenetic relationships in a taxonomically difficult group (Lobariaceae, Ascomycota). Sci. Rep. 2019, 9, 8968. [Google Scholar] [CrossRef] [Green Version]
- DeSalle, R.; Goldstein, P. Review and interpretation of trends in DNA barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, J.R.; Karol, K.G.; Ohmura, Y.; Pogoda, C.S.; Keepers, K.G.; McMullin, R.T.; Lendemer, J.C. Mitochondrial genomes in the iconic reindeer lichens: Architecture, variation, and synteny across multiple evolutionary scales. Mycologia 2023, 115, 187–205. [Google Scholar] [CrossRef]
- Allen, J.L.; McKenzie, S.K.; Sleith, R.S.; Alter, S.E. First genome-wide analysis of the endangered, endemic lichen Cetradonia linearis reveals isolation by distance and strong population structure. Am. J. Bot. 2018, 105, 1556–1567. [Google Scholar] [CrossRef]
- Alonso-García, M.; Grewe, F.; Payette, S.; Villarreal, J.C.A. Population genomics of a reindeer lichen species from North American lichen woodlands. Am. J. Bot. 2021, 108, 159–171. [Google Scholar] [CrossRef]
- Alors, D.; Grande, F.D.; Cubas, P.; Crespo, A.; Schmitt, I.; Molina, M.C.; Divakar, P.K. Panmixia and dispersal from the Mediterranean Basin to Macaronesian Islands of a macrolichen species. Sci. Rep. 2017, 7, 40879. [Google Scholar] [CrossRef]
- Werth, S.; Gugerli, F.; Holderegger, R.; Wagner, H.H.; Csencsics, D.; Scheidegger, C. Landscape-level gene flow in Lobaria pulmonaria, an epiphytic lichen. Mol. Ecol. 2007, 16, 2807–2815. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.L.; McMullin, R.T.; Wiersma, Y.F.; Scheidegger, C. Population genetics and biogeography of the lungwort lichen in North America support distinct Eastern and Western gene pools. Am. J. Bot. 2021, 108, 2416–2424. [Google Scholar] [CrossRef]
- Menges, E.S.; Hawkes, C.V. Interactive effects of fire and microhabitat on plants of Florida scrub. Ecol. Appl. 1998, 8, 935–946. [Google Scholar] [CrossRef]
- Quintana-Ascencio, P.F.; Morales-Hernández, M. Fire-mediated effects of shrubs, lichens and herbs on the demography of Hypericum cumulicola in patchy Florida scrub. Oecologia 1997, 112, 263–271. [Google Scholar] [CrossRef]
Figure 1.
Distribution of the Florida Perforate Cladonia (FPC) in Florida, USA, based on herbarium records available on the Consortium of Lichen Herbaria (https://lichenportal.org/portal/; accessed on 24 April 2023). Grey circles indicate historic FPC records, and the black squares in southeastern Florida on the Atlantic Coast Ridge indicate the three sites sampled for this study—shown in the inset.
Figure 1.
Distribution of the Florida Perforate Cladonia (FPC) in Florida, USA, based on herbarium records available on the Consortium of Lichen Herbaria (https://lichenportal.org/portal/; accessed on 24 April 2023). Grey circles indicate historic FPC records, and the black squares in southeastern Florida on the Atlantic Coast Ridge indicate the three sites sampled for this study—shown in the inset.

Figure 2.
Photographs of habitat and Florida Perforate Cladonia (FPC) at the three sites sampled in southeastern Florida in 2021. (a–c): Jupiter Inlet Lighthouse Outstanding Natural Area (“JILONA”); (d–f): Jupiter Ridge Natural Area (“JRNA”); and (g–i): The Arbors Preserve (“Arbors”). Panels on the left are images of FPC habit, panels in the center depict typical habitat at the sample site, and panels on the right depict typical Cladonia beds. Photo credit: A. DeBolt, except panel ‘b’ from R. Rosentreter.
Figure 2.
Photographs of habitat and Florida Perforate Cladonia (FPC) at the three sites sampled in southeastern Florida in 2021. (a–c): Jupiter Inlet Lighthouse Outstanding Natural Area (“JILONA”); (d–f): Jupiter Ridge Natural Area (“JRNA”); and (g–i): The Arbors Preserve (“Arbors”). Panels on the left are images of FPC habit, panels in the center depict typical habitat at the sample site, and panels on the right depict typical Cladonia beds. Photo credit: A. DeBolt, except panel ‘b’ from R. Rosentreter.

Figure 3.
Phylogenetic inferences based on a 32.1 Mb nuclear phylogenomic matrix (panel (a)) and an alignment of the standard fungal DNA barcode, the ITS (panel (b)). Samples from the three distinct sites are color-coded: Jupiter Inlet Lighthouse Outstanding Natural Area (“JILONA”); Jupiter Ridge Natural Area (“JRNA”); and the Arbors. All relationships in panel (a) were recovered with 100% bootstrap support, and bootstrap values in panel (b) are shown above branches.
Figure 3.
Phylogenetic inferences based on a 32.1 Mb nuclear phylogenomic matrix (panel (a)) and an alignment of the standard fungal DNA barcode, the ITS (panel (b)). Samples from the three distinct sites are color-coded: Jupiter Inlet Lighthouse Outstanding Natural Area (“JILONA”); Jupiter Ridge Natural Area (“JRNA”); and the Arbors. All relationships in panel (a) were recovered with 100% bootstrap support, and bootstrap values in panel (b) are shown above branches.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of nuclear genome of the mycobiont of the Florida Perforate Cladonia (Cladonia perforate—specimen ‘Arbors7’). The size of the assembled fragments is reported, including the number of fragments > 10 kb, >100 kb, and >1 Mb. Annotations results of predicted gene categories from funannotate v. 1.8.7, including carbohydrate-active enzymes (CAZYmes), biosynthetic gene clusters (antiSMASH clusters), biosynthetic enzymes (antiSMASH biosynthetic enzymes), and secondary metabolism gene families (antiSMASH smCOGs).
Table 1.
Summary of nuclear genome of the mycobiont of the Florida Perforate Cladonia (Cladonia perforate—specimen ‘Arbors7’). The size of the assembled fragments is reported, including the number of fragments > 10 kb, >100 kb, and >1 Mb. Annotations results of predicted gene categories from funannotate v. 1.8.7, including carbohydrate-active enzymes (CAZYmes), biosynthetic gene clusters (antiSMASH clusters), biosynthetic enzymes (antiSMASH biosynthetic enzymes), and secondary metabolism gene families (antiSMASH smCOGs).
| # of Contigs | 72 |
|---|---|
| Total size | 33.61 Mb |
| Longest/shortest contig | 1.75 Mb/2 Kb |
| >10 kb/>100 kb/>1 Mb | 53/41/14 |
| N50/L50 | 1.1 Mb/12 |
| GC content | 47.05% |
| BUSCO complete single | 1250/1315 (95.1%) |
| BUSCO complete duplicate | 2/1315 (0.2%) |
| BUSCO fragmented | 40/1315 (3.0%) |
| BUSCO missing | 23/1315 (1.7%) |
| Total annotations | 13,980 |
| CAZYmes | 215 |
| antiSMASH clusters | 48 |
| antiSMASH biosynthetic enzymes | 99 |
| antiSMASH smCOGs | 141 |
Table 2.
Summary of Cladonia perforata multiple sequence alignments. Pairwise identification was not estimated for nuclear genomic data, including coding and BUSCO loci (indicated as “NA”).
Table 2.
Summary of Cladonia perforata multiple sequence alignments. Pairwise identification was not estimated for nuclear genomic data, including coding and BUSCO loci (indicated as “NA”).
| Data Matrix | MSA Size | Genetic Groups | Pairwise Identity (Identical Sites) |
|---|---|---|---|
| Nuclear Genome | 32.1 Mb | 6 | NA |
| Coding loci | 12.1 Mb | 6 | NA |
| BUSCO loci | 1.91 Mb | 6 | NA |
| Cistron | 9777 bp | 2 | 99.7% (99.3%) |
| ITS | 565 bp | 2 (3) 1 | 99.5% (98.1%) |
| Mitochondrial Genome | 59,014 bp | 1 | 100% (100%) |
1. ITS data include the two currently available Cladonia perforata sequences available on GenBank (accession Nos. AF457903 and AY753584); the two GenBank sequences formed a third distinct group, closely related to the sequences generated from specimens from Jupiter Inlet Outstanding Natural Area.
Table 3.
Florida Perforate Cladonia specimens sampled for Illumina high-throughput sequencing. The geographic origin for each of the three populations is coded as: “Arbors” for the Arbors Preserve, “JILONA” for Jupiter Inlet Lighthouse Outstanding Natural Area, and “JRNA” for Jupiter Ridge Natural Area. The total number of Illumina short reads per sample is reported along with the average coverage for the nuclear, mitochondrial (mt), and nuclear ribosomal cistron (nrDNA) of the mycobiont Cladonia perforata per sample.
Table 3.
Florida Perforate Cladonia specimens sampled for Illumina high-throughput sequencing. The geographic origin for each of the three populations is coded as: “Arbors” for the Arbors Preserve, “JILONA” for Jupiter Inlet Lighthouse Outstanding Natural Area, and “JRNA” for Jupiter Ridge Natural Area. The total number of Illumina short reads per sample is reported along with the average coverage for the nuclear, mitochondrial (mt), and nuclear ribosomal cistron (nrDNA) of the mycobiont Cladonia perforata per sample.
| Sample Code | Illumina Reads | Nuclear Coverage | mt Coverage | nrDNA Coverage |
|---|---|---|---|---|
| Arbors1 | 3,204,340 | 9× | 169× | 32× |
| Arbors2 | 5,439,708 | 15× | 251× | 50× |
| Arbors3 | 4,727,072 | 12× | 186× | 65× |
| Arbors4 | 5,475,152 | 14× | 287× | 36× |
| Arbors5 | 1,927,270 | 4× | 61× | 11× |
| Arbors6 | 3,623,778 | 8× | 130× | 35× |
| Arbors7 1 | 3,646,488 | 10× | 159× | 53× |
| JILONA1 | 4,254,622 | 9× | 187× | 54× |
| JILONA2 | 4,915,262 | 12× | 185× | 64× |
| JILONA3 | 6,474,484 | 15× | 271× | 66× |
| JILONA4 | 4,708,534 | 12× | 215× | 52× |
| JILONA5 | 4,270,992 | 11× | 175× | 72× |
| JILONA6 | 4,627,850 | 11× | 209× | 36× |
| JILONA7 | 2,052,574 | 4× | 84× | 26× |
| JRNA1 | 4,086,004 | 9× | 170× | 16× |
| JRNA2 2 | 4,827,906 | 12× | 165× | 94× |
| JRNA3 | 3,255,906 | 8× | 133× | 26× |
| JRNA4 | 5,457,804 | 13× | 230× | 29× |
| JRNA5 | 4,800,882 | 12× | 225× | 30× |
| JRNA6 | 3,589,860 | 8× | 207× | 18× |
| JRNA7 | 5,092,998 | 13× | 245× | 40× |
1 Specimen ‘Arbors7’ was selected for long read de novo shotgun sequencing using the Oxford Nanopore Technologies platform. 2 Specimen ‘JRNA2’ represents Cladonia leporina and was excluded from all analyses.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Leavitt, S.D.; DeBolt, A.; McQuhae, E.; Allen, J.L. Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata). J. Fungi 2023, 9, 698. https://doi.org/10.3390/jof9070698
AMA Style
Leavitt SD, DeBolt A, McQuhae E, Allen JL. Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata). Journal of Fungi. 2023; 9(7):698. https://doi.org/10.3390/jof9070698
Chicago/Turabian StyleLeavitt, Steven D., Ann DeBolt, Ethan McQuhae, and Jessica L. Allen. 2023. "Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata)" Journal of Fungi 9, no. 7: 698. https://doi.org/10.3390/jof9070698
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.