
Polyploid Induction and Identification of Begonia × benariensis
by
 ,
,
Ninghao Xie
1 ,
,
Yi Zhao
1,
Min Huang
1,
Caixia Chen
1,2,
Chuanqu Cao
2,
Jisheng Wang
2,
Zhihua Shi
2 and
Junshan Gao
1,* 1
School of Life Sciences, Anhui Agricultural University, Hefei 230036, China
2
Hefei Hualv Seedling Co., Ltd., Hefei 230041, China
*
Author to whom correspondence should be addressed.
Horticulturae 2024, 10(1), 47; https://doi.org/10.3390/horticulturae10010047
Submission received: 1 December 2023
/
Revised: 26 December 2023
/
Accepted: 28 December 2023
/
Published: 2 January 2024
(This article belongs to the Section Genetics, Genomics, Breeding, and Biotechnology (G2B2))
Abstract
:Begonia × benariensis series varieties have high ornamental and economic value and are excellent varieties for large-area group planting in gardens and green areas. At present, the B. × benariensis series varieties grown in China rely on foreign imports, and the price of imported germplasm resources is expensive, which greatly increases the cost of flower enterprises. In view of this, B. × benariensis (2n = 2x = 34) was used as the material, the callus was treated with colchicine to induce polyploid plants, and the polyploid plants were subjected to morphology, DNA content identification, and stomatal identification, as well as comparisons of the morphophysiological indexes of the plants. The results showed that the polyploid was better induced with the treatment of 0.05% colchicine for 4 h, and the induction rate was 46.67%. Among the mutagenized plants, there were 42 triploids (2n = 3x = 51) and 98 tetraploids (2n = 4x = 68). The stomatal density of diploid leaves was about 1.09 times that of the triploid and 1.67 times that of the tetraploid, the defense cells of the leaves increased gradually, and the stomatal density decreased; with the increase in ploidy, the height of the plants increased, while the crown width decreased; the maximum length and width of the leaves decreased, the color of the leaves became darker, and the chlorophyll content increased. This study provides abundant variation materials and technical support for the selection and breeding of new varieties of B. × benariensis in the future.
1. Introduction
Begonia × benariensis ‘BIG’® Begonia is a perennial herbaceous flower belonging to the genus Begonia in the family Begoniaceae C. Agardh (Begonia L.); it is a hybrid generation of varieties selected and bred by the Benary Seed Company in Germany, with Begonia cucullata and Begonia maculata Raddi as the main parents of the hybrid progeny. The ‘BIG’® series can be divided into green leaf and red flower type, copper leaf and red flower type, and copper leaf and rose-red flower type, bronze leaf and rose-white flower type, and green leaf and white flower type according to the leaf color and flower color [1]. B. × benariensis is an upright plant with a height of 40–50 cm and heart-shaped shiny leaves. The petals are erect and spreading, and the flowers are bright and dazzling in color [2]. The ‘BIG’® series of varieties combines the advantages of many kinds of Begonia and was awarded the “Special Prize at the Moscow International Flower Show 2008”.
The main sources of polyploids are naturally occurring, synthetic, and induced mutations. Natural occurrence is the phenomenon of doubling the number of chromosomes in polyploids, and natural mutation is the more common way of generating polyploid plants and plant evolution in nature. Induced mutation is the most commonly used method in polyploid breeding, and artificial induction of plant polyploids is mainly through the physical induction method, chemical induction method, and biological polyploidy induction method. The objects of physical induction include germinating seeds, seedlings, male and female flower buds, etc., but it has the disadvantages of a low induction rate, poor directionality, serious chimerism, and great harm. The chemical induction method refers to the use of certain chemical reagents to treat the dividing plant organs, tissues, and even cells, to induce the cell chromosome doubling method. Compared with the traditional physical induction method, chemical induction is characterized by a short time, stable progeny, high specificity, and a simple method. Chemical induction of polyploidy is one of the most commonly used and effective methods [3]. At present, more than 200 chemical agents have been found to have an inducing effect on plant polyploidy, and the most widely used and effective agent is colchicine. Because the effect of colchicine is limited to hindering the formation of spindle filaments in the middle of cell division, few other unfavorable mutations occur genetically [4]. The biopolyploidy induction method is a new technology, which is mainly used in the field of plants, such as sexual hybridization, endosperm culture, cell fusion (somatic cell hybridization), tissue culture, etc., and the development of this technology was established on the basis of a tissue culture and protoplast culture.
In the chemical mutation breeding of plants and flowers, colchicine is mainly used to induce the production of polyploids to obtain new varieties. The induced new varieties are often characterized by organ gigantism, adaptability, and stout plants [5], which increase the ornamental and commercial value of flowers, which has been successful in many flowers. Meanwhile, polyploid breeding can produce deformed flowers, and through continuous selection and stabilization of the deformed flowers, new varieties can be quickly created for germplasm innovation.
At present, research on B. × benariensis at home and abroad has only been reported in landscape application, cultivation and management techniques, and cuttings propagation; however, there is no report on polyploidy induction in B. × benariensis. B. × benariensis is still bred through traditional sexual reproduction, and the F1 generation of hybridization has a high hybrid dominance and good growth traits in the field. However, if the seed is kept by itself, the F2 generation will be separated in terms of the traits, and the germination rate of the seed is not high; so, in general, the F2 generation of the seed is not used for sowing. According to market research, the domestic planting of B. × benariensis is mainly dependent on foreign imports, which are expensive, with a seed price of up to CNY 0.7 to 0.8, greatly increasing the cost of floriculture enterprises; so, some small- and medium-sized floriculture enterprises are in a static position. In agriculture, the seeds of tetraploid plants also have a wide range of application possibilities. Tetraploid plants are usually more stress-resistant than diploids, which makes them more capable of surviving in adverse environmental conditions. Tetraploid flower seeds are more likely to survive in arid, cold, saline, and alkaline environments, which not only improves flower yields, but also expands the geographic scope of their application.
In this study, we aimed to establish a polyploid induction system of B. × benariensis with a high induction rate and stable and prominent traits, to provide technical support for breaking the monopoly of foreign germplasm resources and commercial production of B. × benariensis seedlings, and to accelerate the factory production and popularization of the seedlings. At the same time, it also provides guiding ideas for the future establishment of B. × benariensis production lines in China.
2. Materials and Methods
2.1. Plant Material and Explant Preparation
In this experiment, the B. × benariensis ‘BIG’® series of the green leaf and red flower type and copper leaf and red flower type (the seeds were purchased from the Benary Company in Germany) were selected as the experimental materials and planted in the intelligent greenhouse of Hefei Hualü Seedling Co., Ltd. (Figure 1). When the plants grew to 40–50 cm tall, the plants with good horticultural traits were selected as the mother plants. The young leaves of the same size were selected and washed with detergent and then distilled water, placed on the ultra-clean bench, and sterilized with 75% alcohol for 30 s; then, 0.1% HgCl2 was poured onto the leaves for 10 min, the sterilizing solution was removed, and the leaves were washed and soaked with sterilized water about 5 times to obtain the sterile leaves, which were then placed in a 7 cm petri dish lined with sterile filter paper to absorb the water. The leaves were cut into about 1 cm2 pieces with a sterile scalpel for the callus induction.
2.2. Induction of Leaf Callus
The base medium for callus induction was an MS medium with additional concentrations of 1 mg/L of 6-BA, 0.1 mg/L of 2.4-D, and 0.2 mg/L of NAA, and 7 g/L of agar and 30 g/L of sucrose were added to the medium; the pH value of the medium was adjusted to 5.8. A sterilized explant was put into the design of the medium with the leaf side facing up, and 4 leaves were inoculated in each bottle of medium. The inoculated medium was incubated in a group culture room for 20 days, waiting for the production of calluses.
2.3. Induction of Polyploidy in Begonia × benariensis by Colchicine
B. × benariensis calluses were selected as the materials, the colchicine concentration was set at 0.01%, 0.05%, and 0.1%, three gradients, and the treatment time was set at 2 h, 4 h, and 8 h, three gradients, with five bottles for each treatment, four pieces per bottle, and three replicates. On the ultra-clean bench, the calluses were inserted into the sterilized colchicine solution with different concentrations, and then placed in a light-proof environment in the tissue culture room. According to the different treatments, the healing tissues were taken out at the appropriate time, cleaned, and then transferred into the differentiation medium (the same medium as in Section 2.2), observed, and recorded. Subculturing was carried out every 60 d or so. Statistics on the number of mutant plants and mutation rate were recorded.
2.4. Ploidy Identification of Polyploid Begonia × benariensis Plants
First, the morphological identification was carried out by observing the morphological differences of the mutagenized plants, using B. × benariensis undoubled plants as the control. The mutant and control plants were cultured in a rooting medium (the base medium was 1/2MS with an additional concentration of 1.0 mg/L of NAA) for a certain period of time; then, the morphological identification of test tube seedlings was carried out, and the differences in leaf size, plant morphology, and leaf color between the mutant and control plants were analyzed.
Flow cytometry can be used for the rapid screening and early detection of aneuploidy in hybrid seedlings of species. The DNA content of each mutant and wildtype plant was determined using a BD FACSCanto II flow cytometer (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) with an excitation light source at 488 nm and a DNA fluorescent dye, propidium iodide (PI). Each sample was repeated three times, and a minimum of 3000 nuclei were counted in each run. The ploidy of the different samples was initially determined from the mean fluorescence value of the G0/G1 peaks according to the histograms (mean fluorescence value as the horizontal coordinate and number of nuclei as the vertical coordinate) displayed by the BD FACSDiva™ 9.0 (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) software. The specific operation is as follows.
2.4.1. Preparation of Nuclei Suspension
Avoiding the main veins of the leaf blade, we cut about 0.2 g of fresh leaf tissue, placed it in a small petri dish (60 mm × 15 mm), added 1.5 mL of a pre-cooled dissociation solution (10 mmol/L of MgSO4-7H2O, 50 mmol/L of KCl, 3.5 mmol/L of HEPES buffer, 0.3% Triton™ X-100, 1% Polyvinylpyrrolidone K30), and quickly chopped the leaf blade into a homogenate with a sharp single-sided blade, after which the homogenate was filtered through a 38 µm nylon filter into a 5 mL sample uptake tube.
2.4.2. Fluorescence Staining
Here, 200 µL of RNase A (100 µg/mL) and 60 µL of the PI staining solution (100 µg/mL) were added to the sample tube, the tube was gently shaken for 5 s, and then it was placed at room temperature and protected from light for 15 min.
2.4.3. Loading Test
We put the sample tube into the instrument at the sampling place, adjusted the voltage parameters and flow rate, plotted the scatter plot and histogram, adjusted the threshold value, screened out the noise peaks on the left side of the image, and selected the left and right sides of the peaks to generate a gate when the obvious main peaks appeared. When the number of cells counted in the gate reached more than 3000, and the rCV value was less than 5%, we recorded the mean value and the rCV value and saved the image. We tested the other samples using the same parameters.
2.4.4. Stomatal Identification
The leaves of the plants to be tested were selected, and colorless nail polish was applied to the lower epidermis of the leaves, which were left to air-dry; then, the air-dried nail polish was torn off, and the stomata were observed under a microscope at 100× to record the guard cell length, the guard cell width, and the number of stomata in the field of view of the microscope; the stomatal density was calculated. The experimental and control groups were measured in 20 groups, and 10 fields of view were counted for each leaf observation.
2.5. Determination of Morphophysiological Indexes in Polyploid Begonia × benariensis Plants
2.5.1. Measurement of Plant Height
We measured the height of a plant’s collar bud from the root of the plant using a meterstick with a sample size of 10 and calculated the mean.
2.5.2. Measurement of Crown Width
We measured the east–west length of a plant with a meterstick with a sample size of 10 and calculated the mean.
2.5.3. Measurement of Leaves
We determined the leaf length and width in the same development period. The specific operation was as follows: we took the largest leaf blade of the plant as the measurement target and used vernier calipers to measure its leaf length and width. In order to ensure the accuracy of the data, we selected the same period of time and the same growth of ten leaves to repeat the measurement; then, we calculated the average value.
2.5.4. Determination of Chlorophyll Content
Random sampling of leaves from each strain of each generation began at about 15:30 p.m. The leaves taken were fresh leaves 4 to 5 cm from the terminal bud and were stored in an ice box immediately after collection. We weighed 0.2 g of freshly cut leaves, put them into a mortar, added a small amount of quartz sand, calcium carbonate powder, and 2 mL of 95% ethanol, ground them into a pulp, and left them in the dark for about 10 min. We filtered the extract into a 25 mL brown volumetric flask, rinsed the mortar, the rod, the filter paper, and the residue with 95% ethanol several times in order to make the color pigment transfer into the volumetric flask, and then we finalized the volume with 95% ethanol to 25 mL, shook it well, and stored it in a dark place in order to use the spare spectrophotometric method to determine the chlorophyll content. The pigment extract was poured into a colorimetric cup with an optical diameter of 1 cm, and the absorbance was measured at wavelengths of 665 nm, 649 nm, and 470 nm, with 95% ethanol as a control. The concentrations of chlorophyll a and b and their total chlorophyll concentrations were calculated based on the following formula for calculating the pigment concentration in ethanol extracts, respectively.
Note: Chl a and Chl b are the concentrations of chlorophyll a and b, respectively. V is the total amount of the extract (mL), and W is the fresh weight of the sample (g).
Total chlorophyll content = Chl a + Chl b
2.6. Data Analysis
All data were analyzed using SPSS20.0 software (Version 20), and a one-way analysis of variance (ANOVA) was used to analyze the significance of the differences between the morphophysiological indicators. The graphs were plotted with origin 2023b software (Version 2023b).
3. Results
3.1. Effects of Different Concentrations of Colchicine on the Induction Efficiency of Begonia × benariensis
In the polyploid induction test with a variable colchicine concentration of 0.01%~0.1% and treatment times of 2 h, 4 h, and 8 h, it was found that the colchicine concentration of 0.05% with a treatment time of 4 h induced a better effect with the highest mutation rate. From Table 1, with the increase in the colchicine concentration, the survival rate of B. × benariensis leaves decreased significantly; with the 0.01% colchicine solution on a B. × benariensis callus, the injury was less, and the survival rate was higher than 60%. Due to the fact that colchicine itself has a certain degree of toxicity, when the treatment time was too long or the concentration was too high, it led to the death of the plant materials. When the concentration of colchicine treatment was 0.1%, the survival rate of the treated callus ranged from 18.33% to 41.67%, which was generally low, and the callus browned rapidly and died gradually. When a 0% colchicine solution was impregnated for 2 h, the survival rate of the callus was the highest, reaching 80%. When a 0.05% colchicine solution was impregnated for 4 h, the survival rate of the callus reached 60%, and the mutation rate of the callus was the highest at 46.67%. In summary, the most suitable treatment for the polyploidy induction of a B. × benariensis callus was a 0.05% colchicine concentration for 4 h.
3.2. Identification of Polyploid Plants of Begonia × benariensis
3.2.1. Morphological Identification
The identification of polyploids generally followed the research steps of an initial morphological screening followed by cytological confirmation. At the early stage of mutagenesis (Figure 2), the colchicine-treated mutant plants showed slower growth, smaller leaves, darker leaf color, and increased leaf thickness in the early stage compared with control plants (Table 2), and after transplanting (Figure 3), the plant height and crown width were significantly lower than that of control plants, and the leaves were wrinkled and unevenly colored (Table 3).
3.2.2. Flow Cytometry Identification
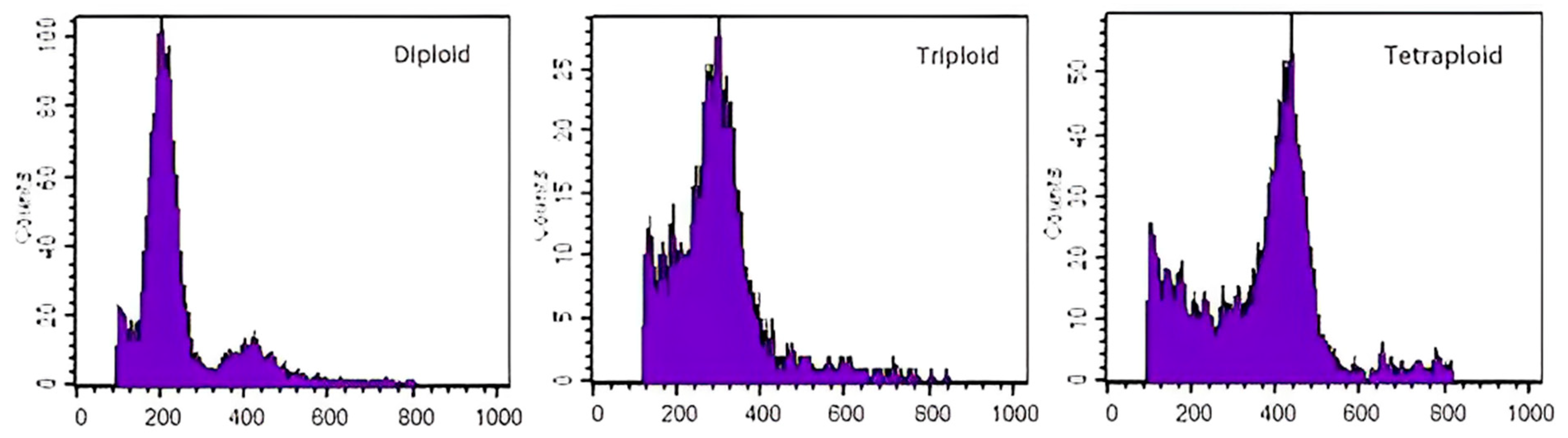
Through subculture and acclimation transplanting of a callus treated with colchicine, 283 regenerated plants were obtained. The untreated diploid calophylla plants were used as the control (CK), and the leaves of CK and mutant plants were tested through flow cytometry. The fluorescence channel value was set to 200, and the ordinate represents the relative cell number of the tested sample. The horizontal coordinate represents the fluorescence channel value, and the ploidy of the tested sample was determined using the position of the peak value. The fluorescence channel of the diploid plants (CK) showed a peak value around 200. The fluorescence channel of the triploid plants peaked at about 300, and the DNA content of the triploid plants was about 1.5 times that of the CK plants. The fluorescence channel of the tetraploid plants peaked at about 400, and the DNA content of the tetraploid plants was about twice that of CK plants (Figure 4). In total, 42 triploids and 98 tetraploids were detected.
3.2.3. Stomatal Identification
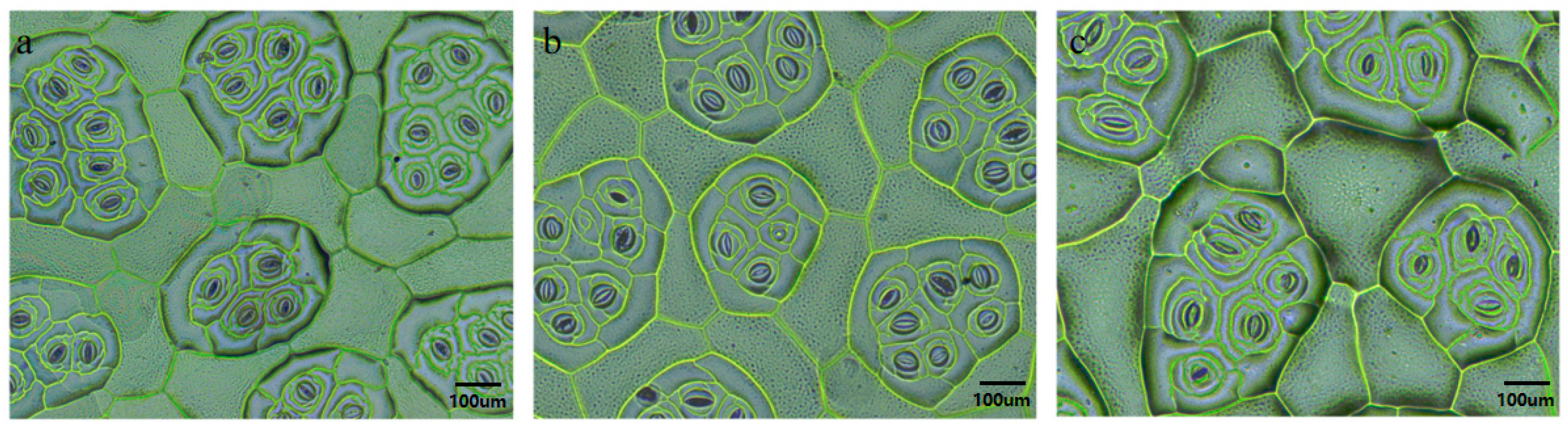
The number of stomata, the length and width of the defense cells, and the stomatal density of plants with different ploidies differed significantly. The lower epidermis of the leaves of untreated B. × benariensis plants (CK) and mutant plants with uniform development was selected for stomatal observation and measured, and the length and width of defense cells and the stomatal density of the mutant plants were enlarged 100 times. Stomatal observation was carried out on the lower epidermis of the leaves after treatment, and the test revealed that the stomata of the lower epidermis of the leaves of the mutant plants were significantly larger than those of the diploid plants, and the density of stomata in the same field of view was significantly less than that of the diploid plants. The average length of the defense cells of the diploid plants was about 58.98 µm, and the average width was about 14.35 µm; the average length of the defense cells of the tetraploid plants was about 85.42 µm, and the average width was about 20.89 µm; the average length of the defense cells of the triploid plants was about 63.12 µm, and the average width was about 14.39 µm; and the stomatal density of diploid plants was about 1.09 times that of the triploid plants and 1.67 times that of the tetraploid plants (Table 4). The results showed that the number of stomata per unit area decreased as the ploidy of doubled plants increased, and the density decreased. The stomatal density was significantly lower in the tetraploids than in diploids (Figure 5).
3.3. Comparison of Morphophysiological Indexes in Doubled Begonia × benariensis Plants
3.3.1. Morphological Differences in Begonia × benariensis Plants with Different Ploidies
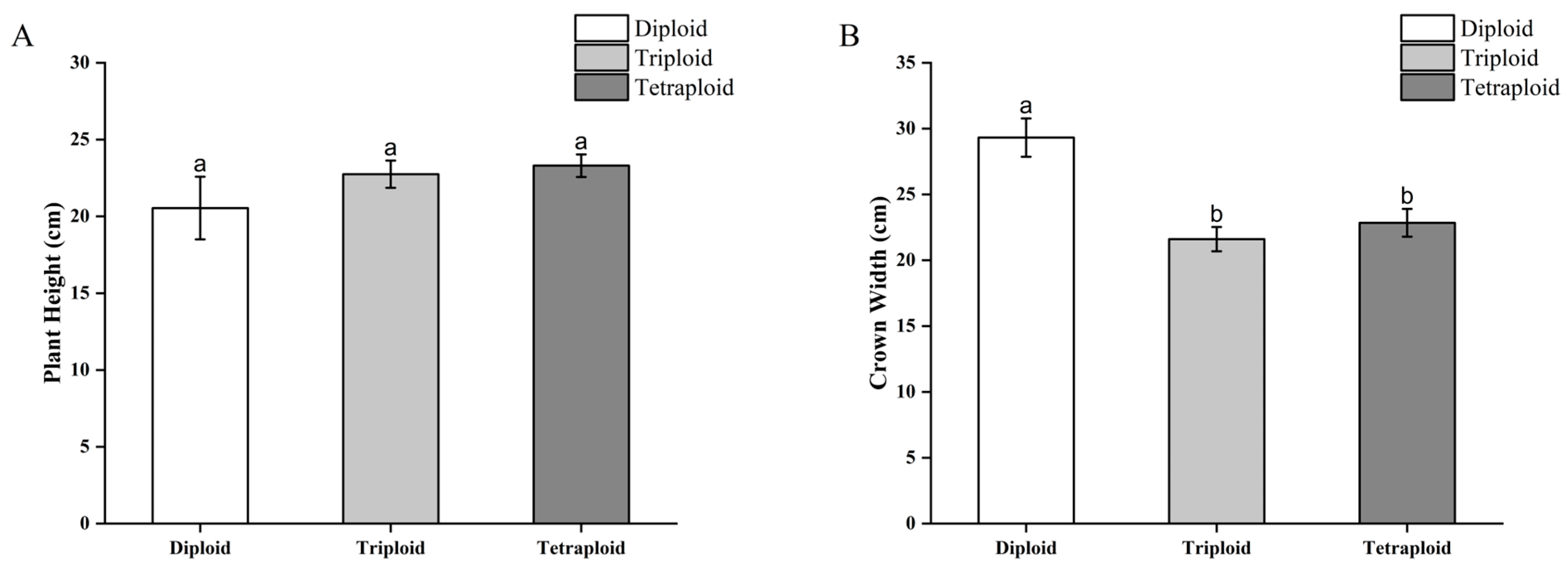
B. × benariensis chromosome-doubled plants and control plants were transplanted and grown for three months, and the differences in plant height and crown width were comparatively analyzed. The average plant height of the diploid plants was 20.54 cm, and the average crown width was 29.32 cm; the average plant height of the triploid plants was 22.75 cm, and the average crown width was 21.60 cm; and the average plant height of the tetraploid plants was 23.30 cm, and the average crown width was 22.85 cm. The differences between the diploids, triploids, and tetraploids were not significant in terms of the plant height (Figure 6A). In terms of the crown width, the diploids had the highest average crown width, and there was a significant difference between the diploids and triploids and tetraploids, while the difference between the triploids and tetraploids was not significant (Figure 6B).
3.3.2. Morphological Differences in Begonia × benariensis Leaf Blades of Different Ploidies
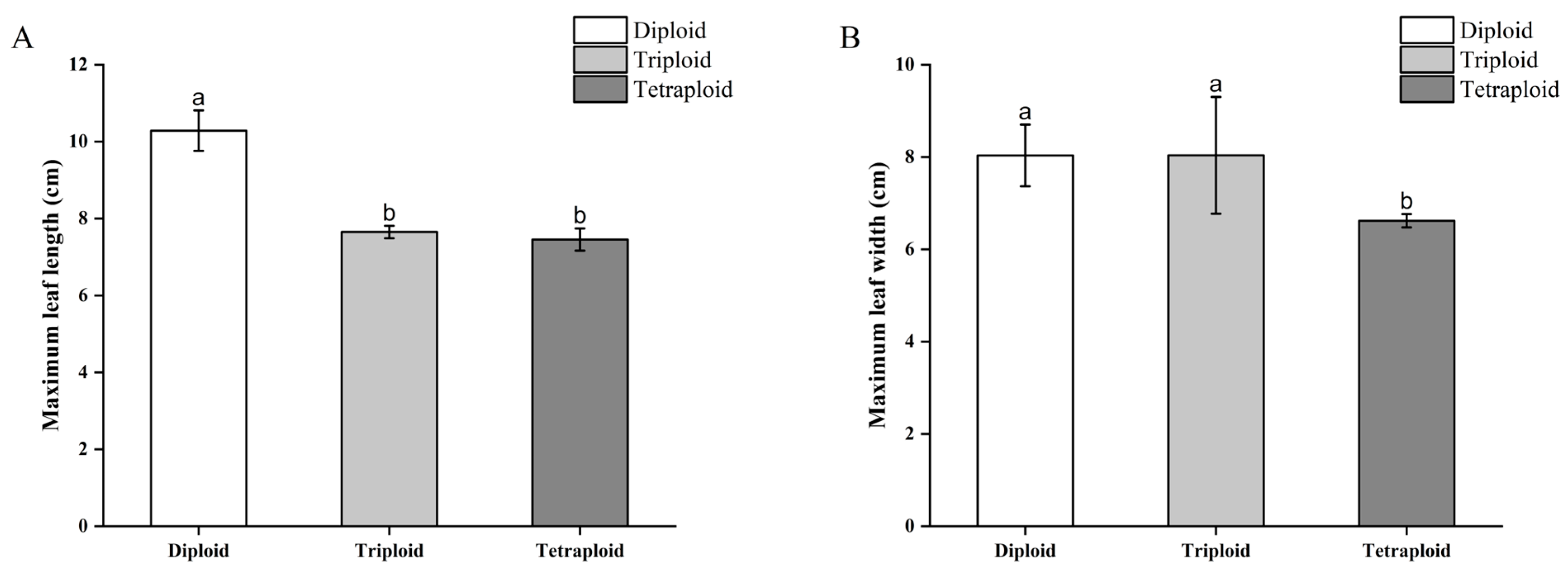
B. × benariensis chromosome-doubled plants and control plants were transplanted and grown for three months, and the differences in the maximum leaf length and maximum leaf width of different ploidy plants were compared. The diploid and triploid plants differed significantly from the tetraploid plants in leaf morphology. The average maximum leaf length of the diploid plants was 10.28 cm, and the maximum leaf width was 8.03 cm; the average maximum leaf length of the triploid plants was 7.65 cm, and the maximum leaf width was 8.04 cm; the average maximum leaf length of the tetraploid plants was 7.45 cm, and the maximum leaf width was 6.62 cm. The leaf blades of the tetraploid plants were smaller than those of the diploid and triploid plants, and the leaf color was darker, the leaf blades were thicker, and the leaf blades were obviously serrated. The leaf blades of the triploid plants were slightly smaller than those of diploid plants, but there was a significant difference in the maximum leaf length between triploid and diploid plants, while the difference in the maximum leaf width was not significant. The maximum leaf length of the tetraploid plants was significantly different from the diploid but not from the triploid, and the maximum leaf width was significantly different from the leaves of the diploid and triploid plants (Figure 7).
3.3.3. Differences in Chlorophyll Content of Begonia × benariensis Leaf Blades of Different Ploidies
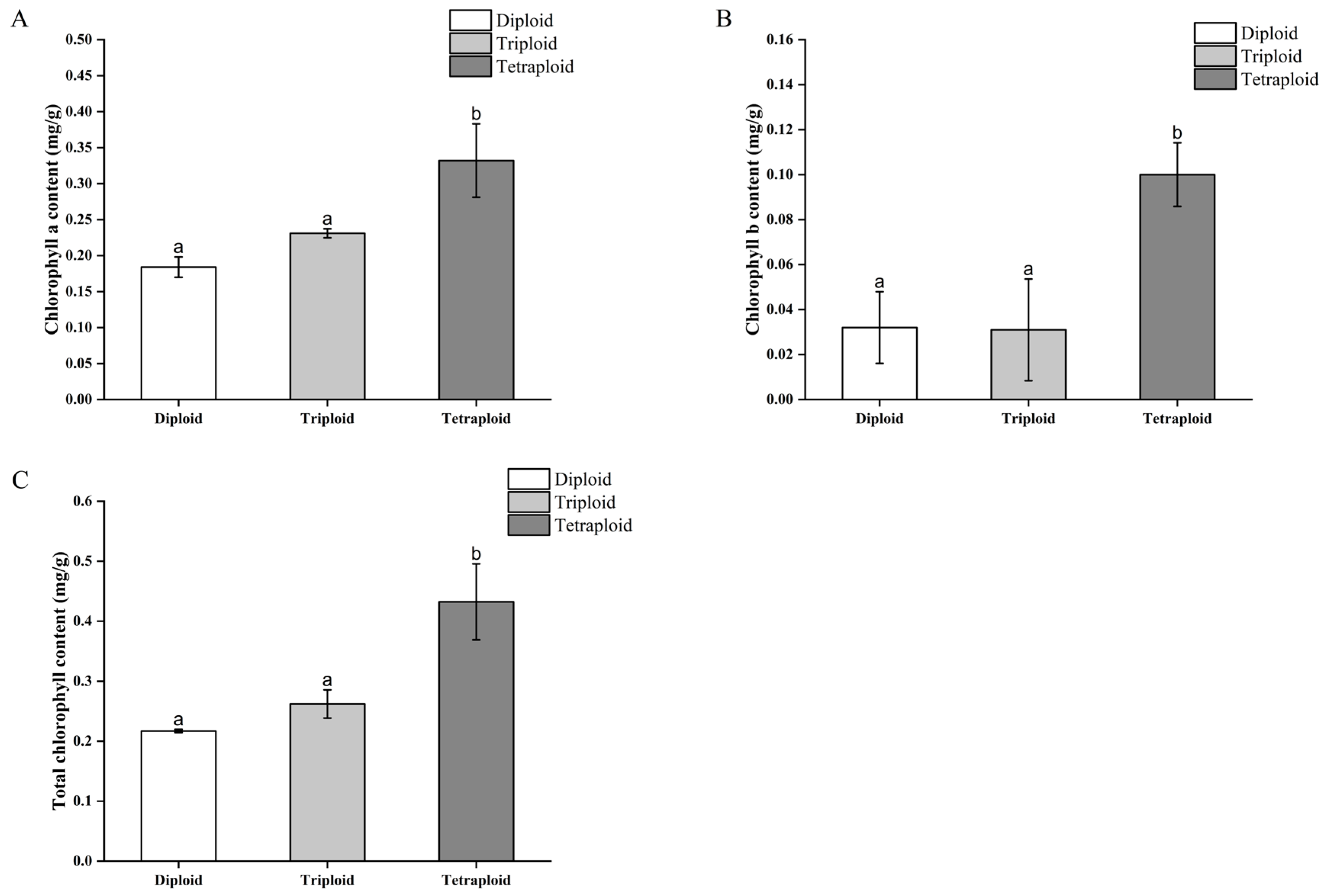
The chlorophyll content of the B. × benariensis chromosome-doubled plants and the control plants was measured three months after transplanting and growing. The content of chlorophyll a, chlorophyll b, and total chlorophyll in the functional leaves of the tetraploid plants was higher than that of the diploid plants, which could be the reason for the dark green color of the leaves of tetraploid plants, and there was a significant difference. However, the chlorophyll content of the triploid plants was not significantly different from that of the diploid plants (Figure 8).
4. Discussion
4.1. Induction of Polyploidy by Colchicine Impregnation
In recent years, the combination of a tissue culture and polyploid breeding has become very common, and in the selection of induction methods, plant polyploids can be obtained through the physical induction pathway, which produces polyploids by damaging the genomic DNA of the cells and altering their genetic material. Sun et al. irradiated ‘Fertility pear’ test tube seedlings with γ-rays and inoculated their terminal buds, lateral buds, and leaves, respectively, and obtained ‘Fertility pear’ triploids and tetraploids after subculturing and screening, as well as the identification of chromosome counts [6]. Zhang et al. obtained a new set of efficient polyploid induction methods by injecting low-energy N+ to treat Astragalus membranaceus (Fisch.) Bge. var. mongholicus (Bge.) Hsiao [7]. However, these methods are prone to form chimeras, have an unstable mutation, and have certain requirements for the mutagenesis apparatus, as well as a high cost. Therefore, this experiment was selected to carry out research through chemical mutagenesis. There are many studies on polyploid induction by colchicine impregnation and mixed cultivation, and many scholars have used both methods to test in order to find the optimal induction method; however, there have been different results in different crops. Previous studies found that the impregnation method was more suitable for polyploid induction in Vaccinium vitis-idaea L. [8] and Vaccinium uliginosum [9] than mixed cultivation; the mixed cultivation method was found to be more suitable for polyploid induction in dragon fruit [10], yellow-flesh Actinidia Chinensis [11], etc., than the impregnation method. In this experiment, we used the impregnation method to induce B. × benariensis polyploid plants and screened the treatment combinations with the best mutagenic ratios with different treatment concentrations and treatment times; through the experimental results, we concluded that the best treatment for B. × benariensis induction was to treat the plants with 0.05% colchicine concentration for 4 h, and the mutagenicity rate reached 46.67%.
4.2. Ploidy Identification of Begonia × benariensis Polyploids
When plants are identified for ploidy, they can be roughly screened using external morphology first, and morphological identification is a more common and simpler identification method for polyploidy. Generally, polyploid plants are larger than normal plants, with broad and thick leaves, thick stems, larger floral organs, and other morphological characteristics [12,13,14]. The gigantism of organs in the polyploid plants is an important characteristic, but plants with hypertrophied organs are not always polyploids, and not all polyploids are definitely hypertrophied.
Further, polyploid stomatal size can be identified at the cytological level, the polyploid stomatal size also increases with the increase in plant ploidy, and the observation of plant stomatal size can be a further screening for polyploid plants [15]. Liu et al. identified the ploidy of Clematis fruticose by observing the stomata and found that tetraploid plants had significantly larger stomata [16]. Observation of the plant chromosome number through root tip chromosome preparation is the most accurate method for identifying plant ploidy. The number of plant chromosomes can clearly determine plant ploidy [17]. Furthermore, the analysis of plant DNA content through flow cytometry is a simple, fast, and accurate method to directly compare the DNA content of different plants [18]. This method has been successfully used to wild Clematis [19], Rhododendron fortunei Lindl [20], Pointed gourd (Trichosanthes dioica Roxb.) [21], pear (Pyrus communis L.) cultivar ‘Fertility’ [22], Paphiopedilum villosum [23] and other plant ploidy identification. In this experiment, B. × benariensis root tip chromosome observation was carried out. Due to the slender root tip, it is difficult to manipulate; hence, three methods, namely, morphology preliminary identification, stomatal identification, and flow cytometry identification, were used to identify the ploidy of doubled plants. The experimental results showed that, although most scholars who studied polyploid plants showed gigantism compared to diploid plants, the polyploid plants often showed gigantism in their organs in terms of the external morphology compared to the diploid plants because of changes in the chromosome numbers in the body, for example, enlarged and thicker leaves [24], thicker and stronger stems, and larger flowers and fruits. In leaf epidermal cell morphology, stomatal size and density show signs of polyploidization [25,26,27].
In this study, we found that B. × benariensis tetraploid plants were stouter than the diploid plants from the mutagenic treatment. The comparison of diploid and tetraploid leaves revealed that the tetraploid plants had a higher leaf thickness than the diploids, a darker leaf color, leaf folds, and other significant differences. Observation of the stomata showed that the tetraploid stomata were larger and of reduced stomatal density compared to the diploids. This is consistent with the morphological characteristics of polyploidy summarized by previous authors [28,29]. However, the plants and leaves were not of a giant type, which is different from the results of many scholars and might be due to the difference in the number of genes.
5. Conclusions
B. × benariensis was induced by soaking the callus with colchicine, and polyploidy was identified using morphology, flow cytometry, and morphophysiological indexes of the treated plants. The results showed that a 0.05% colchicine solution for 4 h was the most effective. Compared with the diploid plants, the successfully induced tetraploid plants showed larger defense cells; lower stomatal density; smaller plant height, crown width, and maximum leaf length and width; and increased chlorophyll content, and the color develops as redder and the petals get thicker and bigger. In conclusion, B. × benariensis tetraploid plants can be successfully induced by treating a B. × benariensis callus with colchicine, which can be used as a reference for selecting and breeding new tetraploid varieties of B. × benariensis.
Author Contributions
N.X.: conceptualization, methodology, and writing—original draft. J.G.: project administration and funding acquisition, validation. M.H., Y.Z., C.C. (Caixia Chen): visualization, supervision, writing—review and editing. C.C. (Chuanqu Cao), J.W.: software, validation. All authors discussed the results and commented on the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded and supported by Hefei Municipal Natural Science Foundation, grant number: 2023045.
Data Availability Statement
The data are contained within the article.
Conflicts of Interest
Caixia Chen, Chuanqu Cao, Jisheng Wang, Zhihua Shi has received research grants from Hefei Hualv Seedling Company.
References
- Zhou, H.; Li, M.; Shi, Z.; Yang, J.; Zhang, S. Selection of formulation of nutrient solutions in soilless culture for Begonia × benariensis. J. Anhui Agri. Sci. 2014, 42, 2565–2567. [Google Scholar] [CrossRef]
- Zhai, Y.; Yan, R.; Li, L.; Zhang, L. Optimization of adventitious bud induction technology of Begonia Benariensis based on response surface methodology. Seed 2022, 41, 137–144. [Google Scholar] [CrossRef]
- Zhong, W.; Ba, W.; Zhu, M.; Wu, Y.; Liang, M.; Zhang, L. Polyploid watermelon induction breeding techniques. Contemp. Hortic. 2022, 45, 25–27. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, X.; Li, M. Research progress on colchicine-induced polyploidy in medicinal plants. Jiangsu Agric. Sci. 2014, 42, 178–181. [Google Scholar] [CrossRef]
- Wu, T.; Jia, R.; Yang, S.; Zhao, X.; Yu, X.; Guo, Y.; Ge, H. Research advances and prospects on Phalaenopsis polyploid breeding. Acta Hortic. Sin. 2022, 49, 448–462. [Google Scholar] [CrossRef]
- Sun, Q.; Sun, H.; Zhu, E.; Li, L. Polyploid induction in pear in vitro treatment with gamma-rays. Acta Hortic. Sin. 2009, 36, 257–260. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Q.; Yang, L.; Wu, C. Effect of N+ implantation on the inducement of polyploid Astragalus membranaceus (Fisch.) Bge. Var. mongholicus (Bge.) Hsiao. Bull. Bot. Res. 2011, 31, 563–568. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Z.; Li, Y.; Wu, L.; Liu, H. Study on polyploid induction of blueberry in vitro with colchicine treatment. J. Northeast. Agric. Univ. 2010, 41, 38–42. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, H.; Yue, S.; Yang, T.; Bao, Q.; Lu, G.; Xie, Y. Effect of colchicine on blueberry chromosome doubling and preliminary identification. J. Kunming Univ. 2015, 37, 72–76. [Google Scholar] [CrossRef]
- Cheng, Z.; Sun, C.; Sun, P.; Guo, S.; Wu, Q. Optimization of polyploidy induction system in dragon fruit. J. Fruit Sci. 2020, 37, 1089–1097. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, X.; Zhang, Y.; Liu, X.; Ye, Q.; Li, Y.; Zhang, H. Polyploid induction and identification of Yellow-flesh Actinidia Chinensis. Mol. Plant Breed. 2020, 18, 4036–4040. [Google Scholar] [CrossRef]
- Premjet, D.; Obeng, A.K.; Kongbangkerd, A.; Premjet, S. Intergeneric hybrid from Jatropha curcas L. and Ricinus communis L.: Characterization and polyploid induction. Biology 2019, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, K.; Zhao, Q.; Yu, J.; Gongga, Y.; Hu, B. Induction of polyploid Malus sieversii (Ledeb.) Roem. and characteristic analysis of its leaves. J. Tianjin Agric. Univ. 2021, 28, 6–10. [Google Scholar] [CrossRef]
- Xi, Y.; Dong, X.; Yang, M.; Meng, Q.; Huang, T. In Vitro polyploid induction and establishment of a clone for Cyclocodon lancifolius (Roxb.) Kurz. Cytologia 2021, 86, 367–374. [Google Scholar] [CrossRef]
- Pachakkil, B.; Midori, I.; Yukiko, K.; Ryo, M.; Hidehiko, K.; Antonio, L.; Hironobu, S. Somatic polyploidization and characterization of induced polyploids of Dioscorea rotundata and Dioscorea cayenensis. Afr. J. Biotechnol. 2016, 15, 2098–2105. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, Y.; Li, M.; Niu, W.; Liu, D. Polyploid induction and identification of Clematis fruticose. For. Ecol. Sci. 2023, 38, 338–344. [Google Scholar] [CrossRef]
- Huang, L.; Yin, X.; Yang, Y.; Li, W.; Yi, Z. Polyploid induction on the hybrid of miscanthus lutarioriparia × Miscanthus sinensis. Acta Bot. Boreali-Occident. Sin. 2015, 35, 50–56. [Google Scholar] [CrossRef]
- Cruz, V.; Lynch, A.; Ray, D.; Niaura, W.; Purdy, P.; Dierig, D. Analysis of mode of reproduction of guayule (Parthenium argentatum A. Gray) using flow cytometry and identification of polyhaploids for breeding. Ind. Crops Prod. 2017, 107, 618–623. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, Y.; Niu, W.; Wang, X.; Li, M.; Liu, D. Polyploid induction and identification of two wild Clematis. North. Hortic. 2022, 18, 52–59. [Google Scholar]
- Mo, L.; Chen, J.; Lou, X.; Xu, Q.; Dong, R.; Tong, Z.; Huang, H.; Lin, E. Colchicine-induced Polyploidy in Rhododendron fortunei Lindl. Plants 2020, 9, 424. [Google Scholar] [CrossRef]
- Hassan, J.; Miyajima, I.; Ozaki, Y.; Mizunoe, Y.; Sakai, K.; Zaland, W. Tetraploid induction by colchicine treatment and crossing with a diploid reveals less-seeded fruit production in pointed gourd (Trichosanthes dioica Roxb.). Plants 2020, 9, 370. [Google Scholar] [CrossRef]
- Sun, Q.; Sun, H.; Li, L.; Bell, R.L. In vitro colchicine-induced polyploid plantlet production and regeneration from leaf explants of the diploid pear (Pyrus communis L.) cultivar, ‘Fertility’. J. Hortic. Sci. Biotechnol. 2009, 84, 548–552. [Google Scholar] [CrossRef]
- Huy, N.P.; Tam, D.T.T.; Luan, V.Q.; Tung, H.T.; Hien, V.U.; Ngan, H.T.M.; Duy, P.N.; Nhut, D.T. In vitro polyploid induction of Paphiopedilum villosum using colchicine. Sci. Hortic. 2019, 252, 283–290. [Google Scholar] [CrossRef]
- Luo, Z.; Brian, J.; Katrina, C. Colchicine-induced polyploidy has the potential to improve rubber yield in Taraxacum kok-saghyz. Ind. Crops Prod. 2018, 112, 75–81. [Google Scholar] [CrossRef]
- Mahpara, K.; Sabbi, J.; Aijaz, A.; Manoj, K. Induction of polyploidy in saffron (Crocus sativus L.) using colchicine. J. Crop Improv. 2021, 36, 555–581. [Google Scholar] [CrossRef]
- Mo, L.; Chen, J.; Chen, F.; Xu, Q.; Tong, Z.; Huang, H.; Dong, R.; Lou, X.; Lin, E. Induction and characterization of polyploids from seeds of Rhododendron fortunei Lindl. J. Integr. Agric. 2020, 19, 2016–2026. [Google Scholar] [CrossRef]
- Liu, Y.; Duan, S.; Jia, Y.; Hao, L.; Xiang, D.; Chen, D.; Niu, S. Polyploid induction and karyotype analysis of Dendrobium officinale. Horticulturae 2023, 9, 329. [Google Scholar] [CrossRef]
- Ren, X.; Wang, X.; Xiao, F.; Zhou, Y.; Wei, L. Polyploidy induction and identification of Hibiscus syriacus. J. West China For. Sci. 2019, 48, 119–125. [Google Scholar] [CrossRef]
- Li, J.; Deng, W.; Xu, Z.; Zhang, Z.; Lu, B. Induction and identification of polyploid from Pistacia chinensis. J. Northeast. For. Univ. 2020, 50, 18–22. [Google Scholar] [CrossRef]
Figure 1.
Test Begonia × benariensis plants. (a) Green leaf and red flower type. (b) Copper leaf and red flower type.
Figure 1.
Test Begonia × benariensis plants. (a) Green leaf and red flower type. (b) Copper leaf and red flower type.

Figure 2.
Growth of tissue cultured seedlings in the early stage of mutagenesis. (a) Control plant. (b) Variant plant.
Figure 2.
Growth of tissue cultured seedlings in the early stage of mutagenesis. (a) Control plant. (b) Variant plant.

Figure 3.
Plant growth after transplanting tissue culture seedlings. (a) Control plant. (b) Variant plant.
Figure 3.
Plant growth after transplanting tissue culture seedlings. (a) Control plant. (b) Variant plant.

Figure 4.
Flow cytometry ploidy analysis chart.

Figure 5.
Comparison of stomata in the lower epidermis of diploid, triploid, and tetraploid leaves of Begonia × benariensis (100×). (a) Diploid. (b) Triploid. (c) Tetraploid.
Figure 5.
Comparison of stomata in the lower epidermis of diploid, triploid, and tetraploid leaves of Begonia × benariensis (100×). (a) Diploid. (b) Triploid. (c) Tetraploid.

Figure 6.
Comparison of differences in plant height and crown width of doubled Begonia × benariensis plants. (A) Plant height. (B) Crown width. Different letters indicate that the parameter values are significantly different at the level of p < 0.05.
Figure 6.
Comparison of differences in plant height and crown width of doubled Begonia × benariensis plants. (A) Plant height. (B) Crown width. Different letters indicate that the parameter values are significantly different at the level of p < 0.05.

Figure 7.
Comparison of leaf morphology of doubled Begonia × benariensis plants. (A) Maximum leaf length. (B) Maximum leaf width. Different letters indicate that the parameter values are significantly different at the level of p < 0.05.
Figure 7.
Comparison of leaf morphology of doubled Begonia × benariensis plants. (A) Maximum leaf length. (B) Maximum leaf width. Different letters indicate that the parameter values are significantly different at the level of p < 0.05.

Figure 8.
Analysis of chlorophyll content in doubled Begonia × benariensis plants. (A) Chlorophyll a content. (B) Chlorophyll b content. (C) Total chlorophyll content. Different letters indicate that the parameter values are significantly different at the level of p < 0.05.
Figure 8.
Analysis of chlorophyll content in doubled Begonia × benariensis plants. (A) Chlorophyll a content. (B) Chlorophyll b content. (C) Total chlorophyll content. Different letters indicate that the parameter values are significantly different at the level of p < 0.05.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Induction of Begonia × benariensis callus using colchicine immersion method.
| Treatment Concentration (%) | Treatment Time (h) | Treatment Quantity | Survival Number | Survival Rate (%) | Mutagenic Number | Mutagenic Rate (%) |
|---|---|---|---|---|---|---|
| 0 | 2 | 60 | 48 | 80.00% | 0 | 0% |
| 4 | 60 | 44 | 73.33% | 0 | 0% | |
| 8 | 60 | 43 | 71.67% | 0 | 0% | |
| 0.01% | 2 | 60 | 41 | 68.33% | 0 | 0% |
| 4 | 60 | 39 | 65.00% | 2 | 3.33% | |
| 8 | 60 | 37 | 61.67% | 0 | 0% | |
| 0.05% | 2 | 60 | 38 | 63.33% | 4 | 6.67% |
| 4 | 60 | 36 | 60.00% | 28 | 46.67% | |
| 8 | 60 | 26 | 43.33% | 0 | 0% | |
| 0.1% | 2 | 60 | 25 | 41.67% | 1 | 1.67% |
| 4 | 60 | 19 | 31.67% | 0 | 0% | |
| 8 | 60 | 11 | 18.33% | 0 | 0% |
Table 2.
Plant performance of tissue cultured seedlings at the early stage of mutagenesis.
| Ploidy | Growth State of Seedlings | Leaf Character |
|---|---|---|
| CK | Normal | Thin, oily, smooth, larger, bright green leaves |
| Variant plant | Stocky and slow growing | Leaf blades thickened and curled, leaf color deepened, leaf blades smaller and darker green |
Note: CK (control check) represents that plants are not treated with colchicine.
Table 3.
Plant growth performance after transplantation.
| Ploidy | Leaf Character | Flower Character |
|---|---|---|
| CK | Thin, oily, smooth, larger, bright green leaves | Thinner and smaller petals |
| Variant plant | Leaf blades thickened and curled, leaf color deepened, leaf blades smaller and darker green | Thickened and enlarged petals, more reddish flower color |
Note: CK (control check) represents that plants are not treated with colchicine.
Table 4.
Comparison of stomata in double plant of Begonia × benariensis.
| Ploidy | Defense Cell Length (µm) | Defense Cell Width (µm) | Stomatal Density (No./mm2) |
|---|---|---|---|
| CK | 58.98 ± 5.87 b | 14.35 ± 2.70 b | 112.37 ± 12.89 a |
| Triploid | 63.12 ± 4.78 b | 14.39 ± 1.10 b | 102.67 ± 11.39 a |
| Tetraploid | 85.42 ± 8.48 a | 20.89 ± 2.44 a | 67.11 ± 6.21 b |
Note: Different letters indicate that the parameter values are significantly different at the level of p < 0.05. The same below.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xie, N.; Zhao, Y.; Huang, M.; Chen, C.; Cao, C.; Wang, J.; Shi, Z.; Gao, J. Polyploid Induction and Identification of Begonia × benariensis. Horticulturae 2024, 10, 47. https://doi.org/10.3390/horticulturae10010047
AMA Style
Xie N, Zhao Y, Huang M, Chen C, Cao C, Wang J, Shi Z, Gao J. Polyploid Induction and Identification of Begonia × benariensis. Horticulturae. 2024; 10(1):47. https://doi.org/10.3390/horticulturae10010047
Chicago/Turabian StyleXie, Ninghao, Yi Zhao, Min Huang, Caixia Chen, Chuanqu Cao, Jisheng Wang, Zhihua Shi, and Junshan Gao. 2024. "Polyploid Induction and Identification of Begonia × benariensis" Horticulturae 10, no. 1: 47. https://doi.org/10.3390/horticulturae10010047
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.