
Age and Growth of Diaphus brachycephalus in the South China Sea Using Sagittal Otolith Microstructure
by
 , ,
, ,
Han Tian
1,2 ,
,
Yane Jiang
1,3, 
Jun Zhang
1,3,
Shannan Xu
1,3,
Zuozhi Chen
1,3,* and
Jiangfeng Zhu
2 1
South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Laboratory for Sustainable Utilization of Open-Sea Fishery, Ministry of Agriculture and Rural Affairs, Guangzhou 510300, China
2
College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China
3
Southern Marine Science and Engineering Guangdong Laboratory, Guangzhou 511548, China
*
Author to whom correspondence should be addressed.
Fishes 2022, 7(4), 169; https://doi.org/10.3390/fishes7040169
Submission received: 14 June 2022
/
Revised: 11 July 2022
/
Accepted: 12 July 2022
/
Published: 14 July 2022
(This article belongs to the Section Biology and Ecology)
Abstract
:To obtain the growth and age characteristics of Diaphus brachycephalus in the South China Sea (SCS), specimens of D. brachycephalus were collected by a mid-water trawl in January and June 2015. The relationship between standard length (L) and body weight (W) was W = 0.00002699L2.8789, with no significant differences between the two survey stations and the sexes. Microscopic observation of the sagittal otoliths revealed that the daily growth increments in each period were divided into the central zone, middle zone, and external zone. The age was determined by summing the daily growth increments of the three areas. The von Bertalanffy growth curve fitted to the relationship between age and L was shown as: L = 65.6[1 − exp{−0.0132(t − 6.94)}], r2 = 0.935. The growth rate decreased from juvenile to adult, with a maximum rate of 0.436 mm day−1. The back-calculated hatching dates of specimens were speculated to be from March to November, but predominantly occurred from April to May and from September to October.
1. Introduction
The South China Sea (SCS), which covers an area of approximately 3.5 million km2 and has an average depth of 1200 m, is the largest semi-enclosed marginal sea in the Pacific Ocean [1,2,3]. Enormous fishery resources in the SCS provide coastal countries and regions with a core fishing economy. However, the fishery supply capacity in the SCS has been declining because of excessive fishing capacity. The exploitation of mesopelagic fish resources in the SCS is still in its infancy but has great potential for future development [4,5,6].
Mesopelagic fishes live in the mesopelagic zone (200–1000 m). They have an estimated resource of 5–15 billion tons and are considered the most abundant vertebrates in the biosphere [7,8,9]. As one of the most dominant groups of mesopelagic fish, lanternfish belong to Myctophiformes; there are 348 species in 34 genera in this family, including the most common and most abundant mesopelagic fish species. They account for at least 20% of total marine fish species [10,11]. Diel vertical migration (DVM) is a well-known habit common to most lanternfish species. Lanternfish establish temporal, spatial, and biological connections between multiple water layers and act as a “biological pump” to facilitate the input of carbon into the deep ocean through DVM [8,12,13]. These fishes also act as a trophic link between zooplankton and top predators, such as seabirds, marine mammals, and other fishes. Both contribute to the energy flow and material cycling of the deep-sea food web and maintain the stability of deep-sea ecosystems [14,15,16].
The life span of lanternfish is short, usually 1–5 years, but they have a rapid rate of regeneration, which is an important reason that they have received attention as a fish resource [17,18]. However, the lack of age and growth information for many mesopelagic fish species limits our knowledge and exploitation of this resource. Their age and growth can be reflected by the otolith microstructure that originates from a primordium, which is the first calcified tissue during embryonic development of osteichthyans [19,20]. Otolith microstructure reveals fish growth in the form of daily annuli formed by the differential deposits of calcium carbonate and protein. Sagittal otoliths are the largest among the three pairs of otoliths; they have more observable structures and therefore are more widely used in fish age growth research [21,22,23]. Moreover, sagittal otoliths are also used to study other biological and ecological features of lanternfish, such as DVM, early life history, and reproduction [22,24,25,26].
As the largest genus of the family, the Myctophidae genus Diaphus contains 77 species, which accounts for more than one-third of the species in Myctophidae [27,28,29,30]. Diaphus brachycephalus, also known as short-headed lanternfish, is widely distributed in tropical and subtropical waters around the world, including the SCS [11,31,32,33,34]. Several surveys conducted in different areas of the SCS had reported the presence of D. brachycephalus and indicated that the dominant distribution of this species is in the SCS. However, research on D. brachycephalus in the SCS is still in its infancy; although some studies have investigated their distribution and DVM, research on their age and growth characteristics is still lacking [34,35,36,37]. In this study, the relationship between age and growth was investigated through the standard length and sagittal otolith microstructure to estimate the growth pattern and biology of D. brachycephalus in the SCS. Thus, it provides a useful reference for further study of D. brachycephalus and scientific guidance for the exploitation of mesopelagic fish in the SCS.
2. Materials and Methods
Specimens of D. brachycephalus were collected from two survey stations in the SCS in January and June 2015 by the R/V Nanfeng (1537 t GT, 66.7 m length, 12.4 m width, and 4.8 m draught) and a mid-water trawl (a cod-end mesh size of 10 mm, estimated mouth opening of approximately 128 m2) with a PI44 monitoring system (Kongsberg Maritime Inc., Kongsberg, Norway) to ensure that the trawl depths remained relatively constant at a 75 m water depth (Table 1). To prevent these fragile specimens from being destroyed, specimens were collected on deck and gently cleaned to remove impurities from the surface of the body. The above steps were completed as soon as possible. Then, specimens were immersed in seawater and temporarily stored at a temperature of −40 °C [38]. Biological characteristics were measured and recorded in an onshore laboratory, which included standard length (L, mm) from the snout tip to the last vertebra (to the nearest 0.1 mm) and body weight (W, g) to the nearest 0.01 g. Sex was determined by macroscopic observation of gonads [24].
Sagittal otoliths were extracted, numbered, and stored in 1.5 mL tubes filled with 75% alcohol to clean the soft tissue and organic material from the surface of the otoliths (Figure 1). The cleaned otoliths were dried for 1 h at 60 °C and then weighed to two decimal places in milligrams. After gluing and fixing with epoxy resin and hardener (EpoHeat, Buehler, Lake Bluff, IL, USA), the otoliths were longitudinally ground by a MetaServ 250 grinding machine (Buehler, Lake Bluff, IL, USA) with 280- and 800-grit sandpaper. They were further polished on top of its viewing surface using 1200- and 2500-grit sandpaper to obtain clear longitudinal sections [23,39,40].
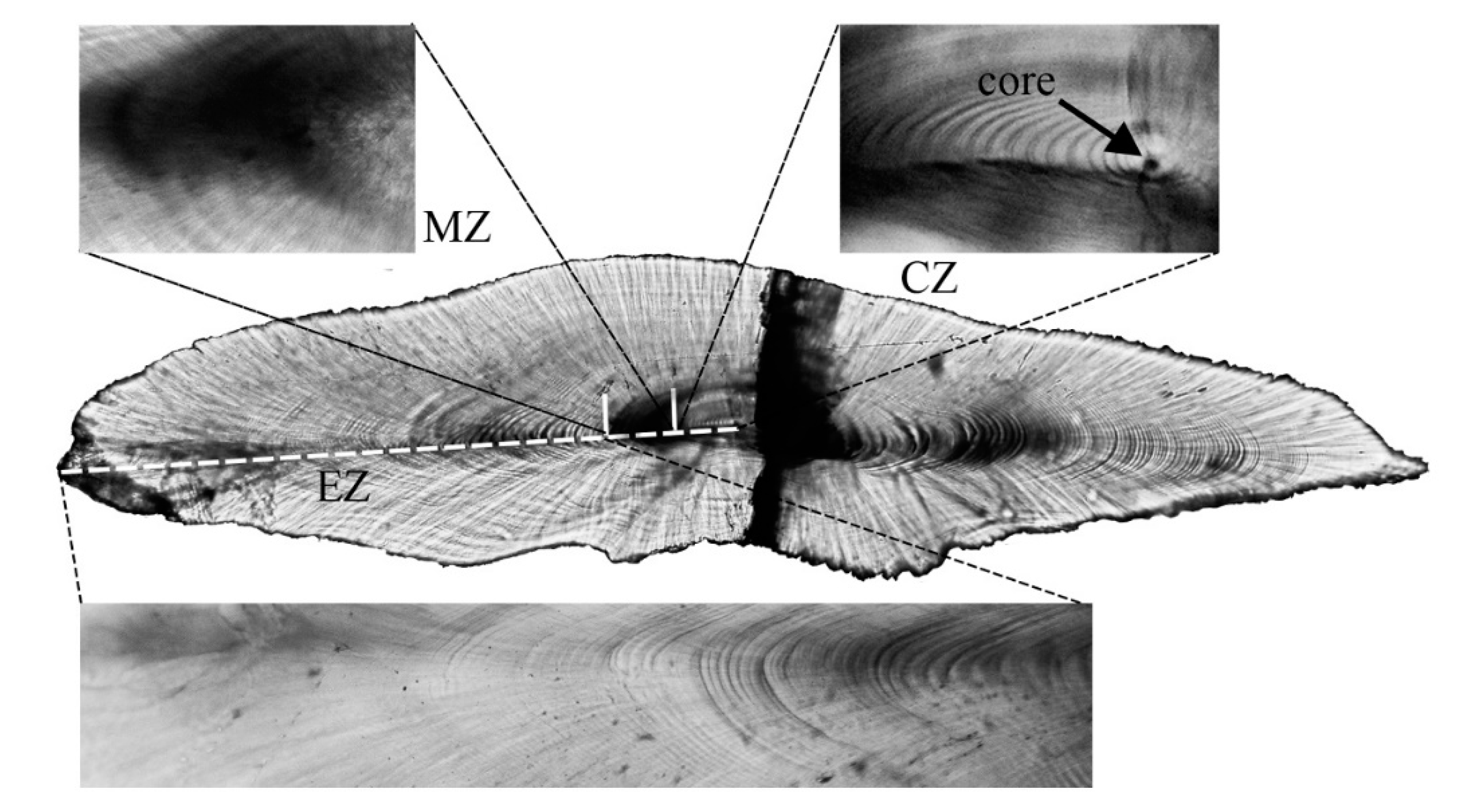
Sagittal otoliths were divided into three regions: the central zone (CZ), middle zone (MZ), and external zone (EZ), as proposed by Gigarosov and cited in subsequent studies [23,41,42,43]. CZ is a bright area that covers the primordium and provides a clearer record of daily growth increments during the larval stage. Among the three regions, pigmentation is most pronounced in the MZ region, such that the otoliths form a white, black, and white sandwich structure between the three regions. The reason for this phenomenon may be related to the change in the perching water layer caused by the metamorphosis of these fish. EZ records the incremental characterization of fish from the completion of metamorphosis until they are harvested, and is usually the area with the clearest, densest, and most abundant incremental characterization. Daily growth increments were counted at least three times and ended with a difference of less than 5% between the average count and each count, unless it needed to be recounted. The above counting experiment was conducted by one person using a light microscope, and the mean value was taken as the final value for analysis. Both growth increments counted in regions of CZ and MZ were extracted and used to obtain biological information at metamorphosis. The summaries of CZ, MZ, and EZ were used for age analysis [20,23,42,43].
Differences between the two regions and between the sexes were tested by one-way ANOVAs. The interaction between the two regions and sexes was also tested. Additionally, the biological characteristics of D. brachycephalus were discussed based on the different results of analyses. The relationship between standard length (L) and body weight (W) was described by the following equation [20,44]:
where L is standard length, W is body weight, and a and b are regression coefficients [45]. The relationship between daily growth increments of CZ and MZ is shown below [22,23]:
where NCZ is the daily growth increment of CZ and NMZ is the daily growth increment of MZ. C and d are regression coefficients. In addition, the daily growth increment ratio of CZ and MZ was calculated to determine the proportion of daily growth between the egg and metamorphic stage. The growth model selection was based on the von Bertalanffy (VB) growth curve, which is expressed as follows [22,23]:
where Lt is the standard length at age t, L∞ is theoretical asymptotic length, k is the growth coefficient, and t0 is the theoretical age when L = 0. The calculation of the above parameters was obtained by maximum likelihood estimation. The growth rate curve was transformed from the VB growth curve [23]:
where dL/dt is the growth rate. The data were analyzed and plotted using Excel 2019, SPSS 21.0, and Origin 9.9.0.225.
W = aLb,
NCZ = cNMZ + d,
Lt = L∞[1 − exp{−k(t − t0)}],
dL/dt = L∞ × k × exp[−k(t − t0)],
3. Results
A total of 87 specimens of D. brachycephalus were collected, of which 46 specimens (9 males and 37 females) were collected in January 2015, and a total of 41 specimens (7 males and 34 females) were collected in June 2015.
3.1. Body Length and Weight
The standard length and body weight of the D. brachycephalus specimens had no significant differences (P > 0.05) between the two regions and between the sexes, as tested by one-way ANOVAs. The interaction between the two regions and sexes was not significant (P > 0.05). Therefore, the male and female specimens of the two regions were analyzed as a whole. Means ± SD for the regions and sexes are shown in Table 2.
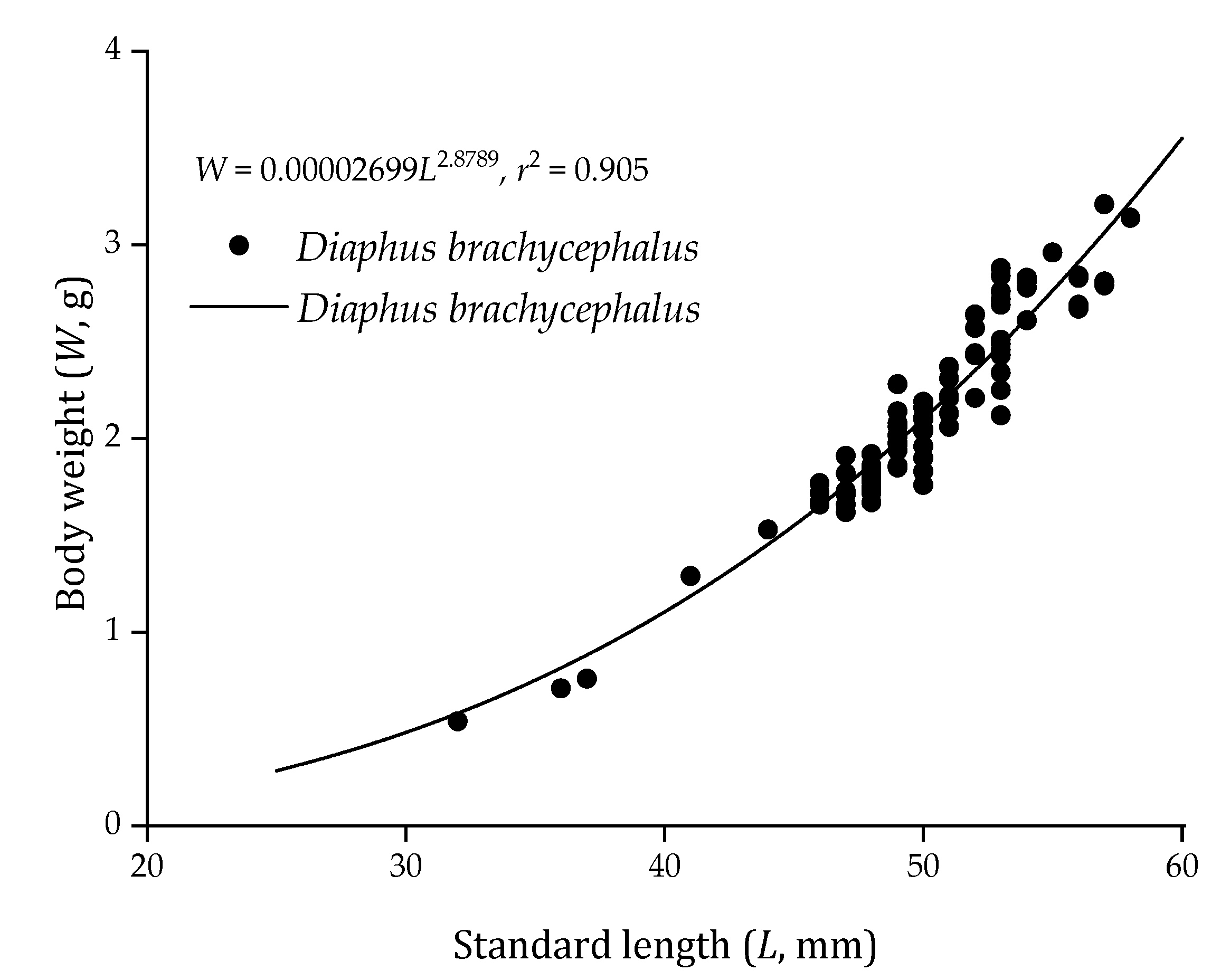
All specimens (87 in total) were used to describe the relationship between L and W (males, 16; females, 71). L of D. brachycephalus in the SCS (n = 87) ranged 32.0–58.0 mm, mean 50.0 ± 4.3 mm, and W of specimens ranged 0.71–3.21 g, mean 2.14 ± 0.51 g. The L–W relationship (Figure 2) of D. brachycephalus was as follows:
W = 0.00002699L2.8789, r2 = 0.905.
3.2. Sagittal Otolith Microstructure
In total, 73 pairs of clear longitudinal sections of the sagittal otoliths were collected because of losses during extraction and grinding. Females accounted for 59 sagittal otoliths, of which 30 were from January 2015 and 29 were from June 2015. Additionally, 7 sagittal otoliths from males were obtained from each region. The one on the right was selected for microscopic observation. Sagittal otolith microstructure of D. brachycephalus is composed of CZ, MZ, and EZ (Figure 3). These three zones were wrapped around from the inside out, which is visible through light micrographs. Moreover, a number of daily growth increments were found in MZ with abnormal dark colors and shapes. Daily growth increments of CZ, MZ, and EZ were counted for all 73 specimens. Additionally, the daily growth increments of CZ and MZ fluctuated within a relatively small range 16–26 (mean = 20.3 ± 2.6) and 12–20 (mean = 16.0 ± 1.9), respectively. The ratio of CZ/MZ was 1.27, and the relationship between daily growth increments of CZ and MZ was fitted as:
NCZ = 0.05029NMZ + 13.9574, r2 = 0.0714, P > 0.05.
Daily growth increments in EZ ranged 18–137. The results showed that daily growth increments of D. brachycephalus did not significantly differ between the regions or between the sexes (One-way ANOVA, P > 0.05).
3.3. Age and Growth
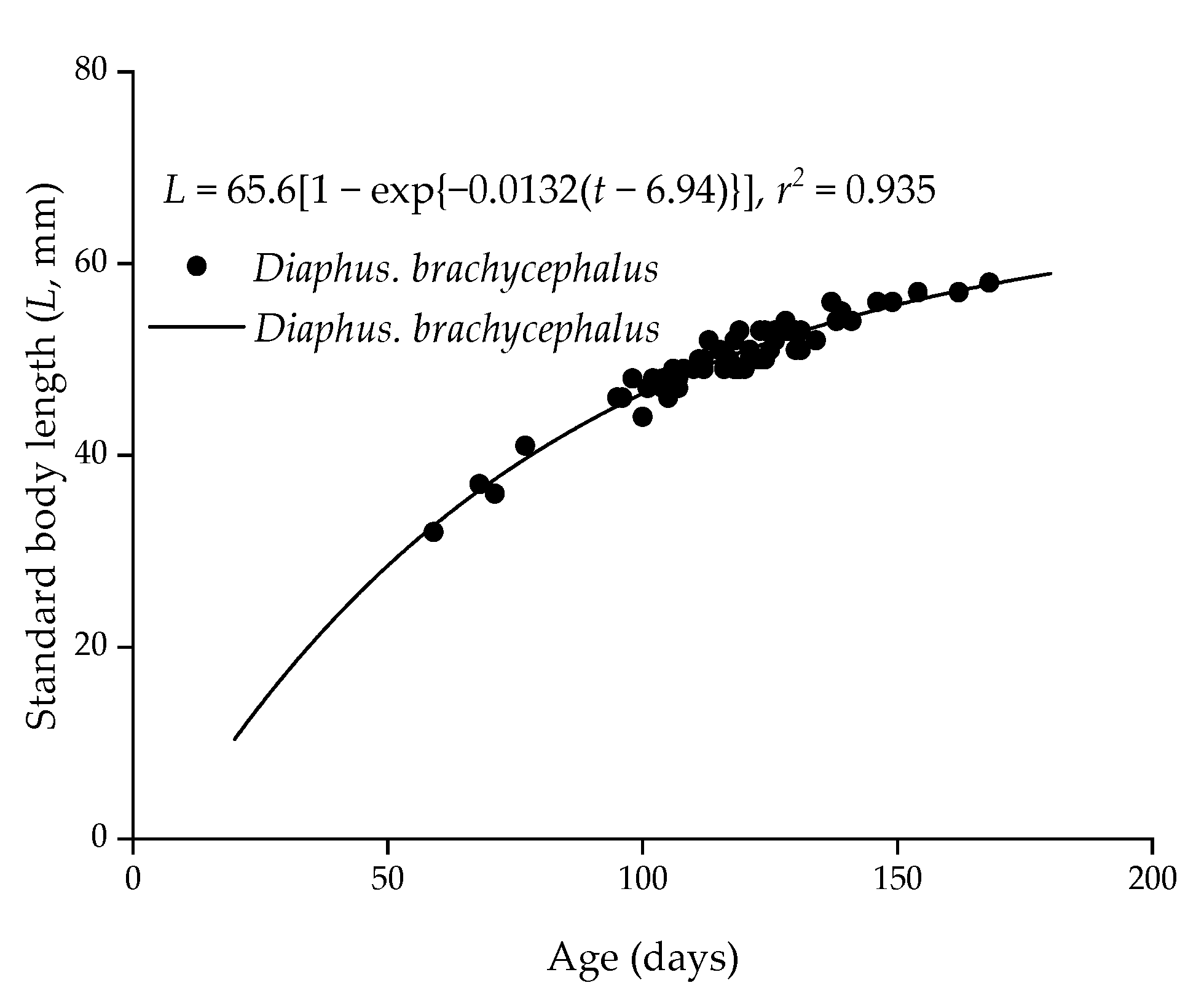
The ages of D. brachycephalus, ranging 59–168, were obtained by summing all of the daily growth increments in CZ, MZ, and EZ. There were non-significant differences between the two regions and sexes (one-way ANOVA, P > 0.05). Additionally, their interaction between the two regions and sexes was not significantly different (P > 0.05). Mean ages of males and females in January 2015 were 115 ± 27 days and 119 ± 25 days, respectively. The mean ages of males and females in June 2015 were 119 ± 15 days and 115 ± 13 days, respectively. Moreover, the interaction between the two regions and sexes was not significant (P > 0.05). The VB growth curve for D. brachycephalus was determined by fitting standard length and age (Figure 4):
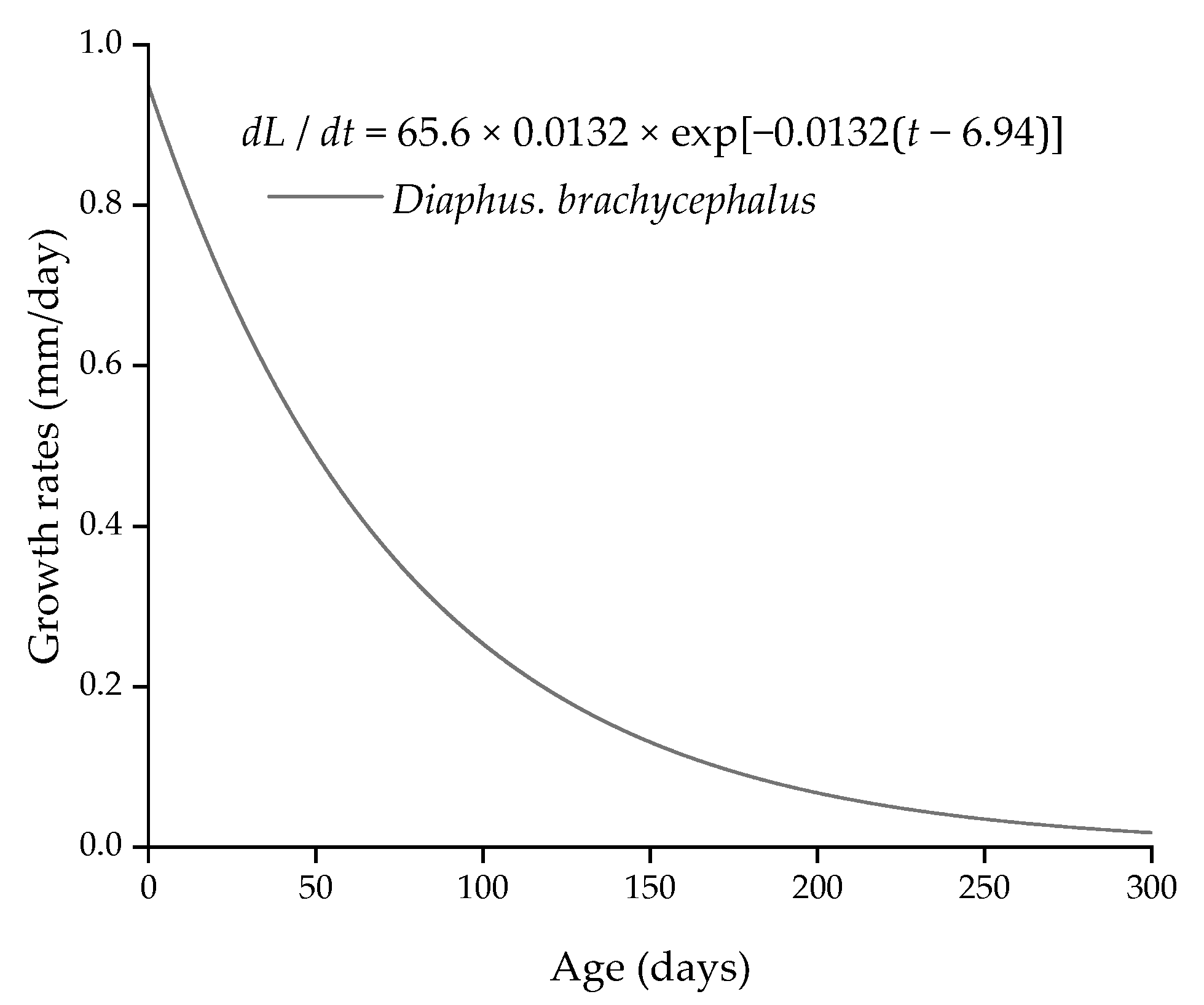
where L is standard length, t is the age in days (summary of daily growth increments in CZ, MZ, and EZ). Fitting the relationship between age and L, the growth rate curve was shown in (Figure 5):
L = 65.6[1 − exp{−0.0132(t − 6.94)}], r2 = 0.935,
dL/dt = 65.6 × 0.0132 × exp[−0.0132(t − 6.94)].
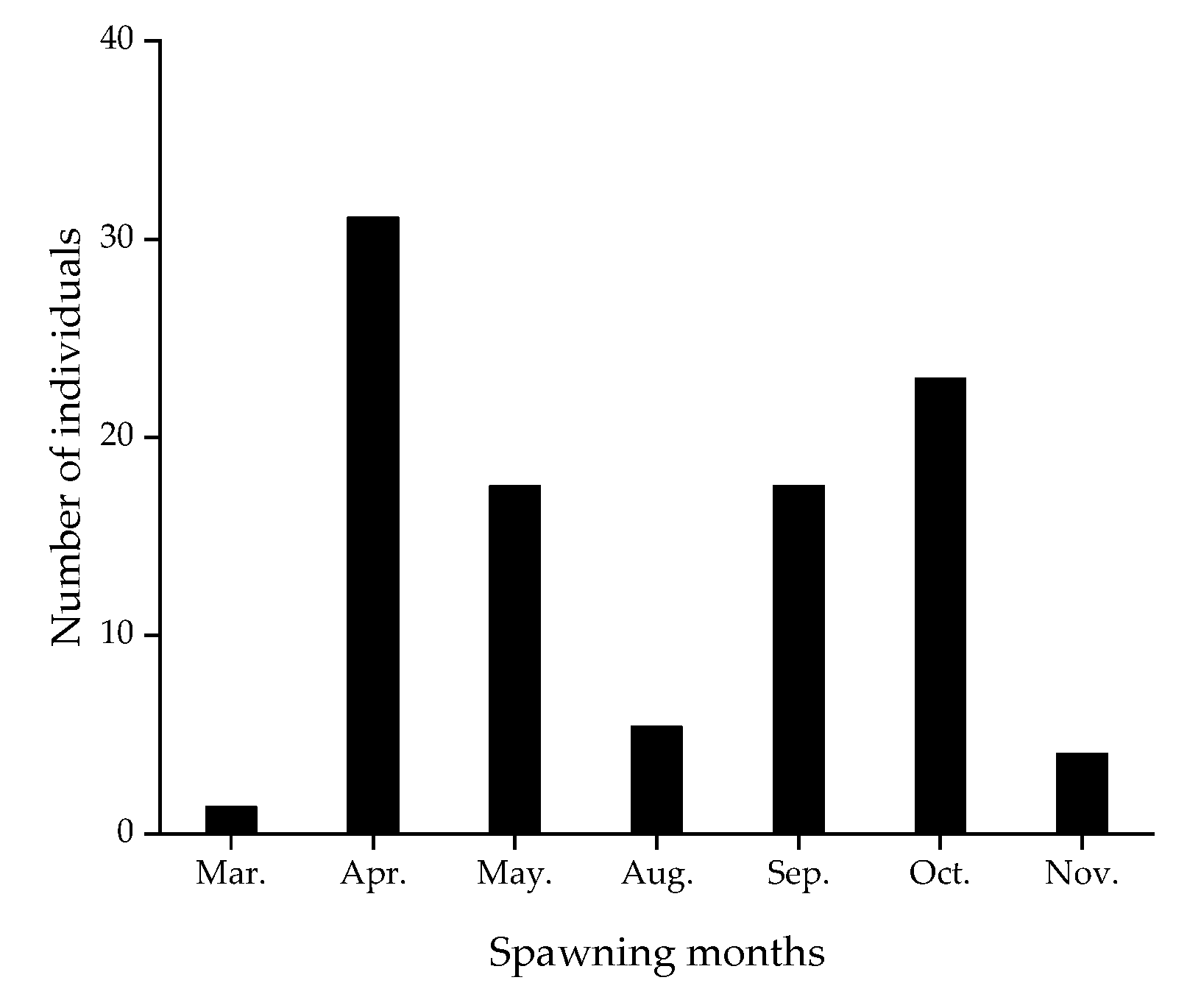
The maximum growth rate was 0.436 mm day−1 at the age of 59 days, when the standard length of D. brachycephalus was 32.0 mm and the body weight was 0.54 g. Based on age and date of capture, the back-calculated hatching dates of D. brachycephalus were distributed from March to May and from August to November, with two peak spawning periods concentrated from April to May and from September to October (Figure 6).
4. Discussion
In this study, there were more female than male D. brachycephalus specimens collected in the SCS, with a ratio of females to males of approximately 4.44. This is a trait thought to be common in many species of lanternfish; a large number of females would produce abundant eggs to guarantee a high population recruitment rate, such as in D. garmani, D. chrysorhynchus, and Myctophum asperum [17,22,46,47,48]. In our study, the range of standard length and body weight of D. brachycephalus (L, 32–58 mm and W, 0.71–3.21 g) were larger than the finding of Lopez-Perez in the Atlantic (L, 12–47 mm and W, 0.02–0.75 g) [49]. Additionally, there were neither sex nor regional differences (P > 0.05) in D. brachycephalus from the SCS. Gibbs also concluded that D. brachycephalus had no sexual dimorphism in body size [50]. Thus, based on the premise that there was no significant difference in body weight and sex, our collection could be used to analyze the relationship between L–W despite only including a few males. These specimens showed that D. brachycephalus had negative allometric growth, whereas most species of mesopelagic fish have isometric growth, such as Benthosema pterotum, Ceratoscopelus warmingii, Hygophum benoiti, and Hygophum hygomii [23,25,51].
Interestingly, our findings were opposite to those of Froese’s survey in the Atlantic [43]. The L–W relationship in the SCS and Atlantic was W = 0.000026999 × L2.8789 and W = 0.00000131 × L3.39, respectively, suggesting that D. brachycephalus had negative allometric growth in the SCS and positive allometric growth in the Atlantic. This phenomenon also occurred in M. punctatum from different regions, with the following relationships between L–W: W = 0.00000057 × L3.36 in the Atlantic and W = 0.000014 × L2.97 in the Mediterranean Sea [25,51]. The differences in the environment and temperature of the ocean may be the main reasons for the differences in the L–W relationships among lanternfish. By sorting out the biological characteristics of D. brachycephalus, we found that the length of juveniles of D. brachycephalus ranged 10–26 mm, subadults 25–40 mm, and adults were generally less than 61 mm [51,52]. Such evidence might explain the discrepancy between our results and those of Lopez-Perez; the Lopez-Perez study might have focused more on the subadult stage of D. brachycephalus and the influence of geographical differences [49].
CZ and MZ document lanternfish developmental processes during their juvenile years. CZ covers the embryonic development of a fertilized egg and ends with hatching. The formation of MZ is still uncertain, but it is generally believed that larval fishes migrate to deep waters where they complete their metamorphosis and thus become subadults [22,23,53]. In the absence of accurate descriptions of egg information for D. brachycephalus, we refer to spawning information for other species of mesopelagic fish in the genus Diaphus. The presence of hydrated oocytes signals that the fish are about to spawn; thus, in the absence of direct collection of eggs from water, the layer of water in which females with hydrated oocytes are collected can be considered the layer in which spawning takes place [46,54,55]. When the gonads of the specimens were examined, 32.4% of the females were found to have hydrated oocytes and the rest were in a state of ovarian development close to imminent spawning. Therefore, D. brachycephalus is likely to spawn in surface waters. Gibbs found D. brachycephalus larvae at depths from 150 m to 250 m, suggesting that MZ formation was influenced by metamorphosis and DVM [50].
In our study, CZ ranged 16–26 days (mean = 20.3 days). CZ varies between different species; for example, 33–43 days in Myctophum nitidulum, 80–139 days in Tarletonbeania crenularis, 22–32 days in B. pterotum, and 14–40 days in C. warmingii [23,41,42,43]. Additionally, MZ ranged 12–20 days (mean = 16 days), which was similar to Lampanyctodes hectoris (5–9 days), Notoscopelus resplendens (mean = 23 days), and M. asperum (mean = 10 days) [22,47,56]. MZ also varies considerably between lanternfish; for example, 51–102 days in Tarletonbeania crenularis and 24–54 days in C. warmingii [42,57]. In addition, there was no significant correlation between the daily increments of CZ and MZ in our study, and the ratio of the mean increments in the CZ/MZ of D. brachycephalus sagittal otoliths was 1.27. The same weak correlation between EZ and MZ was found in other studies, with the ratio between the two areas fluctuating around 1.00 [23,42,43]. For D. brachycephalus, the lower and more concentrated daily growth of CZ and MZ may be due to a combination of the marine environment in the SCS and the reproductive strategy of the population. That is, to maintain a relative balance in population dynamics, small individual fish of D. brachycephalus need to rapidly develop in preparation for entering the reproductive phase as soon as possible.
The VB growth curve objectively reflects the growth of D. brachycephalus. Age and standard length were used as parameters to fit the growth equation and study the growth rate of D. brachycephalus. The L∞ (65.6, mm) of the growth curve was larger than the maximum standard length (58 mm), whereas L∞ was greater than the currently known maximum standard length (61 mm) [52]. The VB growth curve objectively reflected the growth of D. brachycephalus, with r2 = 0.935. Based on the available D. brachycephalus, the growth rate of juveniles was faster than that of adults, with a maximum growth rate of 0.436 mm day−1, which was faster than other lanternfish, such as S. californiensis (0.31 mm day−1), M. asperum (0.3 mm day−1), and C. warmingii (0.35 mm day−1) [22,23,57,58]. However, benefiting from the distribution of specimens at various age stages from larval to adult, especially including the larval stage, Sarmiento-Lezcano argued that the otolith weight–age relationship of N. ressplendens was more in line with the Gompertz curve, whereas the relationship between standard length and age fitted well in both the Gompertz and the VB growth curve [56]. In the future, we need to know more about the distribution characteristics of D. brachycephalus to obtain larval specimens. The spawning of D. brachycephalus was mainly concentrated in April to May and September to October, suggesting that D. brachycephalus had at least two peak spawning periods in one year; however, this is based on the assumption of two surveys. We cannot rule out the possibility of spawning in other seasons, which will require investigation over a longer time span.
There are many gaps in the research on the microstructure of lanternfish otoliths that need to be addressed. In future studies, otolith preservation techniques will need to be fully considered, in addition to the integrity of the specimens at all age stages; a long-time span is required for such investigations. Alcohol is the most commonly used preservation solvent in otolith studies. Because of the narrow shape of the otoliths in D. brachycephalus, they would have fractured under the dehydrating effect of alcohol. Furthermore, Moku found that the preservation techniques of alcohol or formalin, which are popular at this stage, cause the otoliths to shrink and affect the assessment of age [59]. Therefore, the preservation method of otoliths needs to be further improved.
5. Conclusions
This study confirmed the age and growth of D. brachycephalus in the SCS for the first time by analysis of L, W, and sagittal otolith microstructure. The relationship between standard length (L) and body weight (W) was W = 0.00002699L2.8789, with non-significant sex differences. Moreover, D. brachycephalus was determined to have a short hatching period and metamorphosis period on the basis of the different areas of the sagittal otolith (CZ, MZ, EZ). The von Bertalanffy (VB) growth curves fitted to the relationship between age and L were shown as: L = 65.6[1 − exp{−0.0132 (t − 6.94)}], r2 = 0.935. We successfully obtained a faster growth rate during the juvenile period of D. brachycephalus as a result of the VB growth curve fitted with L and age, and the growth rate curve for L was dL/dt. The results indicated that D. brachycephalus is a short-lived small fish, and the microstructure of otoliths could well reflect the age and growth of D. brachycephalus.
Author Contributions
Conceptualization, H.T. and Y.J.; methodology, H.T., J.Z. (Jun Zhang) and Z.C.; software, J.Z. (Jun Zhang) and Z.C.; validation, Z.C., S.X., and J.Z. (Jiangfeng Zhu); formal analysis, H.T.; investigation, H.T. and Y.J.; resources, Y.J. and Z.C.; data curation, H.T. and Y.J. (Jun Zhang); writing—original draft preparation, H.T.; writing—review and editing, H.T., Y.J., Z.C., J.Z. (Jun Zhang), S.X., and J.Z. (Jiangfeng Zhu); visualization, H.T.; supervision, Z.C.; project administration, Z.C.; funding acquisition, Z.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Major Projects of Basic and Applied Basic Research Programs in Guangdong Province (No. 2019B030302004), Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou) (No. GML2019ZD0605), and Central Public-Interest Scientific Institution Basal Research Fund, CAFS (No. 2020TD05).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of the South China Sea Fisheries Research Institute Animal Welfare Committee (approval code: 2019/002 and approval date: 1 March 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the staff of R/V Nanfeng for collecting specimens. We are especially thankful to Yan Wang for his help with microscopic observation of otoliths. In addition, Yuqiong Song advised on writing terms and provided life companionship during the experiment and article preparation. We thank Mallory Eckstut for editing the English text of a draft of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Chen, C.T.A.; Wang, S.L.; Wang, B.J.; Pai, S.C. Nutrient budgets for the South China Sea basin. Mar. Chem. 2001, 75, 281–300. [Google Scholar] [CrossRef]
- Wong, G.T.F.; Ku, T.L.; Mulholland, M.; Tseng, C.M.; Wang, D.P. The South East Asian time-series study (SEATS) and the biogeochemistry of the South China Sea—An overview. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 1434–1447. [Google Scholar] [CrossRef]
- Yang, H.; Yizhuo, Z.; Huiwu, S. Development trends of marine capture fisheries in South China Sea. Agric. Outlook 2015, 11, 51–55, (In Chinese with English summary). [Google Scholar]
- Funge-Smith, S.; Briggs, M.; Miao, W. Regional Overview of Fisheries and Aquaculture in Asia and the Pacific 2012; RAP Publication 2012/26; Asia-Pacific Fishery Commission, FAO Regional Office for Asia and the Pacific: Bangkok, Thailand, 2012. [Google Scholar]
- Huang, H.W.; Chuang, C.T. Fishing capacity management in Taiwan: Experiences and prospects. Mar. Policy 2010, 34, 70–76. [Google Scholar] [CrossRef]
- Teh, L.S.L.; Witter, A.; Cheung, W.W.L.; Sumaila, U.R.; Yin, X. What is at stake? Status and threats to South China Sea marine fisheries. Ambio 2017, 46, 57–72. [Google Scholar] [CrossRef] [Green Version]
- Gjøsaeter, J.; Kawaguchi, K. A review of the world resources of mesopelagic fish. Food Agric. Organ. U. N. 1980, 193, 1–151. [Google Scholar]
- Klevjer, T.A.; Irigoien, X.; Røstad, A.; Fraile-Nuez, E.; Benítez-Barrios, V.M.; Kaartvedt, S. Large scale patterns in vertical distribution and behaviour of mesopelagic scattering layers. Sci. Rep. 2016, 6, 19873. [Google Scholar] [CrossRef] [Green Version]
- Proud, R.; Handegard, N.O.; Kloser, R.J.; Cox, M.J.; Brierley, A.S. From siphonophores to deep scattering layers: Uncertainty ranges for the estimation of global mesopelagic fish biomass. ICES J. Mar. Sci. 2018, 76, 718–733. [Google Scholar] [CrossRef] [Green Version]
- McGinnis, R.F. Biogeography of the lantern fishes (Myctophidae) south of 30° S. Antarctic Res. Ser. 1982, 35, 110. [Google Scholar] [CrossRef]
- Fishbase. Diaphus brachycephalus: List of Point Data. 2022. Available online: https://www.fishbase.se/map/OccurrenceMapList.php?genus=Diaphus&species=brachycephalus (accessed on 22 March 2022).
- Stanton, T.K.; Sellers, C.J.; Jech, J.M. Resonance classification of mixed assemblages of fish with swimbladders using a modified commercial broadband acoustic echosounder at 1–6 kHz. Can. J. Fish. Aquat. Sci. 2012, 69, 854–868. [Google Scholar] [CrossRef] [Green Version]
- Saunders, R.A.; Hill, S.L.; Tarling, G.A.; Murphy, E.J. Myctophid fish (family Myctophidae) are central consumers in the food web of the Scotia Sea (Southern Ocean). Front. Mar. Sci. 2019, 6, 530. [Google Scholar] [CrossRef] [Green Version]
- FAO Fisheries. Lanternfishes: A Potential Fishery in the Northern Arabian Sea. Roma, Italy, 1997. Available online: http://www.fao.org/docrep/003/w4248e/w4248e34.htm (accessed on 26 March 2022).
- Williams, A.; Koslow, J.A.; Terauds, A.; Haskard, K. Feeding ecology of five fishes from the mid-slope micronekton community off southern Tasmania, Australia. Mar. Biol. 2001, 139, 1177–1192. [Google Scholar] [CrossRef]
- Davison, P.C.; Checkley, D.M.; Koslow, J.A.; Barlow, J. Carbon export mediated by mesopelagic fishes in the northeast Pacific Ocean. Prog. Oceanogr. 2013, 116, 14–30. [Google Scholar] [CrossRef]
- Gjøsæter, J. Growth, production and reproduction of myctophid fish Benthosema glaciale from western Norway and adjacent seas. Fiskeridir. Skr. Ser. Havunders. 1981, 17, 79–108. [Google Scholar]
- Catul, V.; Gauns, M.; Karuppasamy, P.K. A review on mesopelagic fishes belonging to family myctophidae. Rev. Fish Biol. Fish. 2011, 21, 339–354. [Google Scholar] [CrossRef]
- Linkowski, T.B. Otolith microstructure and growth patterns during the early life history of lanternfishes (family Myctophidae). Can. J. Zool. 1991, 69, 1777–1792. [Google Scholar] [CrossRef]
- José, I.Z.; Claudia, A.B.; Mauricio, F.L. Larval growth, condition and fluctuating asymmetry in the otoliths of a mesopelagic fish in an area influenced by a large Patagonian glacier. Mar. Biol. Res. 2014, 10, 504–514. [Google Scholar] [CrossRef]
- Carlstrom, D. A crystallographic study of vertebrate otoliths. Biol. Bull. 1963, 125, 441–463. [Google Scholar] [CrossRef]
- Yan, W.; Jun, Z.; Zuozhi, C.; Yane, J.; Shannan, X.; Zhongyi, L.; Xinliang, W.; Yiping, Y.; Xianyong, Z.; Meng, Z. Age and growth of Myctophum asperum in the South China Sea based on otolith microstructure analysis (Article). Deep-Sea Res. Part II 2019, 167, 121–127. [Google Scholar] [CrossRef]
- Jun, Z.; Yan, W.; Zuozhi, C.; Yane, J.; Shannan, X. Age and growth of Ceratoscopelus warmingii (Myctophidae) in the South China Sea based on sagittal otolith microstructure. Mar. Biol. Res. 2021, 17, 733–743. [Google Scholar] [CrossRef]
- Gartner, J.V. Life histories of three species of lanternfishes (Pisces: Myctophidae) from the eastern Gulf of Mexico. Mar. Biol. 1991, 111, 21–27. [Google Scholar] [CrossRef]
- Battaglia, P.; Malara, D.; Romeo, T.; Andaloro, F. Relationships between otolith size and fish size in some mesopelagic and bathypelagic species from the Mediterranean Sea (Strait of Messina. Italy). Sci. Mar. 2010, 74, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Ying, X.; Jian, Y.; Tao, J.; Hongbo, L.; Xiaming, Z.; Jianhua, T. Early life history of the small yellow croaker (Larimichthys polyactis) in sandy ridges of the South Yellow Sea. Mar. Biol. Res. 2017, 13, 993–1002. [Google Scholar] [CrossRef]
- Becker, V.E. Miktofovye Ryby Mirovogo Okeana. In Myctophid Fishes of the World Ocean; Nauka: Moscow, Russia, 1983; pp. 1–246. [Google Scholar]
- Nafpaktitis, B.G.; Robertson, D.A.; Paxton, J.R. Four new species of the lanternfish genus Diaphus (Myctophidae) from the Indo-Pacific. N. Z. J. Mar. Freshw. Res. 1995, 29, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Schwarzhans, W.A. Comparative morphological study of the recent otoliths of the genera Diaphus, Idiolychnus and Lobianchia (Myctophidae). Palaeoichthyologica 2013, 13, 41–82. [Google Scholar]
- Prokofiev, A.M.; Emelyanova, O.R.; Orlov, A.M.; Orlova, S.Y. A New Species of Diaphus Associated with Seamounts of the Emperor Chain, North-Western Pacific Ocean (Teleostei: Myctophiformes: Myctophidae). J. Mar. Sci. Eng. 2022, 10, 65. [Google Scholar] [CrossRef]
- Bullis, H.R.; Thompson, J.R. Collections by the Exploratory Fishing Vessels Oregon, Silver Bay, Combat, and Pelican Made during 1956 to 1960 in the Southwestern North Atlantic; United States Fish and Wildlife Service Special Scientific Reports–Fisheries; U.S. Department of the Interior, Bureau of Commercial Fisheries: Washington, DC, USA, 1965; p. 130. [Google Scholar]
- Backus, R.H.; Craddock, J.E.; Haedrich, R.L.; Shores, D.L. Mesopelagic fishes and thermal fronts in the western Sargasso Sea. Mar. Biol. 1969, 3, 1–9. [Google Scholar] [CrossRef]
- Hulley, P.A.; Quero, J.C.; Hureau, J.C.; Karrer, C.; Post, A.; Saldanha, L. Myctophidae. Check-List of the Fishes of the Eastern Tropical Atlantic (CLOFETA); JNICT: Lisbon, Portugal; SEI: Paris, France; UNESCO: Paris, France, 1990; pp. 398–467. [Google Scholar]
- Wang, X.; Zhang, J.; Zhao, X.; Chen, Z.; Ying, Y.; Li, Z.; Xu, D.; Liu, Z.; Zhou, M. Vertical distribution and diel migration of mesopelagic fishes on the northern slope of the South China sea. Deep-Sea Res. Part II 2019, 167, 128–141. [Google Scholar] [CrossRef]
- Yuan, M.; Chen, Z.; Zhang, J.; Jiang, Y.; Tang, Y.; Xu, S. Community structure of mesopelagic fish species in northern slope of South China Sea. South China Fish. Sci. 2018, 14, 85–91, (In Chinese with English summary). [Google Scholar] [CrossRef]
- Han, T.; Yane, J.; Jun, Z.; Zuozhi, C.; Shannan, X.; Jiangfeng, Z. A preliminary study on the community structure of mesopelagic fish in the cold seep area of Xisha Islands. South China Fish. Sci. 2022. accepted. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Jiang, Y.; Chen, Z.; Zhao, X.; Gong, Y.; Ying, Y.; Li, Z.; Kong, X.; Chen, G.; et al. Species composition and biomass density of mesopelagic nekton of the South China Sea continental slope. Deep-Sea Res. Part II 2019, 167, 105–120. [Google Scholar] [CrossRef]
- Motomura, H. Fish Collection Building and Procedures Manual; Ishikawa, S., Ed.; The Kagoshima University Museum: Kagoshima, Japan; The Research Institute for Humanity and Nature: Kyoto, Japan, 2013; p. 70. ISBN 978-4-905464-01-3. [Google Scholar]
- Beamish, R.J.; Leask, K.D.; Ivanov, O.A.; Balanov, A.A.; Orlov, A.M.; Sinclair, B. The ecology, distribution, and abundance of midwater fishes of the subarctic pacific gyres. Prog. Oceanogr. 1999, 43, 399–442. [Google Scholar] [CrossRef]
- García-Seoane, E.; Meneses, I.; Silva, A. Microstructure of the otoliths of the glacier lanternfish, Benthosema glaciale. Mar. Freshw. Res. 2014, 66, 70–77. [Google Scholar] [CrossRef]
- Gigarosov, V.Y.; Ovcharov, O.P. Age and growth of the lantern fish Myctophum nitidulum (Myctophidae) from the tropical Atlantic. J. Ichthyol. 1992, 32, 34–42. [Google Scholar]
- Bystydzieńska, Z.E.; Phillips, A.J.; Linkowski, T.B. Larval stage duration, age and growth of blue lanternfish Tarletonbeania crenularis (Jordan and Gilbert, 1880) derived from otolith microstructure. Environ. Biol. Fishes 2010, 89, 493–503. [Google Scholar] [CrossRef] [Green Version]
- Hosseini-Shekarabi, S.P.; Valinassab, T.; Bystydzieńska, Z.; Linkowski, T. Age and growth of Benthosema pterotum (Alcock, 1890) (Myctophidae) in the Oman Sea. J. Appl. Ichthyol. 2015, 31, 51–56. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight-length relationships: History meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- King, M. Fisheries Biology: Assessment and Management; Blackwell Science: Oxford, UK, 1995; p. 341. [Google Scholar]
- Clarke, T.A. Sex-ratios and sexual differences in size among mesopelagic fishes from the central Pacific Ocean. Mar. Biol. 1983, 73, 203–209. [Google Scholar] [CrossRef]
- Young, J.W.; Bulman, C.M.; Blaber, S.J.M.; Wayte, S.E. Age and growth of the lanternfish Lampanyctodes hectoris (Myctophidae) from eastern Tasmania, Australia. Mar. Biol. 1988, 99, 569–576. [Google Scholar] [CrossRef]
- Sassa, C.; Tanaka, H.; Ohshimo, S. Comparative reproductive biology of three dominant myctophids of the genus Diaphus on the slope region of the East China Sea. Deep-Sea Res. Part I 2016, 115, 145–158. [Google Scholar] [CrossRef]
- Karuppasamy, P.K.; George, S.; Menon, N.G. Length-weight relationship of Benthosema pterotum (myctophid) in the deep scattering layer (DSL) of the eastern Arabian Sea. Indian J. Fish. 2008, 55, 301–303. [Google Scholar]
- Gibbs, R.H.; Krueger, W.H. Biology of Midwater Fishes of the Bermuda Ocean Acre; Smithsonian Contributions to Zoology: Washington, DC, USA, 1987; pp. 1–187. [Google Scholar] [CrossRef]
- Lopez-Perez, C.; Olivar, M.P.; Hulley, P.A.; Tuset, V.M. Length-weight relationships of mesopelagic fishes from the equatorial and tropical Atlantic waters: Influence of environment and body shape. J. Fish Biol. 2020, 96, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.A. Some Aspects of the Ecology of Lanternfishes (Myctophidae) in the Pacific Ocean near Hawaii. Fish Bull. 1973, 71, 401–434. [Google Scholar]
- Loeb, V.J. Vertical distribution and development of larval fishes in the North Pacific central gyre during summer. Fish Bull. 1980, 77, 777–793. [Google Scholar]
- Lisovenko, L.A.; Prut’ko, V.G. Reproductive biology of Diaphus suborbitalis (Myctophidae) in the equatorial part of the Indian Ocean. 2. Fecundity and reproductive potential. J. Ichthyol. 1987, 27, 1–12. [Google Scholar]
- Gjøsæter, J.; Tilseth, S. Spawning behavior, egg and larval development of the myctophid fish Benthosema pterotum. Mar. Biol. 1988, 98, 1–6. [Google Scholar] [CrossRef]
- Sarmiento-Lezcano, A.N.; Triay-Portell, R.; Castro, J.J.; Rubio-Rodríguez, U.G.; Pajuelo, J.G. Age-based life-history parameters of the mesopelagic fish Notoscopelus resplendens (Richardson, 1845) in the Central Eastern Atlantic. Fish Res. 2018, 204, 412–423. [Google Scholar] [CrossRef]
- Linkowski, T.B. Morphological Variation, Systematics and Speciation of the Ceratosco-pelus townsendi−C. warmingii Complex (Osteichthyes: Myctophidae) Based on the Studies on the Morphology and Microstructure of Otoliths; Morski Instytut Rybacki: Gdynia, Poland, 1997; (In Polish with English summary). [Google Scholar]
- Takagi, K.; Yatsu, A.; Moku, M.; Sassa, C. Age and growth of lanternfishes, Symbolophorus californiensis and Ceratoscopelus warmingii (Myctophidae) in the Kuroshio-Oyashio Transition zone. Ichthyol. Res. 2006, 53, 281–289. [Google Scholar] [CrossRef]
- Moku, M.; Ishimaru, K.; Kawaguchi, K. Growth of larval and juvenile Diaphus theta (Pisces: Myctophidae) in the transitional waters of the western North Pacific. Ichthyol. Res. 2001, 48, 385–390. [Google Scholar] [CrossRef]
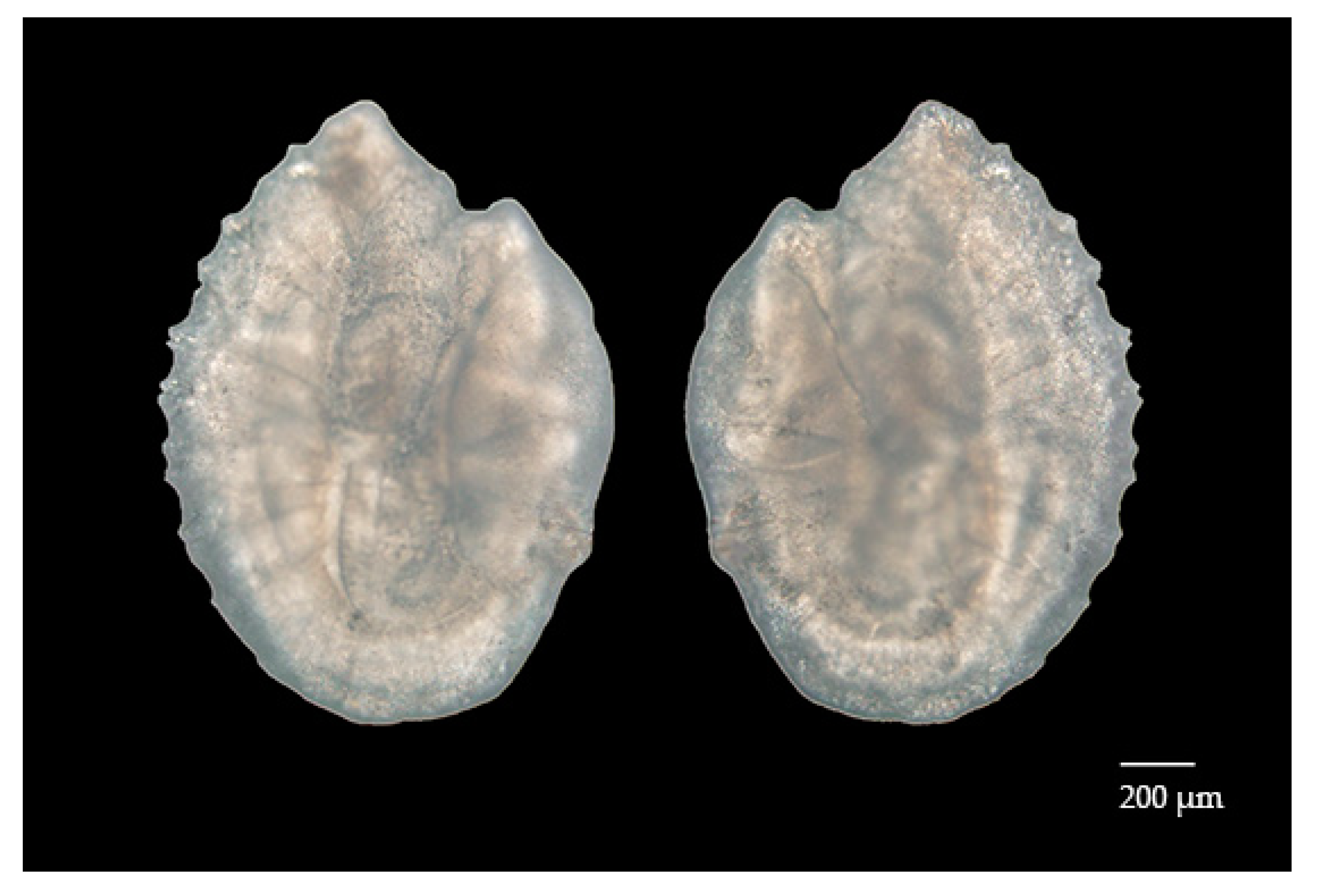
Figure 1.
Sagittal otolith of Diaphus brachycephalus (scale = 200 μm).

Figure 2.
Relationship between standard length and body weight of Diaphus brachycephalus (n = 87) in two regions of the South China Sea.
Figure 2.
Relationship between standard length and body weight of Diaphus brachycephalus (n = 87) in two regions of the South China Sea.

Figure 3.
Light micrographs of a sagittal otolith from Diaphus brachycephalus: central zone (CZ), middle zone (MZ), and external zone (EZ). The shaded area across the otolith on the right was a fracture caused by dehydration due to the action of alcohol when the otolith was preserved and did not affect the observation of daily growth increments.
Figure 3.
Light micrographs of a sagittal otolith from Diaphus brachycephalus: central zone (CZ), middle zone (MZ), and external zone (EZ). The shaded area across the otolith on the right was a fracture caused by dehydration due to the action of alcohol when the otolith was preserved and did not affect the observation of daily growth increments.

Figure 4.
The von Bertalanffy growth curve fitted to the relationship between age and standard length in Diaphus brachycephalus (n = 73) in the South China Sea.
Figure 4.
The von Bertalanffy growth curve fitted to the relationship between age and standard length in Diaphus brachycephalus (n = 73) in the South China Sea.

Figure 5.
Growth rate for standard length as dL/dt for Diaphus brachycephalus in the South China Sea.
Figure 5.
Growth rate for standard length as dL/dt for Diaphus brachycephalus in the South China Sea.

Figure 6.
Speculated spawning times of Diaphus brachycephalus specimens in the South China Sea.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sampling information of Diaphus brachycephalus in the South China Sea.
| Starting Location | Finishing Location | Trawl Depth | Mean Towing Speed | ||
|---|---|---|---|---|---|
| Sample Time | M/D/Y | Lat. (N)/Long. (E) | Lat. (N)/Long. (E) | (m) | (m/s) |
| 18:17~19:51 | 01/27/2015 | 15°21.45′/115°11.07′ | 15°19.93′/115°04.40′ | 75 | 2.3 |
| 21:30~22:30 | 06/20/2015 | 19°48.04′/115°48.90′ | 19°45.44′/115°51.90′ | 75 | 1.8 |
Table 2.
Standard length (L) and body weight (W) of Diaphus brachycephalus.
| M/D/Y | Sexes | L (mean ± SD, mm) | W (mean ± SD, g) | n |
|---|---|---|---|---|
| 01/27/2015 | male | 49.6 ± 6.2 | 1.98 ± 0.62 | 9 |
| 01/27/2015 | female | 50.4 ± 5.1 | 2.27 ± 0.57 | 37 |
| 06/20/2015 | male | 49.6 ± 3.5 | 2.15 ± 0.42 | 7 |
| 06/20/2015 | female | 49.7 ± 2.7 | 2.15 ± 0.42 | 34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tian, H.; Jiang, Y.; Zhang, J.; Xu, S.; Chen, Z.; Zhu, J. Age and Growth of Diaphus brachycephalus in the South China Sea Using Sagittal Otolith Microstructure. Fishes 2022, 7, 169. https://doi.org/10.3390/fishes7040169
AMA Style
Tian H, Jiang Y, Zhang J, Xu S, Chen Z, Zhu J. Age and Growth of Diaphus brachycephalus in the South China Sea Using Sagittal Otolith Microstructure. Fishes. 2022; 7(4):169. https://doi.org/10.3390/fishes7040169
Chicago/Turabian StyleTian, Han, Yane Jiang, Jun Zhang, Shannan Xu, Zuozhi Chen, and Jiangfeng Zhu. 2022. "Age and Growth of Diaphus brachycephalus in the South China Sea Using Sagittal Otolith Microstructure" Fishes 7, no. 4: 169. https://doi.org/10.3390/fishes7040169