
A New Species of Pareas Wagler, 1830 (Squamata, Pareidae) from Northwestern Yunnan, China †

1
Kunming Natural History Museum of Zoology, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming 650223, China
2
Agricultural Service Center of the People’s Government of Lurong Township, Zhenfeng 562203, China
3
Key Laboratory for Conserving Wildlife with Small Populations in Yunnan, Southwest Forestry University, Kunming 650233, China
4
College of Life Science, Leshan Normal University, Leshan 614000, China
5
Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming 650201, China
*
Authors to whom correspondence should be addressed.
†
urn:lsid:zoobank.org:act:8EEA5A22-7041-424A-A931-FCD465E1203A; urn:lsid:zoobank.org:pub:22483816-AC27-41FF-81E9-A0CB378BFA31.
Taxonomy 2023, 3(2), 169-182; https://doi.org/10.3390/taxonomy3020013
Submission received: 20 February 2023
/
Revised: 21 March 2023
/
Accepted: 24 March 2023
/
Published: 31 March 2023
Abstract
:A new species of Pareas Wagler, 1830 is described from Dulongjiang Township, Gongshan County, Yunnan Province, China. Phylogenetically, the new species is most closely related to Pareas kaduri Bhosale, Phansalkar, Sawant, Gowande, Patel and Mirza, 2020; however, the genetic divergence (uncorrected p-distance) of the cyt b gene between the new species and P. kaduri reached 9.4%. Morphologically, the new species can be distinguished from P. kaduri by the absence of preoculars, loreal bordering the orbit, the fusion of subocular and postocular, three rows enlarged vertebral scales, five rows keeled mid-dorsal scales at the middle of the body, having more subcaudals, and having a relatively longer tail. In addition, the new species can be distinguished from all other congeners by a combination of morphological characteristics. Our work brings the total number of recognized species of the genus Pareas to 29, of which 23 occur in China.
1. Introduction
The family Pareidae Romer, 1956 was once considered a subfamily (Pareatinae Romer, 1956) of Colubridae Oppel, 1811 [1,2,3], which encompasses two subfamilies (Pareinae and Xylophiinae Deepak, Ruane and Gower, 2019) and four genera (Aplopeltura Duméril, 1853; Asthenodipsas Peters, 1864; Pareas Wagler, 1830 [4]; and Xylophis Beddome, 1878) [5,6,7]. Poyarkov et al. [8] partitioned the genus Pareas into two subgenera (Pareas and Eberhardtia Angel, 1920).
Evolutionarily, although there is practically no fossil record for Pareinae [9], based on the calibration priors from Deepak et al. [5], Poyarkov et al. [8] inferred that Pareinae originated in Sundaland during the middle Eocene and the most recent common ancestor of Pareinae originated in late Eocene. Pareinae colonized mainland Asia in the early Oligocene, when the genera Asthenodipsas and Pareas started to diversify [8].
The snail-eating or slug-eating snake genus Pareas is a group of snakes that has undergone fast taxonomic changes which is mainly distributed in northeastern India, Myanmar, southern and eastern China, Indochina, and Sundaland [7,8]. Before 2000, there were only 11 recognized species in this genus, namely Pareas boulengeri (Angel, 1920), Pareas carinatus Wagler, 1830; Pareas chinensis (Barbour, 1912); Pareas formosensis (Van Denburgh, 1909); Pareas hamptoni (Boulenger, 1905); Pareas iwasakii (Maki, 1937); Pareas macularius Theobald, 1868; Pareas margaritophorus (Jan, 1866); Pareas monticola (Cantor, 1839); Pareas nuchalis (Boulenger, 1900); and Pareas stanleyi (Boulenger, 1914). Thereafter, P. macularius was synonymized with P. margaritophorus by Huang [10] but was revalidated by Hauser [11]; Guo and Deng [12] described Pareas nigriceps Guo and Deng, 2009 from western Yunnan, China; Vogel [13] described Pareas vindumi Vogel, 2015 from northern Myanmar; You et al. [14] described Pareas atayal You, Poyarkov and Lin, 2015 from Taiwan, China, and resurrected Pareas komaii (Maki, 1931); Ding et al. [15] described Pareas geminatus Ding, Chen, Suwannapoom, Nguyen, Poyarkov and Vogel, 2020 from southern Yunnan, China; Bhosale et al. [16] described Pareas kaduri Bhosale, Phansalkar, Sawant, Gowande, Patel and Mirza, 2020 from northeastern India; Vogel et al. [17] resurrected Pareas andersonii Boulenger, 1888 and Pareas modestus Theobald, 1868; Wang et al. [6] described Pareas menglaensis Wang, Che, Liu, Ki, Jin, Jiang, Shi and Guo, 2020 and Pareas mengziensis Wang, Che, Liu, Ki, Jin, Jiang, Shi and Guo, 2020 from southern and central Yunnan, China, respectively; Liu and Rao [18] described Pareas xuelinensis Liu and Rao, 2021 from western Yunnan, China, resurrected Pareas niger (Pope, 1928), and synonymized P. mengziensis with P. niger; Vogel et al. [19] described Pareas victorianus Vogel, Nguyen and Poyarkov, 2021 from western Chin State, Myanmar; Le et al. [20] described Pareas temporalis Le, Tran, Hoang and Stuart, 2021 from southern Vietnam; Poyarkov et al. [8] described Pareas abros Poyarkov, Nguyen, Pawangkhanant, Yushchenko, Brakels, Nguyen, Nguyen, Suwannapoom, Orlov and Vogel, 2022 and Pareas kuznetsovorum Poyarkov, Yushchenko and Nguyen, 2022 from Vietnam, resurrected Pareas berdmorei Theobald, 1868, and synonymized P. menglaensis with P. berdmorei; and Liu et al. [21] resurrected Pareas yunnanensis (Vogt, 1922) and described Pareas tigerinus Liu, Zhang, Poyarkov, Hou, Wu, Rao, Nguyen and Vogel, 2023 from southwestern Yunnan, China. To date, this genus contains 28 recognized species [21], of which more than one third were described in the past decade [7,21]. However, there are still undiscovered new species and unresolved taxonomic problems in this genus [8,21].
Dulongjiang, a township in Gongshan County, Yunnan Province, China, located in the China–Myanmar border area, which constitutes a long-acknowledged biodiversity hotspot [22,23]. However, previously there were few surveys in Dulongjiang because of its very inconvenient transportation conditions [24]. It was not until recent years that driveways were available there that more surveys were conducted there. However, there are still a lot of new species in Dulongjiang to be discovered.
During our fieldwork in northwestern Yunnan Province, China, in 2015, one specimen of the genus Pareas was collected from Dulongjiang Township, Gongshan County. Morphological comparison and molecular analysis indicate that the specimen belongs to a distinctive taxon and is separated from all other named species of the genus Pareas. Therefore, we describe this specimen as a new species herein.
2. Materials and Methods
The specimen of the new species was collected by hand at night and was euthanized using isoflurane, injected intraperitoneally with 95% ethanol, and then preserved in 75% ethanol. Liver tissue was collected for molecular analysis and stored in 99% ethanol. The specimen was deposited at Kunming Natural History Museum of Zoology, Kunming Institute of Zoology, Chinese Academy of Sciences (KIZ).
Measurements of the body and tail were taken to the nearest 1 mm with a measuring tape and measurements of the head were taken to the nearest 0.1 mm with a digital caliper. Paired meristic characters are given as left/right. The methodology of measurements and meristic counts followed those of Bhosale et al. [16] and Liu et al. [21]. Abbreviations used were as follows: ATem, number of anterior temporals; CL, cephalic length (measured from tip of snout to constriction of neck); DNB, dark nuchal band; DS, dorsal scale rows (counted at one head length behind the head, mid-body, and one head length before the vent); ED, eye diameter (widest diameter of the eye); El, eye to labial height (measured from lowest border of the eye to lower border of the labial); EN, eye to nares distance; ES, eye to snout length; HL, head length (measured from snout tip to the angle of the jaw); HW, head width (measured at the widest part of the head); InfL, infralabials; LoBO, loreal bordering orbit; Max, maxillary teeth; NED, number of enlarged dorsal scale rows at midbody; NKD, number of keeled dorsal scale rows at the middle of the body; NW, neck width (measured at the constriction of the neck); PosO, postoculars; Prec, precloacal plate; PreO, preoculars; PrFBO, prefrontal bordering orbit; PTem, number of posterior temporals; Sc, subcaudals; SPOF, subocular-postocular fused or not; SubO, suboculars; SupL, supralabials; SVL, snout-vent length (measured from tip of snout to posterior margin of cloacal plate); TaL, tail length (measured from posterior margin of cloacal plate to tip of tail); TL, total length; VBTr, vertical dark bars on trunk; Vs, ventrals. For comparison, data for other species were taken from the original and subsequent descriptions [2,6,8,11,12,13,14,15,16,17,18,19,20,21,25,26,27,28,29,30,31,32,33].
The sequence of the mitochondrial gene fragment cytochrome b (cyt b) was generated for the newly collected specimen. Homologous sequences were obtained from GenBank. The new sequence has been deposited in GenBank. Aplopeltura boa (Boie, 1828) was selected as the outgroup, based on the work of Liu et al. [21]. All the GenBank accession numbers for the taxa used in this study are listed in Table 1. Genomic DNA was extracted from liver tissue; the primers L14910/H16064 [34] were used for DNA sequencing. DNA extraction and amplification conditions followed those of Poyarkov et al. [8]. The amplification products were purified and sequenced by Tsingke Biotechnology Co., Ltd., Beijing, China. The sequence was edited and managed using SeqMan in Lasergene 7.1 (DNASTAR Inc., Madison, WI, USA) and MEGA 11 [35].
Sequences were aligned using ClustalW [36] integrated in MEGA 11 [35] with default parameters. Uncorrected pairwise genetic divergences (p-distance) were estimated using MEGA 11 [35]. The best substitution model (GTR + F + I + G4) for Bayesian inference and the best substitution model (GTR + F + R4) for maximum likelihood were selected using ModelFinder [37] in IQ-TREE 1.6.12 [38] under the Akaike Information Criterion (AIC). Bayesian inference was performed in MrBayes 3.2.7 [39]. Four Markov chains were run for 10,000,000 generations sampled every 1000 generations. The first 25% of the sampled trees were discarded as burn-in and the remaining trees were used to calculate Bayesian posterior probabilities. Maximum likelihood analysis was performed using IQ-TREE 1.6.12 [38]. Nodal support was estimated by 1000 ultrafast bootstrap replicates. Nodes with Bayesian posterior probabilities of 0.95 and above were considered strongly supported [40,41,42] and nodes with ultrafast bootstrap values of 95 and above were considered strongly supported [43].
3. Results
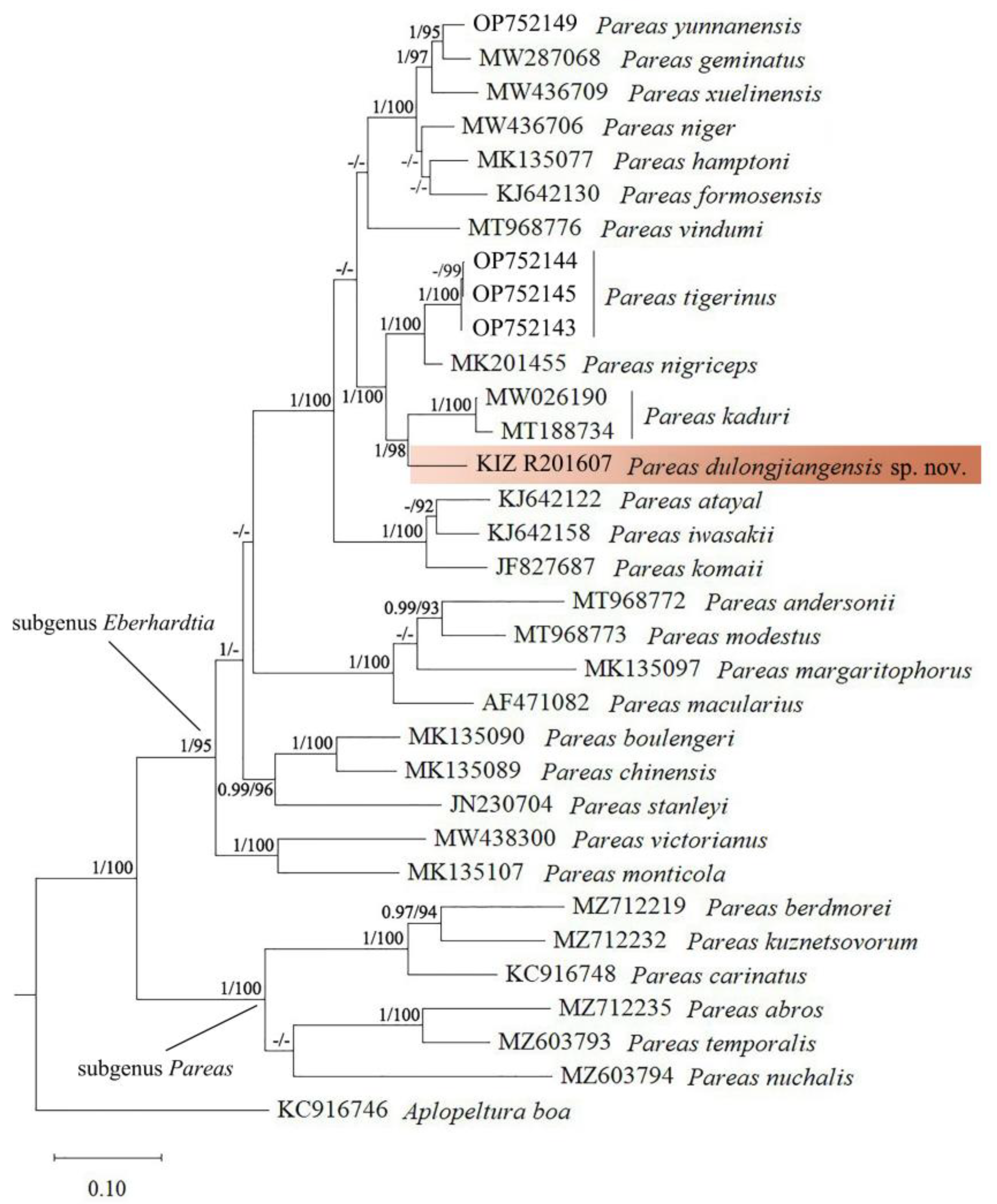
Maximum likelihood analyses and Bayesian inference showed identical topology; the newly collected specimen formed a distinct lineage which is sister to Pareas kaduri with strong support, which means that the newly collected specimen belongs to the subgenus Eberhardtia and is a member of the P. hamptoni species group [8] (Figure 1). The genetic divergence (uncorrected p-distance) between the newly collected specimen and P. kaduri was 9.4%, the genetic divergence (uncorrected p-distance) between the newly collected specimen and other congeners ranged from 10.3% to 25.0% (Table 2).
Class Reptilia Laurenti, 1768
Order Squamata Oppel, 1811
Suborder Serpentes Linnaeus, 1758
Infraorder Caenophidia Hoffstetter, 1939
Family Pareidae Romer, 1956
Subfamily Pareinae Romer, 1956
Genus Pareas Wagler, 1830
Subgenus Eberhardtia Angel, 1920
Pareas dulongjiangensis sp. nov.
urn:lsid:zoobank.org:act:8EEA5A22-7041-424A-A931-FCD465E1203A
Holotype. KIZ R201607, adult female, collected from Kongdang Village, Dulongjiang Township, Gongshan County, Nujiang Prefecture; Yunnan Province, China, on 9 September 2015 by Dingqi Rao (27°50′16″ N, 98°19′30″ E; 1460 m).
Diagnosis. Preocular absent; postocular fused with subocular; loreal bordering orbit; prefrontal bordering orbit; infralabials not fused with chin-shield; chin-shields three pairs, first pair triangular, second pair and third pair elongate; dorsal scales 15 rows throughout without apical pits; three rows vertebral scales enlarged; five rows mid-dorsal scales keeled at middle of body; supralabials6–7; infralabials7–9; precloacal plate undivided; ventral scales182; and subcaudals 76, paired. In preservative, dorsal surface of head chocolate brown, from which two brownish-black longitudinal stripes run on each side of the neck, leaving a pale central portion; lateral surface of head dense with brownish black dusting and two short brownish-black streaks, one on lower anterior temporal and another on last supralabial; almost no mottling on ventral surface of head; sparse brown mottling on ventral surfaces of body and tail; and indistinct vertical dark bars on trunk and tail.
Description. Adult female, SVL 373 mm, TL 115 mm, TaL/SVL 0.308, TaL/TL 0.236; body elongated, slightly compressed laterally; body vertebral ridge developed; head distinct from neck; snout wide and blunt, projecting beyond lower jaw. Rostral approximately as wide as high, not visible from above; nasals undivided; internasals wider than long; prefrontals bordering orbits; frontal anteriorly fused with prefrontals; parietals large, much longer than wide; loreal, single, large, approximately rectangular, broadly bordering orbit; preocular absent; supraocular single, slightly longer than wide; subocular and postocular fused into one thin elongated crescent-shaped scale; anterior temporals two on each side, posterior temporals three on left side and two on right side; supralabials six on left side and seven on right side, all separated from eye; infralabials seven on left side and nine on right side, anterior-most in contact with its opposite between mental and anterior chin-shields; infralabials not fused with chin-shield; chin-shields three pairs, first pair triangular, second pair and third pair elongate, and first pair larger than other two; ventral scales182; precloacal plate undivided; subcaudals 76, paired; dorsal scales 15 rows throughout without apical pits, three rows vertebral scales enlarged, and five rows mid-dorsal scales keeled at middle of body.
Coloration. In preservative over seven years, dorsal surface of head chocolate brown, from which two brownish-black longitudinal stripes run on each side of neck leaving a pale central portion; dorsal surface of body brownish-yellow, and approximately 52 indistinct, vertical, slightly billowing dark bars on trunk, and dark bars on tail more indistinct or invisible; lateral surface of head dense with brownish-black dusting and two short brownish-black streaks, one on lower anterior temporal and another on last supralabial; ventral surface of head light yellow with almost no mottling; and ventral surfaces of body and tail light yellow with sparse brown mottling. Coloration in life unknown.
Comparison. Pareas dulongjiangensis sp. nov. can be distinguished from P. andersonii, P. modestus, P. macularius, and P. margaritophorus by the yellow body background color (vs. body background colors being grey, dark grey, brownish-grey, or completely black).
Pareas dulongjiangensis sp. nov. can be distinguished from P. abros, P. atayal, P. berdmorei, P. carinatus, P. iwasakii, P. komaii, P. kuznetsovorum, P. nuchalis, and P. temporalis by the fusion of subocular and postocular into one thin elongated crescent-shaped scale (vs. subocular and postocular not being fused).
Pareas dulongjiangensis sp. nov. can be distinguished from P. boulengeri and P. stanleyi by the three rows enlarged vertebral scales (vs. vertebral scales not being enlarged).
Pareas dulongjiangensis sp. nov. can be distinguished from P. chinensis, P. formosensis, P. geminatus, P. hamptoni, P. monticola, P. niger, P. vindumi, P. xuelinensis, and P. yunnanensis by the absence of preoculars (vs. preoculars being present).
Pareas dulongjiangensis sp. nov. can be distinguished from P. victorianus by the three rows enlarged vertebral scales (vs. one row vertebral scales being enlarged), having more ventral scales (182 vs. 164), more subcaudals (76 vs. 58), and a relatively longer tail (TaL/SVL 0.308 vs. 0.265).
Pareas dulongjiangensis sp. nov. can be distinguished from P. nigriceps by the absence of preoculars (vs. preoculars being present), loreal bordering the orbit (vs. not bordering), three rows enlarged vertebral scales (vs. one row vertebral scales being enlarged), and having more ventral scales (182 vs. 175).
Pareas dulongjiangensis sp. nov. can be distinguished from P. tigerinus by the absence of preoculars (vs. preoculars being present), loreal bordering the orbit (vs. not bordering), three rows enlarged vertebral scales (vs. one row vertebral scales being enlarged), having more ventral scales (182 vs. 160–171), more subcaudals (76 vs. 62–64), and a relatively longer tail (TaL/SVL 0.308 vs. 0.25–0.26).
Pareas dulongjiangensis sp. nov. is most similar to P. kaduri in morphological characteristics and coloration; however, the new species can be distinguished from P. kaduri by the absence of preoculars (vs. preoculars being present), loreal bordering the orbit (vs. not bordering), the fusion of subocular and postocular (vs. the presence of one subocular and two postoculars), three rows enlarged vertebral scales (vs. one row vertebral scales being enlarged), five rows keeled mid-dorsal scales at the middle of the body in the single female (vs. mid-dorsal scales at the middle of the body being smooth in the single female), having more subcaudals (76 vs. 52–70), and having a relatively longer tail (TaL/SVL 0.308 vs. 0.226–0.262) (see Table 3).
Etymology. The specific epithet dulongjiangensis refers to the Dulongjiang Township, where the new species was discovered. We propose “Dulongjiang Slug-eating Snake” for the common English name and “独龙江钝头蛇” (Pinyin: dú lóng jiāng dùn tóu shé) for the common Chinese name of this species.
Distribution. The new species is currently known only from its type locality in Dulongjiang Township, Gongshan County, Nujiang Prefecture, Yunnan Province, China.
Natural history notes. The specimen of the new species was found at night on the road near a small village by the Dandangluo River, which is a tributary of the Dulongjiang River; there are some rocks, grass, and plantains along the road (Figure 4). More ecological information about this species is unknown. Other herpetological species observed near the type locality of the new species include Elaphe carinata (Günther, 1864); Nanorana chayuensis (Ye, 1977); Odorrana dulongensis Liu, Che & Yuan, 2021; Ovophis zayuensis (Jiang, 1977); Raorchestes dulongensis Wu, Liu, Gao, Wang, Li, Zhou, Yuan & Che, 2021; Rhabdophis leonardi (Wall, 1923); and Zhangixalus burmanus (Andersson, 1939).
4. Discussion
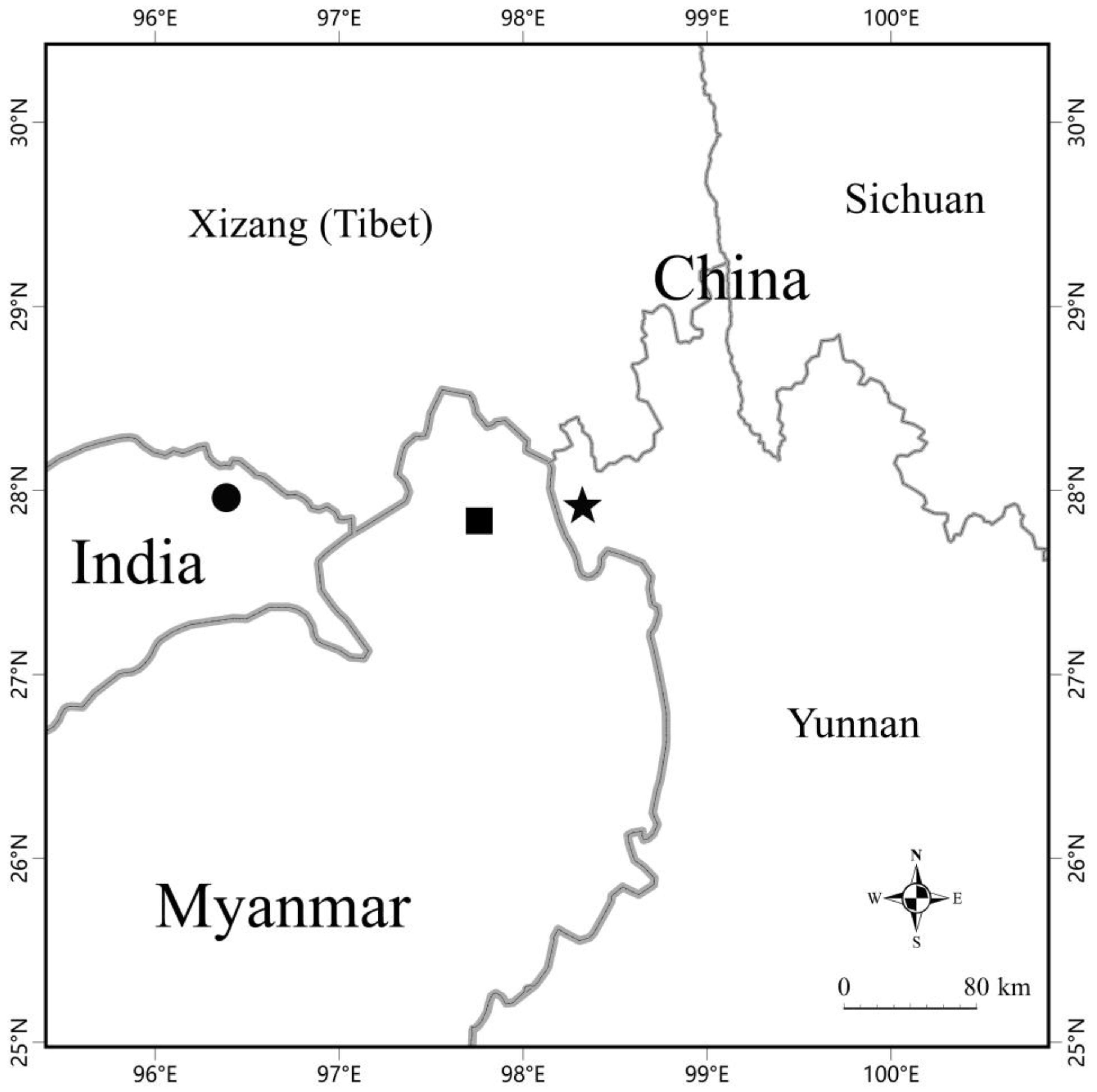
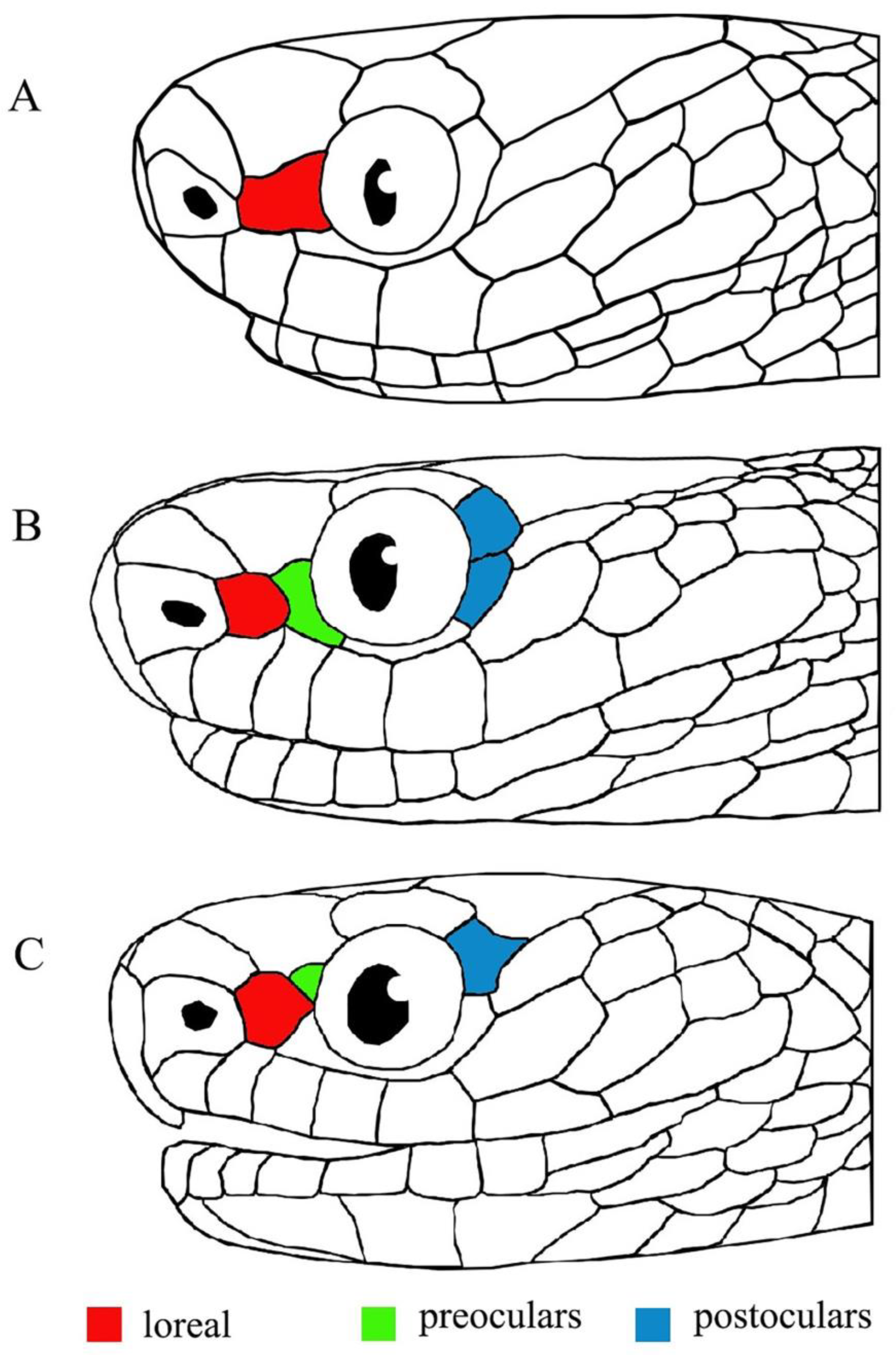
Vogel et al. [19] indicated a potential new species of Pareas from Putao District, Kachin State, northern Myanmar, which lies just between the type localities of P. kaduri and Pareas dulongjiangensis sp. nov. (Figure 5); however, due to the lack of available molecular data for this potential new species, it was not formally described. In terms of morphology, this potential new species also resembles P. kaduri and Pareas dulongjiangensis sp. nov.; nevertheless, it differs from P. kaduri as the loreal contacts the eye (vs. not touching the eye) (see Figure 6), the dorsal scales are keeled in the single female (vs. smooth dorsal scales in the single female), it has slightly more subcaudals (71 vs. 65–70), and has a slightly, relatively longer tail (TaL/TL 0.210 vs. 0.184–0.207); it differs from Pareas dulongjiangensis sp. nov. by the presence of preoculars (vs. preoculars being absent), having one postocular (vs. postocular being fused with subocular), having one row enlarged median vertebral scales (vs. three rows), having fewer subcaudals (71 vs. 76), having a relatively shorter tail (TaL/TL 0.210 vs. 0.236), and having more vertical dark bars on the trunk (60 vs. 52). Although P. kaduri and Pareas dulongjiangensis sp. nov. have the closest phylogenetic relationship, the genetic distance between them is still relatively large (9.4%). We speculate that this potential new species is probably a species sandwiched between P. kaduri and Pareas dulongjiangensis sp. nov. As there is still no molecular data available for this potential new species, this speculation needs to be verified.
Pareas dulongjiangensis sp. nov. is phylogenetically closely related to P. kaduri. Pareas kaduri is distributed near the Lohit River bank, which belongs to the Brahmaputra River system, whereas Pareas dulongjiangensis sp. nov. is distributed near the Dulongjiang River bank, which belongs to the Irrawaddy River system. There are many high mountains between the two river systems [44], which would lead to geographical isolation. Therefore, biogeography also supports that Pareas dulongjiangensis sp. nov. and P. kaduri belong to two different species.
At present, Pareas dulongjiangensis sp. nov. is known only from a single specimen, and nothing is known about the actual distribution range and ecological data of this species. Further research is required to clarify the extent of its distribution, population trends, and conservation status. Moreover, investigation of species diversity in the China–Myanmar border region is far from complete due to the relatively inconvenient traffic conditions; more survey efforts are required to achieve a better understanding of the diversity of Pareas in this region.
Author Contributions
Investigation, D.R.; experiment, M.Y. and J.R.; writing, S.L.; review, Y.G. and D.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Science-Technology Basic Condition Platform from the Ministry of Science and Technology of the People’s Republic of China (grant no. 2005DKA21402), the National Natural Science Foundation Project: Systematic Taxonomy and Biogeographic Study of Pareidae (grant no. 31460558), the National Key Laboratory of Genetic Resources and Evolution and Open Project (grant no. GREKF17-08), and the National Important Research and Development Project: Biodiversity Conservation and Restoration Technology in High Mountain and Valley Regions of Southwestern China (grant no. 2017YFC0505202).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are presented in this article and in GenBank (https://www.ncbi.nlm.nih.gov).
Acknowledgments
The curator and deputy curator of the Kunming Natural History Museum of Zoology, Kunming Institute of Zoology, Chinese Academy of Sciences, are gratefully acknowledged for their support in the taxonomic study. We would like to thank Xiaoyu Zhu for his assistance in the field, Guangyi Lu for providing information about specimen collection, and our workmates for their help and advice. We are also grateful to the reviewers for their valuable comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smith, M.A. The Fauna of British India, Ceylon and Burma, including the Whole of the Indo-Chinese Subregion. Reptilia and Amphibia. Vol. III. Serpentes; Taylor and Francis: London, UK, 1943. [Google Scholar]
- Zhao, E.M.; Huang, M.H.; Zong, Y.; Jiang, Y.M.; Huang, Q.Y.; Zhao, H.; Ma, J.F.; Huang, Z.J.; Wei, G.; Yang, D.T.; et al. Fauna Sinica, Reptilia, Squamata. Serpentes; Science Press: Beijing, China, 1998. [Google Scholar]
- Zhao, E.M. Snakes of China; Anhui Science Technology Publishing House: Hefei, China, 2006; Volume 1. [Google Scholar]
- Wagler, J.G. Natürliches System der Amphibien, mit Vorangehender Classification der Säugthiere und Vogel: Ein Neitrag zur Vergleichenden Zoologie; J.G. Cottasche Buchhandlung Nachfolger: Stuttgart, Germany, 1830. [Google Scholar]
- Deepak, V.; Ruane, S.; Gower, D.J. A New Subfamily of Fossorial Colubroid Snakes from the Western Ghats of Peninsular India. J. Nat. Hist. 2019, 52, 2919–2934. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Che, J.; Liu, Q.; Li, K.; Jin, J.Q.; Jiang, K.; Shi, L.; Guo, P. A Revised Taxonomy of Asian Snail-Eating Snakes Pareas (Squamata, Pareidae): Evidence from Morphological Comparison and Molecular Phylogeny. ZooKeys 2020, 939, 45–64. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. (Eds.) The Reptile Database. Available online: http://www.reptile-database.org (accessed on 16 January 2023).
- Poyarkov, N.A.; Nguyen, T.V.; Pawangkhanant, P.; Yushchenko, P.V.; Brakels, P.; Nguyen, L.H.; Nguyen, H.N.; Suwannapoom, C.; Orlov, N.; Vogel, G. An Integrative Taxonomic Revision of Slug-Eating Snakes (Squamata: Pareidae: Pareineae) Reveals Unprecedented Diversity in Indochina. PeerJ 2022, 10, e12713. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.T.; Georgalis, G.L. The Diversity and Distribution of Palaeogene Snakes—A Review, with Comments on Vertebral Sufficiency. In The Origin and Early Evolution of Snakes; Gower, D., Zaher, H., Eds.; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Huang, Q.Y. Pareas macularius Theobald, 1868 should be a Junior Synonym of Pareas margaritophorus (Jan, 1866). Sichuan J. Zool. 2004, 23, 207–208. [Google Scholar]
- Hauser, S. On the Validity of Pareas macularius Theobald, 1868 (Squamata: Pareidae) as a Species Distinct from Pareas margaritophorus (Jan in Bocourt, 1866). Trop. Nat. Hist. 2017, 17, 25–52. [Google Scholar]
- Guo, K.J.; Deng, X.J. A New Species of Pareas (Serpentes: Colubridae: Pareatinae) from the Gaoligong Mountains, Southwestern China. Zootaxa 2009, 2008, 53–60. [Google Scholar] [CrossRef]
- Vogel, G. A New Montane Species of the Genus Pareas Wagler, 1830 (Squamata: Pareatidae) from Northern Myanmar. Taprobanica 2015, 7, 1–7. [Google Scholar] [CrossRef]
- You, C.W.; Poyarkov, N.A.; Lin, S.M. Diversity of the Snail-Eating Snakes Pareas (Serpentes, Pareatidae) from Taiwan. Zool. Scr. 2015, 44, 349–361. [Google Scholar] [CrossRef]
- Ding, L.; Chen, Z.N.; Suwannapoom, C.; Nguyen, T.V.; Poyarkov, N.A.; Vogel, G. A New Species of the Pareas hamptoni Complex (Squamata: Serpentes: Pareidae) from the Golden Triangle. Taprobanica 2020, 9, 174–193. [Google Scholar] [CrossRef]
- Bhosale, H.; Phansalkar, P.; Sawant, M.; Gowande, G.; Patel, H.; Mirza, Z.A. A New Species of Snail-Eating Snakes of the Genus Pareas Wagler, 1830 (Reptilia: Serpentes) from Eastern Himalayas, India. Eur. J. Taxon. 2020, 729, 54–73. [Google Scholar] [CrossRef]
- Vogel, G.; Nguyen, T.V.; Lalremsanga, H.T.; Biakzuala, L.; Hrima, V.; Poyarkov, N.A. Taxonomic Reassessment of the Pareas margaritophorus-macularius Species Complex (Squamata, Pareidae). Vertebr. Zool. 2020, 70, 547–569. [Google Scholar] [CrossRef]
- Liu, S.; Rao, D.Q. A New Species of the Genus Pareas (Squamata, Pareidae) from Yunnan, China. Zookeys 2021, 1011, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Vogel, G.; Nguyen, T.V.; Zaw, T.; Poyarkov, N.A. A New Species of the Pareas monticola Complex (Squamata: Serpentes: Pareidae) from Chin Mountains with Additions to the Pareas Fauna of Myanmar. J. Nat. Hist. 2021, 54, 2577–2612. [Google Scholar] [CrossRef]
- Le, D.T.T.; Tran, T.G.; Hoang, H.D.; Stuart, B.L. A New Species of Pareas (Squamata, Pareidae) from Southern Vietnam. Vertebr. Zool. 2021, 71, 439–451. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, D.R.; Poyarkov, N.A.; Hou, M.; Wu, L.; Rao, D.Q.; Nguyen, T.V.; Vogel, G. Resurrection of Pareas yunnanensis (Vogt, 1922) with Description of a New Species of Pareas from Yunnan Province, China (Squamata, Pareidae). Eur. J. Taxon. 2023, 860, 1–26. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Marchese, C. Biodiversity hotspots: A Shortcut for a More Complicated Concept. Glob. Ecol. Conserv. 2015, 3, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.R.; Tang, J.S.; Xu, Z.H.; Yang, Y.M.; Chen, Y.S.; Wang, J.H. Goligongshan National Nature Reserve; China Forestry Press: Beijing, China, 1995. [Google Scholar]
- Boulenger, G.A. Descriptions of New Reptiles and Batrachians from Borneo. Proc. Zool. Soc. London 1900, 69, 182–187. [Google Scholar] [CrossRef]
- Boulenger, G.A. Descriptions of Two New Snakes from Upper Burma. Bombay Nat. Hist. Soc. 1905, 16, 235–236. [Google Scholar] [CrossRef]
- Vogt, T. Zur Reptilien-und Amphibienfauna Südchinas. Arch. Für Nat. 1992, 88, 135–146. [Google Scholar]
- Pope, C.H. The Reptiles of China. Turtles, Crocodilians, Snakes, Lizards, Natural History of Central Asia; The American Museum of Natural History: New York, NY, USA, 1935; Volume X. [Google Scholar]
- Grossmann, W.; Tillack, F. On the Taxonomic Status of Asthenodipsas tropidonotus (Van Lidth de Jeude, 1923) and Pareas vertebralis (Boulenger, 1900) (Serpentes: Colubridae: Pareatinae). Russ. J. Herpetol. 2003, 10, 175–190. [Google Scholar]
- Guo, Y.H.; Wu, Y.K.; He, S.P.; Shi, H.T.; Zhao, E.M. Systematics and Molecular Phylogenetics of Asian Snail-Eating Snakes (Pareatidae). Zootaxa 2011, 3001, 57–64. [Google Scholar] [CrossRef]
- Loredo, A.I.; Wood, P.L., Jr.; Quah, E.S.; Anuar, S.; Greer, L.; Norhayati, A.; Grismer, L.L. Cryptic Speciation within Asthenodipsas vertebralis (Boulenger, 1900) (Squamata: Pareatidae), the Description of a New Species from Peninsular Malaysia, and the Resurrection of A. tropidonotus (Lidth de Jude, 1923) from Sumatra: An Integrative Taxonomic Analysis. Zootaxa 2013, 3664, 505–524. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Yeung, H.Y.; Huang, X.Y.; Yang, S.P. First Record of Pareas vindumi Vogel, 2015 (Reptilia: Pareidae) from China with a Revision to Morphology. Taprobanica 2021, 10, 39–46. [Google Scholar] [CrossRef]
- David, P.; Deuti, K. On the Type Specimens of Pareas macularius Theobald, 1868 and Pareas berdmorei Theobald, 1868 with the Designation of a Lectotype for Pareas macularius (Squamata: Serpentes: Pareidae). Zootaxa 2022, 5105, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Lawson, R.; Slowinski, J.B.; Crother, B.I.; Burbrink, F.T. Phylogeny of the Colubroidea (Serpentes): New evidence from Mitochondrial and Nuclear Genes. Mol. Phylogenetics Evol. 2005, 37, 581–601. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment Through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian Inference of Phylogeny and its Impact on Evolutionary Biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, T.P.; Zwickl, D.J.; Heath, T.A.; Hillis, D.M. Phylogenetic Relationships of the Dwarf Boas and a Comparison of Bayesian and Bootstrap Measures of Phylogenetic Support. Mol. Phylogenetics Evol. 2002, 25, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or Bootstrap? A Simulation Study Comparing the Performance of Bayesian Markov chain Monte Carlo Sampling and Bootstrapping in Assessing Phylogenetic Confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Che, J.; Jiang, K.; Yan, F.; Zhang, Y.P. Amphibians and Reptiles in Tibet-Diversity and Evolution; Science Press: Beijing, China, 2020. [Google Scholar]
Figure 1.
Phylogenetic tree of Pareas inferred from cyt b gene. The new species is highlighted in brown. Numbers before slashes indicate values (>0.9) of Bayesian posterior probabilities and numbers after slashes indicate values (>90) of bootstrap support for maximum likelihood analyses.
Figure 1.
Phylogenetic tree of Pareas inferred from cyt b gene. The new species is highlighted in brown. Numbers before slashes indicate values (>0.9) of Bayesian posterior probabilities and numbers after slashes indicate values (>90) of bootstrap support for maximum likelihood analyses.

Figure 2.
Pareas dulongjiangensis sp. nov., holotype in preservative. (A) Dorsal view of the body; (B) ventral view of the body; (C) close-up view of the dorsal side of the head; (D) close-up view of the left side of the head; (E) close-up view of the ventral side of the head.
Figure 2.
Pareas dulongjiangensis sp. nov., holotype in preservative. (A) Dorsal view of the body; (B) ventral view of the body; (C) close-up view of the dorsal side of the head; (D) close-up view of the left side of the head; (E) close-up view of the ventral side of the head.

Figure 3.
Close-up views of the dorsal scales at the middle of the body of the holotype (KIZ R201607) in preservative. (A) Dorsal side; (B) left side; (C) right side.
Figure 3.
Close-up views of the dorsal scales at the middle of the body of the holotype (KIZ R201607) in preservative. (A) Dorsal side; (B) left side; (C) right side.

Figure 4.
The habitat of Pareas dulongjiangensis sp. nov. at the type locality.

Figure 5.
Map showing the type locality (black star) of Pareas dulongjaingensis sp. nov., the type locality (black dot) of P. kaduri, and the locality (black square) of the potential new species in Vogel et al. [19].
Figure 5.
Map showing the type locality (black star) of Pareas dulongjaingensis sp. nov., the type locality (black dot) of P. kaduri, and the locality (black square) of the potential new species in Vogel et al. [19].

Figure 6.
Comparison of head scalation. (A) Holotype of Pareas dulongjiangensis sp. nov. (KIZ R201607); (B) holotype of P. kaduri (BNHS 3574); (C) the potential new species (CAS 235254) in Vogel et al. [19].
Figure 6.
Comparison of head scalation. (A) Holotype of Pareas dulongjiangensis sp. nov. (KIZ R201607); (B) holotype of P. kaduri (BNHS 3574); (C) the potential new species (CAS 235254) in Vogel et al. [19].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sequences (cyt b) used in phylogenetic analysis of this study. “Mt.” = mountain, “Is.” = island, “Hw.” = highway.
Table 1.
Sequences (cyt b) used in phylogenetic analysis of this study. “Mt.” = mountain, “Is.” = island, “Hw.” = highway.
| Species | Locality | Voucher No. | GenBank No. |
|---|---|---|---|
| Pareas abros | Song Thanh, Quang Nam, Vietnam | ZMMU R-16393 | MZ712235 |
| Pareas andersonii | Mt. Natmataung, Chin, Myanmar | CAS 235359 | MT968772 |
| Pareas atayal | N. Cross Is. Hw., Taiwan, China | NMNS 05594 | KJ642122 |
| Pareas berdmorei | Kin Pon Chaung, Mon, Myanmar | CAS 240362 | MZ712219 |
| Pareas boulengeri | Jiangkou, Guizhou, China | GP 2923 | MK135090 |
| Pareas carinatus | Sungai Sedim, Kedah, Malaysia | LSUHC10604 | KC916748 |
| Pareas chinensis | Hongya, Sichuan, China | GP 2383 | MK135089 |
| Pareas formosensis | N. Cross Is. Hw., Taiwan, China | NMNS 05632 | KJ642130 |
| Pareas geminatus | Jiangcheng, Yunnan, China | CIB 118021 | MW287068 |
| Pareas hamptoni | Kachin, Myanmar | YPX 18219 | MK135077 |
| Pareas iwasakii | Ishigaki Is., S. Ryukyu, Japan | I03-ISG1 | KJ642158 |
| Pareas kaduri | Lohit, Arunachal, India | BNHS 3574 | MT188734 |
| Pareas kaduri | Lohit, Arunachal, India | BNHS 3575 | MW026190 |
| Pareas komaii | Taitung, Taiwan, China | HC 000669 | JF827687 |
| Pareas kuznetsovorum | Song Hinh, Phu Yen, Vietnam | ZMMU R-16802 | MZ712232 |
| Pareas macularius | Bago, Myanmar | CAS 206620 | AF471082 |
| Pareas margaritophorus | Cangwu, Guangxi, China | YBU 16061 | MK135097 |
| Pareas modestus | Aizawl, Mizoram, India | MZMU 1293 | MT968773 |
| Pareas monticola | Medog, Tibet, China | GP 2027 | MK135107 |
| Pareas niger | Kunming, Yunnan, China | KIZ 059339 | MW436706 |
| Pareas nigriceps | Mt. Gaoligong, Yunnan, China | SYSr001222 | MK201455 |
| Pareas nuchalis | Belait, Brunei | FK 2626 | MZ603794 |
| Pareas stanleyi | Guilin, Guangxi, China | HM 2007-S001 | JN230704 |
| Pareas temporalis | Da Huoai, Lam Dong, Vietnam | UNS 09992 | MZ603793 |
| Pareas tigerinus | Menghai, Yunnan, China | KIZ 20210703 | OP752143 |
| Pareas tigerinus | Menghai, Yunnan, China | KIZ 20210704 | OP752144 |
| Pareas tigerinus | Menghai, Yunnan, China | KIZ 20210705 | OP752145 |
| Pareas victorianus | Mt. Natmataung, Chin, Myanmar | CAS 235254 | MW438300 |
| Pareas vindumi | Lukpwir, Kachin, Myanmar | CAS 248147 | MT968776 |
| Pareas xuelinensis | Lancang, Yunnan, China | KIZ XL1 | MW436709 |
| Pareas yunnanensis | Dali, Yunnan, China | KIZ 2022036 | OP752149 |
| Pareas dulongjiangensis sp. nov. | Gongshan, Yunnan, China | KIZ R201607 | OQ718498 |
| Aplopeltura boa | Malaysia | LSUHC 7248 | KC916746 |
Table 2.
Uncorrected p-distances (%) amongst the members of Pareas, calculated from cyt b gene sequences. (1) Pareas abros, (2) P. andersonii, (3) P. atayal, (4) P. berdmorei, (5) P. boulengeri, (6) P. carinatus, (7) P. chinensis, (8) P. formosensis, (9) P. geminatus, (10) P. hamptoni, (11) P. iwasakii, (12) P. kaduri, (13) P. komaii, (14) P. kuznetsovorum, (15) P. macularius, (16) P. margaritophorus, (17) P. modestus, (18) P. monticola, (19) P. niger, (20) P. nigriceps, (21) P. nuchalis, (22) P. stanleyi, (23) P. temporalis, (24) P. tigerinus, (25) P. victorianus, (26) P. vindumi, (27) P. xuelinensis, (28) P. yunnanensis, (29) Pareas dulongjiangensis sp. nov.
Table 2.
Uncorrected p-distances (%) amongst the members of Pareas, calculated from cyt b gene sequences. (1) Pareas abros, (2) P. andersonii, (3) P. atayal, (4) P. berdmorei, (5) P. boulengeri, (6) P. carinatus, (7) P. chinensis, (8) P. formosensis, (9) P. geminatus, (10) P. hamptoni, (11) P. iwasakii, (12) P. kaduri, (13) P. komaii, (14) P. kuznetsovorum, (15) P. macularius, (16) P. margaritophorus, (17) P. modestus, (18) P. monticola, (19) P. niger, (20) P. nigriceps, (21) P. nuchalis, (22) P. stanleyi, (23) P. temporalis, (24) P. tigerinus, (25) P. victorianus, (26) P. vindumi, (27) P. xuelinensis, (28) P. yunnanensis, (29) Pareas dulongjiangensis sp. nov.
| (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) | (10) | (11) | (12) | (13) | (14) | (15) | (16) | (17) | (18) | (19) | (20) | (21) | (22) | (23) | (24) | (25) | (26) | (27) | (28) | |
| (1) | ||||||||||||||||||||||||||||
| (2) | 23.5 | |||||||||||||||||||||||||||
| (3) | 22.8 | 20.2 | ||||||||||||||||||||||||||
| (4) | 21.3 | 23.8 | 23.5 | |||||||||||||||||||||||||
| (5) | 23.2 | 19.7 | 18.3 | 23.7 | ||||||||||||||||||||||||
| (6) | 21.8 | 22.9 | 22.6 | 13.8 | 22.2 | |||||||||||||||||||||||
| (7) | 23.7 | 19.1 | 18.4 | 24.7 | 9.0 | 22.6 | ||||||||||||||||||||||
| (8) | 22.9 | 21.7 | 15.1 | 24.9 | 17.2 | 23.9 | 17.4 | |||||||||||||||||||||
| (9) | 22.9 | 22.2 | 14.2 | 23.0 | 17.4 | 23.1 | 18.8 | 8.3 | ||||||||||||||||||||
| (10) | 23.6 | 21.5 | 14.2 | 23.7 | 17.2 | 23.6 | 18.3 | 7.2 | 7.3 | |||||||||||||||||||
| (11) | 23.4 | 20.4 | 7.2 | 23.7 | 16.8 | 23.3 | 17.9 | 14.4 | 14.4 | 13.5 | ||||||||||||||||||
| (12) | 25.0 | 20.7 | 15.6 | 24.9 | 19.8 | 22.6 | 19.3 | 13.3 | 13.7 | 13.0 | 15.2 | |||||||||||||||||
| (13) | 23.3 | 19.5 | 8.5 | 23.9 | 18.1 | 23.9 | 18.3 | 14.7 | 14.9 | 14.5 | 7.9 | 16.1 | ||||||||||||||||
| (14) | 20.9 | 23.8 | 22.9 | 13.0 | 22.6 | 13.0 | 23.0 | 23.7 | 22.8 | 23.3 | 23.9 | 23.1 | 24.2 | |||||||||||||||
| (15) | 23.0 | 13.9 | 19.2 | 22.7 | 17.8 | 22.1 | 17.4 | 19.0 | 20.4 | 19.7 | 18.9 | 19.8 | 18.3 | 22.8 | ||||||||||||||
| (16) | 25.8 | 15.3 | 19.1 | 24.7 | 19.2 | 23.5 | 18.3 | 20.5 | 21.8 | 20.5 | 18.8 | 20.8 | 19.5 | 23.7 | 14.8 | |||||||||||||
| (17) | 23.5 | 12.0 | 18.7 | 24.4 | 19.2 | 24.0 | 18.7 | 20.7 | 19.9 | 19.6 | 19.3 | 19.4 | 17.8 | 24.4 | 11.0 | 13.9 | ||||||||||||
| (18) | 22.6 | 18.9 | 17.3 | 22.0 | 18.7 | 22.8 | 18.1 | 18.9 | 19.7 | 19.0 | 17.8 | 19.0 | 17.9 | 22.5 | 18.1 | 19.7 | 18.2 | |||||||||||
| (19) | 22.7 | 20.4 | 14.3 | 23.8 | 17.5 | 23.0 | 17.8 | 7.2 | 6.8 | 5.6 | 13.7 | 12.5 | 14.9 | 22.7 | 18.9 | 20.1 | 18.9 | 18.5 | ||||||||||
| (20) | 23.6 | 18.8 | 16.2 | 22.9 | 16.9 | 22.6 | 16.2 | 12.6 | 13.5 | 12.6 | 16.1 | 10.1 | 16.2 | 23.9 | 17.8 | 17.9 | 16.4 | 19.1 | 12.5 | |||||||||
| (21) | 21.1 | 24.3 | 23.7 | 21.5 | 24.3 | 21.6 | 24.0 | 24.4 | 25.1 | 24.8 | 24.5 | 25.6 | 23.5 | 20.4 | 23.1 | 26.1 | 24.5 | 21.4 | 25.2 | 23.8 | ||||||||
| (22) | 25.7 | 20.4 | 19.2 | 25.0 | 15.7 | 24.9 | 15.4 | 19.6 | 19.4 | 18.7 | 18.2 | 20.6 | 17.4 | 24.9 | 19.9 | 19.5 | 19.4 | 19.2 | 19.5 | 19.0 | 24.0 | |||||||
| (23) | 12.3 | 23.6 | 23.1 | 20.6 | 22.1 | 19.9 | 21.5 | 24.3 | 23.6 | 23.4 | 23.1 | 24.8 | 23.8 | 20.1 | 24.4 | 24.0 | 23.2 | 21.3 | 23.3 | 23.8 | 19.8 | 23.4 | ||||||
| (24) | 23.1 | 19.4 | 14.7 | 24.2 | 19.0 | 23.3 | 18.6 | 12.3 | 12.3 | 11.8 | 14.1 | 11.2 | 14.0 | 24.3 | 18.6 | 20.4 | 18.1 | 18.9 | 11.4 | 4.3 | 25.2 | 19.4 | 24.6 | |||||
| (25) | 24.3 | 20.6 | 19.6 | 22.8 | 19.1 | 22.8 | 17.4 | 17.8 | 18.4 | 18.6 | 19.7 | 18.9 | 19.4 | 22.9 | 19.1 | 21.5 | 19.3 | 15.1 | 17.9 | 19.1 | 24.7 | 19.0 | 24.2 | 18.1 | ||||
| (26) | 24.5 | 20.8 | 14.9 | 24.7 | 18.4 | 23.8 | 17.5 | 12.1 | 12.4 | 11.4 | 14.7 | 12.9 | 15.2 | 23.7 | 19.3 | 20.5 | 19.9 | 18.3 | 10.8 | 12.3 | 24.7 | 19.4 | 24.9 | 12.0 | 17.8 | |||
| (27) | 23.1 | 21.2 | 13.8 | 25.1 | 16.9 | 24.3 | 18.6 | 8.0 | 5.9 | 8.1 | 13.7 | 13.6 | 14.8 | 24.5 | 19.2 | 21.3 | 20.2 | 19.8 | 7.2 | 12.5 | 25.9 | 19.4 | 24.4 | 12.1 | 18.8 | 12.6 | ||
| (28) | 23.2 | 22.1 | 14.6 | 24.6 | 16.7 | 23.4 | 18.0 | 7.9 | 4.0 | 6.1 | 14.0 | 12.9 | 14.7 | 23.7 | 19.7 | 21.5 | 20.7 | 20.0 | 6.3 | 12.8 | 24.8 | 19.5 | 23.7 | 11.7 | 18.7 | 11.4 | 6.1 | |
| (29) | 24.0 | 19.6 | 14.0 | 24.4 | 18.0 | 23.3 | 17.6 | 12.9 | 12.7 | 12.4 | 13.9 | 9.4 | 13.8 | 24.2 | 19.4 | 18.4 | 18.6 | 16.5 | 12.3 | 10.3 | 24.6 | 19.2 | 25.0 | 10.3 | 18.0 | 12.6 | 13.2 | 12.9 |
Table 3.
Comparison of morphometric and meristic data for Pareas dulongjiangensis sp. nov. and P. kaduri. The data for P. kaduri were obtained from Bhosale et al. [16].
Table 3.
Comparison of morphometric and meristic data for Pareas dulongjiangensis sp. nov. and P. kaduri. The data for P. kaduri were obtained from Bhosale et al. [16].
| Pareas dulongjiangensis sp. nov. Holotype KIZ R201607 | Pareas kaduri n = 4 3, 1 | |
|---|---|---|
| SVL | 373 | 455–550 |
| TaL | 115 | 113–144 |
| TL | 488 | 571–694 |
| TaL/SVL | 0.308 | 0.226–0.262 |
| TaL/TL | 0.236 | 0.184–0.207 |
| HL | 14.0 | 10.4–14.6 |
| CL | 15.4 | 14.3–18.8 |
| HW | 8.1 | 7.1–8.8 |
| ED | 2.6 | 2.7–3.6 |
| El | 1.7 | 1.5–1.8 |
| ES | 4.1 | 3.7–4.3 |
| EN | 1.8 | 2.3–2.7 |
| NW | 3.6 | 3.5–5.8 |
| PrFBO | Yes | Yes |
| PreO | 0 | 1 |
| PosO | Fused | 2 |
| SubO | Fused | 1 |
| SPOF | Yes | No |
| ATem | 2/2 | 2 |
| PTem | 3/2 | 3 |
| SupL | 6/7 | 7 |
| InfL | 7/9 | 7 |
| LoBO | Yes | No |
| Vs | 182 | 160–183 |
| Prec | Undivided | Undivided |
| Sc | 76 | 52–70 |
| Ds | 15-15-15 | 15-15-15 |
| NED | 3 | 1 |
| NKD | 5 | 8 in ♂, 0 in ♀ |
| Max | 5/4 | 6–7 |
| DNB | Yes | Yes |
| VBTr | 52 | 56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, S.; Yang, M.; Rao, J.; Guo, Y.; Rao, D. A New Species of Pareas Wagler, 1830 (Squamata, Pareidae) from Northwestern Yunnan, China. Taxonomy 2023, 3, 169-182. https://doi.org/10.3390/taxonomy3020013
AMA Style
Liu S, Yang M, Rao J, Guo Y, Rao D. A New Species of Pareas Wagler, 1830 (Squamata, Pareidae) from Northwestern Yunnan, China. Taxonomy. 2023; 3(2):169-182. https://doi.org/10.3390/taxonomy3020013
Chicago/Turabian StyleLiu, Shuo, Mingjing Yang, Jingqiu Rao, Yuhong Guo, and Dingqi Rao. 2023. "A New Species of Pareas Wagler, 1830 (Squamata, Pareidae) from Northwestern Yunnan, China" Taxonomy 3, no. 2: 169-182. https://doi.org/10.3390/taxonomy3020013