
Taxonomic Structure and Diversity of Benthic Diatoms (Bacillariophyta) in the Northern Shelf of the Black Sea

A.O. Kovalevsky Institute of Biology of Southern Seas Russian Academy of Sciences, 2 Nakhimov av, Sevastopol 299011, Russia
Phycology 2023, 3(3), 337-355; https://doi.org/10.3390/phycology3030022
Submission received: 8 May 2023
/
Revised: 19 June 2023
/
Accepted: 3 July 2023
/
Published: 7 July 2023
Abstract
:The key role of benthic diatoms in coastal ecosystems highlights the relevance of studying their biodiversity and developing integrated biodiversity conservation measures. Benthic diatom richness in the northern part of the Black Sea was estimated based on both published data and our own data. The checklist of benthic diatoms includes 1100 species and intraspecific taxa, 953 species, 149 genera, 61 families, 32 orders, and 3 classes of Bacillariophyta. More than 280 species and intraspecific taxa, as well as 8 genera new to the diatom flora of the Black Sea and 7 species new to science, were revealed. An interregional analysis of the diversity of benthic Bacillariophyta along the coasts of Bulgaria, Romania, and Crimea, as well as the northwestern and northeastern parts of the Black Sea shelf, was carried out using the Taxonomic Distinctness Indices (TaxDI). The lowest TaxDI value for Crimea was closest to the average value expected for the Black Sea diatom flora, indicating a large proportion of polyspecies branches in the taxocene structure, which resulted in the maximum species richness (species/genus = 7) and led to a decrease in the vertical evenness of hierarchical tree. On the contrary, the highest TaxDI value was detected off the Romanian coast, which was caused by the significant proportion of mono- and oligospecies branches and minimal species saturation in the taxocene structure (species/genus = 4). We also identified some species with a high taxonomic exclusivity rank. If these species disappear, the entire architectonics of the Black Sea hierarchical tree of Bacillariophyta may be significantly altered. Such region-specific species are given the highest protection priority since they have great importance for gene pool conservation.
1. Introduction
Diatoms (Bacillariophyta) account for up to 91–98% of abundance and species richness in the studied marine microphytobenthos [1,2]. These are photosynthetic microorganisms that are mobile or attached and living on any underwater substrate. Benthic diatoms are the most important producers of organic matter and oxygen, representing the food base and initial links in the trophic chains of sublittoral marine ecosystems while also leading in species richness among other groups of algae in the world’s oceans [1,2]. In contrast to those on phytoplankton diatoms, existing studies of benthic microalgae are not as extensive and remain limited to certain regions both globally and in the Black Sea. The key role of benthic diatom algae in coastal ecosystem functioning highlights the relevance of studying Bacillariophyta biodiversity and why their conservation should be prioritised [3,4,5,6]. The combined impact of abiotic factors leads to changes in the structure of the benthic diatom assemblages, which actualises their use as ecological indicators of aquatic environment quality [7,8,9].
Considerable changes in the Bacillariophyta taxonomy system in recent years, as well as the accumulation of extensive but scattered data in the literature on some areas, make it necessary to revise all existing materials and create a unified master list, which is a basic initial step in assessing the diversity of benthic diatoms in the Black Sea. Some of the most recent extensive synopses include collective work on the microalgae of Ukraine [10], a species list of the Black Sea microphytobenthos [11], a checklist of the microphytobenthos of the northwestern Black Sea [12], and a master list of Bacillariophyta in the northern shelf of the Black Sea [13]. The summary on the algoflora of Ukraine [10] considered 4817 species represented by 6004 intraspecific taxa (hereinafter, ISTs), of which Bacillariophyta makes up 15% (714 species, 930 ISTs). Research has mainly been conducted on freshwater species and has not included many genera and families of marine diatoms. Despite tremendous works on the collection and synonymy of the aforementioned species lists, the synopses created by previous authors also does not cover all of the available information. The creation of a complete list of the Black Sea diatom flora seems possible only by combining and thoroughly inventorying the entire existing dataset.
The benthic diatom flora found off the Bulgarian coast (BC) [14], Romanian coast (RC) [15,16], northwestern part of the Black Sea and adjacent estuaries and bays (NWBS) [17,18], Crimean coast (CC) [11,13,18,19,20], and the northeastern part of the Black Sea (NEBS) [13,19,20,21] have been extensively studied. Information on benthic diatoms of the southeastern and southern parts of the Black Sea shelf remains insufficient for a comprehensive comparative analysis of regional floras. For example, 225 species and ISTs were described in works on microplankton found off the Turkish coast, among which approximately 90 are typical planktonic forms of Bacillariophyta [22,23]. There are also reports of new phytoplankton species found off the southern Black Sea [24,25]. Moreover, data on the discovery of new benthic diatom species found off the Black Sea coast of Turkey and Bulgaria appeared at the time of writing this manuscript [26,27,28].
Regional studies have mainly been concerned with the species composition and seasonal dynamics of the diatoms of hard natural and artificial substrates and, to a lesser extent, the structure and composition features of diatom assemblages from soft-bottom substrates. Notably, to date, the challenge of quantifying Bacillariophyta diversity remains almost entirely unresolved. Since historical data are usually represented by simple species lists, it is not possible to quantify the diatom species diversity of the Black Sea by using traditional metrics (e.g., the Shannon, Pielou, and Margalef indices, etc.) that require the number or biomass of individuals of each species. Most widely used methods for measuring biodiversity based on the number of species consider either the number of selected individuals (e.g., the Margalef index of species richness), similarities in the distributions of individuals between species (e.g., Pielou evenness index), or a quantitative assessment of both diversity parameters (i.e., species richness and evenness in the distribution of individuals between species; for example, the Shannon species richness index) [29]. The aforementioned indices can be applied for the comparative assessment of biodiversity in situations where sampling efforts, size, and habitat types are known and comparable, as well as in comparative studies of similar habitats subjected to local external effects. However, the application of such indices is not very effective when analysing heterogeneous datasets that differ significantly in spatial and temporal aspects. In most cases, these indices are highly dependent on sample size and, as a rule, they are of little use in the comparative analysis of data with an unknown (and different) number of replicates during sampling. Therefore, the use of such indices for comparative assessments of biodiversity, especially when the size and number of samples (as well as the types of habitats) differ significantly, is notoriously inefficient or even impossible.
Given this context, when applying only arrays of initial qualitative data, indicators of diversity assessment unrelated to any size and replicates numbers of samples should be used. The Taxonomic Distinctness indices (TaxDI) [30,31,32,33,34,35] allows the estimation of taxonomic aspects of diversity in a biotope based on a taxon’s hierarchical structure by calculating the average degree of phylogenetic similarity between the species. In this case, applying this indices reveals regional differences in the taxonomic structure of benthic diatoms and determines the deviations of index values from the expected average mode calculated for the entire Black Sea using historical data represented by simple species lists.
Thus, the relevance of our work stems from the need for an interregional analysis of the taxonomic diversity of Bacillariophyta in the Black Sea based on the revision and integration of historical and modern data. The present work represents one of the first attempts in marine biology to apply TaxDI to assess the taxonomic diversity of benthic diatoms and summarise information on species richness in the northern shelf of the Black Sea based on data found in the literature and our own data. This work aimed to analyse the reasons for disparities in taxonomic diversity and compare the floristic similarities of benthic diatoms in different regions of the Black Sea.
The results of our calculations based on our study of the Black Sea’s northern shelf should not be considered wholly finalized. Instead, our work is open for supplementation as new data become available. Undoubtedly, with the expansion of the database to include benthic diatoms found off the Southern Black Sea shelf (i.e., the coasts of Bulgaria, Turkey, and the Caucasus), the TaxDI indices and parameters of regional species richness and Bacillariophyta taxonomic diversity are expected to change. Recent publications by colleagues from Turkey and Bulgaria provide some hope for this outcome. Within this context, we believe that our work will help increase people’s awareness of benthic Bacillariophyta diversity and further develop relevant research in the Black Sea region.
2. Materials and Methods
2.1. Sampling Efforts
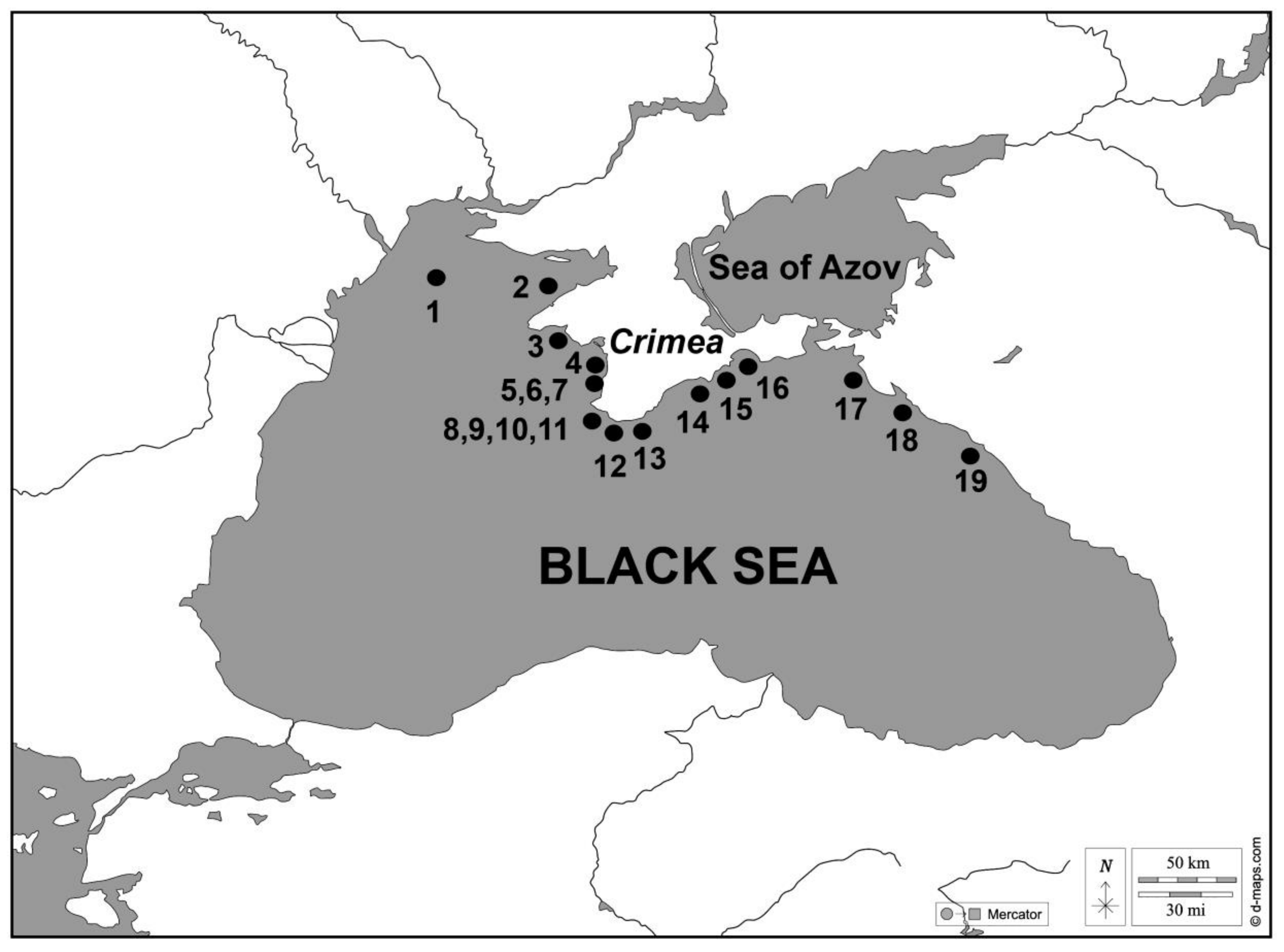
Material for the study of benthic diatoms was sampled at 1 site in the NWBS area (the Zernov Phyllophora field), 15 sites off the Black Sea CC, and 3 sites in the NEBS area at depths ranging from 0.5 to 75 m during a period from 1984 to 2016 (Figure 1). The map presented in Figure 1 is based on data from [©d-maps.com, accessed on 5 March 2022] and has been edited using Adobe Photoshop©. A total of 4 to 37 samples were taken from each site, with 2 replicates each. Overall, 534 samples were processed. Additionally, 6056 microphotographs were taken using a light microscope, and 1365 were taken using scanning electron microscopy (SEM) [13] (Table 1). Sampling was performed by the author and her colleagues from the Benthos Ecology Department of A.O. Kovalevsky Institute of Biology of Southern Seas Russian Academy of Sciences (IBSS RAS), Sevastopol, Russia.
2.2. Biomaterial Analysis
Samples from different types of soft-bottom substrates were collected using a Petersen grab-sampler (0.05 m2) or a sampler for meiobenthos (16 cm2). Substrates were initially treated in an ultrasonic bath for 20 min. Diatom valves were then cleaned using the standard technique, which involves the use of HCl and H2SO4 acids with K2Cr2O7 addition followed by rinsing several times with distilled water. A detailed description of the sampling and processing techniques used for biomaterial is provided in [12,13].
2.3. Microscopy
For light microscopy (LM), cleaned valves were mounted using Meltmount® or Naphrax® [12]. For scanning electron microscopy (SEM), a drop of cleaned valves was applied on Nuclepore Whatman membranes, mounted onto aluminium stubs, and coated with gold-palladium sputter. Samples were treated at the IBSS RAS (Sevastopol, Russia). Light microscopy observations were performed using a Nikon Eclipse 600 equipped with a PlanAPO 100× (Institute of Marine Sciences, University of Szczecin, Szczecin, Poland). SEM micrographs were taken using both a Hitachi S4500 (Goethe University, Frankfurt am Main, Germany) and Hitachi SU3500 (IBSS RAS, Sevastopol, Russia).
The field studies and sampling were performed in non-restricted and freely accessible areas of the Black Sea. Notably, we did not require any permission from the authorities to carry out our research activities.
2.4. Deposited Material
Ethanol-fixed samples, permanent glass slides, and stubs were stored in the respective collections of Dr. Sci. Elena Nevrova (Russia), Prof. Dr. Sci. Andrzej Witkowski (Poland), and Prof. Dr. Sci. Horst Lange-Bertalot (Germany).
2.5. Bacillariophyta Database
To compare the structure of benthic diatom assemblages and the interregional taxonomical diversity in the Black Sea’s northern shelf, a database for benthic Bacillariophyta was constructed using Microsoft Office Access [13]. Multiple changes in the taxonomy of Bacillariophyta and an increase in heterogeneous data meant that combining and revising the material on Black Sea Bacillariophyta was required. In this regard, the inventory of benthic diatoms in the Black Sea combines available sources on five regions of the Black Sea’s northern shelf: the Bulgarian coast (BC), the Romanian coast (RC), the northwestern part of the Black Sea (NWBS), the northeastern part of the Black Sea (NEBS), the Crimean coast (CC). These data were combined with the results of our own studies (1985–2016). The literature data were derived from over 60 publications on the northern shelf of the Black Sea [11,14,15,16,17,18,19,20].
Only the benthic and benthoplanktonic species were included in the database. Generally, planktonic species (e.g., belonging to the genera Chaetoceros Ehrenb., Detonula Schutt, Ditylum Bailey, Leptocylindrus Cl., Pseudosolenia Sundström, Skeletonema Grev., etc.) were not considered. Moreover, the master list did not include more than 100 species and ISTs found only once in one of numerous NWBS estuaries with increased salinity or freshwater ones [17]. Such selectivity was necessary for a more appropriate assessment of the interregional diversity of benthic diatoms [13].
Unfortunately, most of the publications on the Black Sea’s Bacillariophyta do not provide images. Exceptions include the works of C.S. Merezhkovsky, A.I. Proshkina-Lavrenko, and N.E. Guslyakov [17,19,20], which include line drawings or microphotographs of diatoms. In this regard, the question of the existence of some benthic diatom species in the Black Sea remains open. Considering the above, we were forced to take the following path: all benthic diatom findings were included in the master list, and this meant we had to forego the chance to compare images with iconography or our own photographs.
The checklist of Black Sea Bacillariophyta species was based on the taxonomic system of Round, Crawford, and Mann [36], with recent additions [10,12,37,38,39,40,41]. The nomenclature citations of taxa and validity of taxonomic names and synonyms were prepared in accordance with the authors of [10,37,38,39,40,41,42,43]. A taxonomic tree of the diatom flora of the Black Sea’s northern shelf was visualised using Dendroscope3 [44].
2.6. Statistical Analysis
To facilitate a comparative assessment of diatom taxonomic diversity for the five aforementioned regions of the Black Sea’s northern shelf, a taxonomic distinctness indices (TaxDI) were calculated using PRIMERv5 software [45], namely: the average taxonomic distinctness index (AvTD; Δ+) and variability index (VarTD; Λ+). The Δ+ index characterises the vertical taxonomic evenness of the diatom taxonomic tree along ascending levels of hierarchy. The Λ+ index presents the horizontal asymmetry of the taxonomic tree relative to the average expected level (i.e., different representations of lower taxa in higher ones within individual branches of the tree closing at different hierarchical levels) [30,31].
3. Results and Discussion
An updated inventory of the Black Sea’s northern shelf benthic diatoms from the five aforementioned regions (Bulgarian, Romanian, northwestern, Crimean, and northerneastern coasts), which hold 1100 species and infraspecific taxa (ISTs) belonging to 953 species, 149 genera, 61 families, 31 orders, and 3 classes of Bacillariophyta, according to the authors of [36,37,38,39,40,41,42,43], is shown in Table 2. A checklist of benthic diatoms found off the Black Sea’s northern shelf is provided in the Supplementary Materials File S1.
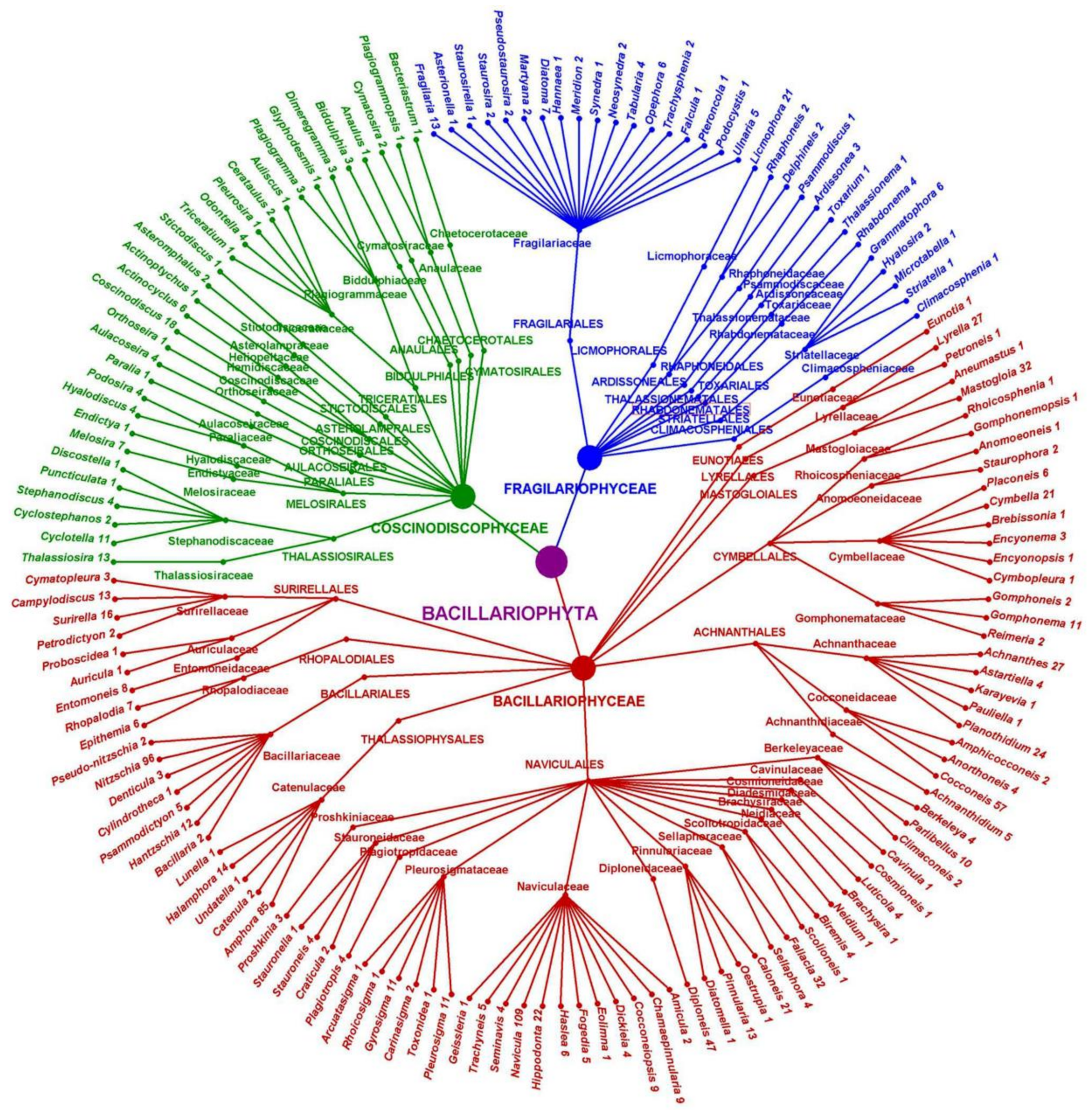
For a visualisation of the architectonics of the Black Sea benthic diatom hierarchical tree, a cyclogram was constructed [44] (Figure 2).
The highest species richness of diatoms ever marked in the Black Sea’s northern shelf was registered off the CC and accounts for 80.2% of the total number of benthic diatom flora. When our results were combined with those from available sources, a total of 882 species and ISTs belonging to 132 genera, 56 families, and 29 orders were observed near the CC. Our own data from the investigated sites along the CC stated that there were 793 species and ISTs pooled into 736 species, 130 genera, 53 families, 27 orders, and 3 classes of Bacillariophyta (Bacillariophyceae: 10 orders; Coscinodiscophyceae: 9 orders; Fragilariophyceae: 8 orders). In other Black Sea regions, the total species richness was much lower. For example, NWBS had 556 species and ISTs (50.5%, excluding species that are inhabitants of brackish-water and hypersaline estuaries), BC had 270 (24.5%), RC had 357 (32.5%), and NEBS had 309 (28.1%) [13]. The low species richness of benthic diatoms found off the BC in comparison to other regions can be explained by the fact that this area is understudied. Notably, the microphytobenthos survey was only performed once along the coastline [14] and was not continued.
Our evaluations based on the Bray–Curtis similarity coefficient indicate that the highest diatom species resemblance was registered between the following region pairs: NWBS–CC (61.1); NWBS–RC (59.9). The lowest resemblance values were detected for the following region pairs: CC–BC (36.3); CC–RC (41.8).
The genera Navicula (109 species and ISTs), Nitzschia (99), Amphora (87), Cocconeis (57), Diploneis (47), Fallacia (33), Mastogloia (31), Lyrella (27), Achnanthes (27), Planothidium (24), Caloneis (21), Licmophora (21), and Cymbella (19) were the most prevalent in the Black Sea benthic diatom flora. Among benthic diatoms, the highest diversity levels at the genus-level and species-level were the polyspecies families Naviculaceae (12 genera/180 species and ISTs), Bacillariaceae (7/124), Catenulaceae (5/106), Cocconeidaceae (3/63), Achnanthaceae (5/58), Fragilariaceae (18/54), Diploneidaceae (1/47), Pinnulariaceae (4/37), Sellaphoraceae (37/2), and Cymbellaceae (36/6). They formed the most saturated polyspecies branches on the hierarchical tree of the Black Sea benthic diatom flora.
During our research, seven new-to-science species were found, with seven new taxonomic combinations being proposed herein: Navicula parapontica Witkowski, Kulikovskiy, Nevrova et Lange-Bert.; N. pontica (Mereschk.) Witkowski, Kulikovskiy, Nevrova et Lange-Bert. [40]; N. petrovii Nevrova, Witkowski, Kociolek et Lange-Bert. [41]; Lyrella abruptapontica Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. pontieuxini Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. ruppelii Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. karayevae Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. pseudolyra Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. dilatata (A.W.F. Schmidt) Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. aestimata (Hust.) Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. rudiformis (Hust.) Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. granulata (Grunow) Nevrova, Witkowski, Kulikovskiy et Lange-Bert.; L. rattrayi (Pant.) Nevrova, Witkowski, Kulilovskiy et Lange-Bert.; L. bacillifera (Pant.) Nevrova, Witkowski, Kulilovskiy et Lange-Bert. [46].
Eight genera new to the Black Sea flora were found: Amicula (Witkowski) Witkowski; Astartiella Witkowski, Lange-Bert. et Metzeltin; Chamaepinnularia Lange-Bert. et Krammer; Cocconeiopsis Witkowski, Lange-Bert. et Metzeltin; Eolimna Lange-Bert. et W. Schiller; Lunella P.J.M. Snoeijs; Rhoicosigma Grunow; Trachysphenia P. Petit [13]. In total, more than 280 newly found species were noted for the Black Sea diatom flora. We failed to identify several taxa at the species-level. However, these species were included in the master list due to their morphological differences from known ones and were documented by taking microphotographs. We will refrain from describing new species until materials relating to various populations accumulate and are studied in detail [13].
The great number of new findings on the Black Sea diatom flora can be explained by the following factors: First, the diverse habitat conditions off the CC (silty-sandy substrates, rocks, boulders covered with macrophytes, etc.) provide many micro-niches for the successful survival of species introduced by ballast waters or as part of ship fouling from various regions of the world’s oceans. The intensive process of Mediterranesation, which is significantly contributing to the replenishment of the flora and fauna of the Black Sea, should also be considered [13,17,47]. It is assumed that a small part of the Black Sea Bacillariophyta is represented by the relics of Ponto-Caspian or Karangate flora that survived drastic changes in the salt regime during the Quaternary period. A more significant part of Black Sea diatom diversity consists of successfully acclimatised freshwater species, species introduced from the Mediterranean basin, and a small but constantly growing group of invasive species recently entered into the Black Sea that is adapting to its various ecological habitat conditions. It is highly likely that some of the species observed are native forms not previously registered in the Black Sea due to rarity or small cell size, as well as the fact that, overall, benthic Bacillariophyta are poorly studied. However, a conclusion on whether the floristic finds of Bacillariophyta are autochthonous or invasive forms can only be made after analysing the phylogenetic relationships and population genotypes [47].
The most common and significant species that contribute to the majority of the structural peculiarity of benthic diatom assemblages in the Black Sea’s northern shelf include Actinocyclus subtilis (W. Greg.) Ralfs ex A. Pritch., Amphora proteus W. Greg., Ardissonea crystallina (C. Agardh) Grunow, Bacillaria paxillifera (O. F. Müll.) Hendey, Berkeleya rutilans (Trentep.) Grunow, Caloneis liber (W. Sm.) Cleve, Campylodiscus thuretii Bréb., Cocconeis pseudocostata O.E. Romero, Cocconeis scutellum Ehrenb., Cylindrotheca closterium (Ehrenb.) Reimann et Lewin, Diploneis smithii (Bréb.) Cleve, Entomoneis paludosa (W. Sm.) Reimer, Grammatophora marina (Lyngb.) Kütz., Halamphora coffeaeformis (C. Agardh) Levkov, Hyalodiscus scoticus (Kütz.) Grunow, Licmophora abbreviata C. Agardh, Licmophora flabellata (Carmichael ex Grev.) C. Agardh, Microtabella delicatula (Kütz.) Round, Navicula parapontica Witkowski, Kulikovskiy, Nevrova et Lange-Bert., Navicula perminuta Grunow ex Van Heurck, Navicula ramosissima (C. Agardh) Cleve, Nitzschia coarctata Grunow, Nitzschia compressa (J.W. Bailey) Boyer, Nitzschia inconspicua Grunow, Nitzschia hybrida Grunow, Nitzschia longissima (Bréb. ex Kütz.) Grunow, Nitzschia sigma (Kütz.) W. Sm., Plagiotropis lepidoptera (W. Greg.) Kuntze, Pleurosigma elongatum W. Sm., Striatella unipunctata (Lyngb.) C. Agardh, Tabularia tabulata (C. Agardh) P.J.M. Snoeijs, Thalassiosira eccentrica (Ehrenb.) Cleve, Trachyneis aspera (Ehrenb.) Cleve (Figure 3A–F’).
A total of 1100 benthic diatom species and ISTs, all found in the Black Sea, were divided into five groups according to how frequently they were mentioned in the literature. Group I consists of 120 species and ISTs found in each of the five BS regions (100% occurrence) and stably present in the regional flora of Bacillariophyta. Groups II, III, and IV comprise species found in four, three, and two regions, respectively. These diatom species are common for diatom taxa of the Black Sea’s northern shelf. Group V is the most numerous and accounts for 517 species, with ISTs being mentioned only once in one of the five regions [13]. Among them, some species were quite abundant in some studied biotopes but not found anywhere else. However, the vast majority of Group V’s representatives are singleton, rare, relic, or invasive species [47].
Conclusions about the endemicity of species from Group V or their confinement to specific habitats can only be made with the expansion of the overall scope and geography of benthic diatom studies of the whole Black Sea basin. Similar conclusions concerning invasive species can also hardly be made due to the small cells of diatoms and insufficient research on the Black Sea basin as a whole. Thus, it cannot be confidently stated that these species did not exist in the Black Sea in the past, nor can it be certainly told that these species were introduced from other oceans in the world.
Thus, approximately 50% of the Black Sea benthic diatom flora consists of sporadic taxa. Among them, as well as among rare species, a group of 33 species belonging to mono- and oligospecies taxa, i.e., to only one branch in the hierarchical tree (one genus, family, or even order), was identified. Each of these species has a high taxonomic exclusivity rank because if the species disappears from the whole regional flora, the corresponding phylogenetic branch will also become extinct. There are 21 such species from the class Coscinodiscophyceae, 10 from Fragilariophyceae, and 2 from Bacillariophyceae (Table 3).
Among the species found only once in one of the five regions, 11 region-specific species belonging to mono- and oligospecies taxa were highlighted and assigned the highest rank of taxonomic exclusivity. Six such species were found off the CC, three off the RC, and two off the NWBS (Table 4). Notably, the species Orthoseira roeseana (Rabenh.) O’Meara and Hannaea arcus (Ehrenb.) Patrick were registered off the RC [15,16] and are freshwater-brackish forms. Therefore, they had a low abundance in the marine microphytobenthos, do not have the highest rank, and are not considered critical to the Black Sea diatom flora.
As previously noted, the elimination of species with the highest rank of taxonomic exclusivity from the regional flora can lead to the disappearance of an entire phylogenetic branch (including genera, families, or orders with specific genomes) from the Black Sea flora, which would cause significant changes in diatom assemblage structure in the region and the architectonics of the hierarchical tree of the Black Sea Bacillariophyta in general. These changes can be assessed based on the TaxDI index (AvTD) and its variability (VarTD) [30,31].
Based on the master list of species from five regions of the Black Sea’s northern shelf, an interregional analysis of the taxonomic diversity and structure of diatom taxocenes was performed, and the average value of the TaxDI index (Δ+) and its variability (Λ+) was calculated. Additionally, the deviation of this indices from the average expected value for the entire Black Sea was estimated [13].
The values of the Δ+ (Figure 4) and Λ+ (Figure 5) indices are presented on graphs within a 95% probability funnel projected onto a plane and narrowing in the direction of increasing the species number. The point’s location in relation to the funnel boundaries characterises the reliability of deviation in the TaxDI value from the average expected level calculated for the master list. The probability funnel outlines the limits of changes in average taxonomic distances calculated between every two species in the diatom taxocene, drawn randomly with 1000-fold pairwise samples for each subset of species.
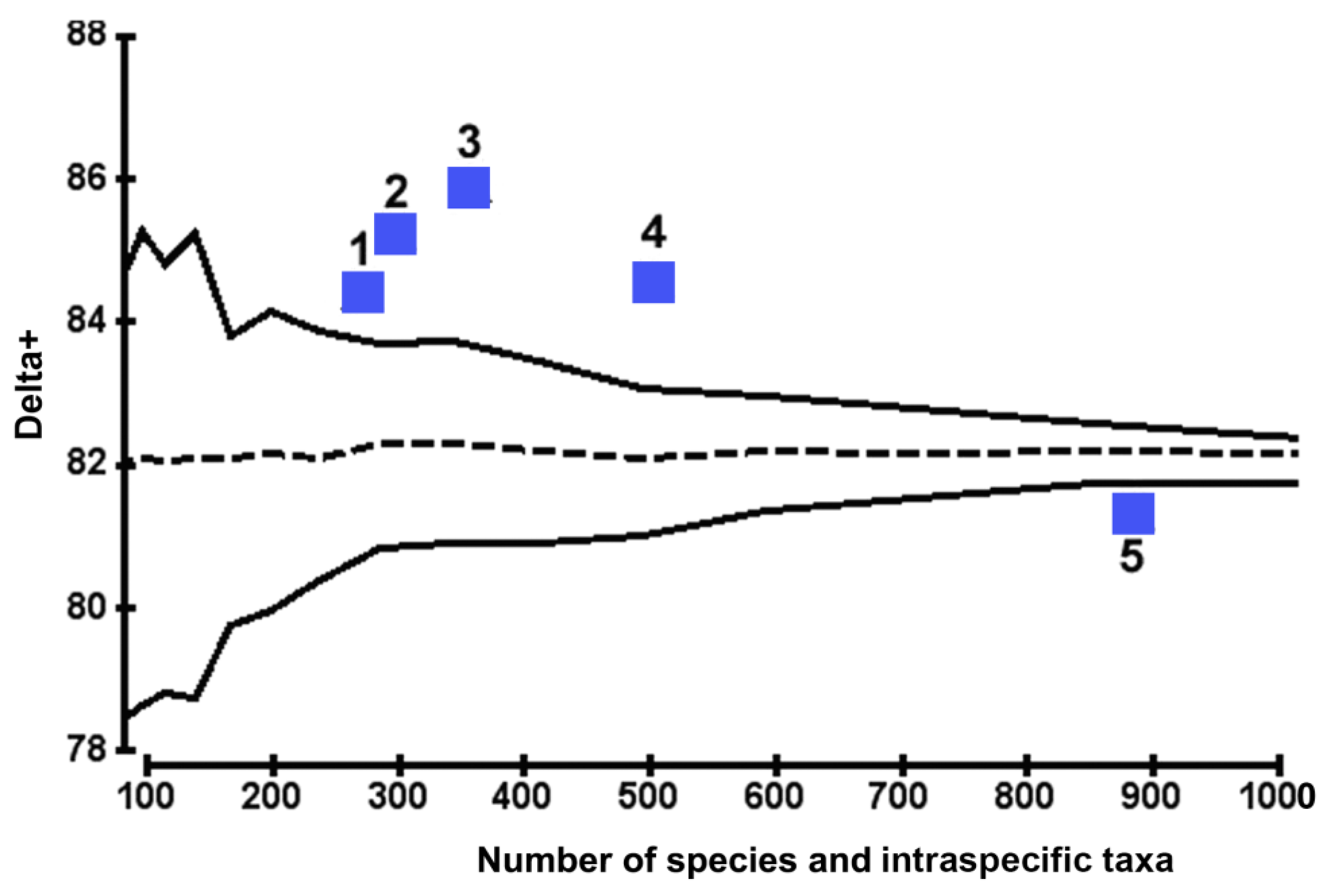
The calculated value of the AvTD index for the diatom assemblage found off the CC was the lowest (81.38 ± 0.2) and almost approached the average expected level for the whole Black Sea Bacillariophyta flora (Δ+ = 82.09). This index value and its location on the graph were due to the large proportion of polyspecies branches in the diatom hierarchical tree, which led to a decrease in vertical evenness in the diatom taxocene structure of the CC region (see Figure 4).
Simultaneously, the Δ+ values for other regions were much higher (NWBS: 84.07 ± 0.2; BC: 84.45 ± 0.6; NEBS: 85.1 ± 0.2), with a maximum (for RC: 85.9 ± 0.2) significantly exceeding the upper limit of the 95% probability funnel (see Figure 4). This fact indicates a large share of oligospecies branches closing at the taxonomic levels of genus or family, resulting in a more flattened model of the taxonomic structure of diatom assemblages found off the coasts of Romania, Bulgaria, and the northeastern part of the Black Sea.
Regarding the structure of diatom assemblages found off the RC, in comparison with other regions, oligospecies taxonomic groups were more represented, thereby affecting the increase in AvTD values. Additionally, in comparison with the average expected level, the higher evenness of the hierarchical structure of diatom assemblages found off the RC was also determined by the large share of species belonging to the IV–V group of occurrences, which formed monotaxon branches sequentially aggregating into one genus, family, or order. These species that are located off the RC are those with a high taxonomic exclusivity rank (i.e., Aulacoseira distans (Ehrenb.) Simonsen, A. granulata (Ehrenb.) Simonsen, A. islandica (O. Müll.) Simonsen, A. italica (Ehrenb.) Simonsen, Asteromphalus flabellatus (Bréb.) Grev., A. robustus Castracane, Cerataulus turgidus Ehrenb., Pleurosira laevis (Ehrenb.) Compère, Endictya oceanica Ehrenb., Biddulphia vesiculosa (C. Agardh) Kütz.) and the highest taxonomic exclusivity (i.e., Orthoseira roeseana (Rabenh.) O’Meara, Hannaea arcus (Ehrenb.) R.M. Patrick, Cavinula lacustris (W. Greg.) D.G. Mann et A. Stickle).
In particular, such monospecies branches are considered the highest conservation priority since their disappearance can lead to the loss of an entire taxon of a high rank (i.e., a family or even an order), which can significantly impoverish the hierarchical tree of regional flora and reduce the Δ+ value for the region. However, as previously noted, the species Orthoseira roeseana and Hannaea arcus were recorded off the RC and do not develop abundantly in the marine microphytobenthos since they are freshwater-brackish forms.
Generally, the taxonomic tree structure of the diatom assemblage found off the RC was characterised by high evenness along with wide variability, which is uncommon. Usually, the relative positions of points Δ+ and Λ+ on such graphs ‘mirror’ each other in nature (see the example of the BC).
Similar trends were revealed for the NEBS, for which the point of Δ+ was also located on the probability funnel boundary (see Figure 4). Such a pattern may be due to the presence of the following diatom species forming monotaxon branches in the regional flora: Asteromphalus flabellatus (Bréb.) Grev.; Cymatosira belgica Grunow; Biddulphia rostrata var. alata Proshk.-Lavr.; Delphineis surirella (Ehrenberg) G.W. Andrews. To study the change in the Δ+ index depending on the presence of species with a high rank in terms of exclusivity in the diatom taxocene structure, the model calculations were carried out [48]. It was shown that with the disappearance of only four of the aforementioned species from the NEBS, the calculated value of AvTD would decrease to 84.8. If we consider that for NEBS, the 95% probability range of calculated Δ+ values for a 1000-fold sample of random pairs of ISTs is within ± 0.2, then a decrease in AvTD will lead to a significant decline in taxonomic diversity and, as a consequence, to a simplification in the hierarchical tree structure of regional diatom flora.
Thus, the presence or absence of the few species forming monotaxon branches and with the highest taxonomic exclusivity rank has a significant impact on the hierarchical tree structure, changing the position of the regional Δ+ point in relation to the probability funnel boundaries on the graph. The ratio of branches with different species saturations on the diatom hierarchical tree of RS makes it distinctive in terms of the components of evenness (Δ+) and variability (Λ+) of taxonomic structure, both in comparison with other regions and with the average expected values of these indicators calculated for the entire Black Sea’s northern shelf.
Values of Δ+ corresponding to the BC (84.45) and NWBS (84.07) are also located on the graph near or slightly above the probability funnel boundary. For the CC, the value of Δ+ (81.38) on the graph is below the funnel boundary and near the average expected level for the Black Sea flora. This implies that the evenness of the diatom assemblage structure of the CC is characterised by values close to the average expected level for the entire Black Sea flora. Along the hierarchical tree, beginning from ISTs, there is the proportionality of the share of lower taxa to the higher ones, with consecutive aggregation to the higher hierarchical levels.
The resulting model of taxonomic diversity is due to the maximum species saturation in the diatom structure of CC (species/genus = 7), in contrast to the minimum value of the RC (4.2). Notably, the presence (or disappearance) of mono- and oligospecies taxa in some regions entails a sufficient shift of points on the Δ+ index graph; however, the variability of the taxonomic structure (Λ+) changes insignificantly.
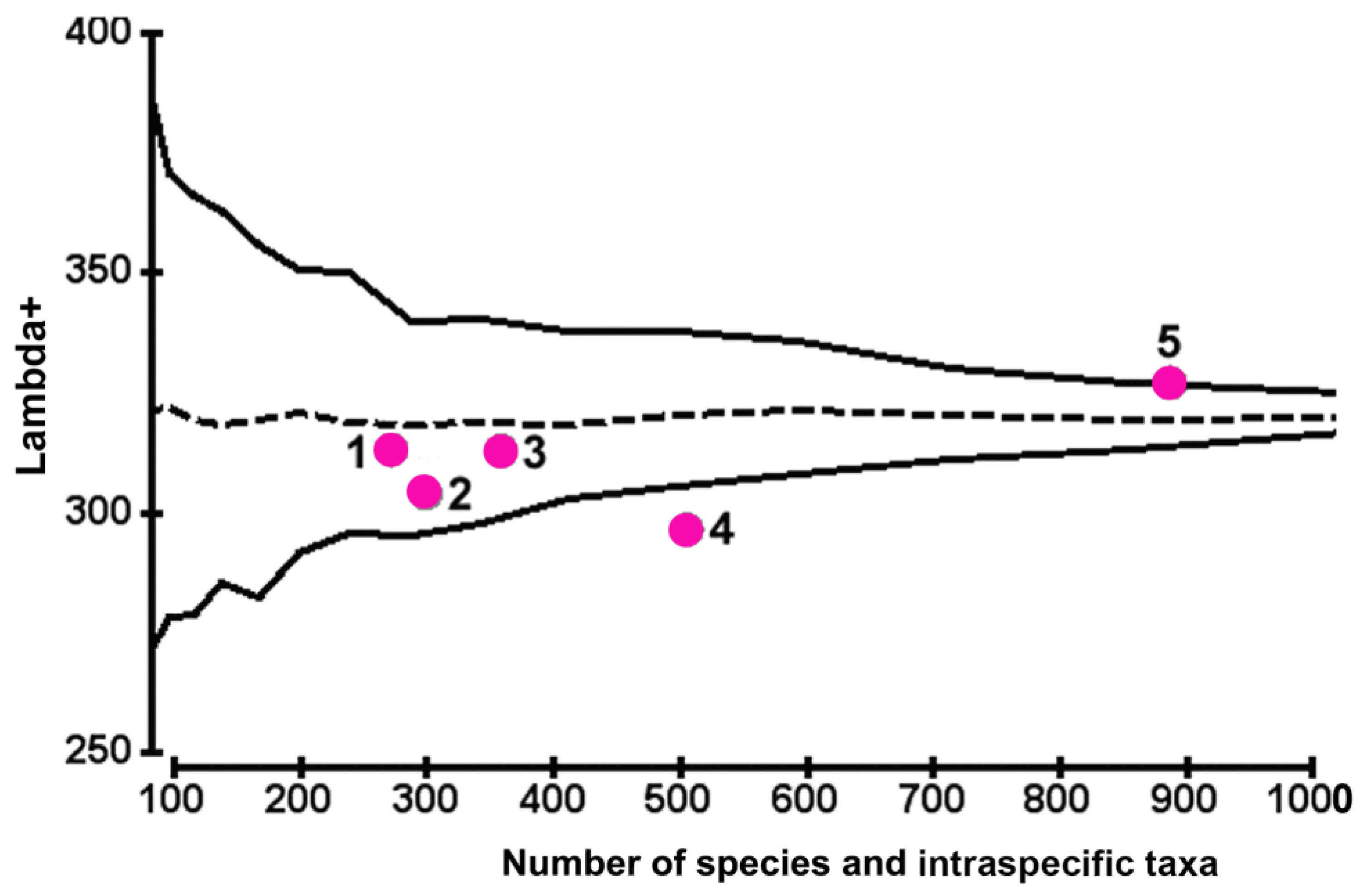
When assessing the variability of taxon distribution along the hierarchical tree, only the points of Λ+ (corresponding to the BC, RC, NWBS, and CC in the graph; Figure 5) were located near the average expected level. Such results imply that the character of aggregation of lower taxa into higher-rank taxa for the above regions is rather similar to the character of taxa aggregation along the hierarchical tree, which corresponds to the general master list for the entirety of Bacillariophyta flora in the Black Sea’s northern shelf.
The Λ+ index, which characterises the variability of TaxDI, was within the 95% funnel for the most studied regions (excluding the NWBS). The low Λ+ value (293.0 ± 0.2) may indicate an insignificant variability in the taxonomic diversity of benthic diatom taxocene in this region. For CC, the Δ+ value (327.13 ± 4.2) was near the upper limit of 95% probability, indicating a wide variability in benthic diatom taxonomic diversity off the Crimean sublittoral area. Our calculations indicate that when considering a small number of taxonomic levels (i.e., three to four), there is an inverse relationship between changes in Δ+ and Λ+ values. As a confirmation, a high value of Δ+ along with a minimum value of Λ+ was observed for the Crimean shelf.
The same trend of the ‘mirrored’ position of the Δ+ and Λ+ points on the plots was revealed for the NWBS and NEBS regions. However, in general, the character of the relationship between these indices is not always described by an inverse relationship and is mainly determined by the features of the hierarchical structure of the taxocene in each region. It has been suggested [49,50,51] that fluctuations in the Δ+ index usually reflect changes in the assemblage structure due to anthropogenic impacts and eutrophication, whereas the variability of the Λ+ index characterises the natural factors effect.
It should be noted that our interregional assessment of diatom species richness is based on the compilation of our own data with a known number of samples and data from other authors (presenting simple species lists). Such heterogeneity of primary data does not allow us to statistically assess the error of calculations of diversity in interregional comparisons. Considering the scarce knowledge of benthic diatoms diversity in the Black Sea regions (i.e., relatively small sampling efforts regarding the region total area consisting of ecologically heterogeneous habitats), we were forced to extrapolate the results of estimating the expected species richness of local samples to a higher regional level without assessing the reliability of samples belonging to a general population, as previously recommended [52]. Nevertheless, the analysis of the taxonomic diversity of benthic diatoms is based on a preliminary prognostic assessment under different habitat conditions and sample numbers at polygon and region scales [53,54,55]. In this case, the results of our calculations—which are based on a limited number of samples and within local seabed areas—are certainly open to further refinement upon new data on Bacillariophyta from unexplored regions being made available.
Notably, although the results of TaxDI analysis considering all benthic diatom species registered in the Black Sea flora allow us to identify the interregional differences in taxonomic structure, they do not provide explanations for the topological reasons leading to deviations in regional points’ Δ+ or Λ+ index values from the average expected level on the plot. Such analysis can be used for a comparative assessment of the long-term environmental impacts on the taxocene hierarchical diversity features. Thus, in the regions for which the values of Δ+ and Λ+ are significantly higher than the average expected level for the entire Black Sea, oligo- and monospecies branches in the diatom assemblage structure are more strongly represented. The presence of such branches can strengthen the component of tree topology vertical evenness, along with a wider variability of the structure due to the existence of polyspecies branches in the taxocene.
The peculiarities of such a taxocene, where the structure of region-specific or phylogenetically isolated species is represented more widely, were apparently caused by the influence of several multidimensional factors. Firstly, this includes stable and relatively homogeneous geomorphological features of the bottom landscape, which often leads to the narrowing of the diversity of ecological niches and the development of a significant number of stenobiont species. Areas with such conditions include the northwestern and northeastern parts of the Black Sea. In the regions of the BC, RC, and NWBS, the shelf is a shoal with a weak gradient and a flat relief that is almost evenly covered with silty-sandy bottom sediments typical of abrasion and accumulation zones and formed due to the outflow of large rivers (i.e., the Dnieper, Dniester, Bug, and Danube). In contrast, the narrow abrasion sublittoral zone has a sharp gradient formed by the rocky substrate of volcanic origin and is practically devoid of soft bottom due to strong underwater currents in the NEBS [56], which is where our work was carried out [13]. The exception is the Colchis Depression area, where the bottom was formed by the sediments of several rivers descending from the Caucasus Mountains [56]; however, this has not yet been explored by studies regarding Bacillariophyta [13].
The Black Sea coast of Crimea presents another picture. Here, many different forms of bottom topography and sediment types are noted [56], which leads to a greater diversification of ecological niches and, as a consequence, an increase in Bacillariophyta taxonomic diversity. In turn, the growth in diversity is characterised by the enhanced branching of the hierarchical tree of benthic diatom taxa, while most of the original taxa are locked on a phylogenetically common node corresponding to the genus or family levels [13,31,32,33,34]. Notably, the share of monotaxon branches in the structure of the hierarchical tree of CC may be reduced by the occurrence of closely related species in this area. In turn, the new finds of taxonomically isolated rare species in some areas of the CC can lead to an increase in the share of mono- and oligospecies branches in the tree architectonics. Thus, the hierarchical tree structure of benthic diatoms found off the CC can be characterised as being closest to the average expected structure of the Black Sea Bacillariophyta based on both the value of the AvTD index and its variability (VarTD) [13,31,32,33,34].
The aforementioned hypothesis agrees well with such well-known patterns of biodiversity formation as ‘the complexity of community decreases with deterioration of an environment quality’ [57] and ‘communities are diverse in spatially heterogeneous habitats’ [29]. In other words, a high-diversity habitat’s heterogeneity and a significant number of other factors are systematically associated with a high number of community elements and can be evaluated based on the analysis of these elements and factors of its evenness [29].
The factor of negative anthropogenic impact should also be considered, which can lead to the elimination of monotaxon branches. As previously mentioned, the disappearance of one species from a monospecies genus, family, or order leads to the loss of the entire taxonomic branch and greatly simplifies the structure of flora. Since such species and ISTs are characterised by being considered high-priority in terms of taxonomic exclusivity, their protection is important for saving the genetic pool of the taxon. In the case of benthic diatoms, one way to preserve high taxonomic diversity may be to maintain pristine ecological conditions and minimise negative anthropogenic effects on its habitats.
4. Conclusions
After compiling data on five regions of the Black Sea’s northern shelf (offshore from Bulgaria, Romania, NWBS, Crimea, and the NEBS) and comparing it to that of our studies (1984–2016), the checklist of Black Sea benthic diatoms has been revised and updated. At present, it combines 1100 species and ISTs belonging to 953 species, 149 genera, 61 families, 31 orders, and 3 classes of Bacillariophyta. The highest number of benthic diatom species was found off the CC and NWBS (about 80% and 50% of the total species richness, respectively). In other Black Sea regions, the species richness was much lower and ranged from 24% to 32%.
The maximum floristic pairwise similarity of benthic diatoms was found for CC–NWBS and NWBS–RC offshore, whilst the lowest similarity coefficient values were observed for CC–BC.
An interregional analysis revealed that the maximum value of the TaxDI index for diatom assemblages was noted off the RC, which is due to minimum species saturation in the taxocene structure. This fact led to an increased proportion of oligospecies branches in the taxonomical tree and resulted in a more flattened hierarchical pattern in the diatom diversity of this region. Simultaneously, the minimum value of Δ+ almost approached the average expected mode for the entire Black Sea northern shelf diatom flora, which was observed off the CC. This may provide evidence of a large proportion of polyspecies branches through to the maximum species saturation, resulting in a lower vertical evenness of the hierarchical tree.
A comparative evaluation of AvTD and VarTD parameters provided additional information on the taxonomic diversity of the Black Sea diatom flora, including the features of the aggregation of lower taxa into higher taxa along phylogenetic tree of Bacillariophyta.
Applying the TaxDI indices allowed us to identify (a) many more aspects of taxonomic diversity in the biotope based on the hierarchical tree of the benthic diatom taxocene, (b) the deviation of index values from the expected average level calculated for the Black Sea’s northern shelf as a whole, and (c) statistically and reliably assess regional differences in taxonomic structure using an array of historical data (mostly presented as simple species lists). The obtained results are recommended for use in the elaboration of comprehensive measures for biodiversity conservation in the context of the current use of natural resources in the Black Sea.
Supplementary Materials
Funding
This research was conducted within the framework of the following
IBSS state assignment: ‘Regularities of the formation and anthropogenic
transformation of the biodiversity and bioresources of the Azov-Black Sea Basin and
other areas of the World ocean (State Reg. NO. 121030100028-0).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author would like to thank her colleagues at the IBSS (Russia), namely, Alexei Petrov for helping with data processing and providing many years of cooperation and friendly support; Olga Akimova for her bibliographic assistance; and Vyacheslav Lishaev for performing SEM microphotography. The author is also sincerely grateful to Andrzej Witkowski and Horst Lange-Bertalot for supervising the manuscript and Manfred Ruppel for performing SEM microphotography. The author very appreciated the anonymous reviewers for improving the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Mann, D.G.; Crawford, R.M.; Round, F.E. Bacillariophyta. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C., Eds.; Springer: Cham, Switzerland, 2016; pp. 205–266. [Google Scholar] [CrossRef]
- Seckbach, J.; Kociolek, J.P. (Eds.) The Diatom World; Springer: Dordrecht, The Netherlands; Berlin/Heidelberg, Germany; London, UK; New York, NY, USA, 2011; 533p. [Google Scholar]
- Blanco, S.; Cejudo-Figueiras, C.; Tudesque, L.; Bécares, E.; Hoffmann, L.; Ector, L. Are diatom diversity indices reliable monitoring metrics? Hydrobiologia 2012, 695, 199–206. [Google Scholar] [CrossRef]
- Borja, A.; Elliott, M.; Andersen, J.H.; Cardoso, A.C.; Carstensen, J.; Ferreira, J.G.; Heiskanen, A.-S.; Marques, J.C.; Neto, J.M.; Teixeira, H.; et al. Good Environmental Status of marine ecosystems: What is it and how do we know when we have attained it? Mar. Pollut. Bull. 2013, 76, 16–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keck, F.; Rimet, F.; Franc, A. Phylogenetic signal in diatom ecology: Perspectives for aquatic ecosystems biomonitoring. Ecol. Appl. 2016, 26, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, M.; Devictor, V.; Schweiger, O. Phylogenetic diversity and nature conservation: Where are we? Trends Ecol. Evol. 2013, 28, 199–204. [Google Scholar] [CrossRef]
- Lavoie, I.; Hamilton, P.B.; Morin, S.; Tiam, S.K.; Kahlert, M.; Gonçalves, S.; Falasco, E.; Fortin, C.; Gontero, B.; Heudre, D.; et al. Diatom teratologies as biomarkers of contamination: Are all deformities ecologically meaningful? Ecol. Indic. 2017, 82, 539–550. [Google Scholar] [CrossRef]
- Rimet, F.; Bouchez, A. Biomonitoring river diatoms: Implications of taxonomic resolution. Ecol. Indic. 2012, 15, 92–99. [Google Scholar] [CrossRef]
- Stenger-Kovács, C.; Tóth, L.; Tóth, F.; Hajnal, E.; Padisák, J. Stream order-dependent diversity metrics of epilithic diatom assemblages. Hydrobiologia 2014, 721, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Tsarenko, P.M.; Wasser, S.P.; Nevo, E. (Eds.) Algae of Ukraine: Diversity, Nomenclature, Taxonomy, Ecology and Geography. 2. Bacillariophyta; A.R.G. Gantner Verlag K.G.: Ruggell, Liechtenstein, 2009; 413p. [Google Scholar]
- Ryabushko, L. Microalgae of the Black Sea Benthos; Kovalevsky Institute of Biology of the Southern Seas, NAS of Ukraine. EKOSI-Gidrophizica: Sevastopol, Russia, 2006; 143p, (In Russian, Abstract in English). [Google Scholar]
- Nevrova, E.L.; Snigireva, A.A.; Petrov, A.N.; Kovaleva, G.V. Guidelines for Quality Control of the Black Sea. Microphytobenthos; Gaevskaya, A.V., Ed.; N. Orianda: Sevastopol, Russia, 2015; 176p, (In Russian, Abstract in English). [Google Scholar] [CrossRef] [Green Version]
- Nevrova, E.L. Diversity and Structure of Benthic Diatom Taxocenes (Bacillariophyta) of the Black Sea; Gaevskaya, A.V., Ed.; A. O. Kovalevsky Institute of Biology of the Southern Seas RAS: Sevastopol, Russia, 2022; 329p, (In Russian, Abstract in English). [Google Scholar] [CrossRef]
- Konsulov, A. Black Sea Biological Diversity: Bulgaria; GEF Black Sea Environmental Series; Institute of Oceanology BAS, United Nations Publications: New York, NY, USA, 1998; Volume 5, 131p. [Google Scholar]
- Bodeanu, N. Structure et dynamique de l’algoflore unicellulaire dans les eaux du littoral Roumain de la mer Noire. Cercet. Mar. 1987, 20, 19–250. [Google Scholar]
- Petranu, A. Black Sea Biological Diversity: Romania; GEF Black Sea Environmental Series; Romanian Marine Research Institute, United Nations Publications: New York, NY, USA, 1997; Volume 4, 314p. [Google Scholar]
- Guslyakov, N.; Zakordonez, O.; Gerasimuk, V. Atlas of Diatom Algae of Benthos of the Black Sea North-West Part and Adjacent Aquatories; Naukova Dumka: Kiev, Ukraine, 1992; (In Russian, Abstract in English). [Google Scholar]
- Zaitsev, Y.; Alexandrov, B.G. Black Sea Biological Diversity: Ukraine; GEF Black Sea Environmental Series; NASU Odessa Branch Institute of Biology of the Southern Seas, United Nations Publications: New York, NY, USA, 1998; Volume 7, 351p. [Google Scholar]
- Proshkina-Lavrenko, A.I. Planctonic Diatoms of the Black Sea; AS USSR: Moscow, Russia, 1955; 222p, [Proshkina-Lavrenko, A.I. Diatomovyie Vodorosli Planktona Chernogo Morya; AN SSSR: Moskva, Russia, 1955; 222s]. (In Russian) [Google Scholar]
- Proshkina-Lavrenko, A.I. Benthic Diatoms of the Black Sea; AS USSR: Moscow-Leningrad, Russia, 1963; 243p. [Proshkina-Lavrenko A. I. Diatomovyie Vodorosli Bentosa Chernogo Morya; AN SSSR: Moskva, Russia, 1963; 243s]. (In Russian) [Google Scholar]
- Komahidze, A.; Mazmanidi, N. Black Sea Biological Diversity: Georgia; GEF Black Sea Environmental Series; United Nations Publications: New York, NY, USA, 1998; Volume 8, 167p. [Google Scholar]
- Öztürk, B. Black Sea Biological Diversity: Turkey; GEF Black Sea Environmental Series; United Nations Publications: New York, NY, USA, 1998; Volume 9, 114p. [Google Scholar]
- Microplankton Species of Turkish Seas. Ege University Faculty of Fisheries Department of Hydrobiology. Available online: http://plankweb.ege.edu.tr/ (accessed on 5 March 2022).
- Baytut, Ö.; Gönülol, A.; Koray, T. New records for marine phytoplankton of Turkish Seas from Southern Black Sea coasts. Ege J. Fish. Aquat. Sci. 2005, 22, 229–231. [Google Scholar]
- Baytut, Ö.; Moestrup, Ø.; Lundholm, N.; Gönülol, A. Contributions to the Diatom flora of the Black Sea from ultrastructural and molecular studies: New records of Skeletonema marinoi, Pseudo-nitzschia pungens var. aveirensis and Chaetoceros tenuissimus for the marine flora of Turkey. Nova Hedwig. 2013, 96, 427–444. [Google Scholar] [CrossRef]
- Dirican, S.; Kaleli, A.; Yilmaz, E.; Özer, A.; Dayioglu, H. New records of diatoms (Bacillariales, Rhopalodiales & Surirellales) with ultrastructure details from the Black Sea coast of Turkey. Aquat. Sci. Eng. 2022, 37, 19–28. [Google Scholar] [CrossRef]
- Kaleli, A.; Akçaalan, R. Checklist of marine diatoms from Turkish coastal waters with updated nomenclature. Aquat. Res. 2021, 4, 88–115. [Google Scholar] [CrossRef]
- Zidarova, R.; Ivanov, P.; Hristova, O.; Dzhurova, B.; Hineva, E. The unexpected diversity in Amphora sensu lato (Bacillariophyta) at Sozopol Bay, the western Black Sea. Phytotaxa 2022, 544, 103–127. [Google Scholar] [CrossRef]
- Protasov, A.A. Biodiversity and its assessment. Conceptual Diversicology; Institute of Hydrobiology of the National Academy of Sciences of Ukraine, Acadeperiodica of NAS of Ukraine: Kyiv, Ukraine, 2002; 106p, [Protasov, A.A. Bioraznoobrazie i Ego Otsenka. Kontseptualnaya Diversikologiya; In-t Gidrobiologii NAN Ukrainyi, Akademperiodika NAN Ukrainyi: Kyiv, Ukraine, 2002; 106s]. (In Russian) [Google Scholar]
- Warwick, R.M.; Clarke, K.R. Taxonomic distinctness and environmental assessment. J. Appl. Ecol. 1998, 35, 532–543. [Google Scholar] [CrossRef]
- Warwick, R.M.; Clarke, K.R. Practical measures of marine biodiversity based on relatedness of species. Oceanogr. Mar. Biol. Annu. Rev. 2001, 39, 207–231. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. A taxonomic distinctness index and its statistical properties. J. Appl. Ecol. 1998, 35, 523–531. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. The taxonomic distinctness measure of biodiversity weighting of step lengths between hierarchical levels. Mar. Ecol. Prog. Ser. 1999, 184, 21–29. [Google Scholar] [CrossRef]
- Clarke KR Warwick, R.M. A further biodiversity index applicable to species list: Variation in taxonomic distinctness. Mar. Biol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E Ltd.: Plymouth, UK, 2001; 154p. [Google Scholar]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; 747p. [Google Scholar]
- Levkov, Z. Amphora Sensu Lato, Diatoms of Europe; A.R.G. Gantner Verlag: Ruggell, Liechtenstein, 2009. [Google Scholar]
- Reid, G. A revision of the family Pleurosigmataceae. In Diatom Monographs; Witkowski, A., Ed.; A.R.G. Gantner Verlag K.G.: Ruggell, Liechtenstein, 2012; Volume 14, pp. 1–163. [Google Scholar]
- Witkowski, A.; Lange-Bertalot, H.; Metzeltin, D. Diatom Flora of Marine Coast 1. Iconographia Diatomologica. VII; A.R.G. Gantner Verlag K.G.: Ruggell, Liechtenstein, 2000; 926p. [Google Scholar]
- Witkowski, A.; Kulikovskiy, M.; Nevrova, E.; Lange-Bertalot, H.; Gogorev, R. The genus Navicula in ancient basins. I. Two novelties from the Black Sea. Plant Ecol. Evol. 2010, 143, 307–317. [Google Scholar] [CrossRef]
- Witkowski, A.; Nevrova, E.; Lange-Bertalot, H.; Kociolek, J. Navicula petrovii sp. nov. (Bacillariophyceae), a naviculoid diatom with amphoroid symmetry and its relationship to Navicula sensu stricto and other naviculoid genera. Nova Hedwig. 2014, 143, 469–484. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase; World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2023; Available online: http://www.algaebase.org (accessed on 5 March 2022).
- Fourtanier, E.; Kociolek, J. Catalogue of Diatom Names; California Academy of Sciences: San Francisco, CA, USA, 2011; Available online: http://researcharchive.calacademy.org/research/diatoms/names/index.asp (accessed on 5 March 2022).
- Dendroscope3 (An Interactive Viewer for Rooted Phylogenetic Trees and Networks). 2015. Available online: https://software-ab.informatik.uni-tuebingen.de/download/dendroscope3/welcome.html (accessed on 5 March 2022).
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; 126p. [Google Scholar]
- Nevrova, E.; Witkowski, A.; Kulikovskiy, M.; Lange-Bertalot, H.; Kociolek, J.-P. A revision of the diatom genus Lyrella Karayeva (Bacillariophyta: Lyrellaceae) from the Black Sea, with descriptions of five new species. Phytotaxa 2013, 83, 1–38. [Google Scholar] [CrossRef]
- Nevrova, E.L. Benthic diatoms of the Black Sea: Rare, invasive, extinct species and assessment of taxonomic exclusivity. Vopr. Sovrem. Algol. 2022, 2, 46–56, (In Russian, Abstract in English). [Google Scholar] [CrossRef]
- Nevrova, E.; Petrov, A. Taxonomic diversity of benthic diatoms of the Black Sea. In Microalgae of the Black Sea: Problems of Biodiversity, Preservation and Biotechnology; Tokarev, U., Gaevskaya, A., Eds.; NAS Ukraine, IBSS. Ekosi-Gidrophyzika: Sevastopol, Russia, 2008; pp. 60–84, (In Russian, Abstract in English). [Google Scholar]
- Leonard, D.; Clarke, K.; Somerfield, P.; Warwick, R. The application of an indicator based on taxonomic distinctness for UK marine biodiversity assessment. J. Environ. Manag. 2006, 78, 52–62. [Google Scholar] [CrossRef]
- Mouliott, D.; Gaillard, S.; Aliaume, C.; Veriaque, M.; Belsher, T.; Troussellier, M.; Chi, T. Ability of taxonomic diversity indices to discriminate coastal lagoon environmental based on macrophyte communities. Ecol. Indic. 2005, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Warwick, R.M.; Ashman, C.M.; Brown, A.R.; Clarke, K.R.; Dowell, B.; Hart, B.; Lewis, R.E.; Shillabeer, N.; Somerfield, P.J.; Tapp, J.F. Inter-annual changes in the biodiversity and commuity structure of the macrobenthos in Tees Bay and the Tees estuary, UK, associated with local and regional environmental events. Mar. Ecol. Prog. Ser. 2002, 234, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Urbach, V.Y. Biometric Methods; High School: Moscow, Russia, 1964; 416p, [Urbah, V.Yu. Biometricheskie Metodyi; Vysshaya Shkola: Moscow, Russia, 1964; 416s]. (In Russian) [Google Scholar]
- Petrov, A.N.; Nevrova, E.L. Evaluation of reproducibility and reliability of benthic diatoms species composition at coastal location of SW Crimea. Mar. Ecol. J. 2012, 11, 79–88, (In Russian, Abstract in English). [Google Scholar]
- Petrov, A.N.; Nevrova, E.L. Prognostic Estimation of Species Richness of Benthic Bacillariophyta. Int. J. Algae 2013, 15, 5–25. [Google Scholar] [CrossRef]
- Petrov, A.N.; Nevrova, E.L. Numerical analysis of the structure of benthic diatom assemblages in replicate samples (Crimea, the Black Sea). Nova Hedwig. 2014, 143, 245–253. [Google Scholar]
- Vylkanov, A.; Danov, X.; Marinov, X.; Vladev, P. Black Sea; Gidrometeoizdat: Leningrad, Russia, 1983; 408p, [Vylkanov, A.; Danov, X.; Marinov, X.; Vladev, P. Chornoe more: Sbornik; Gidrometeoizdat: Leningrad, Russia, 1983; 408s]. (In Russian) [Google Scholar]
- Alimov, A.F. Elements of the Theory of Ecosystem Functioning; ZIN RAN: St-Petersburg, Russia, 2000; 147p, [Alimov, A.F. Elementyi Teorii Funktsionirovaniya Ekosistem; ZIN RAN: St-Petersburg, Russia, 2000; 147s]. (In Russian) [Google Scholar]
- Petrova-Karadjova, V.I. Diatom algae of marine fouling. Izvestiya na Institut po Ribni Resursi 1977, 15, 55–65. [Petrova-Karadzhova V. I. Diatomovye vodorosli morskih obrastanij. Izvestiya na Instituta po Ribni Resursi 1977, 15, 55–65](In Russian) [Google Scholar]
- Cărăus, I. The Algae of Romania: A Distributional Checklist of Actual Algae [Version 2.3]; Studii si Cercetari Biologie, Universitatea Bacau: Bacau, Romania, 2012; 809p. [Google Scholar]
- Nevrova, E.L. Benthic diatoms (Bacillariophyta) at Zernov Phyllophora Field (Northern-Western part of the Black Sea): Taxonomic diversity and structure of taxocene. Mar. Ecol. J. 2014, 13, 47–58, (In Russian, Abstract in English). [Google Scholar]
- Ryabushko, L.I. Microphytobenthos of the Zernov Phyllophora Field; Academy of Sciences Ukrainian SSR, Kovalevsky Institute of Biology of the Southern Seas: Sevastopol, Russia, 1991; 28p, [Ryabushko, L.I. Mikrofitobentos Fillofornogo Polya Zernova; AN USSR, InBUM im. A. O. Kovalevskogo: Sevastopol, Russia, 1991; 28s]. (In Russian) [Google Scholar]
- Guslyakov, N.E.; Nevrova, E.L. Composition of Diatom Algae on Solid Substrates in the Sevastopol Bay Area; Ukrainian Academy of Sciences, A.O. Kovalevsky Institute of Biology of Southern Seas: Sevastopol, Russia, 1987; 14p, [Guslyakov, N.E.; Nevrova, E.L. Sostav Diatomovyh Vodoroslej na Tverdyh Substratah v Rajone Sevastopol’skoj Buhty; AN USSR, Institut biologii yuzhnyh morej im. A.O. Kovalevskogo: Sevastopol, Russia, 1987; 14s]. (In Russian) [Google Scholar]
- Kovalchuk, Y.L.; Nevrova, E.L.; Shalaeva, E.A. Diatom Fouling of Solid Substrates; Russian Academy of Sciences, A.N. Severtsov Institute for Ecology and Evolution, KMK: Moscow, Russia, 2008; 174p, [Kovalchuk, Yu.L.; Nevrova, E.L.; Shalaeva, E.A. Diatomovye Obrastaniya Tverdyh Substratov; Rossijskaya Akademiya Nauk, Institut Problem Ekologii i evolyucii im. A.N. Severcova; KMK: Moskva, Russia, 2008; 174s]. (In Russian) [Google Scholar]
- Kucherova, Z.S. Diatom Algae and Their Role in the Fouling Cenosis of the Black Sea. Ph.D. Thesis, Institute of Biology of Southern Seas, Sevastopol, Russia, 1973; 145p. [Kucherova, Z.S. Diatomovye Vodorosli i ih rol’ v Cenoze Obrastaniya Chornogo Morya. Dissertaciya na soiskanie uchenoj stepeni kandidata biologicheskih nauk, Institute of Biology of Southern Seas, Sevastopol, Russia, 1973; 145s](In Russian). [Google Scholar]
- Nevrova, E.L. Diatom algae of stony substrates of the Black Sea near Karadag (Crimea). Biol. Sci. 1991, 5, 79–86. [Nevrova, E.L. Diatomovye vodorosli kamenistyh gruntov Chornogo morya u Karadaga (Krym). Biol. Nauk. 1991, 5, 79–86].(In Russian) [Google Scholar]
- Nevrova, E.L. Benthic diatom algae in shallow water near Sevastopol (Black Sea). Algologia 1998, 8, 278–285. [Nevrova, E L. Donnye diatomovye vodorosli na melkovodye u Sevastopolya (Chornoe more). Algologia 1998, 8, 278–285](In Russian) [Google Scholar]
- Nevrova, E.L. Benthic diatom algae on soft bottom in the deep-water part of the mouth of Sevastopol Bay. Algologia 1999, 9, 43–54. [Nevrova, E.L. Donnye diatomovye vodorosli ryhlyh gruntov v glubokovodnoj chasti ustyia Sevastopolskoj buhty. Algologia 1999, 9, 43–54](In Russian) [Google Scholar]
- Nevrova, E.L. Structure and taxonomical diversity of benthic diatom at estuarines of Rivers Belbek and Chernaya (South-West Crimea, Ukraine). Algologia 2013, 23, 471–492, (In Russian, Abstract in English). [Google Scholar] [CrossRef]
- Nevrova, E.L. Taxonomic diversity and structure of benthic diatom taxocene (Bacillariophyta) at Sevastopol Bay (the Black Sea). Mar. Ecol. J. 2013, 12, 55–67, (In Russian, Abstract in English). [Google Scholar]
- Nevrova, E.L. Taxonomic diversity and environmental assessment of benthic diatoms at Balaklava Bay (South-Western Crimea, the Black Sea, Ukraine). Algologia 2014, 24, 47–66, (In Russian, Abstract in English). [Google Scholar] [CrossRef] [Green Version]
- Bukhtiyarova, L.N. Diatoms of Ukraine. Inland Waters; National Academy of Sciences of Ukraine, N.G. Kholodny Institute of Botany: Kyiv, Ukraine, 1999; 132p. [Google Scholar]
- Davidovich, N.A.; Gastineau, R.; Gaudin, P.; Davidovich, O.I.; Mouget, J.-L. Sexual reproduction in the newly-described blue diatom. Haslea Karadagensis Fottea 2012, 12, 219–229. [Google Scholar] [CrossRef]
Figure 1.
Sampling design of the Black Sea’s northern shelf: 1—Zernov Phyllophora field; 2—Karkinitsky Bay; 3—vil. Marjino area; 4—r. Belbek and reference site R3; 5, 6, 7—Karantinnaya Bay, Sevastopol Bay, Inkerman Bay; 8, 9, 10, 11—Omega Bay, Golubaya Bay, Cape Fiolent, Balaklava Bay, and reference site R6; 12—Laspi Bay; 13—Cape Sarych; 14—vil. Novyi Svet; 15—Karadag Natural Reserve; 16—Dvujakornaya Bay; 17—Anapa area; 18—Novorossiysk area; 19—Tuapse area.
Figure 1.
Sampling design of the Black Sea’s northern shelf: 1—Zernov Phyllophora field; 2—Karkinitsky Bay; 3—vil. Marjino area; 4—r. Belbek and reference site R3; 5, 6, 7—Karantinnaya Bay, Sevastopol Bay, Inkerman Bay; 8, 9, 10, 11—Omega Bay, Golubaya Bay, Cape Fiolent, Balaklava Bay, and reference site R6; 12—Laspi Bay; 13—Cape Sarych; 14—vil. Novyi Svet; 15—Karadag Natural Reserve; 16—Dvujakornaya Bay; 17—Anapa area; 18—Novorossiysk area; 19—Tuapse area.

Figure 2.
Taxonomic tree of the Black Sea’s northern shelf benthic diatom flora, from the division to genus level. The quantities of species and ISTs were enumerated.
Figure 2.
Taxonomic tree of the Black Sea’s northern shelf benthic diatom flora, from the division to genus level. The quantities of species and ISTs were enumerated.

Figure 3.
(A–F’). The most common species of benthic Bacillariophyta found off the Black Sea (SEM). (A). Thalassiosira eccentrica. (B). Actinocyclus subtilis. (C). Campylodiscus thuretii. (D). Halamphora coffeaeformis. (E). Amphora proteus. (F). Grammatophora marina. (G). Nitzschia compressa. (H). Caloneis liber. (I). Plagiotropis lepidoptera. (J). Entomoneis paludosa. (K). Hyalodiscus scoticus. (L). Bacillaria paxillifer. (M). Diploneis smithii. (N). Trachyneis aspera. (O). Cocconeis pseudocostata. (P). Cocconeis scutellum. (Q). Nitzschia coarctata. (R). Nitzschia inconspicua. (S). Navicula ramosissima. (T). Microtabella delicatula. (U). Berkeleya rutilans. (V). Navicula perminuta. (W). Licmophora abbreviata. (X). Navicula parapontica. (Y). Striatella unipunctata. (Z). Nitzschia hybrida. (A’). Pleurosigma elongatum. (B’). Tabularia tabulata. (C’). Licmophora flabellata. (D’). Ardissonea crystallina. (E’). Nitzschia longissima. (F’). Nitzschia sigma. Scale bar = 10 µm (A–M,Q,W,X–F’), 1 µm (O,P,R–V).
Figure 3.
(A–F’). The most common species of benthic Bacillariophyta found off the Black Sea (SEM). (A). Thalassiosira eccentrica. (B). Actinocyclus subtilis. (C). Campylodiscus thuretii. (D). Halamphora coffeaeformis. (E). Amphora proteus. (F). Grammatophora marina. (G). Nitzschia compressa. (H). Caloneis liber. (I). Plagiotropis lepidoptera. (J). Entomoneis paludosa. (K). Hyalodiscus scoticus. (L). Bacillaria paxillifer. (M). Diploneis smithii. (N). Trachyneis aspera. (O). Cocconeis pseudocostata. (P). Cocconeis scutellum. (Q). Nitzschia coarctata. (R). Nitzschia inconspicua. (S). Navicula ramosissima. (T). Microtabella delicatula. (U). Berkeleya rutilans. (V). Navicula perminuta. (W). Licmophora abbreviata. (X). Navicula parapontica. (Y). Striatella unipunctata. (Z). Nitzschia hybrida. (A’). Pleurosigma elongatum. (B’). Tabularia tabulata. (C’). Licmophora flabellata. (D’). Ardissonea crystallina. (E’). Nitzschia longissima. (F’). Nitzschia sigma. Scale bar = 10 µm (A–M,Q,W,X–F’), 1 µm (O,P,R–V).

Figure 4.
Assessment of benthic diatom diversity in various regions of the Black Sea’s northern shelf, conducted using the AvTD index (Δ+): 1—Bulgarian coast (BC); 2—northeastern part of the Black Sea (NEBS); 3—Romanian coast (RC); 4—northwestern part of the Black Sea (NWBS); 5—Crimean coast (CC).
Figure 4.
Assessment of benthic diatom diversity in various regions of the Black Sea’s northern shelf, conducted using the AvTD index (Δ+): 1—Bulgarian coast (BC); 2—northeastern part of the Black Sea (NEBS); 3—Romanian coast (RC); 4—northwestern part of the Black Sea (NWBS); 5—Crimean coast (CC).

Figure 5.
Assessment of benthic diatom diversity in different regions of the Black Sea’s northern shelf using the VarTD index (Λ+): 1—Bulgarian coast (BC); 2—northeastern part of the Black Sea (NEBS); 3—Romanian coast (RC); 4—northwestern part of the Black Sea (NWBS); 5—Crimean coast (CC).
Figure 5.
Assessment of benthic diatom diversity in different regions of the Black Sea’s northern shelf using the VarTD index (Λ+): 1—Bulgarian coast (BC); 2—northeastern part of the Black Sea (NEBS); 3—Romanian coast (RC); 4—northwestern part of the Black Sea (NWBS); 5—Crimean coast (CC).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sampling effort and species richness of benthic diatoms found off the Black Sea’s northern shelf.
Table 1.
Sampling effort and species richness of benthic diatoms found off the Black Sea’s northern shelf.
| Studied Area | Date | Coordinates | Depth, m | Number of Samples | Substrates | Number of Species and ISTs | |
|---|---|---|---|---|---|---|---|
| Northwestern part of the Black Sea | |||||||
| Zernov Phyllophora field | 5 November 2010 | 45°50′38″ N, 30°44′32″ E | 19–52 | 10 | Silty-sandy | 160 | |
| Black Sea Crimean coast | |||||||
| Karkinitsky Bay | 10 July 2008 | 45°34′31″ N, 32°48′22″ E | 0.5–1 | 4 | Silty-sandy | 22 | |
| vil. Marjino area | 20 July 2010 | 45°20′15″ N, 32°42′12″ E | 6 | 4 | Sandy | 140 | |
| r. Belbek area | 5 November 2009 | 44°39′45″ N, 33°32′31″ E | 6–19 | 20 | Sandy-silty | 243 | 279 |
| Reference site R3 | 5 November 2009 | 44°38′25″ N, 33°31′45″ E | 10–12 | 4 | Sandy | 119 | |
| Karantinnaya Bay | 25 August 1994 | 44°37′05″ N, 33°30′10″ E | 0.5–32 | 44 | Silty-sandy, rocky | 136 | |
| Sevastopol Bay | 11 July 2001 | 44°37′19″ N, 33°31′27″ E | 4–17 | 62 | Silty-sandy | 186 | |
| Inkerman Bay | 6 November 2009 | 44°36′20″ N, 33°35′50″ E | 3–10 | 12 | Silty-sandy | 116 | |
| Omega Bay | 28 July 2004 | 44°35′55″ N, 33°26′56″ E | 1.5–16 | 8 | Sandy-silty | 260 | |
| Golubaya Bay | 15 August 2009 | 44°35′03″ N, 33°22′48″ E | 1.5–6 | 6 | Sandy | 124 | |
| Cape Fiolent | 12 August 2009 | 44°30′53″ N, 33°28′18″ E | 1.5–12 | 16 | Sandy, macrophytes | 290 | |
| Balaklava Bay | 14 October 2006 | 44°29′12″ N, 33°36′53″ E | 6–20 | 34 | Silty-sandy | 191 | 342 |
| Reference site R6 | 14 October 2006 | 44°28′37″ N, 33°37′30″ E | 17–23 | 4 | Sandy | 233 | |
| Laspi Bay | 27 June 1996 | 44°25′10″ N, 33°42′27″ E | 0.5–52 | 74 | Silty-sandy, macrophytes | 217 | |
| Cape Sarych | 20 August 2007 | 44°23′14″ N, 33°44′17″ E | 3–5 | 4 | Sandy, pebbles | 82 | |
| vil. Novyi Svet | 14 August 2009 | 44°49′27″ N, 34°54′25″ E | 0.5–3 | 4 | Sandy | 93 | |
| Karadag Natural Reserve | 10 August 2010 | 44°54′53″ N, 35°13′51″ E | 5–8 | 6 | Sandy, pebbles | 300 | |
| Dvujakornaya Bay | 11 August 2008 | 44°59′28″ N, 35°22′04″ E | 2–9 | 22 | Sandy | 304 | |
| Northeastern part of the Black Sea | |||||||
| Anapa area and Novorossiysk area | 14 October 1999 | 44°52′08″ N, 37°17′44″ E | 0.5–75 | 190 | Silty-sandy, macrophytes | 225 | |
| Tuapse area | 20 August 1990 | 44°04′59″ N, 39°03′04″ E | 25–52 | 6 | Silty | 49 | |
| Total | 534 | ||||||
Table 2.
Representativeness of benthic Bacillariophyta in the Black Sea.
| Class | Order | Family | Genera | Species | ISTs |
|---|---|---|---|---|---|
| Coscinodiscophyceae | 12 | 18 | 30 | 91 | 105 |
| Fragilariophyceae | 9 | 10 | 31 | 88 | 101 |
| Bacillariophyceae | 10 | 33 | 88 | 774 | 894 |
| In total | 31 | 61 | 149 | 953 | 1100 |
Table 3.
Black Sea benthic diatom species with a high taxonomic exclusivity rank.
| Species | Genus | Family | Order | Class | Region of the Black Sea’s Northern Shelf |
|---|---|---|---|---|---|
| Actinoptychus senarius | 1 | 1 | 1 | Coscinodisco- phyceae | BC, NWBS, CC |
| Aulacoseira distans | 1 | 1 | 1 | RC | |
| Aulacoseira granulata | RC, NWBS, CC | ||||
| Aulacoseira islandica | RC | ||||
| Aulacoseira italica | RC, NWBS | ||||
| Asteromphalus flabellatus | 1 | 1 | 1 | RC, NEBS | |
| Asteromphalus robustus | RC, CC | ||||
| Anaulus minutus | 1 | 1 | 1 | BC, NWBS, CC | |
| Auliscus sculptus | 1 | 1 | 1 | NWBS, CC | |
| Cerataulus smithii | 1 | NWBS, NEBS, CC | |||
| Cerataulus turgidus | RC, CC | ||||
| Pleurosira laevis | 1 | RC, NWBS, CC | |||
| Triceratium antediluvianum | 1 | All five regions | |||
| Bacteriastrum hyalinum | 1 | 1 | 1 | NWBS, CC | |
| Paralia sulcata | 1 | 1 | 1 | All five regions | |
| Glyphodesmis distans | 1 | 1 | 1 | All five regions | |
| Cymatosira belgica | 1 | 1 | 1 | CC, NWBS, NEBS | |
| Plagiogrammopsis sp. | 1 | CC | |||
| Endictya oceanica | 1 | 1 | 1 | RC, NWBS, CC, NEBS | |
| Biddulphia rostrata | 1 | 1 | 1 | NWBS, CC, NEBS | |
| Biddulphia vesiculosa | RC | ||||
| Ardissonea crystallina | 1 | 1 | 1 | Fragilario- phyceae | All five regions |
| Ardissonea baculus | All five regions | ||||
| Ardissonea robusta | CC | ||||
| Climacosphenia moniligera | 1 | 1 | 1 | NWBS, BC | |
| Delphineis minutissima | 1 | 1 | 1 | NWBS, CC | |
| Delphineis surirella | NWBS, CC, NEBS | ||||
| Rhaphoneis amphiceros | 1 | CC, NEBS | |||
| Psammodiscus nitidus | 1 | 1 | 1 | All five regions | |
| Thalassionema nitzschioides | 1 | 1 | 1 | All five regions | |
| Toxarium undulatum | 1 | 1 | 1 | All five regions | |
| Anomoeoneis spaerophora | 1 | 1 | 1 | Bacillario- phyceae | NWBS, CC |
| Proschkinia complanatoides | 1 | 1 | 1 | NWBS, CC |
Note: BC—Bulgarian coast, RC—Romanian coast, NWBS—northwestern part of the Black Sea and adjacent estuaries and bays, NEBS—northeastern part of the Black Sea, CC—Crimean coast.
Table 4.
Region-specific diatom species with the highest rank of taxonomic exclusivity.
| Species | Genus | Family | Order | Class | Region of the Black Sea’s Northern Shelf |
|---|---|---|---|---|---|
| Brachysira aponina | 1 | 1 | 1 | Bacillariophyceae | NWBS |
| Proschkinia complanatula | 1 | 1 | NWBS | ||
| Cavinula lacustris | 1 | 1 | RC | ||
| Neidium binodis | 1 | 1 | CC | ||
| Amicula specululum | 1 | 1 | |||
| Astartiella bahusiensis | 1 | 1 | 1 | ||
| Astartiella producta | |||||
| Astartiella sp.1DV | 1 | ||||
| Pauliella taeniata | 1 | ||||
| Orthoseira roeseana | 1 | 1 | 1 | Coscinodiscophyceae | RC |
| Hannaea arcus | 1 | 1 | 1 | Fragilariophyceae |
Note: BC—Bulgarian coast, RC—Romanian coast, NWBS—northwestern part of the Black Sea and adjacent estuaries and bays, NEBS—northeastern part of the Black Sea, CC—Crimean coast.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nevrova, E. Taxonomic Structure and Diversity of Benthic Diatoms (Bacillariophyta) in the Northern Shelf of the Black Sea. Phycology 2023, 3, 337-355. https://doi.org/10.3390/phycology3030022
AMA Style
Nevrova E. Taxonomic Structure and Diversity of Benthic Diatoms (Bacillariophyta) in the Northern Shelf of the Black Sea. Phycology. 2023; 3(3):337-355. https://doi.org/10.3390/phycology3030022
Chicago/Turabian StyleNevrova, Elena. 2023. "Taxonomic Structure and Diversity of Benthic Diatoms (Bacillariophyta) in the Northern Shelf of the Black Sea" Phycology 3, no. 3: 337-355. https://doi.org/10.3390/phycology3030022