

A New Genus of the Microascaceae (Ascomycota) Family from a Hypersaline Lagoon in Spain and the Delimitation of the Genus Wardomyces


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Fungal Isolation
2.2. Phenotypic Study
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Phylogenetic Analysis
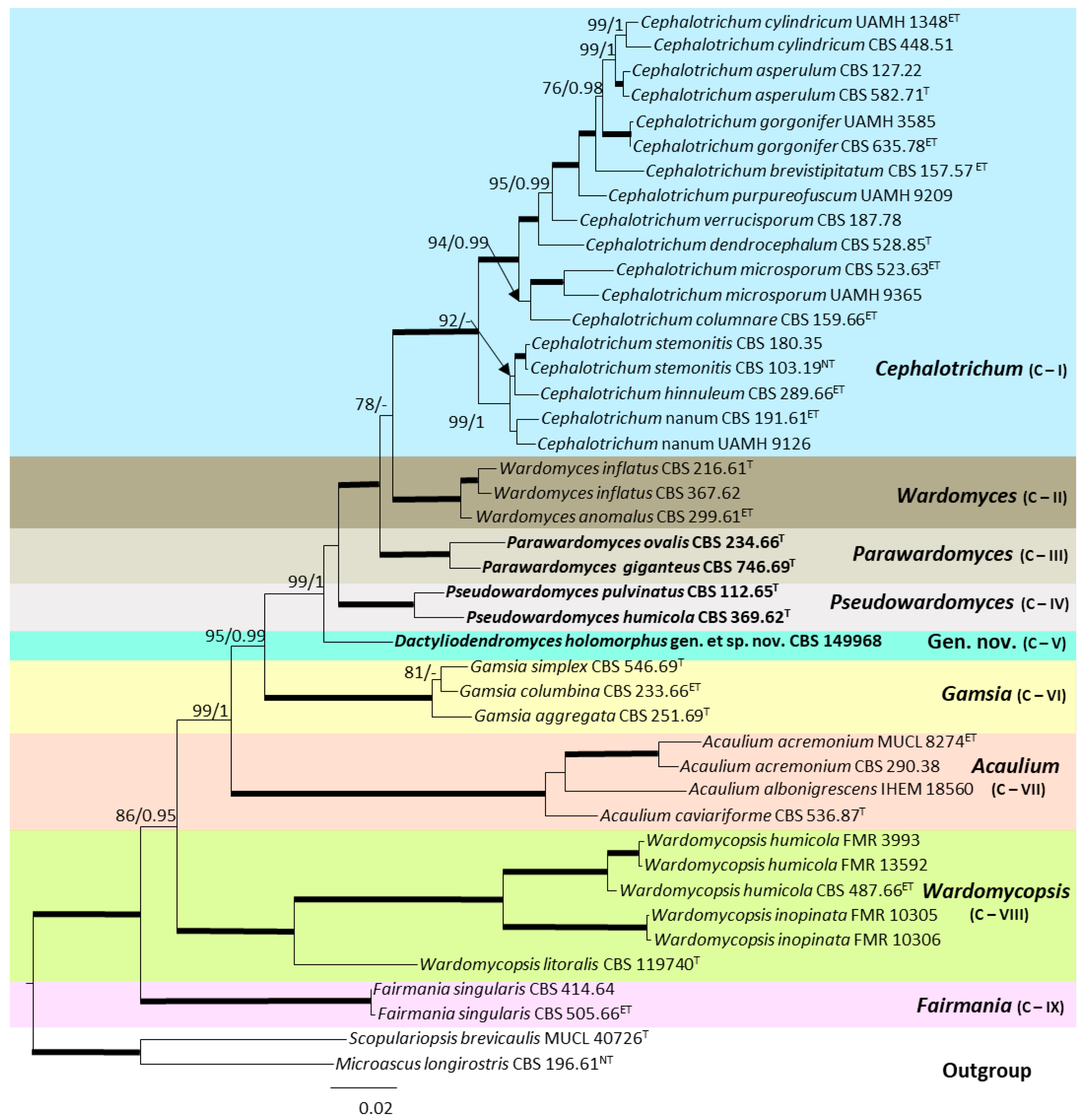
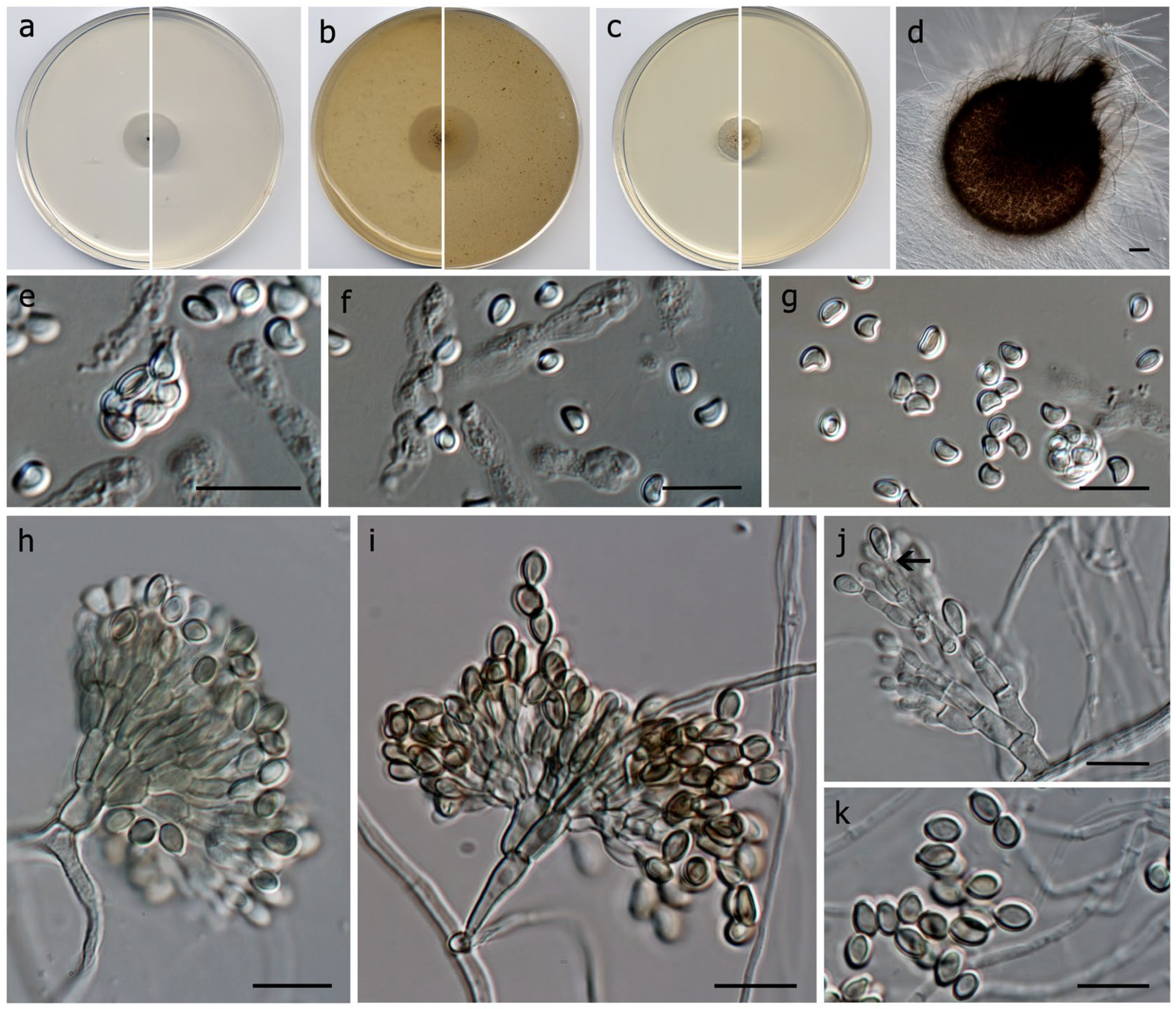
3. Results
Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rampelotto, P.H. Extremophiles and Extreme Environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef] [PubMed]
- D’Elia, L.; del Mondo, A.; Santoro, M.; de Natale, A.; Pinto, G.; Pollio, A. Microorganisms from Harsh and Extreme Environments: A Collection of Living Strains at ACUF (Naples, Italy). Ecol. Quest. 2018, 29, 63–74. [Google Scholar] [CrossRef]
- Santra, H.K.; Banerjee, D. Extremophilic Microbes and their Extremozymes for Industry and Allied Sectors. In Microbiomes of Extreme Environments, 1st ed.; Yadav, A.N., Rastegari, A.A., Yadav, N., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2021; Volume 1, pp. 1–109. [Google Scholar]
- Blachowicz, A.; Chiang, A.J.; Elsaesser, A.; Kalkum, M.; Ehrenfreund, P.; Stajich, J.E.; Torok, T.; Wang, C.C.C.; Venkateswaran, K. Proteomic and Metabolomic Characteristics of Extremophilic Fungi under Simulated Mars Conditions. Front. Microbiol. 2019, 10, 1013. [Google Scholar] [CrossRef] [PubMed]
- Yapiyev, V.; Sagintayev, Z.; Inglezakis, V.J.; Samarkhanov, K.; Verhoef, A. Essentials of Endorheic Basins and Lakes: A Review in the Context of Current and Future Water Resource Management and Mitigation Activities in Central Asia. Water 2017, 9, 798. [Google Scholar] [CrossRef]
- van den Broeck, M.; Waterkeyn, A.; Rhazi, L.; Grillas, P.; Brendonck, L. Assessing the Ecological Integrity of Endorheic Wetlands, with Focus on Mediterranean Temporary Ponds. Ecol. Indic. 2015, 54, 1–11. [Google Scholar] [CrossRef]
- Nicolet, P.; Biggs, J.; Fox, G.; Hodson, M.J.; Reynolds, C.; Whitfield, M.; Williams, P. The Wetland Plant and Macroinvertebrate Assemblages of Temporary Ponds in England and Wales. Biol. Conserv. 2004, 120, 261–278. [Google Scholar] [CrossRef]
- Blanché, C.; Molero, J. Las cubetas arreicas al sur de Bujaraloz (Valle del Ebro). Contribución a su estudio fitocenológico. Lazaroa 1986, 9, 277–299. [Google Scholar]
- Castañeda, C.; Herrero, J.; Conesa, J.A. Distribution, morphology and habitats of saline wetlands: A case study from Monegros, Spain. Geol. Acta 2013, 11, 371–388. [Google Scholar] [CrossRef]
- Castañeda, C.; García-Vera, M.Á. Water Balance in the Playa-Lakes of an Arid Environment, Monegros, NE Spain. Hydrogeol. J. 2008, 16, 87–102. [Google Scholar] [CrossRef]
- Ramsar Convention Secretariat. Designating Ramsar Sites: Strategic Framework and guidelines for the future development of the List of Wetlands of International Importance. In Ramsar Handbooks for the Wise Use of Wetlands, 4th ed.; Ramsar Convention Secretariat: Gland, Switzerland, 2010; pp. 17–116. [Google Scholar]
- Ribera, I.; Blasco-Zumeta, J. Biogeographical links between steppe insects in the Monegros region (Aragón, NE Spain), the eastern Mediterranean, and central Asia. J. Biogeogr. 1998, 25, 969–986. [Google Scholar] [CrossRef]
- Vives i Durán, J.; Vives i Noguera, E. Coleópteros halófilos de Los Monegros. Bol. Asoc. Esp. Entom. 1978, 2, 205–214. [Google Scholar]
- Casamayor, E.O.; Triadó-Margarit, X.; Castañeda, C. Microbial Biodiversity in Saline Shallow Lakes of the Monegros Desert, Spain. FEMS Microbiol. Ecol. 2013, 85, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Azpiazu-Muniozguren, M.; Perez, A.; Rementeria, A.; Martinez-Malaxetxebarria, I.; Alonso, R.; Laorden, L.; Gamboa, J.; Bikandi, J.; Garaizar, J.; Martinez-Ballesteros, I. Fungal Diversity and Composition of the Continental Solar Saltern in Añana Salt Valley (Spain). J. Fungi 2021, 7, 1074. [Google Scholar] [CrossRef] [PubMed]
- Perl, T.; Kis-Papo, T.; Nevo, E. Fungal Biodiversity in the Hypersaline Dead Sea: Extinction and Evolution. Biol. J. Linn. Soc. 2017, 121, 122–132. [Google Scholar] [CrossRef]
- Gostinčar, C.; Gunde-Cimerman, N. Understanding Fungi in Glacial and Hypersaline Environments. Annu. Rev. Microbiol. 2023, 77, 89–109. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, M.L.; Bilanenko, E.N.; Ponizovskaya, V.B.; Kokaeva, L.Y.; Georgiev, A.A.; Efimenko, T.A.; Markelova, N.N.; Kuvarina, A.E.; Sadykova, V.S. Haloalkalitolerant Fungi from Sediments of the Big Tambukan Saline Lake (Northern Caucasus): Diversity and Antimicrobial Potential. Microorganisms 2023, 11, 2587. [Google Scholar] [CrossRef] [PubMed]
- Malloch, D. New Concepts in the Microascaceae Illustrated by Two New Species. Mycologia 1970, 62, 727–740. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Guarro, J.; Cano-Lira, J.F.; Sutton, D.A.; Wiederhold, N.P.; de Hoog, G.S.; Abbott, S.P.; Decock, C.; Sigler, L.; Gené, J. Phylogeny and Taxonomic Revision of Microascaceae with Emphasis on Synnematous Fungi. Stud. Mycol. 2016, 83, 193–233. [Google Scholar] [CrossRef]
- Lackner, M.; de Hoog, G.S.; Yang, L.; Ferreira Moreno, L.; Ahmed, S.A.; Andreas, F.; Kaltseis, J.; Nagl, M.; Lass-Flörl, C.; Risslegger, B.; et al. Proposed Nomenclature for Pseudallescheria, Scedosporium and Related Genera. Fungal Divers. 2014, 67, 1–10. [Google Scholar] [CrossRef]
- Wang, M.M.; Yang, S.Y.; Li, Q.; Zheng, Y.; Ma, H.H.; Tu, Y.H.; Wei, L.; Cai, L. Microascaceae from the marine environment, with descriptions of six new species. J. Fungi 2024, 10, 45. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.; Saxena, R.; Erdoğdu, M.; Selcuk, F.; Rajeshkumar, K.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Gené, J.; Sutton, D.A.; Cano-Lira, J.F.; de Hoog, G.S.; Decock, C.A.; Wiederhold, N.P.; Guarro, J. Redefining Microascus, Scopulariopsis and Allied Genera. Persoonia 2016, 36, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Han, P.J.; Bai, F.Y.; Luo, A.; Bensch, K.; Meijer, M.; Kraak, B.; Han, D.Y.; Sun, B.D.; Crous, P.W.; et al. Taxonomy, Phylogeny and Identification of Chaetomiaceae with Emphasis on Thermophilic Species. Stud. Mycol. 2022, 101, 121–243. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Garcia, A.; Pellon, A.; Rementeria, A.; Buldain, I.; Barreto-Bergter, E.; Rollin-Pinheiro, R.; De Meirelles, J.V.; Xisto, M.I.D.S.; Ranque, S.; Havlicek, V.; et al. Scedosporium and Lomentospora: An Updated Overview of Underrated Opportunists. Med. Mycol. 2018, 56, 102–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, R.Z.; Jiang, H.L.; Schildgen, O. Scedosporium spp. Lung Infection in Immunocompetent Patients: A Systematic Review and MOOSE-Compliant Meta-Analysis. Medicine 2019, 98, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.A.; Halliday, C.L.; Hoenigl, M.; Cornely, O.A.; Meyer, W. Scedosporium and Lomentospora Infections: Contemporary Microbiological Tools for the Diagnosis of Invasive Disease. J. Fungi 2021, 7, 23. [Google Scholar] [CrossRef]
- Su, L.; Zhu, H.; Niu, Y.; Guo, Y.; Du, X.; Guo, J.; Zhang, L.; Qin, C. Phylogeny and Taxonomic Revision of Kernia and Acaulium. Sci. Rep. 2020, 10, 10302. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Zhou, S.Y.; Eurwilaichitr, L.; Ingsriswang, S.; Raza, M.; Chen, Q.; Zhao, P.; Liu, F.; Cai, L. Culturable Mycobiota from Karst Caves in China II, with Descriptions of 33 New Species. Fungal Divers. 2021, 106, 29–136. [Google Scholar] [CrossRef]
- Crous, P.W.; Boers, J.; Holdom, D.; Osieck, E.R.; Steinrucken, T.V.; Tan, Y.P.; Vitelli, J.S.; Shivas, R.G.; Barrett, M.; Boxshall, A.G.; et al. Fungal Planet Description Sheets: 1383–1435. Persoonia 2022, 48, 261–371. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; Dima, B.; Dovana, F.; et al. Fungal Planet Description Sheets: 1112–1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef]
- Preedanon, S.; Suetrong, S.; Srihom, C.; Somrithipol, S.; Kobmoo, N.; Saengkaewsuk, S.; Srikitikulchai, P.; Klaysuban, A.; Nuankaew, S.; Chuaseeharonnachai, C.; et al. Eight Novel Cave Fungi in Thailand’s Satun Geopark. Fungal. Syst. Evol. 2023, 12, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, C. Las Saladas Del Sur de Monegros: Facies, Régimen Hídrico y Estado Actual. Ph.D. Thesis, Universidad de Zaragoza, Zaragoza, Spain, 2004. [Google Scholar]
- Hocking, A.D.; Pitt, J.I. Dichloran-Glycerol Medium for Enumeration of Xerophilic Fungi from Low-Moisture Foods. Appl. Environ. Microbiol. 1980, 39, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Börner, G.V.; Cha, R.S. Induction and Analysis of Synchronous Meiotic Yeast Cultures. Cold Spring Harb. Protoc. 2015, 10, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Kirk, P.M.; Sutton, B.C.; Pegler, D.N. Ainsworth & Bisby’s Dictionary of the Fungi, 8th ed.; CAB International: Oxon, UK, 1995; p. 616. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi, 2nd ed.; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010; pp. 1–475. [Google Scholar]
- Furuya, K.; Naito, A. An effective method for isolation of Boothiella tetraspora from soil. Trans. Mycol. Soc. 1979, 20, 309–311. [Google Scholar]
- Furuya, K.; Naito, A. Stimulation of ascospore germination by phenolic compounds in members of the Sordariaceae. Trans. Mycol. Soc. 1980, 21, 77–85. [Google Scholar]
- Ulfig, K.; Guarro, J.; Cano, J.; Genie, J.; Vidal, R.; Figueras, M.J. General Assessment of the Occurrence of Keratinolytic Fungi in River and Marine Beach Sediments of Catalonian Waters (Spain). Water Air Soil Poll. 1997, 94, 275–287. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978. [Google Scholar]
- Chupp, C. Further notes on double cover-glass mounts. Mycologia 1940, 32, 269–270. [Google Scholar] [CrossRef]
- Müller, F.M.; Werner, K.E.; Kasai, M.; Francesconi, A.; Chanock, S.J.; Walsh, T.J. Rapid extraction of genomic DNA from medically important yeasts and filamentous fungi by high-speed cell disruption. J. Clin. Microbiol. 1998, 36, 1625–1629. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Iturrieta-González, I.; García, D.; Gené, J. Novel Species of Cladosporium from Environmental Sources in Spain. MycoKeys 2021, 77, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR To Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Torres-Garcia, D.; García, D.; Cano-Lira, J.F.; Gené, J. Two Novel Genera, Neostemphylium and Scleromyces (Pleosporaceae) from Freshwater Sediments and Their Global Biogeography. J. Fungi 2022, 8, 868. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA7: Molecular evolutionary genetics analysis version 7.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeifferm, W.; Schwartz, T. The CIPRES science gateway: Enabling high-impact science for phylogenetics researchers with limited resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the Extreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; Association for Computing Machinery: New York, NY, USA, 2012; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Hespanhol, L.; Vallio, C.S.; Costa, L.M.; Saragiotto, B.T. Understanding and Interpreting Confidence and Credible Intervals around Effect Estimates. Braz. J. Phys. Ther. 2019, 23, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Bondarenko, S.A.; Debets, A.J.M.; Bilanenko, E.N. On the Diversity of Fungi from Soda Soils. Fungal Divers. 2016, 76, 27–74. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Jung, H.Y.; Lee, Y.S.; Voigt, K.; Lee, H.B. Phylogenetic Status of Two Undescribed Zygomycete Species from Korea: Actinomucor elegans and Mucor minutus. Mycobiology 2017, 45, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Moubasher, A.H.; Ismail, M.A.; Hussein, N.A.; Gouda, H.A. Enzyme Producing Capabilities of Some Extremophilic Fungal Strains Isolated from Different Habitats of Wadi El-Natrun, Egypt. Part 2: Cellulase, Xylanase and Pectinase. Eur. J. Biol. Res. 2016, 6, 103–111. [Google Scholar]
- Ismail, M.A.; Hussein; Abdel-Sater, M.A.; Sayed, R.M. Evaluation of physiological and biochemical characteristics of Alternaria species isolated from soil in Assiut Governorate, Egypt, in addition to dichotomous key to the encountered species. Assiut Univ. J. Bot. Microbiol. 2020, 49, 34–59. [Google Scholar]
- Kozakiewicz, Z.; Smith, D. Physiology of Aspergillus. In Biotechnology Handbooks, 7th ed.; Smith, J.E., Ed.; Springer: Boston, MA, USA, 1994; pp. 23–40. [Google Scholar]
- Butinar, L.; Zalar, P.; Frisvad, J.C.; Gunde-Cimerman, N. The Genus Eurotium—Members of Indigenous Fungal Community in Hypersaline Waters of Salterns. FEMS Microbiol. Ecol. 2005, 51, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.J.; Hubka, V.; Frisvad, J.C.; Visagie, C.M.; Houbraken, J.; Meijer, M.; Varga, J.; Demirel, R.; Jurjević, Ž.; Kubátová, A.; et al. Polyphasic Taxonomy of Aspergillus Section Aspergillus (Formerly Eurotium), and Its Occurrence in Indoor Environments and Food. Stud. Mycol. 2017, 88, 37–135. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Ding, X.W.; Narsing Rao, M.P.; Zhang, B.; Zhang, Y.G.; Liu, F.H.; Liu, B.B.; Xiao, M.; Li, W.J. Morphological and Transcriptomic Analysis Reveals the Osmo-adaptive Response of Endophytic Fungus Aspergillus montevidensis ZYD4 to High Salt Stress. Front. Microbiol. 2017, 8, 1789. [Google Scholar] [CrossRef]
- Sklenář, F.; Glässnerová, K.; Jurjević, Ž.; Houbraken, J.; Samson, R.A.; Visagie, C.M.; Yilmaz, N.; Gené, J.; Cano, J.; Chen, A.J.; et al. Taxonomy of Aspergillus Series Versicolores: Species Reduction and Lessons Learned about Intraspecific Variability. Stud. Mycol. 2022, 102, 53–93. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Shivas, R.G.; Quaedvlieg, W.; van der Bank, M.; Zhang, Y.; Summerell, B.A.; Guarro, J.; Wingfield, M.J.; Wood, A.R.; Alfenas, A.C.; et al. Fungal Planet Description Sheets: 214–280. Persoonia 2014, 32, 184–306. [Google Scholar] [CrossRef]
- Asgari, B.; Rasoul, Z. The Genus Chaetomium in Iran, a Phylogenetic Study Including Six New Species. Mycologia 2011, 103, 863–882. [Google Scholar] [CrossRef] [PubMed]
- Garmendia, G.; Alvarez, A.; Villarreal, R.; Martínez-Silveira, A.; Wisniewski, M.; Vero, S. Fungal Diversity in the Coastal Waters of King George Island (Maritime Antarctica). World J. Microbiol. Biotechnol. 2021, 37, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I. An Appraisal of Identification Methods for Penicillium Species: Novel Taxonomic Criteria Based on Temperature and Water Relations. Mycologia 1973, 65, 1135–1157. [Google Scholar] [CrossRef] [PubMed]
- Bretzloff, C.W. The Growth and Fruiting of Sordaria fimicola. Am. J. Bot. 1954, 41, 58–67. [Google Scholar] [CrossRef]
- Paiva, D.S.; Fernandes, L.; Pereira, E.; Trovão, J.; Mesquita, N.; Tiago, I.; Portugal, A. Exploring Differences in Culturable Fungal Diversity Using Standard Freezing Incubation—A Case Study in the Limestones of Lemos Pantheon (Portugal). J. Fungi 2023, 9, 501. [Google Scholar] [CrossRef] [PubMed]
- Kis-Papo, T.; Grishkan, I.; Oren, A.; Wasser, S.P.; Nevo, E. Spatiotemporal diversity of filamentous fungi in the hypersaline Dead Sea. Mycol. Res. 2001, 6, 749–756. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Zalar, P. Extremely Halotolerant and Halophilic Fungi Inhabit Brine in Solar Salterns Around the Globe. Food Technol. Biotechnol. 2014, 52, 170–179. [Google Scholar]
- Sayed, A.M.; Hassan, M.H.A.; Alhadrami, H.A.; Hassan, H.M.; Goodfellow, M.; Rateb, M.E. Extreme Environments: Microbiology Leading to Specialized Metabolites. J. Appl. Microbiol. 2020, 128, 630–657. [Google Scholar] [CrossRef]
- Martinelli, L.; Zalar, P.; Gunde-Cimerman, N.; Azua-Bustos, A.; Sterflinger, K.; Piñar, G. Aspergillus atacamensis and A. salisburgensis: Two New Halophilic Species from Hypersaline/Arid Habitats with a phialosimplex-like Morphology. Extremophiles 2017, 21, 755–773. [Google Scholar] [CrossRef]
- Gostinčar, C.; Zalar, P.; Gunde-Cimerman, N. No Need for Speed: Slow Development of Fungi in Extreme Environments. Fungal Biol. Rev. 2022, 39, 1–14. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.M.; Blanchette, R.A.; Held, B.W. New Record of Chaetomium grande Asgari & Zare (Chaetomiaceae) for the Egyptian and African Mycobiota. Phytotaxa 2018, 343, 283–288. [Google Scholar] [CrossRef]
- Mehrabi, M.; Asgari, B. Description of Allocanariomyces and Parachaetomium, Two New Genera, and Achaetomium aegilopis sp. nov. in the Chaetomiaceae. Mycol. Prog. 2020, 19, 1415–1427. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, W.; Cai, L. Polyphasic Characterisation of Chaetomium Species from Soil and Compost Revealed High Number of Undescribed Species. Fungal Biol. 2017, 121, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Silvera-Simón, C.; Gené, J.; Cano, J.; Guarro, J. Wardomycopsis litoralis, a New Soil-Borne Hyphomycete from Spain. Mycotaxon 2008, 105, 195–202. [Google Scholar]
- Sun, B.; Zhou, Y.; Chen, A.J. Two New Microascus Species with Spinous Conidia Isolated from Pig Farm Soils in China. Mycoscience 2020, 61, 190–196. [Google Scholar] [CrossRef]
- Tazik, Z.; Rahnama, K.; Iranshahi, M.; White, J.F.; Soltanloo, H. A New Species of Pithoascus and First Report of This Genus as Endophyte Associated with Ferula ovina. Mycoscience 2020, 61, 145–150. [Google Scholar] [CrossRef]
- Abrantes, R.A.; Refojo, N.; Hevia, A.I.; Fernández, J.; Isla, G.; Córdoba, S.; Dávalos, M.F.; Lubovich, S.; Maldonado, I.; Davel, G.O.; et al. Scedosporium spp. From Clinical Setting in Argentina, with the Proposal of the New Pathogenic Species Scedosporium americanum. J. Fungi 2021, 7, 160. [Google Scholar] [CrossRef]
- Calabon, M.S.; Jones, E.B.G.; Promputtha, I.; Hyde, K.D. Fungal Biodiversity in Salt Marsh Ecosystems. J. Fungi 2021, 7, 648. [Google Scholar] [CrossRef]
- Jalili, B.; Bagheri, H.; Azadi, S.; Soltani, J. Identification and Salt Tolerance Evaluation of Endophyte Fungi Isolates from Halophyte Plants. Int. J. Environ. Sci. Technol. 2020, 17, 3459–3466. [Google Scholar] [CrossRef]
- Dickinson, C.H. The Genus Wardomyces. Trans. Br. Mycol. Soc. 1964, 47, 321. [Google Scholar] [CrossRef]
- Lee, J.H.; Ten, L.N.; Lee, S.Y.; Jung, H.Y. Novel Fungal Species Belonging to the Genus Acaulium Isolated from Riptortus clavatus (Heteroptera: Alydidae) in Korea. Korean J. Mycol. 2021, 49, 477–486. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Meijer, M.; Houbraken, J.; Samson, R.A. Scopulariopsis and scopulariopsis-like Species from Indoor Environments. Stud. Mycol. 2017, 88, 1–35. [Google Scholar] [CrossRef]
- Morelet, M. Micromycètes du var et d'ailleurs (2me Note). Ann. Soc. Sci. Nat. Archèol. Toulon du Var 1969, 21, 104–106. [Google Scholar]



{kind=link}
{kind=link}
| Taxon | Strain Number | Source | Origin | Sequence Accession Number | |||
|---|---|---|---|---|---|---|---|
| LSU | ITS | EF-1α | tub2 | ||||
| Acaulium acremonium | CBS 290.38 | Skin of a horse | København, Denmark | LN851001 | LM652456 | HG380362 | LN851108 |
| Acaulium acremonium | MUCL 8274 ET | Wheat field soil | Schleswig-Holstein, Germany | LN851002 | LM652457 | LN851056 | LN851109 |
| Acaulium albonigrescens | IHEM 18560 ET | Litter treated with urea | Nemuro-shi, Japan | LN851004 | LM652389 | LN851058 | LN851111 |
| Acaulium caviariformis | CBS 536.87 ET | Decaying meat | Flemalle, Belgium | LN851005 | LM652392 | LN851059 | LN851112 |
| Cephalotrichum asperulus | CBS 127.22 | Seed | Wageningen, The Netherlands | LN851006 | LN850959 | LN851060 | LN851113 |
| Cephalotrichum asperulus | CBS 582.71 IT | Soil | Buenos Aires, Argentina | LN851007 | LN850960 | LN851061 | LN851114 |
| Cephalotrichum brevistipitatum | CBS 157.57 T | Tuber | Wageningen, The Netherlands | LN851031 | LN850984 | LN851084 | LN851138 |
| Cephalotrichum columnare | CBS 159.66 T | Dung of hare | Johannesburg, South Africa | LN851010 | LN850963 | LN851064 | LN851117 |
| Cephalotrichum cylindricum | CBS 448.51 | Timber | Bekker, South Africa | LN851011 | LN850964 | LN851065 | LN851118 |
| Cephalotrichum cylindricum | UAMH 1348 ET | Seed of sorghum | KS, USA | LN851012 | LN850965 | LN851066 | LN851119 |
| Cephalotrichum dendrocephalum | CBS 528.85 IT | Cultivated soil | Basrah, Iraq | LN851013 | LN850966 | LN851067 | LN851120 |
| Cephalotrichum gorgonifer | CBS 635.78 ET | Hair | The Netherlands | LN851024 | LN850977 | LN851077 | LN851131 |
| Cephalotrichum gorgonifer | UAMH 3585 | Mushroom compost | Spruce Grove, AB, Canada | LN851025 | LN850978 | LN851078 | LN851132 |
| Cephalotrichum hinnuleum | CBS 289.66 T | Dung of deer | Tasmania, Australia | LN851032 | LN850985 | LN851085 | LN851139 |
| Cephalotrichum microsporum | CBS 523.63 ET | Wheat field soil | Schleswig-Holstein, Germany | LN851014 | LN850967 | LN851068 | LN851121 |
| Cephalotrichum microsporum | UAMH 9365 | Indoor air | Peace River, AB, Canada | LN851015 | LN850968 | LN851069 | LN851122 |
| Cephalotrichum nanum | CBS 191.61 ET | Dung of deer | Richmond Park, SRY, ENG, UK | LN851016 | LN850969 | LN851070 | LN851123 |
| Cephalotrichum nanum | UAMH 9126 | Dung of bison | Elk Island National Park, AB, Canada | LN851017 | LN850970 | LN851071 | LN851124 |
| Cephalotrichum purpureofuscum | UAMH 9209 | Indoor air | Pemberton, BC, Canada | LN851018 | LN850971 | LN851072 | LN851125 |
| Cephalotrichum stemonitis | CBS 103.19 NT | Seed | Wageningen, The Netherlands | LN850952 | LN850951 | LN850953 | LN850954 |
| Cephalotrichum stemonitis | CBS 180.35 | Unknown | Unknown | LN851019 | LN850972 | LN851073 | LN851126 |
| Cephalotrichum verrucisporum | CBS 187.78 | Dune soil | Katijk, The Netherlands | LN851033 | LN850986 | LN851086 | LN851140 |
| Dactyliodendromyces holomorphus | CBS 149968 | Lagoon sediment | Zaragoza, Spain | OR141719 | OR141718 | OR142400 | OR142401 |
| Fairmania singularis | CBS 414.64 | Laboratory contaminant | Tokyo, Japan | LN851035 | LM652442 | LN851088 | LN851142 |
| Fairmania singularis | CBS 505.66 ET | Barrel bottom | Kittery Point, ME, USA | LN851036 | LN850988 | LN851089 | LN851143 |
| Gamsia aggregata | CBS 251.69 IT | Dung of carnivore | Wycamp Lake, MI, USA | LN851037 | LM652378 | LN851090 | LN851144 |
| Gamsia columbina | CBS 546.69 T | Milled Oryza sativa | Osaka, Japan | LN851041 | LM652379 | LN851094 | LN851148 |
| Gamsia columbina | CBS 233.66 ET | Sandy soil | Giessen, Germany | LN851039 | LN850990 | LN851092 | LN851146 |
| Parawardomyces ovalis | CBS 234.66 T | Wheat field soil | Schleswig-Holstein, Germany | LN851050 | LN850996 | LN851101 | LN851155 |
| Parawardomyces giganteus | CBS 746.69 T | Insect frass in a dead log | Coldwater, ON, Canada | LN851045 | LM652411 | LN851096 | LN851150 |
| Pseudowardomyces pulvinatus | CBS 112.65 T | Salt marsh | CHS, ENG, UK | LN851051 | LN850997 | LN851102 | LN851156 |
| Pseudowardomyces humicola | CBS 369.62 IT | Soil in tropical greenhouse | Guelph, ON, Canada | LN851046 | LN850993 | LN851097 | LN851151 |
| Scopulariopsis brevicaulis | MUCL 40726 T | Indoor air | Scandia, AB, Canada | LN851042 | LM652465 | HG380363 | LM652672 |
| Microascus longirostris | CBS 196.61 NT | Wasp’s nest | Kittery Point, ME, USA | LN851043 | LM652421 | LM652566 | LM652634 |
| Wardomyces anomalus | CBS 299.61 ET | Air cell of egg | Ottawa, ON, Canada | LN851044 | LN850992 | LN851095 | LN851149 |
| Wardomyces inflatus | CBS 216.61 IT | Wood, Acer sp. | Sainte-Cécile-de-Masham, QC, Canada | LN851047 | LM652496 | LN851098 | LN851152 |
| Wardomyces inflatus | CBS 367.62 NT | Greenhouse soil | Heverlee, Belgium | LN851048 | LN850994 | LN851099 | LN851153 |
| Wardomycopsis humicola | CBS 487.66 IT | Soil | Guelph, ON, Canada | LM652554 | LM652497 | LN851103 | LN851157 |
| Wardomycopsis humicola | FMR 3993 | Sediment of Ter River | Girona, Spain | LN851052 | LN850998 | LN851104 | LN851158 |
| Wardomycopsis humicola | FMR 13592 | Soil | Reus, Spain | LN851053 | LN850999 | LN851105 | LN851159 |
| Wardomycopsis inopinata | FMR 10305 | Soil | Bagan, Myanmar | LN851054 | LM652498 | LN851106 | LN851160 |
| Wardomycopsis inopinata | FMR 10306 | Soil | Bagan, Myanmar | LN850956 | LN850955 | LN850957 | LN850958 |
| Wardomycopsis litoralis | CBS 119740 T | Beach soil | Castellon, Spain | LN851055 | LN851000 | LN851107 | LN851161 |
| Taxon | Strain Nr 1 | Identity Percentage (%) | GenBank Accession Nr 2 | Markers Used | Source | Extremophilic Features Reported | References |
|---|---|---|---|---|---|---|---|
| Acrostalagmus luteoalbus | 19813 * | 99.69 | KP050692 | ITS | water | Alkali-tolerant | [60] |
| Actinomucor elegans | 19823 * | 99.79 | AY243954 | ITS | water | Thermotolerance (strain-dependent) | [61] |
| Alternaria alternata | 20034 * | 100 | KP124364 | ITS | sediment | Halotolerant; alkali-tolerant | [62] |
| Alternaria chlamydospora | 20037 * | 100 | MG020753 | ITS | sediment | Acidophilic; alkali-tolerant; psychrotolerant; xerotolerant | [63] |
| Aspergillus amstelodami | 20038 * | 99.22 | MT820427 | tub2 | water | Xerophilic; thermotolerant | [64,65] |
| Aspergillus calidoustus | 19423 *, 19820 | 99.7 | LT798990 | tub2 | water | Thermotolerant | [64] |
| Aspergillus intermedius | 19821 * | 100 | LT671082 | tub2 | water | Osmophilic; thermotolerant; xerophilic | [66] |
| Aspergillus montevidensis | 20492 * | 100 | KF499570 | tub2 | sediment | Halotolerant | [67] |
| Aspergillus versicolor | 19427 *, 20659 | 100 | ON807694 | tub2 | sediment | Halotolerant | [68] |
| Cephalotrichiella penicillata | 20498 * | 100 | NR_153893 | ITS | sediment | Not reported | [69] |
| Chaetomium grande | 20036 * | 99.30 | KT214731 | tub2 | sediment | Thermotolerant | [70] |
| Cladosporium europaeum | 19425 *, 20499 | 100 | HM148294 | EF-1α | water | Halotolerant | Our study |
| Dactyliodendromyces holomorphus | 20493 * | On text | ITS, LSU, EF-1α, tub2 | sediment | Not reported | Our study | |
| Epicoccum italicum | 20044 * | 100 | MN983956 | tub2 | sediment | Psychrotolerant; halotolerant | [71] |
| Fusarium culmorum | 20248 * | 99.85 | KT008433 | EF-1α | sediment | Not reported | |
| Malbranchea zuffiana | 20033 * | 98.90 | MH869293 | ITS | sediment | Halotolerant | Our study |
| Ovatospora amygdalispora | 20322 * | 99.41 | MZ343030 | tub2 | sediment | Not reported | |
| Parachaetomium truncatulum | 20041 *, 20495 | 99.77 | HM365298 | tub2 | sediment | Thermotolerant | [70] |
| Penicillium egyptiacum | 20328 *, 20324, 20323, 20331, 20337 | 100 | JX996851 | tub2 | sediment | Psychrotolerant; non-thermotolerant; xerotolerant | [72] |
| Sordaria fimicola | 19587 * | 99.61 | MH860820 | ITS | sediment | Striking stimulation of ascospore germination by acetate | [73] |
| Stachybotrys chartarum | 19808 * | 100 | KU846678 | ITS | sediment | Halotolerant | [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnés-Guirado, M.; Stchigel, A.M.; Cano-Lira, J.F. A New Genus of the Microascaceae (Ascomycota) Family from a Hypersaline Lagoon in Spain and the Delimitation of the Genus Wardomyces. J. Fungi 2024, 10, 236. https://doi.org/10.3390/jof10040236
Barnés-Guirado M, Stchigel AM, Cano-Lira JF. A New Genus of the Microascaceae (Ascomycota) Family from a Hypersaline Lagoon in Spain and the Delimitation of the Genus Wardomyces. Journal of Fungi. 2024; 10(4):236. https://doi.org/10.3390/jof10040236
Chicago/Turabian StyleBarnés-Guirado, María, Alberto Miguel Stchigel, and José Francisco Cano-Lira. 2024. "A New Genus of the Microascaceae (Ascomycota) Family from a Hypersaline Lagoon in Spain and the Delimitation of the Genus Wardomyces" Journal of Fungi 10, no. 4: 236. https://doi.org/10.3390/jof10040236