
An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae)

1
Groningen Institute for Evolutionary Life Sciences (GELIFES), University of Groningen, Nijenborgh 7, 9747 AG Groningen, The Netherlands
2
Kebon Jeruk Baru C 3–12, Kebon Jeruk, Jakarta Barat 11530, Indonesia
3
Marine Biodiversity Group, Naturalis Biodiversity Center, Darwinweg 2, 2333 CR Leiden, The Netherlands
*
Author to whom correspondence should be addressed.
Arthropoda 2024, 2(1), 66-75; https://doi.org/10.3390/arthropoda2010005
Submission received: 22 December 2023
/
Revised: 21 January 2024
/
Accepted: 2 February 2024
/
Published: 18 February 2024
Abstract
:Coral-dwelling gall crabs (Cryptochiridae) are common inhabitants of scleractinian corals. Several species have been described as new in recent years, including Lithoscaptus doughnut, which was described from Hong Kong based on a single female retrieved from the coral Plesiastrea peroni. Here we extend the distribution range of L. doughnut with nine additional localities throughout the Indo-West Pacific, from the Red Sea to the Coral Triangle and Japan. We describe a male specimen of L. doughnut for the first time, based on a specimen from Malaysia, and provide photographs of life and preserved material. Haplotype networks based on COI mtDNA (n = 12) and 16 rRNA sequences (n = 12) were created. We retrieved eleven COI haplotypes and six 16S haplotypes, however no clear geographic distribution pattern was discerned. Intraspecific variation in L. doughnut was 1.4% for COI and 0.2% for 16S. Lastly, the first colour photos and records of associated parasites of this species are provided.
1. Introduction
Tropical coral reefs are among the most biodiverse ecosystems on the planet. A large part of the biodiversity on coral reefs is composed of invertebrates that live in close association with sessile organisms. Reef-building stony corals (Scleractinia) provide habitat to a wide range of species, and their associated fauna has been reviewed in several papers [1,2,3]. Although the exact nature of these associations is not always well understood, with species exhibiting dependencies ranging from facultative to obligate symbiosis, this small majority is an important component of the biodiversity on coral reefs.
Decapod crustaceans form a major part of the coral-associated fauna [3]. The small, coral-dwelling gall crabs of the family Cryptochiridae Paulson, 1875 are obligately symbiotic with scleractinian corals and have been the subject of various recent studies on, for example, their reproductive morphology, occurrence patterns, biogeography, and even fluorescence [4,5,6,7]. Their association with host corals is strong and not affected by intraspecific coral competition [8]. Gall crabs are found on coral reefs worldwide. They are most abundant on shallow-water tropical reefs, but are known to also occur in the deep sea at >500 m depth [9]. There are currently 54 described species across 21 genera [10], however, recent studies highlighted the presence of large (cryptic) diversity in the genera Hapalocarcinus Stimpson, 1859 and Opecarcinus Kropp & Manning, 1987 [7,11]. Cryptochirids show varying degrees of host specificity; most species inhabit a single coral genus or several closely related species [12,13,14]. The Atlantic species Troglocarcinus corallicola Verrill, 1908 is the only host generalist in the family [9,15]. Besides this variation in host-specificity, cryptochirids differ in their geographic distribution. Some species are very widespread, whereas others are only known from a smaller geographic area [7,15]. The Coral Triangle is the biodiversity hotspot for scleractinian corals [16,17], with the Red Sea as a secondary centre of biodiversity [17]. Geographic range shifts are thought to play a critical role in generating the observed species diversity gradients on coral reefs [16], however for many invertebrates, including gall crabs, these distribution ranges need to be studied in more detail [7,18].
Gall crabs of the genus Lithoscaptus A. Milne-Edwards, 1862 inhabit corals of the family Merulinidae H. Milne Edwards & Haime, 1857 [13]. The genus is composite in nature and in need of revision [19,20]. It currently comprises 13 described species; however, more diversity is expected, based on preliminary data [20]. The recently described species L. semperi Van der Meij, 2015, L. tuerkayi Van der Meij, 2017, and L. aquarius Van der Meij, 2023 strictly associate with Trachyphyllia geoffroyi (Audouin, 1826), Caulastrea spp. Dana, 1846, and Catalaphyllia jardinei (Saville-Kent, 1893), respectively, whereas other species have been recorded from multiple host genera [13,21]. Like many other gall crab genera, Lithoscaptus has been recorded throughout most of the Indo-West Pacific (IWP), albeit with patchily known distribution records [13].
Recently, the species Lithoscaptus doughnut Wong, Tsao, Qui & Chan, 2023 was described based on a single female specimen collected from Plesiastrea peroni H. Milne Ewards & Haime, 1857 near Basalt Island (Hong Kong), limiting the species’ known distribution to the type locality. Here, we extend the distribution range of L. doughnut throughout the IWP, based on material collected from the Red Sea to the Coral Triangle and Japan. We illustrate and describe the male of L. doughnut, based on a specimen from Malaysia, provide haplotype networks based on sequence data of the cytochrome c oxidase subunit I gene (COI) and 16S mtDNA and the first colour photographs of the species. Lastly, we comment on the identity of L. cf. doughnut based on 16S data.
2. Material and Methods
2.1. Sampling
Gall crabs were sampled from Plesiastrea peroni during fieldwork between 2007 and 2016 in the Red Sea, Maldives, Malaysia, Indonesia, and Japan (Table 1, Figure 1), and provisionally identified as a new species by the senior author under the placeholder name Lithoscaptus “Plesi”. One sample was collected from a free-living colony (corallith) of Cyphastrea chalcidicum (Forskål, 1775). Most samples were collected from deeper parts of the reef, between 8 and 24 m depth. Based on morphology, COI barcoding, and host data, these samples are now identified as Lithoscaptus doughnut. All specimens are deposited in the scientific collections of Naturalis Biodiversity Center in Leiden (The Netherlands), with the collections coded as RMNH.CRUS.D. All corals were photographed in situ in the field, whereas the crabs were photographed in field laboratories using a digital SLR camera equipped with a macro lens. Abbreviations used: CL: carapace length; P: pereiopod; MXP-3: maxilliped 3; G1: gonopod 1; G2: gonopod 2.
2.2. Haplotype Networks
Sequences of COI mtDNA (COI [22]) and 16S rRNA (16L2 and 16H10 [23]) were obtained following the protocol in Van der Meij [14]. Sequences were assembled in Sequencher 4.10.1 [24] and aligned using ClustalW Multiple Alignment in BioEdit v7.0.5.3 [25], together with the sequences from Wong et al. [26]. Sequences were trimmed to have the same maximum length, 620 bp for COI and 585 bp for 16S. Median-joining haplotype networks were constructed with PopArt 1–7 [27] for both sets of sequences. All samples were assigned to their sampling localities; both networks span 9 locations throughout the IWP (Table 1).
2.3. Intraspecific Divergence
MEGA11 [28] was used to determine the intraspecific divergence of L. doughnut. The best-fitting model of nucleotide substitution was calculated using MEGA11, resulting in the Tamura 3-parameter model for both datasets [29], which was subsequently used to calculate the within-group intraspecific divergence.
3. Results and Discussion
3.1. Distribution and Haplotype Networks
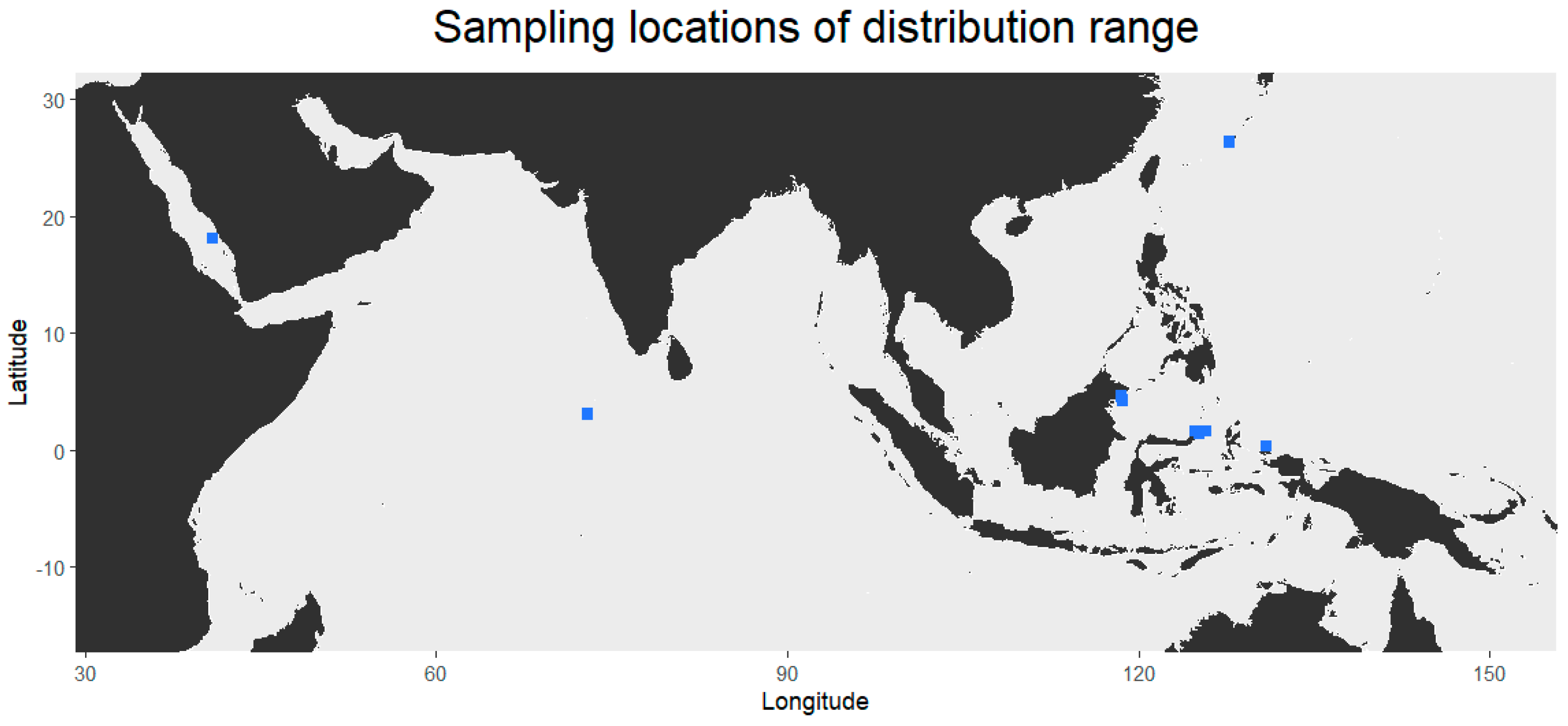
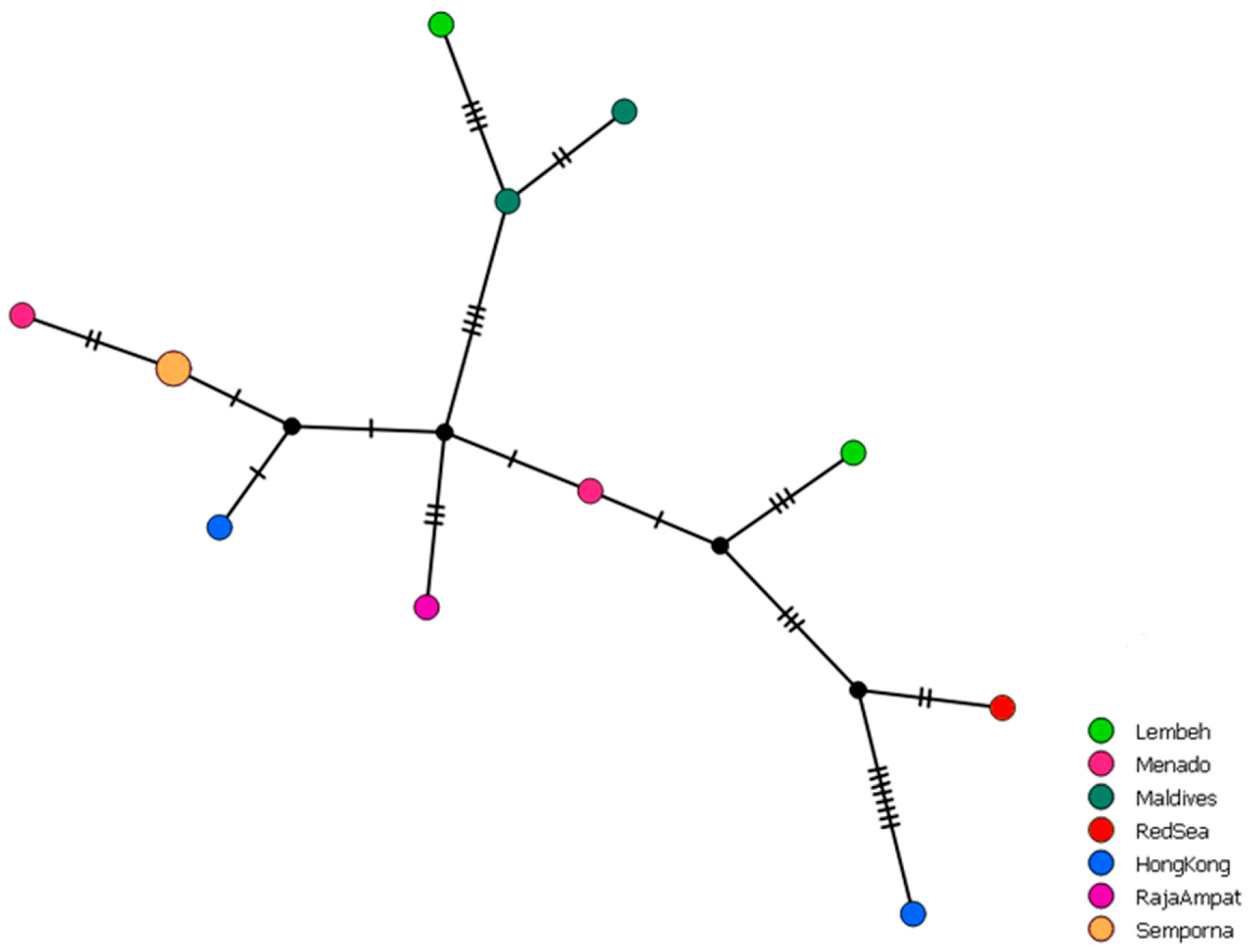
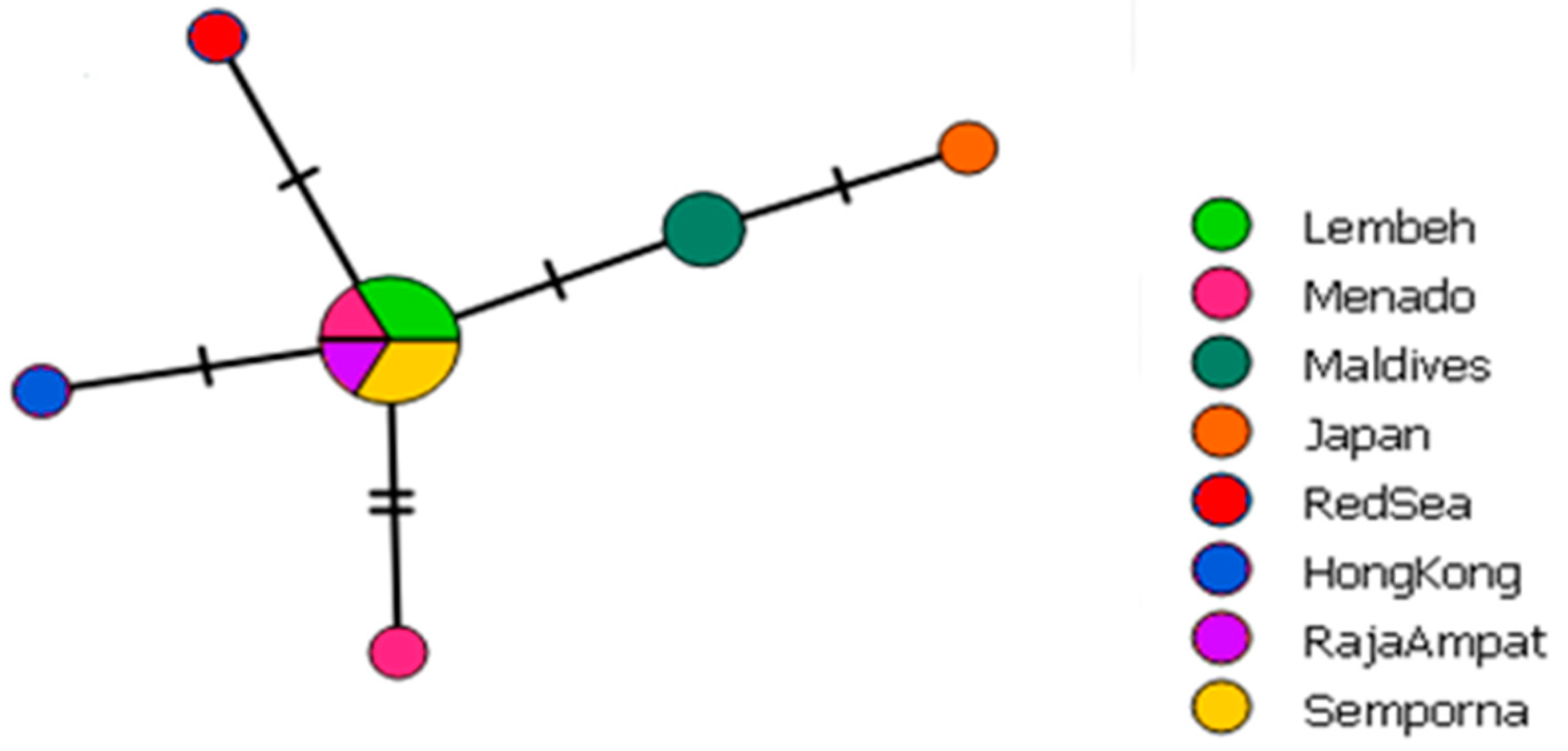
Lithoscaptus doughnut is now recorded from nine localities ranging from the Red Sea to the Coral Triangle and Japan, extending the distribution range of the species from Hong Kong to all throughout the IWP (Figure 1). The distribution is roughly in line with the distribution of its host P. peroni [30]. The Pacific Ocean, where P. peroni also occurs, has not yet been sampled for L. doughnut. The haplotype network based on COI sequences data shows eleven different haplotypes; however, no clear geographical pattern was observed (Figure 2). The 16S haplotype network yields six haplotypes, providing less diversity in comparison to the COI haplotype network (Figure 3). Here, all sequences from the Coral Triangle are included in the main haplotype or differ one or two base pairs from this most common haplotype.
The difference in the number of haplotypes between the COI and 16S networks is supported by the different evolutionary divergence estimates. The intraspecific difference within the sequences is 1.4% for COI and 0.2% for 16S.
3.2. Coral Host
Plesiastrea peroni was identified as the host of L. doughnut by Wong et al. [26]. This is in agreement with our samples, which were retrieved from the same coral host. Earlier studies recorded Plesiastrea versipora (Lamarck, 1816) as the host of this species [31,32]; however, Plesiastrea corals in (sub)tropical waters should be referred to as P. peroni instead [30]. Plesiastrea versipora is restricted to temperate waters, and, so far, no gall crabs have been recorded from this species. One of our samples (from Semporna, Malaysia) was collected from a Cyphastrea chalcidicum corallith. It is not uncommon for host-specific species to occasionally end up in the ‘wrong’ host [33], perhaps caused by the crab needing to settle after reaching the megalopa stage.
Plesiastrea peroni belongs to the Plesiastreidae Dai & Horng, 2009, a family with just two species. Plesiastreidae cluster basal to Montastraeidae Yabe & Sugiyama, 1941 and Diploastraeidae Chevalier & Beauvais, 1987, as well as the species-rich coral families Lobophylliidae Dai & Horn, 2009 and Merulinidae [34]. All other Lithoscaptus species are associated with Merulinidae corals; hence, L. doughnut is currently the only Lithoscaptus species associated with a non-Merulinidae coral. Cyphastrea, the host genus of one of our specimens, does belong to the Merulinidae.
3.3. Phylogenetic Position
Wong et al. [26] retrieved L. doughnut as a sister to L. paradoxus A. Milne-Edwards, 1862 and L. scottae Wong, Tsao, Qiu & Chan, 2022 based on COI phylogeny, albeit without support. Lithoscaptus doughnut was already included in the phylogenetic reconstruction of Van der Meij and Klaus [35] under the placeholder name Lithoscaptus “Plesi”. Based on their three-marker phylogeny, the species was retrieved in a fully supported clade containing Lithoscaptus paradoxus, Lithoscaptus semperi, Lithoscaptus prionotus Kropp, 1994, and Xynomaia sheni (Fize & Serène, 1956). Lithoscaptus aquarius Van der Meij, 2023, associated with Catalaphyllia jardinei, also belongs to this clade [36]. Here, we confidently place L. doughnut as a sister to the above-mentioned taxa.
3.4. Lithoscaptus cf. doughnut
A second specimen from P. peroni obtained by Wong et al. [26] was identified as Lithoscaptus cf. doughnut. The COI sequence obtained from this specimen was shorter in length (567 bp) and had a Kimura 2-parameter (K2P) distance of 2.73%. The authors chose to stay conservative and not fully assign this specimen to L. doughnut, also because this second specimen was damaged and a few morphological differences were observed. The intraspecific divergence falls within the range of our COI sequence data. Based on 16S data, Lithoscaptus cf. doughnut differs by just one base pair from the most common 16S haplotype found in the Coral Triangle (Figure 3). Based on the COI and 16S data presented here, there appears to be enough support to simply regard L. cf. doughnut as L. doughnut. The observed morphological differences could potentially be regarded as intraspecific diversity.
3.5. Colour in Life
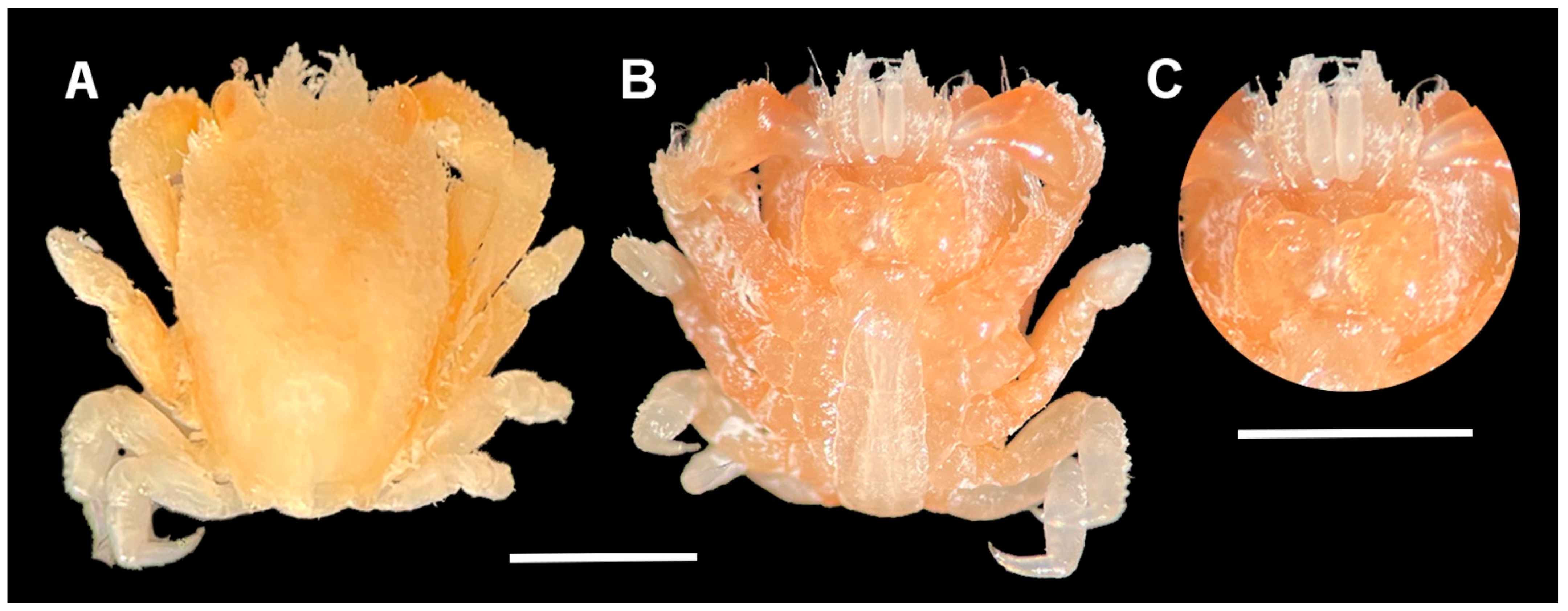
The colour of L. doughnut is overall beige. The deflected anterior one-third of the dorsal surface is a dark brown, whereas the posterior two-thirds are light beige. Granules are a lighter shade (Figure 4A). Chelipeds are dark beige with brown chelae (Figure 4A,B), and another specimen shows clear white markings on the chelae (Figure 4C,D). Pereiopod two is beige with reddish-brown propodus and dactylus, whereas pereiopods three to five are translucent with few off-white markings (Figure 4A–D). The telson from a specimen from the Red Sea is largely translucent (Figure 4C,D), whereas the first and second segments of a specimen from Indonesia have dark brown speckles (Figure 4A,B). The third maxilliped is dark brown, with translucent antennule with white bands, and reddish-brown cornea.
3.6. Description of Male L. doughnut
Material examined: ♂ (1.6 mm × 1.3 mm, RMNH.CRUS.D.53725, Genbank; COI: OR710951; 16S: OR711064), collected from Cyphastraea chalcidium (Figure 5).
Description: Carapace 1.2 times as long as broad, mildly convex, less so than female. Anterior half squaroid, posterior half trapezoid with the shorter base at the posterior end. Anterior half of carapace strongly deflected, with two depressions antero-laterally of mesogastric region. Mesogastric region inflated; lacks distinct longitudinal grooves observed in females. Carapace mildly granulated, well isolated from each other, distribution of granules sparse at posterior half giving a smooth appearance. Antero-lateral edge with spines at outer corners, small spines at inner orbital angle. Lateral edge finely serrated anteriorly, smooth towards posterior end. External orbital angles similar height to internal orbital angle. Orbit broadly V-shaped. Female specimen from the same lot (RMNH.CRUS.D.53725) has an inner orbital angle protruding further than described of the holotype of L. doughnut (see description [26]: Figure 10a). Eye-stalks stout, 1.2 times as long as broad, and more straight than eye-stalks of female. Cornea anterolateral. Pterygostomial region similar to that of female, but lacking granules.
Cheliped stout, merus strongly compressed, half as long as carpus, propodus and merus strongly granulated dorsally, dactylus as long as palms, straight dorsal margin but rounded and open on palmar side, distal tips of fingers touching. Tip of left movable finger is damaged. Several rows of rounded tubercles on dorsal margin of propodus. Carpus armed with small spines, more so than female. Pereiopods decreasing in size: P2 1.3 times longer than P5. Propodus of P2–4 serrated along dorsal margin, dorsal margin of P5 smooth. Proportions of segments similar to female’s. P2-left missing.
Antennulae smooth and tubular. Distal projection of antennular peduncles triangular, strongly toothed and extending beyond eye-stalks, congruent with antennular peduncles of female. Epistome does not show longitudinal crests, unlike female. MXP-3 squaroid with rounded margins, mesial margin of ischium lacking clear granules. Merus with distal external angle. Carpus and propodus grape-like in shape. Propodus with small tuft of setae. Exopod identical to female. Clearly accentuated pleural ridge. Abdomen elongated and straight, approximately three times longer than broad. Abdomen only covers middle of thoracic region, whereas female’s abdomen forms a brood pouch covering the entire thoracic region. Telson rounded. Middle thoracic segments broader than anterior and posterior segments. Anterior plate of thoracic sternites broad, devoid of granules. G1 slightly curved laterally, broad at shoulder, apex pointed. G2 approximately one-third length of G1, inserted into base of G1.
Remarks: colour in life was not recorded.
3.7. Parasites
In a review paper of all known parasites of Cryptochiridae, the bopyrid parasite Carcinione platypleura Bourdon, 1983 was recorded from two Lithoscaptus sp. B specimens from N Sulawesi, Indonesia (RMNH.CRUS.D.54106, RMNH.CRUS.D.54172) [32]. Carcinione platypleura appears to be the most common parasite of gall crabs, with a wide distribution in the Indo-West Pacific. Furthermore, the parasitic epicaridean isopod Cabirnalia nausicaa Boyko & Van der Meij, 2018 was described from material obtained from various hosts, including Lithoscaptus sp. B (RMNH.CRUS.D.57236). Lithoscaptus sp. B was undescribed at the time but is now known as Lithoscaptus doughnut; hence, L. doughnut is an involuntarily host to C. platypleura and C. nausicaa parasites.
3.8. Concluding Remarks
Cryptochiridae show a mix of more widely distributed species and species seemingly confined to smaller geographic areas [7,18]. Here we showed that Lithoscaptus doughnut is a widely distributed species associated with P. peroni across its range. Parts of the range of P. peroni, from the the Red Sea to the Polynesian archipelago in the Pacific Ocean [30], still need to be sampled for the presence of this crab species. One L. doughnut sample was collected from a Cyphastrea chalcidicum corallith. We confidently showed that L. doughnut belongs to a clade with the type species L. paradoxus. The genus remains in urgent need of revision, and the diversity, host associations, and distribution patterns of many Lithoscaptus species are poorly understood. Further research is needed to solve the composite nature of Lithoscaptus and study its divergence across hosts and basins.
Author Contributions
Conceptualization, S.E.T.v.d.M.; Methodology, J.R.C.; Validation, S.E.T.v.d.M.; Formal analysis, J.R.C.; Investigation, J.R.C.; Resources, Y.T. and S.E.T.v.d.M.; Writing—original draft, J.R.C. and S.E.T.v.d.M.; Visualization, J.R.C.; Supervision, S.E.T.v.d.M.; Project administration, all authors; Funding acquisition, J.R.C. and S.E.T.v.d.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The genetic sequence data that supports this study’s finding are available on GenBank.
Acknowledgments
Fieldwork in Indonesia (Raja Ampat, 2007) was organised by Naturalis and the Research Centre for Oceanography (RCO-LIPI), under the umbrella of Ekspedisi Widya Nusantara (E-Win). We thank Farnis Boneka and Markus Lasut of Universitas Sam Ratulangi (Manado) for their help with the fieldwork in Bunaken (2008). Fieldwork in Lembeh Strait (2012) took place during a Marine Biodiversity Workshop based at the Bitung Field Station of RCO-LIPI, co-organized by Universitas Sam Ratulangi. We are grateful to LIPI and RISTEK for granting research permits. The 2010 Semporna Marine Ecological Expedition was jointly organised by WWF-Malaysia, Universiti Malaysia Sabah’s Borneo Marine Research Institute, Universiti Malaya’s Institute of Biological Sciences, and Naturalis. The research permits were granted by the Economic Planning Unit, Prime Minister’s Department, Sabah Parks, and Department of Fisheries Sabah. Fieldwork in the Red Sea (2013) was supported by the King Abdullah University of Science and Technology under the Biodiversity in the Saudi Arabian Red Sea program, award number CRG-1-BER-002 to Michael Berumen. For fieldwork in the Maldives (2015), the help of the University of Milano-Bicocca Marine Research and High Education Centre in Magoodhoo, the Ministry of Fisheries and Agriculture, Republic of Maldives, and the community of Magoodhoo, Faafu Atoll, is gratefully acknowledged. The fieldwork in Okinawa, Japan (2016), was carried out in collaboration with James Reimer of the University of the Ryukyus. The COI and 16S sequences were produced as part of a Naturalis Barcoding project. We thank Tao Xu and Arthur Anker for their excellent photos.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Reaka-Kudla, M.L. The Global Biodiversity of Coral Reefs: A Comparison with Rain Forests. In Biodiversity II: Understanding and Protecting Our Biological Resources; Reaka-Kudla, M.L., Wilson, D.E., Wilson, E.O., Eds.; Joseph Henry Press: Washington, WA, USA, 1997; pp. 83–108. [Google Scholar]
- Hoeksema, B.W.; van der Meij, S.E.T.; Fransen, C.H.J.M. The Mushroom Coral as a Habitat. J. Mar. Biol. Assoc. 2012, 92, 647–663. [Google Scholar] [CrossRef]
- Stella, J.S.; Pratchett, M.S.; Hutchings, P.A.; Jones, G.P. Coral-Associated Invertebrates: Diversity, Ecological Importance and Vulnerability to Disturbance. Oceanogr. Mar. Biol. Annu. Rev. 2011, 49, 43–104. [Google Scholar]
- Vehof, J.; van der Meij, S.E.T.; Türkay, M.; Becker, C. Female Reproductive Morphology of Coral-Inhabiting Gall Crabs (Crustacea: Decapoda: Brachyura: Cryptochiridae). Acta Zool. 2016, 97, 117–126. [Google Scholar] [CrossRef]
- van Tienderen, K.M.; van der Meij, S.E.T. Extreme Mitochondrial Variation in the Atlantic Gall Crab Opecarcinus hypostegus (Decapoda Cryptochiridae) Reveals Adaptive Genetic Divergence over Agaricia Coral Hosts. Sci. Rep. 2017, 7, 39461. [Google Scholar] [CrossRef] [PubMed]
- Bähr, S.; van der Meij, S.E.T. Red Fluorescence in Symbiotic Coral-Dwelling Gall Crabs. Galaxea J. Coral Reef Stud. 2019, 21, 27–28. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Bravo, H.; Paulay, G.; van der Meij, S.E.T. Diversification and Distribution of Gall Crabs (Brachyura: Cryptochiridae: Opecarcinus) Associated with Agariciidae Corals. Coral Reefs 2022, 41, 699–709. [Google Scholar] [CrossRef]
- Bähr, S.; van der Meij, S.E.T.; Terraneo, T.I.; Xu, T.; Benzoni, F. Interspecific Coral Competition Does Not Affect the Symbiosis of Gall Crabs (Decapoda: Cryptochiridae) and Their Scleractinian Hosts. Ecol. Evol. 2023, 13, e10051. [Google Scholar] [CrossRef]
- Kropp, R.K.; Manning, R.B. The Atlantic Gall Crabs, Family Cryptochiridae (Crustacea: Decapoda: Brachyura). Smithson. Contrib. Zool. 1987, 462, 1–11. [Google Scholar] [CrossRef]
- DecaNet Eds. DecaNet. 2023. Available online: https://www.decanet.info (accessed on 3 October 2023). [CrossRef]
- Bähr, S.; Johnson, M.L.; Berumen, M.L.; Hardenstine, R.S.; Rich, W.A.; van der Meij, S.E.T. Morphology and Reproduction in the Hapalocarcinus marsupialis Stimpson, 1859 Species Complex (Decapoda: Brachyura: Cryptochiridae). J. Crustac. Biol. 2021, 41, ruab052. [Google Scholar] [CrossRef]
- Fize, A.; Serène, R. Les Hapalocarcinidés du Viet-Nam. Archives du Museum National d’Histoire Naturelle; Sèptieme Série. 5; Archives de France: Paris, France, 1957; pp. 1–202. [Google Scholar]
- Kropp, R.K. Revision of the Genera of Gall Crabs (Crustacea: Cryptochiridae) Occurring in the Pacific Ocean. Pac. Sci. 1990, 44, 417–448. [Google Scholar]
- van der Meij, S.E.T. Host Relations and DNA Reveal a Cryptic Gall Crab Species (Crustacea: Decapoda: Cryptochiridae) Associated with Mushroom Corals (Scleractinia: Fungiidae). Contrib. Zool. 2015, 84, 39–57. [Google Scholar] [CrossRef]
- van der Meij, S.E.T. Host Species, Range Extensions, and an Observation of the Mating System of Atlantic Shallow-Water Gall Crabs (Decapoda: Cryptochiridae). Bull. Mar. Sci. 2014, 90, 1001–1010. [Google Scholar] [CrossRef]
- Huang, D.; Goldberg, E.E.; Chou, L.M.; Roy, K. The Origin and Evolution of Coral Species Richness in a Marine Biodiversity Hotspot. Evolution 2018, 72, 288–302. [Google Scholar] [CrossRef]
- Briggs, J.C.; Bowen, B.W. A Realignment of Marine Biogeographic Provinces with Particular Reference to Fish Distributions. J. Biogeogr. 2012, 39, 12–30. [Google Scholar] [CrossRef]
- van der Meij, S.E.T.; Berumen, M.L.; Paulay, G. A New Species of Fizesereneia Takeda & Tamura, 1980 (Crustacea: Brachyura: Cryptochiridae) from the Red Sea and Oman. Zootaxa 2015, 3931, 585–595. [Google Scholar] [CrossRef] [PubMed]
- van der Meij, S.E.T. A New Gall Crab Species (Brachyura, Cryptochiridae) Associated with the Free-Living Coral Trachyphyllia geoffroyi (Scleractinia, Merulinidae). Zookeys 2015, 500, 61–72. [Google Scholar] [CrossRef]
- van der Meij, S.E.T. The Coral Genus Caulastraea Dana, 1846 (Scleractinia, Merulinidae) as a New Host for Gall Crabs (Decapoda, Cryptochiridae), with the Description of Lithoscaptus tuerkayi sp. nov. Crustaceana 2017, 90, 1027–1038. [Google Scholar] [CrossRef]
- Kropp, R.K. The Status of Cryptochirus coralliodytes Heller and Lithoscaptus paradoxus Milne-Edwards (Brachyura: Cryptochiridae). Proc. Biol. Soc. Wash. 1988, 101, 872–882. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Schubart, C.D. Mitochondrial DNA and Decapod Phylogenies: The Importance of Pseudogenes and Primer Optimization. In Crustacean Issues 18: Decapod Crustacean Phylogenetics; Martin, J.W., Crandall, K.A., Felder, D.L., Eds.; CRC Press: Boca Raton, FL, USA, 2009; pp. 47–65. [Google Scholar]
- Sequencher; Version 4.10.1 DNA Sequence Analysis Software; Gene Codes Coorporation: Ann Arbor, MI, USA, 2010; Available online: http://www.genecodes.com/ (accessed on 21 December 2023).
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Wong, K.J.H.; Tsao, Y.F.; Qiu, J.W.; Chan, B.K.K. Diversity of Coral-Associated Pit Crabs (Crustacea: Decapoda: Cryptochiridae) from Hong Kong, with Description of Two New Species of Lithoscaptus A. Milne-Edwards, 1862. Front. Mar. Sci. 2023, 9, 1003321. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-Feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Juszkiewicz, D.J.; White, N.E.; Stolarski, J.; Benzoni, F.; Arrigoni, R.; Hoeksema, B.W.; Wilson, N.G.; Bunce, M.; Richards, Z.T. Phylogeography of Recent Plesiastrea (Scleractinia: Plesiastreidae) Based on an Integrated Taxonomic Approach. Mol. Phylogenetics Evol. 2022, 172, 107469. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; van der Meij, S.E.T. Cryptic Marine Biota of the Raja Ampat Island Group; Naturalis Biodiversity Center: Leiden, The Netherlands, 2008. [Google Scholar]
- Boyko, C.B.; van der Meij, S.E.T. A Trifecta of Swiftian Symbioses: Stony Corals, Gall Crabs and Their Parasites (Scleractinia; Brachyura: Cryptochiridae; Isopoda: Epicaridea and Cirripedia: Rhizocephala). Zool. J. Linn. Soc. 2018, 184, 304–329. [Google Scholar] [CrossRef]
- Norton, D.A.; Carpenter, M.A. Mistletoes as Parasites: Host Specificity and Speciation. Trends Ecol. Evol. 1998, 13, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Quek, Z.B.R.; Jain, S.S.; Richards, Z.T.; Arrigoni, R.; Benzoni, F.; Hoeksema, B.W.; Carvajal, J.I.; Wilson, N.G.; Baird, A.H.; Kitahara, M.V.; et al. A Hybrid-Capture Approach to Reconstruct the Phylogeny of Scleractinia (Cnidaria: Hexacorallia). Mol. Phylogenetics Evol. 2023, 186, 107867. [Google Scholar] [CrossRef] [PubMed]
- van der Meij, S.E.T.; Klaus, S. Origin and Diversification of Coral-Dwelling Gall Crabs (Decapoda: Cryptochiridae) In Evolutionary Diversification of Coral-dwelling Gall Crabs (Cryptochiridae). Ph.D. Thesis, Naturalis Biodiversity Center & Leiden University, Leiden, The Netherlands, 2015; pp. 79–86. [Google Scholar]
- van der Meij, S.E.T. Lithoscaptus aquarius sp. nov (Decapoda: Cryptochiridae) Described from a Catalaphyllia jardinei (Scleractinia) out of the Aquarium Trade. Arthropoda 2023, 1, 350–359. [Google Scholar] [CrossRef]
Figure 1.
Map showing sampling locations (in blue) of Lithoscaptus doughnut other than the type locality.
Figure 1.
Map showing sampling locations (in blue) of Lithoscaptus doughnut other than the type locality.

Figure 2.
Haplotype network of Lithoscaptus doughnut based on COI sequences (n = 12). Marks across the branches indicate the amount of different nucleotides between haplotypes. The small coloured circles correspond to a single sequence, circle size increases when sequences are identical.
Figure 2.
Haplotype network of Lithoscaptus doughnut based on COI sequences (n = 12). Marks across the branches indicate the amount of different nucleotides between haplotypes. The small coloured circles correspond to a single sequence, circle size increases when sequences are identical.

Figure 3.
Haplotype network of Lithoscaptus doughnut based on 16S mtDNA (n = 12). Marks across branches indicate the amount of different nucleotides between haplotypes. The small circles correspond to a single sequence, circle size increases when sequences are identical.
Figure 3.
Haplotype network of Lithoscaptus doughnut based on 16S mtDNA (n = 12). Marks across branches indicate the amount of different nucleotides between haplotypes. The small circles correspond to a single sequence, circle size increases when sequences are identical.

Figure 4.
Colour in life of Lithoscaptus doughnut. Non-ovigerous females. (A,B) RMNH.Crus.D.54172; dorsal view and ventral view. (C,D) RMNH.Crus.D.54404; dorsal view and ventral view. Photos not to scale. Photos (A,B) by SET van der Meij; photos (C,D) by A Anker.
Figure 4.
Colour in life of Lithoscaptus doughnut. Non-ovigerous females. (A,B) RMNH.Crus.D.54172; dorsal view and ventral view. (C,D) RMNH.Crus.D.54404; dorsal view and ventral view. Photos not to scale. Photos (A,B) by SET van der Meij; photos (C,D) by A Anker.

Figure 5.
Male Lithoscaptus doughnut (RMNH.CRUS.D.53725): (A) dorsal view (B) ventral view. (C) close-up view of MXP-3. Photos by Tao Xu.
Figure 5.
Male Lithoscaptus doughnut (RMNH.CRUS.D.53725): (A) dorsal view (B) ventral view. (C) close-up view of MXP-3. Photos by Tao Xu.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Overview of material used for genetic analyses.
| Voucher Code | COI | 16S | Host Coral | Country | Locality |
|---|---|---|---|---|---|
| CEL-Hapa-040/ASIZCR | OP103644 | n/a | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Hong Kong | Basalt Island, Sai Kung |
| CEL-Hapa-006 | OP103613 | OP114856 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Hong Kong | Long Ke Tsai, Sai Kung |
| RMNH.CRUS.D.54169 | OR710957 | OR711069 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Tanjung Nanas II, Lembeh Isl., N Sulawesi |
| RMNH.CRUS.D.54172 | OR710956 | OR711070 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Pulau Abadi, Lembeh Isl., N Sulawesi |
| RMNH.CRUS.D.54106 | OR710954 | OR711068 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Lekuan III, Bunaken Isl., N Sulawesi |
| RMNH.CRUS.D.54063 | OR710953 | OR711067 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Tiwoho, Bunaken Isl., N Sulawesi |
| RMNH.CRUS.D.53890 | OR710955 | OR711066 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Mayalibit Bay, E Manil Isl., Raja Ampat, W Papua |
| RMNH.CRUS.D.57236 | NA | OR711074 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Japan | Mizugama, Okinawa |
| RMNH.CRUS.D.53725 | OR710951 | OR711064 | Cyphastrea chalcidicum (Forskål, 1775) | Malaysia | Mabul Isl., Eel Garden, Semporna |
| RMNH.CRUS.D.53740 | OR710952 | OR711065 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Malaysia | Mata Pahi Isl., Semporna |
| RMNH.CRUS.D.58330 | OR710958 | OR711072 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Maldives | Free Climbing, Faafu Atoll |
| RMNH.CRUS.D.58331 | OR710959 | OR711073 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Maldives | Route 66, Faafu Atoll |
| RMNH.CRUS.D.54404 | OR710960 | OR711071 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Saudi Arabia | Shib Radib, offshore of Farasan Banks |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Claassen, J.R.; Tuti, Y.; van der Meij, S.E.T. An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae). Arthropoda 2024, 2, 66-75. https://doi.org/10.3390/arthropoda2010005
AMA Style
Claassen JR, Tuti Y, van der Meij SET. An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae). Arthropoda. 2024; 2(1):66-75. https://doi.org/10.3390/arthropoda2010005
Chicago/Turabian StyleClaassen, Jorn R., Yosephine Tuti, and Sancia E. T. van der Meij. 2024. "An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae)" Arthropoda 2, no. 1: 66-75. https://doi.org/10.3390/arthropoda2010005