
Abstract
Diprotodontians are the morphologically and ecologically most diverse order of marsupials. However, an approximately 30-million-year gap in the Australian terrestrial vertebrate fossil record means that the first half of diprotodontian evolution is unknown. Fossil taxa from immediately either side of this gap are therefore critical for reconstructing the early evolution of the order. Here we report the likely oldest-known koala relatives (Phascolarctidae), from the late Oligocene Pwerte Marnte Marnte Local Fauna (central Australia). These include coeval species of Madakoala and Nimiokoala, as well as a new probable koala (?Phascolarctidae). The new taxon, Lumakoala blackae gen. et sp. nov., was comparable in size to the smallest-known phascolarctids, with body-mass estimates of 2.2–2.6 kg. Its bunoselenodont upper molars retain the primitive metatherian condition of a continuous centrocrista, and distinct stylar cusps B and D which lacked occlusion with the hypoconid. This structural arrangement: (1) suggests a morphocline within Phascolarctidae from bunoselenodonty to selenodonty; and (2) better clarifies the evolutionary transitions between molar morphologies within Vombatomorphia. We hypothesize that the molar form of Lumakoala blackae approximates the ancestral condition of the suborder Vombatiformes. Furthermore, it provides a plausible link between diprotodontians and the putative polydolopimorphians Chulpasia jimthorselli and Thylacotinga bartholomaii from the early Eocene Tingamarra Local Fauna (eastern Australia), which we infer as having molar morphologies consistent with stem diprotodontians.
Similar content being viewed by others



Introduction
Australia is renowned for the uniqueness of its biota, particularly its marsupials. This remarkable endemism is due mainly to their Gondwanan ancestry1,2, and long isolation following the continent’s separation from Antarctica around 45 million years ago (mya)3,4. Much of the morphological and ecological diversity of Australian marsupials is manifested within the order Diprotodontia, which comprises two suborders of primarily herbivorous taxa—Vombatiformes (wombats, koalas and extinct relatives) and Phalangerida (possums and kangaroos)—as well as Thylacoleonidae (marsupial ‘lions’; Diprotodontia incertae sedis5) (Fig. 1). Molecular-clock analyses suggest that Diprotodontia diverged from Agreodontia (Dasyuromorphia + Peramelemorphia + Notoryctemorphia) sometime between the latest Cretaceous and the earliest middle Eocene5,6,7,8,9,10,11,12,13,14. This is important from a biogeographic standpoint because it implies that diprotodontians had diverged before the final separation of Australia and Antarctica. Yet, no definitive fossil diprotodontians from prior to the late Oligocene are known, with the possible exception of diprotodontian-like tarsals from the early or middle Eocene of southern Argentina15.

Cladistic reconstruction of diprotodontian interrelationships. Relationships and divergence estimates for extant lineages follow Duchêne et al.6. Those of extinct lineages follow Beck et al.5, Travouillon et al.102, and Crichton et al.38,39. The divergence estimates for extinct vombatiforms and phalangeroids are speculatory.
Just two Australian fossil assemblages dating between 110 and 25 mya have yielded mammals16,17, leaving a massive gap spanning virtually the entire window of Australian marsupial origins18,19. In this window, taxon descriptions have only been published for metatherians (marsupials and stem-relatives) from the early Eocene (55 mya) Tingamarra Local Fauna (LF) of south-eastern Queensland, all on the basis of isolated dental, periotic and tarsal specimens17,20,21,22,23,24,25,26,27. These elements have been interpreted to lack unequivocal synapomorphies linking them with ordinal-level marsupial taxa known from the Australian late Oligocene1,9,17,19,21. Of these early Eocene taxa, only Djarthia murgonensis has been referred to Australidelphia (Australian marsupials + Microbiotheria of South America)22. Other Tingamarra LF marsupials have been argued to bear greater affinity with bunodont polydolopimorphians from South America23,25,27.
By the time the Australian terrestrial mammal record recommences, near the end of the Oligocene (c. 25 mya), most modern diprotodontian families are represented21,28,29. The distinctiveness of their dental and skeletal morphologies, as well as molecular disparity among modern representatives, suggest long pre-Miocene divergence intervals5,21,30,31. Each family is characterized by a specialized molar dentition that occupies a unique region in a morphospace that includes selenodont (multiple crescents), bunodont (bulbous cusps) and bilophodont (two transverse crests) forms. Identifying homologies between these molar architectures, and reconstructing the transformations that led to their acquisition, has long been considered important to resolving the broader problem of diprotodontian interrelationships5,30,32,33,34,35,36,37,38,39,40. The vombatiform radiation has been of particular interest; the extant representatives (koalas and wombats) capture but a small amount of the total morphological diversity known to have existed.
Koalas (Phascolarctidae) have selenodont molars, characterized by serial crescentic crests that function as leaf cutters during mastication (e.g.,41,42,43). Winge44 considered the molar form of the modern koala to be the least specialized among known diprotodontians, being only slightly modified from that of the archetypal metatherian condition. This view was accepted by Archer32, who suggested that selenodonty in diprotodontians could have arisen directly from a peramelemorphian (bandicoot) type morphology. This was based largely on the observation that modern koala and bandicoot molars lack a continuous crest (centrocrista) connecting the paracone and metacone, with the postparacrista and premetacrista instead linked to a cusp in the stylar cusp C and D positions, respectively32. It has since become evident that this morphology is derived within Peramelemorphia45,46,47; nevertheless, the view that vombatiforms transitioned from selenodonty to lophodonty has been widely supported30,33,34,35.
This study reports newly discovered fossils of koalas, and probable koalas, from the Pwerte Marnte Marnte deposit, southern Northern Territory, the likely-oldest Oligocene terrestrial mammal bearing site on the continent29,48 (Fig. 2). We describe a new genus and species characterized by a distinctive molar morphology that helps clarify evolutionary patterns within Diprotodontia, and may shed light on the phylogenetic affinities of enigmatic early Eocene metatherians from Australia. We also report the presence of two other koala species from the site.

Map of Australia depicting phascolarctid occurrences, modified from Price59 and Black et al.54. Fossil sites that have yielded phascolarctid remains are indicated with black circles. Phascolarctos cinereus former historical distribution is indicated in light green; current distribution in dark green; and translocated distribution in purple.
Systematic palaeontology
Infraclass Marsupialia Illiger, 1811
Order Diprotodontia Owen, 1877
Suborder Vombatiformes Woodburne, 1984
Infraorder ?Phascolarctomorphia Aplin & Archer, 1987
Family ?Phascolarctidae Owen, 1839
Lumakoala blackae gen. et sp. nov. (Fig. 3)
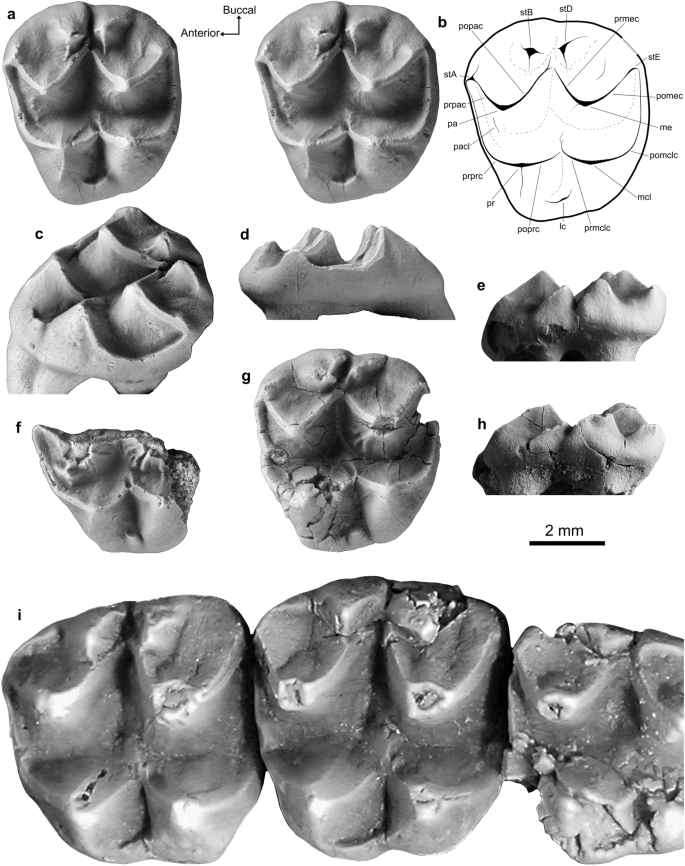
Lumakoala blackae gen. et sp. nov. upper molars. Left M 2? or 3 (Holotype, NTM P12012): (a) stereo occlusal view, with (b) an annotated line drawing; (c) posterolingually offset occlusal view; (d) anterior view; and (e) buccal view. Anterolingual half of LM1 (Paratype, NTM P12014) in (f) occlusal view. Left M ?2 or 3 (Paratype, NTM P12013) in (g) occlusal and (h) buccal views. Priscakoala lucyturnbullae left M1–M3 (Holotype, QM F20203), adapted from Black et al.58. The orientation arrows (Anterior, Buccal) relate only to the specimens in occlusion view. lc lingual cingulum, me metacone, mcl metaconule, pa paracone, pacl paraconule, pomclc postmetaconulecrista, pomec postmetacrista, popac postparacrista, poprc postprotocrista, pr protocone, prmclc premetaconulecrista, prmec premetacrista, prpac preparacrista, prprc preprotocrista, stA stylar cusp A, stB stylar cusp B, stD stylar cusp D, stE stylar cusp E.
Holotype. NTM P12012, left M2 or M3 (Fig. 3a–e).
Paratypes. NTM P12013, left M2 or M3 (Fig. 3g–h); NTM P12014, anterolingual half of left M1 (Fig. 3f).
Locality. Pwerte Marnte Marnte fossil site (24°21′ S 133°43′ E), deriving from an unnamed geological formation on the southern flank of the James Range, Northern Territory, Australia.
Local Fauna, and Age. Initial biochronological assessments have suggested that the Pwerte Marnte Marnte LF is late Oligocene age38,39,48, and may predate the oldest late Oligocene assemblages from the Etadunna and Namba Formations of the southern Lake Eyre Basin, perhaps corresponding to an as-yet-unnamed land mammal age immediately preceding the Etadunnan29,48.
Genus Etymology. Luma is Latin for ‘thorn’, in reference to the morphology of stylar cusps B and D, and their distinction from the postparacrista and premetacrista, respectively. The gender of the genus is feminine.
Species Etymology. Named for Karen Black, whose research has greatly extended our understanding of fossil phascolarctids and other vombatiforms.
Genus Diagnosis. As for the species until another is recognized.
Specific Diagnosis. A new species that appears most similar in upper molar morphology to members of the Phascolarctidae, with: a deep, U-shaped longitudinal valley between the primary buccal and lingual cusps, rather than being shallow and V-shaped as in selenodont vombatomorphians; and a paraconule on M1 (Fig. 3a–h).
Lumakoala blackae is relatively small compared to known phascolarctids, with upper molars that fall within the lower end of the size range of the smallest species of Litokoala. The taxon differs from all phascolarctids in having an M2 with: stylar cusps B and D that are somewhat conical and more structurally independent from the postparacrista and premetacrista, respectively; a postparacrista that continues beyond its junction with the premetacrista towards stylar cusp D; a wider stylar shelf, wherein the junction between the premetacrista and postparacrista is roughly mid-width between the buccal margin and the apices of the paracone and metacone; and more rounded buccal and posterior margins in occlusal view, leading to a generally rounder outline.
Lumakoala blackae differs from all other phascolarctids, unless otherwise noted, in having: proportionately wider molars relative to length (except Litokoala garyjohnstoni); more buccally situated protocone and metaconule apices, with a steeper buccal face of the protocone and metaconule compared with the lingual face (except L. garyjohnstoni); stylar cusp B taller than stylar cusp A on M2 (except Priscakoala lucyturnbullae); a small cuspate paraconule that extends up the anterolingual face of the paracone on M1 (except species of Phascolarctos); lacking a neometaconule (except Priscakoala lucyturnbullae); and lacking a hypertrophied anterobuccal terminus of the preprotocrista on M1 (except Priscakoala lucyturnbullae, species of Madakoala and Perikoala).
Lumakoala blackae is most similar to Priscakoala lucyturnbullae, sharing: simple enamel surface ornamentation without a neometaconule; stylar cusp B taller than stylar cusp A; cuspate stylar cusps B and D; and a wide stylar shelf. In addition to the differences identified with respect to all phascolarctids, L. blackae differs from Priscakoala lucyturnbullae in having: fine crenulations on the lingual faces of the paracone and metacone; a paraconule on M1; a stylar cusp A that is considerably larger, being similar in height to stylar cusp D; a prominent crista that descends from the apex of the metacone down the anterolingual face; a short and sometimes cuspate rather than elongate lingual cingulum; and a metaconule reduced relative to protocone on M2, resulting in an anterior moiety of markedly greater width than the posterior moiety.
Description. The upper molars of Lumakoala blackae (Fig. 3a–h) are low crowned, with fine crenulations on the lingual faces of the paracone and metacone. The molars have bunoselenodont crown morphology composed of four principal cusps, with the two anterior cusps representing the paracone (buccal) and protocone (lingual), and the posterior two representing the metacone (buccal) and metaconule (lingual). Two additional, somewhat conical cusps are present, buccal to the postparacrista and premetacrista, which are referred to as stylar cusps B and D, respectively (see Supplementary 1 for discussion of stylar cusp homologies). The cusp in the anterobuccal corner of the tooth, at the terminus of the preparacrista, is referred to as stylar cusp A.
Specimens NTM P12012 and NTM P12013 are both considered to represent M2, or possibly M3, because: tooth width tapers anteroposteriorly; and there is a prominent buccolingually extensive depression spanning the anterior and posterior faces, deriving from contact with the neighboring upper molars. Based on comparison with the molars of Priscakoala lucyturnbullae, NTM P12012 and NTM P12013 are thought to most likely represent M2, rather than M3 because, in the latter, stylar cusps B and D are markedly reduced. The specimens attributed to the M2 position are trapezoidal from occlusal view with rounded tooth corners, deriving from a strongly convex buccal margin and a lingual margin that curves posterobuccally. The molar proportions taper from anterior to posterior and buccal to lingual (Fig. 3). The anterior width of NTM P12012 is greater than posterior width by 7%, and greater than length by 12% (Table S1). The specimen NTM P12013 is thoroughly fractured, and consequently has slightly inflated dimensions (Table S1). On NTM P12013, there is also damage to the postprotocrista, the apex of the metacone, and the posterobuccal corner of the tooth. Greatest wear is present along the edges of the cristae. Description of the M2 morphology is based primarily on the holotype, NTM P12012, owing to its better preservation.
The metacone is the largest cusp, and the metaconule is the smallest (Fig. 3). The buccal basins of the paracone and metacone are only slightly concave as they slope from the apices to meet the boundaries of the stylar cusps. The buccal margin is open between each of (1) the stylar cusps A and B, and (2) stylar cusp D and the terminus of the postmetacrista (Fig. 3). The paracone apex is positioned slightly anterior relative to the protocone apex, and slightly buccal relative to the metacone apex. A linear preparacrista descends anterobuccally to a slightly raised cuspate structure in the anterobuccal corner of the tooth, representing stylar cusp A. Two cristae descend from the stylar cusp A: one posterobuccally, terminating anterior to stylar cusp B; and the other anterolingually, that is then continuous with the preprotocrista. The latter is not hypertrophied anteriorly into a cuspate structure (referred to as a parastyle in Black et al.49). A linear postparacrista descends posterobuccally, meeting the premetacrista proximate to the lingual extremity of stylar cusps B and D. The postparacrista continues weakly beyond its juncture with the premetacrista towards stylar cusp D before terminating. In NTM P12013, the postparacrista is continuous with a crista descending anterolingually from stylar cusp D, which, together, close off stylar cusp B. The postparacrista and premetacrista are structurally independent from stylar cusps B and D, respectively.
Stylar cusps B and D are somewhat conical;stylar cusp B is the tallest, being only c. 20% shorter than the paracone (Fig. 3e–h). Several short cristae descend from the apices of stylar cusps B and D, differentially expressed between NTM P12012 and NTM P12013 (Fig. 3). In NTM P12012, the most prominent crista descending from the apex of stylar cusp B is oriented anterolingually, weakly parallel to the postparacrista. From stylar cusp D, one crista is oriented posterolingually, parallel to the premetacrista, and a second is oriented posteriorly. By comparison, on NTM P12013, the most prominent crista descending from stylar cusp B is oriented posterobuccally; and from stylar cusp D, one is oriented anterolingually and another posterobuccally, together forming a crest. No structures link the stylar cusps. On NTM P12012, a faint ridge descends the metacone buccal basin anterior to the postmetacrista. The postmetacrista descends posterobuccally from the apex of the metacone, towards the posterobuccal tooth corner where it then arches ventrally, becoming continuous with the postmetaconulecrista. There is a slight swelling at the posterobuccal terminus of the postmetacrista, which represents stylar cusp E. The paracone and metacone are each somewhat conical in profile, wherein the lingual and buccal faces are similarly sloped. A faint crista descends from the apex of the metacone down the anterolingual face, terminating before the longitudinal basin. On NTM P12013, the crista is more prominent and bifurcates midway down the anterolingual face of the metacone.
The protocone is positioned slightly more posteriorly than the paracone, and slightly more lingual than the metaconule (Fig. 3). A crescentic preprotocrista descends anterobuccally from the apex of the protocone, curving as it spans the anterior margin of the tooth. On specimens NTM P12012 and NTM P12013, a faint ridge ascends the anterolingual face of the paracone, representing an incipient paraconule. On NTM P12013, there is also a short crista posterolingual to the apex of the paracone in the longitudinal valley that forms a slight swelling as it intercepts the preprotocrista (Fig. 3). This structure is considered a protostyle, the absence of which on NTM P12012 may be intraspecific variation. A linear postprotocrista descends posteriorly, intercepting the premetaconulecrista at the transverse valley. The premetaconulecrista continues beyond its junction with the postprotocrista before terminating. The postprotocrista and premetaconulecrista are each only slightly buccally inclined, together forming a relatively wide lingual angle of 115 degrees, compared to the 75-degree angle formed by the postparacrista and premetacrista. The metaconule apex is slightly posterior to the metacone apex. A crescentic postmetaconulecrista curves posterobuccally along the posterior margin of the tooth. The buccal faces of the protocone and metaconule are steeper than the lingual faces, with the latter representing roughly a third of the tooth width from occlusal view. A short lingual cingulum closes off the transverse valley at the lingual margin between the protocone and metaconule. On NTM P12013, the lingual cingulum is cuspate.
An anterolingual half of an upper molar (NTM P12014) has generally similar morphology to NTM P12012 and NTM P12013. This molar fragment is considered to represent an M1 on the basis that: the posterior moiety seems to have been wider than the anterior moiety; the paraconule is relatively more prominent; the depression on the anterior face deriving from the point of the contact with (presumably) the posterior face of P3, is small and circular (rather than buccolingually extensive). The paraconule is cuspate, positioned on the anterolingual face of the paracone. In common with specimen NTM P12014, two prominent cristae, aligned parallel one another, descend the anterolingual face of the metacone (Fig. 3). The placement of these two cristae relative to the metacone mirrors that of the paraconule relative to the paracone.
Madakoala sp. cf. M. devisi. Lower molar specimens of the largest phascolarctid in the Pwerte Marnte Marnte LF compare closely to those of Madakoala devisi in size (Table S2, Supplementary 2) and morphology (Fig. 4a–c; and Supplementary 3 for descriptions). Species of Madakoala are otherwise known from the late Oligocene Namba and Etadunna Formations in the southern Lake Eyre Basin50,51. We note that the lower molar specimens (NTM P12015 and NTM P12016) are more similar in morphology to M. devisi than the upper molar specimens (Fig. 4d–g; NTM P14005; NTM P12017), with the latter also similar to species of Perikoala (see Supplementary 3). It is possible that these specimens do not derive from the same taxon. The occurrence of representatives from both genera in a single assemblage has been reported in Ditjimanka LF of the Etadunna Formation50,51. At this stage, we consider it most parsimonious to treat these specimens as belonging to a single species allied to M. devisi.
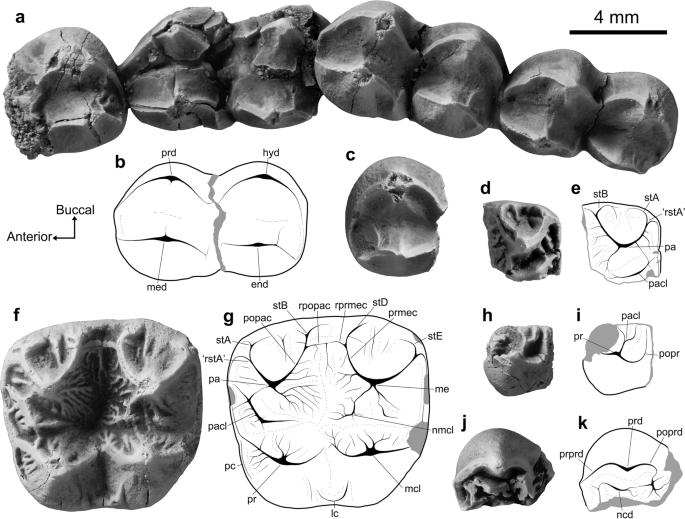
Specimens of Madakoala sp. cf. M. devisi and Nimiokoala sp. indet. from the Pwerte Marnte Marnte site, with annotated line drawings. Madakoala sp. cf. M. devisi: (a, b) partial right lower molar row preserving m1–m4 (NTM P12015); (c) left m3 talonid (NTM P12016); (d, e) paracone from right M 2? (NTM P12017); (f, g) left M1 (NTM P14005). Nimiokoala sp. indet.: (h, i) left protocone (NTM P12018); (j, k) right protoconid (NTM P12019). The orientation arrows (Anterior, Buccal) relate to the right lower molar (a, b, j, k), and left upper molar (f, g, h, i), specimens. end entoconid, hyd hypoconid, lc lingual cingulum, mcl metaconule, me metacone, med metaconid, ncd neomorphic cuspid, nmcl neometaconule, pa paracone, pacl paraconule, pc precingulum, popac postparacrista, popr postprotocrista, poprd postprotocristid, pr protocone, prd protoconid, prmec premetacrista, prprd preprotocristid, rpopac remnant postparacrista, rprmec remnant postmetacrista, rstA remnant stylar cusp A, stA Stylar cusp A, stB stylar cusp B, stD Stylar cusp D, stE stylar cusp E.
Nimiokoala sp. indet. Two molar fragments (NTM P12017, NTM P12018: Fig. 4h–k) compare best with species of Nimiokoala, though both are too fragmentary for systematic assessment beyond generic referral (Supplementary 4). The only other late Oligocene record of the genus, represented by a partial dentary (SAMA P19952) from the Namba Formation52, is not yet referred to a species. The type species, Nimiokoala greystanesi, is known from the Early to Middle Miocene of Riversleigh, north-west Queensland52,53,54,55,56.
Results of phylogenetic analyses. Parsimony analyses with all taxa included, and ordering of states where morphoclines were inferred, generated a strict consensus of 58 most parsimonious trees, each of 498 steps (Fig. S4, Supplementary 7). The strict consensus tree had a consistency index of 0.39 and a retention index of 0.77. Our maximum parsimony (strict consensus) and undated Bayesian analyses (majority rule consensus topology of the post-burnin trees) recovered similar topologies to the equivalent analyses of Crichton et al.39 (Figs. S2–S5, Supplementary 7). This reflects that relatively few modifications were made to the morphological characters of Crichton et al.39. In our unconstrained Bayesian analysis, Lumakoala blackae was weakly supported (BPP = 0.61) as the sister taxon to a moderately supported Phascolarctidae + Vombatomorphia (BPP = 0.82); whereas, under maximum parsimony, it formed a polytomy (BS = 0.88) with Cercartetus lepidus, Thylacoleonidae, and a weakly supported Phascolarctidae + Vombatomorphia (BS = 14%). The placement of L. blackae outside of Phascolarctidae + Vombatomorphia was supported by two character-transformations: stC not linked to paracone on M2 (C.I. = 0.33: char. 41); and metacone not linked to stD on M2 (C.I. = 0.50: char. 47). Thylacotinga bartholomaii consistently formed the sister taxon to Diprotodontia (BBP = 0.97; BS = 0.87).
The alternative hypothesis for the placement of Lumakoala blackae, tested by constraining it into a clade with phascolarctids, yielded 52 most parsimonious trees, each with a length of 499 steps, differing from that of the unconstrained topology by only one step. This was associated with a non-significant (P = > 0.05) difference in character state distribution, as indicated by the one-sided Templeton test (P = 0.28–0.43). As such, the hypothesis that Lumakoala blackae is a phascolarctid cannot be rejected. Strict consensus of the most parsimonious trees from the constrained analysis recovered Phascolarctidae as a polytomy composed of Lumakoala blackae, Priscakoala lucyturnbullae, and a clade comprising the remaining koala taxa (Fig. S5, Supplementary 7).
Discussion
The discovery of phascolarctids in the Pwerte Marnte Marnte fossil site provides the first record of koalas from the Northern Territory in both the historical and fossil records. Assuming Lumakoala blackae is indeed a phascolarctid, the site preserves three koala species, representing the equal highest number recorded from a single deposit, along with Camel Sputum Site from the early Miocene (Faunal Zone B) of Riversleigh57,58. Surprisingly, the fossil sites from the late Oligocene (Faunal Zone A) of Riversleigh have not yielded any phascolarctids55,56. From the late Oligocene Namba and Etadunna Formations of central Australia, phascolarctids are represented in most local faunas, with a single taxon in most, and two in some50. It has been suggested that the high early Miocene koala diversity from Riversleigh may correlate with greater plant diversity in these presumably rainforest communities54. The high phascolarctid diversity in the Pwerte Marnte Marnte LF may thus also indicate the presence of high plant diversity in a wooded paleoenvironment.
If Lumakoala blackae gen. et sp. nov. is a phascolarctid, it is the tenth genus and among the geologically oldest-known. Its discovery further emphasizes the well-noted54,59 phascolarctid turnover from the late Oligocene (Lumakoala, Madakoala, Perikoala, Nimiokoala and Litokoala) to early Miocene (Nimiokoala and Litokoala, Priscakoala). Lumakoala blackae was relatively small, yielding body mass estimates of 2.2–2.6 kg (Table S3, Supplementary 8) based on regression equations60 of M2 and M3 length. This falls well within the size range of the modern brushtail possum Trichosurus vulpecula (1.5–4.5 kg61), and is markedly smaller than other reportedly plesiomorphic phascolarctids (e.g., species of Priscakoala, Madakoala, Perikoala, Koobor and Invictokoala), all of which fall within the size range of the modern koala species (see54: 4.1–13.5 kg62). Indeed, L. blackae is among the smallest-known phascolarctids, together with species of Litokoala (measurements in Table S1, Supplementary 2).
Unlike the selenodont molars of unequivocal phascolarctids, those of L. blackae have a wide stylar shelf with large, conical stylar cusps B and D, that are unconnected from the postparacrista and premetacrista, respectively (Fig. 5). From a functional standpoint, this crown morphology, and the associated wear pattern, suggest no occlusion between the stylar cusps and the hypoconid during the transverse (shearing) phase of the power stroke. Instead, stylar cusps B and D would have overhung the buccal face of the lower molars, presumably abetting puncture-crushing during the initial vertical phase. Direct occlusion between the stylar cusps and the hypoconid may, nonetheless, have been possible as an artifact of high tooth wear later in life history. More generally, the low crown height and poorly developed enamel crenulations are consistent with adaptation for relatively soft foods. We hypothesize that L. blackae had a primarily plant-based diet, although it may have opportunistically consumed insects like many extant possums62.

Concatenated phascolarctid interrelationships, contextualized by upper molars from representative taxa in occlusal view, with annotated line drawings. From top to bottom: Phascolarctos cinereus (M2, FUR 095); Madakoala devisi (M2, FUR unregistered; field number BC 20–112); Priscakoala lucyturnbullae (M2, QM F20203) adapted from Black et al.58; and the probable phascolarctid Lumakoala blackae, gen. et sp. nov. (Left M2 or 3, NTM P12012). Topology based on undated Bayesian analysis, presented as a majority rule consensus, with numbers at nodes representing Bayesian posterior probabilities (Fig. S3, Supplementary 7 for complete tree). Lumakoala blackae was excluded from this analysis; its hypothesized placement within Phascolarctidae is indicated by a dashed line. lc lingual cingulum, me metacone, mcl metaconule, pa paracone, pacl paraconule, popac postparacrista, pr protocone, prmec postmetacrista, prst protostyle, rpopac remnant postparacrista, rprmec remnant postmetacrista, rstA remnant stylar cusp A, stA stylar cusp A, stB stylar cusp B, stD stylar cusp D, stE stylar cusp E. Scale bar equals 2 mm.
The presence of this bunoselenodont molar morphology in a diprotodontian marsupial is phylogenetically significant. This is because the structural independence of stylar cusps B and D, and a continuous centrocrista (postparacrista + premetacrista), are unambiguous symplesiomorphies of the other australidelphian orders (Microbiotheria, Peramelemorphia, Dasyuromorphia and Notoryctemorphia), as well as in Didelphimorphia and many other metatherian clades (e.g., see 5).
Our unconstrained Bayesian phylogenetic analyses imply that L. blackae is a stem vombatiform, sister to Phascolarctidae + Vombatomorphia, though with weak support (BBP = 0.61; for complete trees, see Fig. S2, Supplementary 7). Inclusion of L. blackae within the analyses also resulted in lower support for monophyly of Phascolarctidae and Vombatiformes (Figs. S2 and S3, Supplementary 7), reflecting that the morphology known of the new taxon blurs the boundaries of these clades. This poor resolution may not be surprising given that L. blackae could only be scored for 18% of the morphological characters, as it is known only from upper molars. Furthermore, there are no obvious upper molar synapomorphies of Phascolarctidae, with the possible exception of a paraconule on M1, which is present in all phascolarctids except Priscakoala lucyturnbullae58. Based on a Templeton test, the tree length of the topology in which L. blackae is constrained to form a clade with definitive phascolarctids is not significantly worse than the unconstrained topology in which L. blackae is a stem vombatiform (Figs. S4 and S5, Supplementary 7). At this stage, we consider it most appropriate to consider L. blackae a probable phascolarctid.
Within Phascolarctidae, the upper molar morphology of L. blackae is most similar to that of P. lucyturnbullae. Both are low crowned, have prominent stylar cusps, simple enamel surface ornamentation, and lack a neometaconule (Fig. 5). Priscakoala lucyturnbullae has previously been viewed58 as the most plesiomorphic phascolarctid known to date because it: lacks a paraconule and neometaconule on M1; has a weakly expressed protostylid on m1; and has simple, uncrenulated, selenodont molars. On this basis, the molars of L. blackae could be considered slightly more derived than those of P. lucyturnbullae because a paraconule is present on M1 and also (although only weakly developed) on M2. Nevertheless, we argue that the upper molar morphology of L. blackae is, actually more plesiomorphic than that of P. lucyturnbullae, because it extends phascolarctid molar morphospace from variations on a strict selenodont form, to bunoselenodonty. From the more plesiomorphic to more derived end of this proposed phascolarctid morphocline, we observe: a reduction of the stylar shelf; integration of stylar cusps B and D into the postparacrista and premetacrista, respectively; reduction of stylar cusp B relative to stylar cusps A and D; and increasing obliquity of the angle formed at the postprotocrista–premetaconulecrista juncture (Fig. 5).
The molar morphology of Lumakoala blackae is also intermediate between that of the otherwise basal selenodont vombatomorphians and the archetypal metatherian condition (Fig. 6). In particular, it confirms that some selenodont vombatomorphians retain remnants of a continuous centrocrista, lingual to large cuspate stylar cusps B and D on M1; these include the ilariids Kuterintja ngama (see fig. 263) and Ilaria illumidens (Fig. 6), as well as the ?wynyardiid Muramura pinpensis. The atrophy of these structures occurs in a stepwise manner, from posterior to anterior along the molar row. The postparacrista and premetacrista are lost in more derived vombatiforms (Fig. 6).

Vombatiform interrelationships, contextualized by upper molars from representative taxa in occlusal view, with annotated line drawings. From top to bottom: the diprotodontoid, Raemeotherium yatkolai (RM1, SAMA P43060: photograph mirrored); ?wynyardiid, Namilamadeta crassirostrum (LM1, cast of AR 9745); ilariid, Ilaria illumidens (M1, SAMA P43055); and the probable phascolarctid, Lumakoala blackae, gen. et sp. nov., (isolated left M2 or 3, NTM P12012). The putative polydolopimorphian Chulpasia jimthorselli is represented at the bottom by an upper molar (left M1 or 2, QM F50411). Concatenated topology based on undated Bayesian analysis, presented as a majority rule consensus, with numbers at nodes representing Bayesian posterior probabilities (Fig. S3, Supplementary 7 for complete tree). Lumakoala blackae was excluded from this analysis; its hypothesized placement within Phascolarctidae is indicated by a dashed line. lc lingual cingulum, mcl metaconule, me metacone, pa paracone, pacl paraconule, popac postparacrista, pr protocone, prc precingulum, prmec premetacrista, prst protostyle, stA stylar cusp A, stB stylar cusp B, stD stylar cusp D, stE stylar cusp E. Scale bar equals 2 mm.
It is less clear how metatherian stylar cusp homologies translate to the somewhat bunolophodont molars of thylacoleonids and most early phalangeriforms (Fig. 7; see5,36,38,64 and references therein). Thylacoleonids have been interpreted, by some, as retaining the symplesiomorphic condition of a continuous centrocrista and a distinct stylar shelf38,65,66. It has also been suggested that that the small buccal-most structures on the M1 of thylacoleonids are actually neomorphic, and that the primary buccal cusps, traditionally referred to as the paracone and metacone, are actually stylar in origin5. Many early macropodoids and some phalangeroids bear a small cuspate ridge on the posterobuccal face of the paracone on M138, which has traditionally been identified as stylar cusp C36,67. However, many early phalangeriforms also bear central cusps on their upper molars, most commonly on the posterior moiety of M1. The latter configuration has led several authors to flag the possibility that the cusps traditionally referred to as the paracone and metacone may instead represent stylar cusps B and D, respectively68,69, and that any subsidiary structures buccal to them are neomorphic5.
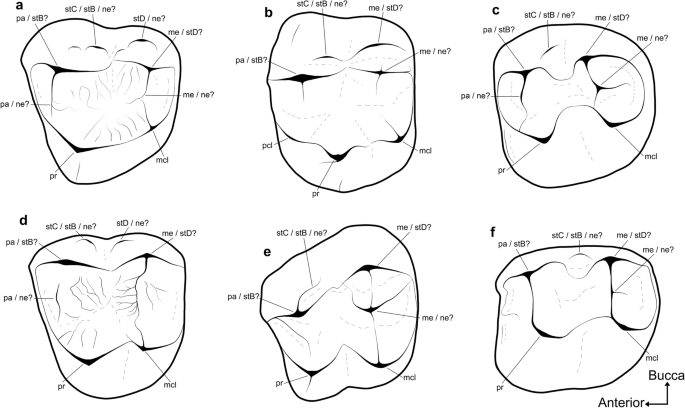
Annotated line drawings of upper molars from representative non-vombatiform diprotodontians, illustrating the ambiguity concerning homology of their buccal structures. The thylacoleonid (a) Lekaneleo roskellyae (RM1, mirrored; QM F23442); phalangeroids (b) Burramys wakefieldi (LM1; SAMA P40931), and (e) Durudawiri inusitatus (RM1, mirrored; QM F31468); macropodoids (c) Palaeopotorous priscus (RM1 or M2, mirrored; NMV P157547), and (f) Nambaroo tarrinyeri (LM1 or M2, 157528); and miminipossumid? (d) gen. et sp. indet. (LM1 or M2, Ditjimanka Local Fauna; SAMA P19856). mcl metaconule, me metacone, ne neomorphic structure, pa paracone, pcl protoconule, pr protocone, stB stylar cusp B, stC stylar cusp C, stD stylar cusp D. Not to scale.
In light of the stepwise morphocline observed between the molar topographies of phascolarctid genera (Fig. 5), and more generally between the vombatiform families (Fig. 6), we posit that the bunoselenodont molar morphology of L. blackae is close to the ancestral condition for Vombatiformes; this is congruent with the stem vombatiform position for this taxon in the unconstrained Bayesian analysis. It is noteworthy that body mass estimates for Lumakoala blackae (2.2–2.6 kg: Table S3, Supplementary 8) also fall within bounds predicted for the ancestor of Vombatiformes (1–5.5 kg)30. With the exception of the relative prominence of the metaconule, this morphology probably does not differ markedly from the ancestral condition for Diprotodontia. These observations imply that crown clade Diprotodontia (Vombatiformes + Phalangeriformes + Thylacoleonidae) retained the symplesiomorphic australidelphian condition of structurally independent stylar cusps B and D, and a continuous centrocrista. If correct, the reduction and/or integration of stylar cusps B and D into proximate structures has occurred convergently in several diprotodontian lineages as they evolved distinct, specialized dentitions.
Most molecular divergence estimates imply that diprotodontians were present when the early Eocene sediments near Murgon were deposited (e.g.,6,7,8,9,10,11,12), but none have been identified from the associated Tingamarra LF21. The plesiomorphic molar form of L. blackae provides an unexpected link between the molar morphologies of the Tingamarran taxa Chulpasia jimthorselli and Thylacotinga bartholomaii and the earliest undoubted diprotodontians (Fig. 6). Correspondingly, these Tingamarran taxa are plausible antecedent diprotodontians. Key molar attributes possessed by these Tingamarran taxa include: prominent and structurally independent stylar cusps B and D; a continuous centrocrista; a small but distinct metaconule; a posterobuccal corner that forms an angle > 60°; and a paracone and protocone that are subequal in size on M2 (see Supplementary 9 for further comparisons).
Thylacotinga bartholomaii and Chulpasia jimthorselli have been reconstructed as omnivores23,25, with body mass estimates of 3.2 kg and 0.2 kg, respectively27; the former being considerably larger than other mammals described to date from the Tingamarra LF. It has been suggested that these chulpasiines became extinct during the Palaeogene23. When included in our primarily vombatiform phylogenetic analyses, Thylacotinga bartholomaii was consistently recovered with strong support as the sister taxon to Diprotodontia (Fig. 6), to the exclusion of the representative agreodontians (for complete trees, Figs. S2–S5 Supplementary 7). A link between these Tingamarran taxa and definitive diprotodontians is attractive in its simplicity, supporting continuity between the early Eocene and late Oligocene Australian marsupial faunas, but we recognize that this may be an artifact of the very poor early Paleogene fossil record from Australia.
Thylacotinga bartholomaii and Chulpasia jimthorselli, together with its congener C. mattaueri from the late Paleocene or early Eocene of Peru, have been placed in the subfamily Chulpasiinae and referred to the primarily South American order Polydolopimorphia23. However, there remains contention concerning the relationship of C. jimthorselli to C. mattaueri, and thus the biogeographic inferences that can be drawn70,71. This is because all three taxa are known from isolated molars23,25,72, and have been grouped together on the basis of generalized bunodont features that are known to be highly homoplastic within Metatheria2,73,74. A formal phylogenetic analysis has yet to be published that tests their relationships to each other and to other bunodont and non-bunodont marsupialiforms. As such, a diprotodontian affinity for Thylacotinga bartholomaii and Chulpasia jimthorselli may not necessitate a pan-Gondwanan distribution for the australidelphian order (as would be implied if these two taxa are closely related to South American taxa). Nevertheless, it is worth recognizing that Lorente et al.15 described isolated australidelphian marsupial tarsals, from the La Barda locality (which now appears to be ~ 43.5 mya or younger75) in southern Argentina, that fell within crown Diprotodontia in their phylogenetic analysis. Putative chulpasiines have not been described from La Barda or a second site with a taxonomically similar mammalian fauna, Laguna Fria76, and it is unclear whether the La Barda tarsals belong to any of the taxa from the site known from dental remains2,15.
It has also been hypothesized, though contentiously, that Diprotodontia may be allied to Polydolopimorphia (the order to which Sige et al.23, referred Chulpasiinae) together deriving from a microbiotherian-like ancestor15,77,78,79,80,81,82,83. However, the phylogenetic affinities of polydolopimorphians are quite controversial, both with respect to their ordinal relationships and their monophyly as a single clade2,74. Furthermore, phylogenomic and retroposon insertion data provides compelling evidence that Microbiotheria is sister to the modern Australian marsupial radiation as whole6,14,84 rather than just Diprotodontia specifically.
Regardless of whether or not Thylacotinga and Chulpasia are in fact early diprotodontians, we hypothesize that their bunodont molar morphology—well suited to an omnivorous diet—is probably very close to the ancestral diprotodontian condition. Unlike tribosphenic metatherians, the power stroke in most extant diprotodontians is divided into a vertical phase and a transverse phase, with the latter accomplished by drawing the lower molars of the occluding side medially across the upper molars85. Interestingly, the upper molar specimens of T. bartholomaii show distinct wear facets on the crests linking stB and stD, presumably due to occlusion with the hypoconid, indicating a strong transverse component to the masticatory stroke. Indeed, much of the required change in molar morphology from the form of Thylacotinga and Chulpasia, to that of Lumakoala and other selenodont diprotodontians, can be explained as adaptations for facilitating a stronger transverse component for specialized herbivory. The formation of continuous transverse crests between the paracone and metacone, and stylar cusps B and D, respectively, increases shearing blade length; whereas loss of the link between the postparacrista and premetacrista allows the hypoconid to move further buccally, forming a larger crushing basin. These changes, alongside cranial adaptions enabling a larger transverse masticatory stroke, probably represented crucial steps in the transition from insectivorous to herbivorous diets in diprotodontian evolution.
Methods
The phascolarctid specimens reported in this study were recovered from ~ 2 tonnes of limestone quarried from the Pwerte Marnte Marnte fossil beds on expeditions in 2014 and 2020, led by A. Couzens and A. Crichton, respectively. The fossiliferous rock was processed during 2020–2022 in the Flinders University Palaeontology Laboratory using a combination of acetic acid (5–10%) etching and mechanical methods, e.g., rock saws and pneumatic micro-jack tools38. For comparative specimens used, see Supplementary 2.
Terminology. Higher-level systematic nomenclature follows Aplin and Archer86, with the exception of: the use of the suborder Phalangeriformes Szalay87, sensu Woodburne88; the superfamily Vombatoidea for the clade that includes Vombatidae + Mukupirnidae + Maradidae following Beck et al.30 and Crichton et al.39; and the subordinal placement of Thylacoleonidae as Diprotodontia incertae sedis following Beck et al.5. We also use ?Wynyardiidae to refer to species within the genera Namilamadeta and Muramura, because their inclusion within the family Wynyardiidae has never been robustly demonstrated33,89,90,91. Biostratigraphic nomenclature follows Woodburne et al.28, Archer et al.92 and Travouillon et al.93. The ages of vertebrate bearing localities from the Namba and Etadunna Formations follows Woodburne et al.28 and Megirian et al.29, and those of Riversleigh World Heritage Area follow Archer et al.92 and Travouillon et al.93 sensu Woodhead et al.53.
Molar position homology follows Luckett94. Molar cusp nomenclature follows Rich et al.95 with the exception of: the structure reported therein as the hypocone, which is here referred to as the metaconule following Tedford and Woodburne96; and the buccal-most cusps on the upper molars in diprotodontians, which are discussed in Supplementary 1.
Institution abbreviations. AR Archer Collection, University of New South Wales, Sydney, New South Wales; AMNH Department of Vertebrate Paleontology, American Museum of Natural History, New York, U.S.A.; FUR Palaeontology Department, College of Science and Engineering, Flinders University, Bedford Park, Adelaide, South Australia; NMV P Palaeontology, Museum Victoria, Melbourne; NTM P Museum of Central Australia, Museum and Art Gallery of the Northern Territory, Alice Springs; QM F, Queensland Museum Fossil Collection, Brisbane; S SAMA P Palaeontology, South Australian Museum, Adelaide, South Australia.
Phylogenetic analyses. To assess the phylogenetic relationships of Lumakoala blackae, we adopted the morphological dataset from Crichton et al.39, built on that of Beck et al.30, comprising 124 craniodental and 20 postcranial characters, which were scored for 46 taxa. We included a further 13 taxa, for a total of 59, namely: Lumakoala blackae gen. et sp. nov., Koobor notabilis, Stelakoala riversleighensis and Invictokoala monticola (Phascolarctidae); Thylacotinga bartholomaii (putative polydolopimorphian); Djarthia murgonensis (Australidelphia); Naraboryctes philcreaseri (Notoryctemorphia); ‘Bulungu spp.’ (Peramelemorphia); Nimbacinus dicksoni, Muribacinus gadiyuli and Badjcinus turnbulli (Thylacinidae); as well as Keeuna woodburnei and Ankotarinja tirarensis (putative dasyuromorphians). To accommodate additional outgroup agreodontians, and changes to the homology of stylar cusp B in Diprotodontia, while minimizing pseudoreplication: three characters were deleted (char. 27, Lophodonty; 37, Preparacrista; 49, Relative placement of stC on M1); 19 were modified; and 17 were added. The resulting character matrix comprised 138 craniodental and 20 post cranial characters (see Supplementary 5 & 6).
Undated Bayesian analysis of the morphological dataset was carried out in MrBayes 3.2.7a 97, using the Markov Chain Monte Carlo (MCMC) approach, with gamma rate variability implemented for morphological data maintaining the assumption that only variable characters were scored. The Bayesian analyses were run for 15 million generations, using four independent runs of four chains (one cold and three heated chains, with the temperature of the heated chains set to the default value of 0.2), sampling trees every 1000 generations and a burn-in fraction of 25%. The post-burn-in trees were summarised using a majority rule consensus of all compatible groups, with Bayesian posterior probabilities as support values.
Maximum parsimony analyses were also performed on the morphological dataset, in TNT version 1.5 98. The tree search involved an initial “new technology” search with sectorial search, ratchet, drift, and tree fusing that was run until the same minimum tree length was found 1000 times. From these saved trees a “traditional” search was applied using the tree bisection resection (TBR) swapping algorithm, with the resulting most parsimonious trees combined into a strict consensus tree. Support values for branch nodes were calculated using 2000 standard bootstrap replicates, implemented using a “traditional” search, which results in output as absolute frequencies.
Additional parsimony analyses were implemented in PAUP 4.0a16999, to test whether there were significant differences in the tree length and character state distribution of an alternative topological hypothesis, which included a positive constraint enforcing Lumakoala blackae within Phascolarctidae. To statistically compare these phylogenetic hypotheses, a single most parsimonious tree was randomly chosen from the unconstrained analyses, and compared against 50 random equal-most parsimonious trees from the constrained analyses using a one-sided Templeton Test100. This differs from a pairwise Templeton test in that the P value is halved101.
Data availability
Data generated and analysed during this study are included in this published article and its supplementary information file (available at https://doi.orgTBA).
References
Beck, R. M. D. In Handbook of Australasian Biogeography, Ch. 13 (ed. Ebach, M. C.) 329–366 (CRC Press, 2017).
Beck, R. M. D. In American and Australasian marsupials: An Evolutionary, Biogeographical, and Ecological Approach, Ch. 1 (eds. Cáceres, N. C. & Dickman, C. R.) 23–87 (Springer, 2023).
White, L. T., Gibson, G. M. & Lister, G. S. A reassessment of paleogeographic reconstructions of eastern Gondwana: Bringing geology back into the equation. Gondwana Res. 24, 984–998 (2013).
Stotz, I. L., Tassara, A. & Iaffaldano, G. Pressure-driven Poiseuille flow inherited from Mesozoic mantle circulation led to the Eocene separation of Australia and Antarctica. J. Geophys. Res. Solid 126, 9945. https://doi.org/10.1029/2020JB019945 (2021).
Beck, R. M. D., Voss, R. S. & Jansa, S. A. Craniodental morphology and phylogeny of marsupials. Bull. Am. Mus. Nat. Hist. 457, 1–352 (2022).
Duchêne, D. A. et al. Analysis of phylogenomic tree space resolves relationships among marsupial families. Syst. Biol. 67, 400–412 (2018).
Mitchell, K. J. et al. Molecular phylogeny, biogeography, and habitat preference evolution of marsupials. Mol. Biol. Evol. 31, 2322–2330 (2014).
Upham, N. S., Esselstyn, J. A. & Jetz, W. Inferring the mammal tree: Species-level sets of phylogenies for questions in ecology, evolution, and conservation. PLoS Biol. 17, e3000494 (2019).
Beck, R. M. D. A dated phylogeny of marsupials using a molecular supermatrix and multiple fossil constraints. J. Mammal. 89, 175–189 (2008).
Meredith, R. W., Krajewski, C., Westerman, M. & Springer, M. S. Relationships and divergence times among the orders and families of marsupialia. Mus. North. Ariz. Bull. 65, 383–406 (2009).
Meredith, R. W., Westerman, M. & Springer, M. S. A phylogeny of Diprotodontia (Marsupialia) based on sequences for five nuclear genes. Mol. Phylogen. Evol. 51, 554–571 (2009).
Meredith, R. W. et al. Impacts of the Cretaceous Terrestrial Revolution and KPg extinction on mammal diversification. Science 334, 521–524 (2011).
Álvarez-Carretero, S. et al. A species-level timeline of mammal evolution integrating phylogenomic data. Nature 602, 263–267 (2022).
Feng, S. et al. Incomplete lineage sorting and phenotypic evolution in marsupials. Cell 185, 1646–1660 (2022).
Lorente, M., Chornogubsky, L. & Goin, F. J. On the existence of non-microbiotherian Australidelphian marsupials (Diprotodontia) in the Eocene of Patagonia. Palaeontology 59, 533–547 (2016).
Hocknull, S. A. et al. Time is of the essence: three case studies from Queensland, Australia, demonstrating responses to fossil site loss through mining and development over differing time scales. J. Vertebr. Paleontol., Program and Abstracts 2019, 118.
Godthelp, H., Archer, M., Cifelli, R., Hand, S. J. & Gilkeson, C. F. Earliest known Australian Tertiary mammal fauna. Nature 356, 514–516 (1992).
Long, J. A., Archer, M., Flannery, T. F. & Hand, S. Prehistoric Mammals of Australia and New Guinea: One Hundred Million Years of Evolution (Univ. of New South Wales Press, Sydney, and Johns Hopkins Univ. Press, 2002).
Archer, M. et al. The evolutionary history and diversity of Australian mammals. Aust. Mammal. 21, 1–45 (1999).
Vickers-Rich, P. In Vertebrate Palaeontology of Australasia (eds. Vickers-Rich, P. et al.) 721–808 (Pioneer Design Studio and Monash University Publications Committee, 1991).
Black, K. H., Archer, M., Hand, S. J. & Godthelp, H. In Earth and Life: Global Biodiversity, Extinction Intervals and Biogeographic Perturbations Through Time (ed. Talent, J. A.) 983–1078 (Springer, 2012).
Beck, R. M. D., Godthelp, H., Weisbecker, V., Archer, M. & Hand, S. J. Australia’s oldest marsupial fossils and their biogeographical implications. PLoS ONE 3, e1858 (2008).
Sigé, B. et al. Chulpasia and Thylacotinga, late Paleocene-earliest Eocene trans-Antarctic Gondwanan bunodont marsupials: New data from Australia. Geobios 42, 813–823 (2009).
Beck, R. M. D. An ‘ameridelphian’ marsupial from the early Eocene of Australia supports a complex model of Southern Hemisphere marsupial biogeography. Naturwissenschaften 99, 715–729 (2012).
Archer, M., Godthelp, H. & Hand, S. J. Early Eocene marsupial from Australia. Kaupia 3, 193–200 (1993).
Godthelp, H., Wroe, S. & Archer, M. A new marsupial from the Early Eocene Tingamarra Local Fauna of Murgon, Southeastern Queensland: A prototypical Australian marsupial?. J. Mamm. Evol. 6, 289–313 (1999).
Beck, R. M. D. A peculiar faunivorous metatherian from the Early Eocene of Australia. Acta Palaeontol. Pol. 60, 123–129 (2013).
Woodburne, M. O. et al. Land mammal biostratigraphy and magnetostratigraphy of the Etadunna Formation (Late Oligocene) of South Australia. J. Vert. Paleontol. 13, 483–515 (1994).
Megirian, D., Prideaux, G. J., Murray, P. F. & Smit, N. An Australian land mammal age biochronological scheme. Paleobiology 36, 658–671 (2010).
Beck, R. M. D. et al. A new family of diprotodontian marsupials from the latest Oligocene of Australia and the evolution of wombats, koalas, and their relatives (Vombatiformes). Sci. Rep. 10, 1–13 (2020).
Black, K. Maradidae: A new family of vombatomorphian marsupial from the late Oligocene of Riversleigh, northwestern Queensland. Alcheringa 31, 17–32 (2007).
Archer, M. Phascolarctid origins and the potential of the selenodont molar in the evolution of diprotodont marsupials. Mem. Queensl. Mus. 17, 367–371 (1976).
Tedford, R. H. et al. The discovery of Miocene vertebrates, Lake Frome area, South Australia. BMR J. Aust. Geol. Geophys. 2, 53–57 (1977).
Black, K. Description of new material for Propalorchestes novaculacephalus (Marsupialia: Palorchestidae) from the mid Miocene of Riversleigh, northwestern Queensland. Alcheringa 30, 351–361 (2006).
Murray, P. Primitive marsupial tapirs (Propalorchestes novaculacephalus Murray and P. ponticulus sp. nov.) from the mid-Miocene of north Australia (Marsupialia: Palorchestidae). Beagle Records Museums Art Galleries Northern Territory 7, 39–51 (1990).
Ride, W. D. L. Jackmahoneya gen. nov. and the genesis of the macropodiform molar. Mem. Assoc. Austral. Palaeontol. 15, 441–459 (1993).
Archer, M. In Vertebrate Zoogeography & Evolution in Australasia (eds. Archer, M. & Clayton, G.) 585–631 (Hesperian Press, 1984).
Crichton, A. I., Worthy, T. H., Camens, A. & Prideaux, G. J. A new ektopodontid possum (Diprotodontia, Ektopodontidae) from the Oligocene of central Australia, and its implications for phalangeroid interrelationships. J. Vert. Paleontol. 2023, e2171299. https://doi.org/10.1080/02724634.2023.2171299 (2023).
Crichton, A. I. et al. A new species of Mukupirna (Diprotodontia, Mukupirnidae) from the Oligocene of central Australia sheds light on basal vombatoid interrelationships. Alcheringa https://doi.org/10.1080/03115518.2023.2181397 (2023).
Ride, W. D. L. On the fossil evidence of the evolution of the Macropodidae. Aust. Zool. 13, 6–16 (1971).
Lanyon, J. M. & Sanson, G. D. Koala (Phascolarctos cinereus) dentition and nutrition. I. Morphology and occlusion of cheekteeth. J. Zool. 209, 155–168 (1986).
Young, W. G., McGowan, M. & Daley, T. J. Tooth enamel structure in the koala, Phascolarctos cinereus: Some functional interpretations. Scann. Microsc. 1, 1925–1934 (1987).
Crompton, A. W., Owerkowicz, T. & Skinner, J. Masticatory motor pattern in the koala (Phascolarctos cinereus): A comparison of jaw movements in marsupial and placental herbivores. J. Exp. Zool. A: Ecol. Integr. Physiol. 313, 564–578 (2010).
Winge, H. The Interrelationships of the Mammalian Genera: Vol. 1 Monotremata, Marsupialia, Insectivora, Chiroptera, Edentata 1–418 (C. A. Reitzels Forlag, 1941).
Muirhead, J. & Filan, S. L. Yarala burchfieldi, a plesiomorphic bandicoot (Marsupialia, Peramelemorphia) from Oligo-Miocene deposits of Riversleigh, Northwestern Queensland. J. Paleontol. 69, 127–134 (1995).
Travouillon, K. J., Beck, R. M. D. & Case, J. A. Upper Oligocene–lower-Middle Miocene peramelemorphians from the Etadunna, Namba and Wipajiri formations of South Australia. Alcheringa 45, 109–125 (2021).
Travouillon, K. J., Gurovich, Y., Archer, M., Hand, S. J. & Muirhead, J. The genus Galadi: Three new bandicoots (Marsupialia, Peramelemorphia) from Riversleigh’s Miocene deposits, northwestern Queensland, Australia. J. Vert. Paleontol. 33, 153–168 (2013).
Murray, P. F. & Megirian, D. The Pwerte Marnte Marnte Local Fauna: A new vertebrate assemblage of presumed Oligocene age from the Northern Territory of Australia. Alcheringa 30, 211–228 (2006).
Black, K. H., Louys, J. & Price, G. J. Understanding morphological variation in the extant koala as a framework for identification of species boundaries in extinct koalas (Phascolarctidae; Marsupialia). J. Syst. Palaeontol. 12, 237–264 (2013).
Rich, T. H. et al. In Vertebrate Palaeontology of Australasia (eds. Vickers-Rich, P. et al.) 1005–1058 (Pioneer Design Studio in cooperation with the Monash University Publications Committee, 1991).
Woodburne, M., Tedford, R., Archer, M. & Pledge, N. In Possums and Opossums: Studies in Evolution (ed. Archer, M.) 293–317 (Surrey Beatty & Sons Pty Ltd, 1987).
Black, K. & Archer, M. Nimiokoala gen. nov. (Marsupialia, Phascolarctidae) from Riversleigh, northwestern Queensland, with a revision of Litokoala. Mem. Queensl. Mus. 41, 209–228 (1997).
Woodhead, J. et al. Developing a radiometrically-dated chronologic sequence for Neogene biotic change in Australia, from the Riversleigh World Heritage Area of Queensland. Gondwana Res. 29, 153–167 (2016).
Black, K. H., Price, G. J., Archer, M. & Hand, S. J. Bearing up well? Understanding the past, present and future of Australia’s koalas. Gondwana Res. 25, 1186–1201 (2014).
Black, K. H. Middle Miocene origins for tough-browse dietary specialisations in the koala (Marsupialia, Phascolarctidae) evolutionary tree: Description of a new genus and species from the Riversleigh World Heritage Area. Mem. Mus. Vic. 74, 255–262 (2016).
Archer, M. et al. Current status of species-level representation in faunas from selected fossil localities in the Riversleigh World Heritage Area, northwestern Queensland. Alcheringa 30, 1–17 (2006).
Travouillon, K. J., Escarguel, G., Legendre, S., Archer, M. & Hand, S. J. The use of MSR (Minimum Sample Richness) for sample assemblage comparisons. Paleobiology 37, 696–709 (2011).
Black, K. H., Archer, M. & Hand, S. J. New Tertiary koala (Marsupialia, Phascolarctidae) from Riversleigh, Australia, with a revision of phascolarctid phylogenetics, paleoecology, and paleobiodiversity. J. Vert. Paleontol. 32, 125–138 (2012).
Price, G. J. In Paleontology in Ecology and Conservation, Ch. 9 (ed. Louys, J.) 171–192 (Springer, 2012).
Myers, T. J. Prediction of marsupial body mass. Aust. J. Zool. 49, 99–118 (2001).
Moore, B. D., Wallis, I. R., Marsh, K. J. & Foley, W. J. In Conservation of Australia’s Forest Fauna (ed. Lunney, D.) 549–575 (Royal Zoological Society of New South Wales, 2004).
Van Dyck, S. & Strahan, R. Mammals of Australia 3rd edn. (New Holland Publishers, 2008).
Myers, T. & Archer, M. Kutjerintja ngama (Marsupialia, Ilariidae): A revised systematic analysis based on material from the late Oligocene of Riversleigh, northwestern Queensland. Mem. Queensl. Mus. 41, 379–392 (1997).
Gillespie, A. K. Two new marsupial lion taxa (Marsupialia, Thylacoleonidae) from the early and Middle Miocene of Australia. Alcheringa, 1–16. https://doi.org/10.1080/03115518.2022.2152096 (2023).
Murray, P., Wells, R. & Plane, M. In Possums and Opossums. Studies in Evolution (ed. Archer, M.) 433–466 (Surrey Beatty & Sons Pty Ltd, 1987).
Gillespie, A. Priscileo roskellyae sp. nov. (Thylacoleonidae, Marsupialia) from Riversleigh, northwestern Queensland. Mem. Queensl. Mus. 41, 321–328 (1997).
Flannery, T. & Rich, T. H. V. Macropodoids from the middle Miocene Namba Formation, South Australia, and the homology of some dental structures in kangaroos. J. Paleontol. 60, 418–447 (1986).
Woodburne, M. O., Pledge, N. & Archer, M. In Possums and Opossums: Studies in Evolution (ed. Archer, M.) 581–602 (Surrey Beatty & Sons Pty Ltd, 1987).
Crosby, K. & Archer, M. Durudawirines, a new group of phalangeroid marsupials from the Miocene of Riversleigh, northwestern Queensland. J. Paleontol. 74, 327–335 (2000).
Wilf, P., Cuneo, N. R., Escapa, I. H., Pol, D. & Woodburne, M. Splendid and seldom isolated: The paleobiogeography of Patagonia. Annu. Rev. Earth Planet. Sci. 41, 561–603 (2013).
Goin, F. J., Woodburne, M. O., Zimicz, A. N., Martin, G. M. & Chornogubsky, L. In A Brief History of South American Metatherians Ch. 3, 77–124 (Springer, 2016).
Crochet, J.-Y. & Sigé, B. Les mammifères de Chulpas (Formation Umayo, transition Crétacé-Tertiaire, Pérou): Données préliminaires. Docum. Lab. Géol. Lyon. 125, 97–107 (1993).
Beck, R. M. D. et al. A bizarre new family of Marsupialia (incertae sedis) from the Early Pliocene of northeastern Australia: Implications for the phylogeny of bunodont marsupials. J. Paleontol. 82, 749–762 (2008).
Beck, R. M. D. The skull of Epidolops ameghinoi from the Early Eocene Itaboraí Fauna, southeastern Brazil, and the affinities of the extinct marsupialiform order Polydolopimorphia. J. Mamm. Evol. 24, 373–414 (2017).
Gosses, J. et al. Facies interpretation and geochronology of diverse Eocene floras and faunas, northwest Chubut Province, Patagonia, Argentina. Geol. Soc. Am. Bull. 133, 740–752. https://doi.org/10.1130/b35611.1 (2021).
Tejedor, M. F. et al. New early Eocene mammalian fauna from western Patagonia, Argentina. Am. Mus. Novit. 2009, 1–43 (2009).
Chornogubsky, L. & Goin, F. J. A review of the molar morphology and phylogenetic affinities of Sillustania quechuense (Metatheria, Polydolopimorphia, Sillustaniidae), from the early Paleogene of Laguna Umayo, southeastern Peru. J. Vert. Paleontol. 35, e983238 (2015).
Goin, F. J., Candela, A. M., Bond, M., Pascual, R. & Escribano, V. Una Nueva “Comadreja” (Mammalia, Marsupialia, ?Polydolopimorphia) del Paleoceno de Patagonia. Argentina. Publ. Electron. Assoc. Paleontol. Argent. 5, 71–78 (1998).
Goin, F. J., Candela, A. M., Abello, M. A. & Oliveira, E. V. Earliest South American paucituberculatans and their significance in the understanding of ‘pseudodiprotodont’ marsupial radiations. Zool. J. Linn. Soc. 155, 867–884 (2009).
Goin, F. J. In Predators with Pouches: The Biology of Carnivorous Marsupials (eds. Jones, M. et al.) Ch. 3, 30–42 (CSIRO (Commonwealth Scientific and Industrial Research Organization), 2003).
Goin, F. J. & Candela, A. M. New Paleogene marsupials from the Amazon Basin of eastern Peru. Nat. Hist. Mus. Los. Angel. Cty. Sci. Ser. 40, 15–60 (2004).
Oliveira, E. V. & Goin, F. J. In Os Marsupiais do Brasil: Biología, Ecología e Evolução (eds. Cáceres, N. C. & Monteiro Filho, E. L. A.) 299–320 (Springer, 2006).
Oliveira, É. V. & Goin, F. J. A reassessment of bunodont metatherians from the Paleogene of Itaboraí (Brazil): Systematics and age of the Itaboraian SALMA. Rev. Bras. Paleontol. 14, 105–136 (2011).
Gallus, S., Janke, A., Kumar, V. & Nilsson, M. A. Disentangling the relationship of the Australian marsupial orders using retrotransposon and evolutionary network analyses. Genome Biol. Evol. 7, 985–992 (2015).
Crompton, A. W. Masticatory motor programs in Australian herbivorous mammals: Diprotodontia. Integr. Comp. Biol. 51, 271–281 (2011).
Aplin, K. P. & Archer, M. In Possums and Opossums: Studies in Evolution (ed. Archer, M.) XV–lxxii (Surrey Beatty & Sons Pty Ltd, 1987).
Szalay, F. S. Phylogenetic relationships of the marsupials. Geobios 15, 177–190 (1982).
Woodburne, M. O. In Mammals: Notes for a Short Course Vol. 8 (ed. Broadhead, T. W.) 48–71 (Knoxville, 1984).
Rich, T. H. V. & Archer, M. Namilamadeta snideri, a new diprotodontan (Marsupialia, Vombatoidea) from the medial Miocene of South Australia. Alcheringa 3, 197–208 (1979).
Pledge, N. S. In Possums and Opossums: Studies in Evolution (ed. Archer, M.) 393–400 (Surrey Beatty & Sons Pty Ltd, 1987).
Megirian, D., Murray, P., Schwartz, L. & Von Der Borch, C. Late Oligocene Kangaroo Well Local Fauna from the Ulta Limestone (new name), and climate of the Miocene oscillation across central Australia. Aust. J. Earth Sci. 51, 701–741 (2004).
Archer, M., Hand, S. J., Godthelp, H. & Creaser, P. Correlation of the Cainozoic sediments of the Riversleigh World Heritage fossil property, Queensland, Australia. Mem. Trav. E.P.H.E. Inst. Montpellier 21, 131–152 (1997).
Travouillon, K. J., Archer, M., Hand, S. J. & Godthelp, H. Multivariate analyses of Cenozoic mammalian faunas from Riversleigh, northwestern Queensland. Alcheringa 30, 323–349 (2006).
Luckett, W. P. In Mammal phylogeny: Mesozoic Differentiation, Multituberculates, Monotremes, Early Therians, and Marsupials (eds Szalay F. S., Novacek M. J., & McKenna M. C.) 182–204 (Springer, 1993).
Rich, T. H., Archer, M. & Tedford, R. H. Raemeotherium yatkolai, gen. et sp. nov., a primitive diprotodontid from the medial Miocene of South Australia. Mem. Mus. Vic. 39, 85–91 (1978).
Tedford, R. H. & Woodburne, M. O. In Possums and Opossums: Studies in Evolution (ed. Archer, M.) 401–418 (Surrey Beatty & Sons Pty, 1987).
Ronquist, F. et al. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542 (2012).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786 (2008).
Swofford, D. L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0b10. (Sinauer Associates, 2002).
Templeton, A. R. Phylogenetic inference from restriction endonuclease cleavage site maps with particular reference to the evolution of humans and the apes. Evolution 37, 221–244 (1983).
Goldman, N., Anderson, J. P. & Rodrigo, A. G. Likelihood-based tests of topologies in phylogenetics. Syst. Biol. 49, 652–670 (2000).
Travouillon, K. J., Butler, K., Archer, M. & Hand, S. J. Two new species of the genus Gumardee (Marsupialia, Macropodiformes) reveal the repeated evolution of bilophodonty in kangaroos. Alcheringa 46, 105–128 (2022).
Acknowledgements
We thank M.-A. Binnie and D. Stemmer, respectively, for providing access to South Australian Museum Palaeontology and Mammal Collections. We are grateful to W. Klein from Orange Creek Station for allowing us to access, and quarry fossiliferous rocks from, the Pwerte Marnte Marnte fossil site. We thank C. Burke, S. Arman, W. Handley, and G. Gully for organizing and/or assistance with the 2014 and 2020 field trips to the site. We acknowledge the Southern Arrernte People, known as the Pwerte Marnte Marnte Aboriginal Corporation, for their custodianship of the lands on which the fossil locality is situated. We also thank G. Price, and an anonymous reviewer, for their thoughtful comments and suggestions that improved this research.
Funding
AIC was supported by The Australian Government Research Training Program Scholarship. Support for the 2014 field trip to the site was provided by a Patterson Memorial Grant from the Society of Vertebrate Paleontology to AMCC.
Author information
Authors and Affiliations
Contributions
A.I.C. contributed reagents/materials/analysis tools, quarried fossiliferous limestone, found and prepared the fossil specimens, conceived the paper, analyzed the data, wrote the paper, and prepared figures and tables. T.H.W reviewed drafts of the paper. A.B.C reviewed drafts of the paper. A.M.C.C quarried fossiliferous limestone, reviewed drafts of the paper and wrote parts of the paper. R.M.D.B reviewed drafts of the paper and wrote parts of the paper. G.J.P. contributed reagents/materials/analysis tools, reviewed drafts of the paper and wrote parts of the paper. All authors contributed to developing the hypotheses presented in the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Crichton, A.I., Beck, R.M.D., Couzens, A.M.C. et al. A probable koala from the Oligocene of central Australia provides insights into early diprotodontian evolution. Sci Rep 13, 14521 (2023). https://doi.org/10.1038/s41598-023-41471-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-41471-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
