S.O.S. - Home
S.O.S. - Home
S.O.S. - Home

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
C O C U Y O<br />
ISSN 1607-2863<br />
COCUYO<br />
C<br />
A<br />
R<br />
T<br />
A<br />
INFO<br />
R<br />
A<br />
M<br />
IV<br />
T<br />
A<br />
Carta Informativa de los Zoólogos de Invertebrados de Las Antillas<br />
NÚMERO 15 OCTUBRE 2005<br />
Editores<br />
Julio A. Genaro<br />
Jorge L. Fontenla<br />
Cristina Juarrero<br />
S.O.S.<br />
La evidencia es abrumadora.<br />
! Los polinizadores están<br />
declinando! El<br />
empobrecimiento de los<br />
ecosistemas y la reducción del<br />
hábitat disminuyen cada vez<br />
más las interacciones vitales.<br />
Para su propia seguridad la<br />
humanidad debe tener en<br />
cuenta a los polinizadores y<br />
las innumerables especies de<br />
plantas que dependen de<br />
ellos..<br />
Polinizadores<br />
CONTENIDO<br />
PROYECTOS ACTUALES<br />
Contribución al estudio de la familia Psychidae<br />
(Lepidoptera)...................................................................3<br />
Nuevo periodo para la Sociedad Cubana de<br />
Zoología……………........................................................…3<br />
NOTAS CIENTIFICAS<br />
Primer hallazgo de Eunicea pinta (Octocorallia) para aguas<br />
cubanas......................................................................….3<br />
El antúrido Accalathura crenulata (Peracarida)………….…..4<br />
Los foraminíferos modernos de Cuba…………....………...5<br />
Leprolochus spinifrons (Araneae) en Las Antillas………...14<br />
Observaciones sobre Cyrtophora citricola (Araneae) en<br />
Haití………..................................................................….15<br />
Macroinvertebrados dulceacuícolas y calidad de las aguas<br />
en estaciones de la región oriental de Cuba.........……….15<br />
Nuevas adiciones a la entomofauna de Mil Cumbres, Pinar<br />
del Río………..................................................................21<br />
Los dípteros (Insecta) de Mil Cumbres, Pinar del Río.…..22<br />
Plagas de los almacenes de fondos de exhibición del<br />
Museo Nacional de Historia Natural de Cuba……………...24<br />
Moluscos del litoral de la bahía de Cienfuegos, Cuba.....27<br />
OBITUARIO<br />
Ha muerto Ernst Mayr (1904-2005)…………..............…...32<br />
M u s e o N a c i o n a l<br />
Historia Natural<br />
Obispo # 61, esquina a<br />
Oficios,Habana Vieja<br />
10 100, Cuba<br />
BIOCOMENTARIOS<br />
Entre lo histórico y lo lógico: el no-diálogo en la<br />
biogeografía..................................................................33<br />
LITERATURA RECIENTE/ 44<br />
cocuyo @ mnhnc.inf.cu
2 COCUYO 15, 2005<br />
Ernst Mayr<br />
Biólogo evolucionista,<br />
ornitólogo y filósofo<br />
filosofo<br />
5 Julio 1904 - 3 Febrero 2005<br />
Alemania<br />
Estados Unidos<br />
El ultimo sobreviviente de la edad<br />
de oro de la síntesis evolucionista
PROYECTOS ACTUALES<br />
La siguiente tesis fue defendida con éxito, el 29 de diciembre,<br />
en la Facultad de Biología, Universidad de La Habana para<br />
obtener el titulo académico de Master ¡FELICIDADES!<br />
Titulo: Contribución al estudio de la familia Psychidae (Insecta:<br />
Lepidoptera) en Cuba.<br />
Autor: Rayner Núñez Aguila.<br />
Tutor: MC. Alejandro Barro Cañamero<br />
Oponente: Dra. Vivian González<br />
Tribunal: Dra. Josefina Cao, Dra. Dania Prieto, Dr. Carlos<br />
Rodríguez<br />
RESUMEN: La familia Psychidae se encuentra entre los<br />
lepidópteros menos conocidos de la región Neotropical. Hasta<br />
el presente, solo se conocían cuatro especies cubanas y<br />
prácticamente no existía información sobre su historia natural.<br />
En este trabajo, se describen cinco especies nuevas para la<br />
Ciencia del género Paucivena, que se registra por primera vez para<br />
Cuba. El género Thyridopteryx y dos especies de Cryptothelea<br />
también constituyeron registros nuevos para la isla. En el<br />
archipiélago cubano sólo está presente una especie del género<br />
Oiketicus, O. kirbyi, que ha variado la forma de construir su<br />
estuche. La hembra del género Paucivena, desconocida hasta la<br />
fecha, así como la larva, la hembra y las pupas de ambos sexos del<br />
género Biopsyche se describen por primera vez a partir de<br />
ejemplares de B. thoracica. Se describen la larva y los adultos de<br />
ambos sexos de Prochalia licheniphilus. P. licheniphilus, Cryptothelea<br />
sp., B. thoracica y O. kirbyi se distribuyeron por toda la isla, en una<br />
amplia variedad de hábitats. Las especies de Paucivena están<br />
restringidas por el momento a los macizos montañosos,<br />
mientras que el resto de las especies se encuentra en una o dos<br />
localidades. Las especies de Paucivena fueron encontradas<br />
alimentándose de musgos (Orthostichidium guyanense) y hepáticas<br />
(Plagiochila sp.) y P. licheniphilus de Physcia alba, un liquen. Seis<br />
especies de plantas superiores constituyen hospederos nuevos<br />
para B. thoracica y tres para O. kirbyi. Avispas de las familias<br />
Chalcididae, Eupelmidae e Ichneumonidae constituyeron los<br />
primeros enemigos naturales registrados para P. licheniphilus y B.<br />
thoracica.<br />
Nuevo período para la Sociedad<br />
Cubana de Zoología<br />
Este es el nuevo Comité Ejecutivo de<br />
la Sociedad Cubana de Zoología, que de<br />
acuerdo a los nuevos estatutos la estará<br />
dirigiendo por cinco años -hasta el 2010. Hirám González<br />
continua reelecto como Presidente, quien ha desempañado un<br />
buen trabajo. Ya la Sociedad cuenta con un boletín electrónico<br />
llamado CartaCuba, de muy buen diseño y amplia circulación. El<br />
editor es Ariam Jimémenez (ariam@fbio.uh.cu)<br />
Nombre<br />
Dr. Hiram González Alonso<br />
MSc. Alejandro Barros Cañamero<br />
Lic. Santos Cubillas Hernández<br />
MSc. Nayla García Rodríguez<br />
Dra. Nereida Novoa Fernández<br />
Dr. Luis de Armas Chaviano<br />
Dr. Erick García Machado<br />
MSc. Roberto Alonso Bosch<br />
Presidente<br />
Vicepresidente Primero<br />
Vicepresidente Segundo<br />
Secretaria<br />
Tesorera<br />
Vocal<br />
Vocal<br />
Vocal<br />
Cargo<br />
NOTAS CIENTÍFICAS<br />
Primer hallazgo de Eunicea pinta (Cnidaria, Octocorallia)<br />
en aguas cubanas<br />
Galia Varona* y Carlos Varela**<br />
*Acuario Nacional de Cuba, 60 y 1ra, Playa, Ciudad de La Habana,<br />
Cuba.<br />
**Centro de Investigaciones Marinas, Universidad de la Habana, 16<br />
no. 114, Playa, Ciudad de La Habana, Cuba<br />
El inicio del estudio sistemático de los octocorales cubanos,<br />
data de la primera mitad del siglo XIX. Desde esa época hasta el<br />
presente, el número de especies registradas para nuestro país se<br />
ha incrementado notablemente con los aportes de Deichmann<br />
(1936), Guitart-Manday (1959), Bayer (1961), Behety (1975),<br />
Alcolado (1981), y García-Parrado y Alcolado (1996; 1998).<br />
Lalana et al., (2001), proporcionan un listado de los octocorales<br />
de nuestras aguas, donde se consignan las 77 especies<br />
pertenecientes a 11 familias, de las cuales la familia Plexauridae<br />
posee el mayor número de especies, con 31. El género Eunicea,<br />
perteneciente a esta familia, es el más rico en la región caribeña,<br />
con 13 especies, de las cuales 11 han sido registradas para Cuba.<br />
El objetivo del presente trabajo es consignar por primera vez la<br />
presencia en Cuba de la especie Eunicea pinta Bayer y Deichmann<br />
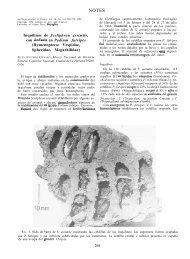
(Fig. 1A-D).<br />
El material estudiado consiste en un fragmento de colonia<br />
colectado mediante buceo autónomo a 15 m de profundidad, en<br />
el arrecife situado en la desembocadura del río Bacunayagua<br />
(239' LN y 8144' LW), al Norte de la provincia de Matanzas, en<br />
Febrero del 2003. Del fragmento colectado se extrajeron<br />
escleritos, mediante la disolución de la parte orgánica en<br />
hipoclorito de sodio al 5%. La composición espicular fue<br />
dibujada con ayuda de la cámara clara y comparada con el patrón<br />
de espículas de las restantes especies del género citadas para la<br />
región según Bayer (1961), y Guzmán y Cortés (1985). Estas<br />
espículas fueron conservadas posteriormente en alcohol en un<br />
tubo de ensayo, para su posterior fijación. El material se<br />
encuentra depositado en la Colección de Invertebrados del<br />
Centro de Investigaciones Marinas, Cnidarios No. 144.<br />
La especie Eunicea pinta fue colectada en el arrecife del<br />
Banco de Gran Bahama (2334' LN y 7633' LW), a una<br />
COCUYO 15, 2005 3
profundidad de 72 m. Esta especie ha sido registrada también<br />
para La Florida (Goldberg, 1973), a 44 m de profundidad y para<br />
Jamaica (Kinzie, 1973), a profundidades por debajo de los 60 m.<br />
Se puede considerar una especie rara, poco frecuente en<br />
muestreos, y de muy baja densidad.<br />
Según Bayer (1961), la forma delgada de la colonia (menor de<br />
3 mm de ancho en las ramas terminales), que la diferencia de las<br />
restantes especies pertenecientes al género, puede deberse a una<br />
preferencia por ambientes tranquilos, lo cual se corresponde con<br />
las condiciones encontradas en el lugar del hallazgo.<br />
Su patrón espicular sugiere una relación cercana con Eunicea<br />
mammosa Lamouroux. Sin embargo, las aberturas caliculares de<br />
E. pinta son mucho más pequeñas que en E. mammosa y los cálices<br />
están ausentes, a diferencia de lo que sucede con las restantes<br />
especies del subgénero Eunicea, que por lo general presentan<br />
cálices muy prominentes. Estas características morfológicas<br />
externas junto al patrón espicular, donde se destacan los<br />
escleritos de la capa externa en forma de mazas muy foliadas en<br />
forma de antorchas (Figura 1D), y los escleritos pequeños y<br />
ligeramente curvados de la armadura antocodial (Fig. 1B), nos<br />
llevan a la identificación de la especie como E. pinta.<br />
Las espículas de la capa axial (Fig. 1C), y la capa media (Fig.<br />
1A), son comunes a varias especies del género Eunicea, aunque su<br />
tamaño y grado de ornamentación varía en algunas de estas, y su<br />
presencia contribuye a ubicar a la especie dentro de este género.<br />
El color del fragmento recién colectado es gris pálido, casi<br />
blanco y se mantiene al secarse la colonia.<br />
A<br />
B<br />
REFERENCIAS<br />
Alcolado, P. 1981. Zonación de los gorgonáceos someros de<br />
Cuba y su posible uso como indicadores comparativos de<br />
tensión hidrodinámica sobre organismos del bentos. Inf.<br />
Cient. Tec., 187: 1-43.<br />
Bayer, F. M. 1961. The shallow water Octocorallia of the West Indian<br />
region. A manual for marine biologists. Martinus Nijhoff, The<br />
Hague. 373p.<br />
Behety, P. A. 1975. Nuevos reportes de gorgonáceos<br />
(Coelenterata) para Cuba. Serie Oceanológica 33:1-9.<br />
Deichman, E. 1936. The Alcyonaria of the Western part of the<br />
Atlantic Ocean. Mem. Mus. Comp. Zool., 53: 1-317.<br />
García-Parrado, P y P. M. Alcolado 1996. Nueva especie de<br />
Eunicea Lamouroux, 1816 (Cnidaria, Gorgonacea)<br />
encontrada en Cuba. Avicennia 4/5: 46-50.<br />
García-Parrado, P. y P. M. Alcolado 1998. Nuevos registros de<br />
Octocorales (Cnidaria) para la plataforma cubana. Avicennia<br />
8/9: 105-108.<br />
Goldberg, W. 1973. The ecology of the coral-octocoral<br />
communities of the southeast Florida coast: geomorfology,<br />
species composition and zonation. Bull. Mar. Sci., 23: 465-<br />
488.<br />
Guitart-Manday, D. 1959. Gorgonias del litoral de la costa Norte<br />
de Cuba. Publ. Acuario Nac., Serie Técnica 1: 1-24.<br />
Guzmán, H. y J. Cortez 1985. Organismos de los arrecifes<br />
coralinos de Costa Rica. IV: Descripción y distribución<br />
geográfica de los octocorales (Cnidaria, Anthozoa) de la<br />
Costa Caribe. Brenesia 24: 125-173.<br />
Kinzie, R. 1973. The zonation of West Indian gorgonians. Bull.<br />
Mar. Sci., 23: 93-155.<br />
Lalana, R.; M. Ortiz y C. Varela. 2001. Lista actualizada y<br />
bibliografía de los Celenterados (Cnidaria) y los Ctenóforos<br />
(Ctenophora), de aguas cubanas. Rev. Biol., 15 (2): 158-169.<br />
0,1 mm<br />
C<br />
Accalathura crenulata (Crustacea: Peracarida), el antúrido<br />
más grande del Archipiélago Cubano<br />
Manuel Ortiz, Rogelio Lalana y Carlos Varela<br />
D<br />
Fig. 1. Eunicea pinta. A: Espículas de la capa media de la rama<br />
terminal. B: Espículas de la armadura antocodial. C: Espículas de<br />
la capa axial de la rama terminal. D: Mazas de la capa externa de la<br />
rama terminal.<br />
Agradecimientos.- Agradecemos las facilidades otorgadas para<br />
la realización de este trabajo por el Proyecto Ramal “Evaluación<br />
ecológica de zonas de colecta del Acuario Nacional de Cuba”.<br />
Centro de Investigaciones Marinas (CIM), Universidad de La<br />
Habana , 16 no. 114, Playa, Ciudad de La Habana, Cuba<br />
El isópodo Accalathura crenulata (Richardson, 1901), fue<br />
descrito como Calathura crenulata, a partir de un ejemplar<br />
colectado en las aguas de las Bahamas, a casi 39 m de<br />
profundidad, entre las islas Nassau y Andros. Es una especie<br />
relativamente común en el Atlántico Occidental Tropical, que ha<br />
sido encontrado en varias localidades del Archipiélago Cubano.<br />
En la descripción original (Richardson, 190I), o en la<br />
monografía de Richardson, (1905), no se precisó su talla. En<br />
Puerto Rico, se ha encontrado una hembra de 14 mm (Menzies<br />
y Glynn, 1968). Kensley y Schotte (1989), lo citan para las aguas<br />
de las Carolinas, Georgia, Cuba, Puerto Rico, Cozumel en<br />
4 COCUYO 15, 2005
México, Carrie Bow Cay en Belize, hasta las costas de la Florida y<br />
el Golfo de México. La talla señalada para una hembra en este<br />
caso, fue de 16 mm. El ejemplar más grande estudiado Müller<br />
(1991), fue una hembra inmadura de 12. 5 mm.<br />
Los ejemplares de mayor talla, según la literatura disponible,<br />
fueron los citados por Schultz (1969), registrando una hembra<br />
de 18 mm, mientras que Koening (1972) al revisar el material<br />
obtenido en varias colectas en las aguas del Brasil, encontró otra<br />
desovada, de igual talla. El objetivo de este trabajo es registrar el<br />
hallazgo del ejemplar conocido, de<br />
mayor talla, en nuestras aguas y<br />
probablemente en el Mar Caribe.<br />
Accalathura crenulata (Fig. 1).<br />
Material estudiado: macho adulto; 20<br />
mm de largo; Playa La Barca, costa Sur<br />
de la Península de Guanahacabibes,<br />
Provincia de Pinar del Río; vii.2003;<br />
depositado en la Colección de<br />
Invertebrados del CIM, No 1289.<br />
Fig. 1. Vista lateral de<br />
Accalathura crenulata.<br />
Agradecimientos.- Nuestro reconocimiento a los estudiantes<br />
de la Facultad de Biología, de la Universidad de La Habana,<br />
participantes del Proyecto de investigación sobre Tortugas<br />
Marinas del CIM, en Playa, por habernos cedido amablemente el<br />
material objeto de este estudio.<br />
REFERENCIAS<br />
Kensley, B. y M. Schotte 1989. Guide to the Marine isopod Crustaceans<br />
of the Caribbean. Smithsonian Institution Press, 308 pp.<br />
Koening, M. L. 1972. Ocurrencia de Accalathura crenulata<br />
(Richardson, 1901) no Brasil (Isopoda- Paranthuridae).<br />
Trab. Oceanogr. Univ. Fed. Pe., Recife, 13: 261-270.<br />
Menzies, R. J. y P. W. Glynn 1968. The common marine isopod<br />
Crustacea of Puerto Rico. Studies on the fauna of Curacao and<br />
other Caribbean Islands, 27: 1-133.<br />
Müller, H. G. 1991. Marine Anthuridea from Martinique, French<br />
Antilles, with redescriptions of some species (Crustacea:<br />
Isopoda). Revue Suisse Zool., 98: 739-768.<br />
Richardson, H. 1901. Key to the isopods of the Atlantic coast<br />
of North America , with descriptions of new and little<br />
known species. Proc. United States Nat. Mus., 23 :493-579.<br />
Richardson, H. 1905. A Monograph on the isopods of North<br />
America. Bull. United States Nat. Mus., 54: 1-747.<br />
Schultz, G. F. A. 1969. How to know the marine isopod crustaceans.<br />
Dubuque, Iowa, Wm. C. Brown. Co., 359 pp.<br />
Relación taxonómica actualizada de los foraminíferos<br />
modernos de Cuba<br />
María E. Chávez Marrero, Lourdes Rivas Rodríguez y María V.<br />
Orozco Yerena<br />
Instituto de Oceanología, Ave. 1ra, No. 18406, e/ 184 Y 186,<br />
Reparto Flores, Playa 12100, Ciudad de La Habana, Cuba.<br />
costa@oceano.inf.cu<br />
ABSTRACT. Actual taxonomic list of Cuba living foraminifera species<br />
systematically ordered by the Loeblich y Tappan (1988) is presented.<br />
This list comprise 614 species beloging to 207 genera, and reorganized<br />
in 230 supragenerics categries.<br />
Los foraminíferos son un grupo trascendental entre los<br />
protistas ameboideos. Están ampliamente ramificados en el<br />
árbol evolutivo Eukariótico. Su primera aparición fue en el<br />
Cámbrico y durante el curso del Fanerozoico invadieron<br />
completamente los ambientes marinos, con una amplia<br />
diversificación y variedad de modos de vida. Son abundantes y<br />
diversos en el océano actual, donde aparecen en ambientes<br />
costeros y mares profundos, tanto en hábitats planctónicos<br />
como bentónicos (Sen-Gupta, 1999).<br />
Estos protistas son generalmente pequeños y algunos<br />
representantes llegan a tener una proporción gigante con<br />
respecto a los demás, como por ejemplo una especie extinta<br />
(Lepidociclyna elephantica) con 14 cm, siendo una de las más<br />
grandes citadas hasta el momento. Son exclusivamente<br />
unicelulares, y efectúan casi todas las funciones principales de la<br />
vida, que desempeñan un gran número de animales<br />
multicelulares, comen, defecan, se mueven, crecen, se<br />
reproducen y responden a los cambios del medio ambiente.<br />
Existen dos rasgos morfológicos que distinguen a los<br />
foraminíferos de otros protistas. Primero, todos poseen<br />
granoreticulopodia, los cuales son finos, filiformes, y<br />
pseudópodos anastosomados. Segundo, casi todos poseen una<br />
concha que rodea el organismo separándolo del entorno<br />
circundante. Otro aspecto a destacar es que se comportan como<br />
una unidad en el ciclo de la vida, caracterizándose<br />
fundamentalmente por la alternación entre generaciones<br />
sexuales y asexuales.<br />
La clasificación taxonómica de los foraminíferos es detallada<br />
y compleja, y varios autores han hecho propuestas, tanto por la<br />
estructura y forma de la pared como por la más reciente<br />
sisemática moleular. Los foraminíferos fueron mencionados por<br />
primera vez en el siglo V a.n.e., por Herodoto. La primera<br />
clasificación la hizo Alcides Dessalines D'Orbigny en 1826,<br />
considerando los foraminíferos como cefalópodos e incluyó<br />
solo cinco Familias, de las que se describen en la actualidad.<br />
En 1835, Dujardin los reorganiza como protozoos,<br />
identificándolos correctamente como ameboides,<br />
removiéndolos así de los cefalópodos donde los había ubicado<br />
D'Orbigny. En 1872 se realiza un crucero alrededor de algunos<br />
mares del mundo (HMS Challenger), donde se colectan gran<br />
número de foraminíferos que permitirían que Brady en 1884<br />
realizara las “Notas taxonómicas de los los foraminíferos<br />
COCUYO 15, 2005 5
Dragdos en el Challenger”, donde usa casi cuarenta categorías<br />
supragenéricas.<br />
Loeblich y Tappan (1964; 1988) confeccionan el “Tratado<br />
de Paleontología de Invertebrados”, donde ubican a los<br />
foraminíferos dentro del Phylum Protozooa, Orden<br />
Foraminifera. Este Tratado ha sido el más usado por todos los<br />
investigadores y Foraminiferólogos hasta el presente, el mismo<br />
refleja el estado de los foraminíferos en las últimas décadas,<br />
llegando a reorganizar 244 categorías supragenéricas y 3000<br />
géneros. Sen-Gupta (1999) hace una nueva clasificación<br />
sistemática de los foraminíferos Modernos divide, la ahora Clase<br />
Foraminífera, perteneciente al Phylum Granuloreticulosa (Lee et<br />
al., 2000) en cuatro Grupos y 16 Ordenes.<br />
RESULTADOS<br />
Los primeros reportes de foraminíferos de Cuba se hicieron<br />
por Alcides Dessalines D'Orbigny en (1826), en la publicación<br />
Historia Física Política y Natural de la Isla de Cuba, donde<br />
describe 121 especies de foraminíferos recientes de las arenas<br />
cubanas. Posteriormente se hicieron algunos intentos por<br />
publicar un Catálogo de los foraminíferos de Cuba incluyendo<br />
tanto las especies fósiles como las recientes, esta propuesta se<br />
realizó en 1946, pero nunca se llegó a publicar solo se tomaron<br />
hojas sueltas del famoso Catálogo de foraminíferos Elliss y<br />
Messina, y se hizo una recopilación de los que aparecían en Cuba.<br />
Más adelante en 1950, el famoso investigador cubano Pedro J.<br />
Bermúdez, en su publicación Contribución a los foraminíferos<br />
del Cenozoico de Cuba, menciona 1511 especies de<br />
foraminíferos y de ellas 545 especies corresponden a los<br />
foraminíferos Recientes.<br />
Después de las investigaciones de Bermúdez se han hecho<br />
algunos trabajos e investigaciones sobre los foraminíferos que<br />
viven actualmente en nuestros mares pero han sido insuficientes,<br />
debido fundamentalmente a la escasez de especialistas en esta<br />
temática.<br />
En la última década nos hemos dado a la tarea de recopilar la<br />
información existente de los foraminíferos recientes de Cuba y<br />
unido a las nuevas investigaciones que hemos realizado, nos han<br />
permitido reorganizar la sistemática de los foraminíferos citados<br />
para Cuba hasta 2003.<br />
Se realizó una amplia y profunda revisión bibliográfica, sobre<br />
todo lo que ha sido publicado de foraminíferos para el Caribe,<br />
que incluyese a Cuba. Se hizo una exhaustiva revisión y<br />
comprobación de las colecciones de foraminíferos recientes que<br />
existen en Cuba.<br />
Para la clasificación sistemática se adoptó la Clasificación<br />
del Tratado de Paleontología de Invertebrados (Loeblich y<br />
Tappan, 1989), la cual brinda un ordenamiento sistemático, que<br />
tiene en cuenta los antecedentes de varias clasificaciones<br />
taxonómicas, así como aspectos evolutivos de estos organismos.<br />
Como resultado obtuvimos que los foraminíferos recientes<br />
de Cuba estén representados por 614 especies, distribuidas en<br />
207 géneros y 230 categorías supragenéricas. De ellas, 11<br />
aparecen representadas en colecciones. A continuación se<br />
expone la relación sistemática de acuerdo con las fuentes de<br />
información existente, utilizando las siguientes abreviaturas:<br />
Referencias: 1. Zernetzkyi (1975) 2. Bermúdez 1950), 3.<br />
Brodermann (1946), 4. Borro y Alvarez (1979), 5. D' Dorbigny<br />
(1839), 6. Bermúdez P.J (1935), 7. Bandy (1964), 8. Buzas (1982).<br />
Colecciones: O. Colección del Instituto de Oceanología. I.<br />
Colección del Instituto de Geología y Paleontología, ME.<br />
Revisados por María E. Chávez.<br />
Lista de los foraminíferos de Cuba<br />
Orden FORAMINIFERA Eichwald, 1830<br />
Suborden ALLOGROMIINA Loeblich y Tappan, 1961<br />
Familia ALLOGROMIIDAE Rhumbler, 1904<br />
Subfamilia ARGILLOTUBINAE Avnimelech. 1952<br />
Género Argillotuba Avnimelech, 1952<br />
A.vermiformis (Göes) var. occidentalis Zerneztki, 1975 1<br />
Género Nodellum Rumbler, 1913<br />
N. rufescens (Thalman y Bermúdez, 1952) I,3<br />
Suborden TEXTULARIINA Delage y Hérouard, 1896<br />
Superfamilia ASTRORHIZACEA Brady, 1881<br />
Familia BATHYSIPHONIDAE Avnimelech, 1952<br />
Género Bathysiphon M. Sars, 1872<br />
B. filiformis M. Sars, 1872 2<br />
B. rufun Folin, 1886 O, 2, ME<br />
Género Rhabdamminella de Folin, 1887<br />
R. cylindrica (Brady, 1882) 2<br />
Familia RHABDAMMINIDAE Brady, 1884<br />
Subfamilia RHABDAMMININAE Brady, 1884<br />
Género Marisipella Norman, 1878<br />
M. elongata Norman, 1878 2<br />
Género Oculosiphon Avnimelech, 1952<br />
O. linearis (Brady, 1879) O, 3, ME<br />
Género Rhabdammina M. Sars, 1869<br />
R. abyssorum M. Sars, 1869 2<br />
R. discreta Brady, 1881 O, 3, ME<br />
Género Rhizammina Brady, 1879<br />
R. indivisa Brady, 1884 3<br />
Subfamilia DENDROPHRYNAE Haeckel, 1894<br />
Género Psammatodendron Norman, 1881<br />
P. arborecens Norman, 1881 3<br />
Familia PSAMMOSPHAERIDAE Haeckel, 1894<br />
Subfamilia PSAMMOSPHAERINAE Haeckel, 1894<br />
Género Psammosphaera Schultze, 1875<br />
P. fusca Schultze, 1875 3<br />
P. parva Flint, 1899 I, 2<br />
Género Storthosphaera Schulze, 1875<br />
S. albida Schulze, 1875 I, 3<br />
Familia SACCAMMINIDAE Brady, 1884<br />
Subfamilia SACCAMMININAE Brady, 1884<br />
Género Saccammina Carpenter, 1869<br />
S. sphaerica Brady, 1871 3<br />
Familia HEMISPHAERAMMINIDAE Loeblich y Tappan, 1961<br />
Subfamilia HEMISPHAERAMMININAE Loeblich y Tappan,1961<br />
Género Tholosina Rhumbler, 1895<br />
T. vesicularis (Brady) var. erecta Heron Allen y Earland, 1961 2<br />
Superfamilia HIPPOCREPINACEA Rhumbler, 1895<br />
Familia HIPPOCREPINIDAE Rhumbler,1895<br />
Subfamilia HYPERAMMININAE Eimer y Fickert, 1899<br />
Género Hyperammina Brady, 1878<br />
H. elongata Brady, 1878 I, 3<br />
H. friabilis Brady, 1884 I,3<br />
H. laevigata (Wright, 1891) 3<br />
Género Saccorhiza Eimer y Fickert, 1899<br />
S. ramosa (Brady, 1879) 3<br />
Superfamilia AMMODISCACEA Reuss, 1862<br />
Familia AMMODISCIDAE Reuss, 1862<br />
Subfamilia AMMODISCINAE Reuss, 1862<br />
Género Ammodiscoides Cushman, 1909<br />
6 COCUYO 15, 2005
A. turbinatus Cushman, 1909 O, 3, ME<br />
Género Ammodiscus Reuss, 1862<br />
A. anguillae Höglund, 1947 2<br />
A. howelli Acosta, 1940 3<br />
A. incertus (D'Orbigny, 1840) O, 3, ME<br />
A. planorbis Höglund, 1947 2<br />
A. tenuis Brady, 1884 3<br />
Subfamilia TOLYPAMMININAE Cushman, 1928<br />
Género Ammolagena Eimer y Fickert, 1899<br />
A. clavata (Parker y Jones, 1860) I, 2<br />
Género Tolypammina Rhumbler, 1895<br />
T. vagans (Brady, 1879) O,2,ME<br />
Subfamilia AMMOVERTELLININAE Saidova, 1981<br />
Género Glomospira Rzchak, 1885<br />
G. charoides (Parker y Jones, 1860) O, 2, ME<br />
G. gordialis (Jones y Parker, 1860) O, 2, ME<br />
Superfamilia HORMOSINACEA Haeckel, 1894<br />
Familia HORMOSINIDAE Haeckel, 1894<br />
Subfamilia REOPHACINAE Cushman, 1910<br />
Género Hormosinella Shchedrina, 1969<br />
H. distans (Brady, 1881) 4<br />
Género Reophax de Mortfort, 1808<br />
R. arayanensis Bermúdez y Seiglie, 1963 1<br />
R. armatus Göes, 1896 3<br />
R. bacillaris Brady, 1881 I, 3<br />
R. bilocularis Flint, 1899 I, 2<br />
R. dentaliformis Brady, 1881 3<br />
R. guttifer Brady, 1881 4<br />
R. irregularis Parker, 1954 4<br />
R. mariae Acosta, 1940 3<br />
R. procerus Göes, 1894 2<br />
R. scorpiurus Montfort, 1808 O, 2, ME<br />
R. spiculifer Brady, 1879 O, 3, ME<br />
Género Subreophax Saidova, 1975<br />
S. aduncus (Brady, 1882) 3<br />
Subfamilia HORMOSININAE Haeckel, 1894<br />
Género Hormosina Brady, 1879<br />
H. globulifera Brady, 1879 I, 2<br />
H. monile Brady, 1881 2<br />
H. ovicula Brady, 1879 I, 2<br />
Género Pseudonodosinella Saidova, 1970<br />
P. nodulosa (Brady, 1879) 4<br />
Superfamilia LITUOLACEA de Blainville, 1827<br />
Familia HAPLOPHRAGMOIDIDAE Maync, 1952<br />
Género Haplophragmoides Cushman, 1910<br />
H. canariensis (D'Orbigny, 1839) I, 2<br />
H. coronatum (Brady, 1884) 3<br />
H. emaciatum (Brady, 1884) O, 3, ME<br />
H. rigens (Brady, 1879) 3<br />
H. rotulatum (Brady, 1879) 3<br />
H. scitulum (Brady, 1881) O, 2, ME<br />
H. subglobusum Sars, 1910 2<br />
Familia LILUOLIDAE de Blainville, 1827<br />
Subfamilia AMMOMARGINULININAE Podobina, 1978<br />
Género Ammobaculites Cushman, 1910<br />
A. agglutinans Cushman, 1910 O, 7, ME<br />
A. calcareum (Brady, 1884) I, 2<br />
A. cylindricus Cushman, 1910 I, 3<br />
A. exiguus (Acosta, 1940) 2<br />
Género Ammomarginulina Wiesner, 1931<br />
A. foliacea (Brady, 1881) 2<br />
Familia PLASCOPSILINIDAE Rhumbler, 1913<br />
Subfamilia PLACOPSILININAE Rhumbler, 1913<br />
Género Placopsilina D' Orbigny, 1850<br />
P. cenomana D' Orbigny, 1850 2<br />
Superfamilia HAPLOPHRAGMIACEA Eimer y Fickert, 1899<br />
Familia AMMOSPHAEROIDINIDAE Cushman, 1927<br />
Subfamilia AMMOSPHAEROIDININAE Cushman, 1927<br />
Género Ammosphaeroidina Cushman, 1910<br />
Ammosphaeroidina sphaeroidiniformis (Brady, 1884) O, 2, ME<br />
Género Cystammina Neumaya, 1889<br />
C. aguayoi Bermúdez y Acosta, 1940 2<br />
C. pauciloculata (Brady, 1879) O, 3, ME<br />
Subfamilia RECURVOIDINAE Alekseychik Mitskevich, 1973<br />
Género Recurvoides Earland, 1934<br />
R. clenchi Bermúdez, 1939 I, 2<br />
R. obsoletum (Göes, 1896) I, 2<br />
R. turbinatus (Brady, 1881) 3<br />
Género Thalmannammina Pokorny, 1951<br />
T. conglobata (Brady, 1884) 2<br />
Superfamilia COSCINOPHRAGMATACEA Thalmann, 1951<br />
Familia HADDONIIDAE Saidova, 1981<br />
Género Haddonia Chapman, 1898<br />
H. torresiensis Chapman, 1898 3<br />
Familia COSCINOPHRAGMATIDAE Thalmann, 1951<br />
Género Bdelloidina Carter, 1877<br />
B. aggregata Carter, 1877 3<br />
Superfamilia LOFTUSIACEA Brady, 1884<br />
Familia CYCLAMMINIDAE Marie, 1941<br />
Subfamilia CYCLAMMININAE Marie, 1941<br />
Género Cyclammina Brady, 1879<br />
C. brady Cushman, 1910 I, 2<br />
C. cancellata Brady, 1879 I, 2<br />
Superfamilia SPIROPLECTAMMINACEA Cushman, 1927<br />
Familia SPIROPLECTAMMINIDAE Cushman, 1927<br />
Subfamilia SPIROPLECTAMMININAE Cushman, 1927<br />
Género Spiroplectammina Cushman,1927<br />
S. carinata (D'Orbigny, 1846) I, 2<br />
S. gramen (D'Orbigny, 1839) O, 2<br />
S. floridana (Cushman, 1922) 4<br />
Subfamilia VULVULININAE Saidova, 1981<br />
Género Vulvulina D'Orbigny, 1826<br />
V. garciai Zernetzki, 1975 1<br />
V. pennatula (Batsch, 1791) O, 3, ME<br />
Familia NOURIIDAE Chapman y Parr, 1936<br />
Género Nouria Heron Allen y Earland, 1914<br />
N. harrisii Heron Allen y Earland, 1914 2<br />
N. johnsoni Cushman, 1935 3<br />
N. polymorphinoides Heron Allen y Earland, 1914 2<br />
Superfamilia TROCHAMMINACEA Schwager, 1877<br />
Familia TROCHAMMINIDAE Schwager, 1877<br />
Subfamilia TROCHAMMININAE Schwager,1877<br />
Género Tritaxis Schubert, 1921<br />
T. fusca (Williamson, 1858) 8<br />
Género Trochammina Parker y Jones, 1859<br />
T. globigeriniformis (Parker y Jones) O, 2, ME<br />
T. rotaliformis Wright, 1911 3<br />
Familia REMANEICIDAE Loeblich y Tappan, 1964<br />
Subfamilia REMANEICINAE Loeblich y Tappan, 1964<br />
Género Remaneica Rhumbler, 1938<br />
R. kelletae (Cushman y Kellet, 1922) 3<br />
Superfamilia VERNEUILINACEA Cushman, 1911<br />
Familia PROLIXOPLECTIDAE Loeblich y Tappan, 1985<br />
Género Karrerulina Finlay, 1940<br />
K. aplicularis (Cushman, 1911) 3<br />
Familia VERNEUILINIDAE Cushman, 1911<br />
Subfamilia VERNEUILINAE Cushman, 1911<br />
Género Gaudryina D'Orbigny, 1839<br />
G. pseudofiliformis (Berthelin, 1880) O, 8, ME<br />
G. flintii (Cushman, 1911) O, 2, ME<br />
G. quadrangularis Bagg var. antillea Bermúdez y Acosta, 1940 O, 3,<br />
ME<br />
Subfamilia BARBOURINELLINAE Saidova, 1981<br />
Género Barbourinella Bermúdez, 1940<br />
B. atlantica (Bermúdez, 1939) O, 3, ME<br />
Superfamilia ATAXOPHRAGMIACEA Schwager, 1877<br />
Familia GLOBOTEXTULARIIDAE Cushman, 1927<br />
Subfamilia GLOBOTEXTULARIINAE Cushman, 1927<br />
Género Globotextularia Eimer y Fickert, 1899<br />
G. anceps (Brady, 1884) 3<br />
COCUYO 15, 2005 7
Subfamilia LIEBUSELLINAE Saidova, 1981<br />
Género Liebusella Cushman, 1933<br />
L. intermedia Vander Broeck, 1876 2<br />
L. soldanii (Parker y Jones, 1860) 1<br />
Familia TEXTURARIELLIDAE Grönhagen y Luterbacher, 1966<br />
Género Textulariella Cushman, 1927<br />
T. barrettii (Jones y Parker, 1876) I, 2<br />
Superfamilia TEXTULARIACEA Ehrenberg, 1838<br />
Familia EGGERELLIDAE Cushman, 1937<br />
Subfamilia DOROTHIINAE Balakhmatova, 1972<br />
Género Dorothia Plummer, 1931<br />
D. bradyi Cushman, 1936 3<br />
D. caribaea Cushman, 1936 O, 2, ME<br />
D. curta (Cushman, 1922) 2<br />
D. exilis Cushman, 1936 3<br />
D. pseudoturris (Cushman, 1922) O, 3, ME<br />
D. scabra (Brady, 1884) O, 2, ME<br />
Subfamilia EGGERELLINAE Cushman, 1937<br />
Género Eggerella Cushman, 1935<br />
E. propincua (Brady, 1884) O, 2, ME<br />
Género Karreriella Cushman, 1933<br />
K. bradyi Cushman, 1911 O, 2, ME<br />
K. hovangliae (Cushman, 1922) 3<br />
K. caribaea (Acosta, 1940) I, 2<br />
Género Martinottiella Cushman, 1933<br />
M. antillarum (Cushman, 1936) 3<br />
M. nodulosa (Cushman, 1935) 2<br />
M. occidentalis (Cushman, 1935) 2<br />
Familia TEXTULARIIDAE Ehrenberg, 1838<br />
Subfamilia TEXTULARIINAE Ehrenberg, 1838<br />
Género Bigenerina D'Orbigny, 1826<br />
B. irregularis Phleger y Parker, 1951 O, 8, ME<br />
B. lytta Lalicker y Bermúdez , 1941 I, 2<br />
B. nodosaria D'Orbigny, 1826 1<br />
B. textularoidea (Göes, 1894) I, 2<br />
B. perna Lalicker y Bermúdez, 1941 2<br />
Género Textularia Defrance, 1824<br />
T. bermudesi Cushman y Todd 3<br />
T. calva Lalicker, 1935 3<br />
T. candeiana D' Orbigny, 1839 O, 2, ME<br />
T. caribaea D'Orbigny, 1839 3<br />
T. catenata Cushman, 1911 3<br />
T. conica D'Orbigny, 1839 3<br />
T. concava (Karrer, 1868) 3<br />
T. heterostoma (Fornasini, 1896) 6<br />
T. corrugata (Heron - Allen y Earland, 1915) I, 2<br />
T. flintii Cushman, 1911 2<br />
T. flintii Cushman var. curta Cushman, 1922 3<br />
T. lateralis Lalicker, 1935 I, 2<br />
T. luculenta Brady,1884 2<br />
T. lythostrota (Schwager, 1866) O, 3, ME<br />
T. mexicana Cushman, 1922 O, 3, ME<br />
T. mexicana var. cubana Palmer y Bermúdez, 1935 2<br />
T. saulcyana D'Orbigny, 1839 5<br />
T. schencki Cushman y Valentine, 1930 2<br />
T. sicca Lalicker y Bermúdez, 1941 2<br />
T. subplana Cushman, 1922 3<br />
T. wiesneri Earland, 1933 3<br />
Subfamilia SIPHOTEXTULARIINAE Loeblich y Tappan, 1985<br />
Género Karrerotextularia Y. Le Calvez, de Klasz y Brun, 1974<br />
K. albatrossi (Cushman, 1922) 3<br />
Familia PSEUDOGAUDRYNIDAE Loeblich y Tappan, 1985<br />
Subfamilia PSEUDOGAUDRYNINAE Loeblich y Tappan, 1985<br />
Género Pseudogaudryina Cushman, 1936<br />
P. atlantica (Bailey, 1851) 2<br />
Familia VALVULINIDAE Berthelin, 1880<br />
Subfamilia VALVULININAE Berthelin, 1880<br />
Género Clavulina D'Orbigny, 1826<br />
C. angularis D'Orbigny, 1826 3<br />
C. mexicana (Cushman, 1922) O, 3, ME<br />
C. nodosaria D'Orbigny, 1839 O, 2, ME<br />
C. tricarinata D'Orbigny, 1839 I, 1<br />
Género Cribrogoesella Cushman, 1935<br />
C. robusta (Brady, 1881) 3<br />
Género Cylindroclavulina Bermúdez y Key, 1952<br />
C. bradyi (Cushman, 1911) 4<br />
Género Goessella Cushman,1933<br />
G. flintiana (Cushman, 1936) 3<br />
Género Valvulina D'Orbigny, 1826<br />
V. fusca (Williamson, 1858) I, 2<br />
V. oviedoiana D'Orbigny, 1839 3<br />
Subfamilia TRITAXILININAE Loeblich y Tappan, 1986<br />
Género Tritaxilina Cushman, 1911<br />
T. atlantica Cushman, 1922 I, 1<br />
Familia GLAUCOAMMINIDAE Saidova, 1981<br />
Género Glaucoammina Seiglie y Bermúdez, 1969<br />
G. trilateralis (Cushman, 1935) 2<br />
Suborden SPIRILLININA Hohenegger y Piller, 1975<br />
Familia SPIRILLINIDAE Reuss y Fritsch, 1861<br />
Género Spirillina Ehrenberg, 1843<br />
S. decorata Brady, 1884 6<br />
S. denticulata Brady, 1884 2<br />
S. luciada Sidebottom, 1908 6<br />
S. ornata Sidebottom, 1908 6<br />
S. tuberculolimbata Chapman, 1899 6<br />
S. vivipara Ehrenberg, 1840 6<br />
Familia PATELLINIDAE Rhumbler, 1906<br />
Subfamilia PATELLININAE Rumbler, 1906<br />
Género Patellina Williamson, 1858<br />
P. corrugata Williamson, 1858 3<br />
Suborden CARTERININA Loeblich y Tappan, 1981<br />
Familia CARTERINIDAE Loeblich y Tappan, 1955<br />
Género Carterina Brady, 1884<br />
C. spiculotesta (Carter, 1877) I, 2<br />
Suborden MILIOLINA Delage y Hérouard, 1896<br />
Superfamilia CORNUSPIRACEA Schultze, 1854<br />
Familia CORNUSPIRIDAE Schultze, 1854<br />
Subfamilia CORNUSPIRINAE Schultze, 1854<br />
Género Cornuspira Schultze, 1854<br />
C. crassisepta (Brady, 1882) 2<br />
C. foliacea (Philippi, 1844) O, 2, ME<br />
C. involvens (Reuss, 1850) 2<br />
C. planorbis (Shultze, 1854) 1<br />
C. selseyensis (Heron - Allen y Earland, 1905) 2<br />
Familia FISCHERINIDAE Millett, 1898<br />
Subfamilia FISCHERINELLINAE Saidova, 1981<br />
Género Fischerinella Loeblich y Tappan, 1962<br />
F. dubia (D'Orbigny, 1839) 5<br />
Subfamilia NODOBACULARIELLINAE Bogdanovich, 1981<br />
Género Nodobaculariella Cushman y Hanzawa, 1937<br />
N. cassis (D'Orbigny, 1839) O, 3, ME<br />
Género Wiesnerella Cushman, 1933<br />
W. auriculata (Eggel, 1893) O, 2, ME<br />
Familia NUBECULARIIDAE Jones, 1875<br />
Subfamilia NUBECULARIINAE Jones, 1875<br />
Género Nubecularia Defrance, 1825<br />
N. lucifuga Defrance, 1825 2<br />
Subfamilia OPHTHALMIDIIDAE Wiesner, 1920<br />
Género Cornuloculina Burbach, 1886<br />
C. inconstans (Brady, 1879) O, 2, ME<br />
Superfamilia MILIOLACEA Ehrenberg, 1839<br />
Familia SPIROLOCULINIDAE Wiesner, 1920<br />
Género Flintia Schubert, 1911<br />
F. robusta (Brady, 1884) I, 3<br />
Género Planispirinoides Parr, 1950<br />
P. bucculenta (Brady, 1884) 2<br />
Género Spiroloculina D'Orbigny, 1826<br />
S. acicularis (Batsch, 1791) I, 1<br />
S. antillarum D'Orbigny, 1839 2<br />
S. asperula Karrer, 1868 1<br />
8 COCUYO 15, 2005
S. arenata Cushman, 1921 I, 3<br />
S. depressa D'Orbigny, 1826 2<br />
S. ornata D'Orbigny, 1839 2<br />
S. poeyana D'Orbigny, 1839 2<br />
S. robusta Brady, 1884 I, 3<br />
S. vivipara (Ehrenberg, 1843) 1<br />
Familia HAUERINIDAE Schwager, 1876<br />
Subfamilia SIPHONAPERTINAE Saidova, 1975<br />
Género Dentostomina Carman, 1933<br />
D. aguayoi Pérez Frafante, 1939 3<br />
D. bermudiana Carman, 1933 3<br />
D. bermudiana Carman, var. trilongides Pérez, 1939 2<br />
Género Schlumbergerina Munier Chalmas, 1882<br />
S. alveoliniformis var. occidentalis (Brady, 1879) O, 2, ME<br />
Subfamilia HAUERININAE Schwager, 1876<br />
Género Hauerina D'Orbigny, 1839<br />
H. howelli Bermúdez, 1935 2<br />
Género Massilina Schlumberger, 1893<br />
M. crenata (Karrer, 1868) O, 6, ME<br />
M. inaequalis Cushman, 1921 8<br />
M. secans (D'Orbigny, 1826) 6<br />
Género Quinqueloculina D'Orbigny, 1826<br />
Q. ackneriana D'Orbigny, 1846 2<br />
Q. agglutinans D'Orbigny, 1839 O, 5, ME<br />
Q. auberiana D'Orbigny, 1839 3<br />
Q. antillarum D'Orbigny, 1839 3<br />
Q. barbouri Bermúdez, 1935 3<br />
Q. bicostata D'Orbigny, 1839 1<br />
Q. bidentata D'Orbigny, 1939 3<br />
Q. bosciana D'Orbigny, 1839 O, 5, ME<br />
Q. bradyiana Cushman, 1917 2<br />
Q. candeiana D'Orbigny, 1839 O, 2, ME<br />
Q. columnosa Cushman, 1922 2<br />
Q. cuvieriana D'Orbigny, 1839 2<br />
Q. enoplostoma D'Orbigny, 1839 5<br />
Q. funafutienses (Chapman, 1902) 2<br />
Q. kerimbatica var. philipinensis Cushman, 1921 3<br />
Q. laevigata D'Orbigny, 1839 1<br />
Q. lamarckiana D'Orbigny, 1839 O, 5, ME<br />
Q. linneiana D'Orbigny, 1839 1<br />
Q. parkeri (Brady) var. occidentalis Cushman, 1921 3<br />
Q. poeyana D'Orbigny, 1839 O, 5, ME<br />
Q. polygona D'Orbigny, 1839 O, 5, ME<br />
Q. planciana D'Orbigny, 1840 5<br />
Q. pulchella D'Orbigny , 1839 O, 2, ME<br />
Q. qualteriana D'Orbigny, 1839 5<br />
Q. reticulata (D'Orbigny, 1826) 3<br />
Q. riveroae Bermúdez y Seiglie, 1963 O, 4, ME<br />
Q. sagra D'Orbigny, 1839 5<br />
Q. semicancellata Zernietkii, 1975 1<br />
Q. seminula (Linné, 1767) O, 2, ME<br />
Q. subarenarea Cushman, 1917 2<br />
Q. subpoeyana Cushman, 1922 2<br />
Q. tricarinata D'Orbigny, 1839 O, 5, ME<br />
Q. torrei Acosta, 1939 5<br />
Subfamilia MILIOLINELLINAE Vella, 1957<br />
Género Affinetrina Ruczkowska, 1972<br />
A. planciana (D'Orbigny, 1839) 5<br />
Género Biloculinella Wiesner, 1931<br />
B. eburnea (D'Orbigny, 1839) 2<br />
Género Cribopyrgo Cushman y Bermúdez, 1946<br />
C. robusta Cushman y Bermúdez, 1946 3<br />
Género Cruciloculina D'Orbigny, 1839<br />
C. triangularis D'Orbigny, 1839 2<br />
Género Flintina Cushman, 1921<br />
F. clenchi Bermúdez, 1939 2<br />
Género Miliolinella Wiesner, 1931<br />
M. circularis Bornenman, 1855 4<br />
M. fichteliana (D'Orbigny, 1839) O, 1, ME<br />
M. labiosa (D'Orbigny, 1839) O, 1, ME<br />
M. suborbicularis Hofker 1<br />
M. subrotunda (Montagu, 1803) 1<br />
Género Neopateoris Bermúdez y Seiglie, 1963<br />
N. cumanaensis Bermúdez y Seiglie, 1963 1<br />
Género Pyrgo Defrance, 1824<br />
P. bradyi (Schlumberger) 2<br />
P. bulloides D'Orbigny, 1826 O, 2, ME<br />
P. carinata D'Orbigny, 1826 5<br />
P. comata (Brady, 1884) I, 2<br />
P. denticulata (Brady, 1884) O, 2, ME<br />
P. depressa (D'Orbigny, 1826) 2<br />
P. elongata D'Orbigny, 1826 2<br />
P .jonsoni Cushman, 1935 3<br />
P. murrhina (Schwager, 1866) 8<br />
P. saccata (Göes, 1894) 2<br />
P. striolata (Brady, 1884) I, 2<br />
P. subphaericus (D'Orbigny, 1839) O, 3, ME<br />
P. oblonga (D'Orbigny, 1839) I, 3<br />
Género Triloculina D'Orbigny, 1826<br />
T. bermudezi Acosta, 1940 2<br />
T. bertheliana (Brady, 1884) 3<br />
T. bicarinata D'Orbigny, 1839 O, 3, ME<br />
T. brongniartiana D'Orbigny, 1826 O, 5, ME<br />
T. carinata D'Orbigny, 1839 I, 2<br />
T. fichteliana D'Orbigny, 1826 5<br />
T. fiterrei Acosta, 1940 3<br />
T. fiterrei var meningoi Acosta, 1940 3<br />
T. gualteriana D'Orbigny, 1839 5<br />
T. gracilis D'Orbigny, 1839 8<br />
T. insignis (Brady, 1884) 2<br />
T. laevigata D'Orbigny, 1826 4<br />
T. linneiana D'Orbigny, 1839 5<br />
T. oblonga (Montagu, 1803) 3<br />
T. quadrilateralis D'Orbigny, 1839 5<br />
T. rotunda D'Orbigny, 1893 3<br />
T. sidebottomi (Mortinotti, 1920 ) 1<br />
T. suborbicularis D'Orbigny, 1839 5<br />
T. schrebeiriana D'Orbigny, 1839 5<br />
T. transversestriata (Brady, 1881 O, 3, ME<br />
T. tricarinata D'Orbigny, 1826 O, 3, ME<br />
T. trigonula (Lamarck, 1804) O, 2, ME<br />
T. viguerasi Bermúdez, 1935 3<br />
Subfamilia SIGMOILINITINAE Luczkowska, 1974<br />
Género Nummoloculina Steinmann, 1881<br />
N. contraria (D'Orbigny, 1846) I, 2<br />
N. irregularis (D'Orbigny, 1839) 2<br />
Género Sigmoihauerina S. Y. Zheng, 1979<br />
S. bradyi (Cushman, 1917) O, 2, ME<br />
Género Sigmoilina Schlumberger, 1887<br />
S. arenata (Cushman, 1921) 2<br />
S. celata (Costa) 1<br />
S. costata Silvestri, 1904 2<br />
S. juareci Zernietkii, 1975 1<br />
S. schlumbergeri Silvestri, 1904 O, 3, ME<br />
S. sigmoidea (Brady, 1884) O, 2, ME<br />
S. tenuis Czjzck, 1848 2<br />
Subfamilia TUBINELLINAE Rhumbler, 1906<br />
Género Articulina D'Orbigny, 1826<br />
A. antillarum Cushman, 1922 I, 3<br />
A. lineata Brady, 1884 3<br />
A. mayori Cushman, 1922 3<br />
A. mexicana Cushman, 1922 3<br />
A. mucronata (D'Orbigny, 1839) 2<br />
A. sagra D'Orbigny, 1839 O, 5, ME<br />
Género Parrina Cushman, 1931<br />
P. zonacenzis Zernetzkii, 1975 1<br />
Género Tubinella Rhumbler, 1906<br />
T. funalis (Brady, 1884) O, 2, ME<br />
Familia RIVEROINIDAE Saidova, 1981<br />
Género Pseudohauerina Ponder, 1972<br />
COCUYO 15, 2005 9
P. occidentalis (Cushman, 1946) O, 2, ME<br />
Género Riveroina Bermúdez, 1939<br />
R. caribaea Bermúdez, 1939 3<br />
Superfamilia ALVEOLINACEA Ehrenberg, 1839<br />
Familia RHAPYDIONINIDAE Keijzer, 1945<br />
Subfamilia RHAPYDIONININAE Keijzer, 1945<br />
Género Ripacubana Loeblich y Tappan, 1964<br />
R. conica (D'Orbigny, 1839) 5<br />
Familia ALVEOLINIDAE Ehrenberg, 1839<br />
Género Borelis de Montfort, 1808<br />
B. pulchra (D'Orbigny, 1839) O, 3, ME<br />
Familia PENEROPLIDAE Schultze, 1854<br />
Género Dendritina D'Orbigny, 1826<br />
D. antillarum D'Orbigny, 1826 2<br />
D. elegans (D'Orbigny, 1846) 4<br />
Género Laevipeneroplis Sulc, 1936<br />
L. bradyi Cushman, 1930 O, 3, ME<br />
L. carinatus D'Orbigny, 1839 3<br />
L. proteus (D'Orbigny, 1839) O, 5, ME<br />
Género Monalysidium Chapman, 1900<br />
M. politum (Chapman, 1902) O, 3, ME<br />
Género Peneroplis Montfort, 1808<br />
P. antillarum D'Orbigny, 1839 5<br />
P. elegans D'Orbigny, 1839 O, 5, ME<br />
P. pertusus Forskal, 1775 1<br />
Género Spirolina Lamarck, 1804<br />
S. acicularis (Batsch, 1791) O, 3, ME<br />
Familia SORITIDAE Ehrenberg, 1839<br />
Subfamilia ARCHAIASINAE Cushman, 1927<br />
Género Archaias de Montfort, 1808<br />
A. angulatus (Fichtel y Moll, 1803) O, 3, ME<br />
Género Cycloputeolina Seiglie y Grove, 1977<br />
C. bocki Seiglie y Grove, 1977 2<br />
Género Cyclorbiculina A. Silvestri, 1937<br />
C. compressa (D'Orbigny, 1839) 2<br />
Subfamilia SORITINAE Ehrenberg, 1839<br />
Género Amphisorus Ehrenberg, 1839<br />
A. hemprichii Ehrenberg, 1839 O, 3, ME<br />
Género Sorites Ehrenberg, 1839<br />
S. marginalis (Lamarck, 1816) I,3<br />
Suborden LAGENINA Delage y Hérouard, 1896<br />
Superfamilia NODOSARIACEA Ehrenberg, 1838<br />
Familia NODOSARIIDAE Ehrenberg, 1838<br />
Subfamilia NODOSARIINAE Ehrenberg, 1838<br />
Género Chrysalogonium Schubert, 1908<br />
C. asperum Cushman y Stainforth,1945 4<br />
C. brodermanni Bermúdez y Acosta, 1940 2<br />
C. ritae Acosta, 1940 3<br />
Género Dentalina Risso, 1826<br />
D. comunnis (D'Orbigny, 1826) I, 2<br />
D. farcimen Reuss, 1863 I, 2<br />
D. flintii Cushman, 1923 I, 8<br />
D. pauperata D'Orbigny, 1846 2<br />
D. subsoluta (Reuss, 1851) I, 2<br />
Género Grigelis Mikhalevich, 1981<br />
Grigelis pyrula (D'Orbigny, 1826) I, 2<br />
Género Nodosaria Lamarck, 1812<br />
N. advena Cushman, 1923 3<br />
N. albatrossi Cushman, 1923 I, 2<br />
N. candei D'Orbigny, 1839 5<br />
N. catesbyi D'Orbigny, 1840 O, 2, ME<br />
N. comatula Cushman, 1923 3<br />
N. fistuca (Schwager, 1866) 2<br />
N. japonica Cushman, 1913 3<br />
N. longiscata D'Orbigny, 1846 2<br />
N. millettii Cushman, 1917 2<br />
N. obliquus (Linneus, 1752) 2<br />
N. perversa Schwager 3<br />
N. rapharistrum (Linné, 1757) 2<br />
N. rugosa D'Orbigny, 1840 2<br />
Subfamilia LINGULININAE Loeblich y Tappan, 1961<br />
Género Lingulina D'Orbigny, 1826<br />
L. carinata D'Orbigny, 1826 5<br />
L. seminuda (Hatken, 1881) O, 3, ME<br />
Subfamilia FRONDICULARIINAE Reuss, 1860<br />
Género Frondicularia Defrance, 1826<br />
F. saggitula Marrie, 1941 I, 2<br />
F. saggitula var lanceolata (Brook) 2<br />
Subfamilia PLECTOFRONDICULARIINAE Cushman, 1927<br />
Género Proxifrons Vella, 1963<br />
P. advena (Cushman, 1923) 2<br />
Familia VAGINULINIDAE Reuss, 1860<br />
Subfamilia LENTICULININAE Chapman, Parr y Collins, 1934<br />
Género Lenticulina Lamarck, 1804<br />
L. calcar (Linné, 1767) 2<br />
L. formosa (Cushman, 1923) I, 2<br />
L. gibba (D'Orbigny, 1839) I, 2<br />
L. iota (Cushman) I, 3<br />
L. limbosa (Reuss) 2<br />
L. peregrina (Schwager, 1866) I, 2<br />
L. rotulata Lamarck , 1804 2<br />
L. sublituus D'Orbigny, 1826 3<br />
L. subaculeata var. glabrata (Cushman, 1923) I, 3<br />
Género Saracenaria Defrance, 1824<br />
S. acutiauricularis (Fitchtel y Moll, 1798) 2<br />
S. italica Defrance, 1824 I, 2<br />
S. latifronts (Brady, 1884) O, 2, ME<br />
Subfamilia MARGINULININAE Wedekind, 1937<br />
Género Marginulina D'Orbigny, 1826<br />
M. bachei Bailey, 1851 2<br />
M. crepidula Fitchel y Moll, 1798 2<br />
M. hirsuta D'Orbigny, 1826 3<br />
Subfamilia VAGINULININAE Reuss, 1860<br />
Género Vaginulina D'Orbigny, 1826<br />
V. americana Cushman, 1923 3<br />
V. elegans D'Orbigny, 1826 2<br />
V. legumen (Linné, 1758) 2<br />
V. legumen var. arquata Brady, 1884 2<br />
V. punctata D'Orbigny, 1839 3<br />
V. spinigera Brady, 1881 I, 3<br />
Familia LAGENIDAE Reuss, 1862<br />
Género Lagena Walker y Jacob, 1798<br />
L. ampulladistoma (Rymer y Jones, 1872) 2<br />
L. auriculata Brady, 1881 2<br />
L. costata (Williamson, 1913) 2<br />
L. desmosphora (Rymer y Jones) 3<br />
L. distoma Parker y Jones, 1864 2<br />
L. elongata Ehrenberg, 1844 2<br />
L. fasciata var. spinosa Sidebottom, 1912 2<br />
L. fimbriata Brady, 1881 3<br />
L. flintiana Cushman, 1923 I, 2<br />
L. quadralata Brady, 1881 3<br />
L. laevis var. nebulosa Cushman, 1923 2<br />
L. perlucida (Monyagu, 1883) 2<br />
L. plumigera Brady, 1881 3<br />
L. striata (D'Orbigny, 1839) 2<br />
L. substriata Williamsom, 1848 2<br />
L. sulcata (Walker y Jacob, 1798) 2<br />
L. sulcata var. striatupunctata Parker y Jones, 1865 3<br />
L. sublagenoides Cushman, 1913 2<br />
Género Procerolagena Puri, 1954<br />
P. gracilis (Williamson, 1848) 2<br />
Familia POLYMORPHINIDAE D'Orbigny, 1839<br />
Subfamilia POLYMORPHININAE D'Orbigny, 1839<br />
Género Globulina D'Orbigny, 1839<br />
G. gibba D'Orbigny, 1826 2<br />
G. inaequalis Reuss, 1850 3<br />
G. inaequalis var. caribaea Cushman y Ozawa, 1930 2<br />
Género Guttulina D'Orbigny, 1839<br />
G. caribaea D'Orbigny, 1839 5<br />
10 COCUYO 15, 2005
G. problema D'Orbigny, 1826 2<br />
G. pulchella D'Orbigny, 1839 3<br />
G. regina (Brady, Parker y Jones, 1870) I, 3<br />
G. spicaeformis var. australis (D'Orbigny) I, 2<br />
G. vitrea D'Orbigny, 1839 3<br />
Género Polymorphina D'Orbigny, 1826<br />
P. irregularis D'Orbigny, 1839 5<br />
P. rochefortiana D'Orbigny, 1839 5<br />
P. rugosa D'Orbigny, 1839 5<br />
Género Pyrulina D'Orbigny, 1839<br />
P. extensa (Cushman) I, 3<br />
P. gutta D'Orbigny, 1826 3<br />
Género Sigmoidella Cushman y Ozawa, 1928<br />
S. elegantissima (Parker y Jones) 2<br />
Género Sigmomorphina Cushman y Ozawa, 1928<br />
S. pearceyi Cushman y Ozawa, 1930 3<br />
Subfamilia RAMULININAE Brady, 1884<br />
Género Ramulina Jones y Wright, 1875<br />
R. globulifera Brady, 1879 O, 2, ME<br />
R. proteiformis Cushman, 1899 2<br />
Familia ELLIPSOLAGENIDAE A. Silvestri, 1923<br />
Subfamilia OOLININAE Loeblich y Tappan, 1961<br />
Género Oolina D'Orbigny, 1839<br />
O. botelliformis (Brady, 1884) 2<br />
O. botelliformis var. rugosa (Sidebottom,1912) 2<br />
Subfamilia ELLIPSOLAGENINAE A. Silvestre, 1923<br />
Género Fissurina Reuss, 1850<br />
F. lucida (Williamson, 1848) 2<br />
F. marginata (Walter y Boys, 1784) I, 2<br />
F. marginoperforata Sequenza, 1880 2<br />
F. orbignyana (Cushman, 1923) I, 2<br />
Familia GLANDULINIDAE Reuss, 1860<br />
Subfamilia GLANDULININAE Reuss, 1860<br />
Género Glandulina D'Orbigny, 1839<br />
G. laevigata var. occidentalis Cushman, 1923 I, 2<br />
G. laevigata var. torrida Cushman, 1929 2<br />
G. radicula (Linné) 3<br />
Suborden ROBERTININA Loeblich y Tappan, 1984<br />
Superfamilia CERATOBULIMINACEA Cushman, 1927<br />
Familia CERATOBULIMINIDAE Cushman, 1927<br />
Subfamilia CERATOBULIMINIDAE Cushman, 1927<br />
Género Ceratobulimina Tonla, 1915<br />
C. pacifica Cushman y Harris, 1927 2<br />
Género Lamarckina Berthelin, 1881<br />
L. atlantica Cushman, 1931 I, 2<br />
L. scabra (Brady, 1884) 2<br />
Superfamilia ROBERTINACEA Reuss, 1850<br />
Familia ROBERTINIDAE Reuss, 1850<br />
Subfamilia ALLIATININAE McGrowran, 1966<br />
Género Cushmanella Palmer y Bermúdez, 1936<br />
C. brownii (D'Orbigny, 1839) 3<br />
Subfamilia ROBERTININAE Reuss, 1850<br />
Género Robertina D'Orbigny, 1846<br />
Robertina sp. 2<br />
Suborden GLOBIGERININA Delage y Hérouard, 1896<br />
Superfamilia GLOBOROTALIACEA Cushman, 1927<br />
Familia GLOBOROTALIIDAE Cushman, 1927<br />
Género Globorotalia Cushman, 1927<br />
G. hirsuta (D'Orbigny, 1839) I, 2<br />
G. fimbriata (Brady, 1884) O, 2, ME<br />
G. menardii D'Orbigny, 1884<br />
G. oceanica Cushman y Bermúdez, 1923 2<br />
G. praemenardii Cushman y Stanfort, 1945 2<br />
G. punctulata (D'Orbigny, 1839) 2<br />
G. truncatulinoides (D'Orbigny, 1839) O, 2, ME<br />
G. tumida (Brady) O, 2, ME<br />
Familia PULLENIATINIDAE Cushman, 1927<br />
Género Pulleniatina Cushman, 1927<br />
P. obliqueloculata (Parker y Jones, 1865) O, 2, ME<br />
Familia CANDEINIDAE Cushman, 1927<br />
Subfamilia CANDEININAE Cushman, 1927<br />
Género Candeina D'Orbigny, 1839<br />
C. nitida D'Orbigny, 1839 O, 5, ME<br />
Superfamilia GLOBIGERINACEA Carpenter, Parker y Jones, 1862<br />
Familia GLOBIGERINIDAE Carpenter, Parker y Jones, 1862<br />
Subfamilia GLOBIGERININAE Carpenter, Parker y Jones, 1862<br />
Género Globigerina D'Orbigny, 1826<br />
G. bulloides D'Orbigny, 1826 O, 2, ME<br />
G. conglobata Brady, 1879 2<br />
G. conglomerata Schwager, 1866 3<br />
G. cretacea D'Orbigny, 1840 6<br />
G. dutertrei D'Orbigny, 1839 5<br />
G. eggeri Rhumbler, 1901 O, 2, ME<br />
G. siphoniofera D'Orbigny, 1839 5<br />
Género Globigerinoides Cushman, 1927<br />
G. rubra (D'Orbigny, 1839) I, 3<br />
G. sacculifera (Brady, 1884) 3<br />
Género Sphaerodinella Cushman, 1927<br />
S. bulloides D'Orbigny, 1826 3<br />
S. dehiscens (Parker y Jones, 1865) O, 2, ME<br />
Subfamilia ORBULININAE Schultze, 1854<br />
Género Orbulina D'Orbigny, 1839<br />
O. bilobata (D'Orbigny, 1846) O, 2, ME<br />
O. universa D'Orbigny, 1839 3<br />
Familia HASTIGERINIDAE Bolli, Loeblich y Tappan, 1957<br />
Género Hastigerina Thomson, 1876<br />
H. aequilateralis ( Brady, 1879) O, 2, ME<br />
H. pelagica (D'Orbigny, 1839) 2<br />
Suborden ROTALIINA Delage yHérovard, 1896<br />
Superfamilia BOLIVINACEA Glaessner, 1937<br />
Familia BOLIVINIDAE Glaessner, 1937<br />
Género Bolivina D'Orbigny, 1839<br />
B. alata (Sequenza, 1862) 2<br />
B. canimarensis Palmer y Bermúdez, 1936 2<br />
B. compacta (Sidebothom, 1905) 2<br />
B. cubana Bermúdez, 1935 3<br />
B. goesi Cushman, 1922 O, 2, ME<br />
B. gramen (D'Orbigny, 1840) 3<br />
B. imbricata Cushman, 1925 3<br />
B. lowmani Phleger y Parker, 1951 1<br />
B. plicatella Cushman y Ponton 8<br />
B. prorrectum (Brady) 2<br />
B. pulchella (D'Orbigny, 1839) O, 3, ME<br />
B. punctata D'Orbigny, 1839 3<br />
B. spinecens Cushman, 1911 2<br />
B. striatula Cushman, 1922 I, 3<br />
B. subspinenscens (Reuss, 1862) 2<br />
B. tortuosa Brady, 1881 3<br />
B. variabilis Williamson, 1856 3<br />
Superfamilia LOXOSTOMATACEA Loeblic y Tappan, 1962<br />
Familia LOXOSTOMATIDAE Loeblich y Tappan, 1962<br />
Género Loxostomun Ehrenberg, 1854<br />
L. limbatum var. costulatum (Cushman, 1922) 2<br />
Familia BOLIVINELLIDAE Hayward, 1980<br />
Género Bolivinella Cushman, 1927<br />
B. eleggans Parker, 1932 3<br />
Superfamilia BOLIVINITACEA Cushman, 1927<br />
Familia BOLIVINITIDAE Cushman, 1927<br />
Género Bolivinita Cushman, 1927<br />
B. quadrilatera (Schwager, 1866) 2<br />
B. rhomboidalis (Milleti) 2<br />
Superfamilia CASSIDULINACEA D'Orbigny, 1839<br />
Familia CASSIDULINIDAE D'Orbigny, 1839<br />
Subfamilia CASSIDULININAE D'Orbigny, 1839<br />
Género Cassidulina D'Orbigny, 1826<br />
C. braziliensis Cushman, 1922 3<br />
C. crassa D'Orbigny, 1826 2<br />
C. palmerae Bermúdez y Acosta, 1940 O, 3, ME<br />
C. subglobosa Bandy, 1881 O, 3, ME<br />
Género Cassidulinoides Cushman, 1927<br />
COCUYO 15, 2005 11
C. bradyi (Norman, 1880) 3<br />
C. brazilensis (Cushman, 1922) 2<br />
Superfamilia BULIMINACEA Jones,1875<br />
Familia SIPHOGENERINOIDIDAE Saidova, 1981<br />
Subfamilia SIPHOGENERINOIDINAE Saidova, 1981<br />
Género Rectobolivina Cushman, 1927<br />
R. advena (Cushman, 1922) 2<br />
R. dimorfa (Parker y Jones, 1865) 2<br />
R. limbata 8<br />
R. mayoris (Cushman, 1922) 2<br />
R. nibilis (Hantken, 1876) I, 2<br />
R. raphana (Parker y Jones, 1865) I, 2<br />
Subfamilia TUBULOGENERININAE Saidova, 1981<br />
Género Sagrina D'Orbigny, 1839<br />
S. pulchella (D'Orbigny, 1839) 5<br />
Familia BULIMINIDAE Jones, 1875<br />
Género Bulimina D'Orbigny, 1826<br />
B. affinis D'Orbigny, 1839 5<br />
B. buchiana D'Orbigny, 1845 2<br />
B. elegans D'Orbigny, 1826 2<br />
B. inflata mexicana (Sequenza, 1862) 2<br />
B. ovata Cushman, 1922 8<br />
B. ovula D'Orbigny, 1839 2<br />
B. Pupoides Cushman, 1922 8<br />
B. pyrula var. spirecens Brady, 1884 3<br />
Familia BULIMINELLIDAE Hofker, 1951<br />
Género Buliminella Cushman, 1911<br />
B. elegantissima var. seminuda Brady, 1884 2<br />
B. elegantissima (D'Orbigny, 1839) O, 6, ME<br />
B. parallela Cushman y Parker, 1931 6<br />
B. silviae Bermúdez y Seiglie, 1963 3<br />
Familia UVIGERINIDAE Haeckel, 1894<br />
Subfamilia UVIGERININAE Haeckel, 1894<br />
Género Siphouvigerina Parr, 1950<br />
S. ampullacea (Brady, 1824) 2<br />
S. fimbriata (Sidebottom, 1918) 6<br />
Género Uvigerina D'Orbigny, 1826<br />
U. asperula Czjzck, 1848 2<br />
U. auberiana glabra Millet, 1898 6<br />
U. bermudezi Acosta, 1940 3<br />
U. canariensis D'Orbigny, 1839 3<br />
U. flinti Cushman, 1923 I, 2<br />
U. occidentalis Cushman, 1923 3<br />
U. peregrina Cushman, 1923 2<br />
U. pygmea (D'Orbigny, 1826) O, 3, ME<br />
U. rustica Cushman y Edwards, 1923 2<br />
U. selseyensis Heron- Allen y Earland, 1909 6<br />
Subfamilia ANGULOGERININAE Galloway, 1933<br />
Género Trifarina Cushman, 1923<br />
T. bradyi (Cushman, 1923) O, 2, ME<br />
T. carinata (Cushman, 1927) O, 2, ME<br />
Familia REUSSELLIDAE Cushman, 1933<br />
Género Acostina Bermúdez, 1949<br />
A. piramidalis (Acosta, 1940) 2<br />
Género Reusella Galloway, 1933<br />
R. aguayoi Bermúdez, 1939 I, 2<br />
R. spinulosa (Reuss, 1850) I, 3<br />
FamiliaTRIMOSINIDAE Saidova, 1981<br />
Género Mimosina Millert, 1900<br />
M. echinata Heron Allen y Earland, 1915 2<br />
Superfamilia FURSENKOINACEA Loeblich y Tappan, 1961<br />
Familia FURSENKOINIDAE Loeblich y Tappan, 1961<br />
Género Fursenkoina Loeblich y Tappan, 1961<br />
F. punctata D'Orbigny, 1839 5<br />
F. squamosa (D'Orbigny, 1826) 2<br />
Superfamilia PLEUROSTOMELLACEA Reuss, 1860<br />
Familia PLEUROSTOMELLIDAE Reuss, 1860<br />
Subfamilia PLEUROSTOMELLINAE Reuss, 1860<br />
Género Ellipsoglandulina A. Silvestri, 1900<br />
E. antillea Bermúdez, 1939 2<br />
E. multicostata (Galloway y Morrei, 1929) 2<br />
Género Nodosarella Rzehak, 1895<br />
N. annulata (Terquem y Berthelin) 3<br />
Género Pleurostomella Reuss, 1860<br />
P. acuminata Cushman, 1922 2<br />
P. brevis Schwager, 1866 I, 2<br />
P. rapa var. recens Dervieux, 1899 2<br />
P. schuberti Cushman y Harris, 1927 2<br />
Superfamilia STILOSTOMELLACEA Finlay, 1947<br />
Familia STILOSTOMELLIDAE Finlay, 1947<br />
Género Orthomorphina Stainforth, 1952<br />
O. candei (D'Orbigny, 1839) 5<br />
Género Siphonodosaria A. Silvestri, 1924<br />
S. abyssorum (Brady, 1881) 2<br />
Género Stilostomella Guppy, 1894<br />
S. obsoleta (Acosta) 3<br />
Familia BAGGINIDAE Cushman, 1927<br />
Subfamilia SEROVAININAE Sliter, 1968<br />
Género Cancris de Montfort, 1808<br />
C. auricula (Fitchtel y Moll, 1798) O, 3, ME<br />
C. sagrai (D'Orbigny, 1839) O, 2, ME<br />
Género Physalidia Heron- Allen y Earland, 1928<br />
P. earlandi Bermúdez, 1935 2<br />
Género Valvulineria Cushman, 1926<br />
V. araucana (D'Orbigny, 1839) I, 2<br />
V. inaequalis (D'Orbigny, 1839) I, 3<br />
Familia EPONIDIDAE Hofker, 1951<br />
Subfamilia EPONIDINAE Hofker, 1951<br />
Género Eponides de Montfort, 1808<br />
E. antillarum (D'Orbigny, 1839) I, 3<br />
E. concentrica (Parker y Jones, 1864) 6<br />
E. repandus (Fichtel y Moll, 1798) 5<br />
E. tuberculata (Balkwill y Wright) 3<br />
E. umbonatus (Bailey) 3<br />
Género Poroeponides Cushman, 1944<br />
P. lateralis (Terquem, 1878) 2<br />
Familia DISCORBIDAE Ehrenberg, 1838<br />
Género Discorbis Lamarck, 1804<br />
D. aguayoi Bermúdez, 1935 3<br />
D. corrugata Millett 2<br />
D. cushmani Palmer y Bermúdez, 1936 I, 2<br />
D. erecta (Sidebottom, 1908) 3<br />
D. floridana Cushman, 1922 I, 3<br />
D. globosa (Sidebottom) 3<br />
D. obtusus (D'Orbigny) 1<br />
D. orbicularis var. antillea Bermúdez, 1935 3<br />
D. pulvinata (Brady) 3<br />
D. rosea (D'Orbigny, 1826) 1<br />
D. torrei Bermúdez, 1935 3<br />
D. valvulata var. granulosa (Heron-Allen y Earland) 3<br />
Familia ROSALINIDAE Reiss, 1963<br />
Género Neoconorbina Hofker, 1951<br />
N. crustata (Cushman) 4<br />
N. orbicularis (Terquem) 8<br />
Género Rosalina D'Orbigny, 1826<br />
R. advena Cushman, 1931 2<br />
R. antillea Bermúdez 2<br />
R. candeiana D'Orbigny, 1839 5<br />
R. concinna (Brady, 1884) 2<br />
R. floridana (Cushman, 1922) 1<br />
R. floridensis Cushman, 1931 2<br />
R. globularis D'Orbigny, 1826 2<br />
R. obtusa D'Orbigny, 1846 1<br />
R. semistriata D'Orbigny, 1839 5<br />
R. subaraucana (Cushman, 1922) 2<br />
Género Tretomphalus Möbius, 1880<br />
T. atlanticus Cushman, 1934 2<br />
T. bulloides (D'Orbigny, 1839) 2<br />
T. milletti (Heron-Allen y Earland) 2<br />
T. planus Cushman, 1924 1<br />
12 COCUYO 15, 2005
Familia SPHAEROIDINIDAE Cushman, 1927<br />
Género Sphaeroidina D'Orbigny, 1826<br />
S. bulloides (D'Orbigny, 1826) 3<br />
S. dehiscens (Parker y Jones) 3<br />
Superfamilia GLABRATELLACEA Loeblich y Tappan, 1964<br />
Familia GLABRATELLIDAE Loeblich y Tappan, 1964<br />
Género Glabratella Dorreen, 1948<br />
G. brasiliensis Boltovskoy, 1959 8<br />
G. erecta (Sidebottom, 1908) 2<br />
G. mirabilis Bermúdez y Seiglie, 1965 6<br />
Superfamilia SIPHONINACEA, Cushman, 1927<br />
Familia SIPHONINIDAE, Cushman, 1927<br />
Subfamilia SIPHONININAE, Cushman, 1927<br />
Género Siphonina Reuss, 1850<br />
S. bradyiana Cushman, 1927 O, 3, ME<br />
S. pulchra Cushman, 1919 O, 2, ME<br />
S. tenuicarinata Cushman I, 2<br />
Género Siphoninella Cushman, 1927<br />
S. soluta (Brady, 1884) O, 6, ME<br />
Subfamilia SIPHONINOIDINAE Loeblich y Tappan, 1984<br />
Género Siphoninoides Cushman, 1927<br />
S. echinatus (Brady, 1879) O, 8, ME<br />
S. glabra (Heron-Allen y Earland, 1915) 3<br />
Superfamilia DISCORBINELLACEA Sigal, 1952<br />
Familia DISCORBINELLIDAE Sigal, 1952<br />
Subfamilia DISCORBINELLINAE Sigal, 1952<br />
Género Discorbinella Cushman y Martin, 1935<br />
D. floridensis (Cushman, 1931) 4<br />
Género Laticarinina Galloway y Wissler, 1927<br />
L. pauperata (Parker y Jones, 1865) O, 2, ME<br />
Superfamilia PLANORBULINACEA Schwager, 1877<br />
Familia PLANULINIDAE Bermúdez, 1952<br />
Género Planulina D'Orbigny, 1826<br />
P. ariminensis D'Orbigny, 1826 O, 2, ME<br />
P. caribaea Cushman, 1931 6<br />
P. depressa (D'Orbigny) 6<br />
P. Edwarsiana (D'Orbigny, 1839) I, 5<br />
P. faveolata (Brady, 1884) O, 2, ME<br />
Familia CIBICIDIDAE Cushman. 1927<br />
Subfamilia CIBICIDINAE Cushman. 1927<br />
Género Cibicides de Montfort, 1808<br />
C. advena (D'Orbigny, 1839) 5<br />
C. floridanus (Cushman, 1918) 6<br />
C. io Cushman, 1931 2<br />
C. lobatulus (Cushman, 1935) 1<br />
C. mayori Cushman, 1924 1<br />
C. pseudoungerianus Cushman, 1931 O, 8, ME<br />
C. robertsonianus (Brady, 1884) O, 7, ME<br />
C. welestorfi (Schwager, 1866) 7<br />
Subfamilia STICHOCIBICIDINAE Saidova, 1981<br />
Género Dyocibicides Cushman y Valentine, 1930<br />
D. biserialis Cushman y Valentine, 1930 2<br />
Familia PLANORBULINIDAE Schwager, 1877<br />
Subfamilia PLANORBULININAE Schwager, 1877<br />
Género Planorbulina D'Orbigny, 1826<br />
P. acervalis Brady, 1884 O, 2, ME<br />
P. mediterranensis D'Orbigny, 1826 O, 6, ME<br />
Familia CYMBALOPORIDAE Cushman, 1927<br />
Subfamilia CYMBALOPORINAE Cushman, 1927<br />
Género Cymbaloporetta Cushman, 1928<br />
C. squamosa (D'Orbigny, 1839) O, 8<br />
C. poeyi (D'Orbigny, 1839) 5<br />
Familia VICTORIELLIDAE Chapman y Crespin, 1930<br />
Subfamilia CARPENTERIINAE Saidova, 1981<br />
Género Carpenteria Gray, 1858<br />
C. candei (D'Orbigny, 1839) 6<br />
C. monticularis Carter, 1877 3<br />
C. proteiformis Göes, 1882 O, 2, ME<br />
Superfamilia ACERVULINACEA Schultze, 1854<br />
Familia ACERVULINIDAE Schultze, 1854<br />
Género Acervulina Schultze, 1854<br />
A. inhaerens Schultze, 1854 I, 1<br />
Género Gypsina H.J. Carter, 1877<br />
G. discus (Göes, 1896) I, 3<br />
G.plana 8<br />
Género Sphaerogypsina Galloway, 1933<br />
S. globulus (Reuss, 1848) 2<br />
Subfamilia RUPERTININAE Loeblich y Tappan, 1961<br />
Género Rupertina Loeblich y Tappan, 1961<br />
R. stabilis (Wallich, 1877) 3<br />
Familia HOMOTREMATIDAE Cushman, 1927<br />
Género Homotrema Hickson, 1911<br />
H. rubrum (Lamarck, 1816) O, 2, ME<br />
Superfamilia ASTERIGERINACEA D'Orbigny, 1839<br />
Familia EPISTOMARIIDAE Hofker, 1954<br />
Subfamilia PALMERINELLINAE Loeblich y Tappan, 1984<br />
Género Palmerinella Bermúdez, 1934<br />
P. palmerae Bermúdez, 1934 O, 6, ME<br />
Familia ALFREDINIDAE S.N. Singh y Katia, 1972<br />
Género Epistomaroides Uchio, 1952<br />
E. atlantis Bermúdez, 1939 I, 2<br />
E. coronata Parker y Jones, 1857 I, 2<br />
E. flinti Cushman, 1931 O, 2<br />
E. flinti var. adelinensis Palmer y Bermúdez, 1934 2<br />
E. semipunctata (Bailey, 1851) O, 3, ME<br />
Familia ASTERIGERINIDAE D'Orbigny, 1839<br />
Género Asterigerina D'Orbigny, 1839<br />
A. carinata D'Orbigny, 1839 O, 8, ME<br />
A. lobata D'Orbigny, 1839 5<br />
Familia AMPHISTEGINIDAE Cushman, 1927<br />
Género Amphistegina D'Orbigny, 1826<br />
A.gibbosa D'Orbigny, 1839 O, 5, ME<br />
Superfamilia NONIONACEA Schultze, 1854<br />
Familia NONIONIDAE Schultze, 1854<br />
Subfamilia NONIONINAE Schultze, 1854<br />
Género Nonion de Montfort, 1808<br />
N. grateloupi (D'Orbigny) O, 3, ME<br />
N. stelligera (D'Orbigny) 2<br />
N. umbilicatulus (Walker y Jacob) 3<br />
Género Nonionella Cushman, 1926<br />
N. auris (D'Orbigny, 1826) 2<br />
N. grateloupi (D'Orbigny, 1826) 2<br />
Subfamilia ASTRONONIONINAE Saidova, 1981<br />
Género Astrononion Cushman y Edwards, 1937<br />
A. brownii (D'Orbigny, 1839) 5<br />
A. grateloupi (D'Orbigny, 1839) 5<br />
A. sloanii (D'Orbigny, 1839) 5<br />
A. steliger (D'Orbigny, 1839) 2<br />
A. tumidum Cushman y Edwards, 1937 2<br />
Subfamilia PULLENIINAE Schwager, 1877<br />
Género Melonis de Montfort, 1808<br />
M. pompiloides (Fichtel y Moll, 1798) 7<br />
Género Pullenia Parker y Jones, 1862<br />
P. riveroi Bermúdez, 1936 3<br />
P. obliqueloculata Parker y Jones, 1865 6<br />
Superfamilia CHILOSTOMELLACEA Brady, 1881<br />
Familia GAVELINELLIDAE Hofker, 1956<br />
Subfamilia GAVELINELLINAE Hofker, 1956<br />
Género Gyroidina D'Orbigny, 1826<br />
G. alfiformis R.E y C.K. Stewart, 1921 2<br />
G. soldanii D'Orbigny, 1826 O, 8, ME<br />
Familia TRICHOHYALIDAE Saidova, 1981<br />
Género Trichohyalus Loeblich y Tappan, 1953<br />
T. aguayoi Bermúdez, 1935 2<br />
Superfamilia ROTALIACEA Ehrenberg, 1839<br />
Familia ROTALIIDAE Ehrenberg, 1839<br />
Subfamilia ROTALIINAE Ehrenberg, 1839<br />
Género Rotalia Lamarck, 1804<br />
R. deformis (D'Orbigny, 1839) 5<br />
R. murrayi Heron-Allen y Earland, 1915 2<br />
COCUYO 15, 2005 13
Familia CALCARINIDAE Schwager, 1876<br />
Género Calcarina D'Orbigny, 1826<br />
C. calcar D'Orbigny, 1839 O, 5, ME<br />
Familia ELPHIDIIDAE Galloway, 1933<br />
Subfamilia ELPHIDIINAE Galloway, 1933<br />
Género Elphidium de Montfort, 1808<br />
E. advenum (Cushman, 1922) O, 2, ME<br />
E. articulatum (D'Orbigny) I, 6<br />
E. craticulatus (Fitchel y Moll, 1798) 3<br />
E. crispum (Linné, 1758) 2<br />
E. discoidale (D'Orbigny, 1839) O, 2, ME<br />
E. frimbiatulum (Cushman, 1918) 2<br />
E. lanieri (D'Orbigny, 1839) O, 6, ME<br />
E. lobatum Galloway y Heminway, 1941 4<br />
E. milletti (Heron-Allen y Earland, 1898) 6<br />
E. morenoi Bermúdez, 1935 I, 6<br />
E. poeyanum (D'Orbigny, 1839) I, 6<br />
E. sagrum (D'Orbigny, 1839) I, 6<br />
Superfamilia NUMMULITACEA de Blainville, 1827<br />
Familia NUMMULITIDAE de Blainville, 1827<br />
Género Heterostegina D'Orbigny, 1826<br />
H. antillarum D'Orbigny, 1839 O, 6, ME<br />
REFERENCIAS<br />
Alvarez García, N y P. Borro 1979. Los Foraminíferos del Golfo<br />
de Batabanó. Investigaciones Marinas 8 (47): 47-68.<br />
Bermúdez, P.J. 1935. Foraminíferos de la costa Norte de Cuba.<br />
Mem. Soc. Cubana Hist. Nat., Felipe Poey 9 (3): 129-224.<br />
Bermúdez, P. J. 1950. Contribución al estudio del Cenozoico<br />
Cubano. Mem. Soc. Cubana Hist. Nat., Felipe Poey 19 (3): 205-<br />
375.<br />
Borro, P. Inédito. Nota sobre los Foraminíferos recientes de<br />
Cuba.<br />
Buzas, M.1982 Distribution of the recent foraminifera in the<br />
Carribbean region. Special publication Smithsonian<br />
Institution.<br />
Broderman, J. Inédito. Catálogo de Foraminíferos de la Isla de<br />
Cuba. 1946<br />
Lee et al. 2000. An Illustrated guide to the Protozoa. Organisms<br />
traditionally refered to as Protozoa, or newly discovered<br />
Groups. Vol, II: 620-1432. Kansas. 66044. USA. Second<br />
Edition.<br />
Loeblich, A.R. Jr. y H. Tappan. 1964. Treatise on Invertebrate<br />
Paleontology. Part. C. Protista 2, vol. 1 y 2. Moore.<br />
Loeblich, A.R., Jr., y H. Tappan. 1988. Foraminiferal genera and their<br />
classification. Van Nostrand, Reinhold Co. Nueva York, (2<br />
vols.). 2047 pp.<br />
Orbigny, A. d' 1839. Foraminíferos. In: Ramón de la Sagra,<br />
Historia física, política y natural de la isla de Cuba. París.<br />
Arthur Bertran: 180<br />
Sen-Gupta Barun K., 1999. Modern Foraminifera. Kluwer<br />
Academic Publishers. Louisiana State University.<br />
Wright Baker R. 1960 Taxonomic notes, on the species figured<br />
by H. B. Brady in his report on the foraminiera dreged by<br />
H.M.S. Challenger during the years 1873, 1876 SEPM, no. 9,<br />
Oklahoma.<br />
Zernetzkyi F. B. 1975. Sedimentos de los mares de América<br />
central. Instituto de Ciencias geológicas. A.C. URSS. Kiev.<br />
Primer registro de Leprolochus spinifrons (Araneae:<br />
Zodariidae) para Las Antillas<br />
Giraldo Alayón García<br />
Museo Nacional de Historia Natural, Obispo no. 61,<br />
Habana Vieja 10100, Cuba<br />
ABSTRACT. The zodariid spider Leprolochus spinifrons Simon, 1892,<br />
previously known from Panama to Venezuela is reported for the first<br />
time for the West Indies, based in two females collected at Bequia, St.<br />
Vincent and the Grenadines. Some natural history data are also given.<br />
La familia Zodariidae tiene, hasta el presente, una especie<br />
registrada para Las Antillas (Jocqué, 1991; Alayón, 2000):<br />
Antillorena polli (Simon) que se ha encontrado en Aruba, Bonaire<br />
y Curacao (Antillas Neerlandesas), Las Bahamas y en Isla de la<br />
Juventud (antigua Isla de Pinos), Cuba.<br />
El género Leprolochus Simon, 1892 fue revisado por Jocqué<br />
(1988) y tiene siete especies: L. spinifrons Simon (Panamá,<br />
Colombia, Trinidad y Venezuela); L. stratus Jocqué & Platnick<br />
(Venezuela); L. birabeni Mello-Leitao (Brasil, Paraguay y<br />
Argentina), L. parahybae Mello-Leitao, L. levergere Lise, L. oeiras<br />
Lise y L. mucuge Lise (Brasil). Este género de arañas se caracteriza<br />
por la presencia de una línea de macrosetas transversas en frente<br />
de los ojos, margen anterior del prosoma (Jocqué, 1988; 1991;<br />
Jocqué & Platnick, 1989).<br />
Desde hace poco más de dos años el Padre Mark da Silva se<br />
encuentra colectando, fotografiando y acopiando información<br />
sobre la historia natural de los arácnidos en las islas Granadinas,<br />
principalmente en Bequia y Mayreau y ha colectado un número<br />
apreciable de ejemplares. Recientemente, tuve la oportunidad de<br />
visitar San Vicente y Las Granadinas, colectar y examinar parte<br />
de la colección de arañas atesoradas por él. En el material<br />
examinado encontré un ejemplar perteneciente a la familia<br />
Zodariidae (L. spinifrons Simon) que constituye el primer registro<br />
de este género y especie para las Antillas.<br />
El ejemplar fue colectado el 22 de junio de 2004. Se<br />
identificó por la forma y estructura del epigino, y por la forma<br />
cilíndrica de las macrosetas del borde del prosoma; la longitud de<br />
una de las hembras fue de 3.25 mm (prosoma: longitud, 1.35;<br />
anchura, 1.03). Posterior a mi visita se colectó en diciembre de<br />
2004, un segundo ejemplar, en el mismo sitio, presumiblemente<br />
de la misma especie.<br />
Estos ejemplares fueron capturados en el suelo del patio de<br />
la iglesia católica de la villa de Hamilton en la isla de Bequia, entre<br />
las piedras, bien camufladas por su color, apariencia e<br />
inmovilidad (cuando se les perturba se alejan con rapidez y<br />
adoptan una postura inmóvil). Una de las hembras está<br />
depositada en las colecciones del Departamento de Biología de<br />
la Universidad de West Indies, Barbados y la otra en las<br />
colecciones del Museo Nacional de Historia Natural de Cuba,<br />
Ciudad de La Habana. La presencia de esta especie en San<br />
Vicente y Las Granadinas nos hace pensar que pudiera también<br />
encontrarse en otras islas más al sur como Granada.<br />
Agradecimientos.- Se agradece al Padre Mark da Silva el ejemplar<br />
14 COCUYO 15, 2005
donado, a Julia Horrocks, Departamento de Biología de la<br />
Universidad de West Indies (Barbados) por los fondos para mi<br />
visita a San Vicente y Las Granadinas y a Marlon Mills-Browne<br />
y Paul Lewis las atenciones y amabilidades durante mi estancia.<br />
REFERENCIAS<br />
Alayón García, G. 2000. La arañas endémicas de Cuba. Rev.<br />
Ibérica Aracnol., 2: 1-48.<br />
Jocqué, R. 1988. An updating of the genus Leprolochus (Araneae:<br />
Zodariidae). Stud. Neotrop. Fauna Environm., 23: 77-87.<br />
Jocqué, R. 1991. A generic revision of the spider family<br />
Zodariidae (Araneae). Bull. American Mus. Nat. Hist., 201: 1-<br />
160.<br />
Jocqué, R. y N. I. Platnick. 1989. On Venezuelan Leprolochus<br />
(Araneae: Zodariidae). J. New York Entomol. Soc., 97: 471-<br />
474.<br />
mientras que apenas se vio en los árboles relativamente limpios,<br />
más adentro. Según Alayón (2003), se cree que las grandes telas<br />
compuestas que teje esta araña, pueden perjudicar la salud de<br />
los árboles, al bloquear la luz. Además de este efecto directo,<br />
una tela puede atrapar hojas muertas, que son aún más opacas<br />
que la tela misma. Este factor se observó en el presente caso.<br />
En particular, dos árboles -un mango y un samán- con extensas<br />
y densas coberturas de tela, se mostraron significamente<br />
desfoliados. Otro araneido -no identificado- se halló en la<br />
periferia de muchas telas de C. citricola.<br />
REFERENCIAS<br />
Alayón G., G. 2003. Cyrtophora citricola (Araneae: Araneidae),<br />
registro nuevo de araña para Cuba. Cocuyo 13:14.<br />
Alayón G., G.; L. F. de Armas y A. J. Abud 2001. Presencia<br />
de Cyrtophora citricola (Forskål, 1775) (Araneae: Araneidae)<br />
en las Antillas. Rev. Ibérica Arachnol., 4:9-10.<br />
.<br />
Soares Álvares, E. S. y M. de Maria 2004. First record of<br />
Cyrtophora citricola (Forskål) in Brazil (Araneae, Araneidae)<br />
Rev. Brasileira Zool., 21: 155-156.<br />
Observaciones sobre Cyrtophora citricola (Araneae:<br />
Araneidae) en Haití<br />
Christopher K. Starr<br />
Dep't of Life Sciences<br />
University of the West Indies<br />
St Augustine, Trinidad & Tobago. ckstarr99@hotmail.com<br />
El género Cyrtophora Simon, 1864 incluye 36 especies del<br />
Viejo Mundo y Australia. Cyrtophora citricola (Forskål, 1775) es<br />
una especie común que ha sido introducida en la región<br />
Neotropical region: Colombia, Cuba, Hispaniola y Brasil<br />
(Alayón et al. 2001; Alayón 2003; Soares Álvares y de Maria,<br />
2004).<br />
Durante una corta visita en el sur de Haití, Hispaniola en<br />
enero de 1998, encontré a C. citricola muy abundante en algunas<br />
partes de Puerto Príncipe y sus alrededores, especialmente en el<br />
cámpus de la Universidad Quisqueya, en el centro de la ciudad.<br />
Una muestra de 46 árboles tuvo 157 telas ocupadas. De estas,<br />
65 (41%) fueron telas compuestas, o sea, construidas y usadas<br />
por varios individuos. Las telas estuvieron a 3.1 m sobre el nivel<br />
del suelo, como promedio. Aparentemente los árboles de<br />
mangos y otras leguminosas presentaron más telas que otras<br />
plantas.<br />
En esa época, las abundantes palmeras reales (Roystonea<br />
regia) estaban en plena florescencia y supuestamente atraían<br />
gran cantidad de insectos. Sin embargo, 19 palmas estudiadas,<br />
tuvieron una sola tela de C. citricola. Tampoco se hallaron en un<br />
grupo de arbustos bajos.<br />
Estas observaciones se hicieron durante la estación seca,<br />
cuando la ciudad tuvo mucho polvo. Es notable que dentro del<br />
cámpus C. citricola se halló sobre todo hacia el perímetro,<br />
Macroinvertebrados dulceacuícolas y calidad de las<br />
aguas de tres estaciones de Sierra de Nipe y Sierra<br />
Cristal, Región Oriental de Cuba<br />
1 1<br />
Dany D. González Lazo , Adrián Trapero Quintana , Carlos<br />
1 2<br />
Naranjo López y Pedro López Del Castillo<br />
1 Departamento de Biología de la Universidad de Oriente. Santiago<br />
de Cuba. Cuba. atrapero@cnt.uo.edu y naranjo@cnt.uo.edu<br />
2 Empresa Nacional para la Protección de la Flora y la Fauna.<br />
Parque Nacional Turquino. Cuba. ffturquino@enet.cu y<br />
plopezc77@yahoo.es<br />
RESUMEN. Se realizó un inventario de los macroinvertebrados<br />
dulceacuícolas y se determinó la calidad del agua de tres estaciones de<br />
los ríos La Chivera, Cirino y Grande, en Sierra de Nipe y Sierra de<br />
Cristal (región oriental de Cuba). Se detectó un número bajo de<br />
especies, familias e individuos, con relación a otras áreas geográficas.<br />
La mayor cantidad de especies (N=14) fue colectada en la estación de<br />
mayor altitud (La Chivera), a 405 m, que a su vez fue la más<br />
heterogénea en cuanto a características ecológicas. Se comprobó que<br />
en esta zona predominan los rabiones, por lo que las especies más<br />
abundantes fueron las adaptadas a este microhábitat. El resultado del<br />
BMWP-Cu, indica que la estación de mejor calidad del agua fue La<br />
Chivera, sin embargo ninguna de las estaciones alcanzó la categoría de<br />
“Buena”, a pesar de ser localidades con bajos niveles de<br />
antropización, debido fundamentalmente a la escasez de familias e<br />
individuos en las colectas, sobre todo en los Ríos Cirino y Grande,<br />
resultado de las fuertes corrientes, la homogeneidad del fondo, la<br />
ausencia de vegetación acuática y de orilla y la completa exposición al<br />
sol de estas estaciones durante todo el día. Los valores de los índices<br />
empleados, variaron de forma similar, lo que sugiere una estrecha<br />
COCUYO 15, 2005 15
elación entre el número de especies, la diversidad biológica y el grado<br />
de contaminación de las aguas en las estaciones analizadas.<br />
En las comunidades de agua dulce, encontramos plantas,<br />
animales y otras formas de vida adaptadas a vivir y reproducirse<br />
en la corriente de los arroyos, ríos y en las aguas inmóviles de los<br />
lagos y los estanques. Las aguas corrientes o hábitat lóticos,<br />
incluyen todas las partes del curso de los ríos, arroyos y<br />
manantiales de su cabecera, la zona central del valle, con sus<br />
remansos y sus rápidos, la zona de la llanura aluvial, y los<br />
estuarios en los que vierten sus aguas al mar. Al evaluar la calidad<br />
de las aguas mediante el estudio de la composición y estructura<br />
de comunidades y organismos surge el término calidad<br />
biológica. Se considera que un medio acuático presenta una<br />
buena calidad biológica cuando tiene características naturales<br />
que permiten que en su seno se desarrollen las comunidades de<br />
organismos que le son propias (Alba-Tercedor, 1996).<br />
Los medios acuáticos son colonizados por animales y<br />
vegetales cuya estructura responde en condiciones normales a<br />
un equilibrio. Cuando sobreviene una perturbación en el medio,<br />
ya sea de origen natural o artificial, se altera la estructura de las<br />
poblaciones; trastornos que se manifiestan en un cambio en la<br />
dominancia relativa de las especies, acompañado de<br />
sustituciones en la fauna y flora dependiendo de lo amplio o<br />
estrecho que sea su gama de resistencia frente a esa perturbación,<br />
pudiendo llegar a la destrucción completa de la biocenosis,<br />
cuando es demasiado drástica (Burrillo, 1997).<br />
Producto de las fuertes cargas de contaminantes a que han<br />
sido sometidos en los últimas décadas los ríos cubanos, en<br />
especial las partes bajas, ha aumentado la preocupación por<br />
conocer la calidad de sus aguas y las comunidades de organismos<br />
que en ellos habita y así determinar con exactitud su grado de<br />
deterioro. Es por ello que se toman medidas para un uso<br />
racional de los recursos hídricos y de esta forma no alterar las<br />
condiciones naturales que conlleven a la pérdida de la<br />
biodiversidad. Teniendo en cuenta lo anterior nos trazamos el<br />
objetivo de inventariar las comunidades de macroinvertebrados<br />
existentes en tres ríos de Sierra de Nipe y Sierra Cristal y<br />
determinar la calidad de las aguas de estas estaciones.<br />
El muestreo se desarrolló en el período comprendido entre<br />
el 23 de marzo al 2 de abril de 2004, en la altiplanicie Pinares de<br />
Mayarí y Sierra Cristal, provincia Holguín, macizo Nipe-Sagüa-<br />
Baracoa, región oriental de Cuba. Para la investigación se<br />
seleccionaron tres estaciones de muestreo, en tres ríos, las cuales<br />
pasamos a describir brevemente: Estación 1. Río La Chivera.<br />
Esta área está ubicada a los 405 m de altitud, (20º24'58'' N;<br />
75º49'58'' W) y muy cercana poblado Pinalito, ladera sur de la<br />
altiplanicie de Pinares de Mayarí, Sierra de Nipe. El ancho del río<br />
fue de 6 m tanto en remansos como en rabiones, presentó un<br />
fondo pedregoso-arenoso, la vegetación de la orilla fue escasa y<br />
el cauce totalmente cubierto por un bosque en galería, que<br />
provocó poca penetración de los rayos solares y una baja<br />
temperatura (20ºC). La profundidad del área de colecta fue de 15<br />
cm y la transparencia del agua total. Estación 2. Río Cirino. Esta<br />
estación estuvo ubicada a 20 m de su entronque con el río<br />
Grande, a los 265 m de altitud, (20º33'35'' N; 75º24'57'' W),<br />
2<br />
Sierra de Cristal. Se muestreó en un área de 25 m con una<br />
profundidad máxima de 40 cm. El fondo estuvo formado<br />
únicamente por piedras, predominando la roca conocida como<br />
“china pelona”. No presentó vegetación de orilla y el cauce<br />
estuvo descubierto, por tanto la incidencia de los rayos solares<br />
fue total, provocando un incremento en la temperatura del agua<br />
(23º C). En esta estación el agua fue, al igual que la anterior,<br />
completamente transparente. Estación 3. Río Grande. El<br />
muestreo en esta estación fue realizado en el río Grande a los 294<br />
m de altitud (20º38'28''N; 75º24'50''W), en la intersección del<br />
camino a Pico Mambrú con el río del mismo nombre, también en<br />
Sierra de Cristal. El ancho del río fue de 60 m, cauce totalmente<br />
descubierto, el fondo pedregoso, la profundidad máxima fue de<br />
70 cm. La temperatura del agua registrada fue de 25 ºC. En la<br />
orilla del río, la vegetación fue escasa. La incidencia de los rayos<br />
solares fue completa y el agua transparente .En todas las<br />
estaciones, fundamentalmente en las dos últimas, el hábitat<br />
predominante fue el de rabiones.<br />
La colecta de ejemplares se realizó empleando los métodos<br />
que se describen a continuación: Levantamiento de piedras.<br />
En cada estación se levantaron 25 piedras y se revisaron<br />
cuidadosamente, extrayendo con un pincel o con pinzas<br />
entomológicas blandas las larvas adheridas a ellas, evitando el<br />
daño de algunas estructuras como patas y branquias,<br />
importantes para la identificación del material. El material se<br />
depositó en alcohol al 90%. Red de colecta en contracorriente<br />
en rabiones y remansos. Se realizó un arrastre con red<br />
entomológica en dirección contraria al sentido de la corriente del<br />
río, tanto en zonas de rabiones como en remansos. Para la<br />
aplicación de este método participaron dos personas, una de las<br />
cuales se desplazó removiendo el substrato, mientras que la otra<br />
avanzó a continuación arrastrando la red contra el fondo. El<br />
contenido se vertió en una bandeja blanca, se extrajo con pinzas<br />
entomológicas y se depositó en alcohol al 90 %. Red de colecta<br />
en orilla. Se utilizó la misma red entomológica que para<br />
contracorriente, pero el arrastre se hizo en 5 m lineales de orilla y<br />
tratando de introducir el mismo en las raíces, troncos, rocas y<br />
detritos. Posteriormente, se vertió el contenido en una bandeja<br />
blanca y se procedió igual que con los métodos anteriores. Los<br />
ejemplares colectados fueron transportados hacia el laboratorio<br />
de Entomología del Departamento de Biología, Universidad de<br />
Oriente, donde se realizó su determinación taxonómica para la<br />
cual se consultaron diferentes claves: Alayo (1968a y b, 1974);<br />
Peters (1971); Naranjo (1986); Kluge (1991, 1992a y b; 1993);<br />
Kluge y Naranjo (1990; 1994); Botosaneanu (1994); Merrit y<br />
Cummins (1996) y Westfall (1996).<br />
Evaluación de la calidad de las aguas<br />
Para la evaluación de la calidad de las aguas mediante el<br />
empleo de la composición y estructura de los organismos que en<br />
ellas habitan, se empleó el BMWP' de Alba-Tercedor y Sánchez<br />
(1988) adaptado a las condiciones de nuestro país por Naranjo et<br />
al. (2003) nombrado como BMWP-Cu para Cuba. Se utilizaron<br />
además el índice de Shanon-Weaver y el de estructura específica,<br />
para tener un criterio más certero sobre la calidad del agua.<br />
BMWP-Cu = T1+T2+T3...+TN; donde T es el valor de<br />
tolerancia de cada familia (Tabla 1).<br />
16 COCUYO 15, 2005
H= Pi log Pi; donde n = número de ejemplares; N = número<br />
2<br />
total de ejemplares en la muestra; Pi = n/N.<br />
Estructura específica: es el número de especies presentes en la<br />
muestra.<br />
RESULTADOS Y DISCUSIÓN<br />
Las colectas realizadas arrojaron 98 individuos<br />
pertenecientes a 26 taxa infragenéricos, distribuidos en 20<br />
familias de ocho órdenes, tres Clases y tres Phyla (Tabla 2). El<br />
Phylum Arthropoda fue el mejor representado con 23 especies<br />
(88.5 % del total), todas de la Clase Insecta, y 88 ejemplares. De<br />
los Phyla Platyhelminthes y Mollusca se colectaron 10<br />
ejemplares pertenecientes a tres especies (11.5 % del total)<br />
(Tabla 2). Se encontraron dos ejemplares del género Antillopsyche<br />
(Polycentropodidae: Trichoptera), en Río Grande.<br />
Anteriormente estaba registrado sólo para el occidente y centro<br />
del país (Botosaneanu, 1979, 1980; Naranjo y González, en<br />
prensa). Otro resultado interesante lo constituyó el hallazgo de<br />
las muy raras especies del orden Ephemeroptera, Hagenulus<br />
(Hagenulus) caligatus (23 larvas) y H. (Turquinophlebia) grandis (tres<br />
ejemplares) en Río Grande y Río Cirino (Tabla 2); la primera está<br />
registrada para los sectores occidental y oriental (Sierra Maestra)<br />
y la segunda sólo para la Sierra Maestra (Peters 1971; Kluge,<br />
1993; Naranjo y Cañizares, 1999), por lo que se citan por primera<br />
vez en este trabajo para el subsector nororiental de Cuba. De las<br />
26 especies encontradas, nueve fueron endémicos de Cuba (34.6<br />
% de endemismo), de ellos, cuatro: Cloeodes superior, H. (T.)<br />
grandis, H (Careospina) H. sierramaestrae y Calosopsyche cubana, de la<br />
Tabla 1. Valores de tolerancia asignados a las 69 familias de macroinvertebrados acuáticos en Cuba, para la<br />
obtención del BMWP-Cu, agrupadas según la categoría taxonómica superior.<br />
Ephemeroptera Trichoptera Amphipoda<br />
Baetidae 7 Hydrobiosidae 10 Gammaridae 1<br />
Leptophlebiidae 9 Calamoceratidae 8<br />
Leptohyphidae 6 Glossosomatidae 9 Decapoda<br />
Caenidae 4 Helicopsychidae 8 Palaemonidae 6<br />
Euthyplociidae 9 Hydropsychidae 5 Pseudotelphusidae 6<br />
Hydroptilidae 7 Atyidae 5<br />
Hemiptera Leptoceridae 8<br />
Corixidae 2 Odontoceridae 10 Mollusca<br />
Pleidae 2 Philopotamidae 8 Thiaridae 7<br />
Gerridae 3 Polycentropodidae 8 Neritidae 7<br />
Hydrometridae 3 Xiphocentronidae 9 Ampullaridae 4<br />
Mesoveliidae 3 Ecnomidae 10 Ancylidae 6<br />
Belostomatidae 4<br />
Veliidae 6 Diptera Turbellaria<br />
Notonectidae 7 Blephariceridae 10 Dugesidae 7<br />
Nepidae 6 Ceratopogonidae 5<br />
Chironomidae 4 Coleoptera<br />
Odonata Dolichopodidae 7 Dryopidae 5<br />
Aeshnidae 8 Empididae 5 Gyrinidae 3<br />
Coenagrionidae 5 Psychodidae 4 Hydrophilidae 5<br />
Gomphidae 8 Simullidae 5 Elmidae 6<br />
Lestidae 3 Tabanidae 6 Psephenidae 7<br />
Libellulidae 3 Tipulidae 6 Carabidae 8<br />
Protoneuridae 4 Dixidae 7 Noteridae 4<br />
Megapodagrionidae 9 Stratiomyidae 7 Halliplidae 4<br />
Culicidae 2 Dytiscidae 4<br />
Lepidoptera Scirtidae 7<br />
Pyralidae<br />
Acarina<br />
5 Hydracaridae 3 Hirudinea<br />
Hydrochidae 3 Gnathobdellidae 1<br />
Hydraenidae 5<br />
COCUYO 15, 2005 17
egión oriental. Como endémico antillano se colectó Hypolestes trinitatis, del orden Odonata, la única especie de este<br />
género en nuestro archipiélago (Tabla 2).<br />
Tabla 2. Lista de los taxa infragenéricos por estaciones de muestreo colectados en Sierra de Nipe y Sierra Cristal. *<br />
especies endémicas de Cuba. ** especies endémicas antillanas. E1= La Chivera; E2 = Río Cirino; E3 = Río Grande.<br />
PHYLUM CLASE ORDEN FAMILIA ESPECIES<br />
Estación de<br />
colecta<br />
E1 E2 E3<br />
Platyhelminthes Turbellaria Tricladida Dugesidae 1. Dugesia cubana (Codreanu y Balsesco,<br />
1973)<br />
4<br />
Mollusca Gastropoda Basomantophora Ancylidae 2. Sp. 3<br />
Mesogastropoda Thiaridae 3. Tarebia granifera (Lamarck, 1816) 3<br />
Arthropoda Insecta Odonata Coenagrionidae 4. Enallagma coecum (Hagen, 1861) 1<br />
Megapoda- 5. Hypolestes trinitatis Gundlach, 1888 **<br />
grionidae<br />
3<br />
Libellulidae 6. Perithemis domitia (Drury, 1773) 1<br />
7. Macrothemis celeno (Selys, 1857) 1<br />
Ephemeroptera Baetidae 8. Cloeodes superior Kluge, 1991* 1<br />
9. Cloeodes inferior Kluge, 1991 * 2<br />
10. Hagenulus (Hagenulus) caligatus Peters &<br />
Alayo, 1971*<br />
23<br />
Leptophlebiidae 11. Farrodes bimaculatus Peters & Alayo, 1971<br />
*<br />
3<br />
12. Hagenulus (Turquinophlebia) grandis Kluge,<br />
1993 *<br />
2 1<br />
13. Hagenulus (Careospina) hespera<br />
sierramaestrae Kluge, 1993 *<br />
6<br />
Caenidae 14. Caenis sp 3<br />
Trichoptera Calamoceratidae 15. Phylloicus chalybeus (Hagen, 1861)* 3<br />
Hydrobiosidae 16. Atopsyche sp. 1<br />
Glossosomatidae 17. Cubanoptila purpurea Sykora en Bots. y<br />
Syk., 1973 *<br />
1 2<br />
Hydropsychidae 18. Calosopsyche cubana (Flint, 1962)* 8<br />
19. Smicridea sp. (mínima , Flint) 1 3<br />
Helicopsychidae 20. Helicopsyche sp. (near comosa) 1<br />
Polycentro- 21. Antillopsyche sp. 1<br />
podidae<br />
2<br />
Coleoptera Psephenidae 22. Pheneps sp. 6 1<br />
Elmidae 23. Phanocerus sp. 2<br />
Gyrinidae 24. Dineutus longimanus Och, 1938 2<br />
Diptera Simuliidae 25. Psilopelmia haematopotum (Malloch,1914) 1<br />
Chironomidae 26. Sp-4 7<br />
Total de ejemplares capturados en cada estación 34 24 40<br />
La estación La Chivera fue la mejor representada en cuanto al número de especies con 14 (53.8 %). Además, fue<br />
la única donde se encontraron ejemplares de los Phyla Platyhelminthes y Mollusca (Tabla 2), y la estación más<br />
heterogénea, en cuanto a condiciones ecológicas, presentando una alternancia entre los dos microhábitats típicos<br />
(rabiones y remansos). Río Cirino al igual que río Grande, presentó ocho especies para 23.3 % del total; sin<br />
embargo, en esta última estación, se encontró la mayor cantidad de ejemplares de la muestra (40 individuos, 40.8 %<br />
del total). Esto se debe a que fue encontrada H. caligatus, la especie más abundante para toda la muestra (Tabla 2)<br />
18 COCUYO 15, 2005
Scapanea frontalis fue observada volando en las tres estaciones<br />
de muestreo, pero no se incluyó en este análisis debido a que no<br />
fueron encontradas sus larvas. La especie más abundante fue H.<br />
caligatus con 23 ejemplares, capturados en río Grande (Tabla 2), le<br />
sigue por su orden Calosopsyche cubana (ocho ejemplares en río<br />
Grande), Pheneps sp (seis ejemplares en río Cirino y uno en río<br />
Grande) y Chironomidae sp 4 (siete ejemplares en río La Chivera).<br />
Las especies menos abundantes resultaron: Enallagma coecum,<br />
Perithemis domotia, Macrotemis celeno, Cloeodes superior, Atopsyche sp.,<br />
Helicopsyche sp. near comosa, y Psilopelmia haematopotum, todas con<br />
un solo ejemplar capturado.<br />
Como se observa en este análisis (excepto Chironomidae sp 4<br />
que fue colectada en zona de remanso de la estación La Chivera)<br />
las especies más abundantes, están adaptadas a vivir en zonas de<br />
rabiones y fueron colectadas en las estaciones de mayor<br />
velocidad de corriente (río Cirino y río Grande). Con respecto a<br />
las especies de menor abundancia hay que destacar que se<br />
correspondieron con las adaptadas a vivir en zonas de remanso<br />
(excepto Helicopsyche sp. near comosa), fundamentalmente<br />
asociadas a la vegetación de las márgenes, como es el caso de los<br />
odonatos. Este resultado es propio de torrentes montañosos<br />
donde el hábitat predominante es el de rabiones, y<br />
evidentemente la mayoría de los taxones están adaptados a estas<br />
condiciones.<br />
En los muestreos realizados se detectó un número bajo de<br />
especies y ejemplares, por localidades. Aldana y Fonseca (2001)<br />
citaron para la cuenca media del río Mayarí (considerando que<br />
por su altura, las estaciones analizadas en este trabajo, se<br />
encuentran ubicadas en la cuenca media de varios ríos) un<br />
promedio de 32 especies y 209 ejemplares, por estaciones, en<br />
época de seca. Este resultado puede tener su explicación en los<br />
siguientes aspectos: 1. Las fuertes corrientes y la presencia, sobre<br />
todo en los ríos Cirino y Grande, de un fondo pedregoso muy<br />
homogéneo lo que favorece únicamente a las especies con<br />
adaptaciones ecomorfológicas (cuerpo depreso y/o<br />
mecanismos de adhesión a las rocas) para vivir encima o debajo<br />
de las piedras y no ser arrastrados por la corriente. Tal es el caso<br />
de los leptoflébidos, los tricópteros y las especies H. trinitatis y<br />
Pheneps sp., presentes en las estaciones antes mencionadas. 2. La<br />
escasa vegetación de orilla en río La Chivera y su ausencia total en<br />
los ríos Cirino y Grande, que juega un rol muy importante en la<br />
emergencia de los subimagos, imagos, en el apareamiento, la<br />
oviposición, como refugio de algunas especies (Odonatos) y<br />
sitio de alimentación. 3. La<br />
completa exposición a los<br />
rayos solares de las<br />
estaciones Río Grande y Río<br />
Cirino, producto de la<br />
ausencia de vegetación de<br />
cubierta y de orilla,<br />
provocando un incremento<br />
de la temperatura del agua. 4.<br />
La ausencia de plantas<br />
acuáticas, muy importantes<br />
para el suministro de<br />
alimento y oxígeno al medio<br />
acuático y como refugio.<br />
Clases Calidad BMWP-Cu<br />
(Valor)<br />
I. Buena >150<br />
101-120<br />
Las estaciones analizadas, a pesar de no estar ubicadas a<br />
grandes altitudes, presentaron bajos niveles de antropización, lo<br />
que sugiere intuitivamente una buena calidad del agua. La<br />
estación con mejor calidad de las aguas, según el BMWP-Cu, en<br />
la categoría de “aceptable” y significado de “evidentes algunos<br />
efectos de contaminación” fue La Chivera, que se encuentra a<br />
mayor altitud (405 m), mientras que con la calidad de “dudosa”<br />
(contaminación evidente) quedaron Río Cirino y Río Grande<br />
(Tablas 3 y 4). En el caso del análisis basado en el índice de<br />
Shannon-weaver los resultados obtenidos se comportaron de<br />
forma similar que en el BMWP-Cu, arrojando a la estación La<br />
Chivera como la de mejor calidad (aguas no contaminadas). Las<br />
estaciones Río Cirino y Río Grande, quedaron en la categoría de<br />
“medianamente contaminadas”. La estructura específica se<br />
comportó igual que en los índices anteriores, donde el mayor<br />
valor fue en La Chivera y las restantes mantuvieron un valor<br />
constante muy bajo (Tabla 3). Como se observa en las Tablas 3 y<br />
4, ninguna estación alcanzó la categoría de “Buena”, lo que<br />
resulta raro en zonas con bajos niveles de antropización.<br />
Pensamos que este resultado se debe fundamentalmente a la<br />
escasez de familias e individuos en las colectas, sobre todo en Río<br />
Cirino y Río Grande, resultado de las fuertes corrientes, la<br />
homogeneidad del fondo, la ausencia de vegetación acuática y de<br />
orilla y la completa exposición al sol de estas estaciones durante<br />
todo el día. Los valores del BMWP-Cu oscilaron de forma<br />
similar a los de riqueza específica y diversidad biológica (Fig. 1),<br />
lo que sugiere una estrecha relación entre los dos últimos índices<br />
y la calidad del agua en las estaciones muestreadas.<br />
Tabla 3. Resultados de los índices BMWP-Cu, Shannon-Weaver<br />
(H) y Estructura Específica (E), en las estaciones.<br />
Indices La Chivera Río Rÿí o Cirino Río R ÿíoGrande Grande<br />
BMWP-Cu 80 47 55<br />
H 3.51 2.76 1.95<br />
E 14 8 8<br />
Tabla 4. Clases de calidad, significación de los valores del BMWP-Cu y colores a utilizar para la<br />
representación cartográfica en ríos de Cuba.<br />
Significado<br />
Aguas muy limpias, aguas no<br />
contaminadas o no alteradas de modo<br />
sensible<br />
COCUYO 15, 2005 19<br />
azul<br />
Color<br />
II. Aceptable 61-100 Evidentes algunos efectos de verde<br />
contaminación<br />
contaminaciÿó n<br />
III. Dudosa 36-60 Aguas contaminadas amarillo<br />
IV. Crÿí Crítica<br />
16-35 Aguas muy contaminadas naranja<br />
V. Muy crítica crÿí<br />
Valor de los índices<br />
100<br />
80<br />
60<br />
40<br />
20<br />
0<br />
1 2 3<br />
Estaciones<br />
Fig. 1. Resultados de los índices de BMWP-Cu , Estructura<br />
Específica y diversidad de Shannon-Weave r<br />
CONCLUSIONES<br />
• Se colectaron 98 individuos pertenecientes a 26 taxa<br />
infragenéricos, distribuidos en 20 familias de ocho<br />
órdenes, tres Clases y tres Phyla.<br />
• La Clase Insecta fue la mejor representada, con 23<br />
especies y 88 ejemplares.<br />
• Las especies más abundantes fueron las adaptadas a<br />
vivir en rabiones, mientras que las menos abundantes<br />
fueron las típicas de remansos. Este resultado es propio<br />
de zonas montañosas donde el hábitat predominante es<br />
el de rabiones.<br />
• Se citan para la zona de estudio nueve especies<br />
endémicas de Cuba y una de las Antillas.<br />
• La mejor calidad de las aguas se obtuvo en la estación La<br />
Chivera, la de mayor altura.<br />
• Se observó una relación estrecha entre el número de<br />
especies, la diversidad biológica y la calidad del agua, en<br />
las estaciones estudiadas.<br />
REFERENCIAS<br />
Alayo, P. 1968a. Las Libélulas de Cuba (Insecta-Odonata).<br />
Torreia, Nueva Serie 2: 3-102.<br />
Alayo, P. 1968b. Las Libélulas de Cuba. (Insecta Odonata)<br />
Torreia, Nueva Serie 3: 3- 54<br />
Alayo, P. 1974. Los Hemípteros acuáticos de Cuba. Torreia,<br />
Nueva Serie 36: 9-64.<br />
Alba-Tercedor, J. y A. Sánchez-Ortega. 1988. Un método rápido<br />
y simple para evaluar la calidad biológica de las aguas<br />
corrientes basado en el de Hellawell (1978). Limnética 4: 51-<br />
55.<br />
Aldana, M y L. Fonseca. 2001. caracterización ecológica de la<br />
macrofauna del río Mayarí. Trabajo de Diploma (Inédito).<br />
Universidad de Oriente. Pp. 48-68.<br />
Botosaneanu, L. 1979. The Caddiflies (Trichoptera) of Cuba<br />
and Isla de Pinos. A Synthesis. Studi. Faun. Curacao and other<br />
Carib. Islands 29: 33-62.<br />
Botosaneanu, L. 1980. Trichopteres adultes de Cuba collectés<br />
par zoologistes cubains (Trichoptera). Mitteilungen der<br />
Münchner Entomol., Gesellschaft 69: 91-116.<br />
Botosaneanu, L. 1994. A study of the larvae of caddisflies<br />
(Trichoptera) from Cuba. Trop. Zool., 7: 451-475.<br />
Burillo-Borrego, L. 1997. La calidad de las aguas en los<br />
humedales: los indicadores biológicos. Universidad de<br />
Valencia. Departamento de Biología Animal. España.<br />
Kluge, N. 1991. Efímeras cubanas de la familia Baetidae<br />
(Ephemeroptera) 1: Géneros Callibaetis, Cloeodes y Paracloeodes.<br />
Zool. Journal , Rusia Vol.12.<br />
Kluge, N. 1992a. Efímeras cubanas de la familia Baetidae<br />
(Ephemeroptera) II: subgéneros Caribaetis y Americabaetis<br />
del género Baetis. Zool. Journal, Rusia 4: 13-20.<br />
Kluge, N. 1992b. Efímeras cubanas de la familia Baetidae<br />
(Ephemeroptera) III: subgénero Fallceon del género Baetis.<br />
Zool. Journal, Rusia. 5: 38-47.<br />
Kluge, N. 1993. A revision of Leptophlebiidae from Cuba<br />
(Ephemeroptera). Zoosytemática Rossica 2(2):247-285.<br />
Kluge, N. y C. Naranjo. 1990. Efímeras de la familia<br />
Leptohyphidae (Ephemeroptera) de Cuba. Rev. Entomol.<br />
URSS. 49: 564-576.<br />
Kluge, N. y C. Naranjo. 1994. Una especie peculiar de efímera<br />
del género Euthyplocia Eaton (Ephemeroptera:<br />
Euthyplociidae) de Cuba. Entomol. Obozrenie, Rusia. 73: 777-<br />
781.<br />
Merritt, R. y K. Cummins. 1996. An introduction to the aquatic<br />
insects of North America.Kwendall/Hunt Publ. Company: 3a.<br />
Edition. Pp 164-211.<br />
Naranjo, C. 1986. Análisis ecológicofaunístico de los insectos<br />
anfibióticos del Parque Nacional Sierra Maestra. Tesis de<br />
Doctorado. Univ. Est. Leningrado: pp: 32-46.<br />
Naranjo, C. y M. Cañizares. 1999. Situación actual de<br />
Ephemeroptera (Insecta). Cocuyo 8:17-19.<br />
Naranjo, C.; G. Garcés; D. González; S. Muñóz e Y. Musle.<br />
2003. Una metodología rápida y de fácil aplicación para la<br />
evaluación de la calidad del agua utilizando el índice BMWP-<br />
CU para ríos cubanos. Evento de la facultad de Ciencias<br />
Naturales de la Univ. de Oriente, ISBN 959-207-093-8.<br />
Naranjo, C. y D. González. En prensa. Situación actual del<br />
orden Trichoptera en Cuba. Biodiversidad de Cuba Oriental.<br />
Peters, W. L. 1971. A Revision of The Lepthophlebiidae of the<br />
West Indies (Ephemeroptera) Smithsonian Contr. Zool., No<br />
67.<br />
Westfall, J. R. y M. J. May. 1996. Damselflies of North America.<br />
Scientific Publishers: pp 112-570.<br />
20 COCUYO 15, 2005
Nuevas adiciones a la entomofauna del Area Protegida<br />
“Mil Cumbres”, Pinar del Río, Cuba<br />
Ileana Fernández García* y Jorge L. Fontenla **<br />
* Instituto de Ecología y Sistemática, A.P. 8029, Ciudad de La<br />
Habana, 10800, Cuba<br />
** Museo Nacional de Historia Natural, Obispo no. 61, Habana<br />
Vieja 10100, Cuba<br />
Las Areas Protegidas representan un elemento fundamental<br />
para la conservación de la biodiversidad pero para lograr este<br />
propósito, es necesario conocer primero las especies de la flora y<br />
fauna que lo integran.<br />
A partir de 1998, comenzó a estudiarse la entomofauna de la<br />
Cordillera de Guaniguanico, provincia de Pinar del Río,<br />
incluyendo entre las localidades al Área Protegida Mil Cumbres<br />
(Armas et al., 2000; Coy et al. 2000; Fernández, 2001, Hidalgo-<br />
Gato y Rodríguez-León, 2000, Rodríguez, 2005 ).<br />
Posteriormente, continuaron realizándose inventarios y<br />
monitoreos en Mil Cumbres, los que permitieron registrar<br />
especies nuevas de insectos para esta localidad. Fernández y<br />
Lozada (en prensa) informaron sobre los coleópteros, mientras<br />
que Rodríguez (2005) y M. Hidalgo-Gato (com. pers.) han<br />
encontrado otras especies de dípteros y homópteros<br />
auquenorrhincos, respectivamente.<br />
En este trabajo se relacionan 54 nuevos registros de insectos<br />
para Mil Cumbres, pertenecientes a los órdenes Neuroptera (una<br />
especie), Orthoptera (una familia, cuatro géneros y cinco<br />
especies), Heteroptera (11, 26 y 26) e Hymenoptera (7, 15, 22).<br />
La información se obtuvo mediante la revisión de las colecciones<br />
entomológicas del Instituto de Ecología y Sistemática (IES) y de<br />
recolectas realizadas durante los meses de junio y septiembre del<br />
2001 y enero y febrero del 2002. Los ejemplares recolectados se<br />
encuentran depositados en la colección entomológica del IES.<br />
Lista de insectos<br />
ORTHOPTERA<br />
Tettigoniidae<br />
Neoconocephalus maxillosus (Fabr.): San Diego de los Baños<br />
Neoconocephalus nietoi (Saussure): San Diego de los Baños<br />
Caulopsis cuspidatus (Scudder): San Diego de los Baños<br />
Conocephalus fasciatus (Degeer): San Diego de los Baños<br />
Stilpnochlora couloniana (Saussure): San Diego de los Baños<br />
NEUROPTERA<br />
Chrysopidae<br />
Ceraeochrysa smithi (Navás): San Diego de los Baños<br />
HETEROPTERA<br />
Belostomatidae<br />
Belostoma boscii (Lepeletier & Serville): San Diego de los Baños<br />
Corixidae<br />
Sigara jarmanae Hungfd: Sierra La Güira<br />
Cydnidae<br />
Rhytidoporus indentatus Uhler: Forneguera, Sierra Chiquita<br />
Lygaeidae<br />
Xyonysius basalis (Dallas): Pan de Guajaibón, Forneguera<br />
Cymonimus notabilis (Distant): San Diego de los Baños<br />
Kleidocerys championi (Distant): San Diego de los Baños<br />
Lygaeus pulchelus Fabr.: Pan de Guajaibón, Forneguera, Sierra<br />
Chiquita<br />
Ortholomus jamaicensis Dallas: Sierra La Güira<br />
Oncopeltus fasciatus (Dallas): Forneguera<br />
Paromius longulus (Dallas): Pan de Guajaibón, Sierra Chiquita<br />
Pamphantus vittatus Bruner: Pan de Guajaibón<br />
Mesoveliidae<br />
Mesovelia mulsanti White: Sierra La Güira<br />
Miriidae<br />
Collaria oleosa (Distant): Forneguera<br />
Creontiades rubrinervis (Stal): Pan de Guajaibón<br />
Dagbertus olivaceus (Reuter): Pan de Guajaibón<br />
Jovertus chryselectrus Distant: Cueva Los Portales, Sierra La Güira<br />
Polymerus testaciepes (Stal)): Pan de Guajaibón, Forneguera<br />
Pycnoderes vanduzeei Reuter: Forneguera<br />
Reuteroscopus ornatus (Reuter): San Diego de los Baños<br />
Pentatomidae<br />
Proxys punctulatus (Palisot de Beauvois): Forneguera<br />
Thyanta perditor (Fabricius): Forneguera<br />
Pyrrhocoridae<br />
Dysdercus mimus (Say): Pan de Guajaibón, Forneguera<br />
Reduviidae<br />
Metapterus fraternus (Say): San Diego de los Baños<br />
Ghinallelia sp.: Pan de Guajaibón<br />
Thyreocoridae<br />
Allocoris minuta (Uhler): Forneguera<br />
Scutelleridae<br />
Diolcus variegatus (Herrich-Schaffer): Pan de Guajaibón<br />
HYMENOPTERA<br />
Bethylidae<br />
Scleroderma macrogaster: (Asmead): Cajálbana<br />
Brachonidae<br />
Chelonus sp.: Pan de Guajaibón<br />
Chalcidae<br />
Conura sp. 1: Forneguera<br />
Conura sp. 2: Pan de Guajaibón<br />
Formicidae<br />
Atta insularis Guérin: San Diego de los Baños, Forneguera<br />
Brachymyrmex obscurior Forel: Pan de Guajaibón, Forneguera<br />
Camponotus planatus Roger: Pan de Guajaibón, Ruta Ecológica,<br />
Sierra Chiquita<br />
Leptothorax squamifera (Roger): Pan de Guajaibón<br />
Leptothorax barbouri (Aguayo): Mil Cumbres<br />
Leptothorax iris (Roger): Pan de Guajaibón<br />
Monomorium floricola (Jerdon): Forneguera, Pan de Guajaibón<br />
Paratrechina longicornis (Latrielle): Pan de Guajaibón<br />
Pheidole megacephala (Fabricius): Sierra Chiquita<br />
Pseudomyrmex pazosi Santschi: Forneguera<br />
Pseudomyrmex cubaensis Forel: Pan de Guajaibón<br />
Solenopsis geminata (Fabricius):Forneguera<br />
Solenopsis sp.: Forneguera, Pan de Guajaibón, Sierra Chiquita<br />
Tetramorium bicarinatum (Nylander): Forneguera<br />
Wasmannia auropunctata (Roger): Pan de Guajaibón<br />
Ichneumonidae<br />
Eiphosoma sp.: Forneguera<br />
Scelionidae<br />
Macroteleia sp.: Forneguera<br />
Crabronidae<br />
Trypoxylon subimpressum (Smith):Sierra Chiquita, Ruta Ecológica<br />
COCUYO 15, 2005 21
Agradecimientos.- Agradecemos la colaboración brindada en la<br />
realización de este trabajo a los compañeros del Instituto de<br />
Ecología y Sistemática que participaron en los muestreos, a<br />
Horacio Grillo por la revisión de la nomenclatura de las especies<br />
del orden Heteroptera y J. A. Genaro la identificación de algunos<br />
himenópteros.<br />
REFERENCIAS<br />
Armas, L. F. de; M. M. Hidalgo-Gato; I. Fernández; R.<br />
Rodríguez- León; D. Rodríguez; N. Mestre y col. 2000.<br />
Diversidad de la fauna de invertebrados de la Sierra del Rosario.<br />
Informe Final. Agencia de Medio Ambiente, Depositado en<br />
la Biblioteca del Instituto de Ecología y Sistemática, 123 pp.<br />
Coy A. et al. 2000. Biodiversidad de la Sierra de los Órganos. Informe<br />
Final, Agencia de Medio Ambiente Depositado en la<br />
Biblioteca del Instituto de Ecología y Sistemática, 272pp.<br />
Fernández, I. 2001. Composición taxonómica de los<br />
coleópteros de la Sierra del Rosario, Pinar del Río, Cuba.<br />
Poeyana 481-483: 20-33.<br />
Fernández, I. y A. Lozada. En prensa. Adiciones a la<br />
coleopterofauna presente en el Area Protegida Mil<br />
Cumbres, Pinar del Río, Cuba. Poeyana 473: 13-14.<br />
Hidalgo-Gato, M. M. y R. Rodríguez-León 2000. Los<br />
homópteros (Homoptera: Auchenorrhyncha) de la<br />
cordilera de Guaniguanico, Cuba. Brenesia 54: 51-60.<br />
Rodríguez, D. 2005. Dípteros (Insecta: Diptera) del Area<br />
Protegida “Mil Cumbres”, Pinar del Río, Cuba. Cocuyo<br />
15: 22-24.<br />
Dípteros (Insecta: Diptera) del Area Protegida “Mil<br />
Cumbres”, Pinar del Río, Cuba<br />
Dely Rodríguez Velázquez<br />
Instituto de Ecología y Sistemática, A.P. 8029, Ciudad de La Habana,<br />
10800, Cuba<br />
Las montañas son ecosistemas importantes donde está<br />
representada la ecología compleja e interdependiente de nuestro<br />
planeta. El actual empobrecimiento de la biodiversidad es en<br />
gran parte resultado de la actividad humana y constituye una<br />
grave amenaza para su propio desarrollo. La pérdida de<br />
diversidad biología por eliminación de áreas forestales es uno de<br />
los grandes problemas a resolver y es preciso tomar<br />
urgentemente medidas decisivas para conservar y mantener los<br />
genes, las especies y los ecosistemas con miras al ordenamiento y<br />
la utilización sostenible de los recursos biológicos (Programa 21,<br />
1992). Cuba presenta alrededor de 8 312 especies conocidas de<br />
insectos, pertenecientes a 29 órdenes, donde los grupos<br />
predominantes son los coleópteros (30.6%), lepidópteros<br />
(18.5%), himenópteros (12.9 %) y dípteros (12.0 %), lo cual se<br />
corresponde con el porcentaje de estos grupos a escala mundial.<br />
La diversidad más novedosa y conspicua está apareciendo en los<br />
núcleos montañosos cubanos (Genaro y Tejuca, 1999).<br />
Entre los estudios sobre biodiversidad referidos a los<br />
insectos en Cuba, se encuentran los realizados en<br />
agroecosistemas cañeros (Novoa et al., 1990; 1995), los del<br />
Archipiélago Sabana Camagüey (ICGC, 1990; Alcolado et al.<br />
1999 y Rodríguez-León et al., 2000), en el macizo montañoso<br />
Sagua- Nipe- Baracoa (Hidalgo-Gato et al., 1998; Garcés y<br />
Portuondo, 1998; Viña et al., 1998), el estudio de la<br />
biodiversidad de Cuba (Vales et al., 1998) y más recientemente<br />
estudios en las Sierras del Rosario (Armas et al., 2000), de los<br />
Órganos (Rodríguez y Mestre. 2002) y la Cordillera de<br />
Guaniguanico (Hidalgo-Gato y Rodríguez-León, 2002).<br />
El objetivo de este trabajo es presentar el primer<br />
inventario de los dípteros del Área Protegida “Mil Cumbres”,<br />
por lo que la información que se brinda es el resultado de la<br />
revisión de las colecciones de insectos ubicada en la Colección<br />
Zoológica del Instituto de Ecología y Sistemática; de la<br />
bibliografía, (Alayo y García, 1983; Alayo y Garcés, 1989;<br />
Gregor et al., 1973; Crúz y García, 1974; González-Broche y<br />
García, 1981; Thompson, 1981; Garcés y Rodríguez, 1998) y<br />
de muestreos realizados en los años 2001 y 2002, con redes<br />
entomológicas y trampas de Malaise, en las siguientes<br />
localidades: Forneguera, Sierra Chiquita y Pan de Guajaibón.<br />
Los ejemplares se identificaron utilizando las claves<br />
propuestas por Bates (1934), Aczél (1955), Borror et al. (1976);<br />
McAlpine et al. (1981), Thompson (1981), Alayo y Garcés<br />
(1989), Scarbrough y Knutson (1989) y Foote et al. (1993);<br />
comparando con ejemplares ubicados en la colección<br />
entomológica del Instituto de Ecología y Sistemática y con<br />
métodos especializados de disección de genitales internos.<br />
Para el Area Protegida “Mil Cumbres” se registraron 53<br />
especies de dípteros, pertenecientes a 15 familias. De las especies<br />
registradas, tres fueron endémicas: Udamopyga cubana,<br />
Stenotabanus melliflus y Tabanus bifloccus. Tomosvaryella spangleris<br />
constituyó un registro nuevo para Cuba; Trupanea ageratae y<br />
Xanthaciura tetraspina ampliaron su rango de distribución, la<br />
primera solamente se conocía del Cañón del río Santa Cruz<br />
(Pinar del Río) y la segunda para Cojímar (Ciudad de La Habana)<br />
(Rodríguez et al., 2001).<br />
Lista de Diptera del Area Protegida “Mil Cumbres ”. * especies<br />
endémicas de Cuba. ** nuevos registros para Cuba.<br />
Asilidae<br />
Atomosia sp.<br />
Efferia sp.<br />
Ommatius sp.<br />
Bombyliidae<br />
Ligyra cerberus (Fabricius, 1794)<br />
Villa lucifer (Fabricius, 1775)<br />
Villa sp.<br />
Ceratopogonidae<br />
Culicoides sp.<br />
Culicidae<br />
Anopheles grabhamii Theobald<br />
A. albimanus Wiedemann, 1821<br />
Culex carcinophilus Dyar y Knad, 1906<br />
C. corniger Theobald, 1903<br />
C. I nicaroensis Duret, 1967<br />
22 COCUYO 15, 2005
C. janitor Theobald 1903<br />
Ochlerotatus mediovittatus (Coquillet, 1906)<br />
O. taeniorhynchus (Wiedemann, 1821)<br />
Toxorhynchites portoricensis (Roeder, 1885)<br />
Urotaenia sapphirina (Osten-Sacken, 1868)<br />
Dolichopodidae<br />
Condylustylus sp.<br />
Ephidridae<br />
Notiphila fulvimana Cresson, 1917<br />
Lauxaniidae<br />
Physegenua sp.<br />
Otitidea<br />
Euxesta sp.<br />
Pipunculidea<br />
Tomosvaryella spangleris Scarbrough y Knutson, 1989 **<br />
Sarcophagidae<br />
Helicobia bethae Dodge, 1965<br />
Helicobia rabbí Dodge, 1965<br />
Udamopyga cubana López, 1940 *<br />
Simulidae<br />
Psilopelmia sp.<br />
Stratiomyidae<br />
Ptecticus cyanifrons (Rondani, 1848)<br />
Peltina sp.<br />
Syrphidae<br />
Copestylum apicale (Loew, 1866)<br />
Palpada sp.<br />
Pseudodoros clavatus (Fabr., 1794)<br />
Toxomerus arcifer (Loew, 1866)<br />
T. floralis (Fabricius, 1798)<br />
T. pictus Macquart, 1842<br />
Toxomerus sp.<br />
Tabanidae<br />
Chrysops variegatus (De Geer, 1776)<br />
Lepiselaga crassipes (Fabr., 1805)<br />
Stenotabanus melliflus (Bequaert, 1940) *<br />
Tabanus bifloccus Hine, 1925 *<br />
T. dorsifloccus Szilády, 1926 *<br />
Tephritidae<br />
Acrotaenia testidinea (Loew, 1873)<br />
Anastrepha obliqua (Macquart, 1835)<br />
Dioxina sororcula (Wiedemann, 1830)<br />
Dioxina sp.<br />
Dyseuaresta sp.<br />
Tetreuaresta obscuriventris (Loew, 1873)<br />
Tetreuaresta sp.<br />
Tomoplagia sp.<br />
Toxotrypana curvicauda Gerstaecker, 1860<br />
Trupanea agérate Benjamín, 1934<br />
Xanthaciura insecta (Loew, 1862)<br />
X. tetraspina (Phillips, 1923)<br />
Xanthaciura sp.<br />
Agradecimientos.-Agradezco a Raúl González Broche la<br />
información referida a Culicidae, lo que contribuyó a enriquecer<br />
los resultados.<br />
REFERENCIAS<br />
Aczél, M. L. 1955. Fruit flies of the genus Tomoplagia Coquillett<br />
(Diptera: Tephritidae). Proc. U. S. Nat. Mus., Washington,<br />
104 (3343): 321-411.<br />
Alayo, P. e I. García. 1983. Lista anotada de los dípteros de Cuba. Ed.<br />
Cient.-Técn. La Habana, 201 pp.<br />
Alayo, P. y G. Garcés. 1989. Introducción al estudio del orden Diptera<br />
en Cuba. Editorial Oriente. Santiago de Cuba. 224 pp.<br />
Alcolado, P. et al. 1999. Protección de la Biodiversidad y<br />
establecimiento de un desarrollo sostenible en el ecosistema<br />
Sabana-Camagüey. Ed. CESYTA España. 221pp.<br />
Armas, L. F. et al. 2000. Diversidad de invertebrados de la Sierra<br />
del Rosario. Informe Final. Programa de Colecciones de la<br />
Agencia de Medio Ambiente.<br />
Bates, M. 1934. Notes on American Trypetidae. IV. Acrotaenia<br />
and similar genera. Rev. Entomol., 4:1-17.<br />
Borror, D. J.; P. M. De Long y C. H. Triplehorn. 1976. An<br />
introduction to the study of insect. Ho H, Rinehart & Winston,<br />
Nueva York, 4ta ed., pp. 536-607.<br />
Cruz, J. e I. García. 1974. Los Tábanos de Cuba. Poeyana 125:1-9.<br />
Foote, R. H.; F. L. Blanc y A. L. Norrbom. 1993. Handbook of the<br />
fruit flies (Diptera: Tephritidae) of America north of Mexico.<br />
Cornell Univ. Press, Ithaca Nueva Cork. 571 pp.<br />
Garcés, G. y D. Rodríguez. 1998. Lista de los sírfidos (Diptera:<br />
Syrphidae) de Cuba. Cocuyo 7:7-8.<br />
Garcés G. y E. Portuondo, 1998. Insecta En: Informe Final<br />
Proyecto Biodiversidad del macizo montañoso Nipe-<br />
Sagua-Baracoa., CITMA<br />
García, I. 1977. Fauna cubana de mosquitos y sus criaderos típicos. Ed.<br />
Academia. Ciudad de La Habana. 106 pp.<br />
Genaro, J. A. y A. E. Tejuca. 1999. Datos cuantitativos,<br />
endemismo y estado del conocimiento en insectos. Cocuyo<br />
8:24-28.<br />
González Broche, R. e I. García. 1981. Estudio y distribución de la<br />
familia Phlebotomidae. Dos especies y una subespecie nuevas para<br />
Cuba. Ed. Científico-Técnica. La Habana. 64 pp.<br />
Gregor, F.; V. Cerný y J. de la Crúz. 1973. On the avian<br />
Hippoboscidae (Diptera) of Cuba. Folia Parasitol., (Praga)<br />
20:183-187.<br />
Hidalgo-Gato, M.; R. Rodríguez-León y L. F. de Armas. 1998.<br />
Homópteros auquenorrincos (Homoptera:<br />
Auchenorrhyncha) de la Altiplanicie Sagua-Baracoa,<br />
provincia de Holguín. Poeyana 468:1-7<br />
Hidalgo-Gato, M. y R. Rodríguez-León. 2002. Los homópteros<br />
auquenorrincos (Homoptera: Auchenorrhyncha) de la<br />
Cordillera de Guaniguanico, Cuba. Brenisia 54:51-60.<br />
ICGC 1990: Estudio de los grupos insulares y zonas litorales del<br />
Archipiélago cubano con fines turísticos. Cayo Guillermo, Coco y<br />
Paredón Grande. Ed. Científico-Técnica. La Habana. 181pp.<br />
McAlpine J. F.; B. V. Peterson; G. E. Shewell; H. J. Teskey; J. R.<br />
Vockeroth et al., 1981. Manual of Neartic Diptera. Res Agri.<br />
Canada, Vol. 1 Monograph No. 27. 674p.<br />
Novoa, N.; M. M. Hidalgo-Gato; I. Fernández; R. Rodríguez-<br />
León; L. M. Hernández. et al., 1995. Taxonomía y Ecología<br />
de los principales grupos animales en la Caña de Azúcar y la<br />
vegetación circundante al cultivo. Proyecto de investigación.<br />
Informe Final, depositado en el Instituto de Ecología y<br />
COCUYO 15, 2005 23
Sistemática, CITMA.<br />
Novoa, N.; R. Rodríguez-León; M. M. et al., 1990. Entomofauna<br />
de la caña de azúcar. Proyecto de investigación. Informe<br />
Final, depositado en el Instituto de Ecología y Sistemática,<br />
CITMA<br />
Programa 21. 1992. Informe de la Conferencia de las Naciones<br />
Unidas sobre Medio Ambiente y Desarrollo. 503 pp.<br />
Rodríguez, D.; A. M. Fernández y V. Hernández. 2001. Catálogo<br />
de los tefrítidos (Diptera: Tephritidae) de Cuba. Fitosanidad<br />
5(4):7-14.<br />
Rodríguez, D. y N. Mestre. 2002. Collembola e Insecta<br />
(Coleoptera, Dermaptera, Dictyoptera, Mantodea,<br />
Diptera, Hymenoptera) de la Sierra de los Organos. Cocuyo.<br />
12:6-10<br />
Rodríguez-León R et al., 2000. Presencia de insectos en 12 cayos<br />
del Archipiélago de Sabana Camagüey. Poeyana 476-480:23-<br />
28<br />
Scarbrough, A.G. y L. V. Knutson. 1989. Asilidae, Bombyliidae,<br />
Conopidae and Pipunculidae (Diptera) of Dominicana,<br />
West Indies. Florida Entomologist. 72(3):519-537.<br />
Thompson, F. C. 1981. The flower flies of the West of Indies<br />
(Diptera: Syrphidae). Entomol. Soc. Washington 9:1-200.<br />
Vales, M; A. Alvarez; L. Montes y H. Ferrás (Eds). 1998. Estudio<br />
Nacional de Biodiversidad. Proyecto GEF/PNUMA. CITMA.<br />
La Habana.<br />
Viña, N. et al., 1998. Biodiversidad del macizo montañoso Nipe-<br />
Sagua-Baracoa. Informe Final Proyecto de Desarrollo<br />
Sostenible de la Montaña. BIOECO. CITMA.<br />
Registros de plagas en los almacenes de Fondos de<br />
Exhibición del Museo Nacional de Historia Natural de<br />
Cuba<br />
Idia A. Vivar Rodríguez, Ana E. Tejuca y Raymil Fuentes García<br />
Museo Nacional de Historia Natural, Obispo no. 61, Habana Vieja<br />
10100, Cuba. cocuyo@mnhnc.inf.cu<br />
Las colecciones de historia natural sirven como un archivo<br />
histórico de la vida sobre la tierra, y un recurso para estudios de<br />
sistemática, ecología y evolución (Duckworth et al. 1993). Los<br />
especímenes, las notas de campo y otros datos alojados en<br />
museos constituyen un recurso importante para las<br />
investigaciones de conservación y manejo (Miller, 1985).<br />
Desafortunadamente, muchas de las prácticas tradicionales de<br />
museos que intentan preservar las colecciones de historia natural<br />
y los datos asociados -en realidad- causan daños a los ejemplares.<br />
El biodeterioro es una de las causas de deterioro de los<br />
ejemplares. Se refiere a las acciones dañinas de plagas como<br />
insectos, artrópodos, mohos, bacterias, roedores, enzimas y<br />
otros procesos bioquímicos. El crecimiento de moho en una<br />
hoja de herbario o una colección de insectos atacada por<br />
escarabajos derméstidos son dos ejemplos de biodeterioro.<br />
La mayoría de las colecciones de historia natural están<br />
expuestas a daños o a la total destrucción por las plagas. Los<br />
especímenes de estas colecciones pueden sufrir daños rápidos e<br />
irreparables.<br />
Otro problema es el concerniente al daño potencial causado<br />
por los pesticidas químicos en el medio ambiente, por lo que la<br />
fumigación se ha convertido en un problema actual y existen<br />
regulaciones estrictas en el uso de sustancias tóxicas. Son muy<br />
pocos los productos que legalmente se pueden usar en el control<br />
de plagas en los museos, aunque se realiza un esfuerzo por<br />
incrementar el número de pesticidas recomendables, para esto<br />
son necesarias: 1. Identificar pesticidas nuevos o alternativos con<br />
máxima efectividad, que minimicen los daños al hombre, al<br />
medio ambiente y a los objetos de colecciones. 2. Determinar los<br />
efectos de los pesticidas en los objetos y materiales con los cuales<br />
entran en contacto. 4. Determinar el grado de concentración<br />
mínima para cada producto químico usado con resultados<br />
efectivos en la erradicación de las plagas, y como reducir los<br />
niveles de exposición del personal que trabaja directamente con<br />
los especimenes. 5. Explorar varios métodos no químicos y<br />
económicamente factibles, como el control biológico que ha<br />
dado muy buenos resultados en otros campos.<br />
El trabajo se realizó en el almacén de fondos de exhibiciones<br />
del Museo Nacional de Historia Natural, que atesora ejemplares<br />
de aves, reptiles, anfibios y peces. Se comenzó por determinar la<br />
o<br />
variación climática (humedad relativa % y temperatura C)<br />
mediante un termohigrómetro digital modelo 5566. Los<br />
resultados del monitoreo fueron graficados y se determinó el<br />
valor promedio de la humedad relativa y la temperatura para los<br />
horarios de la mañana y de la tarde. Se realizó un análisis “t de<br />
student” para determinar la existencia de diferencias<br />
significativas entre los valores obtenidos.<br />
Para detectar la presencia de plagas se realizaron<br />
inspecciones visuales mensuales del área; aislándose en<br />
cuarentena los ejemplares contaminados para la posterior<br />
identificación taxonómica de las plagas encontradas.<br />
Se evaluó un posible control de las plagas mediante la<br />
aplicación de tres tratamientos consecutivos, en un periodo de<br />
90 días (uno cada mes). Dichos tratamientos consistieron en<br />
2<br />
una gota de 0.5 mm del gel Imidacloprid 2.15% como cebo,<br />
realizándose el conteo de individuos eliminados por la trampa<br />
(Fig. 1). Este producto es de 5ta generación, biodegradable, y<br />
plantea el proveedor, un alto grado de inocuidad al medio<br />
ambiente y al personal que trabaja directamente con los<br />
ejemplares.<br />
2<br />
Fig. 1. Una gota de 0.5 mm del gel Imidacloprid 2.15%<br />
utilizado como cebo, y ejemplo de un punto de control.<br />
24 COCUYO 15, 2005
RESULTADOS Y DISCUSIÓN<br />
Los resultados de la variación de la humedad relativa y la<br />
temperatura para el horario de la mañana y la tarde se encuentran<br />
representados en las Figs. 2 y 3. Se observaron fluctuaciones en<br />
la humedad relativa y la temperatura, siendo las fluctuaciones de<br />
ambos factores de deterioro, aspectos negativos para la<br />
conservación de ejemplares de historia natural.<br />
HUMEDAD RELATIVA %<br />
Fig 2. Variación de la humedad relativa.<br />
El valor promedio de la humedad relativa fue de 53.3 ±<br />
7.8% en el horario de la mañana y 53.0 ± 8.3% en la tarde;<br />
dichos valores se encuentran en el rango de humedad (55 %)<br />
registrado como adecuado para la conservación de colecciones,<br />
sin embargo, la alta desviación estándar calculada, corrobora las<br />
fluctuaciones de humedad que se produjeron en el almacén<br />
TEMPERATURA GRADOS CELSIOS<br />
0,7<br />
0,6<br />
0,5<br />
0,4<br />
0,3<br />
0,2<br />
0,1<br />
0<br />
29<br />
28<br />
27<br />
26<br />
25<br />
24<br />
23<br />
22<br />
1 3 5 7 9 11 13 15<br />
DIAS<br />
1 3 5 7 9 11 13 15<br />
DIAS<br />
Humedad relativa am<br />
Humedad relativa pm<br />
Temp am<br />
Temp. pm<br />
Con relación a la temperatura, el valor promedio para el<br />
horario de la mañana fue de 26.8 ± 1ºC y el del horario de la tarde<br />
26.7 ± 1ºC, no existieron diferencias significativas entre las<br />
temperaturas para los dos horarios del día, pero ambos fueron<br />
significativamente superiores a 18° C, necesarios para la<br />
adecuada conservación. Se determinó que aunque también<br />
existieron fluctuaciones en la temperatura, estas fueron menos<br />
pronunciadas que en el caso de la humedad relativa<br />
Respecto al biodeterioro, y como resultado del análisis de las<br />
condiciones climáticas anteriormente señaladas, se observó en<br />
las inspecciones visuales realizadas, que cuando existió presencia<br />
de plagas, se encuentran en las partes óseas de los ejemplares un<br />
polvillo amarillento, lo que puede ser considerado como un<br />
índice de infestación.<br />
Apoyados en nuestra observación visual, en el almacén de<br />
fondos de exhibición se detectó la presencia de dos tipos de<br />
plagas, y se comprobó que las mismas comieron, perforaron,<br />
hicieron túneles y cortes fundamentalmente en las patas y picos<br />
de los ejemplares de aves. Las plagas identificadas fueron<br />
socópteros y el escarabajo Lasiderma serricorne (Figs. 4 y 5). Ambas<br />
plagas, sobre todo los socópteros, fueron muy difíciles de<br />
controlar, por ser resistentes a diferentes tratamientos.<br />
Durante los tres controles<br />
realizados se pudo detectar que casi<br />
todos los ejemplares se encontraron<br />
infestados por socópteros. En el<br />
caso del escarabajo L. serricorne (Fig.<br />
5) solo se le detectó en algunos<br />
ejemplares, principalmente de aves.<br />
Como se observa en la Tabla 1,<br />
cada cebo atrapó un promedio de<br />
seis individuos en el primer control,<br />
25 en el segundo control y cuatro<br />
Fig. 4. Ejemplo de una<br />
especie de socóptero<br />
(Trogium pulsatorium, tomado<br />
de Edwards et. al, 1980).<br />
ejemplares en el tercero, como<br />
promedio. Este resultado tiene una<br />
gran importancia, ya que de forma<br />
preliminar podemos observar,<br />
aumento del número de individuos<br />
muertos hasta el segundo control y<br />
una disminución en el tercero, por debajo del promedio<br />
obtenido en el primer control. Este resultado puede observarse<br />
mejor en la Fig. 6.<br />
Fig 3. Variación de la temperatura.<br />
durante el experimento. También se comprobó que no<br />
existieron diferencias significativas entre los valores promedios<br />
obtenidos en los horarios del día.<br />
Fig.5. Lasioderma serricorne (tomado de Edwards et. al, 1980).<br />
De corroborarse, podríamos contar con un tratamiento<br />
efectivo para este tipo de plagas y cuyas consecuencias para los<br />
ejemplares son despreciables ya que no se aplica directamente<br />
en los mismos.<br />
COCUYO 15, 2005 25
Tabla 1. Resultados de los métodos de control utilizados.<br />
Puntos de<br />
control<br />
Primer<br />
control<br />
Segundo<br />
control<br />
Tercer<br />
control<br />
Individuos muertos<br />
1. 1 32 6<br />
2. 0 32 5<br />
3. 1 17 0<br />
4. 5 11 4<br />
5. 5 23 4<br />
6. 10 34 6<br />
7. 5 35 2<br />
8. 10 19 0<br />
9. 5 28 1<br />
10. 7 19 3<br />
11. 12 52 4<br />
12. 3 4 3<br />
13. 3 44 4<br />
14. 21 26 8<br />
15. 4 33 4<br />
16. 4 4 4<br />
17. 0 19 5<br />
18. 0 19 3<br />
19. 0 21 7<br />
20. 0 32 13<br />
Promedio 6 25 4<br />
Conclusiones. Debido a problemas en el control de las<br />
condiciones ambientales de los almacenes de fondos de<br />
exhibición (hermetización de los techos, acondicionador de aire<br />
deficiente y hacinamiento de los ejemplares) se detectó la<br />
presencia de plagas: Lasioderma serricorne y sp no identificada<br />
(Psocoptera), que han afectado la integridad de ejemplares<br />
fundamentalmente de aves, que requirieron restauración y en<br />
algunos casos darle de baja como ejemplar de exhibición, e<br />
incorporar sus huesos al grupo de Geología y Paleontología.<br />
Preliminarmente se pudo comprobar que el gel,<br />
Imidacloprid 2.15% como cebo, es capaz de controlar las plagas<br />
detectadas en los ejemplares de historia natural del Almacén de<br />
Fondos de Exhibición.<br />
Recomendaciones. El producto utilizado provoca la muerte de<br />
las plagas que se encuentran en los alrededores del cebo; se<br />
deben continuar los estudios para determinar el grado de<br />
control, así como la residualidad del producto activo.<br />
REFERENCIAS<br />
Duckworth, W.D.; H. H. Genoways y C.L. Rose (eds.). 1993.<br />
Preserving natural science collection: chronicle of our<br />
environmental heritage. National Institute for the<br />
Conservation of Cultural Property, Inc. Washington D.C.<br />
iii+140 páginas.<br />
Edwards S. R. et. al. 1980. Pest control in museums: a status<br />
report (1980) by the Association of Systematics Collections.<br />
Museo de Historia Natural, Universidad de Kansas.<br />
Jessup W. C. 1995 Pest Management. Storage of Natural<br />
History Collections: A preventive Conservation Approach.<br />
Pp: 211-220. Eds. Carolyn L. Rose, Catharine A. Hawks,<br />
Hungh H. Genoways. Society for the Preservation of<br />
Natural History Collections. Vol I. Universidad de Iowa,<br />
Iowa.<br />
Jeremy F. J. 1995 Pest Monitoring Case Study: A preventive<br />
Conservation Approach. Pp 221-231. Ed. Carolyn L. Rose,<br />
Catharine A. Hawks, Hungh H. Genoways. Society for the<br />
Preservation of Natural History Collections. Vol I.<br />
Universidad de Iowa, Iowa.<br />
Miller, E.H. (ed.). 1985. Museum collection: their roles and<br />
future in biological research. British Columbia Provincial<br />
Museum, Occasional Papers Series 25.<br />
Simmons J.E. 1999. Colecciones de historia natural:<br />
almacenamiento de colecciones y datos a largo plazo. Apoyo<br />
9: 2.<br />
White R.E. 1964. How to distinguish between the cigarette and<br />
drug-store beetles. Entomology section. Entomology Circular<br />
24: 1-37.<br />
No. individuos muertos<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
Control 1<br />
Control 2<br />
Control 3<br />
1 5 9 13 17<br />
Cebo<br />
Fig 6. Individuos muertos por cebo.<br />
26 COCUYO 15, 2005
Moluscos del litoral de la bahía de Cienfuegos, Cuba<br />
Lisbet Díaz Asencio, lisbet@ceac.perla.inf.cu*, Miguel Gómez<br />
Batista, miguel@ceac.perla.inf.cu, Raúl Fernández Garcés,<br />
rfg@ceagrn.perla.inf.cu y Ángel Moreira Gonzáles,<br />
angel@ceacgrn.perla.inf.cu<br />
* Autor para la correspondencia<br />
Centro de Estudios Ambientales de Cienfuegos, Carretera a Castillo<br />
de Jagua, Km 1 ½, CP: 59350, Ciudad Nuclear, Cienfuegos, Cuba<br />
RESUMEN. Se estudió la composición de especies de moluscos en el<br />
litoral de la bahía de Cienfuegos, Cuba, durante el año 2003. Fueron<br />
identificadas 54 especies, de ellas 3 pertenecieron a la clase<br />
Polyplacophora, 10 a la clase Bivalvia y 41 a Gastropoda. Se estudió la<br />
similitud entre estaciones y especies haciendo uso del análisis de<br />
clasificación numérica (Cluster). La matriz de agrupación directa<br />
mostró una tendencia a la formación de dos grupos principales,<br />
segregándose en el mismo las estaciones ubicadas en los lóbulos norte y<br />
sur de la bahía; mientras la matriz inversa mostró una tendencia a la<br />
formación de asociaciones de especies características de zonas del área<br />
de estudio. Estos resultados permitieron plantear la hipótesis de la<br />
existencia de comunidades de moluscos características de cada lóbulo<br />
de la bahía, lo cual refuerza los criterios establecidos de la marcada<br />
diferencia que entre estas dos áreas existe, desde el punto de vista<br />
hidroquímico, geomorfológico y de contaminación antropogénica.<br />
haciendo una división en dos lóbulos naturales, norte y sur, a la<br />
vez que ejerce gran influencia en la circulación de las masas de<br />
agua de la Bahía. Por otra parte, el lóbulo norte está expuesto a<br />
un impacto antropogénico significativo proveniente de la ciudad<br />
de Cienfuegos, del polo industrial y de dos ríos Damují y Salado.<br />
Por su parte, el lóbulo sur está expuesto a un menor impacto,<br />
debido solamente al arrastre de dos ríos Arimao y Caonao<br />
(Villasol et al, 1990).<br />
El área de estudio comprende siete puntos del litoral de la<br />
bahía (Fig. 1). Estos puntos se agruparon en dependencia de su<br />
biotopo característico (Tabla 1).<br />
Biótopo I: litoral rocoso abierto con influencia marina,<br />
presencia de diente de perro y lagunas de salpicaduras.<br />
Biótopo II: zona de transición con gran cantidad de piedras<br />
lisas cubiertas de vegetación en dependencia de la época del año.<br />
Biótopo III: litoral rocoso interior (presencia de diente de<br />
perro romo) con alternancia de pequeños tramos de playa<br />
interior arenofangosa.<br />
Biótopo Vl: zona con predominio de manglares y aportes de<br />
agua dulce provenientes de manantiales.<br />
Biótopo V: playa interior arenofangosa con presencia de<br />
mangles por tramos.<br />
No fue posible emplear un método de muestreo homogéneo<br />
para todos los puntos que permitiera obtener datos cuantitativos<br />
comparables, por lo que los datos recogidos fueron de presenciaausencia.<br />
En cada punto los mayores esfuerzos de colecta<br />
fueron dedicados al supra y meso litoral, sin embargo, también<br />
La bahía de Cienfuegos constituye un complejo sistema<br />
estuarino ubicado en la región centro sur de Cuba. Por su<br />
importancia como recurso natural ha sido objeto de numerosos<br />
estudios entre los que se destacan los relacionados a la diversidad<br />
biológica que en ella habita. Las macroalgas (Moreira et al, 2003)<br />
así como los peces (González- Sansón et al, 1996) y aves (Lalana y<br />
Ortiz, 1990) han sido estudiados y catalogados, sin embargo no<br />
existe una lista de invertebrados para la bahía. En este sentido lo<br />
que más se acerca son los resultados de un estudio realizado por<br />
Lalana y Ortíz (1992), sobre la fauna asociada al mangle<br />
(Rhizophora mangle) en la Laguna de Guanaroca, y los realizados<br />
por Morales et al, (1993).<br />
N<br />
Río<br />
Damují<br />
7<br />
Río Salado<br />
6<br />
4<br />
Ciudad de<br />
Cienfuegos<br />
5<br />
CUBA<br />
22°0 7’N<br />
Río Caonao<br />
Por esta razón el presente trabajo pretende hacer un estudio<br />
la composición de especies de moluscos en la zona supra y<br />
mesolitoral de la bahía de Cienfuegos.<br />
1<br />
2<br />
3<br />
4<br />
Laguna<br />
Guanaroca<br />
MATERIALES Y MÉTODOS<br />
Área de estudio: La bahía de Cienfuegos es una bahía típica de<br />
2<br />
bolsa con un área superficial de 90 Km y profundidad media de<br />
14 km. Está conectada al Mar Caribe por un estrecho canal de 3<br />
Km de longitud. Un banco rocoso se localiza desde Cayo<br />
Carenas hasta Punta La Cueva a una profundidad de un metro,<br />
Mar<br />
Caribe<br />
80°28 ’ W<br />
Fig 1 Área de estudio. 1: Afueras; 2: Cañón de entrada;<br />
3: La Milpa; 4: Jucaral; 5: Punta La Cueva; 6: Reina;<br />
7: Tetas de Tomasa.<br />
COCUYO 15, 2005 27
fueron registradas especies infaunales sin el empleo de arrastres.<br />
Las especies colectadas fueron examinadas a través de un<br />
microscopio estereo e identificadas con la ayuda de la literatura<br />
especializada (Abbott, 1974; De Jong y Coomans, 1988 y<br />
Espinosa et al, 1994). Se determinó la riqueza de especies, dado<br />
por el número total de especies (S). Para analizar la similitud<br />
entre estaciones se calculó el índice cualitativo de Sorensen<br />
(Sorensen, 1948, tomado de Herrera et al. 1987) y con las<br />
matrices de similitud directa e inversa obtenidas se realizó un<br />
análisis de clasificación numérica (dendrograma), usando el<br />
promedio simple entre grupos como estrategia aglomerativa.<br />
Lista de especies<br />
RESULTADOS<br />
Se identificaron 54 especies, de ellas, 3 pertenecieron a la<br />
clase Polyplacophora, 10 a Bivalvia y 41 a la clase Gastropoda<br />
(Tabla 1). Las familias Littorinidae y Neritidae fueron las mejor<br />
representadas con 11 y ocho especies, respectivamente. Fueron<br />
identificadas 21 familias representadas solo por una especie. Las<br />
especies Crassostrea virginica, Brachidontes domingensis e Isognomon<br />
alatus fueron registradas en todas estaciones de colecta, mientras<br />
que se identificaron 26 especies presentes solo en uno de los<br />
puntos de muestreo, la mayoría de ellas colectadas en Reina y en<br />
las Afueras de la bahía. No se reportan nuevos registros para<br />
Cuba. El mayor valor de riqueza de especies se registró en las<br />
Afueras de la bahía y en el Cañón de entrada con S = 22; en<br />
Jucaral y Punta La Cueva se registró el menor valor con S = 12<br />
(Tabla 1).<br />
Análisis de datos<br />
Los análisis de clasificación permitieron comparar<br />
cualitativamente los puntos de muestreo y sobre todo aquellos<br />
que representan biótopos similares. El dendrograma realizado<br />
con la matriz directa para analizar la similitud entre estaciones,<br />
indicó la formación de dos grupos principales (Fig. 2). El<br />
primero agrupó a las estaciones de La Milpa, Cañón de entrada y<br />
las Afueras (biotopos I, II y III). La estación de las Afueras se<br />
separó de las otras dos, lo cual responde a las diferencias en la<br />
composición de especies. Este biotopo presentó características<br />
distintivas con respecto al resto, y, aunque compartió especies<br />
con las estaciones de La Milpa y el Cañón, presentó algunas que<br />
solamente fueron identificadas en este lugar, como: Fisurella<br />
barbadensis, Cittarium pica, Leucozonia nassa, Lottia leucopleura,<br />
Nodilittorina angustior, Nodilittorina dilatata, Nodilittorina mordax y<br />
Thais deltoidea (Tabla 1). El segundo grupo contiene las restantes<br />
cuatro estaciones (biotopos VI y V) ubicadas todas dentro de la<br />
bahía (Figs. 1 y 2). El elemento unificador en este grupo fueron<br />
las especies Littoraria nebulosa, Melampus coffeus, Melongena<br />
melongena, Neritina virginea Littorina angulifera y Onchidella floridana,<br />
estas dos últimas características de los hábitats de manglares. La<br />
20 40 60 80 100<br />
estación de Reina, aunque muestra características que ameritan<br />
su inclusión dentro del Biótopo V, presentó fuertes rasgos de<br />
antropización dada por la influencia directa de asentamientos<br />
poblacionales e industriales. Fueron identificadas un conjunto<br />
de especies no encontradas en otra estación como: Schwartziella<br />
catesbyana, Anachis crassilabris, Bittiolum varium, Bulla striata,<br />
Haminoea antillarum, Meioceras cornucopiae, Meioceras nittidum (Tabla<br />
1), lo cual probablemente justifique la separación de esta<br />
estación del resto, dentro de este segundo agrupamiento. En el<br />
resto de las estaciones, con excepción de La Milpa, también se<br />
identificaron especies características aunque en menor<br />
proporción.<br />
El dendrograma realizado con la matriz inversa permitió<br />
analizar la similitud entre especies, en dependencia de las<br />
estaciones donde fueron identificadas. Este análisis indicó la<br />
formación de dos grandes agrupamientos (Fig. 3). En el primero<br />
de ellos (A) se agruparon casi exclusivamente las especies<br />
identificadas en los biótopos I, II y III, mientras que el segundo<br />
(B) agrupó a las especies identificadas en los biotopos IV y V, y<br />
cuyas estaciones se ubicaron dentro del área interior de la bahía.<br />
En cada uno de estos grupos resultaron de especial interés los<br />
subgrupos con similitud máxima A1 y B1, que se estuvieron<br />
restringidos a las estaciones de las Afueras y Reina<br />
respectivamente, como se analizó con anterioridad.<br />
Los análisis de similitud reflejaron la marcada diferencia<br />
que existió entre las asociaciones de especies encontradas en las<br />
estaciones dentro de la bahía, y el resto.<br />
DISCUSIÓN<br />
Tetas de Tomasa<br />
Jucaral<br />
Punta La Cueva<br />
Reina<br />
La Milpa<br />
Cañón de entrada<br />
Afueras<br />
Fig. 2 Dendrograma que muestra<br />
la similitud entre estaciones de colecta.<br />
El número de especies registrado fue notablemente inferior<br />
al mencionado por otros autores. Sin embargo, es de destacar<br />
que el mismo no tuvo como objetivo hacer una lista exhaustiva<br />
de la fauna malacológica de la bahía de Cienfuegos, sino<br />
caracterizar la biodiversidad de moluscos en su zona supra y<br />
mesolitoral, por lo que quedó excluída la gran biodiversidad de<br />
organismos infaunales que en ella habita. La naturaleza de este<br />
estudio responde a la necesidad de buscar organismos<br />
indicadores que permitan posteriormente el monitoreo<br />
28 COCUYO 15, 2005
0<br />
20<br />
40<br />
60<br />
80<br />
100<br />
Pedipes mirabilis<br />
Neritina meleagris<br />
Batillaria minima<br />
Angiola lineata<br />
Littoraria tessellata<br />
Puperita tristis<br />
Cittarium pica<br />
Fisurella barbadensis<br />
Leucozonia nassa<br />
Lottia leucopleura<br />
Nodilittorina angustior<br />
Nodilittorina dilatata<br />
Nodilittorina mordax<br />
Thais deltoidea<br />
Chiton viridis<br />
Nerita peloronta<br />
Acanthopleura granulata<br />
Nerita versicolor<br />
Chiton squamosus<br />
Puperita pupa<br />
Nodilittorina mespillum<br />
Nerita tessellata<br />
Cenchritis muricatum<br />
Nodilittorina antoni<br />
Nodilittorina ziczac<br />
Purpura patula<br />
Cerithium atratum<br />
Mytilopsis leucophaeata<br />
Prototaca granulata<br />
Trachicardium muricatum<br />
Phacoides pectinatus<br />
Tagelus divisus<br />
Schwartziella catesbyana<br />
Anachis crassilabris<br />
Bulla striata<br />
Bittiolum varium<br />
Haminoea antillarum<br />
Meioceras cornucopiae<br />
Meioceras nitidum<br />
Mactra fragilis<br />
Melampus coffeus<br />
Melampus monile<br />
Nassarius vibex<br />
Onchidella floridana<br />
Littorina angulifera<br />
Isognomon alatus<br />
Brachidontes domingensis<br />
Crassostrea virginica<br />
Littoraria flava<br />
Littoraria nebulosa<br />
Chione cancellata<br />
Melongena melongena<br />
Nerita fulgurans<br />
Neritina virginea<br />
Fig 3 Dendrograma que muestra la similitud entre las especies.<br />
periódico de este ecosistema.<br />
Moreira et al. (2003) citaron los mayores valores de riqueza<br />
de especies para macroalgas en las estaciones ubicadas en el<br />
lóbulo sur de la bahía. De modo general el presente estudio<br />
muestra resultados similares: los mayores valores de riqueza se<br />
encontraron en las estaciones más próximas al mar abierto,<br />
mientras que los menores, en estaciones ubicadas dentro de la<br />
bahía y que por tanto reciben una mayor influencia de los<br />
residuales provenientes de la ciudad.<br />
Para la bahía de Cienfuegos existen criterios establecidos de<br />
la marcada diferencia que existe entre los lóbulos norte y sur,<br />
desde el punto de vista hidroquímico, geomorfológico y de<br />
contaminación antropogénica (Villasol et al., 1990). El análisis de<br />
similitud por estaciones también refleja esta diferencia: las<br />
estaciones ubicadas dentro y fuera del área de la bahía se separan<br />
notablemente entre sí, así como también los agrupamientos de<br />
especies responden coherentemente a esta división. Estos<br />
resultados permiten plantear la hipótesis de la existencia de<br />
comunidades de moluscos características de cada zona de la<br />
bahía. Esta misma tendencia fue encontrada para las macroalgas<br />
en la bahía de Cienfuegos por Moreira et al (2003).<br />
El análisis de similitud por especies evidenció la existencia<br />
de asociaciones de especies, hecho que resultó interesante ya que<br />
no solo podría definir a una única especie como indicadora de un<br />
proceso, sino a un conjunto de ellas, y a las complejas relaciones<br />
que se establecen. Este fenómeno pudiera ser utilizado en el<br />
monitoreo ecológico de la bahía después de haber realizado un<br />
estudio de la variación temporal de las mismas.<br />
Agradecimientos.- Queremos agradecer al Dr. José Espinosa del<br />
Instituto de Oceanología, Cuba, la exhaustiva revisión de este<br />
trabajo.<br />
COCUYO 15, 2005 29
Tabla 1. Especies de moluscos de acuerdo a los biótopos en cada localidad.<br />
Especies<br />
Clase Polyplacophora<br />
Afueras<br />
Biotopos<br />
I II III IV V<br />
Cañón Caÿñ ÿó n<br />
entrada<br />
Chiton squamosus Linneo L n ÿé1764 x x<br />
C. viridis Spengler, 1797 x x<br />
Acanthopleura granulata (Gmelin, 1791) x x<br />
Clase Gastropoda<br />
Fisurella barbadensis (Gmelin, 1791)<br />
Lottia leucopleura (Gmelin, 1791)<br />
Cittarium pica (L (Linneo)<br />
n ÿé,1758) x<br />
x<br />
x<br />
La Milpa<br />
Jucaral<br />
Punta<br />
La Cueva<br />
Nerita fulgurans Gmelin, 1791 x x x x x<br />
N. peloronta L Linneo<br />
n ÿé,1758 x x<br />
N. tessellata Gmelin, 1791 x x<br />
N. versicolor Gmelin, 1791 x x<br />
Puperita pupa (L (Linneo)<br />
n ÿé,1758) x x<br />
P. tristis (Orbigny, 1842) x<br />
Neritina meleagris Lamark, 1822 x x<br />
N. virginea (L (Linneo)<br />
n ÿé,1758) x x x x<br />
Littorina angulifera (Lamark, 1822) x x<br />
Cenchritis muricatum (Linneo)<br />
n ÿé,1758) x x x<br />
Littoraria flava King y Broderrip, 1832 x x x x<br />
L. nebulosa (Lamark, 1822) x x x x x<br />
L. tessellata (Philippi, 1847) x<br />
Nodilittorina mespillum (Mühlfeld, ÿühlfeld,1824) 1824) x x<br />
N. angustior (Mörch, ÿörch,1876) 1876)<br />
x<br />
N. dilatata (Orbigny, 1842) x<br />
N. ziczac (Gmelin, 1791) x x x<br />
N. antoni (Philippi, 1846) x x x<br />
N. mordax (Bandel y Kadolsky, 1982) x<br />
Schwartziella catesbyana (Orbigny, 1842)<br />
Meioceras cornucopiae (Carpenter, 1858)<br />
M. nitidum (Stimpson, 1851) x<br />
Angiola lineata (da Costa, 1778)<br />
Cerithium atratum (Born, 1778)<br />
Bittiolum varium (Pfeiffer, 1840)<br />
Batillaria minima (Gmelin, 1791)<br />
Thais deltoidea (Lamarck, 1822)<br />
Purpura patula (Linneo)<br />
n ÿé,1758) x x x<br />
Anachis crassilabris (Reeve, 1859)<br />
Nassarius vibex (Say, 1822)<br />
x<br />
x<br />
x<br />
Melongena melongena (Linneo)<br />
n ÿé,1758) x x x<br />
Leucozonia nassa (Gmelin, 1791)<br />
Bulla striata (Gmelin, 1791)<br />
x<br />
x<br />
Reina<br />
x<br />
x<br />
x<br />
x<br />
x<br />
Tetas<br />
Tomasa<br />
x<br />
30 COCUYO 15, 2005
Tabla 1. Especies de moluscos de acuerdo a los biótopos en cada localidad. Continuación...<br />
Haminoea antillarum (Orbigny, 1842)<br />
Onchidella floridana (Dall, 1885) x x<br />
Melampus coffeus (L (Linneo)<br />
n ÿé,1758) x x x<br />
M. monile Bruguiere, 1789 x<br />
Pedipes mirabilis (Mühlfeld, ÿühlfeld,1816) 1816)<br />
x x<br />
Clase Bivalvia<br />
Especies<br />
Brachidontes domingensis (Lamarck, 1819) x x x x x x x<br />
Isognomon alatus (Gmelin, 1791) x x x x x x x<br />
Crassostrea virginica (Gmelin, 1791) x x x x x x x<br />
Phacoides pectinatus (Gmelin, 1791) x x<br />
Trachicardium muricatum (Linneo)<br />
n ÿé,1758) x<br />
Mactra fragilis Gmelin, 1791<br />
Tagelus divisus (Spengler, 1794) x x x<br />
Chione cancellata (Linneo)<br />
n ÿé,1767) x x<br />
Protothaca granulata (Gmelin, 1791)<br />
Mytilopsis leucophaeata (Conrad, 1831)<br />
Afueras<br />
Biotopos<br />
I II III IV V<br />
Cañón Caÿñ ÿó n<br />
entrada<br />
La Milpa<br />
N Número ÿúmerodespecies(S) de especies (S)<br />
22 22 12 12 12 17 17<br />
Jucaral<br />
x<br />
Punta<br />
La Cueva<br />
x<br />
x<br />
Reina<br />
x<br />
Tetas<br />
Tomasa<br />
REFERENCIAS<br />
nd<br />
Abbott, R. 1974. American Seashells. 2 edition Van Nostrand<br />
Reinhold: New York. 663 pp<br />
De Jong, K.M. y H.E. Coomans 1988. Marine Gastropod from<br />
Curacao, Aruba, and Bonaire. E.J. Brill: Leiden. 261 pp.<br />
Espinosa, J.; R. Fernández y E. Rolan 1994. Catálogo actualizado de<br />
los moluscos marinos actuales de Cuba. Sociedad Española de<br />
Malacología. 90 pp.<br />
González-Sansón G.; A. Bosch; E. Guevara y C. Aguilar 1996.<br />
Asociaciones de peces de los sitios de pesca de camarón en la<br />
Bahía de Cienfuegos, Cuba. Rev. Investigaciones Marinas 17 (2-3):<br />
133-141.<br />
Herrera, A.; R. del Valle y N. del Castillo 1987. Aplicación de<br />
métodos de clasificación numérica en el estudio ecológico del<br />
litoral rocoso. Reporte de Investigación, Instituto de Oceanología,<br />
Academia de Ciencias de Cuba, 70: 1-15.<br />
Lalana R. y M. Ortiz 1990. Lista de aves de la Laguna de<br />
Guanaroca, Provincia de Cienfuegos, Cuba. Rev. Investigaciones<br />
Marinas 11 (2): 169 - 173.<br />
Lalana, R . y M. Ortiz 1992. Fauna asociada a mangles de la<br />
Laguna de Guanaroca, Provincia de Cienfuegos, Cuba. Rev.<br />
Investigaciones Marinas 13 (3): 205 214<br />
Morales C. R; G. M. E. Castellanos; R. M. Lara; V. G Acevedo.; P.<br />
J Pardo y N. Vidal 1993. Resultados parciales del monitoreo<br />
permanente en la Bahía de Cienfuegos en el período<br />
correspondiente a 1990-1992. ACC C.M.I.C.T Cienfuegos 19<br />
pp.<br />
Moreira, A. R.; M. Gómez; A.M. Suárez; A.R. León y M.E.<br />
Castellanos 2003. Variación de la composición y abundancia<br />
de macroalgas en la Bahía de Cienfuegos, Cuba. Rev. Invest.<br />
Marinas 24(2): 83-94.<br />
Sorensen, T. 1948. A method of stabilizing groups of equivalent<br />
amplitude in plant sociology based on similarity of species<br />
content and its application of the vegetation of Danish<br />
commons. Biol.Skr., 5(4):1-34.<br />
Villasol A. N; N. Jaime; R. Mederos; H. Quintana; J. Martínez; I.<br />
Fernández y A. I. Tur. 1990. Estudios de la contaminación en<br />
la Bahía de Cienfuegos. Informe Final, Ministerio del<br />
Transporte, Instituto de Investigaciones del Transporte. 66 pp<br />
COCUYO 15, 2005 31
OBITUARIO<br />
Ha muerto Ernst Mayr....<br />
(1904-2005)<br />
En el número anterior de la revista Cocuyo (2004: 14: 32),<br />
nos regocijamos por los 100 años de vida de Ernst Mayr, el<br />
insigne investigador y profesor del Museo de Zoología<br />
Comparativa de la Universidad de Harvard. Lamentablemente<br />
hoy escribo su obituario. El distinguido científico y naturalista<br />
falleció en su apartamento de Bedford, Massachussets el 3 de<br />
Febrero pasado, después de una breve enfermedad.<br />
Escribía Mayr en la revista Science (Julio 2, 2004, vol. 14:<br />
47) que él era el último superviviente de los protagonistas de la<br />
edad de oro de la Síntesis Evolucionaria. Aquella gesta científica<br />
que interpretó, a la luz de los hallazgos genéticos y taxonómicos,<br />
las teorías evolucionarias expuestas por Charles Darwin, casi, 80<br />
años atrás.<br />
Intentar sintetizar sus<br />
contribuciones es muy difícil:<br />
más de 600 artículos científicos y<br />
25 libros, amén de una constante<br />
obra profesoral; por ello quiero<br />
referirme en este artículo a dos<br />
de sus contribuciones que más<br />
me han impactado: el artículo<br />
US<br />
publicado en 1954, como parte del libro “La evolución como<br />
proceso” , intitulado: “Cambio del medio genético y evolución”;<br />
curiosamente este artículo era el preferido de Mayr (así se lo<br />
expresó, en una de sus últimas entrevistas con la periodista<br />
Christine Bahls), y a su contribución a la epistemología<br />
expresada en varios de sus libros sobre las características de los<br />
eventos únicos en los procesos biológicos (en otras palabras el<br />
valor de la “narración histórica” como herramienta científicoconceptual).<br />
Uno de los problemas que no habían sido abordados por<br />
Darwin ni por consiguientes autores fue la correspondencia<br />
entre el registro fósil y los mecanismos probables de la<br />
especiación en pequeñas poblaciones aisladas, los aparentes<br />
“saltos” en las secuencias evolutivas, los desiguales ritmos<br />
1<br />
evolutivos y la aparición de nuevos “tipos” (nuevos taxones).<br />
Según Mayr, estas poblaciones aisladas y pequeñas no eran<br />
objeto de la atención de los taxónomos (por la dificultad de<br />
acceder a ellas y por constituir los extremos de poblaciones<br />
mayores), pero que en éstas operaban los procesos de<br />
reconstrucción genética y de reajustes ecológicos tales como<br />
para iniciar una fuerte divergencia de sus ancestros y ocupar<br />
nuevos horizontes adaptativos que los diferenciaban en<br />
diferentes grados, sólo en ese momento eran objeto de la<br />
atención o es que comenzaban a aparecer sus fósiles respectivos;<br />
encontrando el paleontólogo una población numerosa y<br />
extendida que ha sido reemplazada por otra igualmente<br />
numerosa y extendida y bien diferente a la primera.<br />
u labor contribuyó a la revolución conceptual que<br />
encabezó la síntesis evolucionista moderna de la genética<br />
mendeliana y la evolución del darwinismo, así como el<br />
desarrollo del concepto de especies biológicas entre otros<br />
aportes...<br />
Según el modelo propuesto por Mayr (1954), la evolución<br />
avanza con lentitud en las poblaciones muy grandes, y los<br />
cambios evolutivos más rápidos se dan en las poblaciones<br />
2<br />
pequeñas y aisladas periféricamente (que son las fundadoras) .<br />
Expresado en otros términos, esto parece indicar que las<br />
especies con poblaciones grandes son estables, y que las<br />
poblaciones fundadoras, pequeñas pueden carecer de esta<br />
estabilidad, lo que les permite cambiar rápidamente su fenotipo<br />
mediante una rápida reestructuración genética. Eldredge y<br />
Gould (1972) que introdujeron el concepto de “equilibrio<br />
puntuado”, aceptaron este modelo y propusieron que el<br />
estatismo de las especies populosas puede mantenerse por<br />
millones de años. Este modelo de especiación geográfica fue,<br />
más tarde llamado, por Mayr (1982) “especiación peripátrica”,<br />
porque se desarrolla en la periferia de los límites distribucionales<br />
de las especies parentales, con grandes poblaciones (los<br />
fundadores provienen de la periferia de la especie parental). La<br />
revolución genética que se produce durante la especiación<br />
peripátrica sería, en palabras de Mayr, “un proceso poblacional<br />
gradual aunque enormemente acelerado”. Presumiblemente, el<br />
evento más importante que ocurre en muchos casos de<br />
especiación peripátrica es la<br />
destrucción de la cohesión<br />
previamente existente en el<br />
genotipo y su reemplazo por<br />
un nuevo sistema balanceado.<br />
Igualmente es muy probable<br />
que la adquisición de un nuevo<br />
sistema homeostático facilite<br />
nuevos cambios evolutivos y<br />
esplique el origen de nuevos taxones superiores y de novedades<br />
evolutivas.<br />
Esta explicación, basada en el pensamiento poblacional<br />
introducido por Mayr y Dobzshansky da al traste con las<br />
explicaciones tipológicas (basadas en el esencialismo platoniano:<br />
que desechan la variación por lo que los miembros de una<br />
población se consideran réplicas del “tipo”) de R. B.<br />
Goldschmidt (1940) y seguidores que arguían el conceptos de<br />
3 4<br />
macrogénesis y de mutación sistémica con los llamados<br />
El Dr. Mayr en su oficina del Museo de Zoología<br />
Comparativa, Universidad de Harvard, con el autor<br />
en el otoño de 1991.<br />
32 COCUYO 15, 2005
“monstruos esperanzadores”, al tratar de explicar la<br />
discontinuidad en el registro fósil y las diferencias en los<br />
procesos micro y macro evolucionarios.<br />
Hasta el presente se mantienen candentes discusiones<br />
acerca de la completa certeza de este modelo, ya que aún se<br />
desconocen muchos procesos que se desarrollan en las<br />
poblaciones silvestres que afectan sus propios ritmos evolutivos,<br />
como para afirmar la completa identidad entre macro y micro<br />
evolución; además de que éste no es el único mecanismo<br />
plausible para explicar la especiación, pero, sin dudas, fue Mayr<br />
(1954) el primer autor que elaboró un modelo detallado de la<br />
conexión entre especiación, ritmos evolutivos y<br />
macroevolución.<br />
En muchos libros y ensayos, incluso en su último libro<br />
(Mayr, 2004), Ernst trató los aspectos únicos de los fenómenos<br />
biológicos criticando las aproximaciones reduccionistas que<br />
provenían de la física, y los enfoques ultra-axiomáticos<br />
provenientes de la sistemática filogenética, insistiendo en la total<br />
independencia de la biología como ciencia (vea “Así es a<br />
biología” de Mayr (1998), primera edición en español, editorial<br />
Debate, Pensamiento, 326 pp.). Escribía lo siguiente:“Cuando<br />
un biólogo trata de responder a una pregunta acerca de un<br />
fenómeno único, como, ¿por qué no hay colibríes en el Viejo<br />
Mundo o ¿dónde se originó la especie Homo sapiens, no puede<br />
basarse en leyes universales. El biólogo tiene que estudiar todos<br />
los datos conocidos que tengan que ver con el problema en<br />
cuestión, inferir toda clase de consecuencias a partir de<br />
combinaciones de factores reconstruidos, y después intentar<br />
elaborar un argumento que explique los hechos observados del<br />
caso particular. En otras palabras elabora una narración<br />
histórica”.<br />
La naturaleza de este enfoque difiere notablemente de las<br />
explicaciones causa-efecto, que los filósofos clásicos de la ciencia<br />
(procedentes de la lógica, las matemáticas y la física) lo<br />
consideraron totalmente inadmisible. A pesar de ello, autores<br />
recientes han rechazado la estrechez de la opinión clásica,<br />
planteando no sólo que este enfoque histórico-narrativo es<br />
válido sino que es probablemente el único capaz de explicar,<br />
científica y filosóficamente, los fenómenos únicos, vease<br />
Goudge (1961), Hull, (1975) y Nitecki & Nitecki (1972).<br />
Otros dos aspectos que tienen que ver con la<br />
epistemología desarrollada en los trabajos filosóficos de Mayr es<br />
la fortaleza del método científico inductivo (en las propias<br />
narraciones históricas), fuertemente criticado por la escuela<br />
popperiana (esencialmente hipotética-deductiva) y el<br />
falsacionismo (como garantía para la aceptación científica de<br />
cualquier teoría); este último principio parece tener muchos<br />
problemas al comprobar las teorías probabilísticas (así son casi<br />
todas las teorías biológicas); el surgimiento de excepciones a una<br />
teoría probabilística no la refuta necesariamente....<br />
Ernst Mayr ha muerto, como reza en el título de este<br />
modesto obituario, pero su obra, laboriosidad académica y sus<br />
ideas, permanecerán por mucho tiempo en las universidades,<br />
instituciones y eventos en los cuales se discutan los aspectos<br />
fundamentales de la evolución, las especies y la naturaleza de la<br />
investigación biológica.<br />
REFERENCIAS<br />
Eldredge N. & S. J. Gould 1972. In (T.J. Schopf Ed.). Models in<br />
Paleobiology. San Francisco: Freeman, Cooper, pp. 305-332.<br />
Goldschmidt, R.B. 1940. The material basis of Evolution. Yale Univ.<br />
Press, New Haven.<br />
Goudge, T.A. 1961. The ascent of life. Toronto: Univ. Toronto<br />
Press.<br />
Hull, D.L. 1975. Central subjects and historical narratives.<br />
History and Theory 14: 253-274.<br />
Mayr, E. 1954. Change of genetic environment and evolution.<br />
In J. Huxley, A.C. Hardy y E.B. Ford, eds., Evolution as a<br />
Process. Londres: Allen & Uwin, pp. 157-180.<br />
Mayr, E. 1982. In Mechanism of speciation (Rome Symposium,<br />
1982). Nueva York. Allan R. Liss, pp. 1-19.<br />
Mayr, E. 2004. What Makes Biology Unique Considerations on the<br />
autonomy of a scientific discipline. Cambridge University Press,<br />
Nueva York.<br />
NOTAS<br />
1. Simpson (1944, Tempo and Mode in Evolution): estableció la<br />
evolución cuántica, fenómeno vertical (temporal)<br />
considerada dentro de su definición de especie<br />
evolucionaria.<br />
2. El Principio del Fundador estipula que los fundadores de una<br />
nueva colonia (o población) sólo contienen una pequeña<br />
fracción de la variación genética total de la población (o<br />
especie) parental.<br />
3. Macrogénesis: creación de nuevos tipos por “saltos”.<br />
4. Mutación sistémica: es la que reorganizaría el plasma germinal<br />
y diera origen a tipos completamente nuevos de<br />
organismos.<br />
Giraldo Alayón García,<br />
Museo Nacional de Historia Natural, Obispo no. 61,<br />
Habana Vieja 10100, Cuba<br />
BIOCOMENTARIOS<br />
Entre lo histórico y lo lógico: el no-diálogo en la<br />
biogeografía<br />
Jorge L. Fontenla<br />
Museo Nacional de Historia Natural, Obispo No. 61, Habana Vieja<br />
10100, Cuba<br />
libelula@mnhnc.inf.cu<br />
“Y así son los hombres, que cada uno cree que sólo lo que él piensa y ve es la<br />
verdad, y dice en verso y en prosa que no se debe creer sino lo que él cree […]<br />
cuando lo que ha de hacer es estudiar con cariño lo que los hombres han pensado<br />
y hecho, y eso da un gusto grande”<br />
Martí, 1889<br />
“Fallo de predecir no significa fallo de comprender o explicar. Si estuviéramos<br />
seguros que conociéramos las ecuaciones que gobiernan un sistema caótico,<br />
confiaríamos en comprender su comportamiento, incluyendo nuestra incapacidad<br />
de predecir en detalle su comportamiento a largo plazo”.<br />
Kauffman, 1996<br />
COCUYO 15, 2005 33
Quot capita, tot sensus<br />
Una posible aprehensión de la situación<br />
Kauffman (1993) resumió que el siglo XVIII, bajo el<br />
influjo de la revolución Newtoniana, desarrolló ciencias de la<br />
simplicidad organizada; el siglo XIX, vía la mecánica estadística,<br />
se centró en la complejidad desorganizada; mientras que la<br />
ciencia de finales del siglo XX y durante el XXI, deberá<br />
confrontar la complejidad organizada; pero en ninguna otra<br />
ciencia como en la biología -afirma- se hace notar tanto la<br />
ausencia de herramientas conceptuales para esta confrontación.<br />
Ciertamente, la biología está urgida de tales implementos del<br />
intelecto, pues la gran diversidad de fenómenos que estudian los<br />
biólogos necesita de una diversidad correspondiente de<br />
enfoques epistemológicos en la comprensión de la naturaleza<br />
(Keller, 2003). Najmanovich (2002) aseveró que hemos vivido<br />
bajo el hechizo del método, del que recién comenzamos a<br />
despertar. La tarea -enfatiza- requiere la aceptación de la<br />
incompletitud radical de todo conocer y de abrirse a la<br />
multiplicidad de significados sin despeñarse por lo anárquico.<br />
Pienso que el encantamiento del método ejerce su influjo en el<br />
propio corazón de las polémicas que, de una forma u otra, dan<br />
vida al quehacer cotidiano en la teoría (o mejor decir, teorías,<br />
puesto que no es aceptable ¡por suerte! un dogma centralizado y<br />
rígido) evolucionaria en sentido general, y en la biogeografía<br />
histórica en particular.<br />
Crisci (2001) expuso: “La definición de la biogeografía puede<br />
ser sencilla el estudio de la distribución geográfica de los<br />
organismos- pero esta simplicidad esconde la gran complejidad<br />
del tema. La biogeografía trasciende el clásico tópico de las<br />
áreas, involucra una gama de disciplinas científicas que incluyen<br />
la geografía, geología y biología. Nadie que estudie biogeografía<br />
puede evitar el quedar impresionado, o perplejo, por la<br />
diversidad de enfoques del tema”. Esta gran complejidad<br />
provoca que la disciplina se encuentre en un momento de gran<br />
efervescencia, pletórico de enfoques contrapuestos y pugnas<br />
entre sus hacedores. Martin-Piera y San-Martin (1999) aducen<br />
que la biogeografía es “ameboide”, y Ebach y Humphries (2003),<br />
la tildan de una perspectiva cuyos objetivos todavía permanecen<br />
ambiguos, porque el concepto abarca más de un tópico. Es<br />
posible discrepar de este último criterio, y asumir que la<br />
biogeografía tiene una identidad legítima, traslucida en el<br />
estudio de la percepción, determinación, causalidad e historia de<br />
los patrones de distribución espacial de los organismos. Lo<br />
sugestivo es discernir en lo ameboide algo que se infiltra y se<br />
reticula, que se embebe; y asumir en lo ambiguo su comulgar con<br />
lo indeterminado. Todo ello es coherente con la ontología de lo<br />
complejo.<br />
Brooks et al. (2001) se lamentan que las discusiones recientes<br />
sobre diferentes enfoques para el estudio de la disciplina han<br />
estado confundidas, además de ser confusas. Pienso que el<br />
“hechizo del método” es el acreedor de tal confusión. Los<br />
enfoques se ciernen alrededor de la mejor manera de aplicar una<br />
perspectiva epistemológica, el modelo hipotético-deductivorefutacionista,<br />
la vicarianza como causa eficiente generadora de<br />
los patrones biogeográficos y la metodología cladística. Lo que<br />
sigue es una reflexión somera y preliminar de algunos aspectos<br />
de este quehacer. Por ejemplo, Ebach y Humphries (2003)<br />
señalan que la “plétora” de métodos que existen en la<br />
biogeografía es una consecuencia de que la disciplina no es vista<br />
como un campo en su propio derecho. Ello refleja una ausencia<br />
y consenso de lo que constituye la ontología (especificación d e<br />
conceptualización) de la biogeografía, la cual debe estar ligada al<br />
descubrimiento de relaciones históricas entre áreas, según la<br />
metodología cladística. Wilkinson (2003) impugna estos<br />
señalamientos, y manifiesta que el único método no puede ser<br />
cladística y vicarianza. “La biogeografía -precisa- no necesita<br />
una definición rigurosa más allá del enunciado general que es<br />
acerca de la distribución de los organismos en el espacio y el<br />
tiempo. La biogeografía es justamente uno de nuestros intentos<br />
para interpretar el universo. Tópicos complejos son probables de<br />
producir una 'plétora de métodos' con los diferentes métodos<br />
siendo más o menos adecuados para las diferentes cuestiones<br />
dentro del campo general de indagación”.<br />
El estudio de la evolución le da primacía al devenir de la<br />
forma en el tiempo, mientras que la biogeografía resalta la<br />
dimensión espacial en la evolución de la forma. La distribución<br />
de los organismos se estudia a través de la determinación de<br />
patrones espaciales y de la comprensión de los procesos y<br />
eventos probables a partir de los cuales emergen los patrones. Y<br />
para su comprensión es necesaria una perspectiva sistémica y<br />
transdiciplinar. Este conocimiento es muy poco probable de ser<br />
construido con tan sólo algún método formalizado y<br />
estandarizado. Por su parte, Hovenkamp (1997) precisa que la<br />
biogeografía histórica tiene dos objetivos distintivos: (1)<br />
Dilucidar la historia de la Tierra sobre la base de la evidencia<br />
biológica (historia de las áreas). (2) Dilucidar la historia de uno<br />
o más grupos de organismos sobre la base de una teoría<br />
establecida de la historia de las áreas (historia de los taxones).<br />
Las distinciones más generales dentro de la biogeografía<br />
histórica es llamar Biogeografía de la Vicarianza a las prácticas<br />
de: (1) panbiogeografía (2) biogeografía cladística. Ambas<br />
asumen a la vicarianza como principio central en la generación<br />
de los patrones biogeográficos, pero la panbiogeografía genera<br />
hipótesis a partir de la congruencia espacial de las distribuciones<br />
de diferentes taxones. La segunda perspectiva plantea hipótesis<br />
explícitas de relaciones entre áreas, o cladogramas de áreas, a<br />
partir de cladogramas de taxones. Con relación a esta última<br />
perspectiva, Hovenkamp (2001) realiza dos distinciones: (1)<br />
biogeografía cladística (2) biogeografía de la vicarianza. La<br />
primera distinción busca congruencia entre cladogramas<br />
taxonómicos de áreas de varios grupos monofiléticos. El<br />
objetivo es producir un cladograma general de áreas, el cual<br />
ofrece un patrón común de relaciones entre áreas de varios<br />
grupos no relacionados. Los patrones únicos de distribución<br />
histórica son irrelevantes (Llorente, 2000; Ebach, 2001; Ebach y<br />
Humphries, 2002). Ebach y Humphries (2002) consideraron<br />
que esta disciplina constituye “el arte del descubrimiento”. Por<br />
su parte, la biogeografía de la vicarianza no se preocupa por la<br />
extensión de la distribución del taxón, sino con la posición<br />
geográfica de sus disyunciones y con la evidencia que puede ser<br />
extraída de las distribuciones observadas de los taxones y es<br />
permisible el análisis de grupos particulares.<br />
Brooks y McLennan (2001), reconocen que los patrones de<br />
distribución constituyen fenómenos complejos, porque las<br />
34 COCUYO 15, 2005
iotas de las áreas representan un mosaico de procesos de<br />
especiación alopátrida, simpátrida o por aislados periféricos, o<br />
bien exhiben éstasis evolucionario (no especiación ante<br />
fragmentación de las áreas), dispersión y extinción. Van Veller et<br />
al, (2000, 2002) y Van Veller y Brooks (2001) disciernen dos<br />
tipos de pruebas de hipótesis de procesos causales en la<br />
biogeografía de la vicarianza: (1) a priori (2) a posteriori. La<br />
primera asume a priori diferentes procesos para ajustar los datos<br />
a la hipótesis nula. Busca su corroboración. La segunda sólo<br />
asume vicarianza, e invoca a posteriori otros procesos, como<br />
dispersión y extinción, para explicar incongruencias con la<br />
hipótesis nula. Busca su refutación. Ambos enfoques difieren<br />
ontológicamente: el a priori o de historia- de-las-áreas<br />
representa la simplicidad y el a posteriori o de historia-de-lostaxones,<br />
la complejidad. Este último espera (predicción) que la<br />
hipótesis nula sea falsificada (por la complejidad de la<br />
distribución y la ocurrencia de dispersión, éxtasis, extinción o<br />
especiación simpátrida), y suministra soporte objetivo, a<br />
posteriori, para ello.<br />
Con lo anterior en mente, Van Veller et al. (2003) disciernen<br />
entre: (1) biogeografía cladística (2) biogeografía filogenética<br />
(en artículos previos sólo consideraban a la biogeografía de la<br />
vicarianza: (Van Veller et al., 1999, 2000, 2001, 2002). La primera<br />
ajusta todos los elementos de todos los cladogramas<br />
taxonómicos de áreas a un único grupo de relaciones de áreas,<br />
para mantener la singularidad histórica de estas. La biogeografía<br />
filogenética documenta parsimoniosamente el contexto<br />
geográfico de los eventos de especiación. Estos autores asumen<br />
que la evolución es tan contingente y compleja, que es necesario<br />
analizar tanto patrones comunes como únicos, así como<br />
concebir la posibilidad de áreas con historias reticuladas en<br />
relación con las especies que las habitan. Esta perspectiva, la<br />
filogenética, la consideran como ontológica, en contraste con la<br />
perspectiva epistemológica de la biogeografía cladística. Esta<br />
biogeografía filogenética sería equivalente a la biogeografía de la<br />
vicarianza distinguida por Hovemkamp, con enfoque dehistoria-de-los-taxones.<br />
Así concebido, la biogeografía cladística responde al<br />
enfoque de relaciones-de-áreas, y al método analítico a priori,<br />
mientras que la biogeografía filogenética se identifica con el<br />
enfoque de relaciones-de-taxones y a posteriori,<br />
respectivamente. Van Veller et al. (2003) reiteran que la<br />
biogeografía cladística representa el “arte del descubrimiento” y<br />
al modelo inductivo/verificacionista1; mientras que a la<br />
biogeografía filogenética la tildan de “ciencia del<br />
descubrimiento” y representante del modelo hipotéticodeductivo/refutacionista.<br />
El método analítico líder del “arte del<br />
descubrimiento” es el de los componentes (revisiones recientes<br />
en Espinosa et al., 2002, Morrone, 2004). El método<br />
representante de la “ciencia del descubrimiento”*<br />
es el<br />
* El tratamiento “superior” de ciencia versus arte en las explicaciones, es un<br />
reflejo de las dicotomías y de la falta de diálogo entre modos de saber en nuestra<br />
contemporaneidad. No en balde ya se comienza a hablar de “ciencia-arte”,<br />
como dialógica que articula y armoniza lo que une a ambos. Ambas ramas de<br />
saber precisan de lo intuitivo y lo metafórico, de la pasión creativa y de la soltura<br />
en las ideas. ¿Acaso el laureado físico Bohr no expresó que al átomo es mejor<br />
entenderlo cómo poesía, cómo metáfora<br />
“Análisis de la Parsimonia de Brooks” o BPA (Brooks y<br />
McLennan, 2002, 2003; Brooks y Van Veller, 2003; Brooks et al.,<br />
2001; Dowling, 2002; Green et al., 2002; Van Veller et al., 2000,<br />
2001, 2002, 2003).<br />
Kluge (1997, 1999) discurrió que la biología comparativa es<br />
una ciencia de ciclos de descubrimiento y evaluación. Sobre esta<br />
base, Brooks y McLennan (2001) exponen que el BPA, a la vez<br />
que un método de explicación a posteriori, posibilita interpretar la<br />
complejidad real de la biota en un área, ya que constituye un<br />
método basado en descubrimiento. Brooks et al. (2001)<br />
afirmaron que las confusiones en la biogeografía histórica se<br />
deben en parte a la incapacidad de no distinguir entre los<br />
modelos cognitivos mencionados; mientras que otra razón es la<br />
distorsión o subvaloración del BPA, en especial del BPA<br />
secundario (Apéndice). Van Veller et al. (2003) concluyeron que<br />
los únicos métodos o enfoques meritorios para la biogeografía<br />
histórica son los a posteriori. Ebach et al. (2003) declaran que, a<br />
pesar de lo que Van Veller, Brooks y colaboradores han<br />
enfatizado ad nauseam, el BPA es un método también a priori,<br />
porque realiza modificaciones de los datos originales (la<br />
duplicación de áreas) para obtener un nuevo cladograma. En<br />
adición, concluyen que el BPA no descubre ninguna relación<br />
biogeográfica. La duplicación de áreas del BPA ha sido criticada<br />
por algunos autores, pues su sentido biológico no se encuentra<br />
justificado (Van Welzen et al., 2003). Para Ebach et al. (2003) la<br />
biogeografía filogenética explica historias evolucionarias, pero no<br />
cuestiones biogeográficas; mas bien los patrones biogeográficos<br />
pueden explicar la filogenia. En sus palabras, la Biogeografía se<br />
trata de relaciones proximales de áreas bióticas de endemismo. Los únicos<br />
métodos para ello son la panbiogeografía y la biogeografía<br />
cladística, con su enfoque de “arte de descubrimiento”. Grehan<br />
(2001), es aún más restrictivo, pues afirma que la<br />
panbiogeografía es el único método en la actualidad que es<br />
sólido geográficamente.<br />
Historias y ontología<br />
El análisis cladístico es considerado, en principio, deductivo<br />
(Crother, 2002); aunque algunos lo catalogan de inductivo (ver<br />
Fontenla, 2002). La biogeografía cladística en su sentido más<br />
amplio también es conceptuada como hipotético-deductiva<br />
(Espinosa y Llorente, 1993; Vargas, 1993; Schuh, 2000). De<br />
Queiroz y Poe (2001) exponen que la corroboración de Popper<br />
es basada en el principio general de probabilidad, que es<br />
considerado inductivo. En coincidencia, Kluge (2001) asevera<br />
que el análisis de probabilidad es verificacionista/inductivo,<br />
opuesto al de parsimonia, que es practicado en términos de<br />
sometimiento a prueba de Popper y, por lo tanto, es<br />
falsacionista/deductivo. Así, tanto la biogeografía cladística<br />
como la filogenética, sensu Van Veller et al. (2003), se<br />
corresponderían ambas con este último modelo, ya que están<br />
basadas en la parsimonia. Sin embargo, Farris et al. (2001)<br />
concluyen que la parsimonia es corroboracionista. Por otra<br />
parte, Rieppel (2003) acota que la lógica deductiva no excluye<br />
una actitud verificacionista. Por consiguiente, esta actitud no<br />
tiene que ser equivalente con el inductivismo. Como en la vida<br />
real es absurdo suponer que todas las distribuciones observadas<br />
son vicariantes, los biogeógrafos cladistas han ideado una serie<br />
de enunciados, que permiten una manipulación de los resultados<br />
COCUYO 15, 2005 35
cladísticos para tratar con distribuciones amplias, redundantes y<br />
extinciones.<br />
A estos enunciados se les llama asunciones o suposiciones 0,<br />
1 y 2 (Nelson y Platnick, 1981; Van Veller et al., 2000, 2001).<br />
Suposición 0. Las áreas con el taxón se consideran<br />
monofiléticas. El taxón ampliamente distribuido es una<br />
sinapomorfía (carácter derivado compartido). Suposición 1.<br />
Áreas pueden constituir grupos mono o parafiléticos.<br />
Suposición 2. Sólo una de las ocurrencias es considerada como<br />
evidencia; la otra puede “flotar” en los cladogramas. Las áreas<br />
pueden constituir grupos mono, para o polifiléticos. Estas<br />
suposiciones también exhiben una sintaxis lógico-deductiva, la<br />
cual puede adoptar la siguiente forma generalizada: Suposición<br />
1. Lo que es cierto para T1 (taxón 1) en área A en relación con<br />
T2 y T3 en áreas C y A-C, respectivamente, es también cierto<br />
para T1 en área C. Suposición 2. Lo que es cierto para T1 en<br />
área A en relación con T2 y T3 en áreas A-C respectivamente, no<br />
es necesariamente cierto para T1 en área C. El BPA se<br />
corresponde con la Suposición 0: En esta Brooks y McLennan<br />
(2003) consignan: Debe tratar con todas las especies y todas las<br />
distribuciones en cada entrada filogenética sin modificación, y su<br />
análisis final debe ser lógicamente consistente con todos los<br />
datos entrados. Para ello se genera la convención de la<br />
duplicación de las áreas (Apéndice). La sintaxis lógica es la<br />
siguiente: si cada área es tratada como un taxón único, la<br />
Asunción 0 es violada siempre que un área tenga una historia<br />
reticulada. Esto aparecerá como una homoplasia (convergencia<br />
o paralelismo) en el BPA primario. Bajo tales circunstancias,<br />
duplique las áreas relevantes hasta que la Asunción 0 no se viole<br />
más.<br />
Todas las suposiciones anteriores tienen una estructura<br />
lógica. De hecho, el modelo deductivo tiene sus raíces en la<br />
filosofía Aristotélica y su sintaxis se basa en la construcción de<br />
silogismos2 como el siguiente: Todos los hombres son<br />
mortales/Sócrates es un hombre/Sócrates es mortal. Se obtiene<br />
una inferencia lógica congruente con la realidad ontológica. En<br />
un segundo ejemplo: Todos los mamíferos son<br />
cuadrúpedos/Todos los humanos son mamíferos/Todos los<br />
humanos son cuadrúpedos. La deducción es impecable, pero<br />
errada, porque la premisa mayor, al menos en parte, es falsa y la<br />
inferencia es absurda. Por lo tanto, es obvio que algo<br />
correctamente deducido no tiene que estar correctamente<br />
explicado desde el punto de vista ontológico (Richards, 1998).<br />
Popper (1990) propuso para tratar con fenómenos reales la<br />
aplicación de una lógica proposicional o de probabilidades. La<br />
elegante lógica deductiva tendría una probabilidad = 1. Esa es la<br />
verdad lógica, que tiene una probabilidad absoluta de ocurrencia.<br />
Sin embargo, la verdad en condiciones de evidencias, la verdad<br />
ontológica, responde a una lógica probabilística. La<br />
probabilidad absoluta sería un caso especial de una probabilidad<br />
relativa real, no lo contrario.<br />
Kluge (2002) discrimina entre probabilidad clásica (todos<br />
los eventos son numerados; las probabilidades son conocidas;<br />
existe una predicción) y probabilidad histórica (sólo los eventos<br />
que han ocurrido pueden ser numerados; no existen<br />
predicciones, pero pueden realizarse comparaciones entre<br />
resultados). Por su lado, Grant (2002) aseveró que los biólogos<br />
evolucionistas están obsesionados con las operaciones de<br />
descubrimiento -el prisma a través de los cuales los científicos<br />
ven e interpretan al mundo, el cual es independiente de algún<br />
método en particular- y distingue entre operaciones<br />
nomotéticas (universales, predictivas) e ideográficas<br />
(retroductivas, históricas). Según Salthe (1985), lo nomotético<br />
se enmarca dentro del canon nomológico o hipotéticodeductivo<br />
por cobertura de leyes universales; enfatiza<br />
regularidades en patrones y procesos, mientras que lo<br />
ideográfico enfatiza diferencias: eventos y entidades<br />
individuales, trayectorias históricas.<br />
Historias y ontología<br />
Gould (2002) declara que principios generales pueden<br />
regular gamas de eventos, pero eventos particulares e<br />
impredecibles son los que hacen la historia. La historia no es<br />
una “maldita cosa tras la otra”. La historia es devenir, es<br />
trayectoria, y las trayectorias no devienen ni emergen en pistas<br />
impolutas hasta lo infinito, sino a través de contextos ecológicos,<br />
interaccionantes y co-constructivos. A la vez que la historia es<br />
una emergencia, es, simultáneamente, co-constructora de<br />
hechos. La historia se hace a sí misma de eventos e interacciones,<br />
a la vez que modela y co-determina eventos e interacciones. Lo<br />
que deviene y emerge no es necesariamente deducible, aunque sí<br />
puede ser explicado. La vida no es deducción, es desarrollo y<br />
evolución. Los hechos se desarrollan en contextos ambientales,<br />
naturales o sociales particulares, que constriñen la manera de<br />
expresarse el devenir; pero, además, también imbrican procesos<br />
y eventos casuales, contingentes, que pertenecen a dinámicas<br />
distintas, y que podrían o no haber concurrido para generar un<br />
devenir, una historia particular. La contingencia histórica limita<br />
las opciones evolucionarias (Kluge, 2002). En un contexto<br />
determinado sólo es más probable que ocurran determinados<br />
hechos, los cuales son muy poco probables que ocurran en otro.<br />
Gould (2002) propone que la estructura probabilística de los<br />
fenómenos históricos, su impredicibilidad, es una propiedad<br />
ontológica - consustancial- de la naturaleza. La incapacidad de<br />
predecir antes del hecho, sólo evidencia su complejidad. La<br />
capacidad subsiguiente de explicar con rigor después del hecho,<br />
puede lograr el mismo nivel de confiabilidad que cualquier<br />
resolución física bajo leyes invariantes. Ello es congruente con<br />
las tesis de Kauffman (1993, 1996, 2000) y Goodwin (1998)<br />
acerca de que la evolución es una conjunción de procesos<br />
deterministas intrínsecos de los organismos -considerados<br />
como sistemas dinámicos complejos autoorganizados y<br />
autocausales- y la contingencia histórica.<br />
Brooks, Van Veller y colaboradores enfatizan que la<br />
distribución espacial, su desarrollo histórico, es un fenómeno<br />
complejo, contingente. Por ello consideran que en la<br />
biogeografía histórica el enfoque apropiado es el de la historiade-los<br />
taxones, lo cual impugnan Ebach y Humphries (2003).<br />
Los biogeógrafos aludidos defienden operaciones de<br />
descubrimiento a posteriori. Sin embargo, estos análisis no son<br />
análogos a las explicaciones después del hecho, sensu Gould<br />
(2002), sino que tienen un sentido hipotéticodeductivo/refutacionista,<br />
según el marco epistemológico de<br />
Popper (Brooks y Mc Lennan, 2001; Van Veller et al., 2003). La<br />
36 COCUYO 15, 2005
tarea de la historia es el descubrimiento y la explicación, no la<br />
predicción3 (Siddall y Kluge, 1997; Richards, 1998; Kluge, 2001).<br />
En el BPA secundario se duplican las áreas, para buscar una<br />
congruencia epistemológica, lógica, a la explicación a posteriori;<br />
pero esta duplicación no responde a algún principio biológico<br />
ontológico*, y además se continúa representando mediante un<br />
diagrama jerárquico. Hovenkamp (1997), expuso que las<br />
relaciones genealógicas son divergentes y jerárquicas. Así, la<br />
forma de un cladograma y la historia probable que refleja,<br />
resultan una configuración confiable para representar la historia<br />
de los taxones; pero una historia de reticulación no se representa<br />
con un patrón jerárquico y ramificado. La historia de la tierra<br />
influye en la historia de la vida y la historia de la vida es<br />
divergente, pero no necesariamente la historia de la tierra es<br />
divergente o puede ser representada como un cladograma.<br />
La jerarquía sólo es de esperar cuando existe una<br />
correspondencia directa entre fraccionamiento de áreas y<br />
especiación. Patrones jerárquicos de distribución, como las<br />
verdades lógicas, deben constituir casos particulares de<br />
congruencia entre procesos geológicos, ecológicos y<br />
genealógicos; pero no lo contrario. Como propone Lobo<br />
(1999), la vicarianza es más explicativa a grandes niveles de<br />
inclusividad en el tiempo y en el espacio. Espinosa y Llorente<br />
(1993) reconocieron como un defecto de la biogeografía de la<br />
vicarianza su apego estricto a los arreglos jerárquicos para<br />
expresar las interrelaciones históricas. Martín-Piera y San<br />
Martín (1999) catalogan a estos patrones jerárquicos y<br />
ramificados como reduccionistas, mientras que Legendre y<br />
Makarenkov (2002) proponen un modelo reticulado para<br />
patrones de distribución<br />
Causas y azares<br />
Llorente y Pavavero (1996) llegaron a afirmar que “la<br />
vicarianza es la teoría central de la biología….la elegancia suprema de la<br />
teoría de la vicarianza es que la historia de la Tierra se encuentra escrita en<br />
los cromosomas de los organismos”. Algunos biogeógafos sólo<br />
aceptan a la vicarianza como causa de los patrones de<br />
distribución, y no admiten a la dispersión como evento<br />
generador de patrones biogeográficos. Según este criterio, la<br />
dispersión, en esencia, es un evento aleatorio, y no es posible<br />
considerarlo como hacedor de patrones de distribución. No<br />
obstante, la propia aleatoriedad crea patrones de distribución<br />
reconocibles. El “desbalance” taxonómico de las biotas insulares<br />
oceánicas, o esencialmente oceánicas, con relación a la de las<br />
biotas continentales más cercanas, es una consecuencia,<br />
justamente, de la aleatoriedad del arribo a estas áreas de los<br />
antecesores de su biota actual. Lo cual genera un patrón de<br />
diversidad característico, donde grupos muy diversos en el<br />
continente se encuentran ausentes o pobremente representados<br />
en dichas islas. Se argumenta que la vicarianza sí produce<br />
patrones reconocibles, porque fragmenta biotas completas que<br />
antes se encontraban juntas. Es necesario tener en cuenta que,<br />
desde el momento que dos áreas o biotas quedan separadas, los<br />
* De hecho, todos los casos de duplicación de áreas que se realizan para inferir<br />
la ocurrencia de dispersión o vicarianza, es posible hacerlos en el BPA primario,<br />
analizando la posición de las especies y las áreas en el cladograma.<br />
contextos ambientales de las ahora independientes áreas pueden<br />
variar. Y ello también puede generar un “desbalance”, debido a la<br />
extinción o diversificación diferente de los grupos en las áreas<br />
independientes.<br />
Salthe (1985) distingue entre causa y regulación. “Causa”<br />
4<br />
involucra las causas formal y material , que tienen que ver con el<br />
dinamismo inherente de los sistemas. La “regulación” se refiere a<br />
la causalidad material (en parte), eficiente y final. En otras<br />
palabras: lo causal es intrínseco; lo regulatorio, externo. Como se<br />
aparta de la visión mayoritaria, Salthe reconoce que ello “es su<br />
peor conflicto con los hábitos establecidos”. Usualmente,<br />
pensamos que un cambio ambiental “causa” una<br />
reestructuración de una biota local. El cambio ambiental elimina<br />
a los que no se adaptan o no tienen la posibilidad física de<br />
reajustar sus relaciones espaciales; pero otros sobreviven.<br />
Cabría suponer que la misma causa de la desaparición de unos es<br />
causante de la supervivencia de otros, los cuales de todas<br />
maneras ya sobrevivían sin tal causa. Una alternativa valida es el<br />
asumir que el cambio ha provocado una regulación espacial de las<br />
poblaciones; las ha modificado, al interferir con la dinámica<br />
organismo-entorno preexistente.<br />
Del mismo modo, el aislamiento espacial, la vicarianza, no<br />
causa la especiación, sólo regula o modifica el intercambio de<br />
información genética entre poblaciones. Las causas de la<br />
especiación, o de su antípoda, el éstasis evolucionario, son<br />
intrínsecas. Kampis (1998) declaró que la evolución no debe ser<br />
considerada como un juego contra algo “allá afuera”. Los<br />
cambios vienen desde el sistema en sí mismo, no desde afuera<br />
(Csányi, 1998; Alvárez, 1998). Los organismos no tienen causas<br />
externas, son entidades autocausales (Goodwin, 1998). Así, sus<br />
relaciones de espacio pueden ser reguladas, pero no causadas, por<br />
fuerzas, procesos o eventos extrínsecos. Atlan (1998) enfatiza<br />
que una explicación causal de algo no siempre es lo mismo que la<br />
causa real de tal cosa. Una razón lógica para algo puede ser<br />
diferente a su causa física, como resultado de nuestra frecuente<br />
incapacidad de tener una explicación adecuada para algo<br />
Historias y narrativas<br />
Stamos (1996) expuso, a la vez que criticó, que el basamento<br />
epistemológico de la biología descansa en buena medida en el<br />
legado del filósofo Karl Popper. Ruse (2001), se expresó con<br />
sarcasmo acerca del uso de esta epistemología por parte de los<br />
científicos como soporte a sus ideas. Rieppel (2003) nos hace<br />
percatar que, a partir de la propuesta Popperiana de la lógica<br />
deductiva como el organon del criticismo, se cruzó el puente entre<br />
sentido común filosófico y una teoría plena de conocimiento.<br />
Popper escribió sobre la lógica del descubrimiento científico, no<br />
sobre ciencias aplicadas o como solucionar un problema<br />
específico. Lo lógico y lo hipotético-deductivo no son<br />
antagónicos con la búsqueda de explicaciones sobre la<br />
complejidad del mundo; pero el razonamiento lógico opera en<br />
un espacio abstracto, mientras que la naturaleza es ontológica en<br />
su manifestación material en un espacio y tiempo reales. Lo<br />
universal, lo determinista y lo epistémico, se pueden explicar a<br />
través de derivaciones lógicas. La lógica deductiva es lineal; lo<br />
complejo puede ser heterárquico, emergente o recursivo, pero<br />
nunca lineal. Lo histórico requiere de derivaciones temporales y<br />
COCUYO 15, 2005 37
de la aprehensión de lo contingente y lo contextual. Las<br />
explicaciones históricas se construyen sobre la base de la<br />
interpretación de evidencias, circunstancias y conocimiento de<br />
fondo, con narrativas científicas como salidas explicativas. Las<br />
fórmulas lógicas deductivas no pueden constituir las<br />
herramientas exclusivas para validar la comprensión de<br />
fenómenos complejos del devenir ontológico y emergente de la<br />
naturaleza.<br />
Las narrativas no han tenido una historia feliz en la<br />
biogeografía histórica. Ball (1976) y Humphries y Parenti (1999)<br />
asociaron las narrativas biogeográficas a las explicaciones de<br />
Darwin-Wallace acerca de centros de origen para cada taxón.<br />
Platnick y Nelson (1978) afirmaron que no hay que preocuparse<br />
por distribuciones o patrones particulares, si estos no son<br />
congruentes con un patrón general de relaciones mostrado por<br />
otro grupo de taxones endémicos de las áreas en cuestión.<br />
Nelson (1978) fue lapidario al respecto: “las ideas de Darwin y<br />
Wallace hicieron de la biogeografía la ciencia de lo raro, lo<br />
misterioso, lo milagroso y lo imposible”. Por su parte, Llorente<br />
(2000) expresó que la mejor aproximación para el conocimiento<br />
de la historia espacial de la vida es la congruencia entre áreas, vía<br />
hipótesis cladista o panbiogeográfica, mientras que la<br />
elaboración de narrativas sobre el origen y dispersión de grupos<br />
particulares se está desvaneciendo. Recientemente, Grehan<br />
(2003) la emprendió contra las explicaciones narrativas en la<br />
compilación sobre biogeografía de las Antillas editada por Wood<br />
y Sergile (2001).<br />
Pienso que se han hiperbolizado las reflexiones críticas de Ball<br />
(1976) sobre las asunciones de Darwin y Wallace, ideadas sobre<br />
la concepción de una geografía del mundo estática. Un esquema<br />
similar tuvo la biogeografía filogenética de Hennig (1968) y<br />
Brundin (1972) (esta sería “la otra” biogeografía filogenética).<br />
Ebach y Humphries (2003) afirman que la dispersión no se<br />
integra con la concepción de una tierra dinámica y añaden que<br />
cambios intrínsecos no pueden ocurrir sin un estímulo externo.<br />
Pensar así es considerar a los organismos como objetos a<br />
merced de causas eficientes externas, como en la ortodoxia de la<br />
lógica clásica y de la modernidad. Según este parecer, los<br />
procesos geológicos, como la tectónica de placas, son la fuerza<br />
primaria guiando la evolución; es eso lo que constituye sentido<br />
común en la biogeografía, en su ontología, no el hacer narrativas<br />
5<br />
como explicaciones. Grehan (2000) también expuso un criterio<br />
similar, al afirmar que la capacidad de migración no es una<br />
propiedad intrínseca, sino que surge a través de una relación<br />
particular entre organismo y medio ambiente. Por otra parte,<br />
Wilkinson (2001, 2003), defiende que la dispersión no es sólo<br />
uno de los procesos* fundamentales de la biogeografía, sino<br />
6<br />
además crucial para entender la distribución de los organismos.<br />
El término “narrativa” también tiende a su identificación con<br />
la narrativa literaria, llena de imaginación y figuras retóricas; pero<br />
de lo que se trata es de construir una narrativa histórica para<br />
explicar la complejidad ontológica de la naturaleza. Juarrero<br />
(1999) propone la narrativa hermenéutica, la cual construye<br />
* De una manera más o menos laxa se manifiesta que la dispersión y la<br />
vicarianza son procesos. Considero que existen buenas razones para<br />
considerarlas eventos. Pero eso sería tema para otras reflexiones.<br />
explicaciones de un sistema a partir de las relaciones entre sus<br />
componentes, pues lo deductivo no puede explicar lo que<br />
emerge, lo que deviene. Ciurana (2001.) nos dice que la<br />
hermenéutica es la negación del método, entendido éste como<br />
un conjunto de reglas ideales y universales que nos lleven a un<br />
acuerdo o a descubrir algo. “La hermenéutica es la negación de<br />
un método entendido como punto de vista de Arquímedes desde<br />
el cual dominar las cosas; cómo entidad exterior al mundo. La<br />
lucha de la hermenéutica lo es contra la idea de una<br />
representación exacta del mundo. A cambio se nos propone la<br />
idea de diálogo y de conversación”…La hermenéutica es un<br />
método común a todas las disciplinas que tienen que interpretar<br />
un lenguaje cualquiera (de ahí proviene su afinidad con la<br />
retórica), o que luchan con las dificultades de la interpretación de<br />
lo que parece extraño (de ahí su vinculación con la dialéctica)....<br />
Interpretar es reconstruir un discurso dentro de un contexto de<br />
vida. Frente al afán de conmensurabilidad y de reducción; frente<br />
a la teoría que afirma que la esencia del hombre es descubrir<br />
esencias….. y reflejarlas como en un espejo, con claridad y<br />
distinción, se nos propone, con la hermenéutica, una idea de<br />
cultura como diálogo y conversación”.<br />
La historia es una herramienta para conocer al mundo, para<br />
representarlo e interaccionar con él (Taylor, 1986). Las<br />
narrativas biogeográficas deben ser basadas en la observación de<br />
la distribución de los organismos y en la posterior elaboración de<br />
una historia explicativa consistente (Real y Ramírez, 1992).<br />
Richards (1998) predice que nos percataremos de cuánto más en<br />
la ciencia depende de la eficacia de narrativas históricas que el<br />
subsumir bajo leyes. Las narrativas son irreducibles a otros<br />
modos de representar y explicar; son recursos ineludibles para<br />
ciertas tareas y áreas del conocimiento. El criterio de lo<br />
importante depende de que historia se cuente, de un criterio más<br />
general (Beltrán, 1998). Debe tener en cuenta el “conocimiento<br />
de fondo”, que consiste en teorías y resultados avalados por la<br />
praxis de lo científico y lo racional (Farris, 2000; Kluge, 2001); no<br />
de un pensar racionalizado y enchalecado. Las explicaciones<br />
deben ser contextuales, abarcar lo global dentro de los<br />
desarrollos y trayectorias locales y comprender como lo local<br />
influye en los patrones globales. Hull (1998) afirma que las<br />
narrativas se construyen para aprehender el movimiento de<br />
sucesos cuyo rasgo es el cambio, y agrega que “los filósofos por<br />
no aceptar cambios argumentan que las narrativas históricas no<br />
son explicativas”. Para ser justos, sería correcto acotar que ello<br />
caracteriza sólo a ciertos grupos de filósofos y a ciertos grupos<br />
de indagadores.<br />
Rosen (1988) expuso que la historia no tiene un desarrollo<br />
lógico, por la intervención de eventos contingentes y casuales en<br />
su devenir. También agregó que la ciencia procede en un frente<br />
mucho más abarcador que sólo el método hipotético-deductivo.<br />
Las explicaciones científicas, a la vez que distintamente causales<br />
-como asevera Kluge (2001)- pueden ser racionales y<br />
cualitativas (Pérez, 2000). Si en la selección de una conjunción<br />
de elementos apropiados para el contexto particular de una<br />
narrativa no se puede contar con un criterio “objetivo” externo<br />
al sistema focal y al propio sujeto indagador, ello no señala más<br />
que a la complejidad ontológica de la situación. El sujeto<br />
indagante no puede ser nunca enajenado del sistema indagado<br />
38 COCUYO 15, 2005
(asunción típica del pensamiento neopositivista y racionalista<br />
imperante). La estructuración del sistema cognitivo sujetoobjeto<br />
es unitaria e irreducible. Tampoco existe “objetividad” en<br />
la aplicación de un modelo cognitivo específico, pues el propio<br />
acto de selección es puramente subjetivo y depende del<br />
paradigma aceptado por el sujeto (el sujeto tal vez ni se percate<br />
que se encuentra embebido en un paradigma científico-socialcultural).<br />
La narrativa no es “mala” en sí misma; lo que sí pueden<br />
existir son malos narradores. No existe una historia<br />
ontológicamente “débil”, pero sí es posible construir<br />
narraciones o hipótesis históricas poco robustas, convincentes o<br />
racionales. Del mismo modo que es posible realizar<br />
deducciones impecables en el espacio epistemológico, pero<br />
extravagantes en el espacio de la ontología natural.<br />
Gould (2002) aseveró que tanto leyes inmanentes como<br />
narrativas (hacia las cuales existe una profunda e irracional<br />
reticencia, según sus palabras) son dos formas de conocimiento<br />
objetivo. Al mismo tiempo, abogó por una pluralidad de estilos<br />
explicativos para nuestra comprensión de “generalidades atadas<br />
a leyes y particulares fascinantes”. En torno a la misma idea,<br />
Kauffman (1996) consigna: “¿Cómo puede la vida ser<br />
contingente, impredecible y accidental obedeciendo a la vez<br />
leyes generales La misma pregunta emerge cuando pensamos<br />
acerca de la historia. …Los sistemas vivientes pueden todos<br />
exhibir propiedades sujetas a leyes, y aún estar agraciados con un<br />
bordado de filigrana histórica; esos detalles maravillosos que<br />
podrían fácilmente haber sido de otra manera, y cuya misma<br />
improbabilidad cautiva nuestra admiración hechizada”.<br />
Kauffman (2000) establece otra elegante metáfora entre<br />
narrativa y determinismo, cuando escribe: “Las biosferas<br />
demandan sus Shakespeares tanto como a sus Newtons.<br />
Tendremos que repensar qué es la ciencia en sí misma… Las<br />
humanidades y la ciencia pueden encontrar una inesperada e<br />
inevitable unión”. Por su parte, Pérez (2000) reflexiona que los<br />
esquemas propuestos por los filósofos de la ciencia son camisas<br />
de fuerza y visualiza una necesaria reconstrucción de la filosofía<br />
que considere, junto con la historia, toda la inmensa extensión y<br />
la complejidad de la ciencia contemporánea. Mientras tanto,<br />
evoco la serena y reflexiva pluma de Darwin, el primer<br />
biogeógrafo coherente: “Cuando no miremos ya a un ser orgánico del<br />
modo en el que un salvaje mira un buque, como algo completamente fuera de<br />
su comprensión, cuando consideremos cada producción de la naturaleza como<br />
algo que ha tenido una historia […]; ¡cuánto más interesante, hablo por<br />
experiencia, se volvería nuestro estudio!”<br />
NOTAS<br />
1<br />
En diferentes obras, Popper ha propuesto que la delimitación<br />
entre ciencia y no ciencia estaba dado por el empleo del método<br />
hipotético-deductivo refutacionista, es decir, por el modus tollens<br />
del criticismo. Según Pérez (2000), en el siglo II AC, Crisipo<br />
formula una serie de cinco “silogismos hipotéticos”, en<br />
contraposición a los “silogismos categóricos” de Aristóteles (ver<br />
nota siguiente). El silogismo verificacionista o modus ponens, es<br />
como sigue: Si “P” implica “Q”, y “P” es cierta, entonces “Q” es<br />
cierta. El modus tollens o refutacionista sería: Si “P” implica “Q”, y<br />
“Q” es falsa, entonces “P” es falsa. Rieppel. (2003) utiliza la<br />
siguiente sintaxis: “Si P, entonces Q”, “P”, “por consiguiente<br />
Q”. (modus ponens). “Si P, entonces Q”, “no Q”, “por<br />
consiguiente no P” (modus tollens). Jaramillo (1993) consigna que<br />
el modus tollens, aunque resulta inobjetable lógicamente, no es<br />
aplicable a las ciencias, y se ve la necesidad de la sustitución de<br />
sistemas analíticos lógicos por históricos. La lógica no<br />
monopoliza el ejercicio de la racionalidad ni agota el ámbito de la<br />
reflexión filosófica. Delgado (2001) considera que son<br />
limitaciones en Popper la elevación de la lógica deductiva y el<br />
modus tollens a la calidad de instrumentos lógicos infalibles para<br />
construir teorías científicamente aceptables. Gattei (2002)<br />
expone que la falsabilidad nos advierte que, como seres<br />
humanos racionales, debemos estar alerta acerca de la falibilidad<br />
y crítica de nuestras teorías. Contra ello, es imposible<br />
argumentar. No obstante, y como advierte Pérez (2000), la<br />
aceptación o no de las teorías e hipótesis tienen más que ver con<br />
su aceptación dentro de un paradigma o contexto de intereses y<br />
poder, que de su refutación o corroboración lógicas. Rieppel<br />
(2003) advierte que lógica deductiva no excluye necesariamente<br />
una actitud verificacionista, además de que la deducción ocurre<br />
en un espacio lógico; es un acto racional, epistémico, por lo que<br />
puede estar por completo divorciado de la ontología de la<br />
naturaleza Una conclusión de lo anterior es que lo lógico no debe<br />
tomarse como criterio exclusivo de la demarcación de lo<br />
científico, ni como herramienta necesaria para la comprensión<br />
de lo complejo en la biogeografía.<br />
2<br />
Un silogismo categórico es un enunciado con dos premisas y<br />
una conclusión unidas en forma de inferencia e implicación<br />
(Pérez, 2000). El sujeto de la premisa primera o mayor (S ), 1<br />
aparece como predicado (P) en la premisa segunda o menor (S ) 2<br />
(Rieppel, 2003). La implicación comprende la premisa menor<br />
y el predicado. Así, su sintaxis formal es como sigue: S + P/S<br />
1 2<br />
+ S / S + P; o también: Todos los A son B/Todos los C son<br />
1 2<br />
A/Todos los C son B.<br />
3<br />
Por supuesto, las lecciones de la historia, ya sea esta natural o<br />
humana, contienen elementos de predicción probabilísticas,<br />
pero no constituyen necesidades lógicas, debido a la<br />
contingencia de la complejidad de los ambientes locales y, en el<br />
caso de la humana, porque, al menos en teoría, las sociedades<br />
humanas pueden actuar de manera tal, que no se cumplan ciertas<br />
predicciones, como por ejemplo, evitar una catástrofe ambiental<br />
y social de niveles ecuménicos.<br />
4<br />
Aristóteles describió el desarrollo de las cosas en términos de<br />
una causalidad cuatripartita (tomado de Alvárez, 1998 y Riedl,<br />
1998): Causa material. Es la realidad corpórea y material que<br />
funciona como el sustrato del cambio, donde los cambios<br />
ocurren, a partir de lo cual se desarrollan los componentes. La<br />
unidad de forma y materia es lo que constituye una cosa en su<br />
realidad. Causa formal Es la forma, patrón, configuración,<br />
dentro de lo cual algo es cambiado; es la especificidad que es<br />
transmitida a la materia sobre la base del movimiento. Su<br />
dinámica es integradora, de encerramiento. Puede verse como el<br />
marco dentro del cual acciona la causa eficiente. Causa eficiente.<br />
Desde donde se origina el movimiento o el reposo; es por lo cual<br />
COCUYO 15, 2005 39
ocurre un cambio. Es poder, energía. Es el movimiento que<br />
trasmite la especificidad a la materia, y como tal, es la que realiza<br />
de manera inmediata el desarrollo de las entidades. Su dinámica<br />
es de discriminación, diferenciación; acciona entre componentes<br />
del sistema y entre componentes y el ambiente. Causa final. Es en<br />
aras de lo cual una actividad tiene lugar; representa la finalidad, el<br />
motivo o propósito por lo cual el cambio es producido. Es la<br />
obtención del completo ordenamiento del sistema; conlleva la<br />
máxima cantidad de información, de coherencia. Aristóteles<br />
pensaba que la causa final era la actualización de propiedades<br />
potenciales, que el futuro determinaba el pasado y el presente<br />
(Pérez, 2000). Riedl (1998) considera que Aristóteles concibió<br />
las causas material y formal accionando dentro del sistema,<br />
mientras que la eficiente y la final accionaban desde afuera.<br />
Según Van de Vijver (1998), Aristóteles no aceptó el papel del<br />
azar en el desarrollo y concibió la acción de las causas de manera<br />
lineal y jerárquica. Su concepción se conoce como “el<br />
desarrollismo de uno”. El desarrollo depende básicamente de la<br />
causa eficiente. La ciencia moderna desterró a la multicausalidad<br />
y se quedó sólo con una causalidad eficiente, accionando desde el<br />
ambiente externo. Ahora se concibe la evolución y el desarrollo<br />
bajo el influjo no sólo de procesos deterministas, sino también<br />
del azar, la emergencia y la autoorganización sobre la dinámica<br />
local de los sistemas, conocida como el “desarrollismo de<br />
muchos” Se acepta una multicausalidad, porque sólo una<br />
causalidad eficiente no explica la evolución de sistemas<br />
complejos. Es necesario una causalidad compleja, no lineal y<br />
heterárquica. Con este enfoque y según Matsuno y Salthe (1995)<br />
y Van de Vijver (1998), la causa material se refiere a la singularidad<br />
de la dinámica local y la causa formal a las condiciones de límites<br />
del ambiente. La causa eficiente se concibe como perturbaciones<br />
externas, episódicas, del ambiente, mientras que la causa final<br />
tiene que ver con el aseguramiento, mantenimiento y<br />
conservación de la dinámica local. Causas formal y material<br />
accionan en el interior del sistema y la eficiente y final desde el<br />
exterior Sotolongo (2002) propone que la causa material hace<br />
énfasis en la interacción entre componentes, involucrados como<br />
partes de un todo; la causa formal, con la interacción específica del<br />
todo sobre los componentes; la causa final, con la emergencia de<br />
un orden dinámico complejo, ventajoso dentro del contexto<br />
local, y la causa eficiente con interacciones intrínsecas,<br />
contextuales. Como es posible discernir, existe una diferencia en<br />
el tratamiento moderno de la causa eficiente por parte de estos<br />
autores, ya sea considerándola externa o interna al sistema focal.<br />
Por otra parte, Salthe (1985) sólo reconoce como agentes<br />
causales a los intrínsecos.<br />
5<br />
La primera reflexión que se me ocurre es que no existe ninguna<br />
contradicción entre dispersión y dinamismo de las áreas. Por<br />
ejemplo, Briggs (2003), basado en información reciente<br />
filogenética, paleontológica y paleogeográfica, propone<br />
alternativas dispersalistas para la historia biogeográfica de<br />
grupos de aves y peces, cuya explicación tradicional había sido la<br />
vicarianza del supercontinente Gondwana. Para ello, tuvo en<br />
cuenta, precisamente, el dinamismo de las áreas en cuestión.<br />
Wilkinson (2003) es también acertado cuando expone que las<br />
explicaciones por dispersión o vicarianza responden a<br />
determinadas escalas espaciales y temporales, y de la inclusividad<br />
del nivel taxonómico bajo escrutinio. Lo otro que llama la<br />
atención es la coincidencia de Ebach, Humphries y Grehan con<br />
el pensamiento aristotélico. Según Juarrero (1999), Aristóteles<br />
creía que nada se puede mover, causar o actuar sobre sí mismo;<br />
en especial los organismos. Se necesitan estímulos externos; de<br />
ahí una de las razones para su multicausalidad. Sin embargo, el<br />
gran filósofo Emmanuel Kant (1724-1804) concibió a los<br />
organismos como entidades autocausales y autoorganizadas,<br />
como “ejemplos vivientes de autocausación”. Esta es la<br />
concepción de muchos autores contemporáneos, lo que significa<br />
que, de hecho, los organismos causan su propia evolución, o que<br />
la evolución es causa y efecto de sí misma (Kampis, 1998). Otra<br />
reflexión es que, si aceptamos que la migración no es una<br />
propiedad intrínseca, sino causada por relaciones particulares<br />
de espacio, cabe preguntarse porque no migran todos los<br />
integrantes de una comunidad donde también existen<br />
organismos que sí lo hacen. Es decir, que la causa externa<br />
(relaciones de espacio) de algo (migración) también origina el<br />
efecto contrario a ese algo (sedentarismo o sencillamente no<br />
capacidad migratoria), lo cual no tiene sentido lógico ni<br />
explicativo.<br />
6<br />
De manera apasionada, Wilkinson (2003) consigna que:<br />
“Darwin pudo haber mostrado un grandioso buen sentido<br />
común en enfatizar la importancia de la dispersión, no<br />
meramente porque él no conocía acerca de la tectónica de placas,<br />
sino porque la dispersión es un proceso fundamental en<br />
comprender mucho de la biogeografía”. No menos arrastrados<br />
por sus caldeados intelectos, Van Veller et al. (2003) aseveran:<br />
“Una especie no puede tener dos historias diferentes…por<br />
consiguiente, su ocurrencia en dos áreas distintas no puede<br />
haber sido causada por vicarianza, sino que debe ser el resultado<br />
de dispersión”. La impugnación de Ebach et al. (2003) alcanza<br />
ribetes de ofuscamiento: “Este enunciado, es decir, que todas las<br />
especies sujetas a eventos vicariantes son requeridas que<br />
respondan especiando, refleja una incomprensión de la<br />
biogeografía histórica. Nada en el mundo real requiere que<br />
taxones ampliamente distribuidos sean un resultado de<br />
dispersión”. ¡Pero tampoco nada en el mundo real requiere que<br />
los taxones ampliamente distribuidos sean un resultado de<br />
éstasis evolucionario ante la fragmentación de sus áreas! ¿Por<br />
qué no aceptar ambos procesos como legítimos en la explicación e<br />
interpretación de las distribuciones<br />
REFERENCIAS<br />
Alvarez, J. M. 1998. Self-organization and self-constrruction of<br />
order. En: Evolutionary Systems. Biological and epistemological<br />
perspectives on selection and self-organization. G. Van de Vijver, S. N.<br />
Salthe y M. Delpos (Eds). Kluwer Academic Publishers, pp:<br />
167-78.<br />
Ball, I. R. 1976. Nature and formulation of biogeographical<br />
hypotheses. Syst. Zool., 24: 40-430.<br />
Beltrán, C. L. 1998. Narrativa y explicación en las ciencias<br />
naturales. 1998. En: Historia y explicación en Biología. (Eds: S.<br />
Martínez y A. Barahour). UNAM, México. Pp: 197-211.<br />
Briggs, J. C. 2003. Fishes and birds: Gondwana life rafts<br />
40 COCUYO 15, 2005
econsidered. Syst. Biol., 52: 548-554.<br />
Brooks, D. R. y D. A. McLennan. 2001. A comparison of a<br />
discovery-based and event-based method of historical<br />
biogeography. J. Biogeogr., 28; 757-767.<br />
Brooks, D. R. y D. A. McLennan. 2002. The nature of diversity: an<br />
evolutionary voyage of discovery. University of Chicago Press,<br />
Chicago.<br />
Brooks, D. R. y D. A. McLennan. 2003. Extending phylogenetic<br />
studies of coevolution: secondary Brooks Parsimony<br />
Analysis, parasites, and the Great Apes. Cladistics, 19: 104-<br />
119.<br />
Brooks, D. R. y G. P. Van Veller. 2003. Critique of parsimony<br />
análisis of endemicity as a method of historical biogeography.<br />
J. Biogeogr., 30: 819-826.<br />
Brooks, D. R.; M. G. P.Van Veller y D. A. McLennan. 2001.<br />
How to do BPA, really. J. Biogeogr., 28: 343-358.<br />
Brundin, L. 1972. Phylogenetics and biogeography. Syst. Zool.,<br />
21: 69-79.<br />
Ciurana, E. R. 2001. Antropología Hermenéutica (Sobre Dilthey,<br />
Heidegger y Gadamer). Instituto Internacional para el<br />
pensamiento complejo. USAL, El Salvador.<br />
Crisci. J. V. 2001. The voice of historical biogeography. J.<br />
Biogeogr., 28:157-168.<br />
Crother, B. I. 2002. Is Karl Popper's philosophy of science all<br />
things to all people Cladistics, 18: 445.<br />
Csányi, V. 1998. Evolution: modelo or metaphor En:<br />
Evolutionary Systems. Biological and epistemological perspectives on<br />
selection and self-organization. G. Van de Vijver, S. N. Salthe y M.<br />
Delpos (Eds). Kluwer Academic Publishers, pp: 1-12.<br />
de Queiroz, K. y S. Poe 2001. Philosophy and phylogenetic<br />
inference: a comparison of likelihood and parsimony<br />
methods in the context of Karl Popper's writings on<br />
corroboration. Syst. Bio,l. 50: 305-321.<br />
Delpos (Eds). Kluwer Academic Publishers, pp: 215-232.<br />
Dowling, A. P. G. 2002. Testing the accuracy of TreeMap and<br />
Brooks Parsimony Analyses of coevolutionary patterns using<br />
artificial associations Cladistics, 18: 416435.<br />
Ebach, M. C. 2001. Extrapolating cladistic biogeography: a<br />
brief comment on Van Veller et al. (1999, 2000, 2001).<br />
Cladistics, 17: 383-388.<br />
Ebach, M. C. y C. J. Humphries. 2002. Cladistic biogeography<br />
and the art of discovery. J. Biogeogr., 29: 427-444.<br />
Ebach, M. C. y C. J. Humphries. 2003. Ontology of<br />
biogeography. J. Biogeogr., 30: 959-962.<br />
Ebach, M. C.; C. J. Humphries y D. M. Williams. 2003.<br />
Phylogenetic biogeography deconstructed. J. Biogeogr., 30:<br />
1285-1296.<br />
Espinosa, D. y J. Lorente. 1993. Fundamentos de biogeografías<br />
filogenéticas. Universidad Nacional Autónoma de México.<br />
Facultad de Ciencias.<br />
Espinosa, E.; J. J. Morrone; J. Llorente y O. Flores. 2002.<br />
Introducción al análisis de patrones en biogeografía histórica. Facultad<br />
de ciencias, UNAM, México.<br />
Farris, J. S.; A. G. Kluge y J. C. Carpenter. 2001. Popper and<br />
likelihood versus “Popper”. Syst. Biol., 50: 438-443.<br />
Farris, S. J. 2000. Corroboration versus “strongest evidence”.<br />
Cladistics, 16: 385-393.<br />
Fontenla, J. L. 2002. Biogeografía histórica: reflexión sumaria<br />
acerca de conflictos epistemológicos. Cocuyo 12: 22-30.<br />
Gattei, S. 2002. The positive power of negative thinking.<br />
Cladistics 18 : 446452<br />
Goodwin, B. 1998. Las manchas del leopardo. La evolución de la<br />
complejidad. Tusquets editores. Barcelona.<br />
Gould, S. J. 2002. The structure of evolutionary theory. The Library<br />
of Congress. USA.<br />
Grant, T. 2002. Testing methods: the evaluation of discovery<br />
operations in evolutionary biology. Cladistics, 18: 94-111.<br />
Green, M. D.; M. G. P. Van Veller y D. R. Brooks. 2002.<br />
Assessing modes of speciation: range asymmetry and<br />
biogeographical congruence. Cladistics, 18: 112-124.<br />
Grehan, J. R. 2001. Biogeography and Earth history. The<br />
biogeographer, electronic version Vol 2 (1) Fall 2001.<br />
Grehan, J. R. 2003. Empire of the New World. Cladistics, 19:<br />
372-375.<br />
Hennig, W. 1968. Elementos de una sistemática filogenética. Editorial<br />
Universitaria de Buenos Aires.<br />
Hovenkamp. P. 1997. Vicariance events, no areas, should be used<br />
in biogeographical analysis. Cladistics, 13:67-79.<br />
Hovenkamp. P. 2001. A direct method for the analysis of<br />
vicariance patterns. Cladistics 17: 260-265.<br />
Hull, D. 1998. Sujetos centrales y narrativas históricas. En:<br />
Historia y explicación en Biología. (Eds: S. Martínez y A.<br />
Barahour). UNAM, México. Pp: 247-272.<br />
Humphries, C. J. y L. R. Parenti 1999. Cladistic biogeography. 2<br />
Edition. Clarendon Press. Oxford.<br />
Jaramillo, J. M. 1993. Desarrollos recientes en la filosofía de la<br />
ciencia. Tres momentos significativos. En: Praxis filosófica.<br />
Filosofía de la ciencia. Universidad del Valle. Departamento de<br />
Filosofía. Nueva Serie 4: 68-92.<br />
Juarrero, A. 1999. Dynamic in action. Intentional behavior as a<br />
complex system. Bradford Book. the MIT Press. Cambridge,<br />
Massachusetts.<br />
Kampis, G. 1998. Evolution as its own cause and effect. En:<br />
Evolutionary Systems. Biological and epistemological perspectives on<br />
selection and self-organization. G. Van de Vijver, S. N. Salthe y M.<br />
Delpos (Eds). Kluwer Academic Publishers, pp: 255-266.<br />
Kauffman, S. 1993. The origins of order. New York, Oxford.<br />
Oxford University Press.<br />
Kauffman, S. 2000. Investigations. Oxford University Press.<br />
New York, Oxford.<br />
Kauffman, S. 1996. At home in the universe. The search for the laws of<br />
self-organization and complexity. Oxford University Press. New<br />
York, Oxford.<br />
Keller, R. A. 2003. Book Review: The poverty of Linnean<br />
hierarchy: a philosophical study of biological taxonomy.<br />
Cladistics, 19: 173-175.<br />
Kluge, A. G. 1997. Testability and the refutation and<br />
corroboration of cladistic hypotheses. Cladistics, 13: 81-96.<br />
Kluge, A. G. 2002. Distinguishing ''or'' from ''and'' and the case<br />
for historical identification. Cladistics 18: 585-593<br />
Kluge, A.G. 1999. The science of phylogenetic systematics:<br />
explanation, prediction, and test. Cladistics 15: 429-436.<br />
Kluge, A. G. 2001. Philosophical conjectures and their<br />
refutation. Syst. Biol,. 50 322330.<br />
nd<br />
COCUYO 15, 2005 41
Legendre, P. y V. Makarenkov. 2002. Reconstruction of<br />
Biogeographic and Evolutionary Networks Using<br />
Reticulograms. Syst. Biol., 51: 225-238.<br />
Llorente, J. 2000. Una visión histórica de la biogeografía<br />
dispersionista. Caldasia, 22: 161-184.<br />
Llorente, J. y N. Papavero. 1996. La distribución de los seres vivos y la<br />
historia de la Tierra. Ciencia 148. Fondo de Cultura<br />
Económica. DF, México.<br />
Lobo, J. M. 1999. Individualismo y adaptación espacial: un<br />
nuevo enfoque para explicar la distribución geográfica de las<br />
especies. Bol. S.E.A. 26: 561-572.<br />
Martí, J. 1889. Un paseo por la tierra de los Anamitas. En: La<br />
Edad de Oro. Vol 1 no. 4. Ed: Adacosta Gómez,<br />
Administración 77 William Street, New York. Pp: 98-106.<br />
Martín-Piera, F. y Y. San Martín. 1999. Biogeografía de áreas y<br />
biogeografía de artrópodos holárticos y mediterráneos.<br />
Boletín S.E.A., 26: 535560.<br />
Matsuno, K. y S. Salthe. 1995. Global idealism/local<br />
materialism. Biol. Phil., 10: 309-337.<br />
Morin, E. 1984. Ciencia con conciencia. Editorial Anthopos,<br />
Barcelona.<br />
Morrone, J. J. 2004. Homología biogeográfica. Las coordenadas<br />
espaciales de la vida. Instituto de Biología, UNAM. México.<br />
Morrone, J., y J. V. Crisci. 1995. Historical biogeography:<br />
Introduction to methods. Annu. Rev. Ecol. Syst., 26:373-401.<br />
Najmanovich, D. 2002. La complejidad: de los paradigmas a las<br />
figuras del pensar. En: Primer Seminario Bienal Acerca de las<br />
Implicaciones Filosóficas, Epistemológicas y Metodológicas de la Teoría<br />
de la Complejidad” Instituto de Filosofía, La Habana.<br />
filosof@ceniai.inf.cu<br />
Nelson, G. 1978. From Candolle to Croizat: comments of the<br />
history of biogeography. J. Hist. Biol., 11: 269-305.<br />
Nelson, G. y N. Platnick. 1981. Systematics and biogeography:<br />
cladistics and vicariance. Columbia University Press, New York.<br />
Pérez, R. F. 2000. Existe el método cientfíco. Colegio Nacional y<br />
fondo de Cultura Económica. México.<br />
Platnick, N. y G. Nelson. 1978. A method of analysis for<br />
historical biogeography. Syst. Zool., 27: 1-16.<br />
Popper, K., 1990. A World of Propensities. Thoemmes, Bristol.<br />
Real, R. y J. M. Ramírez. 1992. Fundamento filosófico de los<br />
objetivos y métodos de la biogeografía. Monogr, Herpetol., 2:<br />
21-30.<br />
Richards, R. J. 1998. La estructura de la explicación narrativa en<br />
historia y biología. En: Historia y explicación en Biología. (Eds:<br />
S. Martínez y A. Barahour). UNAM, México. Pp: 212-246.<br />
Rield, R. 1998. Dealing with complex systems or how to<br />
decipher language and organisms. En: Evolutionary Systems.<br />
Biological and epistemological perspectives on selection and selforganization.<br />
G. Van de Vijver, S. N. Salthe y M. Delpos (Eds).<br />
Kluwer Academic Publishers, pp: 267-280.<br />
Rieppel, O. 2003. Popper and systematics. Syst. Biol., 2: 259-270.<br />
Rosen, B. R. 1988. Biogeographic patterns: a perceptual<br />
overview. En: Analytical biogeography. an integrated approach to the<br />
study of animal and plant distribution. (A. A. Millers y P. S. Gillers,<br />
eds.). pp: 23-56.<br />
Ruse, M. 2001. The evolutions wars. A guide to the debates. Rutgers<br />
University Press. New Jersey and London.<br />
Salthe, S. N. 1985. Evolving hierarchical systems. New York.<br />
Columbia University Press.<br />
Schuh, R. 2000. Biological systematics. Principles and application.<br />
Cornell University Press. Ithaca and London.<br />
Siddall, M. E. & A. G. Kluge. 1997. Probabilism and<br />
phylogenetic inference. Cladistics, 13: 313-336.<br />
Sotolongo, P. 2002. Bioética y contemporaneidad. Acerca de<br />
algunos fundamentos cosmovisivos y epistemológicos de la<br />
bioética. En: Bioética para la sustentabilidad. (Ed. J. R. Acosta)<br />
Publ. Acuario. Centro Felix Varela. La Habana. Pp: 69-86.<br />
Stamos, D. N. 1996. Popper, falsibiality, and evolutionary<br />
biology. Biol. Phil., 11: 161-191.<br />
Taylor, P. 1986. Historical versus selectionist explanations in<br />
evolutionary biology. Cladistics 3: 1-13<br />
Van de Vijver, G. 1998. Evolutionary systems and the four<br />
causes: a real Aristotelian story. En: Evolutionary Systems.<br />
Biological and epistemological perspectives on selection and selforganization.<br />
G. Van de Vijver, S. N. Salthe y M. Delpos (Eds).<br />
Kluwer Academic Publishers, pp: 243-255.<br />
Van Veller, M. G. P.; D. J. Kornet y M. Zandee. 2000. Methods<br />
in vicariance biogeography: assessment of the<br />
implementations of assumptions zero, 1 and 2. Cladistics 16:<br />
319-345.<br />
Van Veller, M. G. P.; D. J. Kornet y M. Zandee. 2002. A posteriori<br />
and a priori methodologies for testing hypotheses of causal<br />
processes in vicariante biogeography. Cladistics, 18: 207-217.<br />
Van Veller, M. G. P.; D. R. Brooks y M. Zandee. 2003. Cladistic<br />
and phylogenetic biogeography: the art and the science. J.<br />
Biogeogr., 30: 319-329.<br />
Van Veller, M. G. P.; M. Zandee y D. J. Kornet. 2001. Measures<br />
for obtaining inclusive sets of area cladograms under<br />
assumptions zero, 1, and 2 with different methods for<br />
vicariance biogeography. Cladistics 17: 248-259.<br />
Van Veller, M. G. P.; M. Zandee y D. J. Kornet. 1999. Two<br />
requirements for obtaining valid common patterns under<br />
different assumptions in vicariance biogeography. Cladistics<br />
15: 393-406.<br />
Van Veller, M. G. y D. R. Brooks. 2001. When simplicity is not<br />
parsimonious: a priori and a posteriori methods in historical<br />
biogeography. J. Biogeogr., 28: 1-11.<br />
Van Welzen, P. C.; H. Turner y P. Hovenkamp. 2003. Historical<br />
biogeography of Southeast Asia and the west Pacific, or the<br />
generality of unrooted area networks as historical<br />
biogeographic hypotheses. J. Biogeogr., 30: 181-192.<br />
Vargas, J. M. 1993. Escuelas y tendencias en biogeografía<br />
histórica. Monogr, Herpetol., 2: 107-136.<br />
Wilkinson, D. M. 2001. Dispersal biogeography. Encyclopedia of<br />
life science. Http://www.els.net.<br />
Wilkinson, D. M. 2003. Dispersal, cladistics and the nature of<br />
biogeography. J. Biogeogr., 30: 1779-1780.<br />
Woods C. A. y F. E. Sergile (eds). 2001. Biogeography of the West<br />
Indies. Patterns and perspectives. CRC Press<br />
42 COCUYO 15, 2005
APÉNDICE<br />
Ejemplo de BPA secundario (Brooks et al., 2001). La Fig.<br />
representa un árbol filogenético para las especies hipotéticas 1-<br />
6. Los nodos internos del cladograma se asumen como<br />
antecesores hipotéticos y se numeran para proceder a su<br />
codificación.<br />
Tabla 1<br />
Áreas Taxa Codificación Binaria<br />
A 1 10000000001<br />
B 2,5,6 01001111111<br />
C 3 00100000111<br />
D 4 00010001111<br />
La Tabla 1 muestra la distribución de las especies en las áreas A-<br />
D y su codificación. Las áreas se codifican de acuerdo con la<br />
presencia (1) o ausencia (0) en ellas de especies o sus<br />
antecesores hipotéticos. Luego se realiza un análisis de<br />
parsimonia y se obtiene un cladograma de áreas.<br />
En la siguiente Fig. se observa que el área B contiene tres especies que no descienden de un antecesor común inmediato, por<br />
lo que falsifican la hipótesis de la vicarianza. Así, se multiplica el área B para cada una de las especies 2, 5 y 7, y se construye<br />
una nueva matriz de distribución (Tabla 2).<br />
Tabla 2<br />
Áreas Taxa Código Binario<br />
A 1 10000000001<br />
B 1 2 01000000011<br />
C 3 00100000111<br />
D 4 00010001111<br />
B 2 5 00001011111<br />
B 3 6 00000111111<br />
Se realiza un nuevo análisis de parsimonia, el cual constituye<br />
el BPA secundario. El área B 1 se encuentra en una posición<br />
relativamente basal, mientras que B 2+ B 3son áreas hermanas<br />
en una posición relativamente derivada.<br />
Brooks et al. (2001) concluyen que las especies 5 y 6 ocurren en<br />
el área B, porque su antecesor, la especie 7, que conecta las áreas<br />
B 2+ B 3 , se dispersó desde el área D hacia la B, donde especió y<br />
dio origen a 5 y 6.<br />
Es posible arribar a la misma conclusión desde el BPA primario.<br />
B es un área basal en el cladograma. Por lo tanto, lo más<br />
parsimonioso es asumir que el antecesor común de las especies<br />
5 y 6, que desciende de la especie 8, a su vez antecesor común de<br />
4 y 7, se originó en el área D, desde donde se dispersó hacia la B.<br />
donde especiaron 5 y 6.<br />
COCUYO 15, 2005 43
LITERATURA<br />
RECIENTE<br />
Alayón, G. 2004. Notas sobre la familia Ctenidae (Arachnida:<br />
Araneae) en La Hispaniola, con la descripción de tres nuevas<br />
especies. Rev. Ibérica Aracnología 9: 277-283.<br />
Alayón, G. 2004. Especie nueva de Camillina (Araneae:<br />
Gnaphosidae) del occidente de Cuba Solenodon 4: 27-29.<br />
Albert, S. H & C. A. Núñez. 2004. Distribution of the Asian<br />
citrus psyllid, Diaphorina citri Kuwayama (Rhynchota:<br />
Psyllidae) in the Caribbean basin. Florida Entomologist 87(3):<br />
401-402.<br />
Andrade de, M. L. 1998. First description of fossil Acanthostichus<br />
from Dominican amber (Hymenoptera: Formicidae). Mitt.<br />
Schweiz. Entomol. Ges., 71: 269-274.<br />
Andrade de, M. L. 1998. Fossil and extant species of<br />
Cylindromyrmex (Hymenoptera: Formicidae). Rev. Suisse Zool.<br />
105: 581-664.<br />
Andrade de, M. L. 2001. A remarkable Dominican amber species<br />
of Cylindromyrmex with Brazilian affinities and additions to the<br />
generic revision (Hymenoptera: Formicidae). Beitr. Entomol.,<br />
51: 51-63.<br />
Andrade de, M. L. 2003. First descriptions of two new species<br />
amber species of Cyphomyrmex from Mexico and the<br />
Dominican Republic (Hymenoptera: Formicidae). Beitr.<br />
Entomol., 53: 131-139.<br />
Andrade de, M. L., C. Baroni Urbani & C. R. F. Brandão. 1999.<br />
Two new species of Leptothorax "Nesomyrmex" fossils in<br />
Dominican amber (Hymenoptera: Formicidae). Beitr.<br />
Entomol., 49(1): 133-140.<br />
Backer, M. S. et al. 2005 Lepidopteran soral crypsis on Caribbean<br />
Ferns. Biotropica 37(2): 314316.<br />
Baranowski, R. M. & J. Brambila 2001. Caymanis, a new genus of<br />
Antillocorini from the Cayman islands (Hemiptera:<br />
Lygaeidae). Florida Entomologist 84: 117-118.<br />
Becker, O. V. 1999. Family reassignments and synonymy of<br />
some taxa of Neotropical Microlepidoptera. Rev. Brasileira<br />
Zool., 16: 141-170.<br />
Becker, O. V. 2001. A new Antillean moth of the genus<br />
Enigmogramma Lafontaine & Poole (Noctuidae, Plussinae).<br />
Rev. Brasileira Zool., 18: 145-148.<br />
Becker, O. V. 2002. The Noctuoidea (Lepidoptera) from Cuba<br />
described by Herrich-Schaffer and Gundlach in the Gundlch<br />
collection, Havana. Rev. Brasileira Zool., 19: 349-392.<br />
Becker, O. V. 2002. The Geometroidea (Lepidoptera) from Cuba<br />
described by Herrich-Schaffer in the Gundlch collection,<br />
Havana. Rev. Brasileira Zool., 19: 393-418.<br />
Braby, M. F. 2005. Provisional checklist of genera of the Pieridae<br />
(Lepidoptera: Papilionoidea). Zootaxa 832: 116.<br />
Brooks, S. E. 2005. Systematics and phylogeny of<br />
Dolichopodinae (Diptera: Dolichopodidae). Zootaxa 857:<br />
1158.<br />
Carvalho, C.J.B. de; M.S. Couri; A.C. Pont; D. Pamplona & S.M.<br />
Lopes. 2005. A Catalogue of the Muscidae (Diptera) of the<br />
Neotropical Region. Zootaxa 860: 1282.<br />
Cervantes Peredo, L. 2004. Cholula minuta, a new species of<br />
Myodochini (Lygaeoidea: Rhyparochromidae) from Jamaica.<br />
Florida Entomologist: 173-175.<br />
Davies, N. & E. Bermingham. 2002. The historical<br />
biogeography of two Caribbean butterflies (Lepidoptera:<br />
Heliconidae) as inferred from genetic variation at multiple<br />
loci. Evolution 56: 573- 589.<br />
Eger, Jr, J. E. & R. M. Baranowski. 2002. Diolcus variegatus<br />
(Heteroptera: Pentatomoidea: Scutelleridae), a Caribbean<br />
species established in South Florida. Florida Entomologist<br />
85:267.<br />
Engel, M. 2004. On the identity of Halictus cubensis Spinola, 1851<br />
(Hymenoptera: Halictidae). Entomological News 115: 169-<br />
170.<br />
Falin, Z. H. 2004. Revision of three New World Macrosiagon<br />
Hentz species (Coleoptera: Ripiphoridae: Ripiphorinae) with a<br />
discussion of phylogenetic relationships within the<br />
Macrosiagonini. The Coleopterist Bull., 58: 1-19.<br />
Fernández-Triana, J. L. 2005. The taxonomy and biogeography<br />
of Cuban Ophioninae (Hymenoptera: Ichneumonidae).<br />
Zootaxa 1007: 1-60.<br />
García, N. & J. L. Fontenla. 2004. Especies nuevas de nemátodos<br />
(Nematoda: Rhigonematida) asociados con Anadenobolus a.<br />
arboreus (Diplopoda:Rhinocricidae) de Puerto Rico<br />
Solenodon 4: 6-10.<br />
Garrido, O. H. 2004.Especie nueva que representa el primer<br />
registro del género Diastolinus<br />
(Coleoptera:Tenebrionidae, Pedinini) para Jamaica.<br />
Solenodon 4:37-39.<br />
Garrido, O. H. 2004.Tres especies nuevas de Diastolinus<br />
(Coleoptera:Tenebrionidae, Pedinini) para la Hispaniola.<br />
Solenodon 4:40-45.<br />
Garrido, O. H. 2004.Especie nueva de Diastolinus<br />
(Coleoptera:Tenebrionidae)para Cuba. Solenodon 4:46-48.<br />
Garrido, O. H. 2004.Comentarios sobre el género Strongylium<br />
(Coleoptera: Tenebrionidae,<br />
Strongyliini), con la descripción de tres especies nuevas para<br />
Cuba Solenodon 4:49-55.<br />
Garrido, O. H. 2004. Dos especies nuevas de Meralius<br />
(Coleoptera: Zopheridae) de Cuba, con comentarios sobre<br />
el género. Solenodon 4: 56-62.<br />
Garrido, O. H. & E. Gutiérrez. 2004. A new species of Talanus<br />
(Coleoptera:Tenebrionidae) from Puerto Rico, with<br />
commentaries on Talanus cribrarius Solenodon 4: 63-66.<br />
Genaro, J. A. 2004. Dos especies nuevas de euméninos de los<br />
géneros Euodynerus y Parancistrocerus para Cuba<br />
(Hymenoptera: Vespidae). Solenodon 4: 67-73.<br />
Genaro, J. A. 2004. A new species of Cerceris from Hispaniola,<br />
West Indies (Hymenoptera: Crabronidae: Philanthinae). J.<br />
Kansas Entomol. Soc., 77: 761764.<br />
González, J. M.; J. A. Genaro & R. W. Matthews. 2004. Species<br />
44 COCUYO 15, 2005
of Melittobia (Hymenoptera:Eulophidae) established in Cuba,<br />
Costa Rica and Hispaniola. Florida Entomol. 87: 619-620.<br />
Guidetti, R. & R. Bertolani. 2005. Tardigrade taxonomy: an<br />
updated check list of the taxa and a list of characters for their<br />
identification. Zootaxa 845: 146.<br />
Gutiérrez, E. 2004. Las cucarachas del género Eurycotis<br />
(Dictyoptera: Blattaria: Blattidae)en Puerto Rico, con la<br />
descripción de una especie nueva Solenodon 4: 30-36.<br />
Kondratieff, B. C. & D. E. Perez-Gelabert. 2004. A new species<br />
of Nemomydas and a new record for Baliomydas gracilis (Diptera:<br />
Mydidae) from Hispaniola. Florida Entomol., 87: 380-382.<br />
Krecek J. & R. H. Scheffrahn. 2001 Neotermes platyfrons, a new<br />
dampwood termite (Isoptera, Kalotermitidae) from the<br />
Dominican Republic. Florida Entomologist 84: 70-76.<br />
Leistikow, A. & J.W. Wagele. 1999. Checklist of the terrestrial<br />
isopods of the New World (Crustacea, Isopoda, Oniscidea).<br />
Rev. Brasileira Zool., 16: 1-72.<br />
Lorente, J. & J. J. Morrone (eds). 2001. Introduccion a la<br />
biogeografia en Latinoamerica, teorias, conceptos, metodos<br />
y aplicaciones. Las Prensas de Ciencias, Fac. Ciencias,<br />
UNAM. 277 pp.<br />
Maceira F., D. 2003. Las especies de la familia Veronicellidae<br />
(Mollusca, Soleolifera) en Cuba. Revista Biol. Tropical 51<br />
(Suppl. 3): 453-461.<br />
Manley, D. G. & G. O. Poinar. 2003. A new specimens of fossil<br />
Mutillidae (Hymenoptera) from Dominican amber. Proc.<br />
Entomol. Soc. Washington 105: 1069-1071.<br />
Metz, M. A. & M. E. Irwin. 2002. Revision of Lindneria Kröber,<br />
with the description of Two new genera Insulatitan and<br />
Ambradolon, a fossil from Dominican amber<br />
(Diptera:Therevidae:Therevinae). Ann. Entomol. Soc.<br />
America 93: 977-1018.<br />
Molina-Ochoa, J.; J. E. Carpenter; E. A. Heinrichs & J. E. Foster.<br />
2003. Parasitoids and parasites of Spodoptera frugiperda<br />
(Lepidoptera: Noctuidae) in the Americas and Caribbean<br />
basin: an inventory. Florida Entomologist 86: 254-289.<br />
Morrone J. J. & J. Lorente (eds). 2003. Una perspectiva<br />
latinoamericana de la biogeografia. Las Prensas de Ciencias,<br />
Fac. Ciencias, UNAM. 307 pp.<br />
Moure, J. S. & D. Urban. 2002. Catálogo de Apoidea da Região<br />
Neotropical (Hymenoptera, Colletidae). III. Colletini. Rev.<br />
Brasileira Zool., 19: 1-30.<br />
Myartseva, S. N. & J. M. Coronado-Blanco. 2002. A new<br />
parasitoid of whiteflies from Mexico, with a key to new world<br />
species of the genus Encarsiella (Hymenoptera:<br />
Aphelinidae).Florida Entomologist 85: 620-624.<br />
Nelson, G. H. & C. L. Bellamy. 2004. A revision of the genus<br />
Paratyndaris Fisher, 1919 (Coleoptera: Buprestidae:<br />
Polycestinae). Zootaxa 683: 1-80.<br />
Olmi. M. 2004. Embolemus kheeli: a new species of Embolemidae<br />
(Hymenoptera: Chrysidoidea) from the Dominican<br />
Republic. Zootaxa 500: 16.<br />
Olmi. M. & E. Portuondo. 2003. A new species of Embolemus<br />
from Cuba (Hymenoptera, Embolemidae). Entomologist´s<br />
Monthly Magazine 139: 135-137.<br />
Ortíz, M.; R. Lalana & C. Varela 2004. Especie nueva del género<br />
Sphaeromopsis (Crustacea: Isopoda:Sphaeromatidae) de la<br />
plataforma Sur oriental de Cuba Solenodon 4: 1-5<br />
Peck, S. B. 2005. A checklist of the beetles of Cuba with data on<br />
distributions and bionomics (Insects: Coleoptera).<br />
Arthropods of Florida and Neighbouring Land Areas 18: 1-<br />
241.<br />
Pérez-Asso, A. R. 2004. Dos géneros nuevos y cinco especies<br />
nuevas de milpiés (Diplopoda:Spirobolida: Rhinocricidae)<br />
para la Hispaniola Solenodon 4: 11-26.<br />
Pérez-Gelabert, D. E. 2004. The mantids of Hispaniola, with<br />
description of two new species (Mantodea). Bol. Soc.<br />
Entomol. Aragonesa 34: 35-48.<br />
Pérez-Gelabert, D. E. 2004. En torno a la presencia de Melanoplus<br />
sanguinipes (Orthoptera: Acrididae) en República Dominicana.<br />
Bol. Soc. Entomol. Aragonesa 34: 221.<br />
Pérez-Gelabert, D. E. 2003. Antillotettix nanus, a new genus and<br />
species of tetrigid (Orthoptera: Tetrigidae: Cladonotinae)<br />
from Dominican Republic, Hispaniola. J. Orthoptera<br />
Research 12(2): 111-114.<br />
Pérez-Gelabert, D. E. & D. Otte. 2004. Melanoplus sanguinipes<br />
(Fabricius) (Orthoptera: Acrididae: Melanoplinae), a pest<br />
grasshopper introduced to Dominican Republic, Hispaniola.<br />
Trans. American Entomol. Soc., 129: 543-548.<br />
Pérez-Gelabert, D. E. & R. H. Bastardo. 2004. Synoditella bisulcata<br />
(Kieffer) (Hymenoptera: Scelionidae) parasitizing Orphulella<br />
punctata (DeGeer) (Orthoptera: Acrididae) in Dominican<br />
Republic. Proc. Entomol. Soc. Washington 106: 931-932.<br />
Platnick, N. I. & D. Penney. 2004. A revision of the widespread<br />
spider genus Zimiris (Araneae, Prodidomidae). American<br />
Museum Novitates 3450: 112.<br />
Razowski, J. & O. V. Becker. 1999. A review of the New World<br />
Chlidanotini (Lepidoptera, Tortricidae). Rev. Brasileira Zool.,<br />
16: 1149-1182.<br />
Rightmyer, M. G. 2004. Phylogeny and classification of the<br />
parasitic bee tribe Epeolini (Hymenoptera: Apidae,<br />
Nomadinae). Sci. Papers Nat. Hist. Mus. Univ. Kansas 33: 1-<br />
51.<br />
Robbins, R.K. & G. Lamas. 2002. Nomenclatural Changes in the<br />
Neotropical Eumaeini (Lepidoptera, Lycaenidae,<br />
Theclinae). Revta Bras. Zool. 19 (Supl. 1):197-214.<br />
Roth, L. M. 2003. Systematics and phylogeny of cockroaches<br />
(Dictyoptera: Blattaria). Oriental Insects 37: 1-186.<br />
Sanborn, A. F. 2001. Distribution of the cicadas (Homoptera:<br />
Cicadidae) of the Bahamas. Florida Entomologist 84: 733-<br />
734.<br />
Scheffrahn, R. H.; J. Krecek & N. Y. Su . 2000. Redescriptions of<br />
the dampwood termites Neotermes jouteli and N. luykxi<br />
(Isoptera: Kalotermitidae) from Florida, Cuba, Bahamas,<br />
and Turks and Caicos Islands. Ann. Entomol. Soc. America<br />
93: 785-794.<br />
Schiefer, T. L. 2004. A. new record of an endemic Cuban tiger<br />
beetle, Cicindela (Brasiella) viridicollis (Coleoptera: Carabidae:<br />
Cicindelinae), from the Florida keys. Florida Entomologist<br />
87: 551-553.<br />
Schuh, R. T. & M. D. Schwartz. 2004. New genera, new species,<br />
new synonyms, and new combinations in North America<br />
COCUYO 15, 2005 45
and Caribbean Phylinae (Heteroptera: Miridae). American<br />
Museum Novitates 3436: 136.<br />
Sikes, D. S. & S. B. Peck. 2000. Description of Nicrophorus<br />
hispaniola, new species, from Hispaniola (Coleoptera:<br />
Silphidae) and a key to the species of Nicrophorus of the New<br />
World. Ann. Entomol. Soc. America 93: 391-397.<br />
Slater, J. A. & R. M. Baranowski. 2000. Discocoris dominicanus, a<br />
new species of palm bug from dominican amber.<br />
(Heteroptera: Thaumastocoridae). Florida Entomologist 83:<br />
349-353.<br />
Slater, J. A. & R. M. Baranowski. 2001. Melanopleuroides<br />
dominicanus, a new lygaeine genus and species from the<br />
Dominican Republic (Heteroptera: Lygaeidae) Florida<br />
Entomologist 84: 131-132.<br />
Snelling, R. R. & J. A. Torres. 2004. The spider wasps of Puerto<br />
Rico and the British Virgin Islands (Hymenoptera:<br />
Pompilidae). J. Kansas Entomol. Soc., 77: 356376.<br />
Stange, L. 2004. A systematic catalog, bibliography and<br />
classification of the world antlions (Insecta: Neuroptera:<br />
Myrmeleontidae). Mem. American Entomol. Inst., 74: 1-565.<br />
Szwedo, J.; T. Bourgoin & F. Lefebvre. 2004. Fossil planthoppers<br />
(Hemiptera: Fulgoromorpha) of the world. An annotated<br />
catalogue with notes on Hemiptera classification. Studio 1,<br />
Warsaw, Polonia. 199 pp.<br />
Urban, D. & J. S. Moure. 2001. Catálogo de Apoidea da Região<br />
Neotropical (Hymenoptera, Colletidae). II. Diphaglossinae.<br />
Rev. Brasileira Zool., 18: 1-34.<br />
Urban, D. & J. S. Moure. 2002. Catálogo de Apoidea da Região<br />
Neotropical (Hymenoptera, Colletidae). IV. Hylaeinae. Rev.<br />
Brasileira Zool., 19: 31-56.<br />
Vardy, C. R. 2000. The New World tarantula-hawk wasp genus<br />
Pepsis Fabricius (Hymenoptera: Pompilidae). Part. 1.<br />
Introduction and the P. rubra species group. Zooologische<br />
Verhandelingen 332: 1-86.<br />
Vardy, C. R. 2002. The New World tarantula-hawk wasp genus<br />
Pepsis Fabricius (Hymenoptera: Pompilidae). Part. 2. The P.<br />
grossa to P. deaurata groups. Zooologische Verhandelingen<br />
338: 1-135.<br />
Wetterer, J. K. & C. O. Brandon. 2002. Ants (Hymenoptera:<br />
Formicidae) of the Dry Tortugas, the outermost Florida keys.<br />
Florida Entomologist 85(2): 303-307.<br />
Whitfield, J. B. & M. T. Oltra. The Neotropical species of<br />
Deuterixys Mason (Hymenoptera: Braconidae). J.<br />
Hymenoptera Research 13: 134-148.<br />
Yoder, M. J. 2004. Revision of the North American species of<br />
the genus Entomacis (Hymenoptera: Diapriidae). Tha<br />
Canadian Entomol., 136: 323-405.<br />
Zavortink, T. J. & G. O. Poinar, J. 2000. Anopheles (Nyssorhynchus)<br />
dominicanus sp. n. (Diptera: Culicidae) from Dominican<br />
Amber. Ann. Entomol. Soc. America 93: 1230-1235.<br />
46 COCUYO 15, 2005