К фауне Janiridae (Isopoda, Asellota) морей СССР
К фауне Janiridae (Isopoda, Asellota) морей СССР
К фауне Janiridae (Isopoda, Asellota) морей СССР

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
ТРУДЫ ЗООЛОГИЧЕС<strong>К</strong>ОГО ИНСТИТУТА А<strong>К</strong>АДЕМИИ НАУ<strong>К</strong> <strong>СССР</strong><br />
1962 XXX<br />
О. Г. <strong>К</strong>усакан<br />
<strong>К</strong> <strong>фауне</strong> <strong>Janiridae</strong> (<strong>Isopoda</strong>, <strong>Asellota</strong>) <strong>морей</strong> <strong>СССР</strong><br />
О. G. Kussakin. ON THE FAUNA OF JANIRIDAE (ISOPODA, ASELLOTA)<br />
FROM THE SEAS OF THE USSR<br />
Настоящая статья представляет собой результат обработки материалов<br />
из дальневосточных и северных <strong>морей</strong> <strong>СССР</strong>, собранных различными<br />
экспедициями и хранящихся в коллекциях ЗИН АН <strong>СССР</strong>. Всего было<br />
просмотрено более 1740 экз., относящихся к 3 родам семейства <strong>Janiridae</strong>:<br />
Janiralata Menzies, laniropsis G. О. Sars и Caecijaera Menzies. Из 16 изученных<br />
видов 9 являются новыми для науки. <strong>К</strong>роме того, в статью включено<br />
описание нового вида Janiralata из Японии, также хранящегося<br />
в коллекциях ЗИН АН <strong>СССР</strong>. Один из трех рассматриваемых родов —<br />
Caecijaera впервые отмечается для фауны Советского Союза. Другие роды<br />
этого семейства ревизии не подвергались.<br />
На рисунках приняты следующие обозначения:<br />
Ant I — антенна I. Рр I — перейопод I пары.<br />
Ant II — антенна II. Ор $> — крышечка (II плеопод самки).<br />
Md — мандибула. Ор 6 — крышечка (I плеопод самца)-<br />
Мх I — максилла I. Pip II —V — плеоподы II—V нар.<br />
Мх II — максилла II. Up — уропод.<br />
Мхр — мансилллпеда (ногочелюсть). 1. т. II (IV) t. s. — боковой край II<br />
или IV грудного сегмента.<br />
Тотальные рисунки новых видов выполнены художником Б. А. Гирсгуном.<br />
Род JANIRALATA MENZIES<br />
Род Janiralata был выделен Р. Мензисом в 1951 г. и в первоначальном<br />
объеме включал 10 северотихоокеанских видов, многие из которых относились<br />
ранее к родам Ianira Leach и Jolella Richardson. Просмотр определенных<br />
и неопределенных коллекций ЗИН АН <strong>СССР</strong> позволил нам<br />
изучить большую часть видов, которых не имел в своем распоряжении<br />
Р. Мензис, а также прибавить к списку видов этого рода несколько новых.<br />
Для сравнения были просмотрены также имеющиеся в коллекциях ЗИН<br />
Ianira alta (Stimpson), /. maculosa Leach и Jolella laciniata (G. 0. Sars).<br />
В результате проделанной ревизии подтвердилась правильность выделения<br />
Р. Мензисом особого рода Janiralata, однако число признаков, отличающих<br />
этот род от других, значительно сократилось. Наиболее стойкими<br />
признаками, характеризующими род Janiralata, оказались наличие<br />
зубчиков на проксимальной части внутреннего края проподита I перейопода<br />
и направленных в стороны, довольно длинных задне-боковых отростков<br />
I плеопода самца. Другие признаки, как например наличие видимых<br />
сверху коксальных пластинок на II—VII грудных сегментах,<br />
2 Труды зин: т. ххх
— 18 —<br />
вогнутости на заднем крае II плеопода самки и диагонального ряда щетинок<br />
в ди стальной трети 2-го членика щупика мандибул, или выражены<br />
не у всех видов рода Janiralata, или же встречаются у представителей<br />
других родов и не могут служить родовыми диагностическими признаками.<br />
В результате обработки материала установлено также, что Ianira<br />
tricornis (<strong>К</strong>геуег) должна быть перенесена в род Janiralata; J. alascensis<br />
(Benedict), возможно, является синонимом /. tricornis; японские виды<br />
Jolella chuni Thielemann и Janiralata shiinoi sp. п. значительно ближе<br />
к видам рода Janiralata, чем к Ianira или Jolella. С 6 новыми число видов,<br />
входящих в род Janiralata, увеличилось до 17 (если считать/, alascensis<br />
синонимом /, tricornis). Из 17 видов 14 являются тихоокеанскими бореальными,<br />
2 вида (/. chuni и J. shiinoi) — западнотихоокеанскими субтропическими<br />
и 1 (/. tricornis) — арктическим, заходящим в тихоокеанские<br />
бореальные воды. В целом род Janiralata имеет несомненно тихоокеанское<br />
бореальное происхождение.<br />
Диагноз рода по Мензису. 1—3-й членики щупика ногочелюстей расширенные,<br />
примерно такой же ширины, как внутренняя пластинка, 4-й<br />
и 5-й членики щупика узкие; внутренняя пластинка с 2—4 соединительными<br />
крючками. Спинная поверхность тела лишена шипов. <strong>К</strong>оксальные<br />
пластинки на II—VII грудных сегментах имеются и обычно видны сверху,<br />
реже не видны на II—III сегментах. Глаза имеются, расположены на<br />
спинной стороне головы, приближены к ее боковым краям. II антенна<br />
с чешуйкой; ее жгутик длинный, многочлениковый. I плеопод самца с длинными,<br />
направленными в стороны задне-боковыми отростками, как у Ianiropsis,<br />
обычно несет на дистальном конце более или менее развитые лопасти<br />
или выросты, усаженные по краям щетинками. Экзоподит II плеопода<br />
самца с широким, часто как бы срезанным задним краем, несущим<br />
щетинки. II плеопод самки широкоовальный или округлый, обычно<br />
со слегка вогнутым посредине, реже выпуклым задним краем. Уроподы<br />
с 2 ветвями, превышают половину длины плеотельсона; внутренняя ветвь<br />
такой же длины или несколько длиннее наружной. Зубной отросток цилиндрической<br />
формы, щупик 3-члениковый и несет на ди стальной трети<br />
2-го членика 2 большие зазубренные щетинки и диагональный ряд щетинок<br />
меньшего размера; 3-й членик щупика скручен. Проксимальная<br />
часть внутреннего края проподита I перейопода зазубрена; карпоподит<br />
I перейопода расширеи и несет значительное количество раздвоенных<br />
шипов по внутреннему краю; дактилоподит с 2 коготками. Дактилоподиты<br />
II—VII перейоподов с 3 коготками. Брюшной отдел состоит из короткого<br />
переднего сегмента и крупного заднего, или плеотельсона.<br />
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА<br />
JANIRALATA MENZ1ES<br />
1 (2). Рострум длинный, значительно превышает половину длины головы;<br />
края плеотельсона бахромчато изрезаны. Арктика, Берингово<br />
море и побережье сев. <strong>К</strong>урильских о-вов<br />
J. tricornis (<strong>К</strong>гоуег, 1846—1849).<br />
2 (1). Рострум, если имеется, не превышает половину длины головы;<br />
края плеотельсона гладкие.<br />
3 (10). Задне-боковые утлы плеотельсона плавно закруглены, лишены<br />
лопастей.<br />
4 (5). Передне-боковые отростки головы отсутствуют. <strong>К</strong>алифорния . .<br />
J. rajata Menzies (1951b).<br />
5 (4). Передне-боковые отростки головы имеются, треугольной формы.
— 19 —<br />
6 (7). Рострум очень короткий, широкий; коксальные пластинки II<br />
и III грудных сегментов видны сверху; задний край крышечки<br />
самки слегка вогнут посредине. <strong>К</strong>алифорния .<br />
J. davisi Menzies (1951b).<br />
7 (6). Рострум не выражен; коксальные пластинки II и III грудных<br />
сегментов не видны сверху; задний край крышечки самки слегка<br />
выпуклый.<br />
8 (9). Спинная поверхность тела имеет скульптуру в виде маленьких<br />
бугорков на голове, 3 передних грудных сегментах и плеотельсоне<br />
и вдавлений на 4 задних грудных сегментах. Тихий океан,<br />
побережье о. Хонсю J. chuni (Thielemann, 1910).<br />
9 (8). Спинная поверхность тела лишена скульптуры. Восточно-<strong>К</strong>итайское<br />
море, у о. <strong>К</strong>юсю J. shiinoi Kussakin, sp. и.<br />
10 (3). Задне-боковые углы плеотельсона образуют лопасти или отростки.<br />
11 (22). Задне-боковые углы плеотельсона образуют широкие, сзади закругленные,<br />
реже тупо заостренные лопасти, примерно такой же<br />
ширины, как медиальная лопасть.<br />
12 (17). Боковые и медиальная лопасти заднего края плеотельсона примерно<br />
равной длины, короткие, сзади плавно закруглены.<br />
13 (14). Рострум хорошо развит, выдается за передне-боковые отростки<br />
головы, треугольной формы. Тихий океан, побережье южн. <strong>К</strong>урильских<br />
о-вов . . . . J. kurilensis Kussakin, sp. п.<br />
14 (13). Рострум почти не выражен, вместо него имеется лишь маленькое<br />
заострение или выпуклость посредине переднего края головы,<br />
15 (16). Выпуклый передний край головы слегка заострен посредине;<br />
передняя лопасть коксальной пластинки II грудного сегмента значительно<br />
короче передней лопасти сегмента и примерно такой же длины,<br />
как его задняя лопасть; медиальные лопасти дистального края<br />
I плеопода самца ясно выражены, каждая из них посредине слегка<br />
вогнута; задний край крышечки самки вогнут посредине. Берингово<br />
море и Тихий океан, у вост. побережья <strong>К</strong>амчатки и сев. <strong>К</strong>урильских<br />
о-вов J. erostrata (Richardson, 18ЭЭ).<br />
16 (15). Выпуклый передний край головы закруглен посредине; передняя<br />
лопасть коксальной пластинки II грудного сегмента почти<br />
равна длине передней лопасти сегмента и значительно длиннее<br />
задней лопасти сегмента; медиальные лопасти дистального края I<br />
плеопода самца слабо обособлены; крышечка самки со слегка выпуклым<br />
задним краем. Японское и Охотское моря<br />
J. soldatovi (Gurjanova, 1933а, 19336).<br />
17 (12). Боковые лопасти заднего края плеотельсона значительно длиннее<br />
медиальной, сзади закруглены или тупо заострены.<br />
18 (21). Рострум очень короткий, значительно не достигает уровня<br />
краев передне-боковых отростков головы или почти не выражен<br />
вовсе.<br />
19 (20). Рострум почтя не выражен, передний край головы посредине<br />
образует лишь закругленную спереди выпуклость; длина медиальной<br />
лопасти заднего края плеотельсона составляет несколько более<br />
половины длины боковых; вутренняя пластинка ногочелюстей с 4<br />
соединительными крючками. Тихий океан, побережье южн. <strong>К</strong>урильских<br />
о-вов J. gurjanovae Kussakin, sp. п.<br />
20 (19). Рострум, хотя и очень короткий, но ясно намечен, спереди тупо<br />
заострен; длина медиальной лопасти плеотельсона составляет менее<br />
половины длины боковых; внутренняя пластинка ногочелюстей<br />
с 3 соединительными крючками. Охотское море -<br />
.о J. ochotensis Kussakin, sp. п.<br />
2*
— 20 —<br />
21 (18). Рострум короткий, но хорошо выражен, треугольной формы,<br />
почти достигает уровня краев передне-боковых отростков головы.<br />
Охотское море и Тихий океан, у сев. <strong>К</strong>урильских о-вов . . . .<br />
J. vitjazi Kussakin, sp. п.<br />
22 (11). Задне-боковые углы плеотельсона вытянуты в заостренные<br />
отростки, значительно более узкие, чем медиальная лопасть.<br />
23 (24). Боковые края головы с каждой стороны снабжены глубокой<br />
вырезкой. Тихий океан, <strong>К</strong>алифорния . . . .<br />
J. triangulata (Richardson, 1899).<br />
24 (23). Боковые края головы без вырезок.<br />
25 (26). Передне-боковые углы головы раздвоены, образуя по два небольших<br />
отростка с каждой стороны; Тихий океан, юго-вост. Аляска<br />
, , . J. holmesi (Richardson, 1905b).<br />
26 (25). Передне-боковые углы головы тупые или снабжены отростками,<br />
но цельные, не раздвоенные.<br />
27 (28). Передне-боковые отростки головы почти не выражены. Берингово<br />
море, Алеутские о-ва J. sarsi (Richardson, 1905а).<br />
28 (27). Передне-боковые отростки головы хорошо выражены, более или<br />
менее треугольной формы.<br />
29 (30). Рострум хорошо развит, удлиненный, спереди заостренный.<br />
Тихий океан, побережье Сев. Америки от Аляски до Вашингтона<br />
J. solasteri (Hatch, 1947).<br />
30 (29). Рострум очень короткий, широкотреугольный или почти не<br />
выражен.<br />
31 (32), Задне-боковые отростки плеотельсона узкие, шиповидные, загнуты<br />
несколько внутрь, значительно не достигают заднего края<br />
медиальной лопасти; передне-боковые отростки головы не достигают<br />
уровня переднего края головы; боковые края I грудного сегмента<br />
трехлопастные. Тихий океан, побережье Сев. Америки от Вашингтона<br />
до <strong>К</strong>алифорнии J. occidentalis (Walker, 1898).<br />
32 (31). Задне-боковые отростки плеотельсона сравнительно широкие,<br />
треугольные, направлены назад, значительно выдаются за край<br />
медиальной лопасти; передне-боковые отростки головы выдаются<br />
вперед за уровень переднего края головы в средней части; боковые<br />
края I грудного сегмента двухлопастные. Тихий океан, побережье<br />
<strong>К</strong>урильских о-вов и юго-вост. <strong>К</strong>амчатки<br />
J. pilosa Kussakin, sp. п.<br />
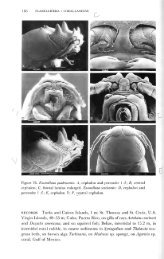
Janiralata tricornis (<strong>К</strong>тоует) (рис. 1)<br />
Henopomus tricornis <strong>К</strong> г в у е г, 1846—1849:372—379, 380; 1849 : табл. XXX,<br />
фиг. 2а—q. — Janira tricornis, Hansen, 1888 : 190—191; Richardson, 1900 :<br />
300; 1901 : 555; 1905a : 474—475, фиг. 529—530; Stebhing, 1900 : 14; О h 1 i n,<br />
1901:30—31; Hansen, 1916:17—19, табл. I, фиг. За—3b; Boone, 1920 : 7d;<br />
Гурьянова, 1932 : 24, табл. Ill, 14; 1933a : 400; 1952 : 175; Stephensen,<br />
1943 : 37—38, фиг. 7. — Jolanthe libbeyi, О r t m a n n, 1900 : 39. — Tole (опечатка,<br />
надо tale) libbeyi, О г t m a n n, 1901 : 157. — Janthe libbeyi Stephensen,<br />
1912 : 583; 1913 : 70, табл. 3. — lolella libbeyi Richardson, 1905a : 463—464,<br />
фиг. 518. — f.Jolella, alascensis Richardson, 1905a : 463—464. фиг. 518. — ?<br />
Janira alascensis Гурьянова, 1932 : 24, табл. Ш, рис. 13. — ? Janiralata alascensis<br />
M e n z i e s, 1951b : 138.<br />
В коллекциях ЗИН АН <strong>СССР</strong> имеются 16 экз. этого вида, из них<br />
5 экз. из района Земли Франца-Иосифа и о. Белый, 3 — из Берингова<br />
пролива, 5 — из зап. части Берингова моря и 3 — с юго-вост. побережья<br />
о. Парамушир. Сравнение особей с зап. сектора Арктики и сев'.-зап.<br />
части Тихого океана показало, что все они относятся к одному виду и<br />
весьма сходны с экземпляром J. tricornis с побережья зап. Гренландии,<br />
изображенным Стефенсеном (Stephensen, 1913, табл. III).
— 21 —<br />
/. tricornis ио всем признакам подходит к роду J antra Ша. Она отличается<br />
от представителей рода Janira (J. maculosa Leach и /. alta Stimpson)<br />
зазубренностью проксимальной части внутреннего края проподита<br />
I перейопода (Stephensen, 1913, табл. III) и строением I плеопода самца,<br />
задне-боковые углы которого сильно оттянуты в стороны (рис. 1).<br />
В оригинальных рисунках Henopomus tricornis (=J. tricornis) <strong>К</strong>ройер<br />
(Kroyer, 1849, табл. XXX, фиг. 2а—q), допустил опечатки; под одним номером<br />
2р дал рисунки I плеоподов самцов различных видов. Эти рисунки<br />
были без исправлений воспроизведены Ричардсон (Richardson,<br />
1905а, рис. 530). Между тем только нижний из изображенных <strong>К</strong>ройером<br />
Рис. 1. Janiralata tricornis (Krayer), I плеопод самца.<br />
о — экземпляр из района Земли Франца-Иосифа; б ~ экземпляр<br />
из 4-го <strong>К</strong>урильского пролива.<br />
илеоподов относится к J. tricornis, тогда как на верхнем рисунке изо-<br />
Г»ражен I плеопод Janira maculosa, которая онисана и иллюстрирована<br />
<strong>К</strong>ройером там же под названием Henopomus muticus.<br />
Janiralata alascensis (Benedict) из вост. части Чукотского моря (Richardson,<br />
1905а) весьма близка к /. tricornis и, возможно, является ее<br />
синонимом. Отличия между /. alascensis и Iolella libbeyi (один из синонимов<br />
/. tricornis) почти незаметны, если сравнивать рисунок J. alascensis<br />
не с рисунками Ортмана (Ortmann, 1900) и Ричардсон (1905а,<br />
рис. 518), а с гораздо более четким рисунком Стефенсена (Stephensen,<br />
1913, табл. III). Передне-боковые углы головы на рисунке Стефенсена<br />
более короткие, чем на рисунке Ортмана, и лишь незначительно более<br />
длинные, чем у /. alascensis. Задняя медиальная лопасть плеотельсона<br />
/. tricornis на рисунке Стефенсена точно так же несколько короче и уже<br />
боковых, как иу ). alascensis. Просмотренные нами особи с Земли Франца-<br />
Иосифа и из Берингова моря были вполне сходны с изображенной на<br />
рисунке Стефенсена. Единственными отличиями, которые можно отметить<br />
для обоих видов, являются наличие причленешшх к вершинам<br />
передне-боковых углов головы шипиков и крошечных вырезок на заднебоковых<br />
краях плеотельсона у ,/. tricornis. Оба эти характерных для<br />
/. tricornis признака не были отмечены для /. alascensis, но следует учесть<br />
что на единственном поврежденном, лишенном II антенн и уропод экземпляре<br />
/. alascensis, по которому был описан вид, эти признаки могли быть
_ 22 —<br />
не замечены. Среди изученных экземпляров из коллекции ЗИН АН <strong>СССР</strong><br />
встречаются такие, у которых шипик с одной стороны головы обломан;<br />
у других же, наоборот, нежные маленькие лопасти по бокам плеотельсона<br />
частично повреждены и боковые края плеотельсона на первый взгляд<br />
кажутся гладкими.<br />
Зап. сектор Арктики: зап. побережье Гренландии и Дэвисов пролив<br />
от 61 до 76° с. ш., вост. Гренландия до 65° с. ш., Ян-Майен, Шпицберген,<br />
о. Белый и Земля Франца-Иосифа до 81°14.5' с. ш., Берингов пролив;<br />
юго-зап. часть Берингова моря к востоку от о. Медный; Тихий океан,<br />
кшн. побережье о. Парамушир; Чукотское море у м. Барроу (?). Арктический<br />
сублиторальный вид, обитает на глубинах от 7 до 167 м, на илистых,<br />
песчаных и галечных грунтах. Обнаружен при температурах от<br />
—1.42 до +0.71° в зап. секторе Арктики и от —1.07 до +2.56° в сев.<br />
части Тихого океана.<br />
Janiralata erostrata (Richardson) (рис. 2)<br />
lanthe erostrata Richardson, 1899 : 858—859; 1900 : 299. — Jolella erostrata<br />
Richardson, 1905a : 465, фиг. 520. — J'antra erostrata Гурьянова, 1936a :<br />
41—42, фиг. 8; 1950 : 281. — Janiralata erostrata M e n z i e s, 1951b : 138.<br />
Рис. 2. Janiralata erostrata (Rich,) (особи с восточного побережья <strong>К</strong>амчатки),<br />
плеоиоды и боковой край II грудного сегмента.<br />
/. erostrata описана Ричардсон по одному неполноценному экземпляру<br />
с Алеутских о-вов и недостаточно подробно. Поэтому определение дальневосточного<br />
материала вызывает затруднения. В коллекциях ЗИН АН
— 23 —<br />
<strong>СССР</strong> имеется 9 экз. с вост. побережья <strong>К</strong>амчатки и сев. <strong>К</strong>урильских о-вов,<br />
которые мы относим к этому виду.<br />
Длина тела в 2.2—2.5 раза превосходит ширину; длина максимального<br />
из просмотренных нами экземпляров около 5.7 мм. Формой тела<br />
дальневосточные особи подходят под описание и рисунок Ричардсон,<br />
за исключением характера коксальных пластинок, которые по Ричардсон<br />
имеют каждая по одной небольшой лопасти, тогда как у наших экземпляров<br />
они состоят из двух лопастей по краю каждого из сегментов,<br />
начиная со II. Передняя лопасть коксальной пластинки II грудного сегмента<br />
значительно короче передней лопасти сегмента и примерно такой же<br />
длины, как его задняя лопасть. Передние лопасти коксальных пластинок<br />
несколько крупнее задних, края обеих лопастей закруглены. I антенна<br />
довольно короткая, ее жгутик состоит из 7—9 члеников; II антенна<br />
длинная, примерно равна длине тела. Внутренний край проподита I<br />
перейопода несет в проксимальной части 6—8 зубчиков. Зазубренная<br />
часть занимает менее J4 длины внутреннего края проподита. Дистальные<br />
боковые углы I плеопода самца сильно оттянуты в стороны; медиальные<br />
лопасти дистального края I плеопода ясно выражены, широкие, каждая<br />
из них посредине слегка вогнута. Задний край крышечки самки с ясной<br />
широкой выемкой посредине. Наружная ветвь уропода незначительно<br />
короче внутренней.<br />
Сев. часть Тихого океана: о. Атту, бухта Чичагова, Алеутские о-ва,<br />
1 $ (голотип) — по Ричардсон (Richardson, 1899, 1905а); вост. побережье<br />
<strong>К</strong>амчатки: у м. <strong>К</strong>амчатский, сборы на э/с «Витязь», 1950 г., ст, 523,<br />
глуб. 105 м, 2 9?; у м. Поворотный, сборы <strong>К</strong>амчатской станции, 7 1X1935,<br />
ст. 608, глуб. 125 м, 3 93, 1 (5 и 2 juv.; к юго-востоку от о. Парамушир,<br />
сборы на р/т «Лебедь», 7 VII 1954, ст. 58, глуб. 196—230 м, t воды у дна<br />
0.7°, грунт — песок и ил, 1 juv. Самки с эмбрионами в сумках были встречены<br />
в сентябре. Число эмбрионов у одной из изученных самок равно 29.<br />
Все остальные указания местонахождения J. erostrata, приводимые<br />
различными авторами, по-видимому, относятся к другими видам.<br />
Janiralata soldatovi (Gurjanova) (рис. 3)<br />
Janira soldatovi, Гурьянова, 19336 ; 8i, рис. 4; 1936 a : 42, фиг. 9. — Janiralata<br />
soldatovi, M e n z i e s, 1951b : 138.<br />
В коллекциях ЗИН АН <strong>СССР</strong> хранится 23 экз. этого вида. По форме<br />
тела J. soldatovi очень близка к J. erostrata, но отличается от последней<br />
несколько более короткими отростками передне-боковых углов головы,<br />
плавно закругленным в средней части передним краем головы и относительно<br />
более крупными глазами. Более четкими признаками, разделяющими<br />
эти виды, являются формы коксальных пластинок на II грудном<br />
сегменте, I плеопода самца и крышечки самки. Верхняя лопасть коксальной<br />
пластинки II грудного сегмента у /. soldatovi значительно длиннее<br />
нижних лопастей как коксальной пластинки, так и самого сегмента и<br />
почти равна длине верхней лопасти сегмента. Дистальные боковые углы<br />
I плеопода самца оттянуты в стороны несколько сильнее, чем у J. erostrata,<br />
и заострены на концах; медиальные лопасти почти не выражены<br />
и снабжены каждая маленькой дополнительной лопастью. Задний край<br />
крышечки самки у /. soldatovi в отличие от всех остальных видов этого<br />
рода лишен выемки и, наоборот, слегка выпуклый. Щетинки на вентральной<br />
поверхности плеотельсона у основания уропод заметно более длинные,<br />
чем у /. erostrata, и иногда видны сверху, так как просвечивают<br />
через плеотельсон. Длина тела у /. soldatovi в 2.3—2.7 раза превосходит
Рис. 3. Janiralata soldatoei (Gurjan.) (котипы), Японское море.
— 25 —<br />
*то ширину. Максимальная длина самки 5.6 мм, самца — 4.6 мм. I антенны<br />
довольно короткие, их жгутик состоит из 6—10 члеников. II антенны<br />
обычно несколько длиннее тела, их жгутик содержит около 100 члеников.<br />
Проксимальная часть внутреннего края проподита 1 перейоцода<br />
несет 8—10 зубчиков. Наружная ветвь уропода заметно короче внутренней.<br />
Охотское море и сев. часть Японского моря, на глубинах 34—144 м.<br />
Побережье Приморья, сборы на шхуне «Россинанте», 16 XI 1930: ст. 19,<br />
глуб. 62—65 м, t воды 3.45', грунт — галька с илом, 1 6 и 2 $д; ст. 141,<br />
глуб. 34 м, t воды 1.2°, грунт — песок с илом, галькой и гравием, 5 котипов<br />
(1 6 и 4 ^з с эмбрионами на I стадии). Зал. Петра Великого: сборы<br />
на шхуне «Сторож», 14 VIII 1908, ст. 10, глуб. 60 м, грунт — песок,<br />
6 66 и 4 $$ и 1 juv.; сборы на шхуне «Альбатрос», 21 VI 1957, ст. 40,<br />
глуб. 60 м, грунт — песок, 2 66 и 1 9 с оостегитами. Охотское море,<br />
зал. Терпения, сборы на р/т «Топорок», 1947 г., ст. 67, глуб. 144 м, 1 6.<br />
Jaruralata gurjanovae Kussakin, sp. п. (рис. 4—6)<br />
Самец. Тело уплощенное, широкоовальное, длина его в 2—2.3 раза<br />
превосходит ширину (длина голотипа 4.1 мм, ширина 1.8 мм). Спинная<br />
поверхность тела гладкая; боковые края головы, грудных сегментов и<br />
плеотельсона несут щетинки. Голова широкая,<br />
ее ширина более чем вдвое превосходит длину.<br />
Рострум очень короткий, спереди плавно закруглен;<br />
боковые лопасти головы крупные, их края<br />
слегка зазубрены в местах отхождения щетинок;<br />
передне-боковые углы головы сильно оттянуты<br />
вперед, значительно выдаются за пределы рострума,<br />
тупо заострены на конце. 3 передних грудных<br />
сегмента примерно равной длины, 4 задних<br />
сегмента несколько более короткие, боковые части<br />
3 передних сегментов направлены несколько вперед,<br />
3 последних — несколько назад. Боковой<br />
край каждого из 3 передних сегментов снабжен<br />
2 длинными лопастями. У I грудного сегмента<br />
передние лопасти значительно уже задних и<br />
сильно суживаются на конце, задние лопасти почти<br />
прямоугольной формы, со срезанными краями.<br />
Задние лопасти II—III сегментов несколько<br />
шире и короче передних лопастей; края всех<br />
лопастей срезаны. 4 задних грудных сегмента<br />
снабжены только передними лопастями; ширина<br />
лопастей у IV сегмента составляет не более половины<br />
ширины сегмента, у V—VII сегментов<br />
почти равна ширине сегментов. Задне-боковые<br />
углы лопастей VI—VII сегментов состоят каждая<br />
из двух лопастей, задние лопасти значительно<br />
короче передних и видны сверху только на<br />
4 передних сегментах. У II—III сегментов коксальные<br />
пластинки расположены в глубоких<br />
вырезках между лопастями сегмента, передние<br />
Рис. 4. Janiralata gurjanovae<br />
Kussakin, sp. п.,<br />
j (аллотип).<br />
лопасти коксальных пластинок лишь незначительно короче задних<br />
лопастей сегментов; у IV—VII сегментов коксальные пластинки расположены<br />
позади лопастей сегментов и значительно короче последних.<br />
Задний край плеотельсона снабжен 3 крупными лопастями, плавно
— 26 —<br />
закругленными на концах; средняя лопасть значительно короче 2 боковых.<br />
I антенны относительно несколько более длинные, чем у /. erostrata;<br />
число члеников жгутика достигает 15—18. II антенны обычно несколько<br />
длиннее, реже немного короче тела. Ротовые части типичного<br />
для рода Janiralata строения. Зубной ряд левой мандибулы состоит<br />
Рнс. 5. Janiralata gurjanovae Kussakin, sp. п. (голотили паратипы),<br />
головные придатки.<br />
примерно из 8 зазубренных щетинок; на внутренней пластинке ногочелюстя<br />
4 соединительных крючка. <strong>К</strong>арпоподит I перейопода расширен,<br />
его выпуклый внутренний край помимо щетинок несет около 30 шипов;<br />
проксимальная часть внутреннего края проподита несет около 12—13 зубчиков,<br />
зазубренная часть занимает около Х А длины внутреннего края.<br />
I плеопод по форме в общем сходен с таковыми у /. erostrata (Rich.) и<br />
/. soldatovi (Gurjan.), но лопасти на дистальном крае, в отличие от<br />
/. soldatovi, четко обособлены и лишены, в отличие от /. erostrata, вогнутости<br />
посредине. Маленькие дополнительные лопасти хорошо различимы,
— 27 —<br />
как и у Л soldatovi. Уроподы немного короче плеотельсона, их ветви<br />
иногда примерно равны по длине, иногда наружная ветвь заметно короче<br />
внутренней.<br />
Рис. 6. Janiralata gurjanovae Kussakin, sp. п. {голотип и паратипы), вдрейоподы><br />
плеоподы и боковые края II и IV грудных сегментов.<br />
Самки внешне сходны с самцами, тело яйценосных самок относительно<br />
несколько более широкое, длина его в 1.8—2 раза превосходит<br />
ширину. Самки в апреле без оостегитов, в августе—сентябре с эмбрионами<br />
на разных стадиях развития. В сумке у самки содержится 30—<br />
60 эмбрионов, длина эмбриона на III стадии 1—1.1 мм, диаметр оплодотворенных<br />
яиц 0.4 мм. Задний край крышечки самки всегда с более<br />
или менее глубокой вырезкой посредине.
— 28 -<br />
Максимальная длина изученных самцов 4.8 мм, самок — 6.5 мм.<br />
Окраска спиртовых экземпляров желтовато-серая, иногда розовато-серая,<br />
с темными пигментными пятнами.<br />
Южн. часть Охотского моря, Южно-<strong>К</strong>урильское мелководье, на<br />
глубине 10—74 м.<br />
О. Шикотан: бухта Открытая (Е. Гурьянова), 18 IX 1949, глуб. около<br />
10 м, 1
— 29 —<br />
не закруглены, а скорее тупо заострены на концах. I антенны, как и<br />
у /. gurjanovae, относительно более длинные, чем у /. erostrata; число<br />
члеников жгутика у голотипа равно 12, у других особей достигает 16—<br />
18. II антенны несколько короче тела. I и II максиллы сходны с таковыми<br />
у /. gurjanovae: зубной ряд левой мандибулы состоит из меньшего числа<br />
(5), но более широких, чем у J. gurjanovae, уплощенных, пильчатых<br />
щетинок; на внутренней пластинке вогочелюстиЗ соединительных крючка.<br />
Рис. 8. Janiralata ochotensis Kussakin, sp. п., 6 (голотип), головные придатки.<br />
<strong>К</strong>арпоподит I перейопода расширен; его выпуклый внутренний край<br />
помимо щетинок несет разное число, но всегда менее 30 раздвоенных на<br />
конце шипов. Проксимальная часть внутреннего края проподита имеет<br />
около 12 зубчиков, зазубренная часть занимает почти 1 А длины внутреннего<br />
края. I плеопод во многих отношениях занимает промежуточное положение<br />
между таковыми у J. soldatovi и /. gurjanovae; лопасти дистального<br />
края обособлены более четко, чем у J. soldatovi, но менее резко,<br />
чем у /. gurjanovae; дополнительные лопасти очень маленькие. Заднебоковые<br />
углы у J. ochotensis сильнее сужены на концах, чем у обоих<br />
сравниваемых видов. Уроподы несколько короче плеотельсона, их внутренняя<br />
ветвь немного длиннее наружной.<br />
Самки внешне сходны с самцами. Задний край крышечки слегка<br />
вогнут посредине. В августе самки с оостегитами, в октябре без оостегитов.<br />
В сумке одной из самок, найденных в августе, были обнаружены яйца,<br />
заполненные паразитическими копеподами.
— 30 —<br />
Окраска обоих полов более светлая, чем у /. gurjanovae, сероватожелтая.<br />
Максимальная длина самца 7.35 мм, самки — 7.9 мм (голотип<br />
и аллотип).<br />
Сев. часть Охотского моря, у входа в Пенжинекий залив, сборы на<br />
э/с «Витязь», VIII 1949, ст. 58, глуб. 92 м, 1 6, голотип (№ 1/39 609)<br />
Рис. 9. Janiralata ochotensis Kussakin, sp. п. (голотип и аллотип), перейопод,<br />
плеоподы и боковые края II и IV грудных сегментов.<br />
и 4 22 (аллотип и паратипы); 24 VIII 1949, ст. 49, глуб. 115 м, 3 99 с оостегитами,<br />
вместе с /. vitjazi; 6 X 1952, ст. 1803, глуб. 124 м, 4 дефектные 99<br />
без сумок и 1 небольшой 6.<br />
Новый вид близок к J. erostrata, J. soldatovi и особенно к /. gurjanovae.<br />
От /. erostrata и /. soldatovi легко отличается сильным развитием боковых<br />
лопастей заднего края плеотвльсона, которые значительно длиннее средней<br />
лопасти. От /. gurjanovae хорошо отличается более короткими передне-боковыми<br />
отростками головы, формой боковых лопастей заднего<br />
края плеотвльсона и задних лопастей II и III грудных сегментов, числом<br />
щетинок зубного ряда мандибул и соединительных крючков ногочелюстей,<br />
а также деталями строения карпоподита I перейопода и I плеопода самца.<br />
От всех трех сравниваемых видов отличается несколько более развитым<br />
рострумом.
— 31 —<br />
Janiralata kurilensis Kussakin, sp. п. (рис. 10—12)<br />
Самец. Тело уплощенное, широкое, его длина вдвое превосходит<br />
ширину (длина голотипа 3.0 мм, ширина 1.5 мм). Спинная поверхность<br />
гладкая; боковые края головы, грудных сегментов и плеотельсона несут<br />
немногочисленные щетинки. Ширина головы менее чем вдвое превосходит<br />
длину. Рострум умеренной длины, треугольный, тупо заострен спереди,<br />
выдается за передние края боковых лопастей головы. Передне-боковые<br />
отростки головы небольшой длины, тупые спереди. Боковые края 3 передних<br />
грудных сегментов снабжены 2 умеренной длины лопастями каждый;<br />
передние лопасти I сегмента заострены, остальные закруглены на<br />
концах; задние лопасти короче передних. 3 задних<br />
грудных сегмента снабжены лишь передними лопастями.<br />
<strong>К</strong>ороткие задние лопасти коксальных пластинок<br />
видны сверху лишь на II и III грудных сегментах;<br />
их передние лопасти длиннее задних, на II<br />
и III сегментах они почти равны по длине или несколько<br />
короче задних лопастей сегментов, на IV<br />
они довольно длинные и лишь незначительно короче<br />
передних лопастей сегмента, на V—VII сегментах<br />
они очень короткие. Задние лопасти плеотельсона<br />
примерно равной длины, широкие, но очень короткие,<br />
значительно короче, чем у /. erostrata (Rich.)<br />
и J. soldatovi (Gurjan.), плавно закруглены на концах,<br />
I антенны относительно короткие, их жгутик<br />
состоит из 9—10 члеников; II антенны короче тела.<br />
Внутренняя лопасть II максилл заметно короче наружных<br />
и относительно более короткая, чем у видов<br />
группы /. erostrata. Зубной ряд левой мандибулы<br />
состоит примерно из 8—9 тесно сближенных,<br />
зубчатых щетинок; внутренняя пластинка ногочелюсти<br />
с 2 соединительными крючками. <strong>К</strong>арпопо-<br />
дит I перейопода расширен, его выпуклый внутренний<br />
край несет около 20 раздвоенных шипов и несколько<br />
щетинок; проксимальная часть внутреннего<br />
края проподита имеет маленькие, едва заметные<br />
зазубрины. Задне-боковые отростки I плеопода<br />
короткие, чем у видов группы /. erostrata<br />
Ряс. 10.<br />
kuri lensis<br />
sp. п., 6<br />
Janira lata<br />
Kussakin,<br />
(голотип).<br />
значительно более<br />
дистальные лопасти слабо<br />
обособлены. Наружная ветвь уроподов заметно короче внутренней.<br />
Самка внешне сходна с самцом, длина ее тела в 2.2 раза превосходит<br />
ширину. <strong>К</strong>рышечка самки широкая, ее задний край с широкой, но неглубокой<br />
выемкой посредине. Длина самки 4.5 мм.<br />
Окраска спиртовых экземпляров обоих полов желтовато-серая, с красно-бурыми<br />
скоплениями пигмента.<br />
Тихий океан, побережье южн. <strong>К</strong>урильских о-вов, сублитораль до<br />
60 м глубины.<br />
О. Шикотан, бухта Открытая (Е. Гурьянова), глуб. 30 м, 18 IX 1949,<br />
2 небольших 66, голотип (№ 1/39 612) и паратип; о. Итуруп, зал. <strong>К</strong>асатка<br />
(Ан. Стрелков и О. <strong>К</strong>усакин), 11 VII 1954, глуб. 50—60 м, биоценоз<br />
губок, 1 9 с эмбрионами на I стадии в выводковой сумке.<br />
J. kurilensis близка к видам группы J. erostrata, куда помимо последнего<br />
вида относятся /. soldatovi, J. gurjanovae Kuss. и J. ochotensis Kuss.<br />
От всех видов этой группы /. kurilensis легко отличается более короткой<br />
внутренней лопастью II максилл и хорошо развитым рострумом, который,<br />
однако, значительно короче, чем у J. tricornis и J. alascensis.
— 32 —<br />
Janiralata vitjazi Kussakin, sp. п. (рис. 13—15)<br />
Самка. Тело уплощенное, широкоовальное, длина его в 1,7—<br />
2.1 раза превосходит ширину. Спинная поверхность тела и боковые края<br />
головы, грудных сегментов и плеотельсона несут многочисленные мелкие<br />
щетинки. Голова широкая, ширина ее примерно вдвое превосходит длину;<br />
рострум умеренной длины, несколько более короткий, чем у J. kurilensis<br />
Kussakin, треугольный, заостренный на конце; боковые отростки головы<br />
широкие, но сравнительно короткие, довольно слабо обособлены. Боковые<br />
лопасти грудных сегментов довольно длинные; передняя лопасть I груд-<br />
Рис, 11. Janiralata kurilensis Kussakin, sp. п.,
— 33 —<br />
7—8 тесно сближенных, зубчатых щетинок; на внутренней пластинке ногочелюсти<br />
2 соединительных крючка. <strong>К</strong>арпоподит I перейопода расширен,<br />
его сильно выпуклый внутренний край несет более 30 раздвоенных шипов<br />
и несколько щетинок; проксимальная часть внутреннего края проподита<br />
Рис. 12. Janiralaia kurilensis Kussakin, sp. п. (голотип и паратипы),<br />
перейопод и плеоподы.<br />
нмеет не более 10 тонких зубчиков. <strong>К</strong>рышечка (II плеопод) самки с глубокой<br />
выемкой посредине заднего края. Наружная ветвь уропода заметно<br />
;ороче внутренней. Окраска в спирту светлая желтовато-серая, более<br />
:-?мная в средней части грудных сегментов из-за просвечивающих мускулатуры<br />
и внутренних органов. Максимальная длина самок (голотип) 5.6 мм.<br />
Самцы не известны.<br />
3 Труды зин, т. ххх
— 34 —<br />
Сев. часть Охотского моря, у входа в Пенжинский залив, сборы на<br />
э/с «Витязь», 24 VIII 1949, ст. 49, глуб. 115 м, 5 дефектных $>$, голотип<br />
(№ 1/39614) и паратипы, вместе с J. ochotensis; 1950 г., ст. 523, глуб. 105 м,<br />
грунт — песок, 1 дефектная $ с эмбрионами на I стадии в выводковой<br />
сумке. Тихий океан, у юго-вое т. побережья о. Парамушир, сборы на<br />
р/т «Лебедь», VIII 1954, ст. 155, глуб. 48 м,<br />
грунт — галька, 1 небольшая j; ст. 165, глуб.<br />
107 м, грунт — песок со следами ила, t у дна<br />
0.9°, 1 $ без оостегитов.<br />
От остальных видов этого рода /. vitjazi отличается<br />
сильно расширенным карпоподитом I перейопода.<br />
Формой рострума /. vitjazi несколько<br />
напоминает /. &wite«sssKussakin, но по ряду признаков<br />
наиболее близка к /. sarsi (Rich.). От последнего<br />
вида отличается формой переднего края<br />
головы, который у /. sarsi практически лишен<br />
рострума и передне-боковых отростков, более короткой<br />
медиальной и более широкими боковыми<br />
лопастями заднего края плеотельсона и некоторыми<br />
другими признаками.<br />
Ряс. 13. Janiralata vitjazi<br />
Kussakin, sp. п., $><br />
(голотип).<br />
Janiralata pilosa Kussakin, sp. u. (рис. 16—18).<br />
Самец. Тело уплощенное, широкоовальное,<br />
его длина в 1.7—1.9 раза превосходит ширину.<br />
Спинная поверхность тела покрыта щетинками;<br />
края головы, плеотельсона и боковые края грудных<br />
сегментов усажены довольно длинными,<br />
иглообразными щетинками. Голова широкая,<br />
ее ширина более чем вдвое превосходит длину; передний край головы<br />
несет 3 короткие, закругленные, примерно равной величины выпуклости,<br />
из которых средняя представляет собой сильно укороченный рострум;<br />
боковые лопасти головы длинные, но относительно узкие; передне-боковые<br />
отростки головы заметно выдаются вперед за пределы рострума. Боковые<br />
лопасти грудных сегментов длинные, узкие, почти прямоугольной формы,<br />
с закругленными концами на I—IV сегментах, довольно широкие, но<br />
значительно более узкие, чем соответствующие боковые лопасти V—<br />
VII сегментов. Задние лопасти II—III грудных сегментов почти такой же<br />
длины, как и передние лопасти. Передние лопасти коксальных пластинок<br />
II и III грудных сегментов удлиненные, но значительно более короткие,<br />
чем лопасти сегментов; задние лопасти короткие. Медиальная задняя<br />
лопасть плеотельсона очень широкая, но короткая, сзади закруглена;<br />
боковые лопасти примерно вдвое длиннее медиальной, гораздо более узкие,<br />
треугольной формы, с более отчетливо, чем у Л vitjazi Kuss,, заостренными<br />
концами. Жгутик I антенн состоит в среднем из 13 члеников.<br />
II антенны значительно короче тела, их жгутик состоит примерно из<br />
60—80 члеников. Внутренняя лопасть II максиллы лишь незначительно<br />
короче наружных; левая мандибула несет не менее 10 тесно сближенных,<br />
зазубренных щетинок зубного ряда; на внутренней пластинке ногочелюсти<br />
2 соединительных крючка. <strong>К</strong>арпоподит I перейопода расширен, несет<br />
около 30 раздвоенных шипов по внутреннему краю; проксимальная часть<br />
внутреннего края проподита имеет 12—16 широких зубчиков; зазубренная<br />
часть занимает около 2 /з длины внутреннего края. Задне-боковые отростки<br />
I плеопода довольно длинные, суживаются, но не заостряются<br />
на концах; дистальные лопасти хорошо выражены, причем каждая из
— 35 —<br />
них выемкой разделена на две, из которых наружная значительно длиннее<br />
внутренней. Уроподы уплощенные, относительно короткие и широкие,<br />
значительно короче плеотельсона; их наружная ветвь немного короче<br />
внутренней.<br />
Самки внешне сходны с самцами. Задний край крышечки слегка<br />
вогнут посредине. В июле и августе встречены самки с эмбрионами на<br />
Рис. 14. Janiralata vitjazi Kussakin, sp. п., $ (голотид), головные придатки.<br />
разных стадиях в выводковых сумках. <strong>К</strong>оличество эмбрионов у 3 просмотренных<br />
самок изменяется в пределах от 25 до 50; диаметр оплодотворенного<br />
яйца около 0.45 мм, длина эмбриона на III стадии около 1 мм.<br />
Окраска спиртовых экземпляров обоих полов светлая желтоватосерая,<br />
с многочисленными довольно крупными пигментвыми пятнами.<br />
Максимальная длина самца 6.7 мм (голотип), самки — 6.6 мм.<br />
Тихий океан, юго-вост. побережье п-ва Лопатка и о. Парамушир,<br />
сборы на р/т «Лебедь», VII—VIII 1954, ст. 140, глуб. 24 м, грунт —<br />
камень, t у дна 6.1°, 3 66, голотпп (№ 1/39 623) и паратипы и 7 g$ (аллотвп<br />
и паратипы); ст. 109, глуб. 44 м, грунт — песок с гравием и галькой,<br />
X у дна 1.4°, 1 $; ст. 64, бухта Васильева, глуб. 31 м, грунт — гравий<br />
3*
— 36 —<br />
с пегком и галькой, t у дна 4.8°, 1 $; юго-вост. побережье о. Итуруи,<br />
сборы на р/т «Топорок», 1948 г., ст. 466, 1 ?.<br />
<strong>К</strong> этому же виду, очевидно, относится и лишенный головы дефектный<br />
экземпляр из 1-го <strong>К</strong>урильского пролива (сборы Смирнова и Бегак<br />
Рис. 15. Janiralata vitjazi Kussakin, sp. п., у (голотип), перейопод, плеоподы<br />
д боковой край грудного сегмента.<br />
в 1907 г.), определенный Е. Ф. Гурьяновой как Janira sarsi (Гурьянова,<br />
1936а, 19366).<br />
J. pilosa наиболее близка к /. vitjazi, но особенно к J. holmesi (Rich.)<br />
и /. sarsi (Rich.). От J. vitjazi J. pilosa отличается прежде всего отсутствием<br />
хорошо выраженного рострума, от J. holmesi и /. sarsi — наличием<br />
сильно развитых передне-боковых отростков головы, а от /. sarsi,<br />
кроме того, отсутствием коксальных пластинок между лопастями I грудного<br />
сегмента.<br />
Janiralatashiiuoi Kussakin, sp. п. (рис. 19—21).<br />
Самец. Тело уплощенное, овальное, длина его менее чем вдвое<br />
превосходит ширину (длина голотипа 5.25, ширина 2.85 мм). Спинная<br />
поверхность гладкая, по бокам тела имеются довольно длинные щетинки.
— 37 —<br />
Голова лишена рострума; передний край головы лишь немного выпуклый;<br />
боковые лопасти головы широкие; передне-боковые отростки головы значительно<br />
выдаются вперед, треугольной формы, тупо заострены на конце.<br />
Глаза относительно небольшие, расположены ближе к заднему, чем к переднему,<br />
краю головы, красно-бурого цвета. Передние боковые лопасти<br />
I грудного сегмента очень короткие, треугольной формы; остальные<br />
боковые отростки всех грудных сегментов довольно длинные, более или<br />
менее закруглены на конце. Передние боковые лопасти И грудного сегмента<br />
значительно уже и короче задних, у III<br />
сегмента эта разница выражена слабее; IV—<br />
VII сегменты несут только передние лопасти.<br />
<strong>К</strong>оксальные пластинки на II—IV грудных<br />
сегментах сверху не видны; на V—VII сегментах<br />
они короткие, но довольно широкие,<br />
выступают за задне-боковые края сегментов.<br />
Медиальная задняя лопасть плеотельсона<br />
широкая, сзади закруглена; задне-боковые края<br />
плеотельсона плавно закруглены, без лопастей.<br />
Жгутнк Г антенны состоит из 9 удлиненных<br />
члеников. II антенна значительно короче тела.<br />
Внутренняя лопасть II максиллы примерно такой<br />
же длины, как наружные; левая мандибула<br />
содержит 10 зазубренных щетинок зубного<br />
ряда; внутренняя пластинка ногочелюстей<br />
с 2 соединительными крючками. <strong>К</strong>арпоподнт<br />
I перейопода расширен; его выпуклый внутренний<br />
край несет около 8 раздвоенных шипов,<br />
несколько длинных и большое количество<br />
коротких щетинок; проксимальная часть внутреннего<br />
края проподита I перейопода имеет<br />
5 зубчиков. Задне-боковые отростки I плеопода<br />
сильно оттянуты в стороны, заострены на<br />
конце; дистальные лопасти четко обособлены от<br />
них, очень широкие и занимают всю остальную<br />
часть каждой половины заднего плеопода.<br />
Уроподы широкие, уплощенные, значительно<br />
Рис. 16, Janiralata pilosa<br />
Kussakin, sp. п., 6 (голотип)<br />
.<br />
короче плеотельсона; обе ветви плоские, удлиненноовальные, почти<br />
равной длины.<br />
Окраска в спирту светло-коричневая, переходящая в средней части<br />
грудных сегментов в темно-коричневую.<br />
Самка внешне сходна с самцом, неполовозрелый экземпляр (аллотип),<br />
имеющийся в нашем распоряжении, несколько отличается от самца<br />
более короткими передне-боковыми отростками головы и боковыми лопастями<br />
грудных сегментов; длина тела вдвое превосходит ширину (длина<br />
аллотипа 2.75 мы, ширина 1.37 мм). Задний край крышечки слегка<br />
выпуклый.<br />
Восточно-<strong>К</strong>итайское море, сборы на э/с «Блюхер», 21 II 1931, трал<br />
23.3142' с. ш., 128°13' в. д., глуб. 180 м, грунт — песок, t у дна 17.3°<br />
1 £, голотип (№ 1/39608) и 1 неполовозрелая g (аллотип).<br />
Л shiinoi рядом существенных признаков отличается от всех известных<br />
видов рода J antra lata и близок к Jolella chuni Thielemann, (1910)'<br />
из зал. Сагами. От J. chuni новый вид отличается отсутствием скульптуры<br />
на спинной поверхности тела, несколько лучше развитыми передними<br />
боковыми лопастями I грудного сегмента, притуплёнными концами<br />
передне-боковых отростков головы и некоторыми другими незна-'
— 38 —<br />
чительными признаками. Сближает оба вида не только сходная форма<br />
тела, но и строение I плеопода самца, отсутствие видимых сверху коксальных<br />
пластинок II и III грудных сегментов, сходная форма боковых лопастей<br />
грудных сегментов, строение уропод, I и II антенн.<br />
Л chuni и /. shiinoi несколько отличаются от представителей родов<br />
Jolella и Janiralata. От типичных Janiralata оба вида отличает отсутствие<br />
видимых сверху коксальных пластинок по бокам II и III грудных<br />
Рис. 17. Janiralata pilosa Kussakin, sp. п. (голотип и алпотип),<br />
головные придатки.<br />
сегментов. Этот признак сближает их с видами рода Jolella, нам же он<br />
представляется формальным, так как коксальные пластинки сходны<br />
у /. shiinoi и у типа рода Janiralata — /. davisi Menzies. Различие по<br />
существу сводится к тому, что у /. shiinoi они не выступают за пределы<br />
выемок между лопастями II и III грудных сегментов и поэтому невидны<br />
сверху, а у J. davisi несколько выдаются и имеют со спинной стороны<br />
вид очень маленьких, закругленных лопастей. Признаки, отличающие<br />
/. shiinoi и /. chu.nl от рода Jolella, более существенны: боковые края<br />
IV грудного сегмента у обоих видов, как и у всех Janiralata, несут по<br />
одной лопасти с каждой стороны, тогда как у Jolella — по две лопасти;
Рис. 18. Janiralata pilosa Kussakin, sp. п. (ГОЛОТИП и паратипы), перейопод, плеопОДИ<br />
и боковой край грудного сегмента.
— 40 —<br />
имеются зубчики на проксимальной части внутреннего края проподита<br />
I перейопода, и I плеопод самца, как и у Janiralata, имеет направленные<br />
в сторону задне-боковые отростки. I плеопод самца J. shiinoi и /. chuni<br />
несколько отличается от остальных Janiralata сильным развитием дистальных<br />
лопастей. Эта особенность, а также отсутствие видимых сверху<br />
коксальных пластинок сближают оба этих вида и ставят их несколько<br />
особняком в роде Janiralata. Выпуклый задний край II плеопода самца<br />
хотя и не типичен для большинства видов рода Janiralata, но встречается<br />
не только у обоих японских видов, но<br />
также у /. soldatovi (Gurjan.). По форме тела<br />
и особенно плеотельсона J. shiinoi и /. chuni<br />
наиболее близки к /. davisi с побережья<br />
<strong>К</strong>алифорнии.<br />
Род IANIROPSIS G. О. SARS<br />
Род Ianiropsis впервые был описан с по<br />
бережья Норвегии. <strong>К</strong> настоящему времени<br />
описано еще 15 видов из сев. части Тихого<br />
океана и из тропических и субтропических<br />
областей. 1 вид (/. breviremis G. О. Sars)<br />
обитает у берегов Зап. Европы, 1 вид<br />
(/. parva Omer-Cooper) — в тропической<br />
части Тихого океана, у о. Фэннинг, 5 видов<br />
(/. kincaidi Rich., I. derjugini Gurjan., /. setifera<br />
Gurjan., /. serricaudis Gurjan. и /. pallidocula<br />
Kuss., sp. n.) — в дальневосточных<br />
морях <strong>СССР</strong>, 8 видов (/. kincaidi Rich.,<br />
/. derjugini Gurjan., /. analoga Menzies,<br />
/. epilittoralis Menzies, /. magnocula Menzies,<br />
/. minuta Menzies, /. montereyensis<br />
Рис. 19. Janiralata shiinoi<br />
Kussakin, зр. п., 3 (голотип).<br />
Menzies и /. tridens Menzies) — на тихоокеанском<br />
побережье Сев. Америки, 1 вид (/. longiantennata<br />
Thielemann) в Японии и, наконец,<br />
2 вида (/. palpalis Barnard и /. bisbidens<br />
Barnard) — у берегов южн. Африки и 3 вида<br />
с побережья Чили.<br />
Диагноз рода Ianiropsis по Мензису (Menzies, 1952). Голова, грудь<br />
и брюшко лишены выступающих лопастей. Голова лишена длинного рострума.<br />
Эпимеральные пластинки видны сверху по крайней мере на II—<br />
VII грудных сегментах. Брюшко состоит из 2 сегментов. Уроподы различной<br />
длины. Щупик ногочелюстей состоит из 5 члеников; 2-й и 3-й членики<br />
расширены и значительно шире внутренней пластинки. Ногочелюсть<br />
с 2 соединительными крючками. I плеопод самца с боковыми<br />
расширениями на конце. Дактилоподит I перейопода с 2 коготками, дактилоподиты<br />
II—VII перейоподов с 3 коготками. Проподит I перейопода<br />
самца гладкий, без зазубрин на внутреннем крае, но обычно несет различное<br />
число раздвоенных на конце щетинок. Зубной отросток мандибулы<br />
расширен на конце, с усеченной вершиной; 2-й членик щупика несет<br />
2 длинные, почти прямые, зазубренные щетинки, между которыми<br />
имеются сильно искривленные, зазубренные щетинки меньшего размера.<br />
У взрослых самцов I перейопод часто почти достигает длины тела. I антенны<br />
с 4-члениковой ножкой. II антенна с 6-члениковой ножкой, несущей<br />
отчетливую чешуйку на боковом крае 3-го членика.
Рис. 20. Janiralata shiinoi Kussakin, sp. п., $ (голотип), голов<br />
ные придатки.
Рис. 21. Janiralata shiinoi Kussakin, sp. п. (голотип и аллотип), перейопод, плеоноды<br />
и боковой край II грудного сегмента.
— 43 —<br />
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА IANIROPSIS G. О. SABS<br />
ИЗ СЕВЕРНОЙ ЧАСТИ ТИХОГО О<strong>К</strong>ЕАНА<br />
1 (12). Боковые кран плеотельсона имеют зазубрины.<br />
2 (5). Плеотельсон несет по 2 или 3 зазубрины с каждой стороны.<br />
3 (4). Плеотельсон несет по 2 зазубрины с каждой стороны. <strong>К</strong>алифорния<br />
I. epilittoralis Menzies (1952).<br />
4 (3). Плеотельсон несет по 3 зазубрины с каждой стороны. <strong>К</strong>алифорния<br />
I. tridens Menzies (1952).<br />
5 (2). Плеотельсон несет более 3 зазубрин с каждой стороны.<br />
6 (9). Длина плеотельсона примерно равна или несколько больше<br />
его ширины.<br />
7 (8). Передний край головы слегка вогнут посередине; задне-боковые<br />
отростки I плеопода самца умеренной длины и ширины, на концах<br />
не раздвоены и закруглены. Приморье<br />
I. serricaudis Gurjanova (19366).<br />
8 (7). Передний край головы выпуклый посередине; задне-боковые отростки<br />
I плеопода самца длинные, узкие, на концах несколько<br />
раздвоены и заострены. Побережье Японии<br />
I. longiantennata Thielemann (1910).<br />
9 (6). Длина плеотельсона значительно меньше его ширины.<br />
10 (11). Глаза нормально пигментированы, темные; передне-боковые<br />
края головы отчетливо угловатые. Тихоокеанское побережье Сев.<br />
Америки от штата Вашингтона до <strong>К</strong>алифорнии<br />
. I. analoga Menzies (1952).<br />
И (10). Глаза слабо пигментированы, светлые; передне-боковые края<br />
головы плавно закруглены. Побережье зап. <strong>К</strong>амчатки<br />
I. pallidocula Kussakin, sp. п.<br />
12 (1). Боковые края плеотельсона лишены зазубрин.<br />
13 (14). Длина уроподов заметно превышает длину плеотельсона. <strong>К</strong>алифорния<br />
I. montereyensis Menzies (1952).<br />
14 (13). Длина уроподов меньше длины плеотельсона.<br />
15 (20). Длина уроподов не превышает половины длины плеотельсона.<br />
16 (17). Плеотельсон с отчетливыми задне-боковыми углами по бокам<br />
выемок для уроподов. Дальневосточные моря <strong>СССР</strong> и тихоокеанское<br />
побережье Сев. Америки . . I. derjugini Gurjanova (1933а).<br />
17 (16). Плеотельсон без отчетливых задне-боковых углов по бокам<br />
выемок для уроподов.<br />
18 (19). Передне-боковые края головы плавно закруглены. <strong>К</strong>алифорния<br />
. I. minuta Menzies (1952),<br />
19 (18). Передне-боковые края головы с заостренными, треугольными<br />
отростками. <strong>К</strong>алифорния I. magnocula Menzies (1952).<br />
20 (15). Длина уроподов превышает половину длины плеотельсона.<br />
21 (22). Поверхность плеотельсона и боковые края грудных сегментов<br />
самок и самцов гладкие или покрыты редкими, мелкими щетинками.<br />
Длина брюшного отдела, включая I брюшной сегмент, несколько<br />
превышает или примерно равна ширине плеотельсона. Дальневосточные<br />
моря <strong>СССР</strong> и тихоокеанское побережье Сев. Америки . .<br />
I. kincaidi Richardson (1904).<br />
22 (21). Поверхность плеотельсона и боковые края грудных сегментов<br />
у самцов покрыты многочисленными длинными щетинками. Длина<br />
брюшного отдела, включая I брюшной сегмент, заметно меньше<br />
ширины плеотельсона. Дальневосточные моря <strong>СССР</strong><br />
I. setifera Gurjanova (1950).
— 44 —•<br />
laniropsis kincaidi Richardson (рис. 22)<br />
laniropsis kincaidi Richardson, 1904 : 221—222, рис. 102—107; 1905a : 456,<br />
рис. 509—514; Гурьянова, 1936a : 44—45, фиг. 12; 1950 : 281-282. — laniropsis<br />
kincaidi kincaidi M в n z i e s, 1952 : 136—139, рис. 56c, 57a—e.<br />
От близких видов /. derjugini Gurjan. и /. setifera Gurjan. отличается<br />
более стройным телом, длина которого в 3.3—3.8 раза превосходит ширину.<br />
От /. derjugini, кроме того, отличается более длинными и стройными<br />
перейоподами, формой заднего края плеотельсона, боковые углы которого<br />
тупо заострены и не образуют отчетливых лопастей, значительно<br />
более длинными уроподами, длина которых превышает половину плеотельсона,<br />
а у самцов почти достигает всей его длины. Из других отличительных<br />
признаков Мензис (Menzies, 1952) указывает на большую<br />
длину 1-го членика II антенны у /. kincaidi по сравнению с таковым<br />
/. derjugini. От /. setifera I. kincaidi отличается относительно менее широким<br />
плеотельсоном, несколько более длинными I антеннами и рядом<br />
других менее существенных признаков.<br />
Авторы, изучавшие эти 3 трудно различимых вида, обращали недостаточно<br />
внимания на половой диморфизм, степенью выраженности которого<br />
они существенно отличаются. У /. derjugini половой диморфизм<br />
выражен слабо, спинная поверхность тела гладкая, степень развития<br />
щетинистого покрова переиоподов и уроподов у обоих полов примерно<br />
одинакова, лишь карпоподит I перейопода у самца несколько сильнее<br />
расширен, чем у самки. У I. kincaidi самцы н самки внешне сходны,<br />
имеют гладкую спинную поверхность тела, но уроподы и особенно перейоподы<br />
самцов несут значительно большее количество более длинных<br />
щетинок, чем у самок. Самцы /. setifera резко отличаются от самок сильно<br />
развитым покровом из длинных, нежных щетинок на конечностях, плеотельсоне<br />
и боковых краях грудных сегментов. Однако неполовозрелые<br />
самцы /. setifera и /. kincaidi по степени развития щетинок очень похожи<br />
на самок соответствующих видов.<br />
Нами просмотрено 557 экз. /. kincaidi, находящихся в коллекциях<br />
ЗИН АН <strong>СССР</strong>, собранных на литорали дальневосточных <strong>морей</strong> от Приморья<br />
и южн. <strong>К</strong>урильских о-вов до вост. <strong>К</strong>амчатки и <strong>К</strong>омандорских<br />
о-вов. Все они отличаются от калифорнийских (любезно присланных<br />
Р. Мензисом) несколько менее широким плеотельсоном. Длина плеотельсона<br />
у наших экземпляров, как правило, немного превышает ширину,<br />
тогда как у калифорнийских он более округлый и его ширина обычно<br />
незначительно превосходит длину. Уроподы самца у дальневосточных<br />
особей относительно несколько более короткие, чем у калифорнийских.<br />
Половой диморфизм у американских и дальневосточных особей выражен<br />
в одинаковой степени. Возможно, здесь мы имеем дело с 2 подвидами одного<br />
вида, но трудно решить, какой из них является основным, соответствующим<br />
голотипу Ричардсон из Якутата (Аляска). По рисункам Ричардсон,<br />
у голотипа длина плеотельсона примерно равна его ширине,<br />
а уроподы относительно более короткие, чем у калифорнийских особей.<br />
С другой стороны, Мензис (Menzies, 1952), сравнивавший калифорнийские<br />
экземпляры с голотипом, не отмечает каких-либо различий между<br />
ними.<br />
Максимальная длина самца 5.3 мм, самки — 4.1 мм. Число эмбрионов<br />
у самок достигает 54, минимальное число их 12; длина молоди, выходящей<br />
из сумок, около 0.84 мм. У дальневосточных особей самки с эмбрионами<br />
в выводковых сумках встречены с июля по начало октября;<br />
в январе—мае самки без оостегитов, данные за июнь, ноябрь и декабрь<br />
отсутствуют.
Рис. 22. laniropsis kincaidi Rich, {особи с восточного побережья <strong>К</strong>амчатки), переиоподы,<br />
плеоподы и уроподы
— \C> —<br />
Японское, Охотское и Берингово моря от Приморья и юго-заи. Сахалина<br />
до вост. <strong>К</strong>амчатки и <strong>К</strong>омандорских о-вов; тихоокеанское побережье<br />
Сев. Америки от Аляски на юг до <strong>К</strong>алифорнии.<br />
Японское море, Приморье (35 экз.) и юго-зап. побережье южн. Сахалина<br />
(1 экз.); Охотское море, побережье южн. <strong>К</strong>урильских о-вов (5 экз.);<br />
Тихий океан, южн. <strong>К</strong>урильские о-ва (283 экз.), <strong>К</strong>омандорские о-ва<br />
(210 экз.), вост. побережье <strong>К</strong>амчатки (18 экз.), побережье Сев. Америки<br />
от южн. Аляски до <strong>К</strong>алифорнии (Ричардсон, 1904 г. и Мензис, 1951 г.;<br />
около 88 экз.). Обитает в литоральной зоне на скалистых, каменистых<br />
и галечных грунтах, преимущественно в ваннах, под камнями, в расщелинах,<br />
в колониях губок, реже среди водорослей. В отличие от /. derjugini<br />
на открытых прибою рифах вне укрытий не встречается.<br />
Iauiropsis derjugini Gurjanova (рис. 23)<br />
Janiropsis derjugini Гурьянова, 1933a : 82—83, рис. 5, 6; 1936a : 45—46,<br />
фиг. 13, а, б. — laniropsis kincaidi derjugini, M e Q z i e s, 1952 : 139—141, рис. 56a, Jt>,<br />
57f—i, 58, 59.<br />
Нами просмотрено 236 экз., имеющихся в коллекциях ЗИН АН <strong>СССР</strong>.<br />
Относительная ширина тела варьирует, но оно всегда более широкое,<br />
чем у /. kincaidi Rich.; длина тела превосходит ширину в 2.5—3.2 раза.<br />
Сравнение больших серий обоих видов показало, что различия между<br />
ними весьма стойкие; особей с переходными признаками не было обнаружено.<br />
Ареалы обоих видов в большей своей части совпадают, и, хотя<br />
экологически виды несколько различаются между собой, их биотопы<br />
частично могут совпадать; в некоторых случаях оба вида были встречены<br />
в одной пробе. Это позволяет нам считать их различными видами, а не<br />
объединять в качестве отдельных подвидов в один вид.<br />
От /. setifera Gurjan. /. derjugini отличается несколько более коренастым<br />
телом с более широкой головой, значительно лучше развитыми<br />
задне-боковыми лопастями плеотельсона, несколько более короткими<br />
уроподами, перейоподами и II антеннами и слабо выраженным половым<br />
диморфизмом.<br />
Максимальная длина просмотренных самцов 4.3 мм, самок — 4.7 мм.<br />
Самки с эмбрионами на разных стадиях развития встречены в июле и<br />
августе, количество эмбрионов достигает 62, диаметр оплодотворенного<br />
яйца около 0.2 мм. С октября по май самки в дальневосточных морях без<br />
оостегитов. Данные за июнь и сентябрь отсутствуют. В сев. <strong>К</strong>алифорнии,<br />
по Мензису, самки с молодью встречены в феврале, мае и июне.<br />
Японское, Охотское и Берингово моря от Приморья до бухты Нагаева<br />
и <strong>К</strong>омандорских о-вов и тихоокеанское побережье сев. Америки на юг<br />
до <strong>К</strong>алифорнии.<br />
Японское море, южн. Приморье, литораль и верхняя сублитораль<br />
(6 экз.). Охотское море, западнокамчатский шельф и бухта Нагаева,<br />
верхняя сублиторалъ (32 экз.). Тихий океан: побережье о-вов Шикотан,<br />
<strong>К</strong>унашир и Итуруп (80 экз.); вост. побережье <strong>К</strong>амчатки (4 экз.), литораль;<br />
побережье Сев. Америки, литораль. Обитает от средней литорали до 2—<br />
3 м глубины, на скалистых и каменистых грунтах, преимущественно<br />
в открытых местах с хорошей аэрацией и водой нормальной солености,<br />
на открытых прибою участках редко встречается на скалах, чаще селится<br />
под валунами и галькой, в расщелинах и ванночках или среди водорослей<br />
Corallina, Iridaea, ризоидов ламинариевых, колоний губок, мшанок и<br />
сложных асцидий. В расщелинах и ваннах литорали встречается в течение<br />
круглого года.
— 47 —<br />
Ianiropsis setifera Gurjanova (рис. 24)<br />
Janiropsis setifera, Гурьянова, 1950 : 282—283, рис. 1.<br />
Основные отличия /. setifera от /. derjugini Gurjan. и /. kincaidi Rich,<br />
были отмечены при характеристике последних. Взрослые самцы /. setifera<br />
легко отличаются от близких видов густым покровом длинных,<br />
тонких щетинок на плеотельсоне и на боковых краях грудных сегментов,<br />
но самки и неполовозрелые самцы, лишенные этого признака, от-<br />
Ряс. 23. Ianiropsis derjugini Gurjan. (котипы), перейоподы, плеоподы и уроподы.<br />
личаются гораздо слабее и иногда относились к /. kincaidi или /. derjugini.<br />
По относительной ширине тела /. setifera занимает промежуточное<br />
положение между указанными видами, хотя чаще в большей степени<br />
приближается к /. derjugini. Длина тела I. setifera в 2.7—3.3 раза превосходит<br />
ширину. <strong>К</strong>ак и у /. derjugini, голова у /. setifera относительно<br />
более широкая, чем у /. kincaidi, и почти такой же ширины, как и плео-
— 48 —<br />
тельсон, а ширина плеотельсона всегда несколько превосходит его длину.<br />
В отличие от I. derjugini, у которого наибольшая ширина плеотельсона<br />
приходится на его переднюю треть, у /. setijera плеотельсон почти округлой<br />
формы и наибольшей ширины достигает примерно в средней части.<br />
Задне-боковые углы плеотельсона у /. setijera меньше оттянуты назад,<br />
Рис. 24, laniropsis setijera Gurjan. (котипы), переноподы, плеоподы и уроподы.<br />
чем у /. derjugini, и в этом отношении описываемый вид больше сходства<br />
имеет с I. kincaidi.<br />
Нами просмотрено 341 экз., хранящихся в коллекциях ЗИН АН <strong>СССР</strong>,<br />
и собранных на литорали дальневосточных <strong>морей</strong> от Приморья до бухты<br />
Нагаева и вост. <strong>К</strong>амчатки. <strong>К</strong>рупные самцы достигают более 5 мм длины.<br />
В августе, сентябре и октябре самки с эмбрионами в выводковых сумках,<br />
данные за другие месяцы отсутствуют. Голотип /. setijera (из вост.<br />
<strong>К</strong>амчатки) утерян; из этого района имеется 6 паратипов (3 вб и 3 £?).<br />
собранных месяцем позже-голотипа. Экземпляры из Приморья отличаются
— 49 —<br />
or всех остальных меньшими размерами л несколько более стройным телом.<br />
Японское море. Приморье, о. Петрова {223 экз.) и о. Путятин (6 экз.).<br />
Охотское море, бухта Нагаева (1 экз.). Тихий океан: Малая <strong>К</strong>урильская<br />
гряда и юго-вост. побережье о. <strong>К</strong>унашнр (96 экз.), вост. <strong>К</strong>амчатка<br />
(6 экз.). Обитает в среднем и нижнем горизонтах литорали, опускается<br />
в самую верхнюю часть суолнторалп до глубины 0.5 м. Селится на скалистых,<br />
валунных п галечных грунтах, под камнями, среди СогаШпа,<br />
Amphiroa, Iridaea и других багрянок, между рнзопдамп ламинариевых,<br />
в ваннах на поверхности рифа. Иногда встречается совместно с /. kincaidi,<br />
а в Приморье — с /. serricaudis Gurjan. В отличие от /. derjugini<br />
и /. kincaidi зимой на южн. <strong>К</strong>урильских о-вах не была найдена.<br />
Ianiropsis serricaudis Gurjanova (рис. 25)<br />
Janiropsis serricaudis Гурьянова, 1936a : 46—47, фиг. 14; 19366 : 251, рпс. 1.<br />
/. serricaudis резко отличается от видов группы /. kincaidi наличием<br />
зазубрин по бокам задней половины плеотельсона, приближаясь к /. Ьгеriremis<br />
Sars, 7. longiantennata Thielemann, Т. analoga Menzies, /. epi-<br />
Рис. 25. Ianiropsis serricaudis Gurjan. (котипы), перейоподы и плеоподы.<br />
littoralis Menzies и /. tridens Menzies. По количеству зазубрин (по 4—7<br />
с каждой стороны) сходен с /. analoga, однако в отличие от последнего<br />
передние зазубрины не достигают середины плеотельсона. <strong>К</strong>роме того,<br />
/. serricaudis отличается от /. analoga слегка вогнутым передним краем<br />
головы, несколько более стройным телом и более удлиненным, широкоовальным<br />
плеотельсоном. От всех видов группы /. breviremis I. serricaudis<br />
отличается более резко выраженным половым диморфизмом, который<br />
4 Труды ЗИН, т. XXX
— 50 —<br />
проявляется в сильном различии формы ногочелюстей (Гурьянова,<br />
1936а, 19366) и I пары перейоподов у обоих полов. У самки ногочелюсти<br />
нормального строения, тогда как у самцов 3—5-й членики щупика сильно<br />
удлинены. Благодаря этому /. serricaudis близок к /. palpalis Barnard<br />
(1914), но особенность /, serricaudis заключается в том, что эти членики<br />
примерно равны по длине. I перейопод самца (рис. 25) сильно превосходит<br />
по длине перейопод самки. Дистальные боковые отростки I плеопода<br />
самца сильнее отогнуты кзади, чем у /. breviremis и видов группы<br />
/. kincaidi. В этом отношении описываемый вид также сходен с /. апаloga,<br />
хотя концы отростков не заострены, как у последнего. <strong>К</strong>рышечка<br />
самки с довольно глубокой вырезкой посредине заднего края.<br />
Максимальная длина самцов 2.9 мм, самок — 2.7 мм. В августе—<br />
октябре самки с эмбрионами в выводковых сумках, данные за другие<br />
месяцы отсутствуют. <strong>К</strong>оличество эмбрионов в сумке колеблется в пределах<br />
от 7 до 32, составляя в среднем 18. Диаметр оплодотворенных<br />
яиц 0.2 мм, длина эмбриона в Ш стадии около<br />
0.3 мм.<br />
Японское море, южн, Приморье, о. Петрова<br />
(Е. Гурьянова), 1934 г., 25 ??, 2 $S и 3 juv.<br />
(котипы); о. Путятин (О. Мокиевский), 1948 г., 1 j.<br />
Обитает в литоральной зоне и верхней части сублиторали<br />
до 2 м глубины, на каменистых и скалистых<br />
грунтах, под камнями, в колониях губок и<br />
сложных асцидий, среди зарослей Amphiroa и<br />
ризоидов ламинариевых, часто вместе ol.setifera<br />
Gurjan.<br />
Рис. 26. laniropsis pal-<br />
Udocula Kussakin, sp.<br />
В.,
— 51 —<br />
10 зазубренных крючков, внутренняя лопасть — 4 толстых и несколько<br />
тонких концевых щетинок; ветви II ыаксиллы примерно равной длины;<br />
2-й и 3-й членики щупика ногочелюстей сильно расширены, 4-й членик<br />
более чем вдвое длиннее 5-го; внутренняя пластинка с 2 соединительными<br />
Ряс. 27. Ianiropsis pallidocula Kussakin, sp. п., в (голотнп), головные придатки.<br />
крючками. <strong>К</strong>арпоподит I перейопода несколько расширен, несет не менее<br />
8 раздвоенных шипов и несколько щетинок по наружному краю.<br />
Задне-боковые углы I плеопода оттянуты в стороны и несколько назад,<br />
.заострены на концах. Длина уропод составляет примерно % длины плеотельсона;<br />
наружная ветвь несколько короче внутренней; база льни ft<br />
членик короче внутренней ветви и примерно такой же длины, как наружная<br />
ветвь.<br />
4*
— 52 —<br />
С а м к и внешне сходны с самцами; длина их тела примерно в 3 раза<br />
превосходит ширину; II антенны короче тела; ногочелюстя сходны с таковыми<br />
самцов; каршшодит I перейопода слабо расширен; крышечка<br />
несколько суживается кзади, задний край ее с широкой выемкой.<br />
Окраска в спирту у обоих полов светлая желтовато-серая, без пятен<br />
темного пигмента. Максимальная длина самца 4.4 мм, самки — 3.4 мм.<br />
Рис. 28. laniropsis pallidocala Kussakin, sp, д. {голопш и аллотип), перейодод<br />
и плеоподы.<br />
Охотское море, западно камчатский шельф, сборы на р/т «Лебедь»,<br />
24 I 1939, ст. 406, верхняя сублитораль, среди ризоидов ламинарий, вместе<br />
с /. derjugini Gurjan., 108 36 (голотипы и паратипы) и 98 $g без оостегитов<br />
(аллотип и паратипы).<br />
От /. serricaudis Gurjan. описанный вид легко отличается формой<br />
переднего края головы, плеотельсона, отсутствием полового диморфизлш<br />
в строении ногочелюстей и другими признаками. Значительно больше<br />
сходства /. pallidocala обнаруживает с /. analoga Menzies, но отличается
— 53 —<br />
от последнего плавно закругленными передне-боковыми краями головы,<br />
более коренастыми перепоподами, сильнее удлиненным 4-м члеником щупика<br />
ногочелюстей и слабой пигментацией глаз.<br />
Род CAECIJAERA MENZIES<br />
Род Caecijaera был установлен Р. Мензисом (Menzies, 1951а) для мелких<br />
<strong>Janiridae</strong> из <strong>К</strong>алифорнии, живущих в ходах лпмнории и, по-видимому,<br />
являющихся ее комменсалами. В отечественных сборах из Баренцева,<br />
Охотского и Японского морен были обнаружены еще 3 новых для науки<br />
вида этого рода, также обитающих в ходах Limnoria. Ниже мы даем описание<br />
этих видов н замечания по характеристике рода Caecijaera.<br />
Caecijaera mirabilis Kussakin, sp. п. (рис. 29—31)<br />
Самец. Тело уплощенное, значительно расширяющееся кзади.<br />
Длина тела в два или в два с небольшим раза превосходит его ширину<br />
(длина голотипа 2.6 мм, ширина 1.25 мм). Поверхность тела покрыта редкими,<br />
короткими щетинками; боковые края головы, грудных сегментов и<br />
плеотельсона усеяны толстыми, игловидными щетинками.<br />
Голова лишена глаз; передний край<br />
головы неровный, образует 5 лопастей, из кото<br />
рых средняя тупо заострена спереди п более других<br />
выдается вперед;'остальные лопасти примерно<br />
равной длины, короткие, расположены ближе<br />
к середине, спереди закруглены; боковые, образующие<br />
передне-боковые углы головы, спереди<br />
тупо заострены. Грудные сегменты примерно<br />
равной длины, ширина их постепенно увеличивается<br />
спереди назад; боковые края I и V —VII<br />
грудных сегментов выпуклые, II — IV сегментов<br />
снабжены неглубокими вырезками, в которых<br />
видны маленькие коксальные пластинки. Брюшной<br />
отдел состоит из 2 сегментов, очень короткого<br />
I брюшного сегмента и широкого, примерно полукруглой<br />
формы плеотельсона. Длина плеотельсона<br />
примерно в 1.5 раза меньше его ширины, его заднебоковые<br />
края плавно закруглены, на заднем крае<br />
по бокам от треугольной медиальной лопасти<br />
имеются две вырезки для уроподов. I антенны<br />
короткие, в отличие от таковых у С. korvathi Menzies,<br />
состоят всего из 5 члеников. Базальные членики<br />
СИЛЬНО расширены И Соприкасаются между рцс, 29. Caecijaera miraсобой<br />
своими внутренними краями; ширина ба- bilis Kussakin, sp. п.,
_ 54 _<br />
новым щупиком; 3-й членик щупика несет по наружному краю 8—9, а 2-й—<br />
2 толстые гребенчатые щетинки. Режущий край мандибулы с 5 зубцами;<br />
подвижная пластинка левой мандибулы с 4—5 зубцами. Зубной ряд щетинок<br />
левой мандибулы состоит из 4 щетинок. Зубной отросток довольно<br />
Рис. 30. Caecijaera mirabilis Kussakin, sp. п. (голотип и аллотип), головные<br />
придатки.<br />
короткий, цилиндрической формы, с усеченным дистальным краем и одной<br />
апикальной щетинкой. Наружная лопасть I максилл несет 6 зазубренных<br />
концевых шипов; внутренняя лопасть широкая, плоская, имеет ряд простых<br />
щетинок. Наружные лопасти II максилл с 4 зазубренными концевыми<br />
щетинками каждая; внутренняя лопасть с 6 простыми концевыми щетинками.<br />
3 первых членика щупика ногочелюстей сильно расширены, каждый
Рис, 31, Caecijaera mirabilis Kussakin, sp. п. (голотип н аллотип), перейоподы,<br />
плеоподы и уропод.
— 56 —<br />
из них значительно шире внутренней пластинки, наружные края 2-го и<br />
3-го члеников щупика оттянуты в широкие, закругленные лопасти. Внутренняя<br />
пластинка несет 2 соединительных крючка. Перейоподы короткие<br />
и крепкие, вооружены незначительным числом щетинок и шипов; 7-е членики<br />
всех перейоподов снабжены 2 концевыми когтями каждый; I перейопод<br />
не отличается значительно от остальных. <strong>К</strong>иль наружного края базиподита<br />
несет 3—4 крепких шипа. I плеопод самца очень широкий и сильно<br />
отличается по форме от такового у С. horvathi; его дистальные боковые<br />
края слегка оттянуты в стороны. II плеопод самца характеризуется<br />
своеобразным устройством копулятивного аппарата — мужской придаток<br />
очень узкий и длинный, свернут по спирали; большая длина его, по-вндимому,<br />
является приспособлением к обитанию в узких ходах лимнории.<br />
Эндоподит III плеопода, как и у С. horvathi, с 5 концевыми перистыми щетинками;<br />
экзоподит более широкий, чем у С. horvathi. Уроподы сходны<br />
с таковыми у С. horvathi, расположены в выемках плеотельсона; протоподит<br />
короткий и широкий, его ширина значительно превосходит длину;<br />
обе ветви уроподов овальной формы, несколько короче протоподита;<br />
внутренняя ветвь шире наружной.<br />
Самка заметно отличается от самца овальными очертаниями тела,<br />
что делает ее сходной по форме с С. horvathi, II плеопод (крышечка) самки<br />
широкий, с широкотреугольным задним<br />
концом. В остальном самки сходны с самцами.<br />
Окраска спиртовых экземпляров обоих<br />
полов желтовато-серая. В октябре и марте<br />
много самок с эмбрионами на разных стадиях<br />
в выводковых сумках, число эмбрионов<br />
в сумке колеблется от 10 до 22.<br />
Охотское море, западнокамчатский<br />
шельф: разрез от устья р. Большой, в древесине<br />
вместе с Limnoria borealis sp. п.,<br />
сборы на р/т «Лебедь», X 1938, 1 6 (голотип),<br />
1 $ (аллотип) и 171 паратип, из-них<br />
42 66, 66 $9 и 63 juv.; ст. 574-575, III<br />
1939, 7 66, И 9$ (из них 3 без сумок и<br />
8 с эмбрионами I—III стадий) и 5 juv.;<br />
VIII 1938, ст. 87, глуб. 32 м, 1 g без сумки.<br />
Рис. 32. Caecijaera derjugini Kussakin,<br />
sp. п., 6 (голотпп).<br />
Caecijaera derjugini Kussakin, sp. п.<br />
(рис. 32—34)<br />
Самец. Тело уплощенное, заметно<br />
расширяющееся кзади. Длина тела в два<br />
или в два с небольшим раза превосходит<br />
его ширину (длина голотипа 2.75, ширина<br />
1.35 мм). Спинная поверхность тела,<br />
особенно плеотельсона, покрыта редкими,<br />
тонкими щетинками; боковые края головы,<br />
грудных сегментов и плеотельсона<br />
усеяны толстыми, игловидными щетинками.<br />
Голова лишена глаз; передний<br />
край головы образует 5 лопастей, из<br />
которых средняя тупо заострена спереди и более других выдается<br />
вперед; передне-боковые лопасти головы несколько длиннее промежуточных;<br />
обе эти пары спереди закруглены. Грудные сегменты при
— 57 —<br />
мерно равной длины; боковые края I и V—VII сегментов выпуклые, II—<br />
IV сегментов сзади снабжены вырезками, в которых расположены маленькие<br />
коксальные пластинки, снабженные по краям игловидными щетинками.<br />
Брюшной отдел состоит нз 2 сегментов: очень короткого I сегмента и широкого,<br />
округлой формы плеотельсона. Длина плеотельсона составляет<br />
Рис. 33. Caecijaera derjugini Kussakin,sp. п., S (голотии), головные<br />
придатки.<br />
около */s его ширины; его задне-боковые края плавно закруглены; на<br />
заднем крае по бокам от округло-треугольной медиальной лопасти имеются<br />
2 вырезки для уроподов. I антенны короткие, состоят всего из 5 члеников.<br />
Базальные членики сильно расширены и соприкасаются менчду собой<br />
внутренними краями; ширина базального членика приблизительно равна<br />
его длине по внутреннему краю. 2-й членик почти вдвое короче и втрое<br />
уже базального. Передние края 1-го и 2-го члеников усеяны шипиками<br />
и игловидными щетинками. 3—5-й членики примерно равной длины,<br />
каждый из них приблизительно вдвое короче 2-го; 3-й примерно в 1.5 раза<br />
уже 2-го; 4-й несколько уже 3-го, а 5-й уже 4-го членика. II антенны также
— 58 —<br />
короткие, длина их немного превышает длину головы. Ножка состоит из<br />
6 члеников. Наружные края 1-го, 2-го и 4-го, а также внутренние края<br />
3-го и 4-го члеников и дистальный край чешуйки, расположенной на 3-м<br />
членике, несут игловидные шипики. Жгутик очень короткий, более чем<br />
вдвое короче ножки, состоит из 10—11, у неполовозрелых особей — из<br />
4—5 члеников. Мандибулы снабжены 3-члениковым щупиком; 3-й членик<br />
щупика несет по наружному краю 10—12 (у неполовозрелых особей 8),<br />
Рис. 34. Caecijaera derjugini Kussakin, sp. п. (голотип и аллотип),<br />
перейоподы, плеоподы и уропод.<br />
а 2-й — 3 толстые гребенчатые щетинки. Режущий край мандибулы с 5 зубцами,<br />
подвижная пластинка левой мандибулы с 4—5 щетинками, зубной<br />
ряд состоит из 3 щетинок. Зубной отросток умеренной длины, почти<br />
цилиндрической формы, несколько расширяется к дистальному краю.<br />
Дистальный край несет 1 шипик и 1 длинную концевую щетинку. Наружная<br />
лопасть I максилл вооружена 6 зазубренными концевыми шипами;<br />
внутренняя лопасть широкая, имеет около 16 простых и зазубренных<br />
щетинок. Наружные лопасти II максилл с 4 зазубренными концевыми щетинками<br />
каждая; внутренняя лопасть несет в среднем 7 щетинок. 3 первых<br />
членика щупика ногочелюстей сильно расширены, каждый из них<br />
значительно шире внутренней пластинки; 4-й и 5-й членики щупика<br />
узкие, примерно в 3 раза уже 3-го членика. Внутренняя пластинка имеет
— 59 —<br />
2 соединительных крючка. Эпинодит значительно более узкий, чем<br />
у С. horvathi Menzies, конец его оттянут в узкий, заостренный отросток.<br />
Перейоподы короткие и крепкие, вооружены незначительным числом<br />
шипиков и щетинок; 7-е членики всех перейоподов снабжены 2 концевыми<br />
коготками каждый, I перейопод не отличается значительно от остальных<br />
Наружные края ишиоподита и базиподита снабжены килями; киль базиподита<br />
несет 4 крепких шипа. I плеопод самца, как и у С. mirabilis Kussakin,<br />
широкий, с оттянутыми в стороны дистальными^боковыми краями.<br />
II плеопод самца, как и у С mirabilis, имеет длинный, узкий, свернутый<br />
по спирали копулятивный придаток, но если у С. mirabilis он образует<br />
по спирали не менее 1.5 оборотов и оканчивается у основания плеопода,<br />
то у С. derjugini Kussakin придаток короче, образует несколько более<br />
1 оборота спирали и оканчивается ниже середины плеопода. У неполовозрелых<br />
особей копулятивный придаток значительно короче и образует<br />
менее 1 оборота. Эндоподит III плеопода несет не менее 8 концевых щетинок;<br />
экзоподит несколько уже, чем у С. mirabilis. Уроподы нормального<br />
для рода строения, с короткими, примерно равной длины ветвями, снабжены<br />
большим количеством шипов и щетинок.<br />
Самка, как ж у С. mirabilis, отличается от самца овальными очертаниями<br />
тела. Длина тела в 2.2—2.6 раза превосходит его ширину на месте<br />
VII грудного сегмента. II плеопод (крышечка) самки округлой формы,<br />
с выступающим округло-треугольным задним концом; ширина его лишь<br />
незначительно превосходит длину. Жгутик II антенн аллотипа состоит<br />
из 9 члеников. Зубной ряд левой мандибуллы состоит из 3, редко 4 щетинок.<br />
Экзоподит III плеопода значительно уже, чем у самца. Длина аллотипа<br />
2.85 мм, максимальная длина самки (экземпляр из зал. Терпения) 3.67 мм.<br />
Окраска спиртовых экземпляров обоих полов серовато-желтая.<br />
Сев.-зап. часть Японского моря, побережье Приморья у м. Егорова,<br />
сборы на э/с «Россинанте», 27 VII 1931, глуб. 91—112 м, ст. 53, в затонувшем<br />
куске древесины, пораженном Limnoria borealis и L. magadanensis,<br />
1 tj (голотип), 4 9? без оостегитов (аллотип и паратипы), 1 неполовозрелый<br />
5 и 37 juv. до 0.9 мм длиной (паратипы). Охотское море, зал. Терпения,<br />
сборы на р/т «Топорок», 1947 г., ст. 99—100, глуб. 27—29 м, 1 { с пустой<br />
выводковой сумкой и 1 § с 32 эмбрионами III стадии длиной около0.7 мм.<br />
Молодь этого вида часто встречается на теле Limnoria sp. вместе с Награсticoida.<br />
Caecijaera borealis Kussakin, sp. п. (рис. 35—37)<br />
Самец. Тело уплощенное, значительно расширяющееся кзади.<br />
Длина тела в 2.2—2.4 раза превосходит его ширину (длина голотипа<br />
2.63 мм, ширина 1.2 мм). Поверхность тела, особенно плеотельсона,<br />
покрыта редкими, тонкими щетинками; боковые края головы, грудных<br />
сегментов и плеотельсона усеяны толстыми, игловидными щетинками.<br />
Голова лишена глаз; передний край ее образует 5 лопастей, из которых<br />
средняя тупо заострена спереди и более других выдается вперед; остальные<br />
лопасти примерно равной длины, короткие, расположены ближе<br />
к середине, тупо заострены, а боковые, образующие передне-боковые углы<br />
головы, спереди закруглены. Грудные сегменты примерно равной длины,<br />
ширина их постепенно увеличивается спереди назад; боковые края I и<br />
V—VII грудных сегментов выпуклые, II—IV сегментов снабжены сзади<br />
вырезками, в которых помещаются небольшие коксальные пластинки,<br />
снабженные игловидными щетинками. Брюшной отдел состоит из 2 сегментов:<br />
очень короткого I сегмента и широкого, округлого плеотельсона.<br />
Длина плеотельсона составляет примерно э /4 ширины; его задне-боковые<br />
края закруглены; на заднем крае, по бокам от округло-треугольной
— 60 —<br />
медиальной лопасти имеются 2 вырезки для уроподов. I антенны короткиег<br />
состоят всего из 5 члеников. Ширина базального членика заметно превышает<br />
его длину по внутреннему краю. 2-й членик примерно вдвое короче<br />
и менее чем в 3 раза уже базального. Передние края 1-го и 2-го члеников<br />
несут шипики и игловидные щетинки. 3-й членик примерно вдвое уже и<br />
короче 2-го; 4-й членик уже, но почти такой же длины, как 3-й, а 5-й уже<br />
и короче 4-го членика. II антенны также короткие, длина их несколько<br />
превышает длину головы; ножка состоит ив<br />
6 члеников; 1 — 5-й членики и хорошо развитая<br />
чешуйка, расположенная на 3-м членике,<br />
несут игловидные шипики. Жгутик короткий,<br />
примерно вдвое короче ножки, состоит<br />
из 8 члеников. 3-й членик щупика мандибул<br />
несет по наружному краю примерно 8Т<br />
а 2-й членик — 2 толстые, гребенчатые щетинки.<br />
Режущий край и подвижная пластинка<br />
левой мандибулы имеют по 5 зубцов. Зубной<br />
ряд щетинок левой мандибулы состоит иа<br />
4 гребенчатых и 1—2 простых щетинок. Дистальный<br />
край цилиндрического, умеренной<br />
длины зубного отростка несет только 1 шипик<br />
и лишен длинной концевой щетинки.<br />
Наружная лопасть I максилл вооружена<br />
6 зазубренными концевыми шипами; внутренняя<br />
лопасть короче наружной, широкая,<br />
несет около 14 простых и зазубренных щетинок<br />
на дистальном крае. Наружные лопасти<br />
II максилл с 4 концевыми щетинками каждая;<br />
внутренняя лопасть несет не менее<br />
16—18 щетинок по дистальному и внутреннему<br />
краям. Ногочелюсти типичного для рода<br />
строения, с 2 соединительными крючками на.<br />
внутренней пластинке, сильно расширенными<br />
3 проксимальными и 2 узкими дистальными<br />
члениками щупика. Эпиподит умеренной ширины,<br />
с почти параллельными боковыми<br />
Рис. 35. Caecijaera borealis<br />
Kussakin, sp. п., с? (голотип).<br />
краями и оттянутым в узкий, заостренный<br />
отросток концом. Перейоподы нормального<br />
для рода строения; киль базиподита имеет<br />
3—4 крупких шипа. I плеопод самца с от<br />
тянутыми в стороны дистальными боковыми краями, напоминает таковой у<br />
С. mirabilis Kussakin и С. derjugini Kussakin. У неполовозрелых самцов дистальные<br />
боковые края оттянуты гораздо слабее. II плеопод, как у С. mi~<br />
rabilis и С. derjugini, снабжен длинным, узким, свернутым по спирали<br />
копулятивным придатком, который у половозрелых самцов относительно<br />
более длинный, чем у С. derjugini, образует значительно более 1 оборота<br />
по спирали и, как у С. mirabilis, оканчивается у проксимального края<br />
плеопода. У неполовозрелых самцов копулятивный придаток короче и<br />
образует менее 1 оборота. Уроподы нормального для рода строения, с ветвями<br />
примерно равной длины.<br />
Самка описываемого вида, как у С. mirabilis и С. derjugini, отличается<br />
от самца овальными очертаниями тела. Длина тела в 2.7—2.9 раза<br />
превосходит его ширину в области VII грудного сегмента. II плеопод<br />
(крышечка) самки широкий, округлый, с выступающим округло-треугольным<br />
задним концом; ширина его значительно превосходит длину. Жгутик
— 61 —<br />
II антенн аллотипа с 8 члениками; 3-й членик щупика мандибул с 8,<br />
2-й с 2 гребенчатыми щетинками. Экзоподит III плеопода уже, чем у самца,<br />
У неполовозрелых самок имеются рудименты I плеоподов, сросшихся<br />
между собой. Число эмбрионов в сумке равно 6—10. Длина аллотипа<br />
2.6 мм, .максимальная длина самки 2.6.5 мм.<br />
Рис. 36. Caecijaera borealis Kussakin, sp, п., с? (голотип), головные<br />
придатки.<br />
Окраска спиртовых экземпляров обоих полов светлая серовато-желтая.<br />
Баренцево море, <strong>К</strong>ольский зал., сборы на э/с «А. <strong>К</strong>овалевский»,<br />
1908 г., ст. 56, 3 3d (голотип и паратипы), 2 JJ с эмбрионами II стадии<br />
{аллотип и паратип), I $ с эмбрионами III стадии длиной 0.68 мм, 1 $ без<br />
оостегитов и 3 неполовозрелые особи длиной 1.3—1.55 мм (паратипы);<br />
сборы Научно-промысловой экспедиции по исследованию Мурмана на<br />
з/с «Андрей Первозванный», VI 1901, ст. 193, 1 9 и ст. 975, 1 $; сборы
— 62 —<br />
Плавучего морского института 7 VII 1925, ст. 259., глуб. 140 м, грунт —<br />
серо-зеленый ил, 1 6 и 3 juv. Все особи найдены в кусках древесины, пораженной<br />
Limnoria borealis.<br />
Описанные виды Caecijaera обнаруживают много черт сходства с единственным<br />
ранее известным видом этого рода С. horvathi Menzies. Сходными<br />
Рис. 37. Caecijaera borealis KussaJsin, sp. п. (голотип, аллотип и паратипы), перейоиоды,<br />
плеоподы и уронод.<br />
признаками являются местообитание этих видов в ходах лимнории,<br />
комменсалами, а, вероятно, частично и паразитами которой они являются,<br />
общее строение тела, ротовых придатков, грудных ножек и уроподов,<br />
а также отсутствие глаз. Вместе с тем 3 описанных намк вида Caecijaera<br />
существенно отличаются от С. horvathi иным строением I и II плеоподов<br />
самца и мандибул. У С. mirabilis Kussakin, С. derjugini Kussakinn С. borealis<br />
Kussakin мандибула еще сохраняет типичное для семейства <strong>Janiridae</strong><br />
строение, имеет хорошо развитую подвижную пластинку, зубной<br />
ряд щетинок и цилиндрический зубной отросток. Последний, правда,<br />
значительно укорочен, и его основание несколько сдвинуто по направле-
— 63 —<br />
нию к подвижной пластинке. У С. horvathi мандибула больше отклоняется<br />
от исходного типа, подвижная пластинка укорочена, зубной ряд щетинок<br />
редуцирован и содержит на левой мандибуле всего 1 щетинку, зубной<br />
отросток сильно уменьшен в размерах и смещен вверх, занимая место<br />
нижней части зубного ряда щетинок на мандибулах нормального строения,<br />
I плеопод самца у С. mirabilis, С. derjugini и С. borealis широкий,<br />
его боковые дистальные углы оттянуты в стороны, тогда как у С. horvathi<br />
I плеопод сильно сужен в дистальной половине, его дистальные боковые<br />
углы оттянуты назад и несколько внутрь. Строение копулятивного придатка<br />
II плеопода самца также значительно различается у этих групп<br />
видов: у С. horvathi он сохраняет обычную длину, у 3 же описанных видов<br />
сильно удлинен, свернут по спирали и значительно превышает длину<br />
плеопода. <strong>К</strong>роме того, у С. horvathi I антенны состоят из 6 члеников,<br />
видимых сверху коксальных пластинок нет, а тело самца имеет овальное<br />
очертание, в то время как у С. mirabilis, С. derjugini л С. borealis I антенна<br />
состоит из 5 члеников, на II—IV грудных сегментах сверху видны небольшие<br />
коксальные пластинки, а тело самца плавно расширяется спереди<br />
назад. Эти отличия, особенно в строении мандибул, выходят за пределы<br />
обычных видовых, и, следовательно, 3 описанных вида следует относить<br />
к отдельному подроду Caecijaerella subgen. п.<br />
Эти 3 вида, хотя и стоят значительно ближе друг к другу, чем к С. horvathi,<br />
все же легко различаются между собой. Помимо отличий в форме<br />
тела, они различаются по следующим признакам. У С. derjugini 2-й членик<br />
щупика мандибулы имеет 3 крупные, гребенчатые щетинки, тогда как<br />
у С. mirabilis и С. borealis только по две; 3-й членик щупика мандибулы несет<br />
у С. mirabilis 8—9, у С. borealis 8, а у С. derjugini 10—12 гребенчатых<br />
щетинок. Жгутик II антенн состоит у С. mirabilis и С. derjugini из 10—<br />
11, а у С borealis из 8 члеников. <strong>К</strong>опулятивный придаток у половозрелых<br />
самцов С. derjugini образует примерно 1 оборот спирали и оканчивается<br />
ниже середины плеопода, тогда как у С. mirabilis и С. borealis он относительно<br />
более длинный, образует значительно более 1 оборота и оканчивается<br />
у основания плеопода. Другие отличия между этими видами отмечались<br />
при их описании.<br />
Диагноз рода Caecijaera, составленный Р. Мензисом (Menzies, 1951а)<br />
на основании рассмотрения всего 1 вида, теперь нуждается в некотором<br />
уточнении. Ниже мы приводим диагноз Р. Мензиса с нашими изменениями.<br />
Диагноз. Глаза отсутствуют. I антенна состоит из 5—6 члеников, ее<br />
базальный членик сильно расширен. II антенна довольно короткая, с 6-члениковой<br />
ножкой, отчетливой чешуйкой на 3-м членике и небольшим жгутиком,<br />
состоящим из 8—14 члеников. Боковые края грудных сегментов<br />
по краям несут игловидные шипики, или же все выпуклые, без видимых<br />
сверху коксальных пластинок, или же края II—IV сегментов сзади снабжены<br />
вырезками, в которых расположены коксальные пластинки. Дистальные<br />
боковые углы I плеопода самца несколько оттянуты в стороны или назад<br />
и внутрь. III плеопод с перистыми щетинками на эндоподите. IV плеопод<br />
состоит из единственной мясистой ветви, лишенной щетинок. Дактилоподиты<br />
всех перейоподов с 2 коготками. 3 первых членика щупика ногочелюсти<br />
не менее широкие, чем внутренняя пластинка; два дистальпых<br />
членика значительно уже проксимальных. I максилла с широкой, уплощенной<br />
внутренней ветвью; наружная ветвь с длинными, зазубренными крючками.<br />
Зубной отросток мандибулы более или менее уменьшен в размерах.<br />
Уроподы расположены в выемках ялеотельсона, разделенных между<br />
собой треугольной лопастью.<br />
Подрод Caecijaera s. str. характеризуется следующими признаками:<br />
I антенна состоит из 6 члеников, зубной отросток мандибулы сильно умень-
— 04 —<br />
пген в размере, зубной ряд щетинок левой мандибулы состоит из единственной<br />
щетинки, копулятивный придаток II плеопода самца короткий,<br />
нормального строения, видимые сверху коксальные пластинки отсутствуют.<br />
Подрод Caecijaerella subgen. п. характризуется следующими признаками:<br />
I антенна состоит из 5 члеников, зубной отросток мандибулы сохраняет<br />
нормальное строение, слабо уменьшен в размере, зубной ряд щетинок<br />
левой мандибулы состоит из 3 или более щетинок, копулятивный придаток<br />
II плеопода самца длинный, значительно длиннее плеопода и свернут по<br />
спирали, на II — IV грудных сегментах имеются видимые сверху коксальные<br />
пластинки.<br />
Тип подрода С. mirabilis Kussakin;<br />
Род Caecijaera, как правильно отмечает Р. Мензис, имеет более всего<br />
черт сходства с родами lais и особенно Jaera, но является несомненно более<br />
специализированным. Такими признаками специализации являются отсутствие<br />
глаз, укороченные II антенны и видоизмененный зубной отросток<br />
мандибул. Оба подрода обнаруживают разную степень развития этих<br />
признаков. Строение мандибул более примитивно у подрода Caecijaerella,<br />
тогда как жгутик II антенн и II плеопод самца, наоборот, у этого подрода<br />
сильнее специализированы, чем у Caecijaera в. str.<br />
ЛИТЕРАТУРА<br />
Гурьянова Е. Ф. 1932. Морские арктические равноногие раки (<strong>Isopoda</strong>). Определит,<br />
по <strong>фауне</strong> <strong>СССР</strong>, изд. ЗИН АН <strong>СССР</strong>, 4, 181 стр., 189 рис.<br />
Г урьянова Е. Ф. 1933а. <strong>К</strong> <strong>фауне</strong> равноногих раков (<strong>Isopoda</strong>) Тихого океана.<br />
II. Новне виды Gnathiidea и <strong>Asellota</strong>. Исслед. <strong>морей</strong> <strong>СССР</strong>, 19: 79—91, рис. 1—15<br />
(Гурьянова Е. Ф.) Gurjanova Е. 19336. Die marinen Isopoden der Arktis.<br />
Fauna arctica. Bd. 6, Lief. 5 : 391—470, 2 fig.<br />
Гурьянова E. Ф. 1936a. Равноногие дальневосточных <strong>морей</strong>. Фауна <strong>СССР</strong>, нов.<br />
сер., Ракообразные, VII, 3, 279 стр., 150 рис.<br />
(Гурьянова Е. Ф.) Gurjanova Е, 19366. Zur Fauna der <strong>Isopoda</strong> des Stilien<br />
Ozeans. Zool. Anz., Bd. 114 : 250—265.<br />
Гурьянова E. Ф. 1950. <strong>К</strong> <strong>фауне</strong> равноногих раков (<strong>Isopoda</strong>) Тихого океана. V.<br />
Иаоподы по сборам <strong>К</strong>амчатской морской станции Государственного гидрологического<br />
института. Исслед. дальневост. <strong>морей</strong> <strong>СССР</strong>, 2 : 281—292, рис. 1—3.<br />
Гурьянова Е. Ф. 1952. Crustacea — Malacostraca Чукотского моря и Берингова<br />
пролива. <strong>К</strong>райний северо-восток <strong>СССР</strong>. II. Фауна и флора Чукотского моря:<br />
169—215, рис. 1—7.<br />
Barnard <strong>К</strong>. Н. 1914. Contributions of the Crustacean Fauna of South Africa. I.<br />
Additions to the Marine <strong>Isopoda</strong>. Ann. South African Mus. X, 7 : 195—230.<br />
Boone P. 1920. <strong>Isopoda</strong>. Report of the Canadian Arctic Exped. 1913—1918. Crustacea,<br />
VII, D : Id—40d.<br />
Hansen H. J. 1888. Oversigt over det vestlige Grcnlands Fauna af Malakostrake<br />
Havkrebsdyr. Vidensk. Meddel. Naturh. Foren. Kjabenh. (Ш7) : 177—198.<br />
Hansen H. J. 1916. Crustacea Malacostraca. III. <strong>Isopoda</strong>. Danish Ingolf-Exped.,<br />
vol. 3, № 5, 262 pp., 16 pis.<br />
Hatch M. 1947. The Chelifera and <strong>Isopoda</strong> of Washington and adjacent regions<br />
Univ. Washington Publ., Biol., 10 : 155—274.<br />
<strong>К</strong> r 0 у e r H. 1846—1849. Karcinologiste Bidrag. Naturh. Tjidskr. (2), II: 1—123, 366-<br />
446.<br />
<strong>К</strong> г 0 у e г H. 1849. Voyages en Scandinavie, en Laponie, au Spitzberg et aux Feme.<br />
. Zoologie, Crustacea. Atlas : pi. XXVIII, figs. 1-2; pi. XXIX, fig. 1.<br />
Menzies R. J. 1951a. A new genus and new species of Asellote Iaopod, Caecijaera<br />
horvathi, from Los Angeles Beach Harbor. Amer. Mus. Novit., № 1542 : 1—7,<br />
figs. 1—3.<br />
Menzies R. J. 1951b. New marine isopods, chiefly from Northern California, with<br />
notes on related forms. Proc. V. S. Nat. Mus., vol. 101, № 3273: 105—156,<br />
figs. 9—33.<br />
Menzies R. J. 1952. Some marine Asellote Isopods from Northern California, with<br />
descriptions of nine new species. Proc. U.S. Nat. Mus., vol. 102, № 3293 : 117—<br />
159, figs. 46-71.
— 65 —<br />
О h 1 i n A. 1901. Arctic Crustacea collected during tbe Swedish Arctic expeditions<br />
1898 and 1899 under the direction of. Prof. A. G. Nathorst. 1. Leptostraca, <strong>Isopoda</strong>,<br />
Cumacea. Bihang. till Kongl. Svenska Vet .-Acad. Handling, XXVI,<br />
Afd. IV, № 12 : 15—40,"pis. I—V.<br />
О г t m a n n A. E. 1900. Preliminary report on the Crustacea and Pycnogonida of tbe<br />
Princeton Arctic expedition, 1899. Princeton Univ. Bull., XI, № 3 : 39—40.<br />
Ortmann A. E. 1901. Crustacea and Pycnogonida collected during the Princeton<br />
Expedition to North Greenland. Proc. Acad. Nat. Sci. Philad : 144—168.<br />
Richardson H. 1899. Key to the Isopods of the Pacific coast of North America,<br />
with descriptions of twenty-two new species. Proc. U. S. Nat. Mus., XXI : 815—<br />
S69.<br />
Richardson H. 1900. Synopses of North-American Invertebrates. VIII. The <strong>Isopoda</strong>.<br />
Amer. Naturalist, XXXIV : 207—230, 295—309.<br />
Hicliardson H. 1901. Key to the Isopods of the Atlantic coast of North America,<br />
with descriptions of new and little-known species. Proc. U. S. Nat. Mus., XXIII:<br />
493—579.<br />
Richardson H. 1904. Isopod crustaceans of the Northwest coast of North America.<br />
Harriman Alasca Exped., Crustacea, 10 ; 213—230,<br />
Richardson H. 1905a. A monograph on the Isopods of North America. Bull.<br />
U. S. Nat. Mus., № 54, LHI 4- 727 pp., 740 figs.<br />
Ri с hardson. H. 1905b. Isopods from the Alasca Salmon Investigations. Bull.<br />
Bureau Fish., 24 : 209—221.<br />
S a rs G. O. 1897—1899. An account of the Crustacea of Norway, II, <strong>Isopoda</strong>, X - 170 pp.,<br />
100 pis.<br />
Stebhing/T. R . R. 1900. Arctic Crustacea : Bruce Collection. Ann. Mag. Nat.<br />
Hist. (7), V : 1 —16.<br />
Stephensen K. 1912. Report on the Malacostraca, Pycnogonida and some Entomostraca<br />
collected Ъу the Danmark-Expedition to N. E. Greenland. Meddel.<br />
Gronl. vol. 45, Л« 11 : 501—618.<br />
Stephensen K. 1913. Account of the Crustacea and the Pycnogonida collected hy<br />
Dr. V. Nordmann in the summer of 19И from Northern Stromfjord and Giesecke<br />
Lake in West Greenland. Meddel. Granl., 51 : 53—77.<br />
Stephensen K. 1943. Leptostraca. Mysidacea, Cumacea, Tanaidacca. <strong>Isopoda</strong><br />
and Euphausiacea. The Zoology of East Greenland. Meddel. Grenl. vol. 121, № 10 :<br />
1-82, 11 figs.<br />
Thielemann M. 1910. Beitrage zur Kenntnis der Isopodenfauna Ost-Asiens.<br />
Abhandl. math.-phys. Kl. K. Bayer. Acad. Wiss., II, Suppl., Bd. 3, Abhandl.:<br />
1-110, 2 Taf., 87 Fig.<br />
W a 1 к e г A. O. 1898. Crustacea collected hy W. A. Herdman in Puget Sound, Pacific<br />
coast of North America. Trans, Liverpool Biol. Soc, XII : 280—281, pi. XV,<br />
figs. 7—10.<br />
7, Труды ЗИН, т. XXX