
免疫系统中的负向调控——髓系来源的抑制性细胞(MDSC)的大作用!
早在上世纪 80 年代 , 人们就在肿瘤患者体内发现了一群具有抑制性的 髓系来源的细胞(Myeloid-derived suppressor cells ,MDSCs), 但是直到最近几年,人们才对这群细胞在免疫系统中的负向调控作用给予了更多关注[1]。下面一起来了解MDSCs吧!
一、MDSCs的生物学特征及诱导产生机制
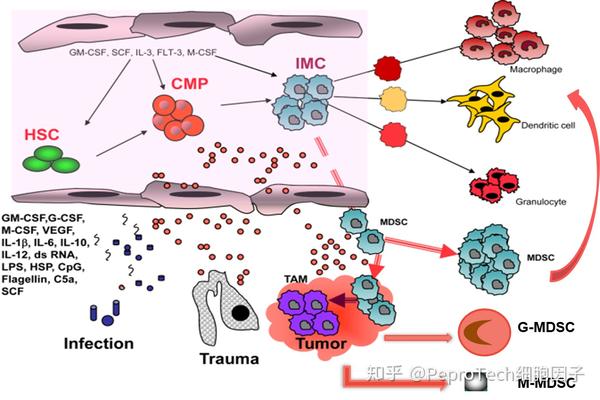
在肿瘤 、细菌或寄生虫感染、脓毒血症、创伤、骨髓移植以及一些自身免疫性疾病等病理状态下, MDSCs 会大量产生和累积,从而对机体的免疫系统起到抑制作用,MDSCs 由共同髓系祖细胞(Common myeloid pro-genitor , CMP)发育分化而来 。造血干细胞分化为共同髓系祖细胞,共同髓系祖细胞再分化为未熟髓系细胞(immature myeloid cells,IMCs),在健康个体中,IMCs进一分化为成熟的中性粒细胞、巨噬细胞或树突状细胞。而当机体发生肿瘤、炎症、感染、创伤等病理情况时,IMCs的正常分化可被炎性因子或肿瘤来源的细胞因子阻滞,诱导其成为MDSCs,并在外周血、骨髓或病变部位募集、扩增、活化,机体内MDSCs细胞比正常的中性粒细胞或单核细胞存活时间短[2]。

*1、MDSCs在小鼠中表型
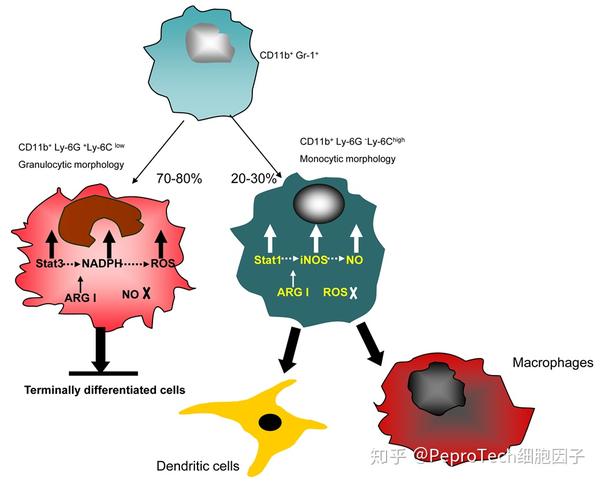
目前研究认为 MDSCs在小鼠中表型为CD11b+Gr1+,根据其表面Ly6G和Ly6C的不同表达可进一步分为单核细胞来源的MDSC(Monocytic-MDSCs)和粒细胞来源的MDSC(Granulocytic-MDSCs)两个亚群。Monocytic-MDSCs(M-MDSCs)的表型为CD11b+Ly6G-Ly6Chigh ,而Granulocytic-MDSCs(G-MDSCs)的表型为CD11b+Ly6G+Ly6Clow[3];
*2. MDSCs在人体中表型
人的MDSCs表型在不同疾病中的表型更为多样化,在人体中MDSCs是表达 Lin-HLA-DR-CD33+或 CD11b+CD14-CD33+的细胞群体[1]。人体内MDSC也可分为粒细胞样MDSC(G-MDSC)和单核样MDSC(M-MDSC),G-MDSCs常见表型为CD11b+CD14-CD15+HLA-DR-CD33+, 而M-MDSCs表型为CD11b+CD14+CD15-HLA-DRlowCD33+[4]。
也有研究者针对不同癌症,通过表面标记区分其亚群。在肾癌病人中,用CD11b+CD14-CD15+ 定义G-MDSC。在黑素瘤病人中,用CD11b+CD14+HLA-DR -/low 定义M-MDSC。
近来研究者还确立了新的表面标记来区分MDSC的亚群: 高水平的CD66b、低水平CD62L 和低表达CD16 等[5]。另外,有研究在脐血[6]或转移性小儿肉瘤患者外周血内[7]发现一个新的MDSC 亚群,即纤维细胞样MDSC ( Fibrocytic myeloid-derived suppres-sor cells,F-MDSC ) ,其表型特点是HLA-DR+、CD33+或CD33-/low[6, 7],通过吲哚胺2,3 双加氧酶( Indoleane-2,3-dioxygenase, IDO) 可诱导调节性T细胞( regulatory T cells,Treg) 产生或直接抑制T细胞增殖。然而在小鼠肿瘤模型或其他肿瘤患者中尚未发现该亚群,还有待进一步研究。人类MDSC亚群的多样性可能与不同肿瘤中MDSC 的诱导和扩增机制有关,目前尚不清楚。由于MDSCs没有特异性标记物,因此对于MDSCs最重要的鉴定方法是明确该群细胞是否具有免疫抑制功能。
二、MDSCs的免疫抑制功能及其临床应用
MDSCs能抑制机体免疫细胞发挥正常固有性和适应性免疫功能,在固有免疫方面,MDSCs 通过膜结合TGF-β使NKG2D的表达下调,从而抑制 NK 细胞的功能[8],MDSCs可诱导 Treg扩增,促进Treg对免疫的负调控作用[9]。在适应性免疫方面,MDSCs可以竞争性消耗体内环境中的半胱氨酸,上调iNOS和Arg-1的活性从而消耗L-精氨酸,使T细胞生成受阻,MDSCs 也可以通过产生活性氧(Reactive oxygen species , ROS)来抑制T细胞免疫应答反应。MDSCs如果与T细胞共培养,可以看到T细胞功能和增殖明显受到抑制[10]。MDSCs 亚群对T细胞功能的抑制机制有所不同,M-MDSCs主要是通过激活STAT1,显著上调iNOS的表达,而对ROS的的调控不明显,G-MDSCs主要是通过激活STAT3,上调NADPH,影响ROS的表达,而不影响iNOS。两种类型的MDSC均可上调Arg-1的活性。使免疫细胞处于无应答或耐受状态,不能及时清除变异的癌前细胞及失去杀伤肿瘤细胞的能力[1]。此外,MDSCs还通过促进肿瘤血管生成、肿瘤细胞侵袭及向远处器官转移等非免疫机制参与肿瘤的生物学行为。近来的研究表明,MDSCs在肿瘤环境中可继续分化成肿瘤相关巨噬细胞,并可分为抑制肿瘤生长的M1亚群和促进肿瘤生长的M2亚群。由于肿瘤组织中生长因子和细胞因子微环境的作用,MDSC主要以M2表型存在,如诱导肿瘤组织向M1表型分化,则可使肿瘤生长减缓、抗肿瘤特异性细胞增加、新生血管生成减少、肿瘤转移受抑制。MDSCs细胞与中性粒细胞相比能产出更高水平的ROS,胞内的溶酶体更多,在肿瘤微环境中更容易被趋化到肿瘤局部,但是其吞噬能力下降[11]。

MDSCs作为肿瘤患者体内发现的一群具有抑制性的髓系来源的细胞,被大量研究阐述了其与肿瘤的发生发展的密切关系。Sun HL等分析了在结肠癌患者肿瘤组织中的MDSCs促进了肿瘤的远端转移[12]。Diaz-Montero等研究表明MDSCs在体内的富集与临床上乳腺癌的分型呈正相关[13],并且肿瘤患者血液中MDSCs比例越高,肿瘤的愈后越差[14]。通过分析实体瘤患者肿瘤组织中MDSCs的富集与生存率的关系证实MDSCs的富集会大大降低患者的生存率[15, 16]。并且外周血中MDSCs已经成为一些循环系统肿瘤的预测指标。近些年,许多针对MDSCs的靶向治疗药物也应用于临床,其可显著提高肿瘤患者免疫细胞的反应,有效抑制肿瘤的生长[17]。
三、MDSCs的体外扩增及其免疫抑制功能检测方法
实验证明,GM-CSF 可以诱导骨髓来源MDSCs 的产生,并且浓度越高诱导产生的MDSCs 速度越快,但是同时也会产生中性粒细胞和树突状细胞( Dendritic cell,DC) [18]。T 细胞、树突细胞和自然杀伤细胞( Natural killer cell,NK) 能分泌高浓度的GM-CSF,导致MDSCs 在炎症部位聚集。G-CSF 在炎症条件下对粒细胞的产生和转移发挥重要作用。实验证明肿瘤模型中G-CSF 的产生与MDSC 的生成直接相关,高浓度的G-CSF 会影响髓系细胞分化与发育,从而导致粒细胞样MDSCs的生成[19]。在人类肾癌细胞系中M-CSF 被发现有与IL-6 一起发挥作用的趋势,能够阻止DC 的分化并引起单核样MDSCs 产生[20]。
研究MDSCs功能机制需要在体外获得大量的MDSC,目前有两种途径可以获得MDSCs,第一种途径是通过分离荷瘤小鼠脾脏或者肿瘤组织中的MDSCs,第二种途径是通过在体外诱导分化髓系祖细胞增殖分化成MDSCs。MDSCs的扩增主要通过JAK和STAT3信号通路的介导,依赖于GM-CSF、M-CSF、IL-6、SCF、VEGF、COX-2等细胞因子的诱导[21, 22],Marigo等人研究报道了40ng/ml GM-CSF和IL-6能够诱导骨髓细胞扩增为MDSCs[23]。许秋桂等人联合GM-CSF和IL-6刺激与单独使用GM-CSF或IL6比较,可以显著提高MDSCs在体外诱导效率[24]。MDSCs体外扩增体系的建立为MDSCs在肿瘤、炎症等疾病中的免疫学机制研究奠定了基础。
MDSCs表型没有特异性,因此我们对于MDSCs的鉴定除了满足表面标记的表达外,最重要的鉴定方法是确定该细胞是否具有免疫抑制功能,在体外我们通常是做T细胞增殖抑制实验来验证MDSCs的功能,
具体实验方案如下:
▶T细胞增殖抑制实验(小鼠MDSCs功能验证)
1. 取OT1小鼠淋巴结(腹股沟、肠系膜)制成单细胞悬液,重悬到2x107/ml。2. 活细胞染色(CFSE):
1)细胞用PBS重悬到1x107/ml。
2) 加入等体积的CFSE染液,使终浓度为5 μM,37℃/10min,避光孵育。
3)加入5倍体积的无安全培养基(含10%FBS)冰上孵育5min。
4)加入完全培养基洗一次,500 g/10min,离心,弃上清。
5)细胞用完全培养基重悬到2x106/ml。
3. 分选的MDSC细胞重悬到1x106/ml及5x105/ml。
4. 将MDSC细胞和T细胞各50ul种于96孔板内(U底)加入100ul刺激剂(5 μg/ml OVA peptide)(SIINFEKL)。5. 将细胞培养3天后收细胞染CD8a-APC,4℃/30min,500g/10min,PBS洗一遍,弃上清,PBS重悬,FACS检测。
四、总结与展望
综上所述,MDSCs是一群异质性较强的髓系细胞群体,本文就MDSCs 生物学特征,诱导产生机制,免疫抑制机制及与疾病的关系等方面都进行了介绍。MDSCs的发现为临床在肿瘤的治疗中提供了很多新的思路,针对MDSCs已经研制出来很多药物,可通过增强免疫系统机能的角度来实现癌症治疗。
尽管如此,MDSCs缺少特异的表面标志,一定程度上阻碍了对MDSCs 复杂发育过程的理解,我们仍然难以确定的是MDSCs 在疾病不同阶段还是其不同亚群介导发挥了其免疫抑制功能,此外对人类MDSCs 亚群的区分,MDSCs 诱导和激活细胞内信号通路的调节机制,MDSCs的表观遗传调控机理及其对其他免疫细胞的影响还不是很清楚,有待进一步研究,解决这些问题将有助于我们利用MDSCs 抑制免疫应答的功能,为基于MDSCs的治疗提供新的干预靶点。
MDSCs鉴定推荐流式染色方案
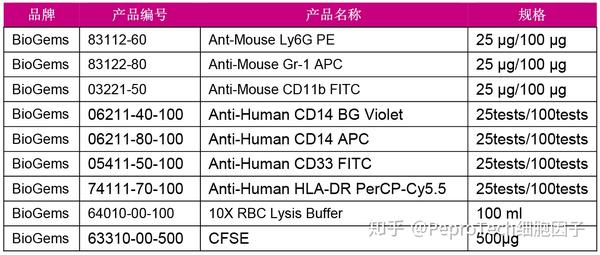

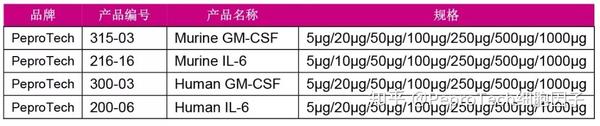
MDSC培养推荐细胞因子

参考文献
1. Talmadge, J.E. and D.I. Gabrilovich, History of myeloid-derived suppressor cells. Nature reviews. Cancer, 2013. 13(10): p. 739-52.
2. Condamine, T., et al., ER stress regulates myeloid-derived suppressor cell fate through TRAIL-R-mediated apoptosis. The Journal of clinical investigation, 2014. 124(6): p. 2626-39.
3. Movahedi, K., et al., Identification of discrete tumor-induced myeloid-derived suppressor cell subpopulations with distinct T cell-suppressive activity. Blood, 2008. 111(8): p. 4233-44.
4. Montero, A.J., et al., Myeloid-derived suppressor cells in cancer patients: a clinical perspective. Journal of immunotherapy, 2012. 35(2): p. 107-15.
5. Dumitru, C.A., et al., Neutrophils and granulocytic myeloid-derived suppressor cells: immunophenotyping, cell biology and clinical relevance in human oncology. Cancer immunology, immunotherapy : CII, 2012. 61(8): p. 1155-67.
6. Zoso, A., et al., Human fibrocytic myeloid-derived suppressor cells express IDO and promote tolerance via Treg-cell expansion. European journal of immunology, 2014. 44(11): p. 3307-19.
7. Zhang, H., et al., Fibrocytes represent a novel MDSC subset circulating in patients with metastatic cancer. Blood, 2013. 122(7): p. 1105-13.
8. Li, H., et al., Cancer-expanded myeloid-derived suppressor cells induce anergy of NK cells through membrane-bound TGF-beta 1. Journal of immunology, 2009. 182(1): p. 240-9.
9. Serafini, P., et al., Myeloid-derived suppressor cells promote cross-tolerance in B-cell lymphoma by expanding regulatory T cells. Cancer Research, 2008. 68(13): p. 5439-5449.
10. Rodriguez, P.C. and A.C. Ochoa, Arginine regulation by myeloid derived suppressor cells and tolerance in cancer: mechanisms and therapeutic perspectives. Immunological reviews, 2008. 222: p. 180-91.
11. Brandau, S., et al., Myeloid-derived suppressor cells in the peripheral blood of cancer patients contain a subset of immature neutrophils with impaired migratory properties. Journal of leukocyte biology, 2011. 89(2): p. 311-7.
12. Sun, H.L., et al., Increased frequency and clinical significance of myeloid-derived suppressor cells in human colorectal carcinoma. World journal of gastroenterology, 2012. 18(25): p. 3303-9.
13. Markowitz, J., et al., Myeloid-derived suppressor cells in breast cancer. Breast cancer research and treatment, 2013. 140(1): p. 13-21.
14. Diaz-Montero, C.M., et al., Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin-cyclophosphamide chemotherapy. Cancer immunology, immunotherapy : CII, 2009. 58(1): p. 49-59.
15. Zhang, B., et al., Circulating and tumor-infiltrating myeloid-derived suppressor cells in patients with colorectal carcinoma. PloS one, 2013. 8(2): p. e57114.
16. Cui, T.X., et al., Myeloid-derived suppressor cells enhance stemness of cancer cells by inducing microRNA101 and suppressing the corepressor CtBP2. Immunity, 2013. 39(3): p. 611-21.
17. Iclozan, C., et al., Therapeutic regulation of myeloid-derived suppressor cells and immune response to cancer vaccine in patients with extensive stage small cell lung cancer. Cancer immunology, immunotherapy : CII, 2013. 62(5): p. 909-18.
18. Kong, Y.Y., et al., Myeloid derived suppressor cells and their role in diseases. Current medicinal chemistry, 2013. 20(11): p. 1437-44.
19. Abrams, S.I. and J.D. Waight, Identification of a G-CSF-Granulocytic MDSC axis that promotes tumor progression. Oncoimmunology, 2012. 1(4): p. 550-551.
20. Menetrier-Caux, C., et al., Inhibition of the differentiation of dendritic cells from CD34(+) progenitors by tumor cells: role of interleukin-6 and macrophage colony-stimulating factor. Blood, 1998. 92(12): p. 4778-91.
21. Saleem, S.J. and D.H. Conrad, Hematopoietic cytokine-induced transcriptional regulation and Notch signaling as modulators of MDSC expansion. International immunopharmacology, 2011. 11(7): p. 808-15.
22. Khaled, Y.S., B.J. Ammori, and E. Elkord, Myeloid-derived suppressor cells in cancer: recent progress and prospects. Immunology and cell biology, 2013. 91(8): p. 493-502.
23. Marigo, I., et al., Tumor-induced tolerance and immune suppression depend on the C/EBPbeta transcription factor. Immunity, 2010. 32(6): p. 790-802.
24 许秋桂, 王文红等, 小鼠髓样抑制细胞体外扩增体系的建立. 现代免疫学, 2015, 35, 2.