第五节 铜

铜(copper,Cu)是在1878年由Fredrig从章鱼血的蛋白质中分离出,并将这种含铜蛋白质称为铜蓝蛋白。1900年发现在喂全奶饲料的动物中出现贫血而不能用补充铁来预防。1928年,Hart报道大鼠贫血只有在补铁同时补充铜才能得到纠正,故认为铜是哺乳动物的必需元素。1912年,Wilson’s病被发现后,首次将人类疾病与铜代谢联系起来。此后报道了营养不良婴儿有铜缺乏症。1962年,Menke’s病被发现后,经过10年研究,该病才被认为是铜吸收紊乱所致。此后,铜的毒性也得到了认识和描述。
一、理化性质
铜是一个过渡金属,原子序数为29,其相对原子质量为63.55。铜的氧化还原化学性质,使它特别适合于释放和接受电子,尤其是将电子直接传递给分子氧。铜在溶液和活的生物体中,多以Cu 2+ 和Cu + 两种价态存在,且以前者为主,两者可以互相转化,而很少作为Cu 3+ 存在。Cu + 在有氧或其他电子接受体存在的情况下很容易被氧化成Cu 2+ ;Cu 2+ 可以从强还原性物质如抗坏血酸盐或还原谷胱甘肽接受电子而还原为Cu + ,这一可逆过程是铜化合物生化反应的功能基础。
人体内的铜主要以含铜蛋白的形式存在,大多数属于含铜氧化酶。这些含铜蛋白可分为三大类:第一类是深蓝色蛋白质,存在于许多含铜的氧化酶中;第二类也存在于多种含铜的氧化酶中,但不是蓝色的,可用电子顺磁共振(electron paramagnetic resonance,EPR)测出;第三类既非蓝色又不能被EPR测出,但也在许多酶中存在。一种酶可以含一种铜蛋白,也可以含多种铜蛋白。生物系统中许多涉及氧的电子传递和氧化还原反应都是由含铜酶催化的,这些酶对生命过程都是至关重要的。
二、吸收与代谢
(一)吸收
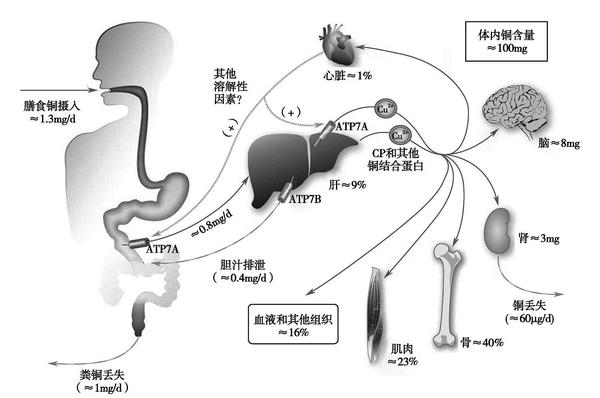
铜主要在十二指肠被吸收,少量由胃和小肠末端吸收。铜吸收率与摄入量成负相关关系,且受饮食中其他因素的影响,在12%~75%范围内波动。每天摄入铜0.4mg时其吸收率为70%;当每天摄入量增加到7.5mg时,吸收率则下降为12%,即吸收量从0.3mg增加到0.9mg。年龄和性别对铜吸收未见明显影响。铜在体内的平衡部分受吸收的调节,而铜的吸收又受机体对铜的需要所调节。当摄入量增加时体内铜贮存量随之增加,摄入量为7~8mg/d时贮存量约1mg/d。植物性食物中铜的吸收率约为33.8%,而动物性食品中铜吸收率约41.2%。膳食中其他营养素对铜的吸收利用可产生影响。如锌摄入过高可干扰铜的吸收,因为过量的锌可以诱导肠道内金属硫蛋白的合成,继而与铜结合将其隔离在肠细胞中,阻碍铜的吸收。但当锌∶铜比值在15∶1或更低时则影响很小。对于婴儿而言,Cu与Fe的协同作用是最为关键的,10.8mg Fe/L配方奶喂养儿与1.8mg Fe/L配方奶喂养儿相比,铜吸收率下降。此外,钼、维生素C、蔗糖和果糖等营养素摄入量对铜的吸收利用也可能产生影响。
膳食中铜水平低时,其吸收机制可能是主动运输;膳食中铜水平高时被动吸收则起作用。膳食中的铜在经过胃和肠道时,释放出铜离子或铜氨基酸复合物,而后与肠道中的高分子量蛋白质(主要是超氧化物歧化酶和金属硫蛋白)结合后运输到肠黏膜上。在Cu 2+ 被还原为Cu + 后被人类铜转运蛋白(hCTR1或DMT1)转运至胞质,并很快被胞质内的蛋白如金属硫蛋白或铜伴侣蛋白螯合,前者起着调节铜储存的作用,后者将铜运送至转运蛋白ATP7A(铜转运P型ATP酶)以备基底外侧流出进入血液,这一过程是肠细胞摄取和短暂储存铜的关键。
(二)转运
铜经胃肠道吸收后,可保留原来形式,也可与其他配体如L-氨基酸、多肽、蛋白质、脂肪酸、胺等进行交换形成其他络合物进入循环。铜主要以三种方式通过血浆运送至器官被摄取和利用:一是血浆铜蓝蛋白,血清中90%~95%铜与血浆铜蓝蛋白结合,较稳定;二是白蛋白结合铜,约5%~10%铜与血浆白蛋白或L-氨基酸结合,这部分铜可交换和转递;三是小分子结合铜。回到肝脏的血浆铜蓝蛋白被新生铜蓝蛋白置换后分解并转运到胆汁排出。剩余的血浆铜约有12%与α2-巨球蛋白结合,约18%与白蛋白结合,其他与小肽和氨基酸结合。铜的转运需要铜伴侣蛋白,铜伴侣蛋白是铜稳态调节的重要实现者,能将所载运的铜精确运送至结合位点,参与目的蛋白装配,同时具有保护机体免受游离铜离子的毒性作用。
(三)分布
成人体内铜含量约为1.5~2.0mg/kg,其中约有50%~70%存在于肌肉和骨骼中,20%在肝内,5%~10%在血液中。各组织中铜的含量最低者不足1μg/kg,高者超过10μg/kg,其中以肝、肾、心、头发和脑中最高,脾、肺、肌肉和骨骼次之,脑垂体、甲状腺和胸腺最低。胎儿和婴儿铜水平与成人不同,出生后头两个月的婴儿铜浓度是以后的6~10倍,这种铜的储存可能为婴儿期所需。
血液中的铜主要分布于红细胞和血浆,在红细胞中约60%的铜存在于铜锌超氧化物歧化酶(SOD1)中,其余40%与其他蛋白质和氨基酸松弛地结合。正常人红细胞中铜约为14.2~15.7μmol(0.9~1.0mg/L)。血浆中铜约有93%牢固地结合于铜蓝蛋白,其余7%与白蛋白和氨基酸结合。与白蛋白疏松结合的铜是运输、吸收、排泄的重要形式和中间环节,也是合成各种细胞蛋白的原料。正常人血清或血浆中铜水平为12.6~18.9μmol/L(0.8~1.2mg/L),其浓度较稳定,不受循环节律和进食等因素影响,女性比男性约高10%,在妊娠后期或口服以雌激素为基础的避孕药,其浓度可升高。妊娠期对铜储存很重要,分娩时产妇肝中铜是一般成人浓度的5~10倍,孕妇所储存的铜可供胎儿生长和母奶喂养婴儿所用。
(四)排泄
一般认为铜不是储存元素,它通常很容易从体内排出,然而多数或所有组织的细胞都能以金属硫蛋白的络合物形式将过多铜储存起来,主要储存在肝中。铜的主要排泄途径是通过胆汁到胃肠道,再与进入胃肠道的铜以及少量来自小肠细菌的铜一起由粪便中排出。由胆汁排泄入胃肠道的铜约10%~15%可被重新吸收。体内对铜的平衡调节,胆汁排泄起着重要作用,所以,对胆管阻塞患者的铜摄入量要严格监测。内源性铜的排泄量明显受铜摄入量的影响,铜摄入量低时几乎没有内源性铜的排泄且铜周转率低,铜摄入量增加时内源性铜的排泄增加且周转加快。健康人每日经尿液排泄铜10~30μg(0.2~0.5μmol),经汗及皮肤通常丢失50μg以下,此外,皮肤、指甲、头发也丢失铜。
综上,哺乳动物铜代谢的主要路径如图1-8-9所示。

三、生理功能
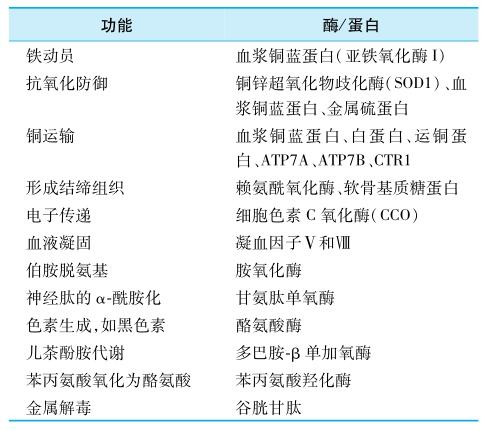
铜参与铜蛋白和多种酶的构成,在人体内发挥重要生理功能。含铜酶有胺氧化酶、亚铁氧化酶I(铜蓝蛋白)、亚铁氧化酶П、细胞色素C氧化酶、多巴胺β-羟化酶、超氧化物歧化酶、单酚单氧酶等。铜结合蛋白有金属硫蛋白、转铜蛋白、凝血因子V、低分子量配体(包括氨基酸和多肽)等。这些铜结合蛋白和酶参与的主要生物学过程如表1-8-10所示。

铜的功能是多方面的,依赖含铜的蛋白质和酶,通过相关生物学过程,实现如下生理功能。
(一)维持正常造血功能
铜参与铁的代谢和红细胞生成。亚铁氧化酶Ⅰ和Ⅱ可使Fe 2+ 氧化为Fe 3+ ,使铁离子结合到运铁蛋白,对生成运铁蛋白起主要作用,并可将铁从小肠腔和贮存点运送到红细胞生成点,促进血红蛋白的形成。故铜缺乏时红细胞生成障碍,可产生寿命短的异常红细胞,表现为缺铜性贫血。正常骨髓细胞的形成也需要铜。缺铜可引起线粒体中细胞色素C氧化酶活性下降,使Fe 3+ 不能与原卟啉合成血红素,可引起贫血。铜蓝蛋白功能缺损也可使细胞产生铁的积聚。
(二)促进结缔组织形成
铜可通过赖酰氧化酶促进结缔组织中胶原蛋白和弹性蛋白的交联,为形成结缔组织所必需。因此,铜在皮肤和骨骼的形成、骨矿化、心脏和血管系统的结缔组织完善中起着重要作用。
(三)维护中枢神经系统的健康
神经髓鞘主要由磷脂形成。磷脂合成依赖于细胞色素氧化酶,故细胞色素氧化酶能促进髓鞘的形成。在脑组织中多巴胺β-羟化酶催化多巴胺转变成神经递质肾上腺素,该酶也与儿茶酚胺的生物合成有关。已报道缺铜可致脑组织萎缩、灰质和白质变性、神经元减少、精神发育停滞、运动障碍等。
(四)促进正常黑色素形成及维护毛发正常结构
酪氨酸酶能催化酪氨酸羟基化转变为多巴并进而转变为黑色素,为皮肤、毛发和眼睛所必需。先天性缺酪氨酸酶,引起毛发脱色,称为白化病。巯基氧化酶具有维护毛发的正常结构及防止其角化,铜缺乏时毛发角化并出现具有铜丝样头发的卷发症,称为Menke’s病。
(五)保护机体细胞免受超氧阴离子的损伤
有三种以上含铜酶具有抗氧化作用,包括广泛分布的超氧化物歧化酶(SOD)、细胞外的铜蓝蛋白和主要在细胞内的铜硫蛋白等。SOD能催化超氧阴离子转变为过氧化物,过氧化物又通过过氧化氢酶或谷胱甘肽过氧化物酶作用进一步转变为水。铜蓝蛋白是几种自由基的清除剂,并可保护特别容易被羟基氧化和破坏的不饱和脂肪酸,在体液中起抗氧化剂的作用。含铜酶在保护机体免受过氧化损伤作用方面是很重要的。
四、缺乏与过量
(一)缺乏
人类铜缺乏较少,认识也较晚。1956年首次报道了人类非缺铁性贫血,同时可见血浆铜含量低的病例。随后,1964年Cordano等报道了营养不良儿童的铜缺乏状况。有报道表明极度营养不良儿童在恢复期100%存在铜缺乏。临床上铜缺乏较少见,铜缺乏常常与其他营养素缺乏同时存在,且症状较轻。
引起铜缺乏的原因可分为先天性和后天性两种,前者主要由遗传性铜代谢紊乱引起,如Menke’s病,后者主要与饮食有关。其他系统紊乱、疾病或治疗亦可增加铜缺乏的风险,如乳糜泻、Crohn’s病、肠道吸收疾病、AIDS和自身免疫病等。长期使用高剂量解酸剂或其他阳离子螯合物会降低人对铜的吸收能力,接受腹膜透析治疗的患者会损失较多的血浆铜蓝蛋白。
1.缺铜性贫血
缺铜时铁转运受阻,一方面使红细胞生成障碍,造血功能下降,另一方面使某些细胞中铁聚集。铜缺乏时人体血红蛋白合成减少,并有短寿命的异常红细胞产生,易发生小细胞低色素性贫血,亦可为正常细胞或大细胞性。网织细胞增加或减少,常低于1500个/mm 3 ,白细胞数亦减少,骨髓象改变。
2.心血管受损
含铜酶是心脏和动脉壁中结缔组织的必要成分,对冠心病的形成起着重要的抑制作用。铜缺乏时可出现心电图异常、心脏收缩功能受损、线粒体呼吸功能受损和心肌肥大等,常伴有高血压和主动脉狭窄等。同时,由于含铜酶合成减少,影响人体心肌细胞的氧化代谢,会导致脂质累积,胆固醇增加。铜缺乏可引起赖氨酰氧化酶活力下降,使弹性蛋白和胶原的生物合成减少而导致心脏和动脉组织强度降低引起破裂,以至死亡。孕妇铜缺乏可导致胎儿心脏、血管发育受损和脑畸形。
3.中枢神经受损
婴儿铜缺乏会引起中枢神经系统的广泛损害。有研究表明铜缺乏导致的氧化性应激可迅速降低老年痴呆症患者的认知能力。动物实验发现母代缺铜可引起子代神经功能紊乱,临床可见运动失调和高死亡率。
4.影响结缔组织功能和骨骼健康
铜缺乏可引起机体骨骼、血管、皮肤中胶原蛋白和弹性蛋白的交联受损,诱发骨质疏松、血管破裂、动脉瘤、皮肤粗糙缺少光泽。发育期缺铜可导致骨畸形,老年缺铜易产生骨质疏松。
5.Menke’s病
又称Menke’s卷发症,是一种先天性铜代谢紊乱疾病,以中枢神经损伤为主,头发卷曲色浅为特征。幼儿Menke’s病多以骨骼缺陷如骨质减少和自发性肋骨骨折为特征。患儿血液、肝和脑中铜含量低,但在某些组织和器官中由于铜无法正常排出细胞而产生聚集,血清铜及血浆铜蓝蛋白含量减少,铜的吸收量降低。含铜酶活性降低是诊断本病的重要依据,给以铜盐(硫酸铜)治疗则可使血清中铜浓度迅速恢复,血浆中铜蓝蛋白浓度渐趋正常,一般情况逐渐好转而康复。
(二)过量
铜对于大多数哺乳动物是相对无毒的。由于人体自身调节机制,Wilson’s病以外的铜中毒在人体中较为少见。人体急性铜中毒偶见于误食铜盐、食用铜污染的食物或饮料,摄入铜量往往超过20g。大剂量铜的急性毒性反应包括:口腔有金属味、流涎、上腹疼痛、恶心、呕吐及严重腹泻。
慢性铜中毒表现为肝脏铜聚集,继而引发结构和生化性质的改变包括慢性间质性肝炎等。慢性铜中毒一般经历两个阶段:第一阶段,没有明显中毒症状,血铜浓度正常或偏高,但肝酶浓度增高明显,这是铜在肝脏中逐渐聚集的反应,可导致富含酸性磷酸酶的肝细胞肿大及含铜枯否细胞(Kupffer细胞)的灶性坏死;第二阶段发生非常迅速,常出现溶血危象,肝脏中会产生广泛变性、点状坏死、炎细胞浸润及胆汁淤积。
慢性铜中毒主要见于Wilson’s病,它与铜在肝及其他组织中达到毒性水平的聚集有关,并非铜摄入量过多所致。Wilson’s病患者常见慢性肝脏损伤和(或)精神损伤,并常伴随有肾功能障碍,眼、血液及骨骼病变也较常见。虽然患者肝铜水平较高,但血浆铜蓝蛋白和血液中铜浓度却较低。限制铜摄入量对该病疗效甚微。目前的治疗方法通常靠口服锌制剂降低铜的吸收和(或)服用螯合剂如D-青霉胺、二巯基丙醇(BAL)或硫代钼酸盐以增加铜的排泄。
肝硬化在印度儿童中发生率较高,研究表明与环境、烹饪方法等导致摄入铜过高有关。其原因是食用铜锅中煮的牛奶,可使铜含量增加0.13~6.35mg/L,以致造成铜中毒。一般发生于6个月到5岁儿童,其特征是肝、尿和血清中铜浓度增加,症状包括腹胀、不规则发热、过度哭泣、食欲改变、黄疸,如不治疗,几个月内发展为肝脾大、腹水、水肿,通常死于出血、继发性感染或肝昏迷。这也可能与曾报道的印度儿童肝硬化的先天性疾病(Indian childhood cirrhosis,ICC)有关,后者是一种基因缺陷的遗传性疾病,在有高铜摄入后才出现铜中毒症状,伴随着在肝中快速积聚,可引起死亡。
五、营养状况评价
经常用来反映铜代谢的血液指标是血清铜和血浆铜蓝蛋白浓度,可有效地诊断Menke’s病、Wilson’s病、中度及重度铜缺乏,但它们属于急性期指标,缺乏灵敏性和特异性以反映边缘性的铜缺乏。含铜酶如红细胞SOD、血小板细胞色素C氧化酶、血浆二胺氧化酶、组织中赖氨酰氧化酶、组织与血浆中肽基甘氨酸α-酰胺化酶(peptidyl glycine α-amidase)的活性都曾被研究用作反映铜缺乏的生物标志,但相关实验证明它们均不足以灵敏而特异地反映早期铜缺乏。有研究表明缺铜大鼠组织和红细胞中的SOD铜伴侣蛋白(CCS1)升高;营养不良儿童的周围单核细胞(peripheral mononuclear cells)实验结果也肯定了这一结论;健康人体单核细胞中的CCS1在中度铜过量时会明显降低。因此铜伴侣蛋白可能会作为既反映缺乏又反映过量的铜营养状况生物标志。
(一)血清铜
是铜缺乏的可靠指标,用于个体则要慎重。正常人血清铜范围为10.0~24.6μmol/L(640~1560μg/L)。女性比男性约高10%。女性妊娠期血清铜可高出一倍。而当发现铜缺乏病例时,血清铜浓度已远低于此下限。补充铜可使血清铜浓度在几天内恢复到正常水平。
(二)血浆铜蓝蛋白(Cp)
正常人血浆Cp水平为180~400mg/L,Cp浓度经常与血清中铜浓度相平行,也是铜缺乏的常用指标之一,但Cp对轻度铜缺乏不灵敏,铜缺乏病例Cp已远低于180mg/L。Cp对补充铜反应很快,但也不能反映膳食中铜摄入量。一般情况下,由于Cp会随雌激素、妊娠和服用避孕药而升高,男性血浆铜蓝蛋白含量低于女性。Cp也是一种急性反应蛋白,可因炎症、感染、类风湿、恶性肿瘤、心肌梗死或传染性疾病等疾病而升高,这种情况下血清铜和铜蓝蛋白水平不能用作诊断铜缺乏指标。此外,血浆铜蓝蛋白还与年龄和季节变化有关,因此,不能用于评估铜的过量。
(三)红细胞中超氧化物歧化酶(SOD)
红细胞中Cu/Zn SOD-1活性受膳食中碳水化合物的影响,当食物成分以果糖为主时可采用SOD-1作为生物标志,以玉米淀粉为主食时不适合。铜耗尽时SOD-1活性下降,但补充铜却未见其上升。另外一个不利因素是缺乏标准方法,临床测试和实验室比对困难,SOD-1也是一种急性相反应物,同样易受健康状况与应激反应的影响。
(四)血小板中细胞色素C氧化酶(CCO)
绝经妇女摄入铜0.57mg/d 105天后,摄入2mg/d 35天后,与血清铜、Cp或红细胞SOD-1相比,CCO是变化最为明显的生物标志,但其活性与铜营养状况是否呈平行变化仍未定论。
(五)赖酰氧化酶
肽基甘氨酸-α-酰胺单氧化酶(PG-α-AM或PAM)、二胺氧化酶等也可能是较敏感的评估膳食中铜含量指标,但因实验数据非常少,尚须进一步观察。
(六)铜平衡实验
过去已经有用平衡实验来评估膳食中推荐量的研究,但需要长期适应,在很广的膳食摄入量范围均能获得平衡,所以此方法对铜需要量的研究不适合应用。
(七)其他
采用许多功能试验来评估铜的营养状态可能有价值,例如免疫功能和抗氧化功能,但缺乏特异性。稳定同位素研究对全身铜的测量、铜库交换、铜周转等这些领域的研究仍有待进一步发展。铜代谢分隔舱模型(compartmental model)对评价铜营养状态可能也有帮助。发、指甲或唾液中铜水平不能完全反映铜的营养状况,尿铜个体差异很大,所以至今尚未见一个简单指标足以评估铜的营养状况。然而,将已确定的多个指标结合起来在评估铜的营养状况中是有价值的。
六、膳食参考摄入量
国外多数国家新修订的膳食铜参考摄入量,除婴儿为AI外,均提出了EAR和RNI(RDA)。2013年中国营养学会修订我国居民膳食铜DRIs时,参考了国外研究资料,也提出我国居民膳食铜的EAR和RNI。
(一)AI与RNI
1.成人
国外制定成人膳食铜的EAR时,主要采用了补充试验的方法。由于没有单一指标可以准确评估成人铜的营养状况,因此采用血清铜、血浆铜蓝蛋白、红细胞SOD活性和血小板铜浓度等多项指标综合分析评估,以期确定EAR。美国采用补充试验研究资料显示,膳食铜摄入量为0.7mg/d时,可覆盖男性和女性半数人群的需要,故确定膳食铜的EAR值为0.7mg/d(男女平均体重68.5kg)。日本也采用该方法获得男女成人膳食铜的EAR分别为0.7mg/d和0.6mg/d。我国缺乏铜需要量研究数据,参照美国研究资料,按我国男女平均体重61kg折算,计算出我国成年人铜的EAR为0.623mg/d,取整数处理,修订EAR为0.60mg/d。设定CV为15%推算并取整数处理,修订成年人铜的RNI为0.80mg/d。
2.儿童和青少年
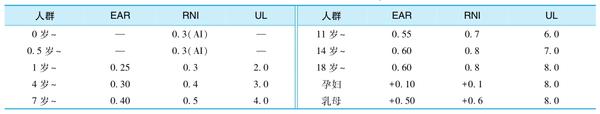
目前缺少儿童或青少年铜的EAR研究数据,中国居民DRIs(2013版)采用成人EAR和代谢体重法推算。设CV=15%,推算RNI,结果见表1-8-11。
3.孕妇和乳母
妊娠期妇女对铜的需要量包括满足自身和满足胎儿生长发育所需的量。国外研究资料显示,以胎儿所需铜总量为13.7mg,妊娠期为280天,妊娠期铜生物利用率为60%,则我国孕妇铜EAR应额外增加0.082mg/d,取整处理,修订为0.1mg/d。CV取值为15%,则孕妇RNI应额外增加0.107mg/d(1.3×0.082mg/d),取整处理,修订为0.1mg/d。
哺乳期妇女铜的EAR需要补加泌乳丢失的铜量0.29mg/d,考虑60%的生物利用率,则乳母铜的EAR应额外增加0.48mg/d,取整处理,修订为0.5mg/d。同样,CV取值为15%,中国居民膳食营养素参考摄入量(2013版)中制定乳母RNI应额外增加0.63mg/d(1.3×0.48mg/d),取整处理,修订为0.6mg/d。
4.婴儿
根据我国部分地区母乳铜含量数据范围在0.27~0.41mg/L之间,平均值为0.38mg/L。0~6月龄婴儿的AI值为0.3mg/d(0.38mg/L×0.75L/d)。7~12月龄婴儿缺乏辅助食物的铜含量,则采用代谢体重法从0~6月龄婴儿的AI值推算为0.39mg/d,从成人代谢体重法推算为0.25mg/d,取其均值0.32mg/d,取整后7~12月龄婴儿AI为0.3mg/d。
(二)UL
根据研究资料,美国食物和营养委员会确定铜的NOAEL为10mg/d。不确定系数UF定为1,结果得到铜的UL为10mg/d。英国和日本成人UL也为此值。欧盟食品科学委员会(SCF)确定成人UL为5mg/d。国际膳食补充剂协会联盟(IADSA)确定UL为9mg/d。WHO建议平均安全范围上限成人男性为12mg/d,女性为10mg/d。
中国营养学会根据以上资料,推测成年人从食物、饮水和膳食补充剂中获得的铜超过上述摄入量上限的可能性非常低。而且铜中毒很少见,即使从食物中摄入的铜水平高到10mg/d左右也未见中毒现象。因此结合我国实际情况,铜的不确定系数UF定为1.2,以铜的NOAEL为10mg/d,得到成人铜的UL为8mg/d。儿童和青少年UL参照与成年人的平均体重比值得出,结果见表1-8-11。
中国居民各年龄组膳食铜参考摄入量见表1-8-11。

七、主要食物来源
铜广泛存在于各种食物中,牡蛎、贝类海产品食物以及坚果(如巴西坚果和腰果)是铜的良好来源,动物的肝、肾,谷类胚芽部分、豆类等次之,植物性食物铜含量受土壤中铜含量及加工方法的影响。奶类和蔬菜含量最低。
在普通膳食中,天然食品如谷类、肉类和鱼类等可以提供50%的铜摄入量。通常成年人每天可以从膳食中得到约2.0mg铜,基本上能满足人体需要。食物中铜吸收平均为40%~60%。牛奶中铜含量很低,人乳含量较高。人乳在产后第一周含量为9.8~14.1μmol/L(0.62~0.89mg/L),几个月后降至2.4~2.7μmol/L(0.1~0.17mg/L)。初生婴儿每天从母乳中可获得铜50g/kg,后来逐渐下降。牛奶铜含量平均为3.8~12.6μmol/L(0.24~0.8mg/L),故人乳铜含量高于牛奶,长期用牛奶进行人工喂养以及母乳喂养后期的婴儿应注意铜的营养状况。
