
【10x Genomics 单细胞转录组Nature文章】在发育性骨化和骨折愈合期间间充质干细胞分化的隔细胞吸收软骨

在发育和再生过程的软骨性骨化中,软骨逐渐被新形成的骨取代。这一过程中,软骨的再吸收对于侵入生长的血管和开始骨化至关重要。循环单核细胞融合产生的破骨细胞会降解骨骼,而负责软骨吸收的细胞身份是一个长期存在且有争议的问题。
2022年1月,来自德国明斯特大学医学院的研究者们在Nature communication上发表了题为Mesenchymal stromal cell-derived septoclasts resorb cartilage during developmental ossification and fracture healing的文章,该文影响因子达17.694,文章使用了共聚焦显微成像,免疫荧光染色和单细胞 RNA 测序 (scRNA-seq)等技术方法系统研究了发育过程中,单细胞分辨率下的再生成骨。并证明了软骨再吸收过程是由少量间充质细胞来源的 FABP5+ 细胞(隔细胞,Septoclasts)介导的。该研究结果将有助于理解骨骼的再生过程,也有助于制定改善骨折愈合的治疗策略。


文章详细内容如下:
1 在软骨-骨边界,吸收细胞的形态学特征
(Morphological characterization of resorptive cells at the chondro-osseous border)
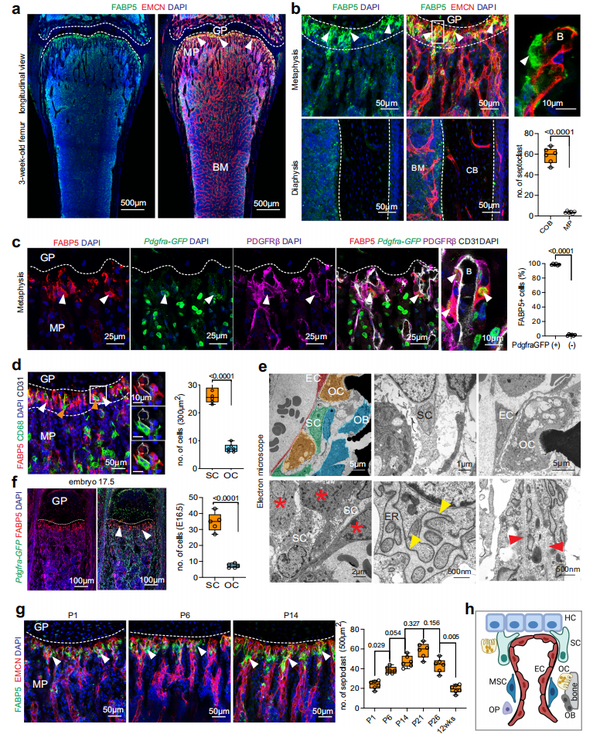
为了深入了解生长板的吸收过程,作者用一系列标记对 3 周龄的小鼠股骨切片进行了免疫染色,发现 FABP5+ 细胞主要位于软骨边界区(图 1a、b )。FABP5+ 细胞与远端血管芽处的 ECs 相关,位于脉管系统的前端,因此可能代表隔细胞。特别需要注意的是,这些 FABP5+ 细胞表达成熟的血管周围间充质细胞标志物,例如血小板衍生的生长因子受体 β(PDGFRβ)和 PDGFRα(图 1c )。以 CD68 和 V 型质子 ATP 酶亚基 B (vATPase) 共表达为标志的骨吸收性多核破骨细胞广泛分布于整个干骺端,可见于软骨-骨边界区但缺乏 FABP5 表达 (图 1d )。此外,FABP5+ 细胞在软骨-骨界面处明显比破骨细胞丰富(图 1d)。电子显微镜 (EM) 下股骨干骺端异常的细胞群与小血管密切相关,可能代表隔细胞。形态学上,这些细胞呈纺锤形,细长和单核,表现出强肌动蛋白丝,众多水泡结构和扩大的粗面内质网,这一特征在浆细胞中也可见,与高水平的蛋白质产生相关(图 1e )。这些细胞与肥大的软骨细胞和骨细胞、破骨细胞和 ECs 进行物理接触(图 1e),但与周细胞不同,它们不嵌入内皮下基底膜中。免疫染色显示 PDGFRα+ 和 PDGFRβ+ 界定的隔细胞表达蛋白多糖 NG2,这是一种周细胞和其它间充质细胞群的标志物,但缺乏 CD146/MCAM 的表达,CD146/MCAM 是周细胞和血管平滑肌细胞的另一种标志物。总之,上述超微结构特征、FABP5 表达及其在软骨-骨边界的定位与隔细胞一致,而 PDGF 受体和 NG2(许多间充质细胞群的标志物)的表达需要进一步研究。EdU 标记表明 FABP5+ 细胞在出生后早期长骨形成期间高度增殖。在胚胎期 (E) 16.5 至 17.5 的胎儿骨骼发育过程中,FABP5+ 细胞主要位于初级骨化中心 (POC) 的血管前部,而 CD68+ 细长的 OCs 遍布整个 POC,而没有在生长板边缘积累(图 1f )。FABP5+ 细胞主要在靠近生长板软骨细胞的干骺端远端和近端的血管前部检测到,并且在出生后长骨发育到第 21 天数量逐渐增加到最大(图 1g )。颅骨是在没有软骨细胞的情况下,通过膜内骨化形成的扁平骨 。值得注意的是,FABP5+ 细胞在 3 周大的颅盖骨中很少见,并且与穿透颅骨的血管有关。这些数据表明骨中的 FABP5+ 细胞主要是与软骨内成骨相关,并在软骨-骨界面处代表一个动态细胞群,将它们与破骨细胞区分开来 (图 1h)。

2 FABP5+细胞的分子特性和分化
(Molecular properties and differentiation of FABP5+ cells)
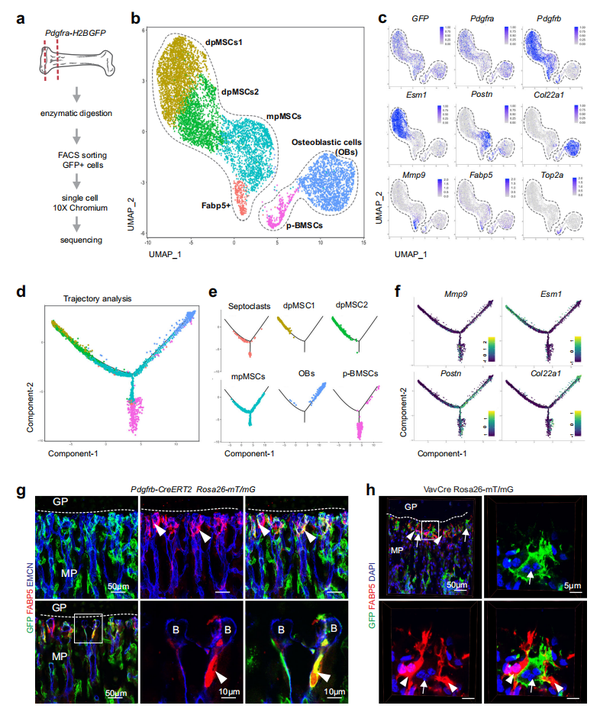
FABP5+细胞的起源和分子特性,假定是隔细胞(Septoclasts,SCs),是小鼠出生后长骨中非常罕见的细胞群,仍有待阐明。因此,作者使用 10× Genomics Chromium 平台进行了单细胞 RNA 测序 (scRNA-seq)。利用 FABP5+ 群体的 PDGFRα 表达,作者通过流式细胞分选(FACS) 从 3 周大的 Pdgfra-GFP 报告小鼠的干骺端 - 骨骺区分离出表达增强型绿色荧光蛋白 (GFP) 的细胞 (图 2a)。基于液滴的 scRNA-seq(参见“方法”)产生了 11,242 个 GFP+ 细胞,属于几个不同的亚群(图2b)。GFP+细胞包括骨间充质基质细胞(BMSCs)和成骨细胞(OBs),可以通过表达 Pdgfrb 和 Col22a1来区分,还有 Acan+ 软骨细胞和 S100a4+ 成纤维细胞(图 2b、c )。BMSC 和 OB 种群(7857 个细胞)的重新聚类确定 6个亚群,即骨干间充质基质细胞1 (dpMSCs1;#2382 细胞)和dpMSCs2(#1659 细胞)表达 Pdgfrb 和 Esm1,干骺端间充质基质细胞 (mpMSCs;#1642 细胞)表达 Pdgfrb 和 Postn,Col22a1+ OB(#1629 细胞),表达Top2a+ 和 Mki67+的增殖性 BMSC(p BMSC;#302 细胞)和少量细胞(#243 细胞) 表达 Fabp5+ 和基质金属蛋白酶 Mmp9 (图 2b、c 和补充图 4d)。轨迹分析表明, BMSC 和 OB 细胞群分为三个独立的方向,包含 dpMSCs1 、dpMSCs2、OBs 和 p-BMSCs 细胞,而 mpMSCs 形成中心 (图 2d,e)。需要注意的是,Fabp5+ 界定的 SCs ,仍然接近 mpMSCs 。伪时间基因表达分析表明,确实来自这个群体(图2d-f)。为了研究这个问题,作者结合了免疫染色和遗传命运追踪他莫昔芬诱导型 Pdgfrb-CreERT2 系,通过 GFP 的表达可以标记 PDGFRβ+ BMSCs 。在 P21 处对靶向 GFP+ 细胞的分析显示了 FABP5+ 细胞的标记(图 2g 和补充图 5a、b)。相反,FABP5+ 假定的 SCs 在 Vav1-Cre R26-mTmG 小鼠中未标记,其在造血细胞中显示出广泛的 GFP 表达,因此也在 CD68+ 破骨细胞中表达(图 2h )。这些数据表明,与破骨细胞不同,软骨-骨边界处的 FABP5+ 细胞并非来自造血谱系,而是来自干骺端的 PDGFRβ+ 间充质基质细胞。鉴于 FABP5+ 细胞群表现出SCs的主要特征,该研究结果表明SCs来源于骨髓间充质干细胞,因此与破骨细胞不同,破骨细胞来自造血细胞。

3 Septoclasts 在长骨形成中的作用
(Function of septoclasts in long bone formation)
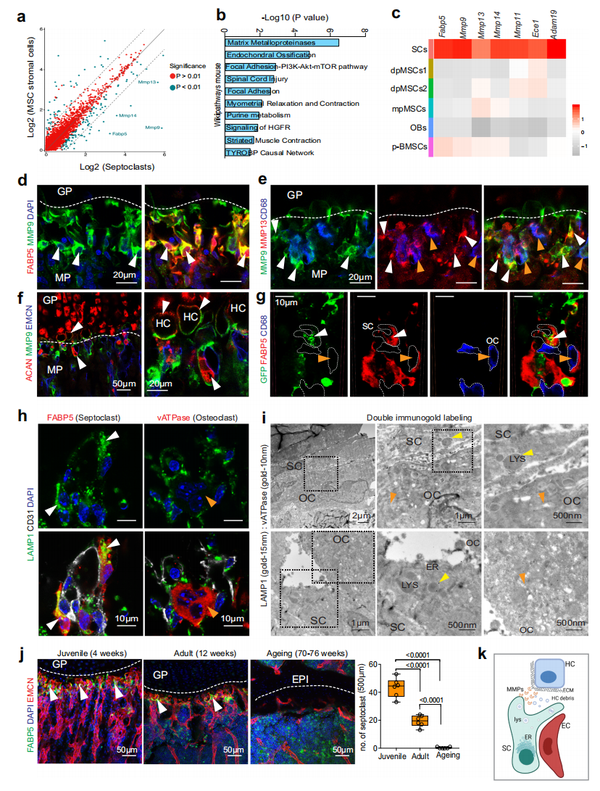
利用scRNA-seq 数据,对 FABP5+ 界定的 SCs 的生物学功能和分子特性进行深入研究(图 3a)。GO富集分析显示细胞外基质组织、细胞外基质分解和蛋白水解是这些细胞参与的生物过程。细胞通路分析表明,与 MMP、软骨内骨化和粘着斑信号通路相关的基因也很丰富(图 3b)。对 scRNA-seq 数据分析表明蛋白酶基因Mmp9、Mmp13、Mmp14、Mmp11、Ece1、Adam19和Ctsb(编码组织蛋白酶 B)在 Fabp5+细胞群中高度表达,认定它确实代表吸收细胞, 因此是隔细胞(SCs)(图 3c )。其它高表达基因与 Rho 家族 GTP 酶(Rhod、Rhoj)、细胞骨架(Tubb2a、Myh9、Vasp、Actn1、Tln1、Flna)、 和细胞 - 基质相互作用(Fn1,Lama4,Itgb5,Itgav)相关。与上述发现一致,MMP9 免疫信号是血管生成和成骨的重要调节剂,修饰软骨-骨边界区的 FABP5+ 细胞以及同一区域的 OC(图 3d、e )。同样,MMP13 和 MMP14 也集中在含有 FABP5+ SCs 的软骨-骨边界区。低抗 MMP13 信号在产后干骺端也标记其它细胞 (图3e)。以往的研究表明,编码基质金属蛋白酶的基因失活,即 (Mmp9、 Mmp13、Mmp14)和编码金属肽酶的基因失活 (Adam19),会导致肥大软骨细胞的扩张和软骨内骨化受损 。与 MMP9 和 MMP13 在软骨胶原蛋白和蛋白聚糖降解中的已知作用一致,MMP9 免疫信号修饰肥大软骨细胞的表面(图 3f),其将经历细胞凋亡以促进骨生长。在软骨-骨界面,分析 AcanCre 标记的表达 GFP 的软骨细胞,结果 显示 在FABP5+ 和 MMP9+ SCs 内存在 GFP+ 碎片,但在同一区域的 CD68+ 破骨细胞内不存在 (图 3g )。此外,通过免疫染色,溶酶体相关膜蛋白 1 (LAMP1)在FABP5+ SCs 中显示较强,它是溶酶体的主要成分,负责介导生物分子的分解。相比之下,软骨-骨边界处的破骨细胞显示出相对较少的 LAMP1+ 信号 (图 3h)。电子显微镜结合对 LAMP1和 vATPase 的免疫金单和双标记(见“方法”)。SCs 对 LAMP1 染色呈阳性,但对 vATPase 呈阴性,而在破骨细胞中恰好相反,对 LAMP1 染色呈阴性,但对 vATPase 呈阳性(图 3i )。此外,当隔细胞( SCs)和软骨细胞(CHO)在体外培养时,SCs 在 SC-CHO 边界区显示出强烈的 MMP9 表达。对来自这些共培养物的分选细胞的分析表明,Fabp5、Mmp9和Mmp14转录物主要存在于 SCs 中,而 Acan+ 富集在预期的 CHO 中,并且Mmp13表达不限于一种细胞类型。SCs 表现出强烈的细胞形状变化,延伸出许多突起,显示出更强的肌动蛋白细胞骨架染色,并且能够在一定程度上从预涂层组织培养表面去除磷酸钙 。进一步分析认为 SCs 分泌 MMPs 从而降解软骨基质和吞噬肥大软骨细胞, 当肥大软骨细胞在骨化期间降解时,高丰度的 FABP5+ 和 MMP9+ SCs存在,反映了积极的生长板重塑 。相反,成年长骨中的 SCs 的 MMP9 表达降低,并且在生长板已转化为骨骺线的衰老动物中不存在免疫信号(图 3j )。上述数据共同表明 FABP5+ SCs 不同于经典破骨细胞, 通过提供分泌的蛋白酶和吞噬作用降解骨细胞碎片参与软骨退化(图3k)。

4 内皮细胞(ECs) - 隔细胞(SCs)相互作用
(Endothelial–septoclast interactions)
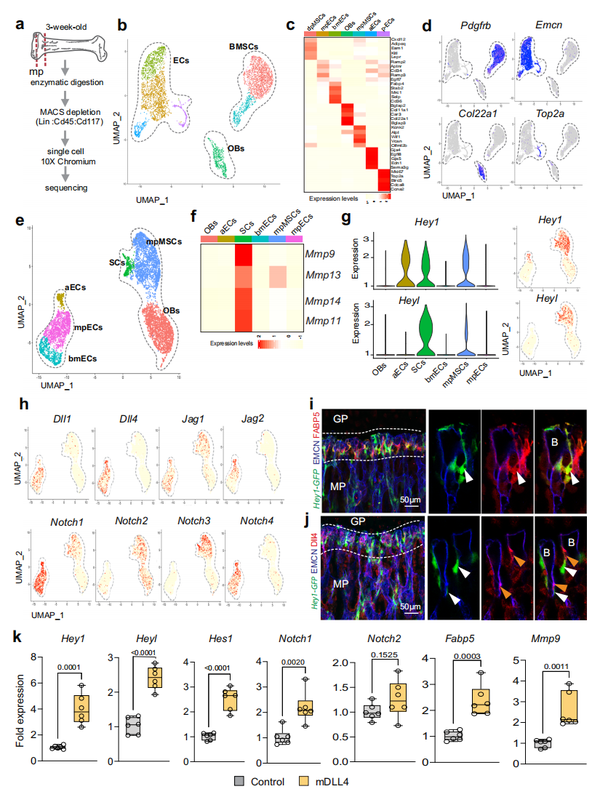
为了深入了解 SCs 和周围细胞之间的相互作用,作者将3 周大的野生型长骨干骺端区域分离出来,并去除造血细胞系和祖细胞后进行 scRNA-seq(参见“方法”)(图. 4a)。作者对 4386 个干骺端细胞进行了测序,这些细胞根据基因表达分群为ECs、BMSCs和OBs (图 4b-d)。接下来,作者将这个数据集和 Pdgfra-GFP scRNA-seq 结果(参见“方法”)合并,然后从合并的数据集中提取 SC、mpMSC、OB 和 EC 细胞群体以进行进一步分析(图 4e )。簇特异性标记基因表达确定了 3 个不同的 EC 子簇,即动脉 ECs (aECs)、干骺端 ECs (mpECs) 类似于之前发布的 CD31high EMCNhigh(类型 H) 内皮亚群和骨和正弦脉管系统的骨髓 ECs (bmECs) ,是在BM腔除了 Fabp5+ SCs、mpMSCs 和 OBs之外的细胞。确认了之前的结果,相对于其它细胞群体,基质金属蛋白酶 Mmp9、Mmp13、Mmp14 和 Mmp11 高度表达于 Fabp5+ SCs细胞群(图 4f)。作者还观察到 Notch 通路基因 Hey1和Heyl在SCs中高度表达(图4g)。Notch 配体Dll1、Dll4、Jag1、Jag2的转录本主要在 ECs 中表达,而 Jag1 也存在于 mpMSCs 中(图 4h)。与Notch信号在脉管系统中的重要作用一致,受体Notch1和Notch4的转录产物在EC中表达丰富,而Notch3以及Notch1和Notch2的转录产物在SCs表达(图4h)。此外,骨切片染色表明Hey1-GFPNotch 报告基因的表达除了 mpMSCs中,还有 SCs (图 4i )。Hey1-GFP+ FABP5+ SCs 靠近内皮芽,生长在 CD31high EMCNhigh 血管的最远端突起, 直接靠近生长板,芽表达EC处免疫染色显示有大量 Dll4 (图 4j )。进一步讨论 Notch 信号的作用, FACS 分离得到Hey1-GFP+ 干骺端细胞体外固定化重组DLL4蛋白,使得 Notch 靶基因表达显著增加,Fabp5和Mmp9转录物的表达也显著上升(图 4k)。在体内,成年小鼠的 ECs Dll4+ 芽和 Hey1-GFP+ SCs 均减少并在衰老过程中消失。作者之前的工作表明,EC 中Dll4的失活会损害骨血管生成和成骨 (图 5a)。表明 Dll4+ ECs 和Hey1-GFP+ SCs 之间的直接互作,在EC特异性的Dll4突变体中,GFP 报告信号丢失并且 FABP5+ 细胞显著减少(图 5b-d )。FABP5+SC 数量的减少伴随着Dll4突变体中 MMP9 的表达显著降低, 而在干骺端肥大的软骨细胞和未被吸收的软骨细胞碎片增加(图5e)。相反,在Dll4突变体骨样品中CD68+ 破骨细胞没有丢失。这些发现表明,SCs被表达 Dll4-的ECs 通过Notch信号通路控制(图5g)。

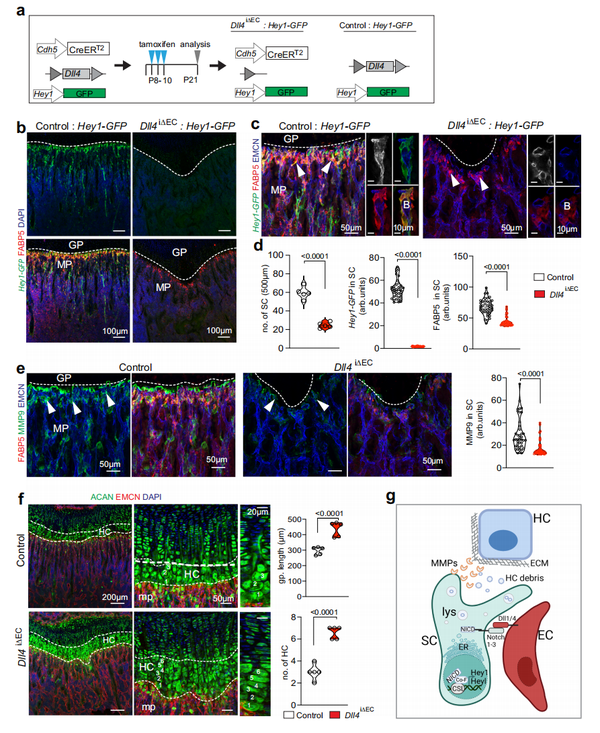

5 骨折愈合中的隔细胞
(Septoclasts in fracture healing)
骨折长骨的修复是一个多步骤的过程,包括血肿形成、血管生成、软骨细胞和富含基质的软骨痂形成,并逐渐转化为新骨。利用scRNA-seq 对骨折后第 14 天 (PFD14) 和年龄相配未断裂的对照骨修复阶段非造血基质细胞进行分析 ,共产生了 45,321 个细胞的数据 (图 6a )。簇特异性标记基因用于表征不同的细胞类型并定义基质异质性和骨折修复期间变化的图(图 6b-e)。在 PFD14 时,骨折骨中的软骨细胞和成纤维细胞明显增加,而 dpMSCs 相对于其它群体减少。在 PFD14 骨骼和对照样本中,EC 数量相当,而在骨折的骨基质中增殖细胞的数量显著增加(图 6d,e )。与成年和老年小鼠中 SCs 的下降一致, 对照中mpMSC 簇的Fabp5+Mmp9+ 细胞稀少 ,但在骨折修复期间增加(图6f,g)。这通过 mpMSCs 的亚聚类分析得到证实,该分析还显示在PFD14 样本中Fabp5和Mmp9在 SCs 中的表达上调。(图 6h、i )。相比于对照,在骨折骨中Mmp13 表达显著增加并扩展到不同细胞群。在PFD14中, scRNA-seq 数据中细胞的轨迹分析与 dpMSCs 转化为 mpMSCs 一致,进而产生成骨细胞谱系细胞、成纤维细胞、软骨细胞和 SC (图 7a、b )。与 scRNA-seq 数据一致,PFD14 骨的免疫染色证实了愈伤组织中软骨细胞和 PDGFRβ+ 间充质细胞的广泛积累以及活跃的血管生长(图 7c、d )。PFD14 样本和3周龄股骨干骺端共有的共同特征包括 CD31high EMCNhigh 血管的芽状形态侵入愈伤组织的无血管、富含软骨细胞的区域,远端血管与 PDGFRβ+ 间充质细胞的关联,以及生长的脉管系统周围的 Osterix + 成骨细胞的丰度(图 7e )。与 scRNA-seq 结果中看到的增加一致,SCs 大量用于骨折愈合并与愈伤组织中软骨细胞附近的血管芽相联系(图7f-h)。在骨折修复骨切片中的同一区域也显示出 MMP9 和 MMP13 的高表达(图 7h、i )。虽然 CD68+ 破骨细胞出现在 SCs 和软骨细胞附近,但这些细胞在整个新的形成的骨,不集中在软骨血管处 ,并显示出相对较弱的 MMP9 免疫染色 (图 7j)。此外,转录因子Runx2和血管生成生长因子VEGF-A的表达在BMSCs和愈伤组织的软骨细胞中强烈增加。这些数据支持再生骨,主要包括发育成骨,包括 FABP5+ SCs 的重新出现、血管生成和血管相关 Osterix+ 细胞的高丰度存在。


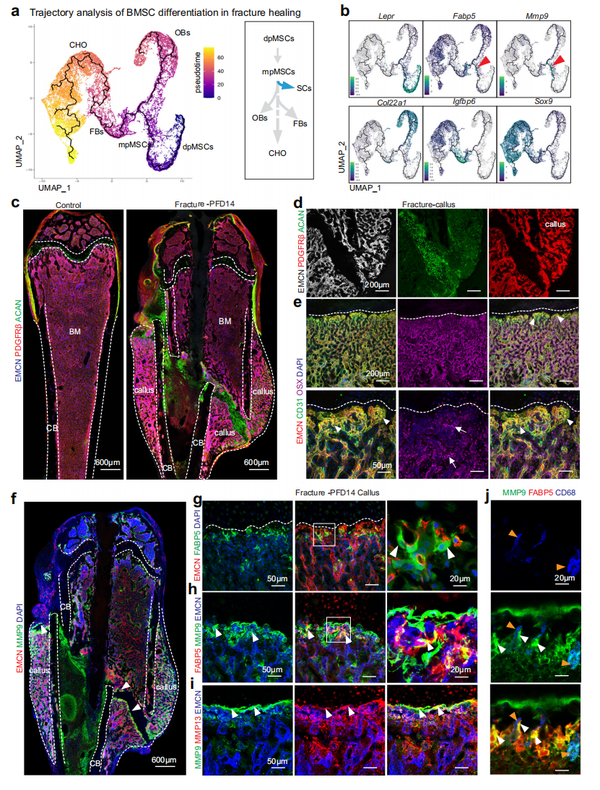

小结
由于外伤或者骨质疏松症导致的骨折,已经影响到很多人的正常生活了。骨折愈伤组织中软骨的退化对于骨化至关重要,但在再生和发育成骨过程中软骨吸收细胞的性质,一直不清楚。本文提出软骨吸收是由表达 FABP5+的单核隔细胞介导的,隔细胞是发育和再生软骨内成骨的一个标志细胞。该研究还提供了骨折修复过程中非造血细胞的图谱,可以深入了解关键细胞群和相互转化过程的分子图谱。该研究结果将有助于理解骨骼的再生过程,也有助于制定改善骨折愈合的治疗策略。
参考文献
Sivaraj,K.K. Majev,P.G.& Jeong,H.W. Mesenchymal stromal cell-derived septoclasts resorb cartilage during developmental ossification and fracture healing[J].Nat Commun. 2022; 13: 571.
附:骨基质细胞的制备
为了富集 septoclasts,作者取3周大的 Pdgfra-H2B-GFP 杂合雄性小鼠的股骨和胫骨,清除附着的周围组织,解剖干骺端区域并收集在消化酶溶液(I型和IV型胶原酶,2mg/ml)中。接下来,骨头被切成小块,用研钵和杵压碎。样品在 37 °C 下消化 20 分钟。将消化后的样品转移到带有70 μm 过滤器的50ml管中以获得单细胞悬液,将其重悬于封闭溶液(1% BSA,1 mM EDTA 的 PBS 溶液,不含 Ca2+/Mg2+)中,300 × g 离心 5 分钟,用冰上预冷的封闭溶液洗涤 2-3 次,并通过 50 μm 过滤器过滤。最终将得到的细胞用封闭溶液分别重悬起来。对于非造血骨基质细胞的 scRNA-seq,对 3 周龄 C57BL/6J 雄性小鼠干骺端节段进行空间解剖,利用如上方法制备单细胞悬液。对于骨折骨的scRNA-seq 实验,取 PFD14 和对照两个样本的股骨,清除周围组织,并从骨折样本中取出钉插入物。利用如上方法,制备对照和 PFD14 两个样本的骨单细胞悬浮液和非造血基质细胞,用于后续ScRNA-Seq实验。
关于北京易研科技有限公司
易研科技专注于为生命科学研究、基础医学研究等领域提供全球最先进的产品及科技服务。
易研科技是10x Genomics的官方授权代理商,可为客户提供Chromium单细胞平台、Visium空间平台以及Xenium原位检测平台相关产品的技术讲解、仪器试剂销售及使用培训,以及相关测序数据的生物信息分析培训。
同时易研科技也提供动物解剖、组织解离、流式细胞分析与分选、质谱流式分析、蛋白多因子检测(Olink平台、Isoplexis平台、luminex平台、Ella平台)、RNA原位杂交(RNAscope平台)、多色荧光免疫组化实验、共聚焦显微成像、组织样本病理检测等全流程实验服务。
更多资讯,欢迎关注“易研科技”公众号
了解更多关于易研科技,欢迎通过以下方式联系我们:
电 话:010-63051149
网 址:http://www.easyresearch.com.cn
邮 箱:service@easyresearch.com.cn
联系地址:北京市丰台区海鹰路8号院3号楼B座702室
