
Nature Metabolism|脂肪生成的关键调节因素

本期我们分享的是2023年5月4日发表在 Nature Metabolism 杂志上的综述文章,文章总结了脂肪生成的组织特异性生理和病理影响以及控制生脂途径的调节机制。


摘要
脂类是生物体的基本代谢物。在产生热量的分子中,脂类具有最高的能量密度,这为其能量的储存和消耗提供了巨大的优势。此外,由于其疏水特性,脂类有助于划定细胞边界,并作为质膜的关键成分发挥作用。此外,最近的研究发现,一些脂质代谢物作为信号媒介调节免疫和代谢的平衡(方框1)。脂质由各种类型的分子组成,如脂肪酸、三酰甘油(TAGs)、胆固醇、磷脂和鞘脂。由于脂质代谢物的不同作用,细胞已经进化到可以生物合成不同的脂质种类。脂肪生成是一个将碳水化合物和其他底物转化为脂肪酸的过程,然后作为TAGs储存起来。参与脂肪生成的关键酶在激素和营养状态下受到转录和翻译后修饰的精确调控。
BOX1
脂质信号媒介
脂质代谢物作为信号分子在能量和免疫稳态中发挥着关键作用。这些脂质介质参与许多代谢和免疫过程,它们的失调与一些疾病有关,如代谢并发症、炎症性疾病、动脉硬化和癌症。脂质信号介质主要分为羟基脂肪酸的支链脂肪酸酯 (FAHFAs)、特异性促炎症消退介质(specialized pro-resolving mediators,SPMs)、氧化脂质和前列腺素,以及鞘脂类。FAHFAs和SPMs发挥抗炎和抗糖尿病的作用。据报道,给5-PAHSA和9-PAHSA(可能是通过反酰基反应催化的),通过增强胰岛素信号,抑制脂肪酸释放和通过G蛋白偶联受体(GPR)40和GPR120产生的肝脏葡萄糖,改善葡萄糖不耐受。此外,FAHFAs通过调节免疫细胞抑制炎症。SPMs来自于多羟基的PUFAs,并由脂氧素(Lipoxins)、消退素(Resolvins)、保护素(Protectins)和 Maresins组成。由于巨噬细胞吞噬死亡细胞和病原体以解决炎症,脂氧素通过PI3K-AKT和细胞外信号调节激酶(ERK)-核因子红细胞2相关因子2(NRF2)途径增加巨噬细胞的寿命,从而促进炎症的解决。Resolvins、protectins和maresins来源于二十碳五烯酸和二十二碳六烯酸,可调节单核细胞、巨噬细胞和中性粒细胞的迁移和激活。值得注意的是,寒冷刺激会促进SPMs的合成和分泌,特别是maresin 2及其异构体,它们部分通过靶向肝脏中的巨噬细胞来改善肥胖症的肝脏炎症。血浆中12,13-二羟基-9-十八碳烯酸(12,13-diHOME)的水平与体重和胰岛素抵抗呈负相关。最近,棕色和米色脂肪细胞已被证明可以产生和分泌12,13-diHOME,并调节葡萄糖和脂质的平衡以及对运动和冷暴露的生热活动。包括神经酰胺和鞘磷脂在内的鞘磷脂调节细胞凋亡、细胞周期控制、细胞衰老和细胞分化的应激反应。神经酰胺水平的改变与代谢性疾病有关,如脂肪组织炎症、非酒精性脂肪性肝炎、肥胖和胰岛素抵抗。
脂肪生成在许多组织中发挥着独特的作用,并以组织特异性的方式进行调节。例如,在脂肪组织中,脂肪生成(Lipogenesis)有助于脂肪细胞生成(adipogenesis)、全身糖脂稳态以及产热。慢性营养过剩时,脂肪生成明显下调,这导致异位脂质积累和胰岛素抵抗。因此,脂肪组织的生脂活性的异常降低被认为是肥胖和胰岛素抵抗之间的病理联系之一。另一方面,肝脏作为脂肪生成的另一个中心组织,也通过脂质的合成、储存以及通过脂蛋白颗粒分配到临近组织来参与脂质平衡。与脂肪组织Lipogenesis不同,肝脏Lipogenesis在肥胖症中是上调的,并导致代谢并发症,如非酒精性脂肪肝(NAFLD)和非酒精性脂肪性肝炎(NASH)。此外,由于脂质是细胞膜的重要组成部分,生脂活性对神经元和免疫细胞特别重要,在细胞增殖和分化过程中,对细胞膜的需求大大增加。因此,失调的生脂作用已被证明参与到代谢并发症免疫调节、神经元活动、衰老和癌症进展中。在此,我们重点讨论Lipogenesis的组织特异性生理和病理影响。此外,我们还讨论了最近在了解控制生脂途径的调节机制方面取得的进展,并讨论了Lipogenesis不足或过量的病理后果。此外,我们还描述了调控Lipogenesis的治疗意义。由于胆固醇生物合成途径和调节过程已被广泛描述,本文不加以赘述。
生脂途径Lipogenic pathways
生脂途径的概述
将碳水化合物转化为脂肪的过程,即脂肪生成(Lipogenesis),是由多种酶以一种复杂和协调的方式介导的。在餐后状态下,葡萄糖和胰岛素水平的提高促进了葡萄糖的摄取和随后的糖酵解,产生丙酮酸,然后被运送到线粒体,作为三羧酸(TCA)循环的底物(图1)。葡萄糖被葡萄糖转运体(GLUT)吸收并立即被葡萄糖激酶(GK)磷酸化。在TCA循环中,柠檬酸盐产生并从线粒体输出,由ATP柠檬酸盐裂解酶(ACLY)和乙酰-CoA羧化酶(ACC)催化为乙酰辅酶A(CoA)和丙二酰-CoA。之后,脂肪酸合成酶(FASN)与丙二酰-CoA进行一系列脱羧的克莱森缩合反应,生成棕榈酸(16:0)。脂肪生成所需的高能电子由NADPH提供,NADPH由磷酸戊糖途径中的葡萄糖-6-磷酸(G6P)脱氢酶(G6PD)和磷酸葡萄糖酸脱氢酶(PGD)、添补反应中(anaplerosis)的苹果酸酶(ME)和丝氨酸分解代谢中的几种酶产生。
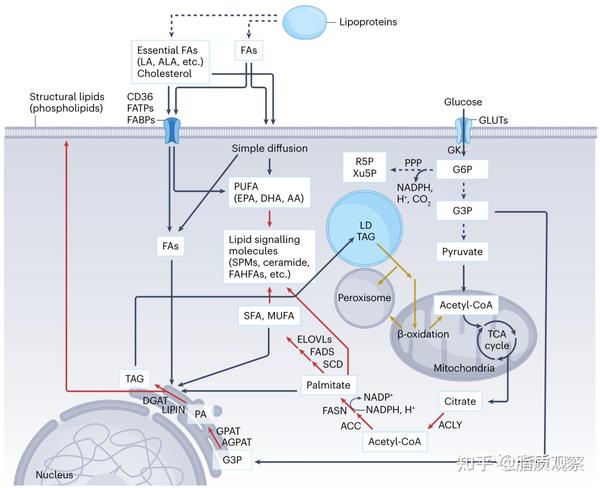

图1 - 脂肪生成概述。脂质代谢的主要途径概述,涉及脂质吸收、脂质生成和通过脂肪酸氧化进行脂质降解。脂肪酸通过线粒体中的糖酵解和TCA循环从碳水化合物转化而来,并通过各种途径吸收,如脂蛋白和脂肪酸结合蛋白的被动扩散和运输。细胞的脂肪酸以TAGs的形式储存在LD中,某些脂肪酸作为信号媒介和结构脂质发挥作用。储存在LD中的脂质通过与线粒体和过氧化物酶体的相互作用被分解以满足能量需求。橘色箭头代表脂类的合成代谢途径,而灰色箭头表示分解代谢途径。AA,花生四烯酸;ACC,ACC1;ACLY,ATP柠檬酸合成酶;AGPAT,1-酰基甘油-3-磷酸-O-酰基转移酶;ALA,α-亚麻酸;CD36,分化群36;DHA,二十二碳六烯酸;EPA,二十碳四烯酸;ELOVLs,极长链脂肪酸蛋白的延伸;FAs,脂肪酸;FABPs,脂肪酸结合蛋白;FAHFA,羟基脂肪酸酯;FATPs,脂肪酸转运蛋白;G3P,3-磷酸甘油酯;LA,亚油酸;MUFA,单不饱和脂肪酸;PA,磷脂酸;PPP,磷酸戊糖途径;R5P,5-磷酸核糖;SPM,专门的促解介质。
棕榈酸酯是其他脂肪酸的关键前体。通过脂肪酸延长酶(ELOVLs)和去饱和酶(硬脂酰-CoA去饱和酶(SCD)和脂肪酸去饱和酶(FADS)),棕榈酸被延长和/或转化为饱和脂肪酸(SFA)和单不饱和脂肪酸。棕榈酸及其衍生物,如硬脂酸(18:0)和油酸(18:1n-9)是啮齿动物和人类脂肪组织、肝脏和血清中的主要脂肪酸种类(约占酯化和游离脂肪酸种类的80%)。多不饱和脂肪酸(PUFAs),如亚麻酸(18:2n-6)和α-亚麻酸(18:3n-3),在后生动物中不能从头合成,由食物提供,并进一步转变为花生四烯酸(20:4n-6)、二十碳五烯酸(20:5n-3)和二十二碳六烯酸(22:6n-3)。虽然PUFAs在脂肪酸种类中占相对较小的比例,但有人认为这些脂质种类和衍生物作为信号媒介和构件发挥着关键作用。
由于过量的脂肪酸可诱发细胞毒性(脂毒性),大多数细胞以TAG的形式在脂滴(LD)中储存脂肪酸。TAG的合成始于甘油-3-磷酸酰化酶(GPAT)对甘油-3-磷酸的连续酰化,生成溶血磷脂酸,进一步酯化并转化为磷脂酸。随后,磷脂酸通过磷脂酸磷酸酶lipin1(LPIN1)和二酰基甘油酰基转移酶(DGAT)被重新定向到不同磷脂的合成或TAG。由于LD不仅储存中性脂质,而且通过与过氧化物酶体和线粒体的相互作用提供能量来源,LD似乎整合了细胞的合成代谢和分解代谢(方框2)。
BOX2
脂滴
脂滴是细胞器,在调控脂质代谢方面发挥着关键作用。脂滴储存中性脂质,如TAGs和胆固醇酯,用于代谢能量和组装原料。LDs由疏水性的中性脂质核心组成,被磷脂单层所包围;它们还含有合成和分解中性脂质的酶。LDs从ER膜产生,出芽,随后通过与负责TAG合成的酶如GPAT和DGAT结合而生长。LD周围的Perilipins (PLIN1-PLIN5)是维持脂滴结构和脂质平衡所必需的。PLIN1主要在脂肪细胞中表达,调节禁食诱导的脂肪分解,抑制脂肪流失及脂肪组织中的促炎症反应。PLIN3在许多组织中普遍表达,参与LD的生物生成和脂肪分解,为脂肪细胞的产热提供内源性PPAR-α激活剂和/或配体。PLIN5在氧化性细胞和组织中高度表达,如心脏、肝脏、肌肉和BAT,在那里它调节脂肪分解以获得能量供应。LDs选择性地、动态地与其他细胞器如过氧化物酶体、线粒体、ER和内体相互作用,以维持细胞的能量平衡。例如,当能量需求增加时,分解代谢过程随着脂肪分解过程的启动而开始:在空腹时,过氧化物酶体以驱动蛋白家族成员C3(KIFC3)依赖的方式迅速与LDs结合,过氧化物酶体蛋白PEX5通过刺激ATGL转位到LDs上,介导空腹诱导的脂肪分解。此外,LDs与线粒体有物理接触,在此基础上,围脂滴线粒体的丙酮酸氧化、电子运输和ATP合成能力增加。从全基因组筛选、成像工具和LD蛋白质组学中积累的证据,发现了决定LD形态和功能的基因,正在迅速扩大我们对LD生物学的理解。此外,最近推出的脂滴知识门户(https://lipiddroplet.org)为这些数据的使用提供了便利。对LD生物学的全面研究将为脂质的合成、储存和利用,以及治疗方法的开发提供机会。
调节脂肪生成的关键因素
脂肪生成的积极因素
Lipogenesis受到营养物质和代谢激素(如胰岛素、胰高血糖素和糖皮质激素)的严格和复杂的调节。在餐后状态下,血清葡萄糖和胰岛素的增加通过信号级联激活脂肪生成的转录因子,如甾醇调节元件结合蛋白(SREBP)1c、碳水化合物反应元件结合蛋白(ChREBP)、肝X受体(LXR)-α和上游转录因子(USF)(图2)。遗传动物模型研究已经确定了它们在脂肪生成中的关键作用(表1)。除转录因子外,由AKT-哺乳动物雷帕霉素(mTOR)复合体(mTORC)2介导的胰岛素信号级联通过Ser455的磷酸化增加ACLY的活性。
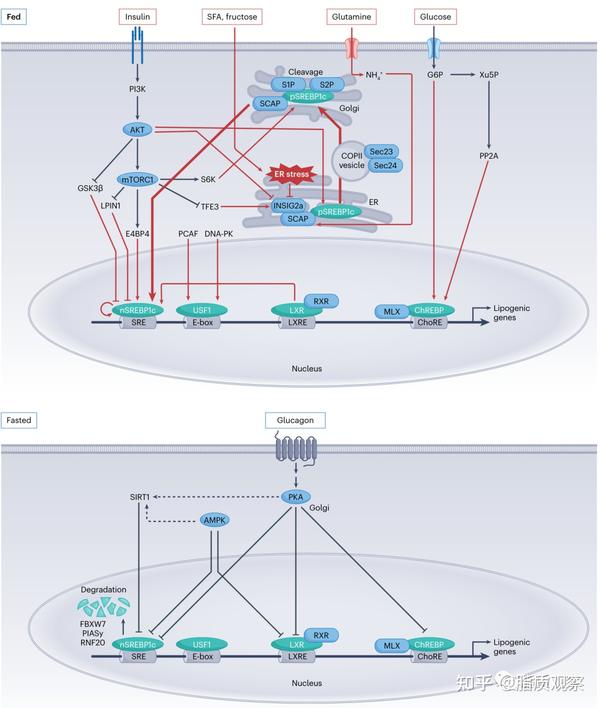

图2 - 激素和营养刺激后脂肪生成的转录和翻译后调节的信号途径。在激素和营养刺激下,大多数脂肪生成的关键转录因子通过转录和翻译后修饰受到严格的调节。合成代谢状态激活脂肪生成的转录因子,如SREBP1c、ChREBP、LXR-α和USF。胰岛素,一种关键的合成代谢激素,刺激生脂转录因子的表达和活性。胰岛素刺激的PI3K-AKT途径和mTOR途径激活并调节SREBP1c。不仅葡萄糖激活ChREBP,其他营养物质如SFA、果糖和谷氨酰胺也调节SREBP1c的活性,导致Lipogenesis的增加。在空腹状态下,胰高血糖素,一种关键的分解代谢激素,通过PKA依赖的转录和翻译后调节,抑制了大多数生脂转录因子。红色箭头表示脂肪生成的刺激作用,蓝色箭头表示抑制作用。ChoRE,碳水化合物反应元件;DNA-PK,DNA依赖性蛋白激酶;E4BP4,E4启动子结合蛋白4;FBXW7,含F-box和WD重复域7;GSK3β,糖原合成酶激酶3β;LXRE,LXR反应元件;MLX,Max-like protein X;PP2A,蛋白磷酸酶2A;RXR,视黄醇X受体;nSREBP1c,核SREBP1c;pSREBP1c,前体SREBP1c;SRE,甾醇调节元件。
除了葡萄糖这一Lipogenesis的主要碳源外,据报道,其他营养物质,如柠檬酸盐、果糖、氨基酸和脂肪酸也积极参与脂肪生成。这些营养物质通过参与酶和转录调节器的加工以及作为脂质合成的底物来调节脂肪生成的转录因子。例如,ACACA(ACC异构体1,ACC1)在柠檬酸盐的存在下被聚合,柠檬酸盐增加其酶活性,而ACACA被丙二酰-CoA和棕榈酰-CoA抑制,形成一个负反馈环。另外,ACLY被柠檬酸异生激活,而其产物乙酰CoA和草酰乙酸则抑制酶的活性。在高葡萄糖水平下,ACLY被乙酰化转移酶PCAF乙酰化,并通过阻止其泛素化和降解而得到稳定。此外,某些营养物质如PUFAs会抑制SREBP1c和ChREBP的活性。
脂肪生成的负面因素
在能量匮乏的状态下,如禁食,非必要的Lipogenesis需要被抑制。胰高血糖素是一种关键的激素,它通过环磷酸腺苷和蛋白激酶A(PKA)信号途径调节对禁食的系统反应。例如,激活的PKA信号促进转录和翻译后的调节,以下调SREBP1c。此外,PKA直接磷酸化ChREBP以抑制其核输入和转录活性。同时,PKA通过磷酸化抑制ACACA的酶活性。AMP激活的蛋白激酶(AMPK)是能量匮乏状态下的另一个重要调节器,它通过在Ser80处磷酸化人类ACACA来抑制ACACA的聚合和激活,从而抑制Lipogenesis。
Lipogenesis中复杂的激素激活剂
糖皮质激素对脂肪生成的贡献是复杂的,取决于激素状态。在没有胰岛素或空腹的情况下,糖皮质激素会抑制葡萄糖的摄取和Lipogenesis。相反,在有胰岛素的情况下,糖皮质激素通过抑制AMPK的活性增强Lipogenesis,从而诱导脂肪生成的基因表达。由于11β-羟类固醇脱氢酶1是一种将无激素活性的可的松(cortisone)转化为氢化可的松(cortisol)的酶,与人类的皮下脂肪组织相比,它在内脏脂肪组织中高度表达,库欣综合征中高水平的糖皮质激素有可能导致内脏肥胖。因此,几种激素通过对关键的生脂转录因子的精密调控,调节脂质的平衡。
脂肪生成的转录调节器
SREBPs
SREBP1c的发现。 在过去的几十年里,关键的 lipogenic 转录因子的发现和对其在脂质代谢中作用的阐明,使人们能够深入了解全身能量平衡。1993年,SREBP1c(又称脂肪细胞决定和分化因子1,ADD1)被两个独立的研究小组用不同的方法克隆出来,他们接下来的工作成为破译脂肪生成的分子控制的基础。由于大多数basic螺旋环螺旋(bHLH)转录因子与E-box(CANNTG)结合,Spiegelman小组用E-box作为探针克隆了adipogenic bHLH转录因子,并确定了ADD1,该因子在脂肪细胞分化过程中被上调。ADD1在脂肪代谢活跃的脂肪组织和肝脏中富集,特别是最近的单细胞和单核RNA测序研究表明,ADD1在脂肪细胞和肝细胞中高度表达,这表明ADD1将对脂肪生成很重要。同年,Brown, Goldstein和他们的同事确定了几个转录因子与甾醇调节元件(CACCCCAC)结合,这是胆固醇代谢的一个关键顺式元件,并将这些因子命名为SREBP1和SREBP2。SREBP1有两个异构体,SREBP1a和SREBP1c,具有不同的N端结构域和不同的启动子用途,而后者与ADD1相同。与其他bHLH转录因子不同的是,已经发现ADD1(SREBP1c)具有独特的双链DNA结合特异性,通过ADD1(SREBP1c)基本结构域(EKRY)中的一个非典型酪氨酸残基与甾醇调节元件和E-box结合(以下称ADD1(SREBP1c)为SREBP1c)。
SREBPs是定位在内质网(ER)膜上转录因子,经过翻译后的处理,进入细胞核以进行目的基因表达。简而言之,SREBPs的前体形式在ER膜上由SREBP裂解激活蛋白(SCAP)负责向高尔基复合体的转运,而SCAP被胰岛素诱导基因(INSIGs)抑制。在高尔基复合体中,SREBPs被两种蛋白裂解酶,即site 1 和site 2蛋白酶(S1P和S2P)。蛋白裂解后,成熟形式的SREBP转入细胞核并激活其靶基因表达。
SREBP1c及其同种异形体SREBP2的调节
迄今为止,不断积累的证据清楚地表明,SREBP1c对大多数lipogenic基因的表达至关重要,而SREBP2对胆固醇的稳态很重要。另外,SREBP异构体的基因表达和激活的调节机制似乎是不同的。例如,SREBP1c受胰岛素和胰高血糖素的严格调节,而SREBP2则受甾醇水平的调节。合成代谢条件和胰岛素刺激SREBP1c的表达,它也被LXR、SP-1、核转录因子Y(NF-Y)和SREBP1c本身与SREBP1c启动子的结合所调节。此外,一些媒介如AKT、mTORC1及其下游蛋白p70S6K、LPIN1和mTORC2促进SREBP1c蛋白加工、核定位和蛋白稳定性。此外,据报道,SREBP1c的加工也受一些营养物质的调节,如葡萄糖、谷氨酸衍生的氨、果糖和一些SFA和PUFAs。尽管作用不同,积累的证据表明,SREBP1c的耗竭在一定程度上被SREBP2所补偿。因此,这些类型的调控机制很可能对反映生物体的能量状态至关重要,并且很可能是在长期能量丰富的状态下储存额外的能量而进化出来的。在合成代谢中,SREBP1c刺激lipogenic基因的表达,包括GK、ACLY、ACACA、FASN、ELOVL6和SCD。此外,SREBP1c通过增强GPAM、LPIN1和DGAT1的表达而增强TAG的合成。此外,SREBP1c通过调节ME1、G6PD和PGD来提高NADPH池,这是脂肪生成的关键还原代谢物。在脂肪组织中,SREBP1c的表达在脂肪细胞分化过程中增强,以促进lipogenic基因的表达,并产生过氧化物酶体增殖剂激活受体(PPAR)-γ的内源配体。除了Lipogenesis外,SREBP1c还与生物现象的各个方面有关。例如,SREBP1c调节胰岛β细胞和癌细胞的细胞增殖,以及肝脏葡萄糖的产生。最近,有研究表明,SREBP1c在肥胖症的脂肪细胞衰老中起着关键作用。当DNA损伤在脂肪细胞中积累时,值得注意的是SREBP1c作为辅助因子与聚(ADP-核糖)聚合酶1(PARP1)结合以介导DNA修复,而不是作为转录因子直接与DNA结合。这是一个重要的发现,表明转录因子不仅会直接与DNA结合以调节其目标基因的表达,而且还会通过与其他蛋白的相互作用发挥 "辅助因子 "的作用。
禁食时,SREBP1c被抑制,以减少不必要的脂肪酸合成。胰高血糖素及其下游的PKA信号会抑制SREBP1c的表达和活性。尽管SREBP1c的核内形式很不稳定并且会迅速降解,但对SREBP1c降解的基本机制还不甚了解。在这方面,已经证明E3泛素连接酶RNF20和激活STAT Y的蛋白抑制剂(PIASy)依赖的小泛素样修饰物(SUMO)化导致SREBP1c的降解。PKA诱导SREBP1c的Ser308磷酸化,这增强了PIASy介导的SUMO化,从而导致SREBP1c的Lys98泛素化和蛋白体降解。此外,一些蛋白质修饰抑制SREBP1c的活性。Sirtuin 1(SIRT1)使SREBP1c去乙酰化,从而抑制SREBP1c在禁食期间与生脂基因启动子的结合,而AMPK直接磷酸化并抑制SREBP1c的转录活性。此外,最近的一项研究表明,禁食诱导的类粘蛋白2(orosomucoid 2)能增强AMPK的活性,抑制SREBP1c的表达和Lipogenesis。这些发现表明,由各种激素和营养刺激介导的信号级联确保SREBP1c的激活,只有在能量丰富的状态下才能有效地储存能量。总之,过去30年的大量研究表明,SREBP1c在Lipogenesis中起着至关重要的作用,并且最近的研究将继续挖掘SREBP1c的新作用和新调节机制。尽管如此,30年的SREBP1c研究很可能还是不足以了解所有的SREBP1c生物学,因此,需要进一步研究SREBP1c的表达、蛋白裂解和激活的调节机制,以及它与SREBP2的关系。
ChREBP
ChREBP在对葡萄糖的反应中能增强Lipogenesis。ChREBP与Max-like蛋白X形成一个异二聚体复合物,后者与碳水化合物反应元件结合。除SREBP1c外,ChREBP还调节编码参与Lipogenesis和糖酵解的基因的表达。ChREBP有两个使用替代第一外显子的异构体:ChREBP-α在脂肪组织和肝脏中均有表达,而ChREBP-β则在脂肪组织中高度表达。ChREBPβ(Mlxipl转录本2,NM_001359237.1)转录本的表达受代谢刺激的调节,如进食、葡萄糖和肥胖,而ChREBPα(Mlxipl转录本1,NM_021455.5)受这些代谢刺激的调节程度不大。在高葡萄糖条件下,5-磷酸木糖(Xu5P)通过Xu5P激活的蛋白磷酸酶导致ChREBP的去磷酸化。此外,G6P通过ChREBP的激活刺激Lipogenesis。从机制上讲,有人认为G6P可能与ChREBP结合,促进ChREBP的异生构象变化,并促进其转位到细胞核。在高葡萄糖水平下,ChREBP也被O-连接-N-乙酰氨基葡萄糖化(O-GlcNAcylated),以稳定ChREBP。此外,p300对ChREBP的乙酰化刺激其被招募到其目标基因的启动子区域。相反,在禁食期间,ChREBP被PKA和AMPK磷酸化,从而阻止ChREBP进入细胞核并减少其与启动子的结合。
SREBP1c和ChREBP的互补关系
有人提出SREBP1c和ChREBP是互补的,前者是Lipogenesis所必需的,并被胰岛素激活,后者是糖酵解和Lipogenesis所必需的,并在碳水化合物的存在下受到刺激,两者是相互补充的,而不是一方占优。通过Insig1的过量表达消耗成熟的SREBP1c会降低ChREBP(Mlxipl)的mRNA水平。另外,SREBP1c诱导GK基因表达,为ChREBP的激活提供G6P。相反,在ChREBP缺失的细胞中,SREBP1c的mRNA和蛋白水平都下降。此外,SREBP1c和ChREBP的重叠但不同的功能使细胞在有胰岛素和碳水化合物的情况下主要合成脂质。在ChREBP缺失后,SREBP1c的过量表达使生脂基因的表达水平正常化,而它不能恢复糖酵解基因的程序。此外,在由Scap敲除影响的SREBP抑制模型中,ChREBP恢复了糖酵解基因的表达,但不能完全挽救lipogenic基因的表达。因此,SREBP1c和ChREBP可能是协调诱导糖酵解基因和lipogenic基因表达以实现适当的lipogenic激活所必需的。
LXR-α
LXR-α是一种被氧化甾醇和胰岛素激活的核激素受体,它参与生脂基因的表达。LXR-α与视黄醇X受体形成异源二聚体,并在有抗氧化剂的情况下与其目标基因启动子中的LXR反应元件结合。LXR-α似乎通过与目标启动子结合直接刺激生脂基因的表达,并通过增加SREBP1c的转录间接地刺激生脂基因的表达。
在进食和高葡萄糖水平下,LXR-α被O-GlcNAcylated,增强其转录活性。同时,在禁食期间,LXR-α被PKA磷酸化,以防止与视黄醇X受体的二聚体化,并招募核共压物1,最终导致LXR-α活性被抑制。此外,PKA磷酸化的LXR-α也会导致LXR-α介导的SREBP1c依赖的脂肪生成被抑制。
USF1
USF1有一个bHLH-亮氨酸拉链结构域,形成同源二聚体或异源二聚体,并与E-box结合,以便在胰岛素的作用下激活生脂基因。USF1是胰岛素激活Fasn启动子的必要条件。此外,有人认为USF1与SREBP1c协同作用于Fasn和Gpam启动子。
由于USF1的表达水平及其与E-box的结合不受禁食和喂食的影响,因此USF1对营养状态的翻译后修饰似乎对生脂基因的表达很重要。进食时,与Fasn启动子结合的USF1被DNA依赖性蛋白激酶磷酸化,并被p300-CBP相关因子乙酰化,从而促进其目标基因的表达。相反,禁食时,USF1被组蛋白去乙酰化酶9去乙酰化,从而阻止USF1的激活。因此,这些研究提供了关于众多转录因子受激素和营养刺激的严格调控以影响脂肪生成和系统脂质平衡的分子基础的见解。