
Elysia cornigera Nuttall, 1989
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4148.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:91353147-FDA8-45CC-A8F1-1DE801C835A6 |
|
DOI |
https://doi.org/10.5281/zenodo.5664209 |
|
persistent identifier |
https://treatment.plazi.org/id/A04A7E6D-9C02-FFD2-46C9-FB1BFCFB1BE8 |
|
treatment provided by |
Plazi |
|
scientific name |
Elysia cornigera Nuttall, 1989 |
| status |
|
Elysia cornigera Nuttall, 1989 View in CoL
( Figs. 6 View FIGURE 6 P, 45–47)
Elysia timida View in CoL [non Risso, 1818] — Ortea, Moro, & Espinosa 1997: 143 –145 (part), fig. 1C; Valdés et al. 2006: 68 –69. Elysia cornigera Nuttall 1989: 302 View in CoL –306, figs. 1–8 (Type locality: Spanish Harbor, West Summerland Key, Florida Keys) — Hess et al. 1994: 161; Clark 1994: 905; Händeler et al. 2009: figs. 6, 7; Krug, Händeler & Vendetti 2011: 481 -484, fig. 1A–D, 2- 3; Redfern 2013: 282, fig. 783A–B; Ortigosa et al. 2013: 65; Christa et al. 2014: figs. 1E, 3; Krug et al. 2015: 990 -991, figs. 3B, 4.
Elysia purchoni View in CoL [non Thompson, 1977] — Ortea et al. 2011: 200 –203, pls. 1–2.
Type material. Elysia cornigera —holotype (USNM 859144) and 5 paratypes (USNM 859145).
Material examined. St. Anne’s Bay, Jamaica, 25 May 2006, 1 specimen ( LACM 173243 View Materials ) ; Bahamas: North Bimini, 21 March 2006, 1 specimen ( CPIC 00127 ), Stocking Island , 15 December 2007, 1 specimen ( CPIC 00015 ), 18 January 2009, 1 specimen ( CPIC 00078 ), 29 January 2009, 2 specimens ( CPIC 00080–81 ), Great Exuma, 29 December 2005, 1 specimen ( LACM 173227 View Materials ), 18 February 2005, 1 specimen ( LACM 172283 View Materials ) .
Additional material examined. Geiger Beach , Florida, USA, August 2007, 1 specimen (isolate Ecor_07Gei01); Discovery Bay, Jamaica, 7 March 2006, 1 specimen (isolate Ecor_06Jam01) ; Little San Salvador, Bahamas, July 2010, 1 specimen (isolate Ecor_10LSS01).
Live animal. Specimens frequently cover their head with anterior parapodial flaps, as if shy. Slugs do not swim, and were not observed to perform the regular head bobbing movement described for E. timida ( Thompson & Jaklin 1988; Wirtz & Anker 2009).
External anatomy. Summarized from Krug et al. (2011): Body small (to 8 mm), color white to grey on parapodia and head, with numerous papillae giving warty appearance ( Fig. 45 View FIGURE 45 A–B). Inside of parapodia densely penetrated by green digestive diverticula, conferring light green tinge to outside of fed specimens ( Fig. 45 View FIGURE 45 C). Foot, sides of head, anterior juncture of parapodia all green ( Fig. 45 View FIGURE 45 D–F). Red granules dot head and rhinophores, with smaller red spots scattered over parapodia ( Fig. 45 View FIGURE 45 D). Rhinophores long, curled; widening medially then tapering to a point; edge of rhinophores uneven, with undulating appearance due to many papillae, hence the etymology behind “horned Elysia ” ( Fig. 45 View FIGURE 45 D–F). Outer surface of rhinophores white with red spots, inner surface yellowgreen with branches of digestive diverticula. Upper edge of oral lobes lined with “moustache” of small black spots ( Fig. 45 View FIGURE 45 B, F).
Parapodia rise into prominent anterior peaks halfway along body, with second posterior set of smaller peaks on large specimens. White papillae in rows along parapodia. Parapodial margin scalloped due to row of white papillae ( Fig. 45 View FIGURE 45 C). Parapodia covered with tiny red sots and larger orange spots. Larger spherical granules of irregular size embedded haphazardly in parapodial surface, ranging from golden-brown to orange, appearing as small craters due to elevated surrounding tissue ( Fig. 45 View FIGURE 45 A–B, D, F).
Pericardium rounded, white with red spots, with one pair of dorsal vessels emerging at posterior end ( Fig. 45 View FIGURE 45 C, Fig. 46 View FIGURE 46 ). Vessels opaque, covered in dense white spots, with lateral branches extending up towards parapodial margin. Short renopericardium followed by a clear furrow running down the midline of most of the remaining body length.
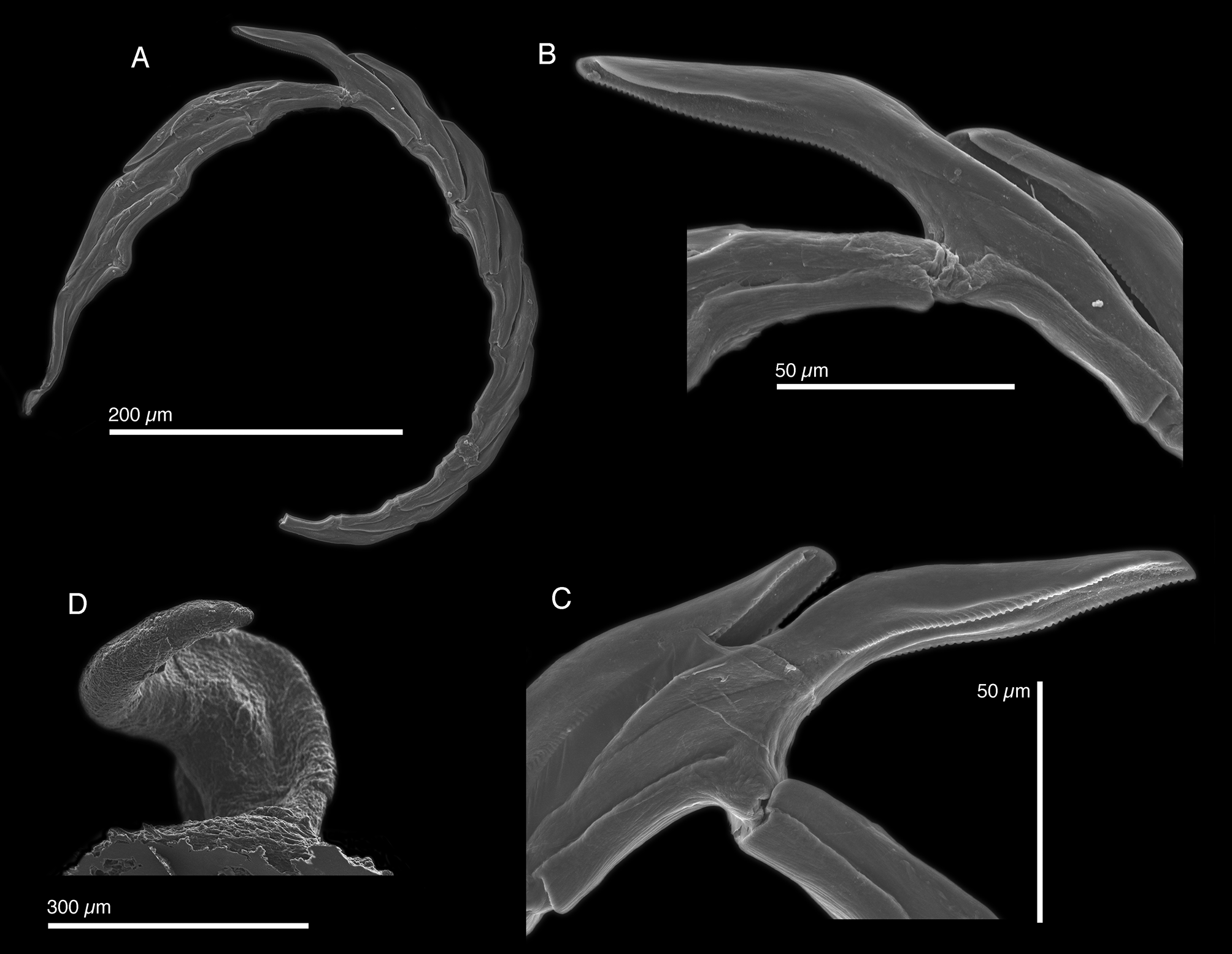
Internal anatomy. Radula with 25 teeth (CPIC 0 0 0 78, CPIC 0 0 0 81, LACM 173243), 4 teeth in ascending limb and 9 teeth in descending limb ( Fig. 47 View FIGURE 47 A). Nuttall (1989) reported 2–5 teeth in the ascending limb and 6–7 teeth in the descending limb. Unlike other Caribbean elysiids, radula of E. cornigera with two cutting edges. Leading tooth elongate with cusp bearing a denticulate keel, one smooth lateral edge ( Fig. 47 View FIGURE 47 B), and one denticulate edge (47C). Nuttall (1989) reported teeth of up to 216 µm in length. Base of teeth thickened and elongate, almost as long as tooth cusp. Ascus (CPIC 00080) containing jumbled heap of discarded teeth (not figured). Ascus with>12 teeth according to Nuttall (1989).
Penis elongate and coiled with with rigid musculature that did not deform after drying, devoid of armature ( Fig. 6 View FIGURE 6 P, 47D). Base of penis wide. Deferent duct short and thin.
Reproduction and development. Development is lecithotrophic. Clutches contain 20–137 ova ~105 µm in diameter ( Nuttall 1989). A flat ribbon of pale yellow or semi-transparent ECY, granular in appearance, lines the upper surface of the egg mass, contacting each capsule ( Fig. 45 View FIGURE 45 G–H). Larvae develop in 15–19 d at 22°C, with most undergoing encapsulated metamorphosis (95.8 ± 7.2%, n = 3 clutches) and emerging as crawl-away juveniles (Krug 2009). Mean larval shell length for two clutches from Florida Keys specimens was 243.6 µm (± 10.0 SD, n = 22 shells) and 253.1 µm (± 6.2, n = 19), small larvae for Caribbean sacoglossans with lecithotrophic development (Krug 2009). The shell aperture has a distinctive extension diagnostic of E. cornigera ( Nuttall 1989) .
Host ecology. The only known host is Acetabularia crenulata . Utricles atop the stipe are individually drained of cytoplasm. Whereas E. timida retains diet-derived chloroplasts for 3–4 weeks, E. cornigera quickly digests chloroplasts, showing a linear decline in photosynthetic activity over 11 d after feeding (Krug et al. 2011).
Phylogenetic relationships. Elysia cornigera was recovered as sister to the Mediterranean species E. timida , with which it was formerly synonymized ( Fig. 4 View FIGURE 4 ). The resurrection of E. cornigera (Krug et al. 2011, 2013) was based in part on the divergence between the two species at COI (11.2% TrN), an inter-specific distance for Elysia spp. No sister group was recovered with support for the clade of ( E. cornigera + E. timida ); the most closely related clade comprised three species, including E. thompsoni , which is similar in appearance and also feeds on Acetabularia , and E. lobata . Within E. cornigera , the Bahamas population was highly genetically differentiated from the Florida population, with the Florida clade of COI haplotypes nesting within a basal grade of Bahamas haplotypes; the lone Jamaica haplotype grouped with Florida (Krug et al. 2011).
Range. Bahamas ( Valdés et al. 2006; Krug et al. 2011; Redfern 2013), Cayman Islands ( Hess et al. 1994; Valdés et al. 2006), Florida, USA ( Nuttall 1989; Krug et al. 2011), Jamaica (present study), Mexico ( Ortigosa et al. 2013).
Remarks. Nuttall (1989) described E. cornigera from specimens collected about 5 km from our Florida collection sites, based on live specimens, radular SEM images, and larval development. Ortea et al. (1997) synonymized E. cornigera with E. timida based on material from Cuba ( Ortea et al. 1997: fig. 1C, pl. 1B), but Ortea et al. (2011) rejected their own earlier synonymy and proposed that Caribbean material previously identified as E. timida could be E. cornigera . Ortea et al. (2011) also suggested the possibility of the existence of two cryptic species under the name E. cornigera . Genetic data in Carmona et al. (2011) indicate that a specimen of E. timida from the Mediterranean was sampled in Florida, whereas most Floridian samples were genetically distinct E. cornigera . It is thus possible that E. timida was introduced to Florida and/or Caribbean populations.
Elysia timida is larger (6–13 mm in body length), with outer parapodia, head and rhinophores colored pure white in contrast to the deep green diverticula visible inside the parapodia. In E. cornigera the head and parapodia are greyer and more papillose, the red spots proportionally smaller, and the distinctive large golden-brown spherical inclusions giving an overall brownish tint. Rhinophores of E. cornigera have an uneven edge, curve backward, and taper to a point, whereas those of E. timida are smooth-edged and terminate in a rounded tip. The rhythmic contraction of rhinophores and head-bobbing of E. timida have not been described from E. cornigera . Developmentally, the ECY of E. cornigera is distinctively semi-transparent, rather than bright yellow as in E. timida , and egg size appears to be smaller in E. cornigera (105 µm) than in E. timida (120 µm).
Ortea et al. (2011) described specimens of E. cornigera as E. purchoni , which we noted previously is a synonym of E. canguzua ; Ortea et al. (2011) further suggest that E. cornigera can be a junior synonym of E. purchoni . There are many similarities shared by E. canguzua (= E. purchoni ) and E. cornigera : both have scattered red spots across the head and body, a “moustache” of black spots on the upper oral lobes, and parapodial margins that widen considerably at the anterior edge. However, the parapodial extensions of E. canguzua (= E. purchoni ) are rounded, and form a pronounced siphonal opening ( Thompson 1977: fig. 25f); in contrast, the anterior parapodial edge of E. cornigera extends into angular, straight-edged side flaps. The shape of the rhinophores and the rounded siphonal opening drawn for E. purchoni are only consistent with E. canguzua , a species that Thompson was evidently unaware of, and not E. cornigera . Although the radular tooth of both species possesses lateral edges on the sides of the cusp, the tooth of E. canguzua (= E. purchoni ) is only finely denticulate (and only evident by SEM), while that of E. cornigera has both a prominently denticulate keel and a serrated lateral edge ( Marcus 1955; Thompson 1977; Nuttall 1989; present study).
Finally, Ortea et al. (2013) state that “Redfern ... illustrates Elysia purchoni and Elysia timida under the name Elysia cornigera Nuttall, 1989 (species 783A and 783B respectively).” We find these remarks incoherent, given that Ortea et al. (2011) proposed E. cornigera may be present in Cuba and was not in fact E. timida , and that E. cornigera could be synonymous with their E. purchoni . In fact, Redfern (2001) correctly identified his material as E. cornigera , as per Nuttall (1989) and Krug et al. (2011).




| LACM |
Natural History Museum of Los Angeles County |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Elysia cornigera Nuttall, 1989
| Krug, Patrick J., Vendetti, Jann E. & Valdés, Ángel 2016 |
Elysia purchoni
| Ortea 2011: 200 |
Elysia timida
| Krug 2015: 990 |
| Redfern 2013: 282 |
| Ortigosa 2013: 65 |
| Krug 2011: 481 |
| Valdes 2006: 68 |
| Ortea 1997: 143 |
| Hess 1994: 161 |
| Clark 1994: 905 |
| Nuttall 1989: 302 |