
Abstract
This manuscript aims to provide a comprehensive review of the work done by Antarctic sclerochronology research across different taxa (arthropods, bivalves, brachiopods, bryozoans, cephalopods, hard and soft corals, gastropods, echinoderms and teleost fish), provide an analysis of current challenges in the discipline and start a discussion of what sclerochronology can offer for Antarctic research in future. The Southern Ocean ecosystem remains largely unstudied in part for its remoteness, extreme climate and strong seasonality. This lack of knowledge, some of it even on basic biological information, it is especially worrying due to ongoing climate-driven changes that the Southern Ocean ecosystem is experiencing. Lack of long-term in situ instrumental series has also being a detriment to understand long-term feedbacks between the physical environment and the ecosystem. Sclerochronology, the study of periodic accretional patterns in the hard body structures of living organisms, has contributed to a wide range of Antarctic research disciplines (e.g. paleoclimate reconstructions, population structure analysis, environmental proxies). This review highlights a disparity in research focus by taxa with some groups (e.g. bivalves, teleost fish) attracting most of the research attention, whereas other groups (e.g. gastropod) have attracted much little research attention or in some cases it is almost non-existent (e.g. echinoderms). Some of the long-lived species considered in this review have the potential to provide the much-needed high-resolution eco-environmental proxy data and play an important role in blue carbon storage in the Sothern Ocean. Another issue identified was the lack of cross-validation between analytical techniques.
Graphic abstract

Similar content being viewed by others

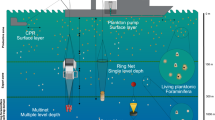
Introduction
Sclerochronology is the study of growth patterns preserved in the hard skeletal structures of living organisms (e.g. shells, bones, coral reefs) (e.g. Clark 1974; Richardson 2001; Gröcke and Gillikin 2008; Oschmann 2009; Schöne and Gillikin 2013; Butler and Schöne 2017; Butler et al. 2019; Gillikin 2019; Trofimova et al. 2020). Transferring much statistical knowledge from dendrochronology (tree-ring analysis), sclerochronology has focussed on a wide range of scientific interests including ontogeny, paleoclimate, archeology and geochemistry. A key concept in sclerochronology is that all individuals within a population respond to common environmental cues/drivers so they display common synchronous growth in addition to their own individual biological growth rhythm. The sclerochronological literature is rich in terminology (e.g. growth increments, growth rings, growth marks, growth checks) which also will be reflected in some part in this review. A distinction should be made in advance that in some sclerochronological records (i.e. fish otoliths and gastropod and cephalopod statoliths the term growth ring) would be preferred over growth increment. Also the term “annuli” is also commonly used in teleost fish otolith research; however, in this review it has been substituted for the term “ring” to provide a reduced terminology and avoid confusion. Another clarification to be made is the distinction between the terms: growth increment and growth check or mark. Growth check or mark is used in this review as the thin line that it is formed in many sclerochronological records at the end of the growing season, whereas growth increment is used in this review as the portion of growth contained within two consecutive growth marks/checks. Statistical techniques used in sclerochronology (many derived from dendrochronological research) such as: detrending and regional curve standardization allow to separate individual and populational biological growth rhythm from the common synchronous growth attributed to common environment drivers in the life history growth pattern (e.g. Briffa and Jones 1990; Esper et al. 2003; Butler et al. 2010; Roman-Gonzalez et al. 2017). Thus allowing to investigate independently both the biological (i.e. individual specific), population and environmentally driven growth (Fig. 1). Some of the research concepts studied by sclerochronology are outlined in Fig. 1, which will be discussed further in the main text of this review. Much sclerochronological research has been undertaken in temperate environments and has focussed on long-lived species (some of which can attain lifespans spanning several centuries) for a number of reasons (e.g. ageing studies, constructions of long proxy records that extend beyond the onset of instrumental dataset). Commercially important shorter-lived species have attracted attention in the context of provide scientific advisory datasets (e.g. population age structure, recruitment, age of maturity) to the respective fisheries’ governing bodies (e.g. Panfili et al. 2002; Hollyman et al. 2018a, b; Hunter et al. 2018).

Sclerochronology can be used with a varied range of marine organisms (e.g. fish, bivalves, gastropods, bryozoans). In addition, sclerochronology can provide an insight into varied fields such as: (1) ecosystem composition and trophic webs, (2) primary production and marine carbon fluxes, (3) population structure and distribution and ageing, (4) current and past ocean temperature and (5) oceanic circulation which in the Southern Ocean are heavily affected by sea ice processes and salinity gradients (*)
Many authors highlight the systemic lack of long-term instrumental records in the Southern Ocean and how this gap limits our understanding of processes and feedbacks in the climate system at southern high latitudes (e.g. Turner and Comiso 2017). Much of this is due to the remoteness and harshness of the environment, which limits the operational life of deployed equipment, and the logistical and economic costs of developing long-term environmental monitoring programmes. Many nations operating Antarctic research programmes do not have a year-round, long-term marine monitoring systems in place, such as: the UK RaTS (est. 1997), USA Palmer-LTR (est. 1990) and McMurdo Oceanographic Observatory (MOO, est. 2017). These few programmes are bound to the location of their respective Antarctic research bases, which does not provide an adequate spatial coverage for the Southern Ocean.
Thus, the great majority of the Southern Ocean research community has been forced either to deal with scarce environmental datasets and/or take advantage of satellite measurements, which offer ample spatial coverage. Since the onset of the satellite era, environmental monitoring by remote sensing has been improving constantly with increasing resolution, accuracy, spatial coverage and range of parameters measured (Rayner et al. 2003). However, these records do not have a long temporal extent, with earliest satellite measurements only going back to the 1970s for the Southern Ocean. In addition, instrumental data collection from coastal areas, where the interactions between terrestrial and marine environments generate complex feedbacks, are largely underrepresented compared to open oceanic areas.
A central question is what can sclerochronology can offer in particular to Antarctic research? As mentioned above, the lack of long-term instrumental records, the close association of those with Antarctic research bases (and therefore with a limited spatial coverage), remoteness and technical sampling difficulties associated with the extreme climate of the Southern Ocean has hindered the understanding of ecological, physical and climatic processes in the Southern Ocean. Therefore sclerochronology has the potential to provide directly biological and ecological information encoded in the growth increment patterns and in the geochemistry of the hard skeletal structures for many Antarctic species. Compared to other proxy records such as ice or sediment cores, sclerochronology can be annually resolved and absolutely dated (i.e. specific calendar years can be assigned to each growth increment) and the records have the potential to extend beyond the onset of in situ instrumental records with crossmatching techniques. In addition, sclerochronological material such as shells, coral skeleton, cephalopod beaks, can be collected from research cruises using a range of sampling gear away from Antarctic research bases and therefore extend the spatial coverage of scientific data. Many of the species suitable for sclerochronological analysis inhabit environments where scientific data are scarce (e.g. deep-sea, littoral areas) and they also have a pan-Antarctic distribution providing the potential to develop networks of local climatic monitoring stations. Data extracted from sclerochronological records can also feed back into global climate models to provide more accurate predictions.
Thus purpose of this manuscript is to provide the research community with a comprehensive review of the research on, and related to sclerochronology in the Southern Ocean and subantarctic islands, provide an analysis of current limitations of Antarctic sclerochronology and a discussion of possible research avenues to expand the use of Antarctic sclerochronology. This review will cover publications on the following topics: chronology construction, ontogeny and age determination, paleoclimate reconstructions and ontogenetic geochemical variability, and on the following taxa: bivalves, gastropods, cephalopods, echinoderms, brachiopods, bryozoans, corals and teleost fish. The common thread in all studies reviewed is the analysis of accretional hard skeletal structures of marine organisms which record the specimens’ life histories. Other general topics (e.g. general biology, physiology), however relevant tangentially to the field, are not explicitly discussed although some are referenced when considered appropriate. A summary table is provided detailing information about species name, taxa, known distribution, maximum estimated lifespan, age determination methods in the literature and relevant citations (Table 1).
State of the art knowledge
Bivalve molluscs
One of the classical major research areas in sclerochronology has been bivalve molluscs. Three modern marine bivalve species have attracted most attention (see Table 1) in the Southern Ocean: Adamussium colbecki (E.A. Smith, 1902), Laternula elliptica (King, 1832) and Aequiyoldia eightsii (Jay, 1839) (previously known in much of the literature as Yoldia eightsi).
The first study which tried to determine the age of A. colbecki, along with L. elliptica, was done undertaking X-ray photographs, which provided lifespans of 6–7 and 12–13 years respectively (Ralph and Maxwell 1977). In addition to X-ray photography, the periodicity of the external striations shell of A. colbecki was also used to determine the age of specimens collected from McMurdo Sound (Stockton 1984). The outcome of this analysis was not conclusive due to the low calcification of the shell and it was determined unlikely that each striation represented 1 year of growth (Stockton 1984). Further analysis of cyclical variation (i.e. wide and narrow) external shell striae pattern in A. colbecki was used for age estimation in a population structure study from the Ross Sea (Berkman 1990). In addition, Berkman (1990) also used a light table to reveal internal banding in the shell, using these two methods it was hypothesised that narrow external striae were formed during the winter and wider striae during the summer. Similar to the work cite above, the A. colbecki population in Terra Nova Bay was investigated by Cattaneo-Vietti et al. (1997). X-ray photography was also successfully used as an age determination technique and annual calcification rates were also found similar to those from McMurdo populations (Cattaneo-Vietti et al. 1997). More recently, an excellent study compared different age estimation methodologies in A. colbecki and discussed the possible caveats of each method (Cronin et al. 2020). The results indicated that X-ray photography seems to provide the most accurate age estimations along with a newly introduced age estimation method in A. colbecki, annuli (defined here as “whitish rings in concave depressions against the brown, convex shell surface”) counting (Cronin et al. 2020). However, the main purpose of this study was to assess the influence of sea ice-related environmental condition upon the growth and longevity of A. colbecki. The results showed that sea ice conditions (i.e. annual vs multiannual) did not have any effect on A. colbecki lifespan, which seems to be controlled by temperature (Cronin et al. 2020). Contrary to the sea ice control over A. colbecki lifespan, sea ice conditions were found to significantly affect growth which was hypothesised to relate to nutrient availability depending on the sea ice conditions (Cronin et al. 2020). Further work with A. colbecki has focussed on understanding the geochemical variability of the shell with respect to growth increment pattern and environmental variability (Barrera et al. 1990; Berkman 1994; Heilmayer et al. 2003; Lartaud et al. 2010; Trevisiol et al. 2013). Three sampling methodologies were used to investigate δ18O and δ13C variability in A. colbecki; the first sampling method drilled several samples along the growing margin (from the anterior to the posterior part of the shell), the second method collected several samples along the dorso-ventral axis (following the maximum growing direction of the shell) and the third method collected samples from several location on the internal shell surface (i.e. in the concave part of the shell) (Barrera et al. 1990). No significant relationship was found in the δ18O values around the growing margin (apart from one sample collected on one sample collected from one auricle); however, variability was significantly higher for δ13C values (Barrera et al. 1990). Subsequent work by the same authors expanded the investigations of lateral δ18O and δ13C variability in L. elliptica shells and found statistical differences in samples collected along the shell margin (Barrera et al. 1994). This consideration of lateral (i.e. along the anterior–posterior axis) geochemical variability done by Barrera et al. (1990, 1994) is often missing in other sclerochronological work based on bivalve species. It is a common practice that bivalve shells are drilled for geochemical studies only along the maximum dorso-ventral axis and it is unknown whether displacing the sampling axis to either the anterior or posterior part of the shell would produce statistically different geochemical data series. Berkman (1994) provided the first δ18O series for A. colbecki which used to determine where the shell was precipitated in equilibrium with the seawater, which was the case. In addition, radiocarbon analysis were used to determine the reservoir age derived from Antarctic bivalves which was determined to be 1400 ± 100 years (Berkman 1994). A. colbecki specimens collected from Terra Nova Bay were studied using X-ray photography and δ18O, providing new data on δ18O variability for this species (Heilmayer et al. 2003). This cross-technique validation study (Heilmayer et al. 2003) is a great example of the much-needed methodological analysis in sclerochronology. It was also the growth marks were formed during the austral winter and that some growth marks may not be visible with x-ray photography (but could be determine using δ18O values), which could lead to the underestimation of true age. Subsequent work compared new δ18O A. colbecki records with environmental parameters (e.g. sea surface temperature, SST, salinity) and determined that the reconstructed temperatures were more similar to the mean summer SST rather than the mean annual SST (Trevisiol et al. 2013). In addition, preliminary work on the trace element (i.e. Li, B, Mg, Mn, Co, Sr, Ba and Pb) composition of A. colbecki specimens, collected from Dumont d’Urville Station, provided a concentration baseline for these elements (Lartaud et al. 2010); although the interpretation of the variability in the element records remains unclear. A more recent study used atom probe tomography (APT) to analyse the organic matrix of A. colbecki, which confirmed the presence of carboxylate proteoglycans associated with biomineralization (Pérez-Huerta et al. 2020). These compounds have a special relevance to sclerochronology since they are nitrogen rich and have the potential to be associated with trace elements (e.g. Mg) and therefore can affect any δ15N and Mg-Ca proxies (Pérez-Huerta et al. 2020). The authors used SEM imaging to identify growth increments associated with external striae (Pérez-Huerta et al. 2020); however, this has proven not the most desired method (Cronin et al. 2020). Nonetheless Pérez-Huerta et al. (2020) reported a set of substriae in the cross-sections of the shell associated with external shell striae, which is the first time that such a feature is reported, although not much further analysis was done.
Laternula elliptica is the largest bivalve species in the Southern Ocean, which in part facilitates the technical work in the laboratories compared to other significantly smaller bivalve species. Pioneering sclerochronological work using L. elliptica established the annual periodicity of the shell growth increment pattern using δ18O and δ13C (Brey and Mackensen 1997). Expanding the research on the δ18O variability of L. elliptica shell revealed differences between seawater temperature instrumental data and δ18Oshell-derived temperatures (Tada et al. 2006). The authors attributed this difference to changes in salinity due to the influence of ice-melt run off and pointed that with better constraints of the parameters influencing ice-melt run off, volumes of ice-melt run off could be calculated based on δ18Oshell (Tada et al. 2006). In addition, differences found in the δ18Oshell between early and late ontogenetic growth increments seems to suggest that L. elliptica grows continuously throughout the year during the first years of growth but ceases to grow during winter as the individual gets older (Tada et al. 2006). Further work on L. elliptica collected from King George Island explored the relationships between shell growth, different environmental parameters (e.g. SST, surface air temperature, turbidity) and climate indices (Southern Annular Mode, SAM, and El Niño Southern Oscillation, ENSO) (Brey et al. 2011). Their results found that the growth of L. elliptica was closely associated with ENSO and surface air temperatures (SAT) but surprisingly no clear relationship was found between L. elliptica growth and sea surface temperatures (Brey et al. 2011). Seasonal variability of SAT was hypothesised to influence sea ice formation and glacial meltwater runoff which was also associated with increased sediment suspension, and which in turn affected indirectly L. elliptica growth (Brey et al. 2011). In addition, a shift in the metabolic maintenance costs (i.e. energy requirements for the organism to keep normal body functions) of L. elliptica was associated with a significant increase in the sediment load at their study site. More recently, high-resolution trace element and δ18O and δ13C datasets derived from L. elliptica shells were used to the impact of glacial melt water impulses on coastal Antarctic areas (Woo et al. 2019). In addition, this study compared L. elliptica specimens collected from the Antarctic Peninsula and from East Antarctica, two regions affected very differently by climate warming, which clearly show a stronger impact of glacial melt in the Antarctic Peninsula population (Woo et al. 2019). In addition, physiological and ontogenetic studies on this species have used sclerochronological age determination methods to assign individual ages for collected specimens. An example of this approach was carried out when comparing the impact of physiological functions on the maximum attainable lifespan of the temperate bivalve Mya arenaria Linnaeus, 1758 and L. elliptica, which occupy similar ecotypes in their respective environments (Philipp et al. 2005). The use of trace metals in bivalve shells as environmental proxies have always been an attractive prospect; however, there has been difficulties in attributing trace metal variability to environmental drivers. Detailed information about the environmental drivers affecting trace metals and high-resolution sampling are key to comprehend trace metal incorporation in the shell. Dick et al. (2007) analysed trace metal concentration (i.e. Al, Fe, Mn, Cu, Pb and U) along the umbo of L. elliptica using sequential laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS). Their main findings were that biology-driven processes acted strongly in the incorporation of these elements into L. elliptica shell as the element ratios did not resemble the surrounding water, sediment, or Earth’s crustal basal composition (Dick et al. 2007). The authors also pointed out that sampling resolution was still a limitation and an improvement in analytical techniques could offer a new understanding of trace element incorporation in the shell (Dick et al. 2007). A modern example can be found in the work done by Wing et al. (2020) in L. elliptica, where 17 trace elements were analysed over shells lifespans ranging from 13 to 25 years. The lifespans of these specimens coincided with an 8-year period of persistent sea ice influenced by the presence of two large icebergs in McMurdo Sound, the area of study (Wing et al. 2020). The authors discussed the relationships between sea ice persistence, marine primary production and trace metal availability and incorporation into the shell. As hypothesised by Dick et al. (2007) an increased sampling resolution in LA-ICP-MS provided Wing et al. (2020) a technical advance in the study trace element incorporation into the shell of L. elliptica. Variability of conservative (Li, Ba, Mg, and Sr), bioactive (Fe, Co, Ni, Zn, and P), and highly scavenged (Al, Cu, and Pb) trace metals in L. elliptica shell seem to be highly coupled with sea ice variability and its cascading effects in the ecosystem (Wing et al. 2020).
Early study of shell growth and the growth increment pattern A. eightsii was carried out using acetate peel replicas and radioactive calcium (45Ca) incorporation into the shell (Nolan and Clarke 1993). This study was the first to report primary (Fig. 2d), secondary and tertiary growth increments and although the primary growth increment pattern was assumed to have an annual periodicity there was no final confirmation (Nolan and Clarke 1993). Additionally, Nolan and Clarke (1993) reported a maximum age of 52 years based on size-frequency data. An exploration of A. eightsii as a sclerochronological record established the first chronology for this species from historical (i.e. 1988) live-collected specimens from the South Orkney Islands and explored the relationships between shell growth and different environmental parameters (e.g. fast-ice duration, SST, chlorophyll concentration, Roman-Gonzalez 2011). In addition, this study used the advantage of repeated specimen sampling throughout the year to assess the formation and periodicity of the primary growth increment pattern observed firstly by Nolan and Clarke (1993), which was confirmed to be annually formed (Roman-Gonzalez 2011). Subsequent development of these results found significant negative correlations between shell growth and fast-ice duration and winter duration and significant positive correlations between shell growth and SST (Roman-Gonzalez et al. 2016). Further work on A. eightsii has focussed on the development of new chronologies and δ18O and δ13C records from live-collected specimens collected near Rothera Research Station, West Antarctic Peninsula, and historical specimens from the South Orkney Islands (Roman-Gonzalez 2018). These geochemical analysis of A. eightsii shells proved for the first time that temporally aligned δ18O and δ13C records from several specimens can be constructed (Roman-Gonzalez 2018). However, an isotopic fractionation was present in the δ18O data shell, modelling of the theoretical environmental conditions (i.e. temperature and salinity) was done by calculating a synthetic δ18O of the seawater (hereafter δ18Osyn) based on the δ18O values from the shell and the instrumental equilibrium equation from the Rothera Biological Time Series (RaTS) (Roman-Gonzalez 2018). The modelling results showed that disequilibrium could be accounted either if: (i) the ambient water was 2.05 fresher (as compared registered in the RaTS dataset), (ii) the ambient water was 2.45 °C warmer or (iii) the ambient water was 1.3 °C and 0.91 fresher (Roman-Gonzalez 2018). Scenarios (i) and (iii) were discarded due to the unrealistic increase in water temperature and it was hypothesised that the disequilibrium was due to the influence of glacial meltwater in the shallow littoral environments where the specimens were collected (Roman-Gonzalez 2018). Lateral fractionation (i.e. along the anterior–posterior axis) of δ18O and δ13C values was also studied in A. eightsii from samples collected from the outer shell surface; significant differences were found but with no clear relationship between isotope values and sampling locations (Roman-Gonzalez 2018). This variability was hypothesised to be due to varying growth increment thickness along the anterior–posterior axis (Roman-Gonzalez 2018) although more research is needed to investigate the lateral δ18O and δ13C variability. In addition, the study of ontogenetic growth patterns, critical for the development of reliable climatic proxies, in A. eightsii and L. elliptica has been explored (Roman-Gonzalez et al. 2017). The authors found a secondary ontogenetic trend (with a periodicity of approximately 9 years) in A. eightsii which may be related to reallocation of energetic resources between somatic growth and gonad development. A statistically significant cyclicity was observed in the residuals after the primary negative exponential growth; starting at around 16 ontogenetic year (Roman-Gonzalez et al. 2017) which coincides approximately with the onset of maturity in A. eightsii (Nolan and Clarke 1993; Peck et al. 2000). It was hypothesised that this secondary cyclicity was due to an innate growth rhythm in A. eightsii, independent from environmental variables, related to the reallocation of energetic resources (Roman-Gonzalez et al. 2017). Subsequent biological work (see Lau et al. 2018), unrelated to sclerochronology, based on A. eightsii provided further confirmation of the hypothesis done by Roman-Gonzalez et al. (2017).

Microphotograph examples of sclerochronological archives from marine species from the Southern Ocean. a Dissostichus eleginoides otolith section, b Neobuccinum sp. statolith section, c Bryozoan Melicerita obliqua colony and d Shell section of the bivalve Aequiyoldia eightsii. Scale bars are provided. (a, b courtesy of Philip Hollyman and c courtesy of David Barnes, both British Antarctic Survey)
The small bivalve Lissarca notorcadensis Melvill and Standen, 1907 has a circum-Antarctic and eurybathic distribution, and is normally associated with sea urchins, hydroids and bryozoan colonies (Brey and Hain 1992). An analysis of the growth increments observable on the external shell surface of L. notorcadensis provided a maximum estimated lifespan of 16 years (Brey and Hain 1992). Similarly to the work done in A. colbecki, the external shell markings of the small eurybathic (from 8 to 2350 m water depth) brooding bivalve Adacnarca nitens Pelseneer, 1903 revealed a maximum estimated lifespan of 20 years (Higgs et al. 2009). Using external shell markings as a method for age estimation is perhaps the simplest of tools available in sclerochronology. However, this approach can be challenging, misleading or impossible depending on factors such as species longevity or shell preservation. For example, this issue was highlighted in studies of A. nitens and L. miliaris as external shell damage led to an underestimation of the number of external shell markings and therefore age (Richardson 1979; Higgs et al. 2009). Although the methodological information provided by Richardson (1979) regarding age determination was unclear and therefore difficult to assess properly.
Work on fossil shells from Antarctic waters has received limited attention to date. The analysis of fossil shells of the Eocene bivalve Cucullaea raea Zinsmeister (1984) revealed an annually formed growth increment pattern based on sectioned shells and stable oxygen and carbon isotope ratios (δ18O and δ13C) with a maximum estimated lifespan of 127 years (Buick and Ivany 2004). The growing season for C. raea was counterintuitively determined to be during the Austral winter when food resources are most scarce; the authors hypothesising spawning events as the reason for summer growth cessation (Buick and Ivany 2004). This research was expanded with geochemical analysis (i.e. δ18O, δ13C, δ 87Sr) of more C. raea specimens and additional fossil species: venerid bivalve Eurhomalea newtoni Wilckens, 1911, naticid gastropod Polinices subtenuis Ihering, 1907 and terebratellidae brachiopod Bouchardia antarctica Buckman, 1910 (Ivany et al. 2008). These analysis provided information concerning the environmental conditions (e.g. progressive marine cooling, possible changes in circulation or in the shallow marine carbon cycle) occurring during the formation of these Antarctic Eocene deposits at Seymour Island, Antarctic Peninsula. Subsequent work on C. raea explored the effects of El Niño Southern Oscillation (ENSO) variability during the Eocene on the growth increment patterns and on driftwood from a co-occurring coniferous tree collected from Seymour Island on the Antarctic Peninsula (Ivany et al. 2011). More recently, another extinct Antarctic bivalve studied has been Lahillia larseni Sharman and Newton, 1897 (Mohr et al. 2020). High-resolution δ18O and δ13C series were used to provide indication of the palaeoenvironment during the Cretaceous-Paleogene boundary. The data suggests a period of strong marine anoxic conditions and two periods of climate warming, one of them concurrent with high volcanic activity (i.e. Deccan traps eruptions, Mohr et al. 2020).
Assessing correctly the marine radiocarbon reservoir is crucial for understanding the stability of ice sheets, ocean circulation and sea-level change. Carbonate material from fossil and modern calcareous marine organisms play an important role in determining the marine radiocarbon reservoir. A study, based on Antarctic bivalves (i.e. A. colbecki, Thracia meridionalis E.A. Smith, 1885, L. elliptica, A. eightsii and Hiatella antarctica (Philippi, 1845), gastropods (i.e. Neobuccinum eatoni (E. A. Smith, 1875) and Nacella concinna (Strebel, 1908); Fig. 2b) and the sea urchin Sterechinus neumayeri (Meissner, 1900), calculated a decrease of nearly 500 years in the marine radiocarbon reservoir during the second half of the twentieth century and suggested a radiocarbon reservoir correction of 1300 ± 100 years for the Antarctic marine ecosystem during the Holocene (Berkman and Forman 1996). Studies like this are important as they provide some information regarding the local variation from the global average (ΔR) marine reservoir effect (e.g. Alves et al. 2018).
Gastropod molluscs
Studies of Antarctic gastropod molluscs are in general very limited and this is especially true for sclerochronological studies focussing main on the Antarctic limpet N. concinna. Distribution, growth and reproduction of the Antarctic limpet, N. concinna, was studied using external line counting for age determination and its relationship with shell size (Picken 1980); however, this technique can be highly inaccurate and can only be used for preliminary assessments. The alternative combined technique of strontium labelling (i.e. Sr enriched water) and SEM proved successful for identifying the growth increment pattern in N. concinna shell (Peck et al. 1996). The study highlighted Sr incorporation pathways into the shell, periodicity of growth increment formation (which was found not to be daily), and minimum residence times of Sr in the seawater before being incorporated into the shell. Similarly, the use of calcein staining, SEM and the study of the internal growth increment patterns of N. concinna, provided lifespan estimations between 3 and 5 years (Clarke et al. 2004), which contrasts with longer lifespans estimated during earlier investigations (i.e. 21 years) (Picken 1980). Further work on N. concinna showed that this species presents technical challenges in obtaining clear profiles of sectioned shells, obtained from shell middens, making it difficult to read the internal growth increment pattern (Roman-Gonzalez 2011). In addition, sectioned shells showed an intricate growth increment pattern in the outer shell layers (Roman-Gonzalez 2011). This shell region showed periodic changes in growth direction, each cycle containing an extended period along the main growth axis and a short growth period perpendicular to the main growth axis (i.e. downwards); this change in growth direction was hypothesised to be related to winter conditions (short growth period) when the limpet mantle was more time retracted into the shell and the animal was clamped to the rock (Roman-Gonzalez 2011). More recently, shell growth of intertidal and subtidal populations of N. concinna were studied to assess the environmental plasticity of this species (Lomovasky et al. 2020). This comparative study found morphological difference in the shell but no differences in the maximum lifespan (both reaching 20 years) between the two environments (Lomovasky et al. 2020). Food availability, available feeding time, predation stress and energy gaps associated with higher mobility were considered to be the most likely environmental drivers affecting shell growth (Lomovasky et al. 2020).
A study on the prosobranch gastropod Laevilacunaria antarctica (Martens, 1885) population from Signy Island over a period of almost 2 years found strong seasonal summer growth thought to be controlled by food availability (Picken 1979). However, methodological information regarding year class calculation is unclear in the study, and there is also a lack of information regarding shell structure and geochemical composition for this species. Similarly, the axial ribs of the small pelagic gastropod Limacina rangii (d’Orbigny, 1835) (previously known as Limacina antarctica) were hypothesised to have a daily formation period; however, the study, which used scanning electron microscopy (SEM), did not provide an estimation of number of ribs per individual (Sato-Okoshi et al. 2010). Another internal calcified structure in gastropods are the statoliths, which are used for gravitational orientation. Preliminary work on statoliths from Neobuccinum sp. (Fig. 2b) collected in coastal fjords along the West Antarctic Peninsula has been done (Hollyman P, British Antarctic Survey, 2019, per. comm.). Statoliths showed concentric rings that may be useful in sclerochronological analysis if their periodicity can be validated.
Cephalopod molluscs
Sclerochronological studies of cephalopods have been traditionally linked, as in the case of fish, to marine resource management. The lack of industrial commercial cephalopod fisheries in Antarctic waters is likely to have had an impact on the scientific literature. Despite this, there are few studies focussed on the growth of Antarctic cephalopods. An extensive review on Antarctic squid research from the Soviet/Russian research programmes provides valuable information for sclerochronologists who wish to develop these records further (Filippova 2002).
The statolith microstructure of seven Antarctic squid species (Moroteuthopsis longimana (Filippova, 1972) referred in the study as Kondakovia longimana, Psychroteuthis glacialis Thiele, 1920, Brachioteuthis sp., Mastigoteuthis psychrophila Nesis, 1977, Galiteuthis glacialis (Chun, 1906), Bathyteuthis abyssicola Hoyle, 1885 and Alluroteuthis antarcticus Odhner, 1923) collected from Prydz Bay, East Antarctica, was analysed in order to provide basic knowledge on population structure of these important members of the Antarctic ecosystem (Jackson and Lu 1994). The number of increments in the statoliths varied and it was not possible to count them in B. abyssicola or in A. antarcticus as the growth increment pattern was too faint (Jackson and Lu 1994). The authors hypothesised that the increments are formed daily in the studied squid species; however no external confirmation of the periodicity is provided. The analysis of the growth increment patterns present in the gladius or squid pen was proposed as an alternative to statolith analysis for squid age determination (Bizikov 1991). The reported lifespan was of about 5 years for the Antarctic squid M. longimana, if the periodicity is to be considered daily (Bizikov 1995). Although longevity is considered to be short (i.e. 1 to 2 years) for most Antarctic squid species where data are available, there are many other species which have not been studied. A particularly interesting species is the colossal squid Mesonychoteuthis hamiltoni Robson, 1925, which exclusively inhabits the Southern Ocean. The longevity of this species remains unknown, although it is thought to be longer than more temperate species (normally ranging between 12 and 18 months; Rosa et al. 2017).
Octopods also possess an internal calcified structure named stylet (a vestigial shell) which is also formed periodically as the animal grows. Literature on age determination and growth increment patterns in Antarctic octopods is also limited. The analysis of the growth pattern in six stylets of the octopus Megaleledone setebos (Robson, 1932) showed the number of increments ranging between 167 and 1077 and although the growth increment pattern was reported to be faint, it was clearly readable (Barratt and Allcock 2010). If the periodicity is to be assumed daily the lifespan of M. setebos is at least 3 years.
Another source of sclerochronological material in octopods can also be found in their beaks. Octopus beaks are chitinous structures that grow with the animal and have several parts. The beaks can be collected from live-collected octopod specimens but they can also be found regularly in stomach contents of their predators (e.g. albatrosses). Regularly formed concentric rings have been found in the beaks of temperate octopus species (Hernández-Lopez et al. 2001), although this feature is poorly cited for Antarctic species. This could indicate that either the concentric rings are a species-specific feature, that there a lack detailed research on the topic or just that it is a piece of information that is commonly underreported in the scientific literature.
Chemical analysis of the beaks of Antarctic octopods [i.e. Adelieledone polymorpha (Robson, 1930) and Pareledone turqueti (Joubin, 1905)] have been used to determine levels of mercury (Hg) in the environment (Matias et al. 2020); however, whole upper and lower parts of the beaks were grounded and homogenized for analysis and no chronological analysis of the beak was done. A chronological analysis of the beak could have provided a detailed history of environmental Hg concentration in the environment that could be associated with either specific calendar years or animal ontogenetic ages. Nonetheless, this study highlights the potential of octopod beaks as sources of material for environmental monitoring. _Another geochemical study of octopod beaks has focussed in the development of stable nitrogen and carbon isotopes ratios (δ15N and δ13C) to determine the trophic levels of the diet of M. setebos (Queiros et al. 2020). The stable isotope series data showed small but significant variations throughout the lifetime of M. setebos, which was attributed to changes in diet, of at least one trophic level, rather than an ontogenetic change in habitats (Queiros et al. 2020). An often cited comment by researchers studying Antarctic cephalopods is the lack of knowledge of the environmental driver(s) producing a daily pattern in the hard accretionary structures at high latitudes, where 24 h light cycles are present during Antarctic spring/summer. Whether the daily growth patterns in cephalopods are formed by either internal physiological clocks independent from the animal’s activity or their environment, or are due to retained evolutionary vertical migrations in the water column, as in the case of pelagic species, is still to be investigated.
Barnacles
The use of barnacle shells in sclerochronological studies has not been explored in detail, even considering temperate and tropical environments. The study of Antarctic barnacles by sclerochronology is limited to one single species: Bathylasma corolliforme (Hoek, 1883), which has shown promising results such as clear growth increment an extended lifespan (up to 73 years), precipitation of the shell in equilibrium with seawater and widespread distribution in the Southern Ocean (Burgess et al. 2010). Stable oxygen and carbon isotope ratios series from B. corolliforme shell was used to confirm that the growth increments were annually formed, although it was not possible to determine whether growth occurred continuously throughout the year (Burgess et al. 2010). Radiocarbon and U-Th analysis were also collected to assess the validity of these techniques in barnacle shells. U-Th dating was not found to provide consistent data, which the authors attributed to an open-system U exchange between the shell and the environment (Burgess et al. 2010).
Echinoderms
Sclerochronological studies of echinoderms (starfish, brittle stars, sea urchins, sea cucumbers and crinoids) have been neglected compared to other taxa, not only for Antarctic species but also in temperate environments. This is despite their widespread spatial and temporal (i.e. in the fossil record) distribution and desirable characteristics for building sclerochronological records. Growth increment patterns in sea urchins are found in the jaws (i.e. half pyramids) of Aristotle’s lanterns (i.e. mouth piece), skeletal and genital plates. To date, the only sclerochronological study of an Antarctic sea urchin, S. neumayeri, showed a light–dark banding pattern in the Aristotle’s lanterns, which was assumed to be formed annually (one light and one dark band per year), and provided a maximum lifespan of almost 40 years (Brey et al. 1995a). However, it could be possible that the age of these specimens was underestimated, as it is expected that the jaws suffer from erosion during grazing and some increments may have been lost.
The technical difficulties of working with brittle stars are shown by a comparative study of two age determination methods (i.e. growth rings readings as increment size data, method A, and a corrected method for overgrowth of the central articulated area, method B) based on growth marks in the ossicles (i.e. calcareous exoskeleton elements) of the Antarctic species Ophionotus victoriae Bell, 1902 (Dahm and Brey 1998). This study provides a basic methodology and reviews the shortcomings of each method such as lack of pith offset (i.e. difference between the firstly formed increment and the first readable increment) determination, introduction of errors due to high variability in the individual growth in method A, and more intense statistical treatment and the introduction of subjective interpretations in the method B.
Brachiopods
Brachiopods are a group of marine invertebrates with a two-valved shell that encapsulates all soft body tissues apart from the pedicle (i.e. a soft tissue stalk), which functions as an anchor point to hard structures on the seabed. Interest in their study has been driven in part by studies on physiological adaptations to the cold and extreme seasonality of the Antarctic environment. However, most of this research focus on brachiopods has been on extinct fossil species and little attention has been given to living species. In the Antarctic, the focus has been on three living species, Liothyrella uva (Broderip, 1833) and Magellania fragilis Smith, 1907 Aerothyris kerguelensis (Davidson, 1878) and exploratory work has been undertaken on Neorhynchia strebeli (Dall, 1908). Early work on L. uva analysed the relationship between growth checks in the shell and radiocarbon (14C) bomb signals (Peck and Brey 1996). Some of the main highlights of this study are that the growth checks of L. uva are not formed annually but with a quasi-biennial periodicity, a 2 to 5 year lag in the shell 14C activity of the terrestrial environment (leading) over the marine environment and that the 14C source is due to atmospheric deposition and accumulation in ice rather than oceanographic processes (e.g. upwelling) (Peck and Brey 1996). This was interpreted as the influence of large resident times of the Antarctic Circumpolar Water with a large carbon pool and contradicted the theory that polar upwelling of deep water (with low 14C) would cause reduced 14C signals (Peck and Brey 1996). This is a clear example of how the combination of the study of the growth increment pattern and the geochemistry of shell (i.e. 14C) can provide results that overspill into other science disciplines (i.e. oceanography). Additional work covering basic investigations of L. uva growth rates, shell structure, morphology, effects of ocean acidification and geochemistry has been carried out by several authors (Peck and Holmes 1989; Peck et al. 1997; Parkinson et al. 2005; Goetz et al. 2009; Brand et al. 2013; Cross et al. 2015). The analysis of the growth increments of M. fragilis revealed a complex structure of punctae and ridges, periodicity of which was assumed to be annual (Brey et al. 1995b). Features such as distance between punctae and the formation of ridges in areas of high punctae density defined the structure of the increment growth pattern (Brey et al. 1995b). In addition, the authors point to the technical challenge of identifying the growth increments, especially when they become increasingly narrow; nevertheless, the best estimate provided a minimum lifespan of 49 years (Brey et al. 1995b). If as in the case of L. uva (Peck and Brey 1996), it is demonstrated that the growth checks of M. fragilis have a biennial (every 2 years) periodicity, this would mean that the lifespan of M. fragilis might extend to a century or longer. The analysis of the external shell marking on N. strebeli revealed that the shell had up to 11 primary increments, which were assumed to be annual, and up to 100 secondary increments (Barnes and Peck 1997). The authors concluded that the lack of an independent age estimation method could lead to under or overestimation of the actual age, especially considering what has been found in other Antarctic brachiopod species, M. fragilis, whose growth increment pattern presented a biennial periodicity. More recently, a comprehensive analysis, considering: growth, geochemical composition (trace elements, stable oxygen and carbon isotopes), form and microstructure of the shell of A. kerguelensis provided novel sclerochronological information for this species (Gaspard et al. 2018). The growth increment pattern of A. kerguelensis was studied using SEM photography and trace element composition (elemental mapping SEM–EDS and electron microprobe WDS) and although the periodicity of the growth increments is still a matter of debate, the data presented in this study seem to indicate that they are annually formed (Gaspard et al. 2018), at least for this species. Other results highlighted by this study revealed that the anterior valve may be better suited to read the growth increments, the presence of microscopic boring organism in the shell and a relation between the growth increment microstructure and the geochemical (i.e. Mg and S bands) composition (Gaspard et al. 2018). The remineralisation of A. kerguelensis shells carried out by the microscopic boring organisms is always a possibility and can modify the geochemical composition of the shell. It was noted that the action of these organisms had an impact on the structure of growth increments and on Mg and S concentration (Gaspard et al. 2018); however, a more detailed study of the possible effects of these boring microorganisms on A. kerguelensis shell geochemistry (e.g. stable isotope fractionation) is needed, if this species wants to be considered as a palaeoenvironmental proxy.
Teleost Fish
Sclerochronological techniques have been used extensively by the research community focussing on fish population structure, especially with applications to fisheries management. Otoliths are small internal calcium carbonate structures inside the head of teleost fish which are used functionally as orientation sensory organs but which provide information on growth patterns, population structure and environmental drivers of fish growth. The literature on the study of Antarctic fish otoliths is extensive and encompasses a range of different age determination methods (see Table 1).
Studies based on fish otoliths providing basic biological information regarding age and population structure are common in all environments. Some examples of these are: the study of Gymnoscopelus nicholsi (Gillbert, 1911) in the South Shetlands, which provided a maximum estimated age of 7 years and gave some insights into the population structure (Linkowski 1985), the study of Pagothenia borchgrevinki (Boulenger, 1902) collected from McMurdo Sound which had a maximum lifespan of 7 years (Pankhurst 1990), the study of Trematomus bernacchii Boulenger, 1902 from the Ross Sea which provided maximum ages of 21 years for females and 16 years for males and insights into the ontogenetic growth of this species (La Mesa et al. 1996) and the study of Pleuragramma antarctica Boulenger, 1902 (previously known as P. antarcticum), Electrona antarctica (Günther, 1878), and Protomyctophum bolini (Fraser-Brunner, 1949) with a maximum calculated ages of 12 and 11 years respectively and no age for P. bolini. For other similar studies refer to the information collated in Table 1.
Recruitment processes are an important part in the study of wild fish populations, which require the analysis of larval stages and to determine age of the specimens (measured in days rather than years). An example such studies can be found in the analysis of otolith growth and RNA/DNA ratios of the Antarctic species: Notolepis coatsi Dollo, 1908, Gobionotothen gibberifrons (Lönnberg, 1905) and Trematomus scotti (Boulenger, 1907) (Morales-Nin et al. 2002). This study represented the first (and only) attempt at using these two methodological techniques in Antarctic teleost fish which provided satisfactory although limited results due to the limited number of samples (Morales-Nin et al. 2002). The authors reported difficulties estimating the age due to lack of contrast in the otoliths and also as no independent validation of the periodicity of the otolith rings was available the authors assumed to be daily (Morales-Nin et al. 2002). This lack of contrast using light microscopy in Antarctic fish otoliths [i.e. G. gibberifrons and Nototheniops larseni (Lönnberg, 1905)] was also highlighted by Ruzicka and Radtke (1995), who suggested the use of SEM photography instead of light microscopy.
La Mesa and Vacchi (2001) provide an excellent review and plenty of information on ageing techniques of notothenioid fish. This review summarises the findings of a series of technical workshop on age determination techniques, which can provide a significant amount of insight to those who want to extend the sclerochronological work in Antarctic fish. Some of the highlights of these workshops are: i) fish scales in some species may provide an underestimation of age and need to be considered with caution and ii) the crucial need of validation of daily increment for many species, especially for commercially exploited species (La Mesa and Vacchi 2001). Age determination techniques and maximum lifespan provided in La Mesa and Vacchi (2001) have also been collated into Table 1.
A comprehensive study of otoliths from the notothenioid fish Trematomus newnesi Boulenger, 1902 found daily but no annual increments (Radtke et al. 1989), with the longest lived specimen being 7 years old. Similarly, an excellent crosscalibration study of otoliths and scales from Notothenia rossii Richardson, 1844 provided age estimations up to 7 years and a solid background methodology for further work on this species (Barrera-Oro and Casaux 1992). Using these data the authors were able to estimate hatching dates and population age structure for N. rossii from Potter Cove, King George Island. The importance of the laboratory treatment of the otolith samples in order to preserve the growth ring pattern and some technical challenges when identifying the growth ring pattern were also highlighted.
Another group of teleost fish are dragonfishes, which are endemic from the Southern Ocean (Eastman 1993). A couple of studies focussing on the genus Parachaenichthys (Kompowski and Rojas 1994; La Mesa et al. 2012). The population and age determination of P. georgianus Fisher, 1885, and some P. charcoti Vaillant, 1906, specimens were analysed by Kompowski and Rojas (1994) from specimens collected from South Georgia. Age determination data on P. georgianus showed a maximum age of 10 years and 5 years for P. charcoti. In addition, age estimation based on otolith ring estimation P. charcoti collected from the tip of Antarctic Peninsula, was determined to be 9+ years (La Mesa et al. 2012). However, the authors highlighted that large specimens were underrepresented in their sampling which could result in the maximum age being underestimated. La Mesa et al. (2018a) provide some insight into the biology, population structure and ontogeny of the rare dragonfish Gerlachea australis Dollo, 1900 from specimens collected in the Weddell Sea. Based on the analysis 64 otolith ring counting (32 females and 32 males), maximum age for G. australis was determined to be 14 years based on (La Mesa et al. 2018a). The work in dragonfish was later expanded by the same authors, who analysed otoliths of relatively unstudied dragonfish species: Akarotaxis nudiceps (Waite, 1916), Bathydraco macrolepis Boulenger, 1907 and Bathydraco marri Norman, 1938, from specimens collected from the Weddell Sea (La Mesa et al. 2018b). The calculated age estimation based on otolith ring counting were: 6–11, 5–11 and 8–11 years for A. nudiceps, B. macrolepis and B. marri, respectively (La Mesa et al. 2018b). Although age estimations were done considering reader repeatability, no independent age determination assessment was carried out. An independent age determination technique such otolith markers, geochemical analysis or sequential seasonal sampling were found necessary in other Antarctic teleost fish species to validate otolith ring counting.
The analysis of otoliths from the Antarctic silverfish, P. antarctica, collected from the Weddell Sea revealed a distinct growth increment pattern, assumed to be daily, which provided a maximum estimated age of over 30 years (Radtke et al. 1993), which was significantly longer than that one provided by Liu and Chen (1995), which was 12 years. P. antarctica otoliths also showed two orders of increments: i) low order (high frequency, assumed to be daily) and ii) high order (low frequency) of an unknown origin and period. In addition, the variability of Sr/Ca profiles along the otolith’s main growth axis may reveal changes in hydrographic conditions during the lifetime of P. antarctica. A related study of N. larseni (previously known as Notothenia larseni) otoliths found a clear growth increment pattern, also assumed to be daily, which also presented cyclic variations in Sr/Ca (Radtke and Targett 1984).
An excellent review of age and growth is available for Antarctic notothenioid fish, providing age estimations for 16 species (e.g. Trematomus sp., P. antarctica, Dissostichus mawsoni Norman, 1937), general biological traits (e.g. age of maturity, size), ageing techniques and their validation and environmental controls of growth (La Mesa and Vacchi 2001). Some of the highlights of this review are: (i) lack of independent validation of ageing techniques, (ii) small size and high longevity are the norm, (iii) scarce data on maturity onset and (iv) low growth rates controlled by complex feedbacks within the environment. Additional work with D. mawsoni was subsequently carried in comparison with a closely related species living at lower latitude (i.e. D. eleginoides) (Horn, 2002). This study provides a detailed visual description of D. mawsoni otolith structure and a maximum age estimation of 35 years compared to 50 years in D. eleginoides. Further validation of translucent ring counting as an age determination technique in D. mawsoni was done by Horn et al. (2003) using length modal analysis and oxytetracycline-tagged fish. The results confirmed translucent ring, which were annually formed, counting in D. mawsoni as a valid method for age determination (Horn et al. 2003). The two most important commercial fisheries in Antarctic waters (besides Antarctic krill) are those that target: D. mawsoni and D. eleginoides. Routine sclerochronological work on otoliths of these two species is carried out to provide scientific data the fisheries management body, Commission for the Conservation of Antarctic Marine Living Resource, CCAMLR (e.g. Mormede et al. 2014; Hanchet et al. 2015; Lee et al. 2018; Parker et al. 2019). Some of the findings of these studies are: the importance of tagging programs to assess the population structure and stock when no fishery-independent estimates are possible (Mormede et al. 2014), and that D. mawsoni populations in southern McMurdo Sound are generally older and larger than the population living in northern McMurdo Sound and that both population may have a limited mixing (Parker et al. 2019).
Although much of the research done with fish otoliths has focussed on age determination, there has been some investigations as well of the geochemical composition of Antarctic fish otoliths as environmental proxy archives. A study of the trace element concentrations in otoliths of several ice fish species (i.e. T. bernacchii, T. pennellii Regan, 1914 and P. borchgrevinki) showed how trace elements can provide valuable insights into the ecology of these species and about patterns in sea ice dynamics (McMullin et al. 2017). Differential trace element concentrations associated between species-specific habitats and between sea ice years and open water years (i.e. when the sea ice broke completely in McMurdo Sound, the area of study) were the main result highlights (McMullin et al. 2017).
A detailed SEM analysis of the otolith microstructure of the Antarctic lanternfish, E. antarctica showed clear primary daily growth increments and provided an estimated lifespan of about 3 years (Greely et al. 1999). However, age estimations of E. antarctica based on otolith ring counting have provided a varied range of estimations (see as well: Linkowski 1987; Hulley et al. 1989), which was also discussed by Greely et al. (1999). More recently, taking advantage of a large collection of E. antarctica specimens from the Southern Kerguelen Plateau, Zhu et al. (2020) did a comprehensive analysis of age determination and provided a much longer maximum estimated lifespan of 9 years, which agrees with early work done by Linkowski (1987). A common research goal highlighted by several authors (Greely et al. 1999; Zhu et al. 2020) is the need of a validation study of the daily formation of otolith increments of E. antarctica such as: otolith markers (e.g. oxytetracycline incorporation) and periodic estimation of growth ring (sequential seasonal sampling).. Other Antarctic lanternfish species recently studied are: Gymnoscopelus braueri (Lönnberg, 1905) and Krefftichthys anderssoni (Lönnberg, 1905) (Saunders et al. 2019). The authors build-up of previous studies from these two species, concurrent in the Scotia Sea, providing detailed description and analysis of the growth patterns in the otoliths. Reported maximum ages for G. braueri is 6 years and two for K. anderssoni (Saunders et al. 2019). One concern raised by the authors is the lack of biological baselines for many species in the Southern Ocean. This is even more significant for non-commercial species and hinders any forecast on population effects from ongoing environmental changes in the region.
The use of SEM for assessing microincrements (daily) in the otoliths seem to provide positive results even when no clear ring can be discerned. An example of this was found in an ontogenetic study of Champsocephalus gunnari Lönnberg, 1905 and Notothenia rossii Richardson, 1844 (previously known as Notothenia rossii marmorata), whose otoliths revealed lifespans of 15 years and 6.5 years respectively (Radtke 1990). Another example was the study of larvae, which have not yet formed any ring, of L. larseni and G. gibberifrons; SEM analysis of the larvae’s otoliths provided age estimations which were then converted into hatch dates (Ruzicka 1996).
Otolith geochemistry has been used more recently in Antarctic fish species to assess their spatial distribution and population connectivity. A study of two populations D. mawsoni, from the Ross Sea and Southeast Pacific Basin, found that otolith nuclei and edge geochemistry indicated that this populations have a common habitat during juvenile stages (environmental homogeneity) but different habitats (environmental heterogeneity) during adult stages (Ashford et al. 2012). A similar more recent study of otolith geochemistry (i.e. element:Ca ratios for Mg, Sr and Ba) of E. antarctica suggested changes in spatial distribution during the lifetime of this organism (Zhu et al. 2018), heterogeneous habitats for juvenile E. antarctica and homogenous habitats for adult specimens. These contrasting results in ontogenetic changes in habitat distribution for two Antarctic fish species shows the importance of understanding species-specific ecological traits. A similarly earlier study found differences in otolith nuclei geochemistry in the Patagonian toothfish, D. eleginoides (Fig. 2a), between South America and Antarctic populations (Ashford et al. 2008), although this analysis did not cover to the entire lifespan of the specimens. The population differentiation during the lifetime of E. antarctica suggested by Zhu et al. (2018) was later on confirmed by the analysis of elemental ratios (i.e. MgCa−1 and SrCa−1) in E. antarctica otoliths between populations across the Antarctic Circumpolar Current (ACC) and the southern Kerguelen Plateau (SKP, Duan et al. 2020). Elemental ratios showed a total differentiation between the two populations during early stages of life and differences in later stages in life possibly due to different feeding habits (Duan et al. 2020).
Bryozoans
Bryozoans are sessile colonial organisms that form hard external structures made of calcium carbonate (calcite). As in corals, the colony can develop for several years even though individual zooids may have short lifespans. Additionally, bryozoan colonies can be either encrusting, forming flat mats covering a hard substrate such a rock, or erect, forming elaborate species-specific erect growing structures. These erect structures can present annually resolved growth increment patterns that can be used to interpret the environmental drivers controlling colonial growth (Stebbing 1971).
Early work determining lifespans of Antarctic bryozoans estimated the maximum lifespan of Isosecuriflustra tenuis (Kluge, 1914), previously known as Alloeflustra tenuis, and Cellarinella watersi Calvet, 1909 from Signy Island to be 26 and 9 years respectively (Barnes 1995). In addition, a comparison of the growth seasonality of A. tenuis, Nematoflustra flagellata (Waters, 1904) and C. watersi, found that despite a slight reduction in winter growth rate, A. tenuis did not present any growth seasonality, whereas the deeper dweller, C. watersi, presented marked growth seasonality. It was not possible to assess the seasonality in N. flagellata; however, it was hypothesised that it may grow continuously year-round because it has ecological similarities with A. tenuis (Barnes 1995). It was also hypothesised that the maximum lifespan attained by Antarctic bryozoans maybe limited by iceberg scouring/damage (Barnes 1995).
A collection of specimens of Melicerita obliqua (Thornely, 1924) (Fig. 2c) from the Weddell and Lazarus Seas revealed a lifespan of up to 25 years (Brey et al. 1998). Seasonal δ18O and δ18C ratios were also analysed and it was determined that the segment nodes were formed during periods of high δ13C values (i.e. high primary production) whereas the segments themselves were formed during periods of low δ13C values (i.e. low primary production). Further work on M. obliqua from the Weddell Sea determined a maximum age of about 45 years for a colony using node counting methods (Bader and Schäfer 2004). In addition, the authors reported an ontogenetic trend in the length of the colony segments (i.e. decreasing length of colony segments with increasing ontogenetic age), a common finding for all long-lived species studied in sclerochronology.
Further work on the Antarctic bryozoan, Cellaria incula Hayward and Ryland, 1993, found that colonies may live at least up to 14 years and have seasonal variability in δ18O and δ13C (Brey et al. 1999). The authors determined that growth in C. incula occurred during periods of low primary production, reflected in the δ13C ratios, and associated this with the selection of nanoplankton as primary food source. It was also reported that even though most of branch segments were annually formed, some segments can form over a period of 2 years (Brey et al. 1999). An ontogenetic study combined with carbonate production assessment of three erect rigid Antarctic bryozoans (i.e. Cellarinella nutti Rogick, 1956, C. nodulata Waters, 1904 and Swanomia belgica Hayward and Ryland, 1993) demonstrated the low annual calcification rates (compared to temperate species) for these species and provided an estimate lifespan for these species of 11–14, 11–18 and 23 years respectively (Smith 2007). Similarly the growth of C. nutti collected from the Weddell Sea was studied by Barnes et al. (2006), who found that calendar year was a highly significant factor when considering growth (irrespective of colony age or sampling location). The authors also provide a detail description of colony growth and calcification rates and also explored the relationship between ENSO and C. nutti growth, although no obvious relationship was found (Barnes et al. 2006). It was also suggested that zooid counting can be a realisable method to assess growth compared to other commonly used techniques such as segment length measurements (Barnes et al. 2006). Studies such as these constitute the basis for improved estimations of carbon sequestration in marine ecosystems, an essential component for more tightly constrained Earth system models. A collection of growth data from six Antarctic bryozoan species (Cellarinella margueritae Rogick, 1956, C. nodulata, C. rogickae Moyano, 1965, C. watersi, M. obliqua and Stomhypselosaria watersi Hayward and Thorpe, 1989) was used to constrain growth variability and assess environmental drivers of growth considering species, genus, age and calendar year (Barnes et al. 2007). When considering environmental drivers of colony growth, the authors concluded in this key study that even though there is interannual variability present in the segment growth series, this seems to be species-specific. The authors attribute this to a disparity in the effect of climatic trends for the region amongst the different species studied (Barnes et al. 2007).
Soft and hard Corals
Coral research occupies a prominent place in sclerochronological investigations in temperate and tropical environments. Zooxanthellate corals are constrained by latitude and depth due to the need to maintain the symbiotic zooxanthelle algae. However, azooxanthellate corals do not have these limitations and can be found at depth and in high latitudes (Stanley and Cairns 1988). In addition to this, alcyonean corals (i.e. soft corals) are more common in the Southern Ocean than scleractinian corals (i.e. stony corals). Because the development of sclerochronological records requires the preservation of the record in the hard structures of the living organisms; thus, scleractinian corals have been traditionally the main focus of the sclerochrological research community. Species diversity of scleractinian corals in the Southern Ocean is very low, fewer than ten species; this is assumed to be accurately estimated and not simply due to a lack of knowledge (Guinotte et al. 2006). This reason has most likely limited, at least in part, the development of coral-based sclerochronology in the Southern Ocean. On the other hand, alcyonean corals are more common in the Southern Ocean than scleractinian corals; however, they remain one of the least studied taxa of benthic Antarctic ecosystems. The scleractian coral Flabellum (Flabellum) impensum Squires, 1962 has attracted a degree of research attention. A metabolic study in combination with external mark of F. (Flabellum) impensum estimated that colonies of about 80 mm could attain a lifespan around 80 years (Henry and Torres 2013). Sclerochronological studies based on alcyonean corals are possible as some species have calcified skeletons that grow incrementally with colony age. A comparative study of two age determination methodologies: radiometric tracing (14C and 210Pb) and ring counting at the basal stem (using stereoscopic microscopy and SEM) from three Antarctic alcyonians (Fannyella (Scyphogorgia) abies (Broch, 1965), Fannyella. (Fannyella) rossii Gray, 1872 and Thouarella (Thouarella) variabilis Wright and Studer, 1889) showed inconsistencies in the number of increments in the basal stem between stereoscopic and SEM analysis (Martinez-Dios et al. 2016). Furthermore, the comparison between radiometric tracing and ring counting revealed that the growth increment periodicity was between 1 and 3 years; which resulted in colony maximum estimated lifespans over three centuries for F. (Scyphogorgia) abies and F. (Fannyella) rossii and over a millennium for T. (Thouarella) variabilis.
Neodymium (Nd) isotopes and 14C have been investigated in deep-sea stony corals, Desmophyllum dianthus (Esper, 1794), collected in the Drake Passage to constrain ocean circulation during the last Heinrich event (H1, 16.7 ka) (Robinson and Flierdt 2009). The authors interpret the data to indicate a twofold reduction (compared to current conditions) in the export of North Atlantic Deep Water (NADW) during H1, considered locally as the NADW component to the Antarctic Circumpolar Current. Investigation of Nd isotopes from fossil deep-sea corals [i.e. D. dianthus, Caryophyllia spp., Paraconotrochus antarcticus (Gardiner, 1929)] from the Drake Passage has been expanded more recently in a study focussing on sea ice control of ocean circulation during the Last Glacial Maximum (LGM, Wilson et al. 2020). Changes in Antarctic sea ice appeared to have an impact in the North Atlantic Deep Water influence in the Southern Ocean during the LGM with implications to deep water formation and consequently oceanic carbon storage (Wilson et al. 2020). Further work on Nd isotopes based on Antarctic scleractinian corals D. dianthus, Balanophyllia (Balanophyllia) malouinensis Squires, 1961, and Flabellum (Flabellum) curvatum Moseley, 1880 collected from the Drake Passage (Struve et al. 2017). The authors carried out a calibration between seawater samples, live and dead-collected material and concluded that D. dianthus and F. (Flabellum) curvatum could be used as marine Nd archives; however, B. (Balanophyllia) malouinensis showed significant differences between seawater and skeletal-derived Nd values and therefore, Nd interpretations based on this species must be done with caution. Furthermore, this study also confirms that aragonite corals present elevated Nd concentrations from those expected from inorganic precipitation experiments, although this finding, in the opinion of the authors, does not compromise the use of aragonitic corals as palaeo Nd archives (Struve et al. 2017). Radiocarbon analysis has been a major tool in the repertoire of the sclerochronology community since its development. It is commonly used in dead-collected material to range find a time frame when the animals were living, determine the marine 14C reservoir age (associated to changes in water masses; Berkman and Forman 1996). Deep-sea stony corals, collected from the Drake Passage, were used to develop a novel ‘reconnaissance’ method for percent-level 14C/12C analysis (Burke et al. 2010). The authors point out how 14C analysis coupled with U-Th analysis from deep-sea corals in the region provides valuable information about the reservoir age of the Southern Ocean. They also used 14C ages to assess the temporal and spatial distribution of the studied species and determined that this was not constant for the time frame considered (i.e. 0–40 ky; Burke et al. 2010). The work done by Burke et al. (2010) was latterly expanded by Margolin et al. (2014), who investigated in detail the distribution of fossil (i.e. as in dead-collected) scleractinian corals across the Drake Passage using radiocarbon and U-Th dating. This study highlighted the transient nature of scleractinian coral species distribution in the Drake Passage and although the environmental drivers controlling their distribution over millennial timescales are still not well understood, it provides some insights of the distribution of some scleractinian coral species over the last 35,000 years (Margolin et al. 2014). Investigating taxa as new palaeoenvironmental archive is always an arduous task that involves many calibration studies. An early study of the geochemical (δ18O and δ13C ratios) composition of deep-sea corals, which considered two Antarctic scleractinian corals (i.e. F. (Flabellum) impensum and P. antarcticus, previously known as Gardineria antarctica) provided some insight into the formation (i.e. in equilibrium or fractionated) of their skeletons (Smith et al. 2000). This study highlighted a significant disequilibria in the δ18O and δ13C ratios for all studied species but despite of this it was possible to obtain a method to calculate palaeotemperatures (Smith et al. 2000). Coral Li/Mg temperature proxy was assessed considering tropical and polar species (i.e. Javania antarctica (Gravier, 1914), F. (Flabellum) impensum, F. (Flabellum) gardineri, P. antarcticus and Caryophyllia (Caryophyllia) antarctica Marenzeller, 1904) (Cuny-Guirriec et al. 2019). The results of this study showed some limitations of Li/Mg proxy thermometry, and specifically related to the Antarctic species, due to the presence of calcite deposits (in: P. antarcticus, F. (Flabellum) impensum, F. (Flabellum) gardineri) within the aragonite skeleton (Cuny-Guirriec et al. 2019). Despite of these calcite deposits the authors suggest a method to correct their effect on coral Li/Mg thermometry using mRaman, optical microscopy and targeted laser ablation ICP-MS; although, the authors suggest that more research needs to be done assessing the impact of organic-bound Mg contamination in this analysis (Cuny-Guirriec et al. 2019).
Samperiz et al. (2020) have done a cutting edge geochemical (i.e. stable oxygen and carbon isotopes) calibration study of stylasterid corals. Stylasterid corals are a widely distributed deep-sea corals with calcite, aragonite or a combination of both skeletons (Samperiz et al. 2020). The authors found internal variability in δ18O and δ13C ratios depending on the sampling location within the coral (i.e. main truck, branch tip, secondary branch), with lower values always occurring at the tips. In addition, calcitic species showed larger departures from seawater equilibrium and therefore was suggested that aragonite-calcifying species could be more apt as seawater temperature proxies (Samperiz et al. 2020). However, no age estimates are available for many stylasterid species, to date there is none available for those found in Antarctic waters, which potentially could limit their use as palaeoenvironmental proxies. Another study of stylasterid corals explored the radiocarbon (C14) variability along sequential sampling of the skeleton (i.e. from tip to bottom), of specimens collected from the Ross Sea, off Wilkes Land and Weddell Sea (King et al. 2018). The authors found a reversal in radiocarbon age with ontogenetic age of the coral, which they attributed to major changes in the oceanographic regime of the Southern Ocean (increased influence of upwelling of Circumpolar Deep Water, CDW) starting around the Little Ice Age event. Determining the time of the onset of the current Antarctic melting, which has been mediated greatly by incursions of the CDW (e.g. Schmidtko et al. 2014), has been the focus of much research due to the impacts in the global climate and the regional ecosystems (e.g. Morley et al. 2020). Thus, the importance and relevance of studies such as the one done by King et al. (2018) on stylasterid corals.
Current challenges
The studies presented in this review have revealed some common issues highlighted across different taxa. Some of these challenges are difficult or even impossible to tackle, whereas others offer opportunities for sclerochronological research in the Southern Ocean.
Collection of marine animal samples for research is economically costly and often logistically complicated. Working in Antarctic marine environments complicates any collection even further due to the remoteness and extreme climatological conditions. In addition, Antarctic calcifying organisms are notorious for low calcification rates (Fabry et al. 2009; Watson et al. 2017), which makes them more fragile to sampling methodologies such as benthic trawling (i.e. significant physical damage can occur within the cod-end net by crushing against other material collected). In theory the use of sampling grabs such as: Van Veen grab, box corers or Hamon grab could collect delicate material without damaging it in the process; however, sampling surface of these pieces of equipment is small and many deployments would be needed. Furthermore, Antarctic shelf and coastal environments are significantly deeper than other parts of the world where sclerochronology has flourished. Collection of organisms by scuba divers has been undertaken from many Antarctic bases and have provided excellent specimens used in sclerochronology; however, this technique has its own drawbacks. Closeness to decompression chambers, maximum diving depth (approximately 30 m), weather conditions and exclusive summer sampling are some of the limiting factors for scuba diving sampling. Developing appropriate sampling methodologies allows the collection of these deep and fragile specimens and also allow an estimation of population densities, is essential for the development of Antarctic ecology and sclerochronology.
Sampling seasonality, biased towards the austral spring/summer, is a predominant characteristic of Antarctic research, irrespective of the specific scientific discipline. This is due mainly to operational season for most Antarctic research programmes as many do not carry out sampling over the austral winter. Sclerochronology could be a priori less affected by this bias if the organism grows continuously throughout the year. The research shows that some Antarctic species can grow continuously throughout the year, albeit at a much reduced rate during the winter. Analysing the winter-formed portions of the hard skeletal parts of the organisms discussed here will be challenging due to the small size of many of these records or may prove impossible due to technical sampling capabilities. With most of the hard skeletal parts being formed over the main growing season during the austral summer and spring, during the austral spring/summer seasons, there will always be an inherent skewed seasonality in sclerochronological proxy records. This characteristic drives the imperative to determine for each species the growing season (period) encapsulated in the hard skeletal parts. This process is critical if subsequent environmental proxies are to be developed from the information contained in the growth increment series.
Aragonite is a calcium carbonate form used by many marine organisms (e.g. many marine bivalves species, some stylasterid and scleractinian corals, pteropods) to construct hard skeletal structures. Aragonite saturation horizon (ASH) is the water depth which demarcates saturated from unsaturated aragonite waters. Above ASH the water is supersaturated with aragonite which allows its precipitation and below ASH the water is undersaturated in aragonite which favours aragonite dissolution. As atmospheric CO2 concentrations increase mean pH level will decrease (become more acidic) and it is predicted that ASH will become shallower globally. However, this phenomenon is expected to disproportionately affect high latitude seas, predicted to reach surface level by 2060 for most parts of the Southern Ocean (e.g. Guinotte et al. 2006; Jones et al. 2017). This poses a direct and significant threat for many marine calcifying organisms, many of which are light calcifiers (Fabry et al. 2009). Based on these predictions, it is likely that preservation of fossil/subfossil carbonate material cannot be taken for granted (McClintock et al. 2009). In addition, it is also likely that there will be a race against chemical dissolution for many species to reach reproductive maturity in order to maintain viable populations.
The lack of species of known extreme longevity, such as Arctica islandica (Linnaeus, 1767) (more than 500 years; Butler et al. 2013) or Glycymeris glycymeris (Linnaeus, 1758) (close to 200 years; Reynolds et al. 2013) from Northern Hemisphere temperate waters, in the Antarctic fauna and the deterioration of fossilised material (i.e. physical breakage or chemical dissolution, see McClintock et al. 2009) may well limit the use of Antarctic marine organisms as long-term scleroclimatological proxies. Based on the current state of knowledge it is difficult to envisage the construction of long-term sclerochronologies spanning more than a thousand years like those created in the northern hemisphere (see Butler et al. 2013) or even more than a couple centuries. However, even if the temporal frame of study is limited to the past 100–200 years, when all the instrumental data were collected, there is still a wealth information potentially available, especially considering the short length and the spatial homogeneity of instrumental records for the Southern Ocean. However, even if it will not be possible to construct annually resolved multicentennial long chronologies from Antarctic molluscs other taxa were found to present extended lifespans, for example alcyonean corals (see Table 1). In addition, sclerochronological research of Antarctic stylasterid corals has developed very recently and perhaps some of these species could provide environmental proxy data spanning over several centuries.
The introduction of possible biases into any sclerochronological records is always a concern for the research community. One avenue of introducing biases is the misidentification, by adding or subtracting, increments. For example, as mentioned in the context of the Antarctic bryozoan C. incula, some branch segments can be formed over a period of 2 years (Brey et al. 1999). Clear and rigorous methodological protocols need to be established, sometimes species-specific, for collecting, analysing and sampling carbonate material from Antarctic species.
For example, the study of the Antarctic bivalve A. colbecki shell showed a significant diagenetic modification of organic compounds in the shell matrix (Pérez-Huerta et al. 2020), which could potentially limit the use of proxy data based on Mg/Ca rations and δ15N. This study highlights the importance of understanding properties of organic compounds in the hard structures used as proxy environmental recorders.
Despite all these challenges and most likely others not covered in this review but known experts in the different disciplines, sclerochronology has contributed to Antarctic research providing a wealth of information. In addition, there are research opportunities associated with these challenges, some of which will be discussed in the next section, and hopefully will start a discussion within Antarctic sclerochronology to push our understanding of the Southern Ocean and its species further.
Scope for the future
Sclerochronology has been contributing to a variety of disciplines in Antarctic research for almost five decades. Despite this, there are still significant knowledge gaps, which offer the basis for future Antarctic sclerochronology research. Sclerochronology can provide ecological and biological context for a significant amount of modern species, which would also be advantageous for the interpretation of information derived from fossil species.
There is a lack of investigations into whether animal populations are growing synchronously or whether growth is individualistic in Antarctic fauna. This applies to all Antarctic bryozoans, brachiopods, cephalopods, bony fish and many bivalve and gastropod mollusc species. Sclerochronology can provide much information about animal and population growth using already established techniques for many marine taxa. Understanding how individuals and populations grow is an essential prerequisite in order to build chronologies that may offer an insights into long-term changes in the environment.
Antarctic bivalve molluscs are one of the taxa that has attracted much of the sclerochronological research in the Southern Ocean (Table 1). Most of the research has used live-collected material for analysis as in comparison with studies of temperate species from the Northern Hemisphere where the use of life and dead-collected material is common. Dead-collected material, once dated and crossmatched, allows the constructions extended master chronologies that have the potential to extend into the past beyond the onset of the instrumental records (e.g. Butler et al. 2010, 2013; Reynolds et al. 2013). The collection of Antarctic bivalve fossil material has perhaps been overlooked or limited due to intense reworking of Antarctic marine sediments by ice scouring (e.g. Sahade et al. 2015) or by chemical dissolution due to ocean acidification (e.g. McClintock et al. 2009; Fabry et al. 2009; Jones et al. 2017;). This is a necessary step, if sclerochronology wants to provide palaeoenvironmental information beyond the instrumental record. Thus the collection and preservation of fossil material should be considered in future Antarctic sampling campaigns.
There are significant knowledge gaps in the ecology, ontogeny and biology of many Antarctic taxa. Very few sclerochronological studies are available for some groups (e.g. brachiopods, echinoderms), whereas others have more studies (e.g. bivalve molluscs) and others show promising results (e.g. bryozoans, cephalopods The literature on sclerochronological studies in many research fields based on scleractinian corals is abundant for tropical and temperate species. Several scleractinian coral species are known to live in the Southern Ocean (e.g. Cairns 1983; Schejter et al. 2016), although little is known for many species regarding lifespan, distribution and their possible uses as palaeoenvironmental archives. Proteinaceous corals are a varied group of species which include among others gorgonians and black corals or antipatharians and which are showing promising results as palaeoenvironmental archives (e.g. Williams 2020). Some of these species among other scleractian corals have been reported in Antarctic waters (Cairns 1983). However, the literature of Antarctic corals is reduced or completely non-existent for certain tax (e.g. antipatharia or black corals). Species such as the black coral, Bathypathes patula Brook, 1889, which found in the Southern Ocean, have shown promising although challenging results for sclerochronology in individuals collected in New Zealand waters (Marriot et al. 2020). To date no studies of B. patula are available for specimens collected in the Southern Ocean. Comparative studies across a latitudinal gradient of species such as B. patula may offer new insights into the biology of the species and may provide a significant amount of information as environmental biorecorders. Some studies discussed in this review have researched the biochemical composition of coral skeletons (e.g. neodymium isotopes, Li/Mg ratios). However, the samples are usually analysed in bulk, meaning that the growth increment pattern was not considered when sampling. Developing annually resolved geochemical series from coral skeletons which possess annual growth increments has the potential to provide valuable palaeoenvironmental information for deep-sea environments (e.g. Kimball et al. 2014). There have been also some advances developing other geochemical analytical techniques such as clumped isotopes in deep-sea corals; however, further investigations are needed to asses possible vital effects and isotopic fractionation (e.g. Kimball et al. 2016; Spooner et al. 2016).
Stylasterid corals have shown promising results as new palaeoenvironmental archives (Samperiz et al. 2020) although there is plenty of research to be done for many species. No age determination is available for any Antarctic stylasterids nor detailed studies of the micro-skeletal structure. The combination of an understanding of the internal micro-structure and detailed geochemical analysis have the potential to provide much-needed palaeoenvironmental proxy records for understudied parts of the ocean such as the deep-sea. Research of Antarctic gastropod molluscs could be extended greatly using the great body of knowledge available from studies of temperate species.
This review has highlighted that to date sclerochronological work based on Antarctic gastropods is limited. A review on sclerochronological techniques for age determination and growth assessment was done by Hollyman et al. (2018a, b). The authors provide a discussion of different methods and their limitations that could be of great use for those researchers who want to embark on the study of Antarctic gastropod molluscs. The study of gastropod shells and statoliths have been investigated to a greater extent in temperate environments. For example, the study of the growth increment patterns, and geochemical composition of gastropod shells have been shown to be reliable high-resolution proxies for seawater temperature (e.g. Fenger et al. 2007; Schöne et al. 2007) and have provide accurate scientific data for stock assessment of commercial species (Hollyman et al. 2018a, b). In principle there is no impediment to extend these lines of work to Antarctic research (see Fig. 2b) and expand it further. Strong salinity gradients due mainly to sea ice formation and ice melting will still be a factor to consider for studies tackling the development of sea surface proxy records but this should not discourage such studies. To date there is no study of Antarctic gastropod statoliths in the Southern Ocean. Researchers interested in this avenue of research will likely find much historically collected material available at museum and Antarctic research institutes around the world that would provide sufficient samples to explore.
Sclerochronological research of cephalopod beaks has a great potential to offer a significant insight into the biology and ecology of many species and provide novel proxy records for environmental monitoring. For example, it is still unknown, or underreported, whether there are any rhythmic growth patterns in the beaks of Antarctic octopods. Further investigation on Antarctic cephalopod beaks could provide information about lifespan and population structure for some of these species. In addition, geochemical analysis (e.g. δ15N and δ13C) of their beaks coupled with a close understanding of the ontogeny of the species (e.g. sampling along known age beak rings) can provide high resolution life history of Antarctic octopod ecology. Diagenetic alteration of organic compounds of the beaks remains unexplored in Antarctic octopods and could have significant impact in the use of δ15N as seen in bivalve species (Pérez-Huerta et al. 2020).
The Antarctic sclerochronological literature based on teleost fish is extensive. Many of the studies reported in this review have provided a much-needed insight into basic biological traits, population structure and distribution, ontogeny for many species. Otolith ring counting (done mainly using light microscopy) seem to be the most used analytical technique. This technique provides valuable ontogenetic information at a relatively low cost. However, for many species there has been no independent age determination methodology applied (e.g. otolith makers like oxytetracycline, sequential seasonal sampling, geochemical analysis). The use of independent age determination methodologies is necessary to validate much of the work done to date with Antarctic teleost fish. In addition, for some species (Protomyctophum spp.; see Saunders et al. 2015) age determination was carried out by length-frequency analysis and no sclerochronological work is available on the scientific literature. These techniques will need to be applied almost on a species-by-species basis since not all techniques will be suitable for all species (e.g. tagging and recapture technique with deep fish species may prove nearly impossible). In addition, the development of geochemical data series (either trace metals or stable isotope ratios) from known age / calendar years (based on otolith ring analysis) has been relatively unused in Antarctic species. The development of geochemical series could provide a wealth of information for many of the species, their role in the ecosystem and of the environment they inhabit during their lifetime. Antarctic teleost fish fisheries are attracting a growing amount of commercial interests (e.g. Brooks et al. 2018). It will be necessary to understand for the commercially targeted species basic biological information (e.g. lifespan, growth rates), population structure and resilience to environmental factors such as climate change (e.g. Brooks et al. 2018; Rayfuse 2018; Wendebourg 2020) so we do not repeat mistakes from the past and assure the ecosystem preservation and a sustainable exploitation. Sclerochronology will undoubtedly play a central role in providing quality scientific information for the Antarctic fisheries management bodies.
Ophiuroids are an important part of Antarctic benthic ecosystems and are likely a significant component contributing to carbon capture at high latitudes (Barnes et al. 2020). Although the use of sclerochronological techniques in ophiuroids have been stablished and applied elsewhere in the world (e.g. Gage 1990; Ravelo et al. 2017), very few studies have been done on Antarctic species. Antarctic ophiuroids can offer a great research opportunity contributing to fields such as: biology, ecology and climatology.
Geochemical analysis, especially stable oxygen and carbon isotopes have played a major role in sclerochronological analysis. However, despite of this there are still major gaps in the understanding of geochemical variability, and not only for Antarctic species, within the carbonate skeletal parts on many species. Work done by Barrera et al. (1990, 1994) have shown the importance of this in A. colbecki and L. elliptica and it should be expanded to as many species as possible. These studies would be of great importance in assessing the effect of sampling location for geochemical analysis (e.g. sectioning transect located on the centre of a valve or positioned towards the anterior or posterior parts of the valve). In addition, it is currently not considered how the different shell layers vary laterally along the anterior–posterior axis. Varying thickness of the different shell layers is especially relevant when shells need to be drilled on its external surface due to limitations of shell thickness (see Roman-Gonzalez 2018). Studying the anterior–posterior variability in the thickness of shell layers should be a relatively straight-forward analysis, involving sequential cuts along the dorso-ventral margin and the comparison of polished sections. The development of long-term series of geochemical proxies (stable isotopes and trace metals) from the carbonate structures from erect Antarctic bryozoans is another example that could bring extra level of detail into the ontogeny and the habitats these Antarctic species inhabit.
Many species considered in this review (e.g. bivalves, gastropods, brachiopods) are known to live under the seasonal winter ice in littoral Antarctic regions. The proxy environmental data extracted from such specimens could provide invaluable data of oceanic processes occurring during the winter, which have been very limited to date due to obvious technical difficulties of working in Antarctica during the winter season. In addition, they can provide as well valuable biological information about physiological processes that these organisms undergo through the winter.
The research community may have been discouraged by the shorter lifespan of many of the Antarctic species investigated compared to the significantly extended lifespan of some temperate Northern Hemisphere species such A. islandica or G. glycymeris, especially if research aims are to establish long-term records that extend beyond the onset of instrumental data series collection. Lifespans of many Antarctic species with calcareous skeletons are still unknown (see Table 1) but some corals present promising results. However, prior to the development of any environmental proxy based on Antarctic marine species, it is necessary to fully comprehend the environmental and biological parameters driving growth. There is an opportunity here for the sclerochronological community to bridge this gap and contribute to the broader Antarctic science community in multidisciplinary programmes.
Antarctic marine ecosystems are complex and they are tightly coupled to the highly seasonal physical environment. Current climatic trends are expected to have strong impacts over multiple ecosystem levels (e.g. Constable et al. 2014); thus, understanding the processes, feedbacks and drivers is essential to better forecast and protect Antarctic marine ecosystems. Much of the research done to date has focussed on understanding single species biological traits and using the information extracted from the calcareous hard bodily structure to develop single environmental proxies that are commonly limited to specific locations. To date chronologies constructed based on Antarctic marine organisms have also been species-specific and provide short snapshots of the environment from single locations. A true network of chronologies developed from either the same species or from a range of species could in theory provide some insight into the regional variability of the climate system (e.g. Reynolds et al. 2017). Sclerochronology in Antarctic science and in polar research in general can offer much more from insights into ecosystem functioning and climate-ecosystem interactions when there is a critical mass of well-developed records (e.g. Black et al. 2014), has the potential to provide quality data for climate models to understand long-term (potentially spanning millennia) variability in the Earth and ocean system (e.g. Halloran et al. 2020). Multidisciplinary analyses are key in tackling ecosystem functioning and drawing expertise from several science disciplines. Discovery of species which grow all year-round would be a very valuable contribution to Antarctic sclerochronology. Such species could provide much-needed climatological data for the austral winter, which in turn could be used in other science disciplines.
Retreating ice shelves and marine terminating glaciers along the coast of West Antarctica are having profound effects on marine ecosystems via complex interactions (e.g. mixed layer depth, seawater temperature and salinity, primary production, iceberg scouring, seabed variability) (e.g. Sahade et al. 2015). As these ice systems peel back further, new seabed is becoming available for colonization by marine organisms (e.g. Lagger et al. 2017), many of which are calcifying species. These newly developed habitats and communities act as atmospheric carbon sinks (i.e. blue carbon) and carbon storage and thus will be one of the few mitigating feedbacks of climate change in the Antarctic (e.g. Barnes et al. 2020). Some species discussed in this review present extended lifespans (e.g. soft corals) and their calcareous skeletons can potentially act as long-term carbon storage component (e.g. Coppari et al. 2019). Marine calcifying organisms play an important role in the development of these new ecosystems and sclerochronology can inform about the paucity, environmental conditions and rhythms of these newly formed marine ecosystems.
Sclerochronology in the Southern Ocean has expanded in recent decades the knowledge on how marine calcifying organisms grow and what environmental drivers are responsible for such growth patterns. Many of these studies are species-specific and there are still major knowledge gaps for many taxa. For many species there is a clear need for technical studies that independently validate and standardize age determination methods in order to provide sound sclerochronological records. A great example of this need was highlighted by Cronin et al. (2020) for the Antarctic bivalve A. colbecki, as depending on the analytical technique used age estimation differs in one order of magnitude (i.e. from around 20 year to 100+ years). Despite this, there is a challenge for the community to refocus this science from species-specific analysis to a better understanding of ecosystem functioning and feedbacks between the ecosystems, habitats and climate forcings.
References
Alves EQ, Macario K, Ascough P, Ramsey B (2018) The worldwide marine radiocarbon reservoir effect: definitions, mechanisms, and prospects. Rev Geophys 56:278–305. https://doi.org/10.1002/2017RG000588
Ashford JR, Jones CM, Hofmann EE, Everson I, Moreno CA, Duhamel G, Williams R (2008) Otolith chemistry indicates population structuring by the Antarctic Circumpolar current. Can J Fish Aquat Sci 65:135–146. https://doi.org/10.1139/f07-158
Ashford J, Dinniman M, Brooks C, Andrews AH, Hoffmann E, Cailliet G, Jones C, Ramanna N (2012) Does large-scale ocean circulation structure life history connectivity in Antarctic toothfish (Dissostichus mawsoni)? Can J Fish Aquat Sci 69:1903–1919. https://doi.org/10.1139/f2012-111
Bader B, Schäfer P (2004) Skeletal morphogenesis and growth check lines in the Antarctic bryozoan Melicerita obliqua. J Nat Hist 38:2901–2922. https://doi.org/10.1080/00222930310001657685
Barnes DKA (1995) Seasonal and annual growth in erect species of Antarctic bryozoans. J Exp Mar Biol Ecol 188:181–198. https://doi.org/10.1016/0022-0981(95)00003-A
Barnes DKA, Clarke A (1994) Seasonal variation in the feeding activity of four species of Antarctic bryozoan in relation to environmental factors. J Exp Mar Biol Ecol 181:117–133. https://doi.org/10.1016/0022-0981(94)90108-2
Barnes DKA, Peck LS (1997) An Antarctic shelf population of the deep-sea, Pacific brachiopod Neorhynchia strebeli. J Mar Biol Assoc 77:399–407. https://doi.org/10.1017/S0025315400071757
Barnes DKA, Webb K, Linse K (2006) Slow growth of Antarctic bryozoans increases over 20 years and is anomalously high in 2003. Mar Ecol Prog Ser 314:187–195. https://doi.org/10.3354/meps314187
Barnes DKA, Webb KE, Linse K (2007) Growth rate and its variability in erect Antarctic bryozoans. Polar Biol 30:1069–1081. https://doi.org/10.1007/s00300-007-0266-2
Barnes DKA, Sands CJ, Cook A, Howard F, Roman-Gonzalez A, Muñoz-Ramirez C, Retallick K, Scourse J, Van Landeghem K, Zwerschke N (2020) Blue carbon gains from glacial retreat along Antarctic fjords: what should we expect? Glob Change Biol 26:2750–2755. https://doi.org/10.1111/gcb.15055
Barratt IM, Allcock AL (2010) Ageing octopods from stylets: development of a technique for permanent preparations. ICES J Mar Sci 67:1452–1457. https://doi.org/10.1093/icesjms/fsq047
Barrera E, Tevesz MJS, Carter JG (1990) Variations in oxygen and carbon isotopic compositions and microstructure of the shell of Adamussium colbecki (Bivalvia). Palaios 5:149–159. https://doi.org/10.2307/3514811
Barrera-Oro ER, Casaux RJ (1992) Age estimation for juvenile Notothenia rossii from Potter Cove, South Shetland Islands. Antarct Sci 4:131–136. https://doi.org/10.1017/S095410209200021X
Barrera E, Tevesz MJS, Carter JG, McCall PL (1994) Oxygen and carbon isotopic composition and shell microstructure of the bivalve Laternula elliptica from Antarctica. Palaios 9:275–287. https://doi.org/10.2307/3515202
Bednaršek N, Tarling GA, Fielding S, Bakker DCE (2012) Population dynamics and biogeochemical significance of Limacina helicina antarctica in the Scotia Sea (Southern Ocean). Deep-Sea Res Part II 59-60:105–116. https://doi.org/10.1016/j.dsr2.2011.08.003
Berkman PA (1990) The Population Biology of the Antarctic Scallop, Adamussium colbecki (Smith 1902) at New Harbor, Ross Sea. In: Kerry KR, Hempel G (eds) Antarctic ecosystems. Springer, Berlin, pp 281–288. https://doi.org/10.1007/978-3-642-84074-6_32
Berkman PA (1994) Geochemical signatures of meltwater in mollusc shells from Antarctic coastal areas during the Holocene. Mem Natl Inst Polar Res 50:11–33
Berkman PA, Forman SL (1996) Pre-bomb radiocarbon and the reservoir correction for calcareous marine species in the Southern Ocean. Geophys Res Lett 23:363–366. https://doi.org/10.1029/96GL00151
Bizikov VA (1991) A new method of squid age determination using the gladius. In: Jereb P, Ragonese S and von Boletzky S (eds) Proceeding of the International Workshop: vol Special Publication No. 1. Squid age determination using statoliths. Instituto di Tecnologia della Pesca e del Pescate, Italy, pp 39–51
Bizikov VA (1995) Growth of Sthenoteuthis oualaniensis, using a new method based on gladius microstructure. ICES J Mar Sci J Cons 199:455–458
Black BA, Sydeman WJ, Frank DC, Griffin D, Stahle DW, Garcia-Reyes M, Rykaczewski RR, Bogard SJ, Peterson WT (2014) Six centuries of variability and extremes in a coupled marine-terrestrial ecosystem. Science 345:1498–1502. https://doi.org/10.1126/science.1253209
Brand U, Azmy K, Bitner MA, Logan A, Zuschin M, Came R, Ruggiero E (2013) Oxygen isotopes and MgCO3 in brachiopod calcite and a new paleotemperature equation. Chem Geol 359:23–31. https://doi.org/10.1016/j.chemgeo.2013.09.014
Brey T, Hain S (1992) Growth, reproduction and production of Lissarca notorcadensis (Bivalvia Philobryidae) in the Weddell Sea, Antarctica. Mar Ecol Prog Ser 82:219–226. https://doi.org/10.3354/meps082219
Brey T, Mackensen A (1997) Stable isotopes prove shell growth bands in the Antarctic bivalve Laternula elliptica to be formed annually. Polar Biol 17:465–468
Brey T, Pearse J, Basch L, McClintock J, Slattery M (1995a) Growth and production of Sterechinus neumayeri (Echinoidea: Echinodermata) in McMurdo Sound, Antarctica. Mar Biol 124:279–292. https://doi.org/10.1007/BF00347132
Brey T, Peck LS, Gutt J, Hain S, Arntz WE (1995b) Population dynamics of Magellania Fragilis, a Brachiopod dominating a mixed-bottom macrobenthic assemblage on the Antarctic shelf. J Mar Biol Assoc 75:857–869. https://doi.org/10.1017/S0025315400038200
Brey T, Gutt J, Mackensen A, Starmans A (1998) Growth and productivity of the high Antarctic Bryozoan Melicerita obliqua. Mar Biol 132:327–333. https://doi.org/10.1007/s002270050398
Brey T, Gerdes D, Gutt J, Mackensen A, Starmans A (1999) Growth and age of the Antarctic bryozoan Cellaria incula on the Weddell Sea shelf. Antarct Sci 11:408–414. https://doi.org/10.1017/S0954102099000516
Brey T, Voigt M, Jenkins K, Ahn I-Y (2011) The bivalve Laternula elliptica at King George Island—a biological recorder of climate forcing in the West Antarctic Peninsula region. J Mar Syst 88:542–552. https://doi.org/10.1016/j.jmarsys.2011.07.004
Briffa KR, Jones PD (1990) Basic chronology statistics and assessment. In: Cook ER, Kairiukstis LA (eds) Methods of dendrochronology—applications in the environmental sciences. Springer, Dordrecht, pp 137–152
Brooks CM, Ainley DG, Abrams PA, Dayton PK, Hofman RJ, Jacquet J, Siniff DB (2018) Antarctic fisheries: factor climate change into their management. Nature 558:177–180. https://doi.org/10.1038/d41586-018-05372-x
Buick DP, Ivany LC (2004) 100 years in the dark: extreme longevity of Eocene bivalves from Antarctica. Geology 32:921–924. https://doi.org/10.1130/G20796.1
Burgess SN, Henderson GM, Hall BL (2010) Reconstructing Holocene conditions under the McMurdo Ice Shelf using Antarctic barnacle shells. Earth Planet Sci Lett 298:385–393. https://doi.org/10.1016/j.epsl.2010.08.015
Burke A, Robinson LF, McNichol AP, Jenkins WJ, Scanlon KM, Gerlach DS (2010) Reconnaissance dating: a new radiocarbon method applied to assessing the temporal distribution of Southern Ocean deep-sea corals. Deep Sea Res Part I 57:1510–1520. https://doi.org/10.1016/j.dsr.2010.07.010
Butler PG, Schöne BR (2017) New research in the methods and applications of sclerochronology. Palaeogeogr Palaeoclimatol Palaeoecol 465:295–299. https://doi.org/10.1016/j.palaeo.2016.11.013
Butler PG, Richardson CA, Scourse JD, Wanamaker AD, Shammon TM, Bennell JD (2010) Marine climate in the Irish Sea: analysis of a 489-year marine master chronology derived from growth increments in the shell of the clam Arctica islandica. Q Sci Rev 29:1614–1632. https://doi.org/10.1016/j.quascirev.2009.07.010
Butler PG, Wanamaker AD, Scourse JD, Richardson CA, Reynolds DJ (2013) Variability of marine climate on the North Icelandic Shelf in a 1357-year proxy archive based on growth increments in the bivalve Arctica islandica. Palaeogeogr Palaeoclimatol Palaeoecol 373:141–151. https://doi.org/10.1016/j.palaeo.2012.01.016
Butler PG, Freitas PS, Burschell M, Chauvaud L (2019) Archaeology and Sclerochronology of Marine Bivalves. In: Small A et al (eds) Goods and services of marine bivalves. Springer, Charm, pp 413–444. https://doi.org/10.1007/978-3-319-96776-9_21
Cairns SD (1983) Antarctic and Subantarctic Scleractinia. Antarct Res Ser 34:1–74. https://doi.org/10.1029/AR034p0001
Cattaneo-Vietti R, Chiantore M, Albertelli G (1997) The population structure and ecology of the Antarctic scallop Adamussium colbecki (Smith, 1902) at Terra Nova Bay (Ross Sea, Antarctica). Sci Mar 61:15–24. https://doi.org/10.1007/978-3-642-59607-0_41
Clark GR (1974) Growth lines in invertebrate skeletons. Annu Revi Earth Planet Sci 2:77–99. https://doi.org/10.1146/annurev.ea.02.050174.000453
Clarke A, Prothero-Thomas E, Beaumont J, Chapman A, Brey T (2004) Growth in the limpet Nacella concinna from contrasting sites in Antarctica. Polar Biol 28:2–61. https://doi.org/10.1007/s00300-004-0647-8
Constable AJ, Melbourne-Thomas J, Corney SP, Arrigo KR, Barbaud C, Barnes D, Bindoff NL, Boyd PW et al (2014) Climate change and Southern Ocean ecosystems I: how changes in physical habitats directly affect marine biota. Glob Change Biol 20:3004–3025. https://doi.org/10.1111/gcb.12623
Coppari M, Zanella C, Rossi S (2019) The importance of coastal gorgonians in the blue carbon budget. Sci Rep 9:13550. https://doi.org/10.1038/s41598-019-49797-4
Cronin KE, Walker SE, Mann R, Chute AS, Chase Long M, Bowser SS (2020) Growth and longevity of the Antarctic scallop Adamussium colbecki under annual and multiannual sea ice. Antarct Sci 32:466–475. https://doi.org/10.1017/S0954102020000322
Cross EL, Peck LS, Harper EM (2015) Ocean acidification does not impact shell growth or repair of the Antarctic brachiopod Liothyrella uva (Broderip, 1833). J Exp Mar Biol Ecol 462:29–35. https://doi.org/10.1016/j.jembe.2014.10.013
Cuny-Guirriec K, Douville E, Reynaud S et al (2019) Coral Li/Mg thermometry: caveats and constraints. Chem Geol 523:162–178. https://doi.org/10.1016/j.chemgeo.2019.03.038
Daniels RA (1983) Demographic characteristics of an Antarctic plunderfish, Harpagifer bispinis antarcticus. Mar Ecol Progr Ser 13:181–187
Dahm C, Brey T (1998) Determination of growth and age of slow growing brittle stars (Echinodermata: Ophiuroidea) from natural growth bands. J Mar Biol Assoc 78:941–951. https://doi.org/10.1017/S0025315400044891
Dick D, Philipp E, Kriews M, Abele D (2007) Is the umbo matrix of bivalve shells (Laternula elliptica) a climate archive? Aquat Toxicol 84:450–456. https://doi.org/10.1016/j.aquatox.2007.07.005
Duan M, Ashford JR, Bestly S, Wei X, Walters A, Zhu G (2020) Otolith chemistry of Electrona antarctica suggests a potential population marker distinguishing the southern Kerguelen Plateau from the eastward-flowing Antarctic Circumpolar current. Limnol Oceanogr 9999:1–17. https://doi.org/10.1002/lno.11612
Eastman JT (1993) Antarctic fish biology: evolution in a unique environment. Academic Press, San Diego
Esper J, Cook ER, Krusic PJ, Peters K, Schweingruber FH (2003) Tests of the RCS method for preserving low-frequency variability in long tree-ring chronologies. Tree-Ring Res 59:81–98
Fabry V, McClintock J, Mathis J, Grebmeier J (2009) Ocean acidification at high latitudes: the bellwether. Oceanography 22:160–171. https://doi.org/10.5670/oceanog.2009.105
Fenger T, Surge D, Schöne B, Milner N (2007) Sclerochronology and geochemical variation in limpet shells (Patella vulgata): a new archive to reconstruct coastal sea surface temperature. Geochem Geophys Geosyst 8:Q07001. https://doi.org/10.1029/2006GC001488
Filippova JA (2002) Review of Soviet/Russian studies on squids in the Antarctic Ocean. Bull Mar Sci 71:255–267
Gage JD (1990) Skeletal growth bands in brittle stars: microstructure and significance as age markers. J Mar Biol Assoc 70:209–224. https://doi.org/10.1017/S0025315400034329
Gaspard D, Aldridge AE, Boudouma O, Fialin M, Rividi N, Lécuyer C (2018) Analysis of growth and form in Aerothyris kerguelenensis (rhynchonelliform brachiopod)—shell spiral deviations, microstructure, trace element contents and stable isotope ratios. Chem Geol 483:474–490. https://doi.org/10.1016/j.chemgeo.2018.03.018
Gillikin DP (2019) Chemical sclerochronology. Chem Geol 526:1–6. https://doi.org/10.1016/j.chemgeo.2019.06.016
Goetz AJ, Griesshaber E, Neuser RD, Lüter C, Hühner M, Harper E, Schmahl WW (2009) Calcite morphology, texture and hardness in the distinct layers of rhynchonelliform brachiopod shells. Eur J Mineral 21:303–315. https://doi.org/10.1127/0935-1221/2009/0021-1922
Greely TM, Gartner JV Jr, Torres JJ (1999) Age and growth of Electrona antarctica (Pisces: Myctophidae), the dominant mesopelagic fish of the Southern Ocean. Mar Biol 133:145–158. https://doi.org/10.1007/s002270050453
Gröecke DR, Gillikin DP (2008) Advances in mollusc sclerochronology and sclerochemistry: tools for understanding climate and environment. Geology 28:265–268. https://doi.org/10.1007/s00367-008-0108-4
Guinotte JM, Orr J, Cairns S, Freiwald A, Morgan L, George R (2006) Will human-induced changes in seawater chemistry alter the distribution of deep-sea scleractinian corals? Front Ecol Environ 4:141–146. https://doi.org/10.1890/1540-9295(2006)004[0141:WHCISC]2.0.CO;2
Halloran PR, Hall IR, Menary M et al (2020) Natural drivers of multidecadal Arctic sea ice variability over the last millennium. Sci Rep 10:688. https://doi.org/10.1038/s41598-020-57472-2
Hanchet S, Sainsbury K, Butterworth D, Darby C, Bizikov V, Rune Godø O, Ichii T, Kock K-H, López Abellan L, Vacchi M (2015) CCAMLR’s precautionary approach to management focusing on Ross Sea toothfish fishery. Antarct Sci 27:333–340. https://doi.org/10.1017/S095410201400087X
Heilmayer O, Brey T, Chiantore M, Cattaneo-Vietti R, Arntz WE (2003) Age and productivity of the Antarctic scallop, Adamussium colbecki, in Terra Nova Bay (Ross Sea, Antarctica). J Exp Mar Biol Ecol 288:239–256. https://doi.org/10.1016/S0022-0981(03)00020-0
Henry LV, Torres JJ (2013) Metabolism of an Antarctic solitary coral, Flabellum impensum. J Exp Mar Biol Ecol 449:17–21. https://doi.org/10.1016/j.jembe.2013.08.010
Hernández-López JL, Castro-Hernández JJ, Hernández-García V (2001) Age determined from the daily deposition of concentric rings on common octopus (Octopus vulgaris) beaks. Fish Bull 99:679–684
Higgs ND, Reed AJ, Hooke R, Honey DJ, Heilmayer O, Thatje S (2009) Growth and reproduction in the Antarctic brooding bivalve Adacnarca nitens (Philobryidae) from the Ross Sea. Mar Biol 156:1073–1081. https://doi.org/10.1007/s00227-009-1154-9
Hollyman PR, Leng MJ, Chenery SRN, Laptikhovky VV, Richardson CA (2018a) Statoliths of the whelk Buccinum undatum: a novel age determination tool. Mar Ecol Prog Ser 598:261–272. https://doi.org/10.3354/meps12119
Hollyman PR, Laptikhovsky VV, Richardson CA (2018b) Techniques for estimating the age and growth of Molluscs: Gastropoda. J Shellfish Res 37:773–782. https://doi.org/10.2983/035.037.0408
Horn PL (2002) Age and growth of Patagonian toothfish (Dissostichus eleginoides) and Antarctic toothfish (D. mawsoni) in waters from the New Zealand Subantarctic to the Ross Sea. Antarctica Fish Res 56:275–287. https://doi.org/10.1016/S0165-7836(01)00325-3
Horn PL, Sutton CP, DeVries AL (2003) Evidence to support the annual formation of growth zones in otoliths of Antarctic toothfish (Dissostichus mawsoni). CCAMLR Sci 10:125–138
Hunt BPV, Pakhomov EA, Hosie GW, Siegel V, Ward P, Bernard K (2008) Pteropods in Southern Ocean ecosystems. Prog Oceanogr 78:193–221. https://doi.org/10.1016/j.pocean.2008.06.001
Hulley PA, Camus P, Duhamel G (1989) Ichthyological results of cruise MD-42/ SIBEX-II. Part 1. Fishes from RMT-8 stations, with additional records of lanternfishes (Myctophidae: osteichthyes) from the Indian and Atlantic sectors of the Southern Ocean. Cybium 13:83–99
Hunter E, Laptikhovsky VV, Hollyman PR (2018) Innovative use of sclerochronology in marine resource management. Mar Ecol Prog Ser 598:155–158
Ivany LC, Lohmann KC, Hasiuk F, Blake DB, Glass A, Aronson RB, Moody RM (2008) Eocene climate record of a high southern latitude continental shelf: Seymour Island, Antarctica. Geol Soc Am Bull 120:659–678. https://doi.org/10.1130/B26269.1
Ivany LC, Brey T, Huber M, Buick DP, Schöne BR (2011) El Niño in the Eocene greenhouse recorded by fossil bivalves and wood from Antarctica: EOCENE EL NIÑO. Geophys Res Lett 38:L16709. https://doi.org/10.1029/2011GL048635
Jackson GD, Lu CC (1994) Statolith microstructure of seven species of Antarctic squid captured in Prydz Bay, Antarctica. Antarct Sci 6:195–200. https://doi.org/10.1017/S0954102094000301
Jones EM, Fenton M, Meredith MP, Clargo NM, Ossebaar S, Ducklow HW, Venables HJ, de Barr HJW (2017) Ocean acidification and calcium carbonate saturation states in the coastal zone of the West Antarctic Peninsula. Deep-Sea Res Part II 139:181–194. https://doi.org/10.1016/j.dsr2.2017.01.007
Kimball JB, Dunbar RB, Guilderson TP (2014) Oxygen and carbon isotope fractionation in calcitic deep-sea corals: Implications for paleotemperature reconstruction. Chem Geol 381:223–233. https://doi.org/10.1016/j.chemgeo.2014.05.008
Kimball J, Eagle R, Dunbar R (2016) Carbonate “clumped” isotope signatures in aragonitic scleractinian and calcitic gorgonian deep-sea corals. Biogeosciences 13:6487–6505. https://doi.org/10.5194/bg-13-6487-2016
King TM, Rosenheim BE, Post AL, Gabris T, Burt T, Domack EW (2018) Large-scale intrusion of circumpolar deep water on antarctic margin recorded by stylasterid corals. Paleoceanogr Paleoclimatol 33:1306–1321. https://doi.org/10.1029/2018PA003439
Kompowski A, Rojas C (1994) On some biological characters of Parachaenichthys georgianus (Fischer, 1885) (Pisces, Notothenioidei, Bathydraconidae) from the shelf waters of South Georgia (Antarctica). Acta Ichthyol Piscat 24:61–75. https://doi.org/10.3750/AIP1994.24.2.08
La Mesa M, Vacchi M (2001) Age and growth of high Antarctic notothenioid fish. Antarct Sci 13:227–235. https://doi.org/10.1017/S0954102001000335
La Mesa M, Arneri E, Giannetti G (1996) Age and growth of the nototheniid fish Trematomus bernacchii Boulenger from Terra Nova Bay, Antarctica. Polar Biol 16:139–145. https://doi.org/10.1007/BF02390434
La Mesa M, Catalano B, Kock K-H, Jones CD (2012) Age and growth of the Antarctic dragonfish Parachaenichthys charcoti (Pisces, Bathydraconidae) from the southern Scotia Arc. Polar Biol 35:1545–1553. https://doi.org/10.1007/s00300-012-1194-3
La Mesa M, Calì F, Donato F, Riginella E, Mazzoldi C (2018a) Aspects of the biology of the Antarctic dragonfish Gerlachea australis (Notothenioidei: Bathydraconidae) in the Weddell Sea, Antarctica. Polar Biol 41:793–803. https://doi.org/10.1007/s00300-017-2240-y
La Mesa M, Riginella E, Donato F, Mazzoldi C (2018b) Life history traits of rare Antarctic dragonfishes from the Weddell Sea. Antarct Sci 30:289–297. https://doi.org/10.1017/S0954102018000317
La Mesa M, Vera-Duarte J, Landaeta MF (2017) Early life history traits of Harpagifer antarcticus (Harpagiferidae, Notothenioidei) from the South Shetland Islands during austral summer. Polar Biol 40:1699–1705. https://doi.org/10.1007/s00300-017-2093-4
Lagger C, Servetto N, Torre L, Sahade R (2017) Benthic colonization in newly ice-free softbottom areas in an Antarctic fjord. PLoS ONE 12:e0186756. https://doi.org/10.1371/journal.pone.0186756
Lartaud F, Chauvaud L, Richard J, Toulot A, Bollinger C, Testut L, Paulet Y-M (2010) Experimental growth pattern calibration of Antarctic scallop shells (Adamussium colbecki, Smith 1902) to provide a biogenic archive of high-resolution records of environmental and climatic changes. J Exp Mar Biol Ecol 393:158–167. https://doi.org/10.1016/j.jembe.2010.07.016
Lau SCY, Grange LJ, Peck LS, Reed AJ (2018) The reproductive ecology of the Antarctic bivalve Aequiyoldia eightsii (Protobranchia: Sareptidae) follows neither Antarctic nor taxonomic patterns. Polar Biol 41:1693–1706. https://doi.org/10.1007/s00300-018-2309-2
Lee B, Brewin PE, Brickle P, Randhawa H (2018) Use of otolith shape to inform stock structure in Patagonian toothfish (Dissostichus eleginoides) in the south-western Atlantic. Mar Freshw Res 69:1238–1247. https://doi.org/10.1071/MF17327
Liu Q, Chen D (1995) Length frequency analysis of Pleuragramma antarcticum, Electrona antarctica, Protomyctophum bolini. Chin J Oceanol Limnol 13:380–384. https://doi.org/10.1007/BF02889475
Linkowski TB (1985) Population biology of the myctophid fish Gymnoscopelus nicholsi (Gillbert, 1911) from the western South Atlantic. J Fish Biol 27:683–698. https://doi.org/10.1111/j.1095-8649.1985.tb03213.x
Linkowski TB (1987) Age and growth of four species of Electrona (teleostei, Myctophidae). In: Kullander SO and Fernholm B (eds) Proceedings of the 5th Congress of European Ichthyologists. Swedish Museum of Natural History Stockholm, Sweden, pp 435–442
Lomovasky BJ, de Aranzamendi MC, Abele D (2020) Shorter but thicker: analysis of internal growth bands in shells of intertidal vs. subtidal Antarctic limpets, Nacella concinna, reflects their environmental adaptation. Polar Biol 43:131–141. https://doi.org/10.1007/s00300-019-02615-z
Margolin AR, Robinson LF, Burke A, Waller RG, Scanlon KM, Roberts ML, Auro ME, van de Flierdt T (2014) Temporal and spatial distributions of cold-water corals in the Drake Passage: insights from the last 35,000 years. Deep-Sea Res II 99:237–248. https://doi.org/10.1016/j.dsr2.2013.06.008
Marriot P, Tracey DM, Bostock H, Hitt N, Fallon SJ (2020) Ageing deep-sea black coral Bathypathes patula. Front Mar Sci 7:479. https://doi.org/10.3389/fmars.2020.00479
Martinez-Dios A, Dominguez-Carrió C, Zapata-Guardiola R, Gili J-M (2016) New insights on Antarctic gorgonians’ age, growth and their potential as paleorecords. Deep Sea Res Part I Oceanogr Res Pap 112:57–67. https://doi.org/10.1016/j.dsr.2016.03.007
Matias RS, Seco J, Gregory S, Belchier M, Pereira ME, Bustamante P, Xavier JC (2020) Antarctic octopod beaks as proxy for mercury concentrations in soft tissues. Mar Poll Bull 158:111447. https://doi.org/10.1016/j.marpolbul.2020.111447
McClintock JB, Angus RA, Mcdonald MR, Amsler CD, Catledge SA, Vohra YK (2009) Rapid dissolution of shells of weakly calcified Antarctic benthic macroorganisms indicates high vulnerability to ocean acidification. Antarct Sci 21:449–456. https://doi.org/10.1017/S0954102009990198
McMullin RB, Wing SR, Reid MR (2017) Ice fish otoliths record dynamics of advancing and retreating sea ice in Antarctica. Limnol Oceanogr 62:2662–2673. https://doi.org/10.1002/lno.10597
Mohr RC, Tobin TS, Petersen SV, Dutton A, Oliphant E (2020) Subannual stable isotope records reveal climate warming and seasonal anoxia associated with two extinction intervals across the Cretaceous-Paleogene boundary on Seymour Island, Antarctica. Geology 48:1131–1136. https://doi.org/10.1130/G47758.1
Morales-Nin B, Palomera I, Busquets X (2002) A first attempt at determining larval growth in three Antarctic fish from otoliths and RNA/DNA ratios. Polar Biol 25:360–365. https://doi.org/10.1007/s00300-001-0351-x
Morley SA, Abele D, Barnes DKA et al (2020) Global drivers on southern ocean ecosystems: changing physical environments and anthropogenic pressures in an earth system. Front Mar Sci 15:507. https://doi.org/10.3389/fmars.2020.547188
Mormede S, Dunn A, Hanchet SM (2014) A stock assessment model of Antarctic toothfish (Dissostichus mawsoni) in the Ross Sea region incorporating multi-year mark-recapture data. CCAMLR Sci 21:39–62
Nolan CP, Clarke A (1993) Growth in the bivalve Yoldia eightsi at Signy Island, Antarctica, determined from internal shell increments and calcium-45 incorporation. Mar Biol 117:243–250. https://doi.org/10.1007/BF00345669
Oschmann W (2009) Sclerochronology: editorial. Int J Earth Sci 98:1–2. https://doi.org/10.1007/s00531-008-0403-3
Panfili J, de Pontual H, Troadec H, Wright PJ (2002) Manual of fish sclerochronology. Brest, France
Parker SJ, Mormede S, Hanchet SM, Devries A, Canese S, Ghigliotti L (2019) Monitoring Antarctic toothfish in McMurdo Sound to evaluate the Ross Sea region Marine Protected Area. Antarct Sci 31:195–207. https://doi.org/10.1017/S0954102019000245
Parkhurst NW (1990) Growth and reproduction of the Antarctic nototheniid fish Pagothenia borchgrevinki. Polar Biol 10:387–391. https://doi.org/10.1007/BF00237826
Parkinson D, Curry GB, Cusack M, Fallick AE (2005) Shell structure, patterns and trends of oxygen and carbon stable isotopes in modern brachiopod shells. Chem Geol 219:193–235. https://doi.org/10.1016/j.chemgeo.2005.02.002
Peck LS, Holmes LJ (1989) Scaling patterns in the Antarctic brachiopod Liothyrella uva (Broderip, 1833). J Exp Mar Biol Ecol 133:141–150. https://doi.org/10.1016/0022-0981(89)90163-9
Peck LS, Brey T (1996) Bomb signals in old Antarctic brachiopods. Nature 380:207–208. https://doi.org/10.1038/380207b0
Peck LS, Baker AC, Conway LZ (1996) Strontium labelling of the shell of the Antarctic limpet Nacella concinna (Strebel, 1908). J Mollus Stud 62:315–325. https://doi.org/10.1093/mollus/62.3.315
Peck LS, Brockington S, Brey T (1997) Growth and metabolism in the Antarctic brachiopod Liothyrella uva. Philos Tran R Soc B 352:851–858. https://doi.org/10.1098/rstb.1997.0065
Peck LS, Colman JG, Murray AWA (2000) Growth and tissue mass cycles in the infaunal bivalve Yoldia eightsi at Signy Island, Antarctica. Polar Boil 23:420–428. https://doi.org/10.1007/s003000050463
Pérez-Huerta A, Walker SE, Cappelli C (2020) In situ geochemical analysis of organics in growth lines of Antarctic scallop shells: implications for sclerochronology. Minerals 10:529. https://doi.org/10.3390/min10060529
Philipp E, Brey T, Pörtner H-O, Abele D (2005) Chronological and physiological ageing in a polar and a temperate mud clam. Mech Ageing Dev 126:598–609. https://doi.org/10.1016/j.mad.2004.12.003
Picken GB (1979) Growth, production and biomass of the antarctic gastropod Laevilacunaria antarctica Martens 1885. J Exp Mar Biol Ecol 40:71–79. https://doi.org/10.1016/0022-0981(79)90035-2
Picken GB (1980) The distribution, growth, and reproduction of the Antarctic limpet Nacella (Patinigera) concinna (Strebel, 1908). J Exp Mar Biol Ecol 42:71–85. https://doi.org/10.1016/0022-0981(80)90167-7
Queiros JP, Cherel Y, Ceia FR, Hilario A, Roberts J, Xavier JC (2020) Ontogenetic changes in habitat and trophic ecology of the giant Antarctic octopus Megaleledone setebos inferred from stable isotope analyses in beaks. Mar Biol 167:56. https://doi.org/10.1007/s00227-020-3666-2
Radtke RL (1990) Age determination of the Antarctic fishes Champsocephalus gunnari and Notothenia rossii marmorata from South Georgia. Polar Biol 10:321–327. https://doi.org/10.1007/BF00237818
Radtke RL, Targett TE (1984) Rhythmic structural and chemical patterns in otoliths of the Antarctic fish Notothenia larseni: their application to age determination. Polar Biol 3:203–210. https://doi.org/10.1007/BF00292624
Radtke RL, Hubold G, Folsom SD, Lenz PH (1993) Otolith structural and chemical analyses: the key to resolving age and growth of the Antarctic silverfish, Pleuragramma antarcticum. Antarct Sci 5:51–62. https://doi.org/10.1017/S0954102093000082
Radtke Rl Targett T, Kellermann A, Bell J, Hill K (1989) Antarctic fish growth: profile of Trematomus newnesi. Mar Ecol pro Ser 57:103–117. https://doi.org/10.3354/meps057103
Ralph R, Maxwell JGH (1977) Growth of Two Antarctic Lamellibranchs: Adamussium colbecki and Latemula elliptica. Mar Biol 42:171–175
Ravelo AM, Konar B, Bluhm B, Iken K (2017) Growth and production of the brittle stars Ophiura sarsii and Ophiocten sericeum (Echinodermata: Ophiuroidea). Cont Shelf Res 139:9–20. https://doi.org/10.1016/j.csr.2017.03.011
Rayfuse R (2018) Climate change and antarctic fisheries: ecosystem management in CCAMLR. Ecol Law Q 45:53–81. https://doi.org/10.15779/Z381834271
Rayner NA, Parker DE, Horton EB, Folland CK, Alexander LV, Rowell DP, Kent EC, Kaplan A (2003) Global analyses of sea surface temperature, sea ice, and night marine air temperature since the late nineteenth century. J Geophys Res 108:4407. https://doi.org/10.1029/2002JD002670
Reynolds DJ, Butler PG, Williams SM et al (2013) A multiproxy reconstruction of Hebridean (NW Scotland) spring sea surface temperatures between AD 1805 and 2010. Palaeogeogr Palaeoclimatol Palaeoecol 386:275–285. https://doi.org/10.1016/j.palaeo.2013.05.029
Reynolds DJ, Richardson CA, Scourse JD, Butler PG, Hollyman P, Roman-Gonzalez A, Hall IR (2017) Reconstructing North Atlantic marine climate variability using an absolutely-dated sclerochronological network. Palaeogeogr Palaeoclimatol Palaeoecol 465:333–346. https://doi.org/10.1016/j.palaeo.2016.08.006
Richardson MG (1979) The ecology and reproduction of the brooding Antarctic bivalve Lissarca miliaris. Br Antarct Surv Bull 49:91–115
Richardson CA (2001) Molluscs as archives of environmental change. Oceanogr Mar Biol 39:103–164
Robinson LF, van de Flierdt T (2009) Southern Ocean evidence for reduced export of North Atlantic Deep Water during Heinrich event 1. Geology 37:195–198. https://doi.org/10.1130/G25363A.1
Roman-Gonzalez A (2011) Climate change in the Antarctic: the search for sclerochronological proxies (MSc. dissertation). Bangor University, School of Ocean Sciences, Bangor (Gwynedd), Wales, United Kingdom
Román-González A (2018) Investigation of shallow marine Antarctic environments using the annual increment growth pattern of the bivalve mollusc Aequiyoldia eightsii (Jay, 1839). (PhD. Thesis). Cardiff University, Cardiff, United Kingdom
Roman-Gonzalez A, Scourse JD, Richardson CA, Peck LS, Bentley MJ, Butler PG (2016) A sclerochronological archive for Antarctic coastal waters based on the marine bivalve Yoldia eightsi (Jay, 1839) from the South Orkney Islands. Holocene 27:271–281. https://doi.org/10.1177/0959683616658525
Roman-Gonzalez A, Scourse JD, Butler PG, Reynolds DJ, Richardson CA, Penk LS, Brey T, Hall IR (2017) Analysis of ontogenetic growth trends in two marine Antarctic bivalves Yoldia eightsi and Laternula elliptica: Implications for sclerochronology. Palaeogeogr Palaeoclimatol Palaeoecol 465:300–306. https://doi.org/10.1016/j.palaeo.2016.05.004
Rosa R, Lopes VM, Guerreiro M, Bolstad K, Xavier JC (2017) Biology and ecology of the world’s largest invertebrate, the colossal squid (Mesonychoteuthis hamiltoni): a short review. Polar Biol 40:1871–1883. https://doi.org/10.1007/s00300-017-2104-5
Ruzicka J (1996) Comparison of the two alternative early life-history strategies of the Antarctic fishes Gobionotothen gibberifrons and Lepidonotothen larseni. Mar Ecol pro Ser 133:29–41. https://doi.org/10.3354/meps133029
Ruzicka J, Radtke RL (1995) Estimating the age of Antarctic larval fish from otolith microstructure using light and electron microscopy. Polar Biol 15:587–592. https://doi.org/10.1007/BF00239651
Sahade R, Lagger C, Torre L et al (2015) Climate change and glacier retreat drive shifts in an Antarctic benthic ecosystem. Sci Adv 1:e1500050. https://doi.org/10.1126/sciadv.1500050
Sato-Okoshi W, Okoshi K, Sasaki H, Akiha F (2010) Shell structure of two polar pelagic molluscs, Arctic Limacina helicina and Antarctic Limacina helicina antarctica forma antarctica. Polar Biol 33:1577–1583. https://doi.org/10.1007/s00300-010-0849-1
Samperiz A, Robinson LF, Stewart JA, Strawson I, Leng MJ, Rosenheim BE, Ciscato ER, Hendry KR, Santodomingo N (2020) Stylasterid corals: a new paleotemperature archive. Earth Planet Sci Lett 545:116407. https://doi.org/10.1016/j.epsl.2020.116407
Saunders RA, Collins MA, Ward P, Stowasser G, Shreeve R, Tarling GA (2015) Trophodynamics of Protomyctophum (Myctophidae) in the Scotia Sea (Southern Ocean). J Fish Biol 87:1031–1058. https://doi.org/10.1111/jfb.12776
Saunders RA, Lourenço S, Vieira RP, Collins MA, Assis CA, Xavier JC (2019) Age and growth of Brauer’s lanternfish Gymnoscopelus braueri and rhombic lanternfish Krefftichthys anderssoni (Family Myctophidae) in the Scotia Sea, Southern Ocean. J Fish Biol 96:364–377. https://doi.org/10.1111/jfb.14206
Schejter L, Bremec CS, Cairns S (2016) Scleractinian corals recorded in the Argentinean Antarctic expeditions between 2012 and 2014, with comments on Flabellum (Flabellum) areum Cairns, 1982. Polar Res 35:29762. https://doi.org/10.3402/polar.v35.29762
Schmidtko S, Heywood KJ, Thompson AF, Aoki S (2014) Multidecadal warming of Antarctic waters. Science 346:1227–1231. https://doi.org/10.1126/science.1256117
Schöne BR, Gillikin DP (2013) Unraveling environmental histories from skeletal diaries—advances in sclerochronology. Palaeogeogr Palaeoclimatol Palaeoecol 373:1–5. https://doi.org/10.1016/j.palaeo.2012.11.026
Schöne BR, Rodland DL, Wehrmann A, Heidel B, Oschmann W, Zhang Z, Fiebig J, Beck L (2007) Combined sclerochronologic and oxygen isotope analysis of gastropod shells (Gibbula cineraria, North Sea): life-history traits and utility as a high-resolution environmental archive for kelp forests. Mar Biol 150:1237–1252. https://doi.org/10.1007/s00227-006-0435-9
Smith AM (2007) Age, growth and carbonate production by erect rigid bryozoans in Antarctica. Palaeogeogr Palaeoclimatol Palaeoecol 256:86–98. https://doi.org/10.1016/j.palaeo.2007.09.007
Smith JE, Schwarcz HP, Risk MJ, McConnaughey TA, Keller N (2000) Paleotemperatures from deep-sea corals: overcoming ‘vital effects.’ Palaios 15:25–32. https://doi.org/10.2307/3515589
Spooner PT, Guo W, Robinson LF, Thiagarajan N, Hendry KR, Rosenheim BE, Leng MJ (2016) Clumped isotope composition of cold-water corals: a role for vital effects? Geochim Cosmochim Acta 179:123–141. https://doi.org/10.1016/j.gca.2016.01.023
Stanley GD Jr, Cairns SD (1988) Constructional azooxanthellate coral communities: an overview with implications for the fossil record. Palaios 3:233–242. https://doi.org/10.2307/3514534
Stebbing ARD (1971) Growth of Flustra foliacea (Bryozoa). Mar Biol 9:267–273. https://doi.org/10.1007/BF00351389
Stockton WL (1984) The biology and ecology of the epifaunal scallop Adamussium colbecki on the west side of McMurdo Sound, Antarctica. Mar Biol 78:171–178. https://doi.org/10.1007/BF00394697
Struve T, Van de Flierdt T, Burke A, Robinson LF, Hammond SJ, Crocket KC, Bradtmiller LI, Auro ME, Mohamed KJ, White NJ (2017) Neodymium isotopes and concentrations in aragonitic scleractian cold-water coral skeletons—modern calibration and evaluation of palaeo-applications. Chem Geol 453:146–168. https://doi.org/10.1016/j.chemgeo.2017.01.022
Tada Y, Wada H, Miura H (2006) Seasonal stable oxygen isotope cycles in an Antarctic bivalve shell (Laternula elliptica): a quantitative archive of ice-melt runoff. Antarct Sci 18:111–115. https://doi.org/10.1017/S0954102006000101
Trevisiol A, Bergamasco A, Montagna P, Sprovieri M, Taviani M (2013) Antarctic seawater temperature evaluation based on stable isotope measurements on Adamussium colbecki shells: kinetic effects vs. isotopic equilibrium. J Mar Syst 126:43–55. https://doi.org/10.1016/j.jmarsys.2012.10.012
Trofimova T, Alexandroff SJ, Mette MJ et al (2020) Fundamental questions and applications of sclerochronology: Community-defined research priorities. Estuar Coast Shelf Sci 245:106977. https://doi.org/10.1016/j.ecss.2020.106977
Turner J, Comiso J (2017) Solve Antarctica’s sea-ice puzzle. Nature 547:275–277. https://doi.org/10.1038/547275a
Watson S-A, Morley SA, Peck LS (2017) Latitudinal trends in shell production cost from the tropics to the poles. Sci Adv 3:e1701362. https://doi.org/10.1126/sciadv.1701362
Wendebourg MR (2020) Southern Ocean fishery management—is CCAMLR addressing the challenges posed by a changing climate? Mar Policy 118:103847. https://doi.org/10.1016/j.marpol.2020.103847
Williams B (2020) Proteinaceous corals as proxy archives of paleo-environmental change. Earth Sci Rev 209:103326. https://doi.org/10.1016/j.earscirev.2020.103326
Wilson DJ, Struve T, van de Flierdt T, Chen T, Li T, Burke A, Robinson LF (2020) Sea-ice control on deglacial lower cell circulation changes recorded by Drake Passage deep-sea corals. Earth Planet Sci Lett 544:116405. https://doi.org/10.1016/j.epsl.2020.116405
Wing SR, O’Connell-Milne SA, Wing LC, Reid MR (2020) Trace metals in Antarctic clam shells record the chemical dynamics of changing sea ice conditions. Limnol Oceanogr 65:504–514. https://doi.org/10.1002/lno.11318
Woo KS, Kim J-K, Lee JI, Lim HS, Yoo K-C, Summerhayes CP, Ahn I-Y, Kang S-H, Kil Y (2019) Glacial melting pulses in the Antarctica: evidence for different responses to regional effects of global warming recorded in Antarctic bivalve shell (Laternula elliptica). J Mar Syst 197:103179. https://doi.org/10.1016/j.jmarsys.2019.05.005
Zinsmeister W (1984) Late Eocene Bivalves (Mollusca) from the La Meseta Formation, collected during the 1974–1975 joint Argentine-American expedition to Seymour Island, Antarctic Peninsula. J Paleontol 58:1497–1527
Zhu G, Duan M, Ashford JR, Wei L, Zhou M, Bestley S (2018) Otolith nucleus chemistry distinguishes Electrona antarctica in the westward-flowing Antarctic Slope Current and eastward-flowing Antarctic Circumpolar Current off East Antarctica. Mar Environ Res 142:7–20. https://doi.org/10.1016/j.marenvres.2018.09.010
Zhu G, Duan M, Wei L, Trebilco R, Bestley S, Walters A (2020) Determination and precision of otolith growth zone estimates of Electrona antarctica in the Southern Kerguelen Plateau region in the Indian sector of the Southern Ocean. Deep-Sea Res 174:104778. https://doi.org/10.1016/j.dsr2.2020.104778
Acknowledgements
This manuscript was possible thanks to the funding provided by the UK-Chile (NERC-CONICYT) collaborative project: Impacts of deglaciation benthic marine ecosystem in Antarctica (ICEBERGS; NE/P003087/1). I want to express my gratitude to Nicolas Duprey, Simone Moretti and one anonymous reviewer whose contribution increased the quality and the scope of this review. In addition, I want to thank my colleagues at University of Exeter (James Scourse and Paul Butler) to provide me with their comments and opinions about the present work.
Funding
This manuscripts was possible thanks to the funding provided by the UK-Chile (NERC-CONICYT) collaborative project: Impacts of deglaciation benthic marine ecosystem in Antarctica (ICEBERGS; NE/P003087/1).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The author does not have any conflict of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Roman Gonzalez, A. Sclerochronology in the Southern Ocean. Polar Biol 44, 1485–1515 (2021). https://doi.org/10.1007/s00300-021-02899-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-021-02899-0