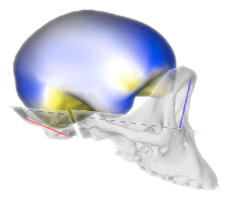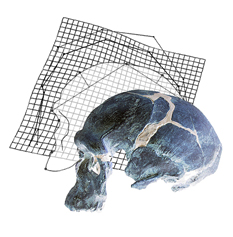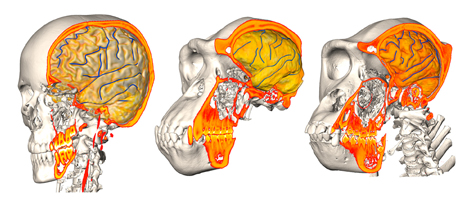
A recent study by José Luis Alatorre Warren and colleagues supplies one of the very few analyses on both skull and brain morphology from the same individuals, in humans, chimpanzees, and gorillas. They analyzed the shape variation of the brain and skull separately, and then at the same time. The study describes the spatial relationships between brain and skull, and the differences between species. The study confirms once more that there is a limited correspondence between cerebral and cranial anatomical boundaries and references. According to their results, the most striking difference between humans and chimpanzees is, in the former, a spatial change of the frontal lobe regions associated with language. They also found a larger parietal cortex in humans when compared with apes but, according to the model they use, these differences would be not so remarkable as suggested by most of the preceding studies. However, their geometrical model is very comprehensive and it includes many distinct and independent elements of the brain and of the skull, that are melted and averaged when all their coordinates are superimposed together. In these cases, punctual or local differences are spread onto the whole global variation, and can be hardly detected. Namely, these extensive shape registrations are excellent to analyze general covariation patterns, but can be tricky when trying to identify the contribution of local regions or of specific anatomical elements in a heterogeneous and multifactorial anatomical complex.
In my opinion, the article suffers, unfortunately, some important conceptual limitations. There is a confusion between Ralph Holloway’s theories on parietal evolution in australopiths (largely based on the position of the lunate sulcus) and my own ones on parietal evolution in modern humans (based on the size and proportions of the parietal lobe). It seems obvious to say, but we are dealing with distinct taxa (australopiths and modern humans) and distinct processes (early hominid evolution and late human specialization), and therefore there is no reason to look for common factors or mechanisms. For example, when taking into account the possibile influence of posture on brain shape, the evolution of bipedalism may be an issue for australopiths or early Homo, but not when comparing modern humans with Neandertals. Also, there seems to be a constant misunderstanding between parietal cortex enlargement (which involves parietal lobe absolute and relative size) and brain globularization (which refers to the rounded shape of the brain). It should be clear that these features can be influenced by (and due to) different factors, although they concern the same anatomical regions. Finally, there is a frequent confusion (and miscitation) between papers and results on parietal bones, and papers and results on parietal lobes. So, in my opinion, we are dealing with a study which supplies an amazing analysis, but a problematic discussion of the results. I think this study would have seriously benefited from a more cautious interpretation of the numerical outputs, and a more careful integration of the literature. Nonetheless, this exceptional database is there, and I hope it will supply in the future more information on the spatial relationships between brain and braincase.