
Mislabeling and nomenclatorial confusion of Typhlotanais sandersi Kudinova-Pasternak, 1985 (Crustacea: Tanaidacea) and establishment of a new genus
- Published
- Accepted
- Received
- Academic Editor
- Antonina Dos Santos
- Subject Areas
- Biodiversity, Marine Biology, Taxonomy, Zoology
- Keywords
- Peracarida, North Atlantic, Deep-sea, Northwest Pacific
- Copyright
- © 2022 Gellert et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2022. Mislabeling and nomenclatorial confusion of Typhlotanais sandersi Kudinova-Pasternak, 1985 (Crustacea: Tanaidacea) and establishment of a new genus. PeerJ 10:e14272 https://doi.org/10.7717/peerj.14272
Abstract
Re-examination of historical collections allowed us to resolve the taxonomic status of Typhlotanais sandersi Kudinova-Pasternak, 1985, originally described based on a single specimen from Great-Meteor Seamount. The holotype of this species was considered lost and the species redescribed based on a second specimen from the type locality by Błażewicz-Paszkowycz (2007a), who placed Ty. sandersi on a newly established genus Typhlamia. Thorough morphological analysis of Typhlamia and Typhlotanais species and recently obtained genetic data of typhlotanaids from N Atlantic and NW Pacific waters allow us to conclude that the redescription of Ty. sandersi by Błażewicz-Paszkowycz (2007a) was based on a wrongly labelled specimen that, rather than a type of Ty. sandersi, represents in fact a new species of Typhlamia. The morphological comparison of the type species of Typhlotanais (Ty. aequiremis) with all ‘long-bodied’ typhlotanaid taxa with rounded pereonites margins (i.e., Typhlamia, Pulcherella, Torquella), and the use of genetic evidence, support the establishment of a new genus to accommodate: Ty. sandersi, Ty. angusticheles Kudinova-Pasternak, 1989, and a third species from N Atlantic waters, that is described here for the first time. Current knowledge on ‘long-bodied’ typhlotanaids with rounded pereonites is summarised and a taxonomical key for their identification provided.
Introduction
Typhlotanais Sars, 1882 (Crustacea: Tanaidacea) is a diverse genus established by Sars (1882) about two decades after Lilljeborg (1864) discovered its first member and type species—Tanais aequiremis Lilljeborg, 1864—on north off Ireland (Lilljeborg, 1864; Sars, 1882). The definition of the genus comprises characters shared by many deep water tanaids such as three-article antennules and complete lack of eyes (Sieg, 1983, 1984). This short and elusive diagnosis is the main reason why Typhlotanais quickly became a hotchpotch taxon comprising a handful of diverse species (Sars, 1899; Hansen, 1913; Vanhöffen, 1914; Kudinova-Pasternak, 1966, 1984, 1990; Lang, 1968; Shiino, 1970). The excellent illustrations of Sars (1899) clearly show that Typhlotanais aequiremis (Lilljeborg, 1864) has a long body with distinctly straight pereonites (Fig. 1A). Morphological differences between the type species and the other putative Typhlotanais species were so apparent that, during the second half of the 20th century, four genera (Meromonakantha Sieg, 1986a, Paratyphlotanais Kudinova-Pasternak & Pasternak, 1978, Peraeospinosus Sieg, 1986a, and Typhlotanoides Sieg, 1983) and the family Typhlotanaidae Sieg, 1984 itself were erected (Kudinova-Pasternak & Pasternak, 1978; Sieg, 1983, 1984, 1986a). Typhlotanais currently includes 53 species (44% of 123 typhlotanaid species known) and it is the most speciose out of 16 typhlotanaid genera (Gellert, Palero & Błażewicz, 2022; WoRMS, 2022; Lubinevsky, Tom & Bird, 2022).
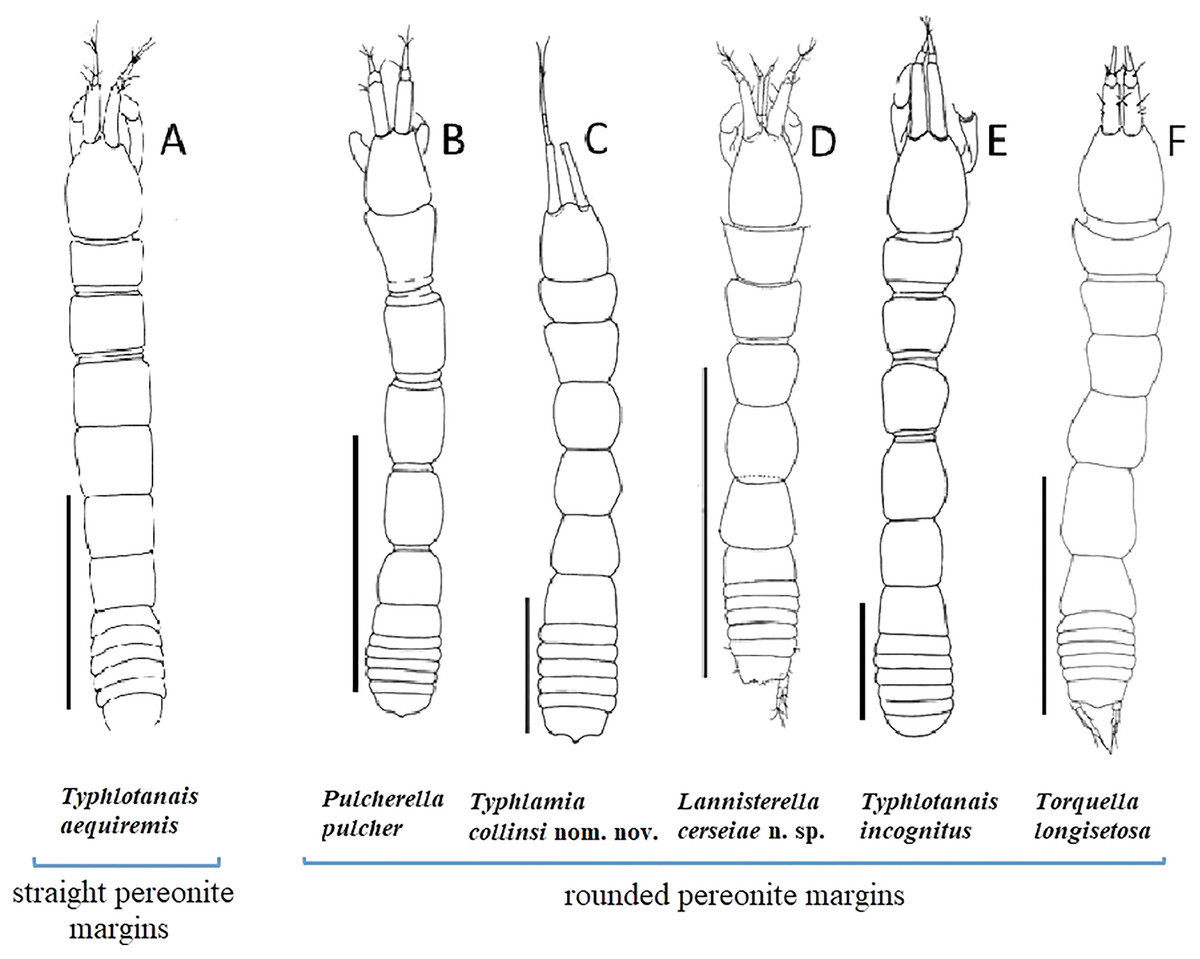
Figure 1: Comparison of body habitus.
(A) Typhlotanais aequiremis (Lilljeborg, 1864). (B) Pulcherella pulcher (Hansen, 1913). (C) Typhlamia collinsi nom. nov. (D) Lannisterella cerseiae n. sp. (E) ‘variabilis’ group: Typhlotanais incognitus Larsen, Błażewicz-Paszkowycz & Cunha, 2006. (F) Torquella longisetosa (Kudinova-Pasternak, 1990) (from: Błażewicz-Paszkowycz, 2007a; Larsen, Błażewicz-Paszkowycz & Cunha, 2006). Scale: 1 mm.{kind=link}
Kudinova-Pasternak (1985) reported four new Typhlotanaidae species from the RV Vitjaz expedition at the base of the Great-Meteor Seamount, North Atlantic, namely Typhlotanais sandersi Kudinova-Pasternak, 1985, Paraleptognathia bacescui Kudinova-Pasternak, 1985, Paratanais hessleri Kudinova-Pasternak, 1985, and Pseudotanais siegi (P. diegi sic!) Kudinova-Pasternak, 1985, and deposited type material for all those taxa at the Museum of Zoology in Moscow (Kudinova-Pasternak, 1985). According to the original description (in Russian), the holotype of Ty. sandersi was fully dissected and kept in slides (cat. No. Mh:4) before drawing its appendages. Those slides could not be found just two decades later though, so Błażewicz-Paszkowycz (2007a) decided to redescribe Ty. sandersi based on a second, undissected specimen, found in a jar labelled as “holotype cat. No. Mh:4” (Błażewicz-Paszkowycz, 2007a: p. 90–93, figs. 50–51). The specimen that Błażewicz-Paszkowycz found and illustrated was morphologically distinct from Ty. aequiremis (type species of Typhlotanais), so she decided to erect a new genus (i.e., Typhlamia) to accommodate what she called Typhlamia sandersi (Kudinova-Pasternak, 1985) along with Tm. mucronata (Hansen, 1913) and Tm. bella Błażewicz-Paszkowycz (2007a).
After a new species of Typhlamia Błażewicz-Paszkowycz, 2007a was collected from Northwest Pacific waters (Gellert, Palero & Błażewicz, 2022), detailed morphological analysis and literature revision has revealed that the original description of Ty. sandersi by Kudinova-Pasternak (1985) differed from the redescription made by Błażewicz-Paszkowycz (2007a). A thorough morphological analysis of ‘long-bodied’ typhlotanaids with rounded pereonite margins (e.g., Typhlamia, Pulcherella, Torquella), all different from Ty. aequiremis, has allowed us to conclude that the putative Ty. sandersi specimen studied by Błażewicz-Paszkowycz (2007a) was wrongly labelled and in fact represents a different species and genus from the Ty. sandersi described and figured by Kudinova-Pasternak (1985). This article aims to correct the taxonomic mixing of Ty. sandersi described by Kudinova-Pasternak (1985) and Tm. sandersi sensu Błażewicz-Paszkowycz (2007a) and to establish a new genus with the support of additional morphological and molecular evidence.
As a result of morphological analysis of long body typhlotanaids with rounded pereonite margins, a new morphological group ‘variabilis’ was distinguished, for which the definition is given here for the first time.
Materials and Methods
Literature data
Data on the morphology and morphometry of the ‘long-bodied’ typhlotanaids (body length ≥8.0 L:W) with rounded pereonites were collected from Bird (2004), Błażewicz-Paszkowycz (2007a), Błażewicz-Paszkowycz, Bamber & Jóźwiak (2013), Błażewicz-Paszkowycz et al. (2014), Gellert, Palero & Błażewicz (2022), Hansen (1913), Kudinova-Pasternak (1966, 1985, 1989, 1990), Kudinova-Pasternak & Pasternak (1978), Lang (1971), Larsen (2011), Larsen, Błażewicz-Paszkowycz & Cunha (2006), Larsen & Shimomura (2007), Sars (1882, 1899). The data gathered are shown in Table 1.
| Genus/morpho-group | Torquella Błażewicz-Paszkowycz, 2007a | Typhlamia Błażewicz-Paszkowycz, 2007a | Pulcherella Błażewicz-Paszkowycz, 2007a | Lannisterella n. gen. | ‘variabilis’ group | |
|---|---|---|---|---|---|---|
| Species included | Tq. angularis, Tq. eltaninae, Tq. galatheae, Tq. grandis, Tq. iberica, Tq. longisetosa, Tq. magdalensis, Tq. parangularis, Tq. rotundirostris | Tm. bella, Tm. genesis, Tm. mucronata, Tm. collinsi | P. juraszi, P. filatovae, P. pulcher, P. spiniventris | L. angusticheles, L. cerseiae, L. sandersi | Ty. incognitus, Ty. variabilis | |
| Carapace | Rounded (1.0–1.2 L:W) | Tapering proximally, short (1.0–1.2 L:W) | Tapering proximally, short (1.1 L:W) | Elongated (1.3 L:W) | Tapering proximally, short (1.2 L:W) | |
| Pereonite-1 proximal corners | Proximally extended (‘collar’) | Proximally not extended | Proximally not extended | Proxilmally extended (‘collar’) | Proximally not extended | |
| Pereonite-1 lateral seta | Present or absent | Absent | Absent | Present | Absent | |
| Antennule inner margin with | Middle and distal setae | Two middle and one distal seta | middle and distal setae | Four long setae | Short middle and distal setae | |
| Antennule article-3 | Short | Long | Short | Short | Short | |
| Antennule with distal spur | Present | Absent | Absent | Present | Absent | |
| Maxilliped | Basis | Long | Short | Short | Long | Long |
| Basal seta | Short | Long | Long | Short | Short | |
| Cheliped carpus ventral margin | Rounded | Straight | Straight | Rounded | Straight | |
| Minute (‘third’) seta | Present | Absent | Absent | Present | Present | |
| Pereopod-1 carpus dorsodistal seta | Short | Long | Long | Long/short | Short | |
| Pereopods 2–3 Propodus dorsodistal seta | Long | Long | Long | Long | Short | |
| Carpus (ornamentation) | With short setae, spines, tubercles, rod setae | With setae and spine(s) | With setae only | With setae and spine(s) | With setae only | |
| Pereopods 4–6 carpus with clinging apparatus | Surrounded by blunt spines | Not surrounded by blunt spines | Not surrounded by blunt spines | Surrounded by blunt spines | Not surrounded by blunt spines | |
| Pereopods 4–6 unguis | Simple | Bifurcated | Bifurcated | Bifurcated | Bifurcated | |
| Uropod | Exopod (article number) | Two-articled | One-articled | One-articled | Two-articled | Two-articled |
| Character | Slender or short | Slender | Slender | Slender | Slender | |
| Exopod/endopod | Shorter | Shorter | Shorter | Shorter | Almost equal | |
Genetic analyses
All sequences used in the molecular studies are from GenBank. Sequences from Gellert, Palero & Błażewicz (2022) were obtained from the University of Lodz Tanaidacea collection (GenBank accession numbers: ON310832-ON310845 for COI and ON255540-ON255555 for 18S rDNA; see Table 2). The molecular tree used in the current study come from publication by Gellert, Palero & Błażewicz (2022).
Terminology and species description
Total body length (BL) was measured along the central axis of symmetry from the frontal margin to the end of pleotelson; body width (BW) was measured perpendicular to the main axis at the widest point of pereonite-3. Width and length of carapace, pereonites, pleonites, and pleotelson were measured on whole specimens. All measurements were taken using a digital camera connected to the microscope (Nikon Eclipse Ci-L) and the NIS-Elements View software (www.nikoninstruments.com).
The clinging apparatus is a system of various hooks, tubercles, thorns, and spines located on the carpus of pereopods 4−6 (Błażewicz-Paszkowycz, 2007a; Gellert et al., 2022). Unspecified setae in taxonomic descriptions are referred here as simple setae (= without ornamentation) by default. Besides, we recognize penicillate setae—with a distal tuft of setules and supracuticular articulation, and rod setae—distally inflated seta and with a terminal pore (Thomas, 1970; Garm, 2004). The short ventral seta situated besides two long setae on the cheliped carpus is called ‘third’ seta. The distal part of the cheliped basis, extending backwards, is referred here as ‘cheliped lobe’ (Larsen, 2005). The term ‘collar’ is used to refer the shape of pereonite-1, with a deeply concave anterior edge and lateral corners extended forwards (e.g., Torquella Błażewicz-Paszkowycz, 2007a). Błażewicz-Paszkowycz (2007a) proposed a classification of typhlotanaids into ‘short-bodied’ (body <6.0 L:W) and ‘long-bodied’ (body ≥8.0 L:W) taxa (see the Key for Typhlotanaidae genera and morpho-groups in Błażewicz-Paszkowycz, 2007a). The neuter is the post-manca stage, that cannot be classified as male or female. Two-letter genus abbreviations are used throughout the text to distinguish between genera: Tq. = Torquella, Tm. = Typhlamia and Ty. = Typhlotanais. Type material for the species described here is deposited in the Museum der Natur Hamburg, Germany (ZMHK) and ICUL/ITan numbers correspond to unique identifiers from the Invertebrates Collection held at the University of Lodz, Poland.
Results and interpretation
The recent discovery of a new species of Typhlamia from the Kuril-Kamchatka Trench (Gellert, Palero & Błażewicz, 2022) and the re-evaluation of Typhlotanais sp. A from Błażewicz-Paszkowycz et al. (2014) uncovered significant genetic and morphological differences between Typhlamia and the morphogroup of species including the original holotype description and drawings of Typhlotanais sandersi Kudinova-Pasternak, 1985. Although types of all taxa described by Kudinova-Pasternak were deposited at the Zoological Museum in Moscow after her retirement (R.K. Kudinova-Pasternak in 2004, personal communication), all microscopic slides made for studying and illustrating morphological details were disposed of, even those with holotype appendages. For this reason, most of the type collection studied by Kudinova-Pasternak (except undissected type material) should be considered lost. This situation is particularly critical for Ty. sandersi Kudinova-Pasternak (1985), of which a single specimen was dissected after the habitus was illustrated.
According to Kudinova-Pasternak (1985), two Typhlotanais specimens were collected from the same station (st. 162) during the RV Vitjaz expedition at the base of the Great-Meteor Seamount: the holotype of Ty. sandersi described and drawn in the text and another individual identified as Typhlotanais mucronatus Hansen, 1913 (currently Typhlamia mucronata (Hansen, 1913)). The latter was not dissected by Kudinova-Pasternak because it was assigned to a known species and, most likely, it was kept intact at the Zoological Museum of Moscow State Lomonosov University within the original jar. Indeed, an intact specimen was later found by Błażewicz-Paszkowycz (2007a) during her revision of Typhlotanaidae taxa in a vial labelled “Typhlotanais sandersi n. sp., holotype, Mh 4”, and Błażewicz-Paszkowycz (2007a) wrongly considered it as type material of Typhlotanais sandersi Kudinova-Pasternak, 1985. In fact, the undissected specimen corresponded to the second Typhlotanais specimen from station 162, identified initially by Kudinova-Pasternak as Typhlotanais mucronatus Hansen, 1913.
A careful comparison of their morphology, based on the original drawings, bring us to the conclusion that the true Typhlotanais sandersi (holotype) and the specimen referred by Błażewicz-Paszkowycz (2007a) as Typhlamia sandersi (Kudinova-Pasternak, 1985) represent two distinct taxa. All species of Typhlamia present an elongated third antennular article with particularly long distal setae (Fig. 1C), and the intact specimen is undoubtedly close to Ty. mucronatus Hansen, 1913; see Błażewicz-Paszkowycz, 2007a: 90–93). The absence of this antennular feature in the holotype of Typhlotanais sandersi illustrated by Kudinova-Pasternak (1985: page 54) drew our attention first, and further analysis revealed other significant differences in the uropods (exopod unarticulated and about half the endopod length in Typhlamia (Fig. 2G), whereas it is clearly biarticulated and subequal to endopod in the holotype) and cheliped carpus or pereonite-1 setation, among other features (see Table 1).
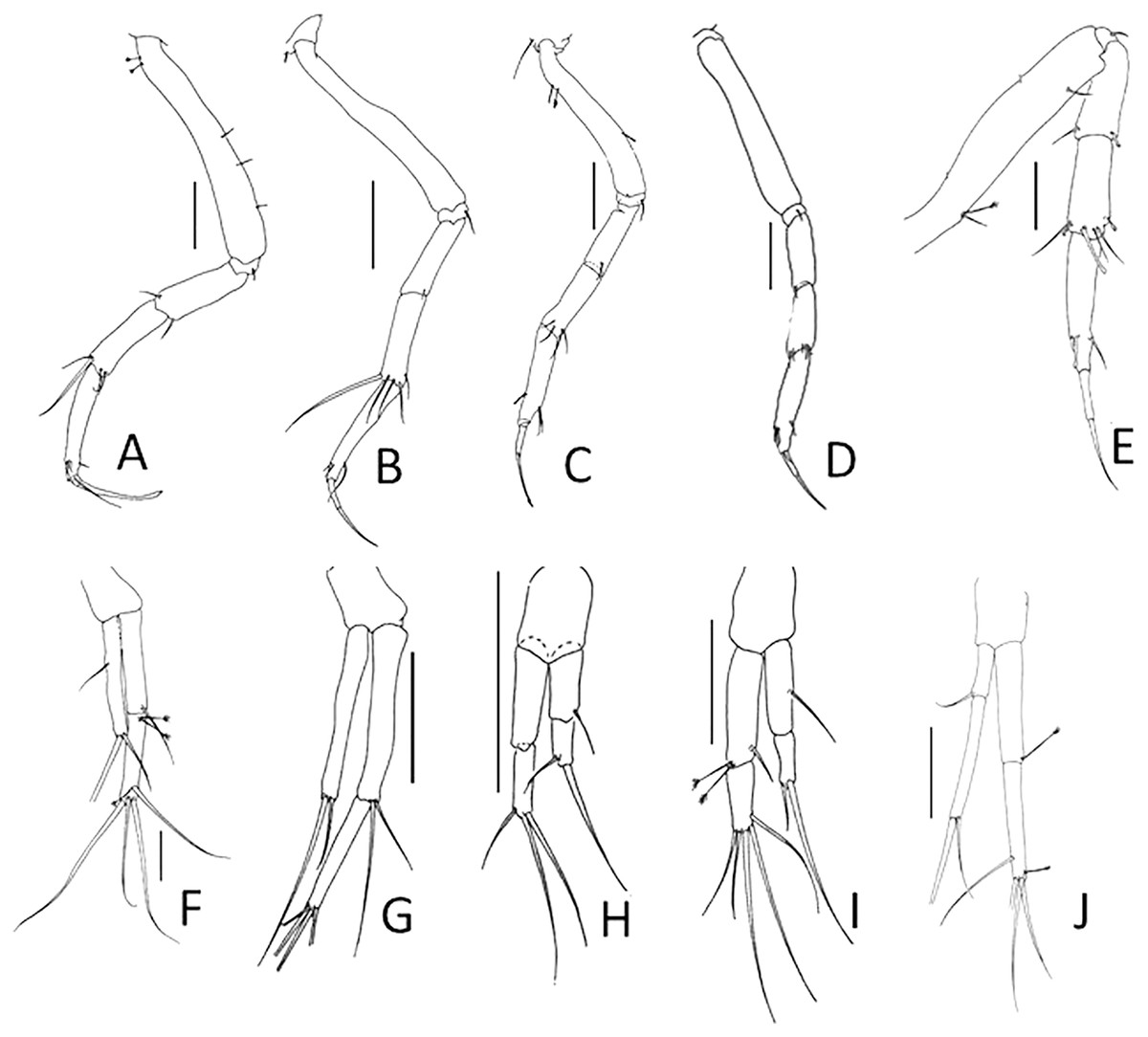
Figure 2: Comparison of pereopod-1 and uropod.
(A, F) Pulcherella pulcher (Hansen, 1913). (B, G) Typhlamia collinsi nom. nov. (C, H) Lannisterella cerseiae n. sp. (D, I) ‘variabilis’ group: Typhlotanais incognitus Larsen, Błażewicz-Paszkowycz & Cunha, 2006. (E, J) Torquella longisetosa (Kudinova-Pasternak, 1990) (from: Błażewicz-Paszkowycz, 2007a; Larsen, Błażewicz-Paszkowycz & Cunha, 2006). Scale: 0.1 mm.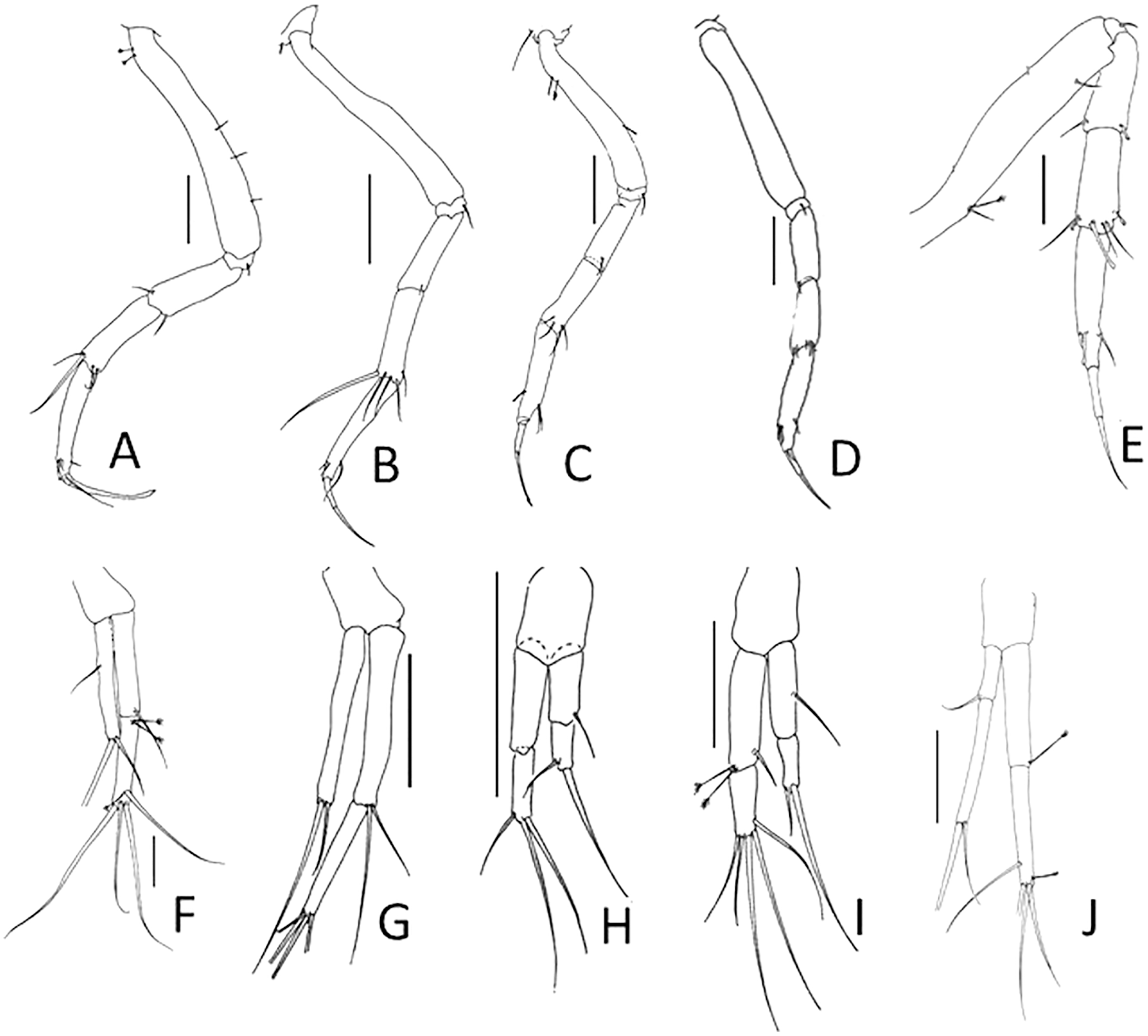{kind=link}
The morphological revision of ‘long-bodied’ typhlotanaids with rounded pereonites revealed that Typhlotanais sandersi Kudinova-Pasternak, 1985 is morphologically close to Typhlotanais angusticheles Kudinova-Pasternak, 1989 and Typhlotanais sp. A (Błażewicz-Paszkowycz et al., 2014) (Table 3). Those three species most likely belong to a new, undescribed, genus, because they are apparently different from both Typhlamia and Typhlotanais aequiremis Lilljeborg, 1864 (type species of Typhlotanais). The combination of features such as the long body and round pereonites distinguishes these three species from other Typhlotanaidae genera. Indeed, the molecular comparison of species of Typhlamia genesis and Typhlotanais sp. A (morphologically close to Ty. sandersi) confirmed that both taxa belong to genetically distinct clades (Fig. 3), giving further support for the establishment of a new genus, herein named Lannisterella n. gen.
| Species | Expedition | Area | Station | Latitude | Longitude | Depth [m] | Reference |
|---|---|---|---|---|---|---|---|
| Tm. bella | ANDEEEP-3 XXII | Weddell Sea | PS67/81-8-E | 70°32.19′S | 14°35.13′W | 4,392–4,385 | Błażewicz-Paszkowycz (2007a) |
| PS 67/81-8-E | 70°32.19′S | 14°35.13′W | 4,392–4,385 | ||||
| ANDEEEP-3 XXII | W off Antarctic Peninsula | PS 67/154-9 | 62°31.36′S | 64°39.25′W | 3,804–3,808 | ||
| Tm. genesis | KuramBio I | Kuril-Kamchatka Trench | 1-10 | 43°57.92′N | 157°23.76′E | 5,423–5,429 | Gellert, Palero & Błażewicz (2022) |
| 6-12 | 42°32.68′N | 154°1.27′E | 5,304–5,307 | ||||
| 9-9 | 40°38.79′N | 150°59.97′E | 5,399–5,408 | ||||
| 9-12 | 40°38.71′N | 150°59.72′E | 5,392–5,397 | ||||
| 10-9 | 41°15.66′N | 150°5.70′E | 5,265–5,643 | ||||
| 10-12 | 41°8.44′N | 150°5.53′E | 5,249–5,262 | ||||
| 12-4 | 39°46.72′N | 147°11.90′E | 5,215–5,228 | ||||
| KuramBio II | Kuril-Kamchatka Trench | 8 | 43°51.698′N | 151°45.85′E | 5,103–5,109 | ||
| 10 | 43°51.810′N | 151°46.54′E | 5,103–5,188 | ||||
| 86 | 44°56.784′N | 151°6.009′E | 5,534–5,630 | ||||
| 87 | 44°58.024′N | 151°5.589′E | 5,640–5,465 | ||||
| Vitjaz | Kuril-Kamchatka Trench | 5617 | 45°32′N | 153°46′E | 6,740–6,710 | Kudinova-Pasternak (1970) | |
| 5624 | 45°22′N | 154°00′E | 5,200–5,240 | ||||
| 5634 | 44°17′N | 149°33′E | 4,690–4,720 | ||||
| Gulf of Alaska | 6109 | 56°14′N | 139°44′W | 3,460–3,450 | Kudinova-Pasternak (1973) | ||
| Tm. mucronata | Ingolf | N Iceland | 120 | 67°29′N | 11°32′W | 1,620 | Hansen (1913) |
| 119 | 67°53′N | 10°19′W | 1,848 | ||||
| Jan Mayen | 177 | 69°13′N | 8°23′W | 1,835 | |||
| BIOGAS | Bay of Biscay | 3 | 47°33.90′N | 09°38.40′W | 3,992–4,260 | Holdich & Bird (1985) | |
| Tm. sandersi | Vitjaz | North Atlantic | 162 | 29°50′N | 28°08′6E | 3,080–3,140 | Kudinova-Pasternak (1985) |
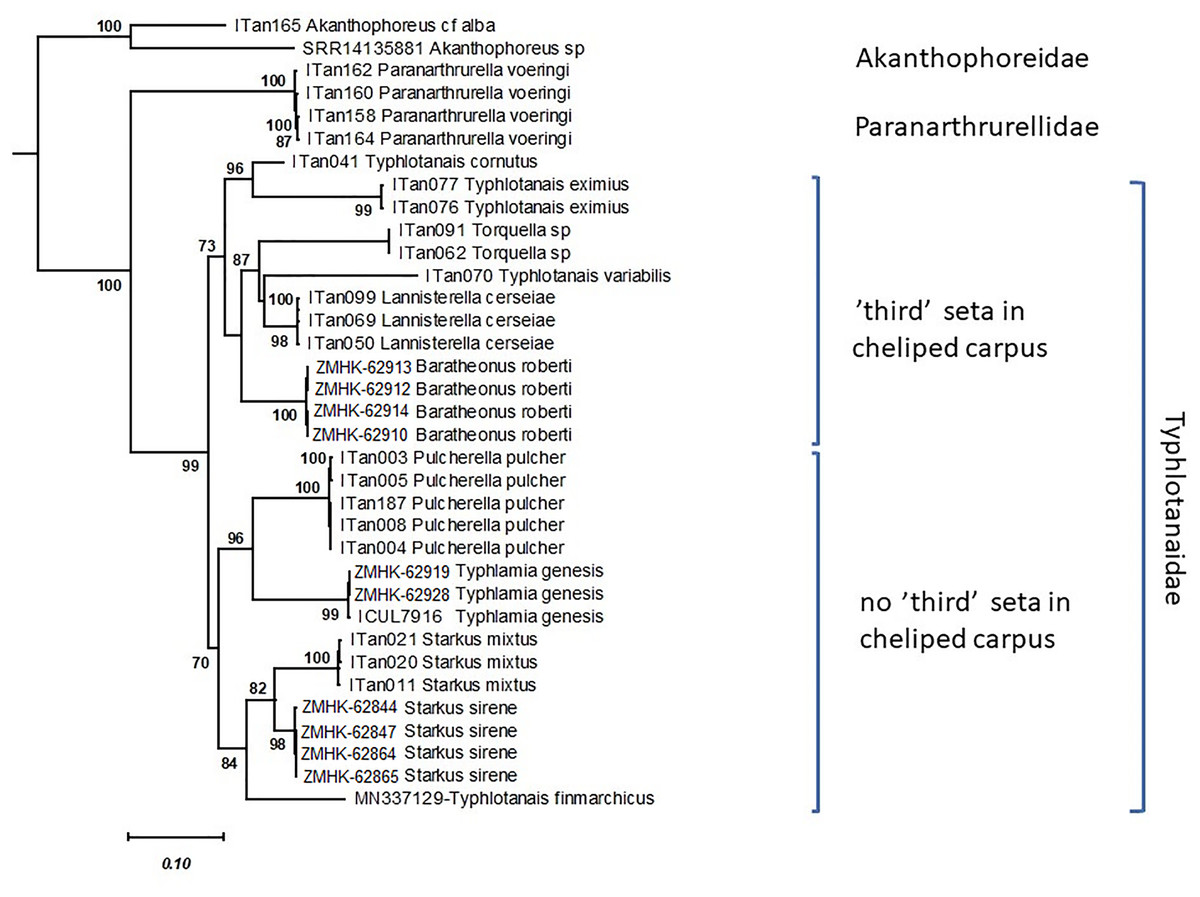
Figure 3: Maximum likelihood evolutionary tree of Typhlotanaidae species inferred from the COI and 18S concatenated alignment.
Only statistically significant bootstrap values (>70%) are shown. Modified after Gellert, Palero & Błażewicz, 2022.{kind=link}
Of the ‘long-bodied’ Typhlotanaidae with a developed clinging apparatus on pereopods 4–6 previously known, only a few taxa have rounded pereonite margins. These include the three genera—Torquella, Pulcherella and Typhlamia, and two Typhlotanais species which have very conservative mouth parts, but share several unique morphological characters—Ty. variabilis Hansen, 1913 and Ty. incognitus Larsen, Błażewicz-Paszkowycz & Cunha, 2006 (‘variabilis’ morpho-group) (Hansen, 1913; Larsen, Błażewicz-Paszkowycz & Cunha, 2006). The morphological features that allow these genera and species-group to be distinguished from Lannisterella n. gen. are summarized in Table 1, and Figs. 1, 2 and 4.
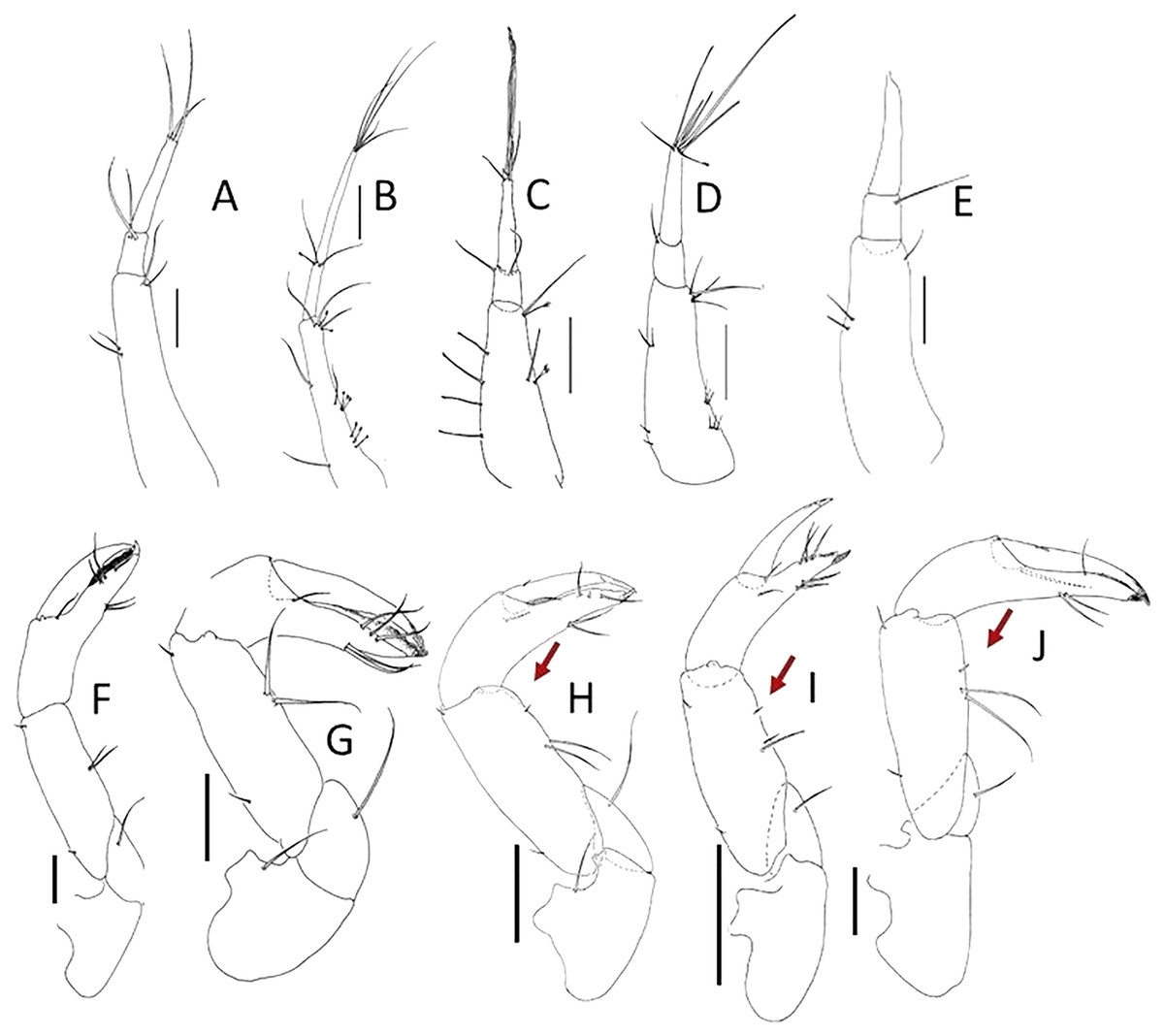
Figure 4: Comparison of antennule and cheliped.
(A, F) Pulcherella pulcher (Hansen, 1913). (B, G) Typhlamia collinsi nom. nov. (C, H) Lannisterella cerseiae n. sp. (D, I) ‘variabilis’ group: Typhlotanais incognitus Larsen, Błażewicz-Paszkowycz & Cunha, 2006. (E, J) Torquella longisetosa (Kudinova-Pasternak, 1990) (from: Błażewicz-Paszkowycz, 2007a; Larsen, Błażewicz-Paszkowycz & Cunha, 2006). Scale: 0.1 mm.{kind=link}
Taxonomy
Genus Typhlamia Błażewicz-Paszkowycz, 2007a
Diagnosis (Błażewicz-Paszkowycz, 2007a; amended): Body long, about 8.0–9.0 L:W. Carapace tapering proximally, short (1.0–1.2 L:W). Proximal corners not extended; lateral seta absent. Antennule inner margin with two medial and one distal setae; article-3 long; distal spur absent. Maxilliped basal seta long. Cheliped carpus ventral margin straight; ‘third’ seta absent. Pereopod-1 carpus dorsodistal with long seta; pereopods 2–3 propodus with long distodorsal seta, carpus with setae and spine(s); pereopods 4–6 carpus with two rounded/blunt cusps instead of prickly tubercles; pereopods 4–6 unguis bifurcated. Uropod slender; exopod one-articled.
Type species: Typhlamia bella Błażewicz-Paszkowycz, 2007a, by designation.
Species included: Typhlamia bella Błażewicz-Paszkowycz, 2007a; Typhlamia genesis Gellert, Palero & Błażewicz, 2022, Typhlamia mucronata (Hansen, 1913), Typhlamia collinsi Błażewicz nom. nov.
Remarks: Of all the ‘long-bodied’ of Typhlotanaidae with rounded pereonite margins, only Typhlamia and Pulcherella have slender uropods with one-articled exopod (Table 1, Figs. 2F, 2G). Furthermore, both genera have long dorsodistal seta on the pereopod-1 carpus. In contrast to Pulcherella, members of Typhlamia have long third article in the antennule provided with long distal setae (Figs. 4A, 4B).
Typhlamia bella Błażewicz-Paszkowycz, 2007a
Typhlamia bella—Błażewicz-Paszkowycz (2007a): 6, 87, 90, 92, 94–98.
Distribution: Species only known from type locality: Weddell Sea, Antarctic (70°32.02′–70°32.19′S 14°35.05′–14°35.13′W) from 4,392–4,385 m depth, which is the deepest record of this species in the Atlantic. Another, much shallower record from 62°31.47′–62°31.36′S 64°39.45′–64°39.25′W at 3,804–3,808 m depth is doubtful (see Błażewicz-Paszkowycz, 2007a).
Typhlamia mucronata (Hansen, 1913)
Typhlotanais mucronatus—Hansen (1913): 37, 131, 132, plate 4, 5; Nierstrasz (1913): 38; Stephensen (1932): 350; Shiino (1970): 275, 289; Holdich & Bird (1985): 445; Larsen, Błażewicz-Paszkowycz & Cunha (2006): 25; Kudinova-Pasternak (1985): 63; Kudinova-Pasternak (1990): 94.
Typhlamia mucronata—Błażewicz-Paszkowycz (2007a): 6, 27, 87–90.
not Typhlotanais mucronatus—Kudinova-Pasternak (1985): 52, 53.
Remarks: Typhlamia mucronata was redescribed by Błażewicz-Paszkowycz (2007a) based on Hansen’s collection from HMS Ingolf cruise (syntypes: CRU 3916, CRU 7423, CRU 3885) held in the Natural History Museum of Copenhagen.
Distribution: North of Iceland (67°29′N 11°32′W and 67°53′N 10°19′W) and Jan Mayen (69°13′N 8°23′W) from 1,618–1,847 m depth (Hansen, 1913; Lang, 1970); Bay of Biscay (47°33.90′N 09°38.40′W) from 3,992–4,260 m depth (Holdich & Bird, 1985), and Walvia Seamount (25°37′6″S 5°19′2″E) from 2,270 m depth (Kudinova-Pasternak, 1990).
Typhlamia collinsi Błażewicz nom. nov.
LSID urn:lsid:zoobank.org:act:EC9D55F0-683F-4F0B-8487-ECEC22A33790
Typhlotanais mucronatus—Kudinova-Pasternak (1985): 52, 53.
Typhlamia sandersi (Kudinova-Pasternak, 1985)—Błażewicz-Paszkowycz (2007a): 6, 87, 90–93.
Remarks: Typhlamia collinsi description was based on a single specimen deposited at the Zoological Museum in Moscow (Błażewicz-Paszkowycz, 2007a) (see results section).
Distribution: Species known only from the type locality at Great-Meteor Seamount—North Atlantic (29°50′N 28°08′6E) at depths of 3,080–3,140 m (Kudinova-Pasternak, 1985).
Typhlamia genesis Gellert, Palero & Błażewicz (2022)
Typhlotanais mucronatus—Kudinova-Pasternak (1970): 348, 379–380; Kudinova-Pasternak (1973): 153.
Typhlamia mucronata—Błażewicz-Paszkowycz (2007b): 102; Stępień, Pabis & Błażewicz (2019): 178: 3; Błażewicz et al. (2020): 474, 485–487.
Typhlamia genesis—Gellert, Palero & Błażewicz (2022).
Distribution: Species known from North Pacific waters. Reports from Kurile-Kamchatka Trench by Kudinova-Pasternak (1970) and Gellert, Palero & Błażewicz (2022) at depths from 4,690 to 6,740 m, and in much shallower waters, between 2,340–3,450 m, in the Gulf of Alaska. The depth range for Typhlamia genesis (as Typhlotanais mucronatus) given by Kudinova-Pasternak (1970) must be incorrect (4,840–6,675 m), but this mistake was repeated in latter papers (Błażewicz-Paszkowycz, 2007b; Stępień, Pabis & Błażewicz, 2019; Błażewicz et al., 2020) (Table 3).
Genus Lannisterella n. gen.
LSID urn:lsid:zoobank.org:pub:508FFE15-7F76-4125-9485-D7974A93BCD9
Diagnosis: Body long (7.0–8.0 L:W). Carapace elongated (1.3 L:W). Proximal corners extended (form ‘collar’); pereonite-1 lateral seta present. Antennule inner margin with four long setae*; article-3 short; distal spur present. Maxilliped basal seta short. Cheliped ventral margin rounded, with small ‘third’ seta. Pereopod-1 carpus short dorsodistal seta; pereopods 2–3 propodus distodorsal seta long; carpus with setae and spine(s); pereopods 4–6 carpus with clinging apparatus surrounded by blunt spines; pereopods 4–6 unguis bifurcated*. Uropod slender; exopod two-articled.
*Figures by Kudinova-Pasternak (1985) do not show setae on antennules, one of the carpal setae is longer than the others, unguis is simple.
Etymology: Named after the Lannister family, one of the Great Houses of Seven Kingdoms, from George R. R. Martin’s novel, Game of Thrones.
Type species: Lannisterella cerseiae n. sp.
Gender: feminine.
Species included: Lannisterella angusticheles (Kudinova-Pasternak, 1989); Lannisterella sandersi (Kudinova-Pasternak, 1985); Lannisterella cerseiae n. sp.
Remarks: The genus Lannisterella is a ‘long-bodied’ typhlotanaid with rounded pereonite margins and 2-article uropods rami (Figs. 1D and 2H). Lannisterella has rounded (not parallel) and smooth (not corrugated) pereonite margins like Typhlamia, Torquella and two species from the genus Typhlotanais (‘variabilis’ group: Typhlotanais variabilis and Typhlotanais incognitus, see below). Lannisterella has uropodal exopod with two articles (one uropod exopod in Typhlamia and Pulcherella).
Lannisterella, Torquella and the ‘variabilis’ group have similar uropods. The ‘variabilis’ group is distinguished by clearly separated pereonites 1–3 (Fig. 1E), lacking the distal spur in antennule article-3 (Fig. 2D), short dorsodistal seta in pereopods 2–3 propodus and simple clinging apparatus on pereopods 4–6 (no blunt spines surrounding prickly tubercles).
Those characters allow to distinguish members of the ‘variabilis’ group from Lannisterella and Torquella, which pereonites 1–3 are not separated (Figs. 1D, 1F), antennule have distal spur (Figs. 4C, 4E), pereopods 2–3 propodus have a long dorsodistal seta and pereopods 4–6 carpal prickly tubercles are surrounded by blunt spines. Lannisterella can be distinguished from Torquella by the presence of four distinct setae on antennule article-1 (Fig. 4C), pereopods 1–3 lack rod setae, and pereopods 4–6 have simple unguis. In contrast, Torquella has few medial and distal setae on antennule article-1 (Fig. 4E), pereopods 1–3 articles bear distinct rod setae and pereopods 4–6 have bifurcated unguis.
Lannisterella angusticheles (Kudinova-Pasternak, 1989) n. comb.
Peraeospinosus angusticheles—Kudinova-Pasternak (1989): 28, 30–33.
Typhlotanais angusticheles—Błażewicz-Paszkowycz (2007a): 5, 46; Błażewicz-Paszkowycz et al. (2014): 443.
Diagnosis: Antennule article-3 subequal to pereonite-2. Cheliped carpus short (2.1 L:W). Antennule article-3 long (10 L:W) (see Table 4). Pereopod-1 with three dorsodistal setae.
| Species | L. cerseiae | L. sandersi | L. angusticheles |
|---|---|---|---|
| Antennule article-3 | Short (7.0 L:W) | Short (6.2 L:W) | Long (10 L:W) |
| Cheliped carpus | Long (2.6 L:W) | Long (2.7 L:W) | Short (2.1 L:W) |
| Pereonite-1 to pereonite-2 | Subequal | Shorter | Subequal |
| Pereopod-1 propodus with dorsodistal seta | One | Two | Three |
| Pereopod-2 carpus ornamentation | Two setae and one spine | Six setae and one spine | Data not available |
| Uropod endopod | 6.0 L:W | 8.0 L:W | Data not available |
Remarks: Lannisterella angusticheles is distinguished from other members of the genus Lannisterella by having cheliped carpus 2.1 L:W (2.7 L:W in L. sandersi and 2.6 L:W in L. cerseiae). Pereopod-1 with three dorsodistal setae in L. angusticheles, two and one respectively in L. sandersi and L. cerseiae.
Lannisterella sandersi (Kudinova-Pasternak, 1985) n. comb.
Synonyms: Typhlotanais sandersi—Kudinova-Pasternak (1985) 120: 52–55, Fig. 1.
Diagnosis: Antennule article-3 shorter than pereonite-2. Cheliped carpus long (2.7 L:W). Antennule article-3 short (6.2 L:W) (see Table 4). Pereopod-1 with two dorsodistal setae. Pereopod-2 carpus with six setae and one spine. Uropod endopod 8.0 L:W.
Remarks: Lannisterella sandersi is distinguished from its two congeners, by a relatively short (7.0 L:W) antennule article-3 (10 L:W in L. angusticheles), long (8.0 L:W) uropodal endopod (6.0 L:W in L. cerseiae) and presence of six setae in pereopod-2 carpus (two setae in L. cerseiae).
Lannisterella cerseiae n. sp.

Figure 5: Lannisterella cerseiae n. sp., neuter.
(A) Dorsal view. (B) Lateral view. (C) Antennule. (D) Antenna. (E) Labrum. (F) Right mandible. (G) Maxillule endite tip. (H) Maxilliped. (I) Labium. (J) Epignath. (K) Uropod. Scale: A–B = 1 mm, C–K = 0.1 mm (from Błażewicz-Paszkowycz et al., 2014).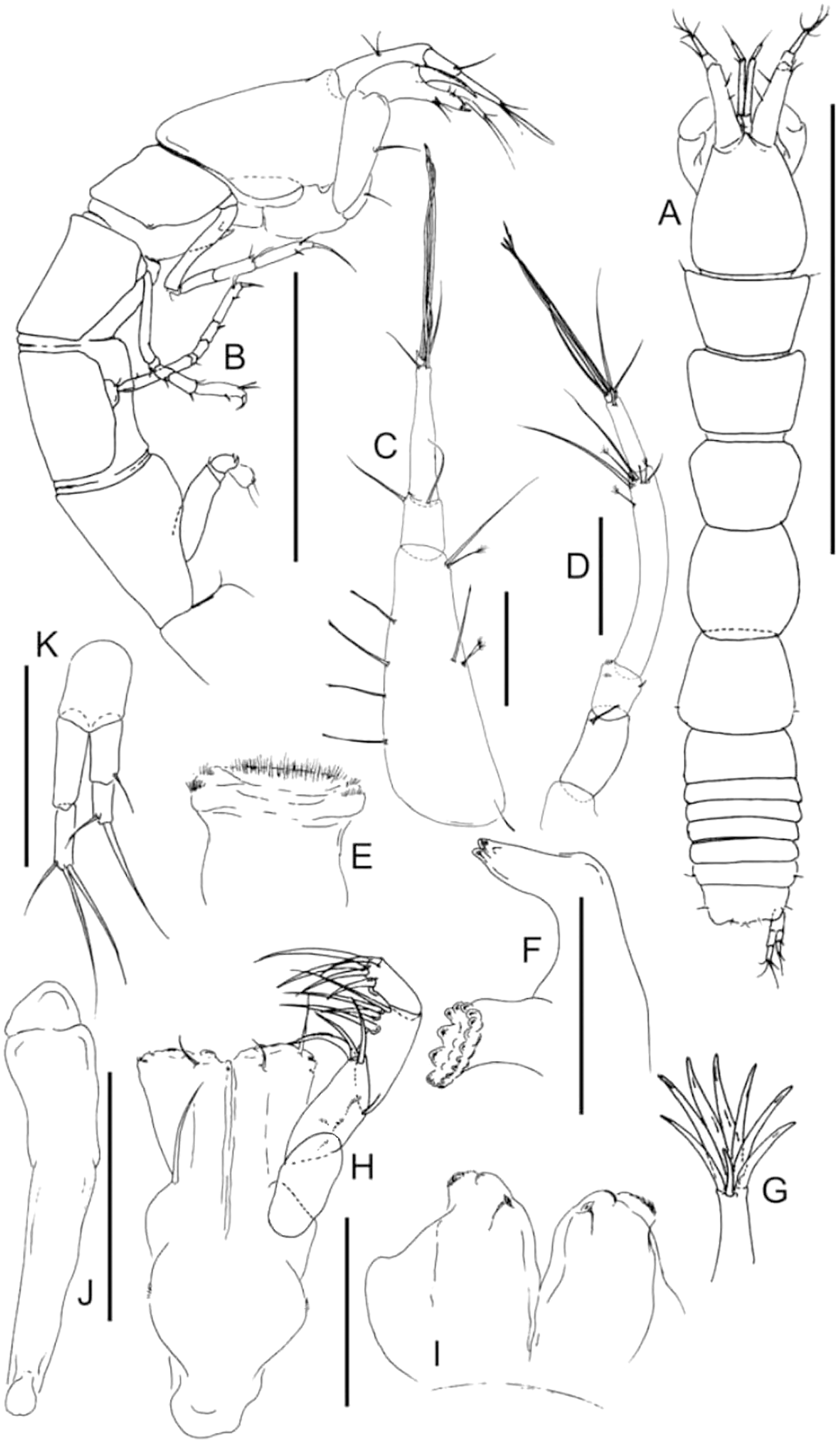{kind=link}
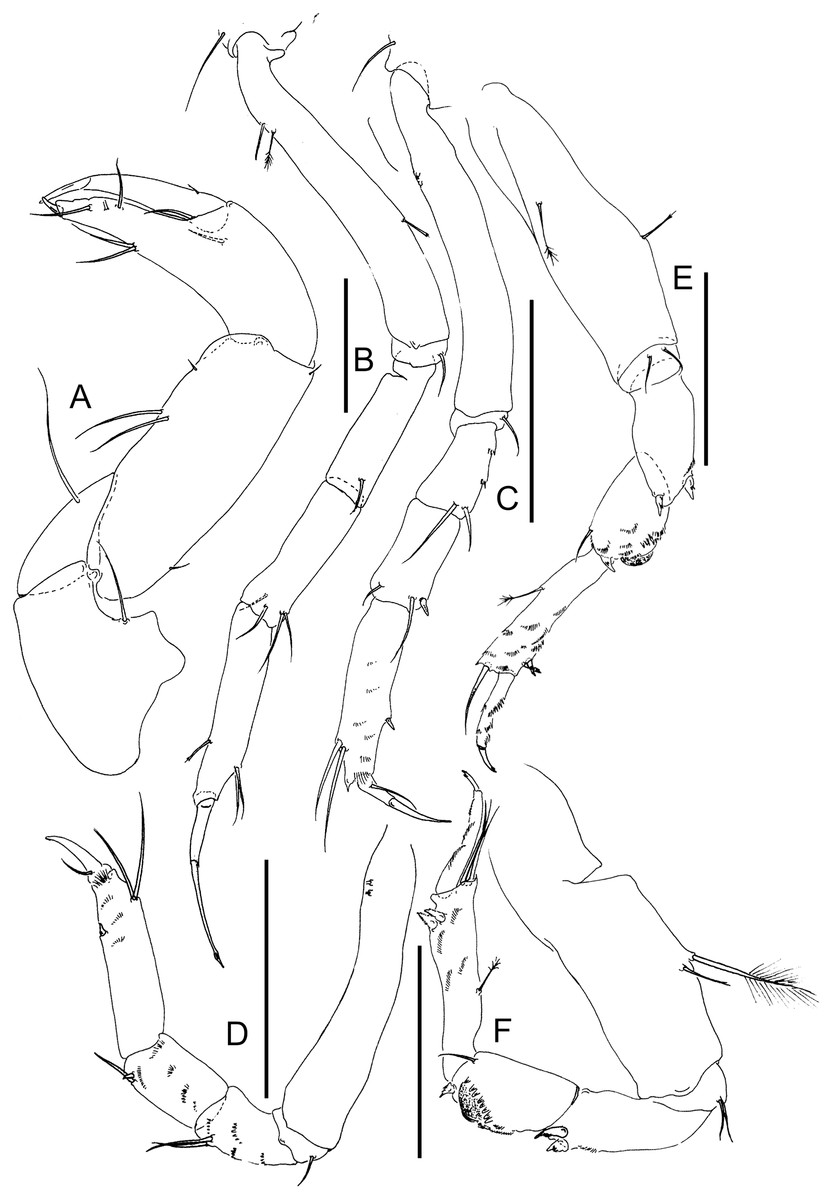
Figure 6: Lannisterella cerseiae n. sp., neuter.
(A) Cheliped. (B) Pereopod-1. (C) Pereopod-2. (D) Pereopod-3. (E) Pereopod-4. (F) Pereopod-6. Scale: = 0.1 mm (from Błażewicz-Paszkowycz et al., 2014).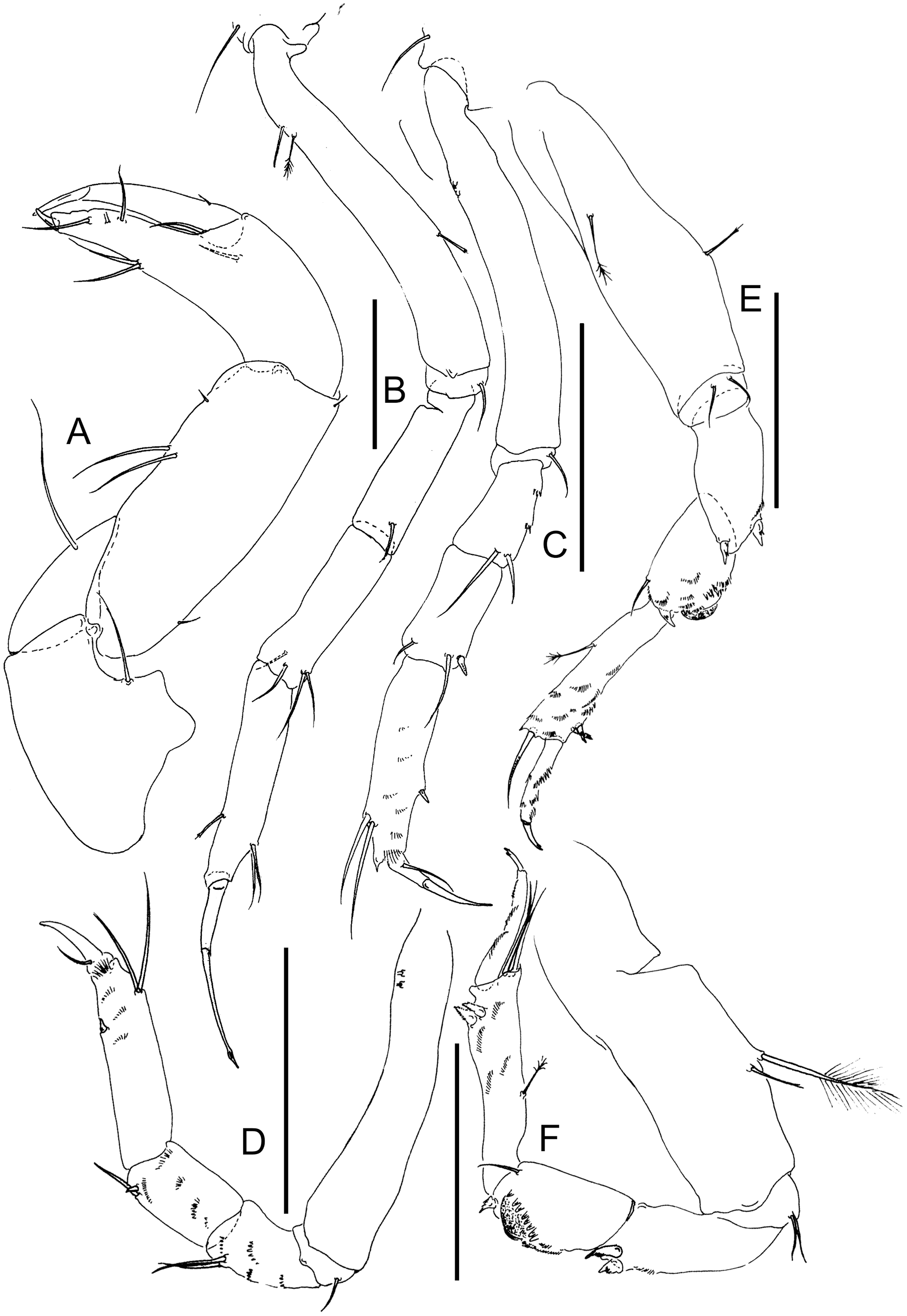{kind=link}
LSID urn:lsid:zoobank.org:pub:508FFE15-7F76-4125-9485-D7974A93BCD9
Typhlotanais sp. A: Błażewicz-Paszkowycz et al. (2014): 418, 421–423, 443–449.
Studied material:
Holotype, neuter (ZMH-K44196), IceAGE St. 1072, 63°00.46′–63°01.10′N 28°04.09′–28°03.15′W, epibenthic sledge, depth 1,569–1,594 m, 9 September 2011.
Paratype: neuter (ZMH-K44194), the locality as for holotype; mature (swimming) male (ZMH-K44190), the locality as for holotype.
Diagnosis: Antennule article-3 subequal pereonite-2. Cheliped carpus 2.6 L:W. Antennule article-3 short (7.0 L:W). Pereopod-1 with one dorsodistal seta. Pereopod-2 carpus with two setae and one spine. Uropod endopod 6.0 L:W.
Etymology: Named after Cersei Lannister, Queen of the Seven Kingdoms from George R.R. Martin’s novel, Game of Thrones.
Description of neuter, length 1.7 mm, paratype (ZMH-K44196). Body (Figs. 5A, 5B) slender, 7.7 L:W. Cephalothorax 1.2 L:W, 1.6× pereonite-1; eyes absent. Pereonites 1−6: 0.7, 0.8, 1.0, 1.0, 0.9 and 0.5 L:W, respectively. Pereonite-1 trapezoidal, subequal pereonite-2, with distinct seta on each side; pereonite-2 rectangular, 0.8× pereonite-3; pereonite-3 square, 0.9× pereonite-4; pereonite-4 square, 1.1× pereonite-5; pereonite-5 trapezoidal, 2.0× pereonite-6; pereonite-6 trapezoidal. Pleon 0.2× total BL; pleonites 1−5: all the same size—0.2 L:W. Pleotelson 2.8× pereonite-6.
Antennule (Fig. 5C) as long as cephalothorax; article-1, 0.7 of antennule length, 3.0 L:W, with four setae along the article, two penicillate setae and one medial long and penicillate and long setae distally; article-2, 1.4 L:W, 0.2× article-1, with two distal setae (inner and outer setae); article-3, 7.0 L:W, 2.3× article-2, with two short and five long setae distally.
Antenna (Fig. 5D) six-articled; article-1 fused with the body; article-2, 2.4 L:W, with distal seta; article-3, 1.4 L:W, 0.6× article-2, with short distal seta; article-4, 6.3 L:W, 3.6× article-3, with two short, two long (longer than article-5) and three penicillate setae distally; article-5, 5.0 L:W, 0.4× article-4, with long seta; article-6 1.0 L:W, with one short and four long distal setae.
Mouthparts. Labrum (Fig. 5E) hood-shaped, with rounded, setose distal margin. Right mandible (Fig. 5F) incisor with two lobes without lacinia mobilis. Labium (Fig. 5I) with distolateral corner finely setose. Maxillule (Fig. 5G) endite with eight terminal spines, one innermost spine shorter than the others. Epignath (Fig. 5J) simple distally. Maxilliped (Fig. 5H) basis with long seta, shorter than endites; endites unfussed, with two small gustatory cusps, seta on outer margin and seta on inner margin; palp with four articles: article-1 naked; article-2 with three inner setae and microtrichia on outer margin and outer seta; article-3 with four inner setae; article-4 with six inner distal and outer setae. Left mandible and maxilla lost during the dissection.
Cheliped (Fig. 6A) basis separated from pereonite-1, slender; 1.9 L:W, with long seta; merus subtriangular, with long ventral seta; carpus 2.7 L:W, with two long setae and short seta ventrally, two short setae on dorsal margin—dorsodistal rod seta and subdistal seta; chela 1.8× carpus, 4.8 L:W; palm 0.7× fixed finger, with two setae near dactylus insertion (inner and outer surface); fixed finger with two long ventral setae; cutting edge with three setae and three very weak, blunt cusps distally; dactylus with a short dorsoproximal seta.
Pereopod-1 (Fig. 6B) walking type, overall 22.5 L:W; coxa with seta; basis 7.8 L:W with three (subproximal, penicillate and rod medial) setae; ischium with ventral seta; merus 3.3 L:W with one ventrodistal seta; carpus 3.7 L:W; 0.8× merus, with four setae distally; propodus 3.0 L:W, 1.2× carpus, with one rod dorsodistal and two ventrodistal setae; dactylus 0.7× unguis; dactylus and unguis 0.6× propodus; unguis simple.
Pereopod-2 (Fig. 6C) walking type; overall 17.5 L:W; coxa with seta; basis naked; ischium with ventral seta; merus 1.9 L:W, with two ventrodistal setae and numerous calcified microtrichia; carpus 1.8 L:W, 0.9× merus, with dorsodistal seta and long seta and spine ventrodistally; propodus 4.2 L:W, 1.2× merus and carpus combined, with two dorsodistal setae and ventrodistal spine; dactylus 2.0× unguis; dactylus and unguis together 0.4× propodus; unguis simple.
Pereopod-3 (Fig. 6D) walking type, overall 15.8 L:W; coxa with seta; basis 6.0 L:W, naked; ischium with ventral seta; merus 1.9 L:W, with two long ventrodistal setae and numerous calcified microtrichia; carpus 1.8 L:W, 0.9× merus, with one spine and seta ventrodistally; propodus 4.2 L:W, 1.8× carpus, with ventrodistal spine, two dorsodistal setae, and numerous microtrichia; dactylus 2.0× unguis, with seta; dactylus and unguis together 0.4× propodus; unguis simple.
Pereopod-4 (Fig. 6E) clinging type, overall 7.8 L:W; coxa absent; basis slender, 4.2 L:W, with penicillate and rod medial seta; ischium with two ventral setae; merus 2.5 L:W, with two spines distally; carpus 1.8 L:W, 0.8× merus, with prickly tubercles moderate size, surrounded by spines and dorsal seta and rod seta distally; propodus 4.4 L:W, with numerous calcified microtrichia, two ventrodistal spines, and penicillate seta dorsally; dactylus 5.0× unguis, both combined 0.7× propodus; unguis simple.
Pereopod-5 as pereopod-4.
Pereopod-6 (Fig. 6F) as pereopod-4, but propodus with three dorsodistal setae.
Uropod (Fig. 5K) basal article 1.6 L:W; exopod two-articled, 5.2 L:W, 0.7 endopod, exopod proximal article 2.7 L:W, with distal seta; distal article 0.6× proximal, one long and one robust seta distally; endopod proximal article 3.0 L:W, naked; distal article 0.6 L:W, tipped by three setae distally.
Swimming male: The description supported with drawings of body habitus and appendages was presented in paper by Błażewicz-Paszkowycz et al. (2014): 443–449.
Remarks: Lannisterella cerseiae n. sp. has antennule article-1 3.6 L:W, what distinguishes it from L. angusticheles (2.9 L:W) and L. sandersi (5.1 L:W). Moreover, it has pereonite-1 subequal to pereonite-2, (clearly shorter in L. angusticheles) and has pereopod-1 propodus with only one dorsodistal seta (two setae in L. sandersi and three setae in L. angusticheles) (Table 4).
Distribution: Type locality off Iceland (63°00.46′–63°01.10′N 28°04.09′–28°03.15′W) at 1,569–1,594 m depth.
Identification key for neuters of Lannisterella
1. Antennule article-1 long (10 L:W). . . . . . .L. angusticheles (Kudinova-Pasternak, 1989)
Indian Ocean3600–5047 m
- Antennule article-1 short (<6.0 L:W). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2
2. Uropod endopod short (6.0 L:W). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . L. cerseiae n. sp.
Iceland (N Atlantic)1569–1594 m
- Uropod endopod long (8.0 L:W). . . . . . . . . . . . . . .L. sandersi (Kudinova-Pasternak, 1985)
Great Meteor Seamount (N Atlantic)3080–3140 m
Typhlotanais ‘variabilis’ group
Diagnosis: Carapace narrowing proximally, short (1.2 L:W). Proximal corners not extended (not form ‘collar’); pereonite-1 lateral seta absent. Antennule inner margin with short medial and distal setae; article-3 short; distal spur absent. Maxilliped basal seta short. Cheliped ventral margin straight; ‘third’ seta present. Pereopod-1 carpus short dorsodistal seta; pereopods 2–3 propodus distodorsal seta short; carpus (ornamentation) with setae only; pereopods 4–6 carpus with clinging apparatus not surrounded by blunt spines; pereopods 4–6 unguis bifurcated. Uropod slender; exopod two-articled.
Species included: Typhlotanais variabilis Hansen, 1913; Typhlotanais incognitus Larsen, Błażewicz-Paszkowycz & Cunha, 2006.
Remarks: The ‘variabilis’ group is proposed for ‘long-bodied’ of Typhlotanaidae with prickly tubercles and rounded pereonite lateral margins (Table 1). At first glance, the ‘variabilis’ group may resemble Lannisterella, which also has slender cheliped carpus with ‘third’ seta and biarticulated uropod rami, and pereonites 1–3 clearly separated by flexible articulations. By this characters, antennule without distal spur, pereonite-1 without lateral seta and pereopods 4–6 carpus with prickly tubercles not surrounded by blunt spines, the members of the morpho-group are distinguished from Lannisterella, which have antennule with distal spur, pereonite-1 with lateral seta and pereopods 4–6 carpus with prickly tubercles surrounded by blunt spines. The first article of the antennule is also less setose, being supplied with fine medial and distal setae, where Lannisterella has four long setae at this article margin.
Currently two species are included within the ‘variabilis’ morpho-group. Both species can be distinguished by length of the dorsodistal seta in pereopods 4–5 propodus, which is long in Ty. variabilis (reaches tip of unguis) and short (reaches half of the dactylus) in Ty. incognitus.
Currently Typhlotanais sensu stricto includes only the species of the genus (Ty. aequiremis). The ‘variabilis’ group can be included in Typhlotanais sensu lato. It is recognised that continued research may in the future provide the basis for the establishment of a new genus
Identification key for neuters of ‘long-bodied’ typhlotanaids with rounded pereonites
1. Uropod exopod two-articled. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .3
- Uropod exopod one-articled. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2
2. Antennule twice long as carapace and long terminal setae. . . . . . . . . . . . . . . . .Typhlamia
- Antennule as long as carapace and short terminal setae. . . . . . . . . . . . . . . . . . . . .Pulcherella
3. Pereopods 2–3 propodus with short dorsodistal seta. . . . . . . . . . . . . . . . .‘variabilis’ group
- Pereopods 2–3 propodus with long dorsodistal seta. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4
4. Pereopods 1–3 with rod setae and pereopods 4–6 unguis simple. . . . . . . . . . . .Torquella
- Pereopods 1––3 without rod setae and pereopods 4––6 unguis bifurcated. . . . . . . . . . . .Lannisterella n. gen.
Genetic data
The Maximum Likelihood tree shows that the typhlotanaid taxa analyzed can be split into two main groups based on the presence or absence of a ‘third’ seta on cheliped carpus (Fig. 3). This small seta is ontogenetically independent (present in all development stages) and observed even in old historical collections. The group without a third seta includes four genera with two subclades. The first subclade comprising Pulcherella and Typhlamia, two genera with rounded pereonite margins, long uropods with unarticulated exopod, slender cheliped, and bifurcated unguis in pereopods 4–6 (see Figs. 1, 2 and 4); and the second subclade including taxa with straight pereonite margins, e.g., Starkus mixtus (Hansen, 1913), Starkus sirene Gellert, Palero & Błażewicz, 2022 and Typhlotanais finmarchicus Sars, 1882. The group with a ‘third’ seta on the cheliped carpus includes species with ‘short-bodied’ (Typhlotanais cornutus and Typhlotanais eximius) and ‘long-bodied’, which are further split into taxa with rounded pereonite margins, two-articled uropod rami and collar-shape of pereonite-1 (Lannisterella, Torquella and ‘variabilis’ group), and Baratheonus roberti Gellert, Palero & Błażewicz, 2022 characterized by straight pereonite margins, simple pereonite-1 and unarticulated uropod rami.
Discussion
The paradigm that the deep-sea ecosystem is continuous (no topographic barriers) and stable over time resulted in the widespread acceptance of large geographic ranges for deep sea taxa (Kudinova-Pasternak, 1970, 1973; Sieg, 1986b). Nevertheless, a new concept of the deep sea as a highly diverse ecosystem has emerged (Zardus et al., 2006; Etter & Bower, 2015), acknowledging that restricted gene flow and population connectivity might redefine the general idea of widely distributed deep-sea species, particularly among Peracarida (Brandt et al., 2012; Hilário et al., 2015; Jakiel, Stępień & Błażewicz, 2018; Jakiel, Palero & Błażewicz, 2019, 2020). The consequences of insufficient knowledge about typhlotanaid taxonomy or inaccurate research methods only become obvious after state-of-the-art methods (e.g., application molecular approach or powerful microscopy) are improved and the data is re-analyzed. Our revision of ‘long-bodied’ typhlotanaids with rounded pereonites showed Kudinova-Pasternak (1985) was not able to notice the fine morphological features that differentiate Typhlamia mucronata (Hansen, 1913) (Typhlotanais mucronatus in Kudinova-Pasternak, 1985) from earlier studied Typhlamia genesis (see synonyms this article) despite both taxa occupy different oceanic basins. Historical collections, which include type material or hold rare specimens from pioneering scientific expeditions exploring the ocean floor (Beddard, 1886; Hansen, 1913) are kept in museums and treated as most valuable objects (Frutos et al., 2022). Because of their rarity, newly discovered deep-sea species are not always described at once, but they are generally drawn or photographed and described without providing a name, waiting for more material to be compared and supporting the erection of a new taxon (Błażewicz-Paszkowycz & Larsen, 2005; Kavanagh, Frutos & Sorbe, 2015; Stępień et al., 2022). Such is the case of Typhlotanais sp. A from Błażewicz-Paszkowycz et al. (2014), which was illustrated, diagnosed, and kept unnamed until more material or new evidence could support the erection of a new species. Morphological revision of ‘long-bodied’ typhlotanaids with rounded pereonites, combined with newly obtained molecular data, allowed us to describe it here as Lannisterella cerseiae n. gen., n. sp.
New, overlooked, or mislabelled taxa can be discovered while working with historical collections. Some species are described only provisionally, waiting for a subsequent revision or the discovery of a sibling species (Corbari, Frutos & Sorbe, 2019; Segadilha, Serejo & Błażewicz, 2019). During revision of high-level taxa and examination of historical collections, re-examination and checking of original labels allow to amend specimen misidentifications and/or to correct the geographical coordinates (Frutos & Sorbe, 2010).
Correct taxonomic identification is a baseline for biological research and the analyses at each level of biological organization (e.g., organism, population, community, ecosystem). Its accuracy determines quality of further analyses, e.g., phylogenetic, biogeographic or ecological, therefore, failure in taxonomic identification may turn in erroneous results and lead to wrong conclusions of next-level analyses (Frutos et al., 2022; Kürzel et al., 2022). An integrative approach combining genetic and morphological data allowed us to reconstruct historical (literature) data for material that has been lost or erroneously reported. Its interpretation in the light of new and improved knowledge also allowed us to propose changes in Typhlotanaidae systematics and clarify several taxonomic uncertainties. A phylogenetic analysis based on morphological traits was attempted. Unfortunately, due to high plasticity and divergence, it was not possible to obtain a tree with acceptable parameters (e.g., Consistency Index, Bremer Support). The molecular analysis of Typhlotanaidae taxa presented here must be taken as a preliminary result that requires further studies, including larger number of genes and taxa. While accelerated climate change and plastics are affecting even the deepest parts of the ocean, an accurate estimation and understanding of the biodiversity in this virtually unknown environment is needed before irreversible changes happen.