
I’ve been thinking about limbs lately. That’s because I was invited to participate in a symposium on that topic at the Annual Meeting of the Society of Vertebrate Paleontology this fall, and abstracts for the meeting were due this past week. Being a South American mammal aficionado, I naturally submitted an abstract about South American mammal limbs. Specifically, I’ll be discussing the feet of two groups of plant-eaters: litopterns and notoungulates, the most diverse and abundant of the native South American “ungulates.”
What makes something an ungulate? By definition, having hooves is the key characteristic. As I’ve mentioned previously, horses are the quintessential modern ungulates, though cattle and their close relatives (including sheep, goats, and a bewildering variety of antelope) are probably a close second. All of these mammals have well-developed hooves, but horses and cattle differ in one fundamental way: horses have a single hoof on each foot, whereas cows have a pair of hooves. This difference is reflected in the names of the larger groups to which these animals belong. Horses are members of the order Perissodactyla (“odd-toed” mammals) and cattle are members of the order Artiodactyla* (“even-toed” mammals).
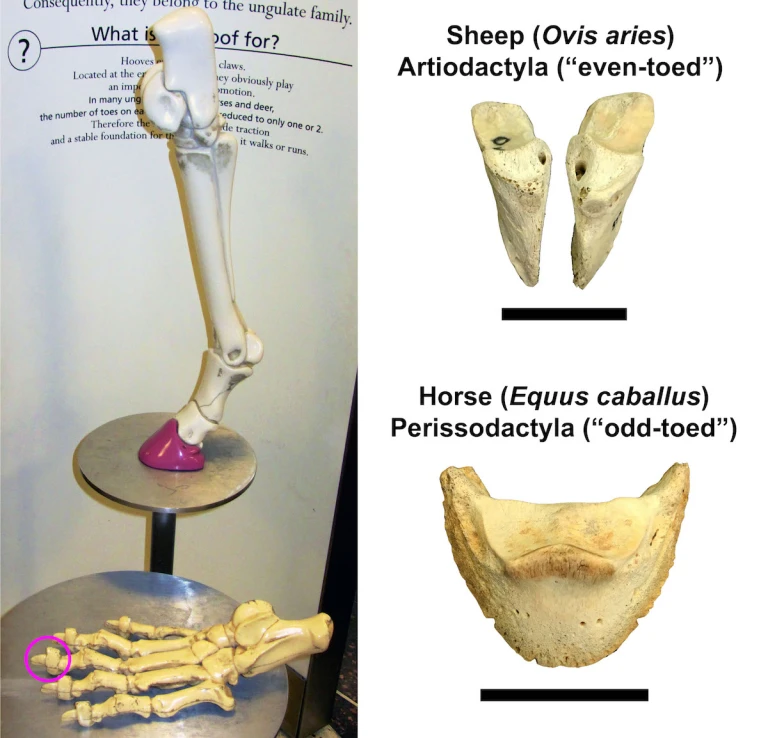
But like all things in nature, being an ungulate isn’t always so clear-cut. Modern hyraxes (order Hyracoidea) have somewhat hoof-like nails, but these aren’t nearly so developed as the hooves of most artiodactyls and perissodactyls. The same is true of elephants (order Proboscidea), which have rather pointed toes (including a sixth one) surrounded by a lot of soft tissue. Accordingly, these groups are sometimes referred to as “subungulates,” though I am sure they do not appreciate being considered something lower than a “true” ungulate.
How about native South American ungulates? (Bear in mind that when I say “native” ungulates, I’m talking about ones that have been there tens of millions of years, not Johnny-come-latelys like camels, deer, and tapirs that immigrated from North America within the past few million years.) Like modern mammals, notoungulates and litopterns varied widely in their foot structure. Some of these species would undoubtedly be classified as ungulates if they were alive today whereas others probably would not.
In overall limb structure, litopterns were more ungulate-like than notoungulates, as they apparently all had hooves. One exceptional little litoptern called Thoatherium minusculum, which was about the size of a tiny deer and lived some 18 million years ago, had very horselike feet, with only a single functional toe on each foot (digit 3, the middle toe) and tiny remnants of digits 2 and 4 on either side. Most of the other members of its family (“gracile” litopterns of the family Proterotheriidae), were slightly larger and had longer side toes that would not have touched the ground when the animal was walking. No mammal alive today has a similar arrangement, but some extinct horses did.
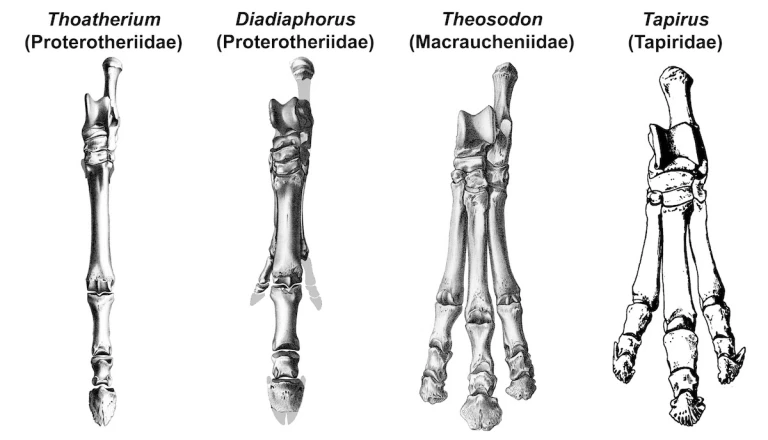
Members of the other well-known litoptern family, Macraucheniidae (robust litopterns), had side toes that were nearly as long as the middle one. Thus, all three toes would have supported the animal when it was walking. Modern rhinoceroses (family Rhinocerotidae) and tapirs (family Tapiridae) have similar hind feet. (Their forefeet can have three or four toes.) But since no other mammals living in South America at the time had a similar arrangement, fossilized mammal tracks with three toes are assumed to have been produced by members of this family. Similarly, one-toed tracks are thought to have been made by proterotheriid litopterns.

The technical term for a foot in which the weight of the animal is primarily borne by the middle (third) toe is mesaxonic. This reflects the fact that the axis of the foot passes directly through its middle. (Mes means “middle” in Greek.) Thus, litopterns were mesaxonic ungulates, just like perissodactyls. The alternative condition, in which the axis of the foot passes between two digits (the third and fourth), is known as paraxonic. (Bar means “beside” in Greek.) Cows, sheep, antelope, pigs and all other artiodactyls are paraxonic. In common parlance, they are known as are cloven-hoofed ungulates.
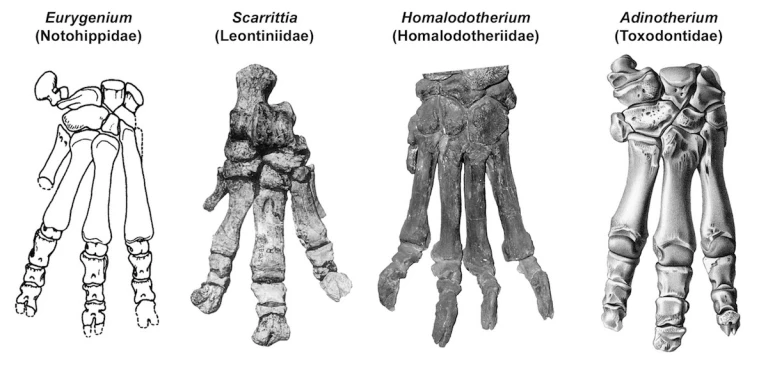
Things are a bit more complicated in notoungulates. The largest notoungulates, toxodontids, had three functional toes on each foot like most modern tapirs and rhinoceroses. And like these animals, their toes ended in hooves. Another group of large notoungulates known as leontiniids (Chaffee 1952) apparently had a similar arrangement, as did notohippids, smaller relatives of both of these groups (Shockey 1997). The outliers among the larger notoungulates are Homalodotherium and its relatives (family Homalodotheriidae), which had four large, claw-bearing digits on each foot, with the claws of the forefeet larger than those of the hind feet. These unusual notoungulates may have been able to support themselves on their hind limbs and use their long, clawed arms to reach up into trees and pull branches down within reach of their mouth – an alternative strategy to having a long neck like a giraffe. This distinctive lifestyle evolved independently in several groups of Northern Hemisphere “ungulates” that are now extinct (Coombs 1983).
Most smaller notoungulates never evolved typical hooves like those of modern ungulates. Rather, they had something between a hoof and a claw that was relatively flat, moderately broad, and sometimes had a cleft tip like the claws of many modern digging mammals (as well as some larger notoungulates; Shockey et al. 2007). But aside from those similarities, the three main groups of typotheres evolved in different directions in terms of their forefeet and hind feet. One group of typotheres known as mesotheres (family Mesotheriidae) probably were habitual diggers, and their forefeet and hind feet had four well-developed digits (only the fifth digit was reduced). The hegetotheres (family Hegetotheriidae), had somewhat mesaxonic forefeet and hind feet earlier in their evolutionary history, with a long third digit, two slightly smaller ones (second and fourth), and one that was smaller still (fifth). Later species relied primarily on just the second and third digits for weight support. This condition has been dubbed the “pseudo-paraxonic” condition because the two digits involved (second and third) are different from the weight-bearing ones in artiodactyls (third and fourth) (Simpson 1945).
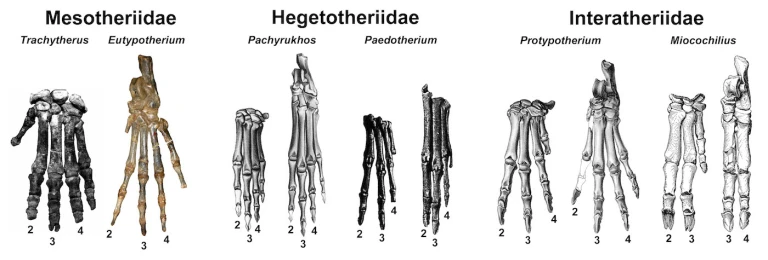
Early interatheres (family Interatheriidae) had forefeet that resembled those of early hegetotheres but more symmetrical hind feet: the third and fourth toes were similar and length as were the second and fifth (though this latter pair was shorter than the former). Thus, their forefeet were pseudo-paraxonic, but their hind feet had the “true” paraxonic condition! This trend was taken to an extreme in the interathere Miocochilius anomopodus, whose specific epithet means “irregular foot” in Greek; its side digits were reduced to a greater degree than in any other typothere, giving its feet a very piglike appearance. At the opposite end of the spectrum from Miocochilius was Interatherium robustum, which had forefeet and hind feet more like mesotheres, with four robust digits of similar size.
As you can see, although notoungulates and litopterns are both referred to as South American ungulates, the term ungulate doesn’t really tell you anything about their hooves (or lack thereof). It also doesn’t give you an appreciation of the diversity of their feet, which varied greatly not only in the number of toes on each foot, but also in whether those toes ended in hooves, claws, or something in between. Neither does the term ungulate tell you anything about the evolutionary relationships of these groups. Some studies (e.g., Welker et al. 2015) have concluded that notoungulates and litopterns are closely related to horses, rhinos, and other “true” ungulates, but other studies (e.g., O’Leary et al. 2013) have concluded that they are more closely to African “sub-ungulates” such as elephants and hyraxes. On top of that, notoungulates and litopterns may not even be closely related to one another.
So why do we refer to these animals as ungulates? It seems that ungulate is the best term we currently have to refer to medium-to-large, plant-eating mammals that don’t climb trees and aren’t rodents (nor marsupials) – characteristics that aren’t tightly linked to the ends of their feet. But even though “ungulate” may not be the most accurate descriptor of these animals, for me, notoungulates and litopterns – rather than llamas and tapirs – will always be South America’s true ungulates.
* I know the name Cetartiodactyla exists. I just choose not to use it.
References Cited
- Chaffee, R. G. 1952. The Deseadan vertebrate fauna of the Scarritt Pocket, Patagonia. Bulletin of the American Museum of Natural History 98:509-562.
- Coombs, M. C. 1983. Large mammalian clawed herbivores: a comparative study. Transactions of the American Philosophical Society, New Series 73:1-96.
- Elissamburu, A. 2010. Estudio biomecánico y morfofuncional del esqueleto apendicular de Homalodotherium Flower 1873 (Mammalia, Notoungulata). Ameghiniana 47:25-43.
- Kraglievich, J. L. 1926. Sobre el conducto humeral en las vizcachas y paquirucos Chapadmalenses con descripción del Paedotherium imperforatum. Anales del Museo Nacional de Historia Natural “Bernardino Rivadavia” 34:45-88.
- Krapovickas, V., P. L. Ciccioli, M. G. Mángano, C. A. Marsicano, and C. O. Limarino. 2009. Paleobiology and paleoecology of an arid-semiarid Miocene South American ichnofauna in anastomosed fluvial deposits. Palaeogeography, Palaeoclimatology, Palaeoecology 284:129-152.
- O’Leary, M. A., J. I. Bloch, J. J. Flynn, T. J. Gaudin, A. Giallombardo, N. P. Giannini, S. L. Goldberg, B. P. Kraatz, Z.-X. Luo, J. Meng, X. Ni, M. J. Novacek, F. A. Perini, Z. S. Randall, G. W. Rougier, E. J. Sargis, M. T. Silcox, N. B. Simmons, M. Spaulding, P. l. M. Velazco, M. Weksler, J. R. Wible, and A. L. Cirranello. 2013. The placental mammal ancestor and the post-K-Pg radiation of placentals. Science 339:662-667.
- Scott, W. B. 1910. Mammalia of the Santa Cruz Beds. Volume VII, Paleontology. Part I, Litopterna; pp. 1-156 in W. B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896-1899. Princeton University, E. Schweizerbart’sche Verlagshandlung (E. Nägele), Stuttgart.
- Scott, W. B. 1912. Mammalia of the Santa Cruz Beds. Volume VI, Paleontology. Part II, Toxodonta; pp. 111-238 in W. B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896-1899. Princeton University, E. Schweizerbart’sche Verlagshandlung (E. Nägele), Stuttgart.
- Scott, W. B. 1937. A History of Land Mammals in the Western Hemisphere, 2nd Ed. MacMillan Co., New York, 786 pp.
- Shockey, B. J. 1997. Two new notoungulates (Family Notohippidae) from the Salla Beds of Bolivia (Deseadan: late Oligocene): systematics and functional morphology. Journal of Vertebrate Paleontology 17:584-599.
- Shockey, B. J., D. A. Croft, and F. Anaya. 2007. Analysis of function in the absence of extant functional analogs: a case study of mesotheriid notoungulates. Paleobiology 33:227-247.
- Simpson, G. G. 1945. A Deseado hegetothere from Patagonia. American Journal of Science 243:550-564.
- Sinclair, W. J. 1909. Mammalia of the Santa Cruz Beds. Volume VI, Paleontology. Part I, Typotheria; pp. 1-110 in W. B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896-1899. Princeton University, E. Schweizerbart’sche Verlagshandlung (E. Nägele), Stuttgart.
- Stirton, R. A. 1953. A new genus of interatheres from the Miocene of Colombia. University of California Publications in Geological Sciences 29:265-348.
- Welker, F., M. J. Collins, J. A. Thomas, M. Wadsley, S. Brace, E. Cappellini, S. T. Turvey, M. Reguero, J. N. Gelfo, A. Kramarz, J. Burger, J. Thomas-Oates, D. A. Ashford, P. D. Ashton, K. Rowsell, D. M. Porter, B. Kessler, R. Fischer, C. Baessmann, S. Kaspar, J. V. Olsen, P. Kiley, J. A. Elliott, C. D. Kelstrup, V. Mullin, M. Hofreiter, E. Willerslev, J.-J. Hublin, L. Orlando, I. Barnes, and R. D. E. MacPhee. 2015. Ancient proteins resolve the evolutionary history of Darwin’s South American ungulates. Nature 522:81–84.
