
Leptophis ahaetulla ( Linnaeus, 1758 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5153.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:A658ADE4-F352-4D16-9DC7-2721BCBE1EEF |
|
persistent identifier |
https://treatment.plazi.org/id/039B220B-FFFC-D15B-FF6B-959DFB76EB87 |
|
treatment provided by |
Plazi |
|
scientific name |
Leptophis ahaetulla ( Linnaeus, 1758 ) |
| status |
|
Leptophis ahaetulla ( Linnaeus, 1758)
( Figs. 1–3 View FIGURE View FIGURE 2 View FIGURE )
Coluber ahaetulla Linnaeus, 1758: 225 View Cited Treatment . Lectotype (UUZM 02; not examined), designated by Savage & Oliver (1956: 149) (cf. Lönnberg 1896, no. 02). Type locality: “Asia, America”; restricted to South America fide Savage & Oliver (1956: 148) and to Surinam fide Hoge (1960 –1962: 60).
Coluber richardi Bory de Saint-Vincent, 1823: 588 . Three syntypes unknown destination (one illustrated in Bory de Saint-Vincent, no page number) (not examined). Type locality: “Guiane” [ Guyana].
Ahaetula (sic) linnei Gray, 1831: 93 (in part). Substitute name for ahaetulla Linnaeus. In : Baron Cuvier (Ed.) 1831.
Leptophis Ahaetulla (sic)— Fitzinger 1843: 26 (in part).
Leptophis ahaetulla — Amaral 1930b: 85; Amaral 1930c: 161; Parker 1935: 520; Emsley 1977: 241 (in part); Kornacker 1999: 97 (in part); Cole et al. 2013: 496 (in part); Wallach, Williams & Boundy 2014: 372 (in part); Torres-Carvajal & Terán 2021: 2.
Leptophis liocerus (sic); Déplanche, 1858: 51.
Leptophis ahaetulla ahaetulla — Oliver 1942: 1; Beebe 1946: 32; Peters & Orejas-Miranda 1970: 161; Cunha & Nascimento 1978: 102; Cunha & Nascimento 1993: 63; Tipton 2005: 161.
Leptophis copei Oliver 1942: 7 . Holotype male (USNM 83564; examined).Type locality: Salto do Huá (0°44’41”N, 66°18’37”W), Brazil-Venezuela boundary; Pérez-Santos & Moreno 1988: 206; Albuquerque (2009: 295) placed it in the synonymy of L. a. ahaetulla .
Thalerophis richardi — Oliver 1947: 64.
Thalerophis richardi richardi — Oliver 1948: 219.
Leptophis ahaetulla [ ahaetulla ]— International Commission of Zoological Nomenclature 1958: 270.
Diagnosis. Leptophis ahaetulla can be distinguished from its congeners by the following unique combination of character states: (1) head scales not edged with black and with no black spots; (2) adult color pattern with no dark dorsal bands; (3) two dorsolateral stripes separated from each other by vertebral stripe one to one and a half scales wide in adults, at least anteriorly; (4) dorsals keeled, except first dorsal row on each side; keels not black; (5) no loreal scale; (6) ventrals 150–179 in males, 154–192 in females; (7) subcaudals 129–183 in males, 131–181 in females; (8) dorsal scales of tail with no keels; (9) maxillary teeth 24–27; (10) TL/ SVL: 95% CI = 0.649 –0.660 (n = 253); (11) small spines at first basal row of hemipenial body; (12) asulcate side of hemipenis similar to sulcate side GoogleMaps .
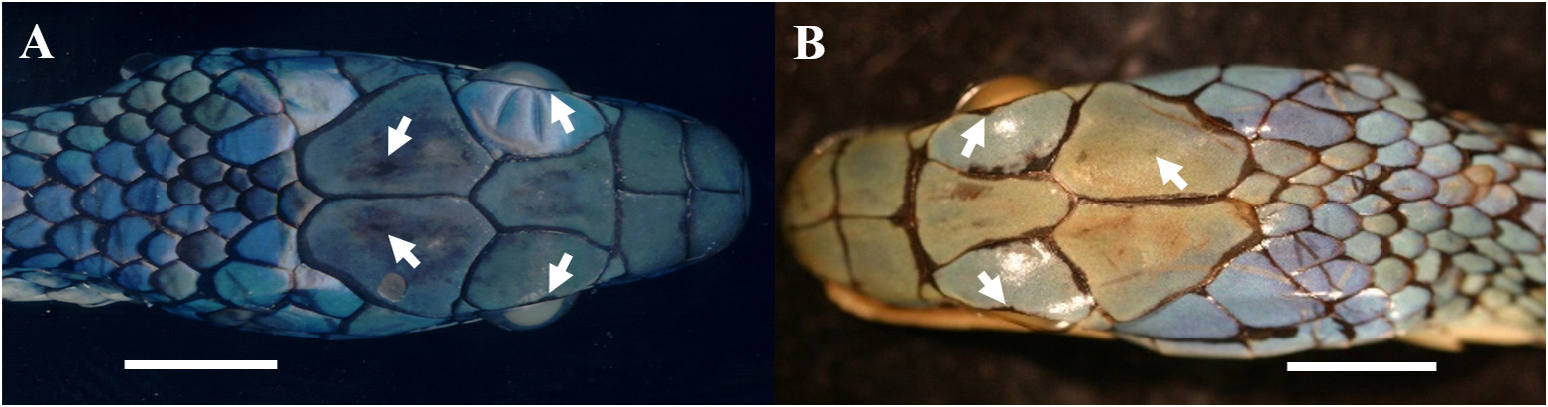
Comparisons. Leptophis ahaetulla is distinguished from all members of the L. ahaetulla complex, except L. dibernardoi , by having two dorsolateral stripes separated from each other by a vertebral stripe extending onto the tail ( Fig. 1 View FIGURE ) (vs. no dorsolateral stripes). Leptophis ahaetulla is similar to L. dibernardoi in its dorsal coloration, with both species sharing the pattern of two green dorsolateral stripes separated by a vertebral stripe, at least anteriorly. However, in life, the first (on the anterior region of trunk) to fourth (middle to posterior region) dorsal scale rows are White to Pale Sulphur Yellow (92) in L. dibernardoi (vs. second to fourth rows Sulphur Yellow [80]). The Jet Black (300) postocular stripe is wider in L. dibernardoi than in L. ahaetulla , occupying the lower half or two-thirds of the anterior temporal and most of the lower posterior temporal (vs. lower edge of the anterior temporal, with lower edge to one-third of lower posterior temporal pigmented). The snout is wider and shorter in L. dibernardoi than in L. ahaetulla . In life, the dorsal background color of L. dibernardoi is Light Emerald Green (142) to Robin’s Egg Blue (161) (vs. Light Grass Green [109] to Light Emerald Green [142] in L. ahaetulla ). Leptophis ahaetulla also differs from the partially sympatric L. coeruleodorsus by showing a vertebral stripe one to one and half scales wide (vs. three to nine scales wide) and by the higher number of ventrals for males 95% CI = 162.7–163.9, and females 95% CI = 165.6–166.8 (vs. 158.1–161 and 162.6–165.2, respectively) ( Table 2). It is further distinguished from the occasionally sympatric L. marginatus by having supracephalic scales not edged with black ( Fig. 2 View FIGURE 2 ) (vs. edged with black, Fig. 19A–B View FIGURE 19 ), and parietal and supraocular scales immaculate (vs. small black spot approximately on center of each parietal scale and usually on outer side of each supraocular, Fig. 19A–B View FIGURE 19 ). Leptophis ahaetulla differs from the sympatric L. nigromarginatus by showing coloration of vertebral stripe spreading out laterally, and occupying scales of paravertebral rows in the middle and posterior portion of the body ( Fig. 3 View FIGURE ) (vs. vertebral stripe absent, Fig. 23C–D View FIGURE 23 ), by having ventrals immaculate (vs. ventrals edged anteriorly and laterally with dark greenish blue in preservative, Fig. 23B View FIGURE 23 , or with Robin’s Egg Blue [161], Fig. 24D View FIGURE 24 ,), and by having head scales with no black spots ( Fig. 2 View FIGURE 2 ) (vs. one or two black spots on center and outer edge of each parietal and supraocular scales, respectively) ( Figs. 23A View FIGURE 23 , 24A–C View FIGURE 24 , 26 View FIGURE 26 ). Males of L. ahaetulla have a higher number of ventrals 95% CI = 162.7–163.9 than males of L. marginatus (95% CI =160.3–161.9), whereas males and females of L. ahaetulla have, respectively, a higher number of subcaudals 95% CI = 158.1–160.8 and 156.1–158.7 than males and females of L. marginatus (95% CI = 139.6–143.4 and 139.3–143.4) ( Table 2).
Variation and sexual dimorphism. Largest male SVL 1126 mm, TL 700+ mm, largest female SVL 885 mm, TL 569+ mm; ventrals 150–179 in males (163.4 ± 5.1, n = 234), 154–192 in females (166.2 ± 5.4, n = 258); subcaudals 129–183 in males (159.6 ± 9.6, n = 156), 131–181 in females (157.5 ± 9.2, n = 164); supralabials 6–10 (8.7 ± 0.5, n = 1000), with fifth–sixth (73.7%, n = 727), fourth–fifth (25.5%, n = 251), or, rarely, fourth–sixth (0.3%, n = 3) and sixth–seventh (0.5%, n = 5) bordering orbit; infralabials 8–12 (10.7 ± 0.6, n = 974), with first 6 (69.1%, n = 675), first 5 (27.8%, n = 272), first 7 (2.5%, n = 24), or, rarely, first 4 (0.5%, n = 5) and first 10 (0.2%, n = 2) contacting first chinshields; preoculars 1–2 (1.0 ± 0.2, n = 988); postoculars 1–4 (2.0 ± 0.2, n = 780); anterior temporal 1 (n = 492) and a single specimen with 2 on the right side; posterior temporal 1–3 (1.9 ± 0.2, n = 988); keels stronger in adult males than females.
In CHUNB 24750 postocular stripes extend to the level of 10 th scale row posterior to ultimate supralabial on each side. Females have more ventrals than males (F 1,490 = 35.2782; P <0.01) but males have more subcaudals than females (F 1,320 = 3.8290; P = 0.0483). The TL/SVL showed no significant difference between females and males (F 1,251 = 2.1720; P = 0.1378).
Hemipenial morphology. Eleven retracted organs examined extending up to eighth subcaudals. Everted hemipenis unilobed, noncapitate; sulcus spermaticus centrolineal, undivided, extending from base to tip of lobe; basal portion bears numerous spines distributed in 5–7 rows approximately encircling the organ; first row bears 5–8 hooked spines; spines in the first row longer than those in other rows (MNRJ 7596, MPEG 17373, UTA 15864), but in some organs (IBSP 64423, MPEG 12031, MPEG 17722) spines in the fourth row longer than most spines of first row; spine on the first row adjacent to sulcus spermaticus is the largest; several spinules widely scattered on basal portion, occurring below first row of spines and between spines; small spines adjacent to sulcus spermaticus as a continuation of spinules present on basal region; these small spines become short and stout papillae on the middle region of hemipenial body, being larger along sulcus spermaticus; papillae gradually decrease in length and number toward distal portion of hemipenis; small calyces ornamented with 7–10 papillae concentrated above most distal row of basal spines whereas large-sized calyces ornamented with 5–7 papillae concentrated along distal portion of hemipenial body; distal portion of lobe bears a few papillate calyces irregularly distributed (MPEG 18630), or nude (MNRJ 1758, MPEG 17755); asulcate side similar to sulcate side.
Coloration in life. Dorsum of the head Light Grass Green (109) to Light Emerald Green (142). Adults with two dorsolateral Light Grass Green (109) to Light Emerald Green (142) stripes separated from each other by Yellow Ocher (14), Cinnamon-Drab (50) or Sulphur Yellow (80) stripe one to one and half scale wide, at least in anterior portion of the body; usually, coloration of vertebral stripe spreads out laterally in posterior portion of the body and tail; rows II–III (occasionally II–V) on anterior portion of the body Sulphur Yellow (80); before body scale reduction from 15 to 11, rows II–III usually suffused with Yellow Ocher (14); rows II–III (occasionally II–IV or II–V) on the middle to posterior portion of the body Yellow Ocher (14) or Cinnamon-Drab (50) posteriorly; preocular Jet Black (300) stripe can be reduced only to black margin of fifth supralabial giving the impression of being absent, or extends up to fifth supralabials ( Fig. 2 View FIGURE 2 ); postocular Jet Black (300) stripe always narrow (but see Remarks below), with upper and lower postoculars ranging from immaculate to almost entirely pigmented; lower edge to two-thirds of anterior temporal pigmented, posterior temporal immaculate (occasionally lower margin pigmented), and upper edge of last three supralabials immaculate; supralabials usually white (upper edge of supralabials Light Grass Green (109) to Light Emerald Green (142) in specimens with ocular stripes reduced); infralabials, chin, and throat white; venter white in anterior portion of the body, changing gradually to tan toward tail.
Color photographs of living specimens appear in Campbell & Lamar (2004; fig. 1171), Cole et al. (2013: fig. 32A) and Fraga et al. (2013: 186), who also described the color pattern of Leptophis ahaetulla . Color descriptions are also available in Beebe (1946: 33) and Cunha & Nascimento (1993: 63).
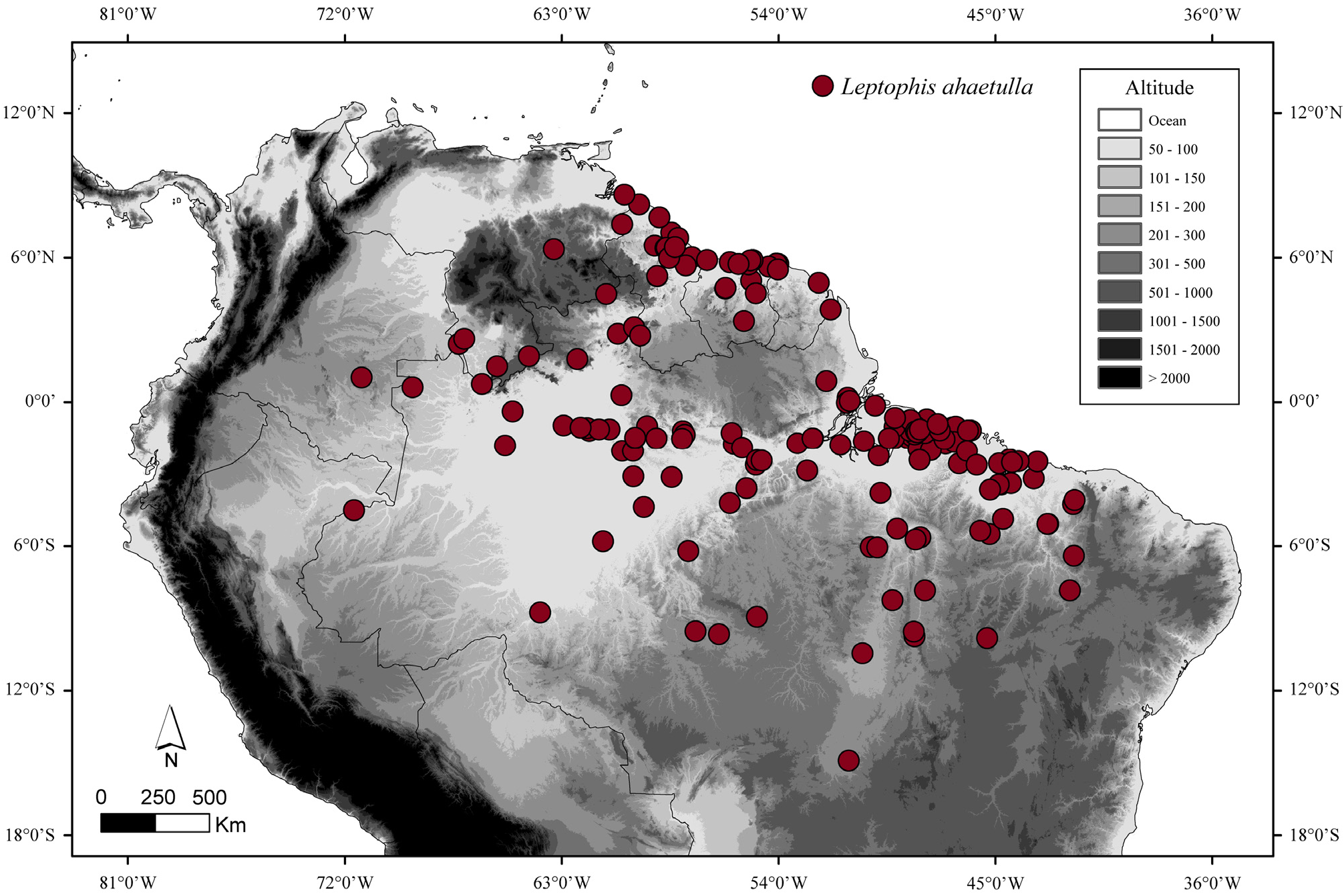
Distribution and natural history. Lowlands of northern Brazil, Guyana, Surinam, French Guiana, southwestern Venezuela and southern Colombia. These snakes were collected up to 240 m asl ( Hoogmoed 1979) ( Fig. 4 View FIGURE 4 ). Occurs mainly in the tropical and subtropical moist broadleaf forests ecoregion, as defined by Olson et al. (2001). Leptophis ahaetulla has a dietary preference for hylids, especially those of the genus Scinax Wagler ( Albuquerque et al. 2007).
Remarks. Oliver (1948) identified three specimens of Leptophis ahaetulla as juveniles, from which two showed a striped pattern like that of adults, whereas the third had a pattern of oblique bands superimposed on the striped pattern. Oliver (1948) did not mention the catalogue number of these specimens, but one of the banded specimens examined in the present study was MZUSP 12127, a female of 573 mm in total length, which has bands in the anterior and middle portion of the body. On the other hand, MPEG 1637, a female of 570 mm in total length, has a striped pattern similar to that of adults.
Although ocular stripes are usually reduced in Leptophis ahaetulla , some specimens from the municipality of Peixe, state of Tocantins, Brazil, that were previsously kept alive at Instituto Butantan, São Paulo State, Brazil, during a visit of the first author at its herpetological collection (23 Feb 2006), had a broad preocular stripe covering upper edge of rostral and nasal scales, upper edge of first, second, third and fourth supralabials, nearly all preocular, half of upper and lower postocular, two-thirds of anterior temporal, nearly all posterior temporal and upper edges of last three supralabials; this stripe extended beyond eight scales onto nuchal region ( Fig. 2F View FIGURE 2 ). Moreover, the supracephalic scales of specimens from Tocantins were Parrot Green (134) whereas their supralabials were Smoky White (261). A specimen from Pium, Goiás (CHUNB 24750), exhibited similar color pattern. These specimens are herein tentatively assigned to Leptophis ahaetulla until a more thorough study of their variation is undertaken.
Leptophis ahaetulla has a confused nomenclatural history because five specimens of two morphologically similar genera, Leptophis and Ahaetulla Link 1807 , were described under the name Coluber ahaetulla Linnaeus, 1758 . The type species, Coluber ahaetulla , was described on the basis of four specimens from South America (currently Leptophis ahaetulla ) and one specimen from Asia (currently Ahaetulla nasuta Lacépède) ( Lönnberg 1896; Andersson 1899), which led Oliver (1947) to propose the name Thalerophis as a replacement name for the South America genus Leptophis . However, Savage (1952) noted that Linnaeus (1758) had listed the ventral and subcaudal counts of only a single specimen (currently the lectotype UUZM 2) ( Fig. 5 View FIGURE ), one of the American specimens, in the original description of L. ahaetulla , corroborating the applicability of Leptophis for the South American species.
Leptophis ahaetulla sensu stricto was recently recovered as paraphyletic by Torres-Carvajal & Terán (2021) based on three vouchers from Guyana, the Brazilian state of Tocantins and “ Brazil ”, respectively. We examined one of the sequenced specimens (AMNH 140899, the Guyana specimen), and identified it as L. ahaetulla . Although these authors showed no further comments on this clade due to reduced sampling, our results corroborate the recognition of L. ahaetulla as a valid species based on the examination of about 500 specimens and also a unique combination of phenotypic (i.e., meristic, color pattern and morphometric) character states.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Leptophis ahaetulla ( Linnaeus, 1758 )
| Albuquerque, Nelson Rufino De & Fernandes, Daniel S. 2022 |
Leptophis ahaetulla
| International Commission of Zoological Nomenclature 1958: 270 |
Thalerophis richardi richardi
| Oliver, J. A. 1948: 219 |
Thalerophis richardi
| Oliver, J. A. 1947: 64 |
Leptophis ahaetulla ahaetulla
| Tipton, B. L. 2005: 161 |
| Cunha, O. R. & Nascimento, F. P. 1993: 63 |
| Cunha, O. R. & Nascimento, F. P. 1978: 102 |
| Peters, J. A. & Orejas-Miranda, B. 1970: 161 |
| Beebe, W. 1946: 32 |
| Oliver, J. A. 1942: 1 |
Leptophis copei
| Albuquerque, N. R. 2009: 295 |
| Perez-Santos, C. & Moreno, A. G. 1988: 206 |
| Oliver, J. A. 1942: 7 |
Leptophis ahaetulla
| Torres-Carvajal, O. & Teran, C. 2021: 2 |
| Wallach, V. & Williams, K. L. & Boundy, J. 2014: 372 |
| Cole, C. J. & Townsend, C. R. & Reynolds, R. P. & MacCulloch, R. D. & Lathrop, A. 2013: 496 |
| Kornacker, P. M. 1999: 97 |
| Emsley, M. G. 1977: 241 |
| Parker, H. W. 1935: 520 |
| Amaral, A. 1930: 85 |
| Amaral, A. 1930: 161 |
Leptophis liocerus
| Deplanche, M. A. 1858: 51 |
Leptophis Ahaetulla
| Fitzinger, L. 1843: 26 |
Ahaetula (sic) linnei
| Gray, J. E. 1831: 93 |
Coluber richardi
| Bory de Saint-Vincent, J. B. G. M. 1823: 588 |
Coluber ahaetulla Linnaeus, 1758: 225
| Savage, J. M. & Oliver, J. A. 1956: 149 |
| Savage, J. M. & Oliver, J. A. 1956: 148 |
| Linnaeus, C. 1758: 225 |