
Halopteris vervoorti Galea, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.1196007 |
|
persistent identifier |
https://treatment.plazi.org/id/03A487AE-FF88-2C14-FC50-89B5FEEF6E66 |
|
treatment provided by |
Plazi |
|
scientific name |
Halopteris vervoorti Galea, 2008 |
| status |
|
Halopteris vervoorti Galea, 2008
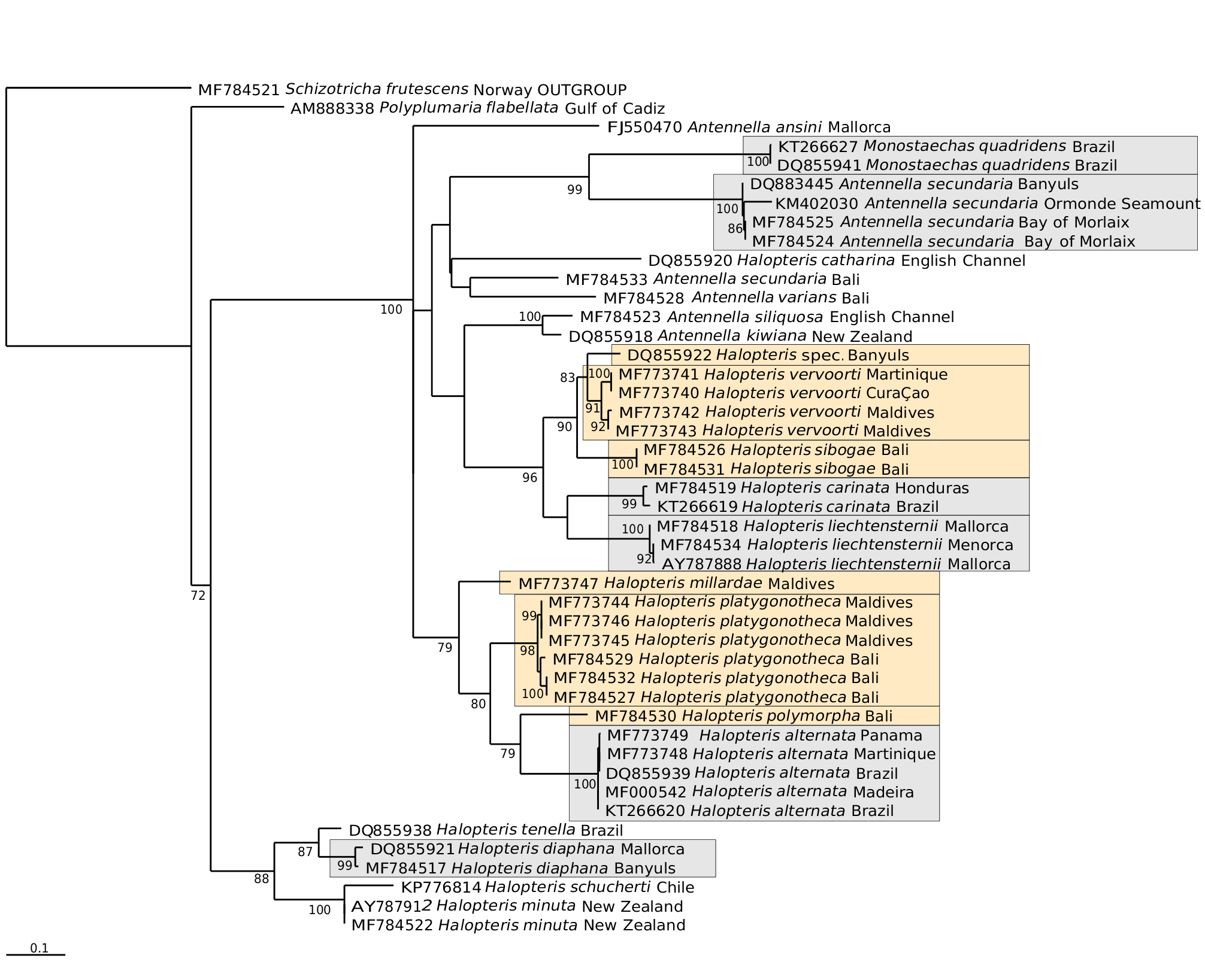
Figs 1D View Fig , 2 View Fig C-E, 5; Tables 3 View Table 3 , 4 View Table 4 ; Appendix 1
Halopteris vervoorti Galea, 2008: 42 , fig. 9; 2010: 3, 4; 2013: 50.
Halopteris polymorpha – Ansín Agís et al., 2001: 167, fig. 70. – (?) Preker & Lawn, 2010: 120 [non Plumularia polymorpha Billard, 1913 (pro parte): 24, fig. 14B, C].
Heterotheca buski – (?) Hirohito, 1974: 30, fig. 14 (incorrect subsequent spelling).
Halopteris buskii – (?) Ryland & Gibbons, 1991: 527, fig. 2.
Halopteris buski – (?) Hirohito, 1995: 244 fig. 82 (incorrect subsequent spelling).
non Plumularia buskii Bale, 1884: 125 , pl. 10 fig. 3, pl. 19 figs 34-35.
Antennella secundaria – (?) Vervoort, 1967: 42, fig. 12 [non Antennella secundaria ( Gmelin, 1791) ].
Material examined: MHNG-INVE-98635; Republic of Maldives, Faafu Atoll, Magoodhoo Island, 3.07606° 72.96159°, 3 m, coll. D. Maggioni and S. Montano; 29.01.2016; many stems, up to 1.3 cm high, some of them bearing female gonothecae; 16S sequence MF773743 View Materials . – MHNG-INVE-98636; Republic of Maldives, Faafu Atoll , 3.07478° 72.96628 °, 10 m, coll. D. Maggioni and S. Montano; 13.04.2016; 4 stems, 5-7 mm high, of which one bears two young female gonothecae; 16S sequence MF773742 View Materials . – MHNG- INVE-97952; Indonesia, Bali, Pemuteran , -8.14338° 114.65805 °, 0.5 m, coll. H. R. Galea; 04.10.2015; several sterile cormoids, up to 1.2 cm high. – MHNG- INVE-98633; Madagascar, Bay of Toliara, Dimadimatsy reef, -23.48234° 43.73285 °, 1-2 m, coll. N. Gravier- Bonnet; 13.09.1969; fertile (monoecious) colony on alga, stems up to 0.8 cm high. – HRG-0337; France, Guadeloupe, Basse-Terre, Petite Anse , 16.09639° -61.77139 °, 0-1 m, coll. H. R. Galea; 22.01.2008; numerous cormoids, up to 1.2 cm high, some bearing male gonothecae. – HRG-0897; France, Martinique, Case-Pilote , 14.64311° -61.14171 °, 5-7 m, coll. H. R. Galea; 27.02.2014; several infertile plumes, up to 1.3 cm high. – HRG-1339; France, Martinique, Case-Pilote , 14.64337° -61.14199 °, 1-2.5 m, coll. R. Ferry; 24.07.2017; numerous cormoids, up to 1.9 cm high, many bearing male gonothecae, and one their female counterpart; 16S sequence MF773741 View Materials . – DM &SM- CU005; Dutch Caribbean, Curaçao I., Piscadera Bay , 12.12139° -68.96917 °, 11 m, coll. S. Montano; 09.06.2017; two sterile stems, ca. 15 mm high; 16S sequence MF773740 View Materials .
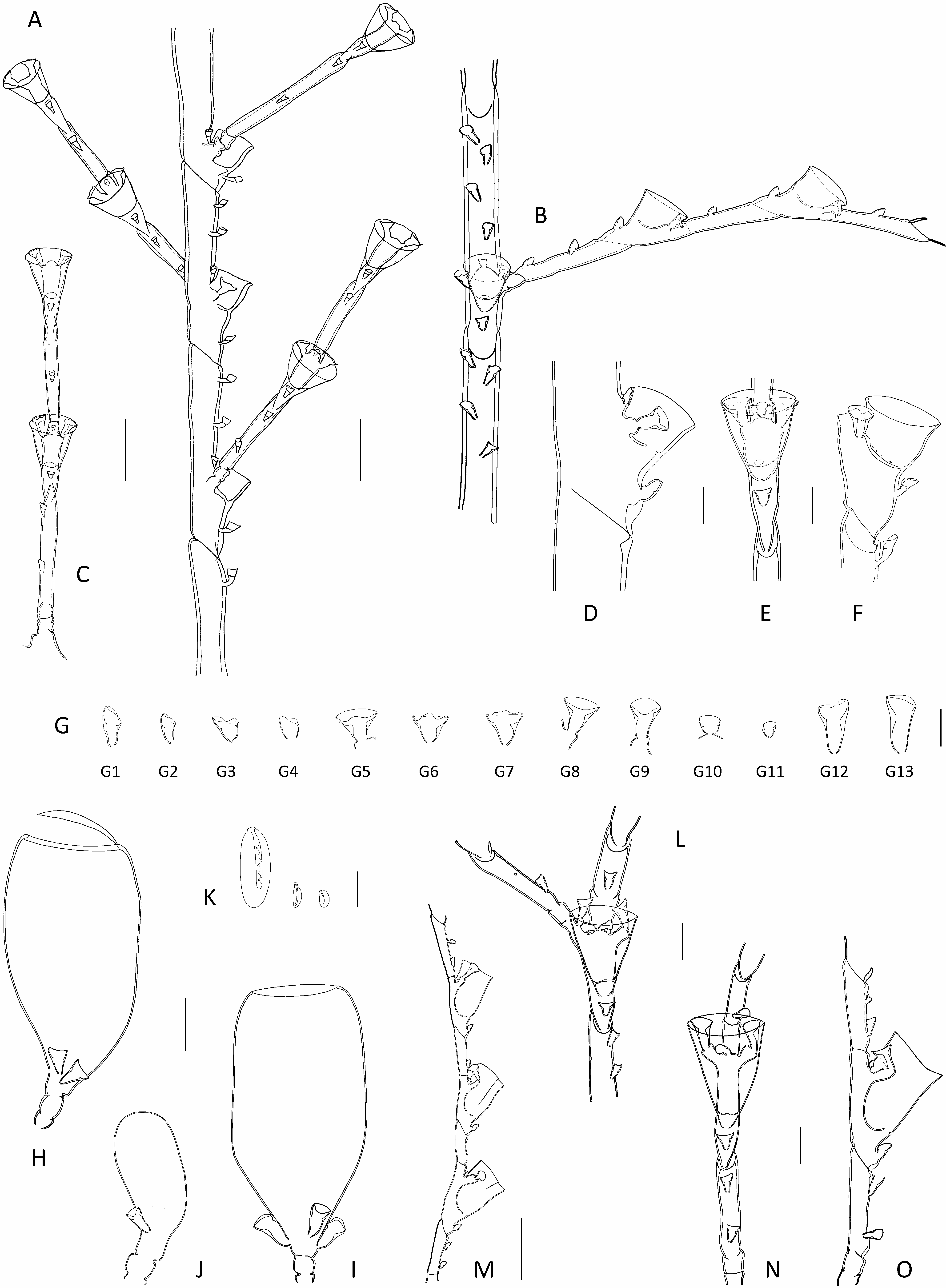
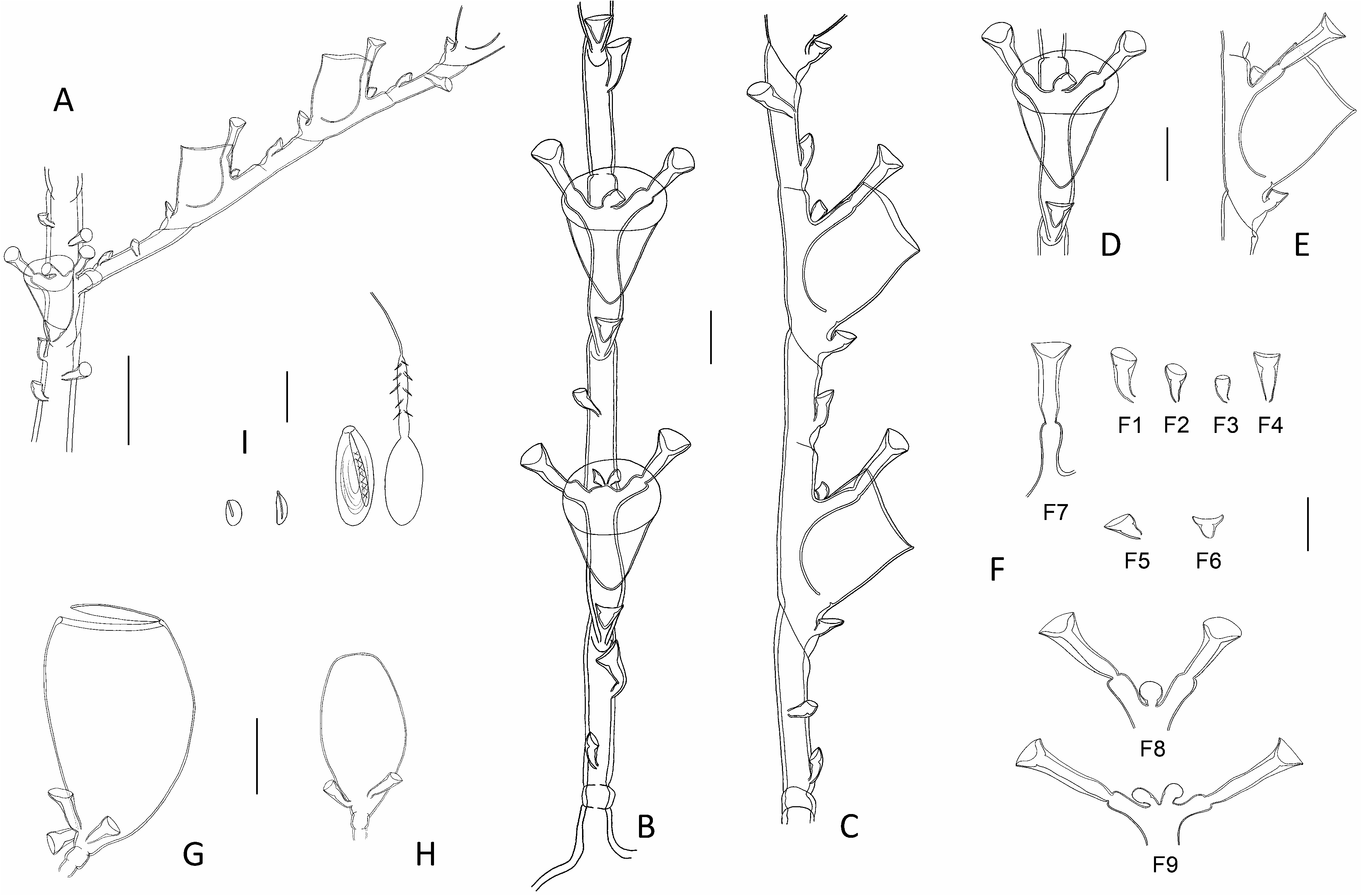
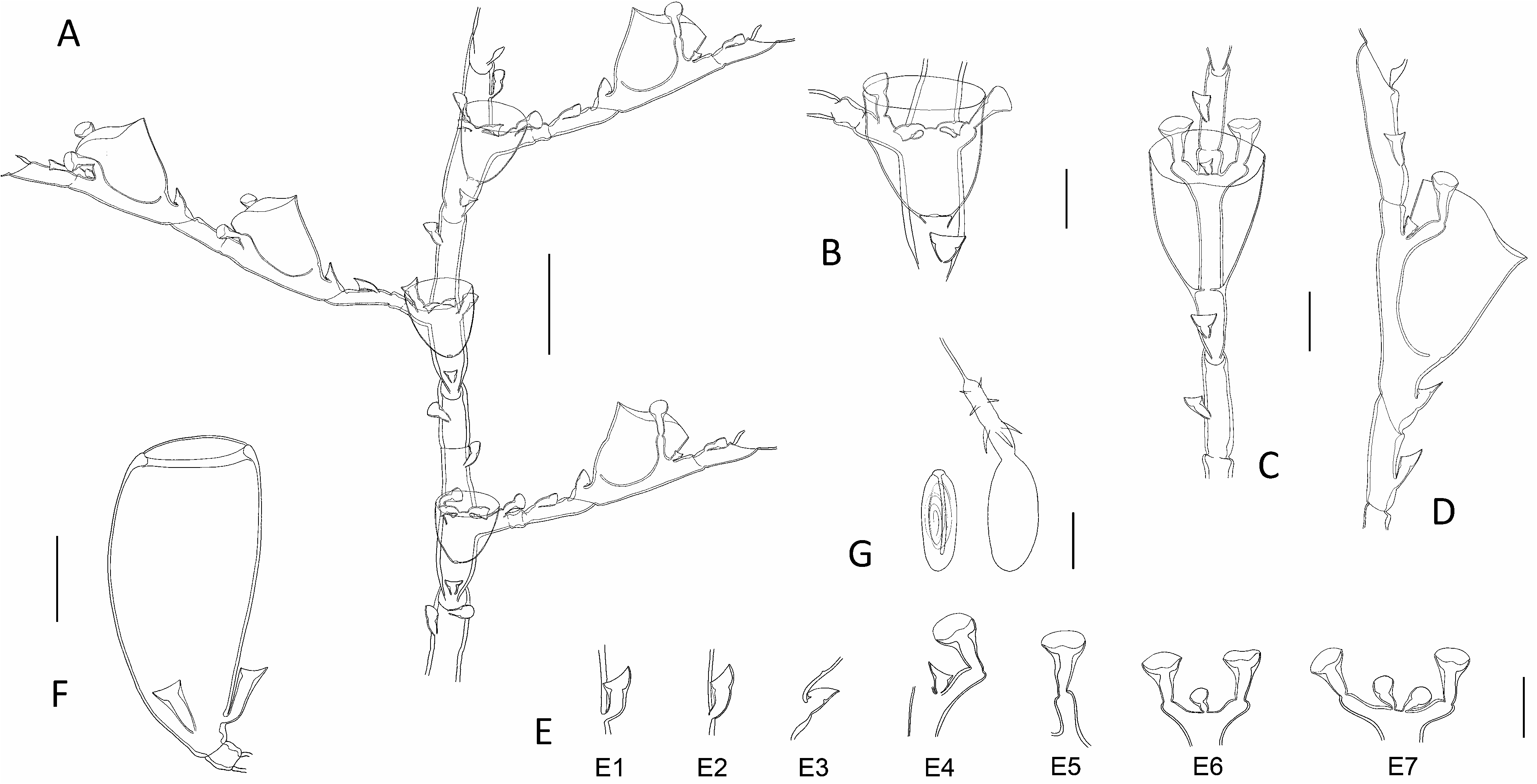
Diagnosis: Small-sized (up to 2.0 cm high) Halopteris with homomerously-divided stems; internodes rather short, comprising a hydrotheca in their lower 2/3rd part, a well-developed lateral apophysis, and up to 7 nematothecae (1 mesial, a pair of laterals, 2 axillar, and 1-2 superior ones in a median row). Cladia alternate, divided heteromerously; hydrothecate internodes comparatively longer than their ahydrothecate counterparts, bearing a centrally-placed hydrotheca and up to 5 nematothecae (1 mesial, a pair of laterals and generally 1, exceptionally 2, axillar nematothecae); ahydrothecate internodes very short, carrying single nematotheca in their lower halves. Hydrotheca deep, tubular, slightly flaring below aperture, rim circular, slightly scooped in lateral view. Lateral nematothecae barely surpassing hydrothecal rim, borne on well-developed apophyses; conical, walls of lower chamber gradually thickening distally, apical chamber shallow, rim from lowered to sinuated adaxially. Male gonothecae elongated-ovoid, borne on short, quadrangular pedicel; 2 basal nematothecae; distally a narrow, circular aperture. Female gonothecae comparatively larger, borne on short, quadrangular pedicel; broadly ovoid, tapering below, with large apical aperture with thickened rim, closed by glass-watchshaped operculum; basally 2-3 nematothecae.
Description: Colonies composed of varied number of cormoids arising from tubular, creeping, branching hydrorhiza, devoid of nematothecae. Stems simple, erect (up to 13 mm high in present material), monosiphonic ( Figs 1D View Fig , 2 View Fig C-E), composed of a basal, ahydrothecate part, and a much longer, distal part bearing hydrothecae and cladia. Basal part of varied length, almost straight, irregularly divided into successive segments of unequal length through up to 6 transverse nodes, bearing a total of up to 18 nematothecae arranged in two parallel, closely-set, longitudinal rows; distalmost segment separated from hydrothecate part of caulus through deeply-incised, oblique node. Upper part of caulus almost collinear proximally, becoming decidedly geniculate distally, divided into up to 15 regular internodes by means of oblique nodes ( Fig. 5A, M View Fig ); distally, the segmentation may occasionally change to heteromerous through the insertion of transverse nodes. Each stem internode composed of a hydrotheca situated in its proximal half, a lateral apophysis (or a pair in the proximal most internode), and up to 7 nematothecae (a mesial, a pair of laterals, a pair of axillar, as well as 1 or, occasionally, 2 superior ones in a median row ( Fig. 5C View Fig ); superior nematothecae confined to a separate, ahydrothecate internode, where caulus segmentation becomes heteromerous; often a pair of laterally-displaced superior nematothecae, inserted at same level, on proximal most stem internode bearing the pair of cladia); internodes slightly bent above hydrotheca in opposite direction to corresponding cladium. Apophyses, exclusive of those of proximal most internode (that are paired), alternate along stem, each supporting a cladium. Cladia composed of a proximal, short, athecate, quadrangular internode, followed by a succession of a- and hydrothecate internodes resulting through a heteromerous segmentation ( Fig. 5B View Fig ). Ahydrothecate internodes with transverse proximal node and oblique distal node, carrying proximally a single nematotheca; internodes very short, though proximal most one slightly longer than subsequent counterparts ( Fig. 5D View Fig ). Hydrothecate internodes, up to 4 per cladium, with oblique proximal node and transverse distal node, rather short, accommodating a centrally-placed hydrotheca and its up to 5 associated nematothecae (1 mesial, a pair of laterals, as well as 1, or exceptionally 2, axillar nematothecae) ( Fig. 5E, N View Fig ). All hydrothecae deep, conical, adnate for about half their adaxial length; free adaxial wall straight, abaxial wall slightly sigmoid (basally imperceptibly concave, distally slightly everted below rim); aperture wide, circular, rim slightly scooped in lateral view ( Fig. 5F, O View Fig ); hydranths with ca. 14 filiform tentacles. Gonothecae borne on both caulus and cladia through short, lateral apophyses arising midway between hydrothecal bases and mesial nematothecae. Female gonothecae large, piriform, borne on short quadrangular pedicels; basally a couple of nematothecae; distally a large, ovoid, transverse aperture, with conspicuously thickened rim, closed by a glass-watch-shaped operculum ( Fig. 5H, I, P View Fig ). Male gonothecae borne on single, quadrangular pedicels; fusiform to elongated-ovoid, with couple of basal nematothecae, and a distal, narrow, circular, transverse aperture, closed by thin perisarc sheet ( Fig. 5J, K, Q View Fig ). All nematothecae bithalamic, including the axillar ones; mesial triangular in frontal view, upper chamber slightly shallower than basal one, with conspicuously lowered rim on adaxial side ( Fig. 5G View Fig 3 View Fig ); laterals borne on well-developed apophyses, conical, moderatelylong ( Fig. 5G View Fig 4 View Fig , 5 View Fig ), not always reaching the hydrothecal rim ( Fig. 5D, F, O View Fig ); basal chamber narrow, tubular, rather tall; upper chamber shallow, wide, margin either lowered or sigmoid on adaxial side; cauline ( Fig. 5G View Fig 1 View Fig ) and cladial ( Fig. 5G View Fig 2 View Fig ) nematothecae, other than those associated to the hydrothecae, as well as those borne by the gonothecae ( Fig. 5G View Fig 8 View Fig , 9 View Fig 9 ), long conical, with tall basal chamber and comparatively shallow upper chamber, with rim slightly lowered adaxially; axillar nematothecae ( Fig. 5G View Fig 6 View Fig , 7 View Fig ) with much thinner perisarc than their exposed counterparts, short, with chambers of nearly the same depth, wall of upper chamber facing adaxial hydrothecal wall distinctly lowered. Cnidome ( Fig. 5L, R View Fig ) composed of 3 types of microbasic mastigophores: large, elongated-ovoid [(16.8-18.2) × (6.5-6.8) μm, in nematophores, as well as scattered in the coenosarc]; small, banana-shaped [(5.6-5.8) × (1.9- 2.1) μm, in tentacles]; small, ovoid capsules [(4.5-4.9) × (2.8-2.9) μm, scattered in the coenosarc]. Color in life from pale yellow to brownish.
Remarks: The segmentation of cauli is generally homomerous, although the insertion of transverse node towards their distal parts introduces a heteromerous segmentation. The cladia are generally divided heteromerously, though an occasional absence of transverse nodes could be noted in material MHNG- INVE-98635, this not supposing a homomerous segmentation throughout the involved cladium. The cauline hydrothecae generally display a pair of axillar nematothecae, though only one, placed medially, may occasionally occur in distalmost hydrothecae. In contrast, generally a single axillar nematotheca is associated to each cladial hydrotheca though, exceptionally, a couple may occur. The stems internodes bear generally 1, occasionally 2, median superior nematothecae (ca. 95% and 5%, respectively, n=150); these are arranged in a single row in ordinary internodes, while they are shifted laterally and occur at the same level in the basalmost internodes giving rise to pairs of cladia. The first cladial internodes bear generally but a single nematotheca; rarely, two nematothecae were observed in some cormoids from MHNG-INVE-98635. The material MHNG-INVE-9833 comprises monoecious stems, while only female gonothecae seem to occur in former.
The trophosomes of the present materials from the Indian Ocean were compared with those of specimens from the Lesser Antilles, and all appear indistinguishable, both morphologically and morphometrically (compare Fig. 5A, B, D and 5M, O View Fig ). In addition, the as yet unknown female gonothecae of the Caribbean population, were found only recently (sample HRG-1339), and do not differ from those of the Indian Ocean counterparts (compare Fig. 5P and 5H & I View Fig ; Table 3 View Table 3 ). Moreover, the molecular study supports the conclusions based on the morphological observations ( Fig. 9 View Fig 9 ).
Several literature records of hydroids conform, through the segmentation of both the stem and cladia, and the number and position of the nematothecae they carry, to the morphology of the present species. However, minor discrepancies could be occasionally noted, although they are thought to reflect rather overlooked details. Thus, Vervoort’s (1967) material from the Red Sea, obviously erroneously assigned to Antennella secundaria ( Gmelin, 1791) , is reportedly said to carry single axillar nematothecae behind the cauline hydrothecae, a situation – indeed – met with only distally in the stems of the present species. Moreover, it should be stated that one nematotheca belonging to a pair, namely that given off on the same side as the corresponding cladium, may be difficult to notice as it is hidden (in frontal view) by the apophysis supporting the lateral nematotheca originating on the same side (see Fig. 5C View Fig – lateral nematotheca on same side as the cladium expressly shown).
The “second pair of lateral nematothecae […] mesial to and much smaller than normal pair”, observed by Ryland & Gibbons (1991) in their Fijian specimens assigned to H. buskii , is nothing else than the couple of cauline axillar nematothecae. Similarly so, besides the commonly seen single cladial axillar nematothecae, “two mediosuperior nematothecae are often present behind the free adcauline wall of the hydrotheca”. These, combined to the heteromerous segmentation of cladia, the shape and size of both hydro-and lateral nematothecae, and the morphology of the female gonotheca, suggest that their specimens are, most probably, conspecific with H. vervoorti .
The Japanese materials assigned to both Heterotheca and Halopteris buski (sic!) by Hirohito (1974, 1995, respectively) are morphologically close to the present species, as their cauli and cladia display the same segmentation, the same number and position of nematothecae, and the monoecious condition of their cormoids. However, a pending genetic study is expected to clarify their relationships undoubtedly.
The presence of H. vervoorti , originally described from the Caribbean ( Galea, 2008), in the Indian Ocean is not entirely surprising. The reverse situation is, indeed, more common at present, with many examples of hydroids, primarily known to occur in the Indo-Pacific that were subsequently recorded from the Caribbean, viz. Sertularia rugosissima Thornely, 1904 ( Galea, 2008) , S. tongensis ( Stechow, 1919) ( Galea, 2010) , S. hattorii Leloup, 1940 ( Galea & Ferry, 2015), Aglaophenia postdentata Billard, 1913 ( Galea, 2013) , and Clytia edentula Gibbons & Ryland, 1989 ( Galea & Ferry, 2015). Whether these species primarily occurred in one geographical area and spread afterwards elsewhere, could not be established at this stage, but it is obvious that only the frequency of the collecting efforts first revealed their presence in one area but not in the other.
Halopteris vervoorti comes close to a number of nominal species through the occurrence of paired axillar nematothecae behind the cauline hydrothecae and the heteromerous segmentation of its cladia, viz. H. australis sp. nov. (see below), H. brasiliensis (see below), H. liechtensternii ( Marktanner-Turneretscher, 1890) , and H. sibogae ( Billard, 1913) . However, unlike H. australis , it does not possess hydrothecae with distinctly sinuated margins or exceedingly-long lateral nematothecae, respectively. On the other hand, H. brasiliensis is a species forming taller cormoids, with thicker stems and cladia, broader hydrothecae, and cauline superior nematothecae distinctly arranged into two longitudinal rows; unlike in H. vervoorti , its cauline hydrothecae occupy nearly the whole length of the internode. Halopteris liechtensternii is a species with comparatively longer stem internodes (and, implicitly, more widely-spaced cladia), provided regularly with 2-3 superior nematothecae arranged in two distinct rows, its female gonothecae are much longer and almost cylindrical, while its males appear dwarfed compared to those of the present species. Halopteris sibogae , besides many common features shared with H. vervoorti (see under the former), is immediately distinguishable through its exceedingly long lateral nematothecae. Additional differences to other congeners are summarized in Appendix 1.
Distribution: Lesser Antilles ( Galea, 2008, 2010, 2013), Cape Verde ( Ansín Agís et al., 2001, as H. polymorpha ), (?) Red Sea ( Vervoort, 1967; as Antennella secundaria ), Madagascar, the Maldives, Indonesia (present study), (?) Australia ( Preker & Lawn, 2010, as H. polymorpha ), (?) Japan [ Hirohito (1974), as Heterotheca buski (sic!); Hirohito (1995), as Halopteris buski (sic!)], (?) Fiji ( Ryland & Gibbons, 1991, as Halopteris buskii ).





| R |
Departamento de Geologia, Universidad de Chile |
| DM |
Dominion Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Halopteris vervoorti Galea, 2008
| Galea, Horia R., Gioia Di Camillo, Cristina, Maggioni, Davide, Montano, Simone & Schuchert, Peter 2018 |
Halopteris vervoorti
| Galea H. R. 2008: 42 |
Halopteris polymorpha
| Preker M. & Lawn I. D. 2010: 120 |
| Ansin Agis J. & Ramil F. & Vervoort W. 2001: 167 |
Halopteris buski
| Hirohito & Emperor of Japan 1995: 244 |
Halopteris buskii
| Ryland J. S. & Gibbons M. J. 1991: 527 |
Heterotheca buski
| Hirohito & Emperor of Japan 1974: 30 |
Antennella secundaria
| Vervoort W. 1967: 42 |
Plumularia buskii
| Bale W. M. 1884: 125 |