A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads


1
Key Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Anthropology, Chinese Academy of Sciences, 142 Xizhimenwai Ave., Beijing 100044, China
2
CAS Center for Excellence in Life and Paleoenvironment, Beijing 100044, China
3
Institute of Paleobiology, Polish Academy of Sciences, Twarda 51/55, 00-818 Warsaw, Poland
*
Author to whom correspondence should be addressed.
Diversity 2020, 12(11), 420; https://doi.org/10.3390/d12110420
Submission received: 25 October 2020
/
Revised: 1 November 2020
/
Accepted: 3 November 2020
/
Published: 5 November 2020
(This article belongs to the Section Phylogeny and Evolution)
Abstract
:The middle Eocene in Nei Mongol (China) was an interval of profound faunal changes as regards the basal Glires and gliriform mammals in general. A major diversification of rodent lineages (ctenodactyloids) and more modern small-sized lagomorphs was accompanied by a decline of mimotonids (Gomphos and Mimolagus) and anagalids. The latter was an enigmatic group of basal Euarchontoglires endemic to China and Mongolia. Here, we describe the first anagalid tooth (a P4) from the Huheboerhe classic site in the Erlian Basin. The tooth, characterized by its unique morphology intermediate between mimotonids and anagalids is semihypsodont, has a single buccal root typical of mimotonids, a large paracone located anteriorly, and a nascent hypocone, characteristic of advanced anagalids. The new finding of neither an abundant nor speciose group suggests a greater diversity of anagalids in the Eocene of China. This discovery is important because it demonstrates the convergent adaptations in anagalids, possibly of ecological significance.
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Glires (rodents and lagomorphs) is the most numerous clade of Euarchontoglires which, in turn, is one of the four main clades of placental mammals, recognized on the basis of molecular and morphological data—e.g., [1,2,3]. The “basal Glires” is an operational term for a paraphyletic group of primitive members of the clade, which do not display clear rodent or lagomorph synapomorphies. However, this group can be partitioned into simplicidentates (Eurymylidae) and duplicidentates (Mimotonidae), based on the number of incisors [4,5,6]. The basal Glires usually cluster in the phylogenetic analyses with Anagalidae [4,7], an enigmatic grouping of basal Euarchontoglires. The concept of “Anagalida” was proposed originally by Szalay and McKenna [8] as including Anagalidae, Pseudictopidae, Zalambdalestidae, and possibly, Didymoconidae; later emended by the addition of Macroscelida and Lagomorpha [9]. Macroscelida in the light of molecular data are considered Afrotheria [10]; thus, the Anagalida sensu McKenna [9] are now disproved. Meng and Wyss [11] defined the term “Gliriformes” as all taxa are phylogenetically closer to Glires than to any other Recent placental group—a notion endorsed, e.g., by Missiaen and Smith [12].
Whether Anagalidae themselves form a monophyletic group or are also a paraphyletic group is still a matter of debate [13]. Nevertheless, with respect to the cranial and especially dental morphology, they can be viewed as a morphological transitional zone in the morphospace between the basal primates and scandentians on one side and basal Glires on the other, but they are closer to the last group [13].
All representatives of Anagalidae and almost all early Glires are known exclusively from the Paleogene of Asia (mainly from China) with the only exception of the rodentiaform Alagomyidae lineage which is also known from North America [14]. Most anagalid taxa are restricted spatio-temporally to the Paleocene of southern and central China [13,15]. The Eocene representatives were confined to northern China and Mongolia [13,16,17,18] and their findings are scarce. Thus, any new discovery of this enigmatic group is of considerable interest to our understanding of evolution of gliriform mammals.
Here we present an upper premolar of an unknown genus and species of the anagalid branch of the Euarchontoglires from the middle Eocene (Irdinmanhan Asian Land Mammal Age) Huheboerhe locality in the Erlian Basin, Nei Mongol, China. The specimen shows strong similarities both to Eocene representatives of basal Glires and some more evolved Anagalidae.
2. Materials and Methods
The isolated gliriform premolar (IVPP V26960) is housed in the collection of the Institute of Vertebrate Paleontology and Paleoanthropology of the Chinese Academy of Sciences, Beijing, China (IVPP). The tooth was examined with a scanning microscope JSM 6100 (at the IVPP). Terminology of dental structures follows Meng and Wyss [11].
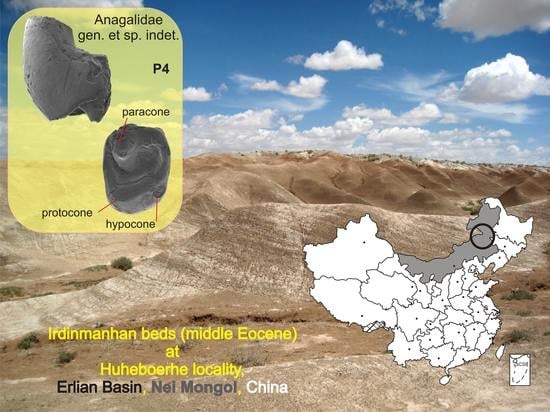
Geological setting: The tooth was found at the Huheboerhe locality (Figure 1), which lies ca. 37 km SW of Erenhot (Nei Mongol, China), close to the China/Mongolia boundary [19]. The site is located at the “Huhebolhe Cliff” (sensu Russell and Zhai [20]) and is also known as “Huhe Bulak” or “locality 7 mi west and southwest of Camp Margetts”, as marked during the Central Asiatic Expedition (CAE) led by the American Museum of Natural History (AMNH) in 1930 [19]. Paleogeographically, Huheboerhe is located in the Erlian Basin [21,22], one of non-marine basins of late Mesozoic origin in northeastern China. The Huheboerhe section includes three sediment series divided by upper and lower hiatuses: the “upper beds”, consisting of a lower part of the Irdin Manha Formation (IM-1) and underlying upper beds of the Arshanto Formation (AS), the “middle beds” including deposits of the Arshanto Formation, and the “lower beds” consisting of the upper Nomogen Formation sediments of the Bumbanian Asian Land Mammal Age [23].
The fossil mammal fauna of Huheboerhe is rich; the lower beds yielded many Gomphos specimens [25], the Arshantan sediments include numerous remains of perissodactyls [19,23], and the Irdinmanhan deposits yielded, among others, artiodactyls, primates, cricetids, ischyromyids, many ctenodactyloids, lagomorphs, and the mimotonid Mimolagus aurorae [19,23,24,26,27,28].
3. Systematic Paleontology
- Mammalia Linnaeus, 1758
- Euarchontoglires Murphy et al., 2001
- Anagalida Szalay and McKenna, 1971
- Anagalidae Simpson, 1931
- Gen. et sp. indet.
- (Figure 2)
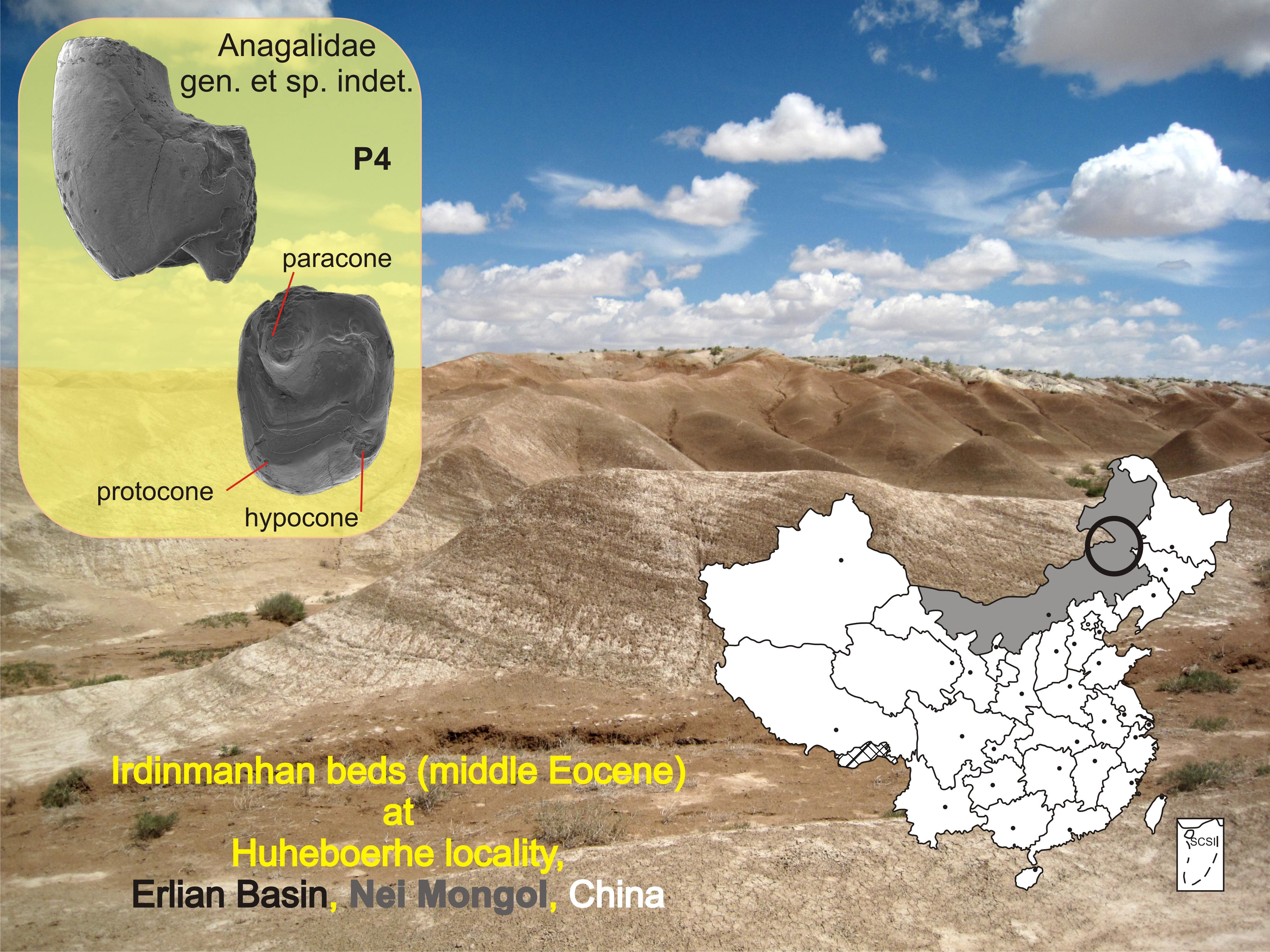
Referred material: An isolated left P4 (IVPP V26960) with a complete tooth crown, and broken buccal and lingual roots.
Locality and age: Huheboerhe, Erlian Basin, Nei Mongol, lower Irdinmanhan beds (IM-1) of the Irdin Manha Formation, middle Eocene (Irdinmanhan ALMA).
Description and Comparisons
IVPP V26960 is a left P4 with two clearly marked roots, with a semicircular cross-section; the buccal root was smaller than the main one, which is typical of the semihypsodont upper premolars. Similar to large Eocene mimotonids, such as Gomphos and Mimolagus [17,24,25], it has only one buccal root. However, overall morphology of the crown (a squarish outline, the paracone located anterobuccally, and an incipient hypocone) more closely resembles that of larger anagalids (such as Anagale, Anagalopsis, and Hsiuannania); thus, comparisons with both groups seem relevant for this study. The tooth size is close to that of large mimotonids (Mimolagus) and middle-sized anagalids such as Anaptogale; it is smaller than the P4 of Anagalopsis, Eosigale, Hsiuannania, and Qipania, but larger than that of Diacronus. The crown is distinctly bilaterally hypsodont, with the lingual side almost twice as high (3.6 mm) as at the buccal one (2.16 mm), which is another feature characteristic of both Mimotonidae and Anagalidae [5,29,30].
The occlusal surface is almost square in outline, with the parallel anterior and posterior margins (the distal one shorter) oriented perpendicularly to the buccal margin. Such arrangement forces the lingual margin of the tooth to be placed askew, with the protocone being the lingual-most prominent element. The total width of the tooth is 4.32 mm and the maximum anteroposterior length is 3.24 mm. Morphologically, the tooth is most similar to that of Anagale or Anagalopsis, which are, however, more square (Figure 2). It differs from the P4 of Eosigale and Diacronus where the outline of the occlusal surface is subtriangular, and from Hsiuannania tabiensis and Qipania yui with a less squarish P4 outline, closer to a trapezoid (Figure 2). The anterior and posterior sides of the tooth are not parallel in Hsiuannania but angled to each other, whereas in Qipania they are parallel but not at the right angle with the buccal margin. In both genera (Hsiuannania and Qipania) the buccal and lingual margins are positioned askew to the parasagittal axis. On the other hand, the P3 and P4 in Gomphos and Mimolagus are much more oval in outline than IVPP V26960 and have somewhat tapered lingual margins [24,25], not flattened ones.
The new tooth shows cuspo-lobate morphology with a large, conical paracone, low and partly worn protocone, and two tiny style-like cuspules at the buccal margin of the tooth, distal to the paracone (Figure 2); whether both are stylar cusps or one of them is a minuscule metacone remains unclear. The paracone is the most prominent feature of the tooth. It is as eminent in the new specimen as in other anagalid genera, although it does not cover as much area of the occlusal surface as in Anaptogale and Eosigale, being more constricted at the base (1.68-mm width and 1.1-mm length). It resembles Anagalopsis and Hsiuannania more closely in the overall proportions of cusp size relative to the whole dental surface. In Gomphos, the paracone, or the “central cusp” sensu Meng et al. [25], is placed centrally at the tooth occlusal surface in both P3 and P4, and is surrounded by the pre- and postprotocrista originating at a poorly distinguished protocone. Such topography of the occlusal structures is different from that of IVPP V26960. The P4 in Mimolagus is poorly known, although the strongly worn tooth of the holotype of M. rodens indicates the centro-buccal position of the paracone (see also [17]). The P4 of Mimolagus aurorae is not known, but the P3 is similar to that of Gomphos in general morphology [24].
IVPP V26960 lacks the paraconule, similarly to Anagalopsis and Gomphos, but unlike in Hsiuannania and Qipania, which have small paraconules. The preprotocrista in the new specimen is long and goes straight anteriorly to the anterior side of the paracone, where it ends, defining the anterior margin of the tooth. In this way, the protocrista is complete in the new specimen, not interrupted by the paraconule as in Hsiuannania. In Anaptogale, however, the preprotocrista is poorly developed and very short, and in Hsiuannania and Qipania the preprotocrista ends at the paraconule. Gomphos does not have the paraconule at P4, and its occlusal structure is relatively simple. The heavily worn upper dentition of Mimolagus rodens (IVPP RV51002.1) does not show any additional cusps, apart from the paracone and remains of the protocone.
The postprotocrista in the new specimen is smooth and meets the posterior margin of the tooth, which can be regarded as the postcingulum. The tooth shows no sign of the anterior cingulum present in Anaptogale, Diacronus, Eosigale, Hsiuannania, and Qipania, and has no buccal cingulum, unlike Diacronus and Eosigale.
There is no clear indication of conules; nevertheless, a wear surface of the protocone (and the posterior part of the protocristae) bears a circular extension, indicating a small cusp distally to the protocone, what may be interpreted as a weakly expressed metaconule.
The joining point of the postprotocrista and the posterior tooth margin also indicates the position of a nascent hypocone. It forms a minute shelf slightly dorsal to the occlusal surface. In this feature the tooth resembles that of Hsiuannania tabiensis (IVPP V4274; Figure 2), and differs markedly from both Gomphos and Mimolagus, which do not have a hypocone on the premolars.
There is no parastyle, whereas such a structure is well-expressed in Anagale and also as a minute cusp in Gomphos (Figure 2). There is no marked hypostria at the shaft between the protocone and the hypocone but the shaft wall shows a shallow groove-like concavity descending along the shaft. Posteriorly and lingually to the paracone there is a large, flat-bottomed basin, which is interpreted as the trigon basin.
4. Discussion
Since the middle Eocene onward, mammalian faunas in central Asia experienced major faunal turnovers coupled with climate cooling and aridification [31,32,33,34]. Specifically, on the Mongolian Plateau, the basal Glires lineages identified with mimotonids and eurymylids gave way to rodents and lagomorphs of a modern aspect; both groups underwent diversification between the middle-to-late Eocene and the early Oligocene [21,24,26,27,28]. The demise of archaic gliriform taxa such as Anagalidae, and ischyromyid rodents [35] coincided with the advent of numerous small perissodactyls [19], primates [36], and tree-shrews [37].
After the middle Eocene the number of known anagalids dropped to three monotypic genera. Anagalopsis kansuensis from Gansu (northern China) and Zofiagale ergilinensis (Mongolia) are known solely from their type material [13,17], while Anagale gobiensis from Nei Mongol (China) is represented by two specimens, each of them consisting of the cranium and associated postcranial remains [16,18]. The only other Eocene finding ascribed to Anagalidae is one unilaterally hypsodont tooth of an unknown species from the Irdinmanhan beds at Wulanhuxiu, Nei Mongol, China [30]. The Eocene record of the basal Glires is slightly more diverse, with four species of Gomphos [25,30], and two of Mimolagus [17,24,38]; interestingly, they show the same pattern of size increase [24]. Thus, during the middle-to-late Eocene the representatives of large mimotonids and anagalids co-occurred in the Erlian Basin.
The dental morphology of IVPP V26960 is intermediate between that of large Eocene mimotonids and advanced anagalids. The outline and structure of the occlusal surface more closely resemble those of anagalids than mimotonids—in particular, Anagale and Anagalopsis, in the shape and cusp arrangement, and Hsiuannania in the presence of the small hypocone, situated on the shaft. On the other hand, the presence of a single buccal root is characteristic of the Eocene mimotonids (but not the Paleocene ones), whereas in most of the anagalid genera the P4 has double buccal roots. The premolar IVPP V26960 is slightly smaller than the corresponding teeth of Anagale, Anagalopsis, and Hsiuannania, but nearly equal in size to that of Mimolagus [24].
The evolution of the dentition in Euarchontoglires is highly mosaic, with frequent examples of convergence or evolutionary parallelisms in more distant or more closely related lineages, respectively [5]. However, the dentition of Anagalidae still lacks, typical of Glires, deletions in their premolar segment, and incisors (up to three pairs) are usually only slightly enlarged and semiprocumbent; their molars display distinct unilateral hypsodonty also typical of early Glires and the whole duplicidentate lineage. In some cases (e.g., Anagalopsis, Hsiuannania or Qipania) the crowns of the molars are higher than those in the co-occurring Glires (see e.g., [30]). Additionally, the molar occlusal surfaces, in most anagalids, show very heavy wear, which obliterates all cuspal pattern but the most buccal structures (see, e.g., [16,17]). Such extensive dental wear suggests an abrasive diet indicative of a high fiber content, demanding long chewing, as seen in duplicidentates—i.e., lagomorphs and larger mimotonids, such as Gomphos or Mimolagus [24].
That said, the taxonomic affiliation of IVPP V26960 is quite enigmatic. One might argue that, morphologically, the presence of a single buccal root advocates a Glires affiliation. However, the patterning mechanism of the root number in placental mammals is regulated developmentally and is highly plastic [39]; thus, a thorough modification of the tooth occlusal structure to resemble that of anagalids would probably require a greater number of the evolutionary transformations in mimotonids than merely the fusion of the buccal roots in anagalids. We are inclined to regard the new finding from the Irdinmanhan beds of Huheboerhe as a representative of a new genus and species of the derived anagalid lineage, probably related closely to Anagalopsis or Anagale. Its progressive characters are, apart from the root structure, the simplification of the occlusal crown pattern, the loss of the anterior and buccal cingula, and a nascent hypocone, indicating a trend towards molarization of the premolars, which parallels Glires [40]. Eventually, in the early Oligocene, the anagalids and basal Glires lost the ecological competition to radiating rodents and lagomorphs of a modern aspect, and small perissodactyls [24,31].
This study suggests that the increased sampling and better taxonomic resolution, especially from the Irdinmanhan and younger sediments, is needed to further address the question of morphological convergence and ecological specializations in late anagalids.
Author Contributions
Ł.F.F. designed the study; methodology: Ł.F.F. and Q.L.; analysis of data: Ł.F.F., Q.L., and A.S.; original draft preparation: Ł.F.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NATIONAL SCIENCE CENTRE (Cracow, Poland) grant number 2015/18/E/NZ8/00637, Chinese Academy of Sciences Strategic Priority Research Program XDB26000000, and National Natural Science Foundation of China grant number 41572013. The fieldwork was also supported by the Special Fund for Fossil Excavation and Preparation, Chinese Academy of Sciences. The APC was funded by grant number 2015/18/E/NZ8/00637 (NSC).
Acknowledgments
We are grateful to Jin Meng, Yuanqing Wang, Xijun Ni, Bin Bai, Xun Jin, Ping Li, Chuankui Li, K. C. Beard, D. L. Gebo, Wei Chen, Wei Zhou, Shejie Li, Qiang Cao, Wei Gao, and Chenkai Sun for their assistance during fieldwork. We thank Wending Zhang (IVPP) for the SEM photos and Aleksandra Hołda-Michalska (Institute of Paleobiology PAS, Warsaw, Poland) for help with the drawing in Figure 1. The comments by two anonymous reviewers greatly improved the final version of the paper.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Murphy, W.J.; Eizirik, E.; Johnson, W.E.; Zhang, Y.P.; Ryder, O.A.; O’Brien, S.J. Molecular phylogenetics and the origins of placental mammals. Nature 2001, 409, 614–618. [Google Scholar] [CrossRef]
- Asher, R.J.; Bennett, N.; Lehmann, T. The new framework for understanding placental mammal evolution. Bioessays 2009, 31, 853–864. [Google Scholar] [CrossRef]
- Hautier, L.; Bennett, N.C.; Viljoen, H.; Howard, L.; Milinkovitch, M.C.; Tzika, A.C.; Goswami, A.; Asher, R.J. Patterns of ossification in southern versus northern placental mammals. Evolution 2013, 67, 1994–2010. [Google Scholar] [CrossRef]
- Asher, R.J.; Meng, J.; Wible, J.R.; McKenna, M.C.; Rougier, G.W.; Novacek, M.J. Stem Lagomorpha and the antiquity of Glires. Science 2005, 307, 1091–1094. [Google Scholar] [CrossRef]
- Fostowicz-Frelik, Ł. Convergent and parallel evolution in early Glires (Mammalia). In Evolutionary Biology: Self/Nonself Evolution, Species and Complex Traits Evolution, Methods and Concepts; Pontarotti, P., Ed.; Springer: Cham, Switzerland, 2017; pp. 199–216. [Google Scholar] [CrossRef]
- Fostowicz-Frelik, Ł. Most successful mammals in the making: A review of the Paleocene Glires. In Evolutionary Biology: A Transdisciplinary Approach; Pontarotti, P., Ed.; Springe: Cham, Switzerland, 2020; in press. [Google Scholar] [CrossRef]
- Meng, J.; Hu, Y.M.; Li, C.K. The osteology of Rhombomylus (Mammalia, Glires): Implications for phylogeny and evolution of Glires. Bull. Am. Mus. Nat. Hist. 2003, 275, 1–247. [Google Scholar] [CrossRef]
- Szalay, F.S.; McKenna, M.C. Beginning of the age of mammals in Asia: The late Paleocene Gashato fauna, Mongolia. Bull. Am. Mus. Nat. Hist. 1971, 144, 273–317. [Google Scholar]
- McKenna, M.C. Toward a phylogenetic classification of Mammalia. In Phylogeny of the Primates. A Multidisciplinary Approach; Luckett, W.P., Szalay, F.S., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 21–46. [Google Scholar]
- Murphy, W.J.; Pringle, T.H.; Crider, T.A.; Springer, M.S.; Miller, W. Using genomic data to unravel the root of the placental mammal phylogeny. Genome Res. 2007, 17, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Wyss, A.R. The morphology of Tribosphenomys (Rodentiaformes, Mammalia): Phylogenetic implications for basal Glires. J. Mammal. Evol. 2001, 8, 1–71. [Google Scholar] [CrossRef]
- Missiaen, P.; Smith, T. The Gashatan (late Paleocene) mammal fauna from Subeng, Inner Mongolia, China. Acta Palaeontol. Pol. 2008, 53, 357–378. [Google Scholar] [CrossRef]
- López-Torres, S.; Fostowicz-Frelik, Ł. A new Eocene anagalid (Mammalia: Euarchontoglires) from Mongolia and its implications for the group’s phylogeny and dispersal. Sci. Rep. 2018, 8, e13955. [Google Scholar] [CrossRef] [PubMed]
- Flynn, L.J. Hystricognathi and Rodentia incertae sedis. In Evolution of Tertiary Mammals in North America. Volume 2: Small Mammals, Xenarthrans, and Marine Mammals; Janis, C.M., Gunnell, G.F., Uhen, M.D., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 498–506. [Google Scholar]
- Li, C.K. Order Anagalida. In Basal Synapsids and Mammals, Volume III, Eulipotyphlans, Proteutheres, Chiropterans, Euarchontans, and Anagalids; Li, C.K., Qiu, Z., Eds.; Science Press: Beijing, China, 2016; pp. 390–411. [Google Scholar]
- Simpson, G.G. A new insectivore from the Oligocene, Ulan Gochu Horizon, of Mongolia. Am. Mus. Novit. 1931, 505, 1–22. [Google Scholar]
- Bohlin, B. Some mammalian remains from Shih-ehr-ma-ch’eng, Hui-hui-p’u area, Western Kansu. Reports from the Scientific Expedition to the North-Western Provinces of China under Leadership of Dr Sven Hedin. The Sino-Swedish Expedition Publication 35, VI. Vert. Paleontol. 1951, 5, 1–48. [Google Scholar]
- McKenna, M.C. New evidence against tupaioid affinities of the mammalian family Anagalidae. Am. Mus. Novit. 1963, 2158, 1–16. [Google Scholar]
- Bai, B.; Wang, Y.Q.; Li, Q.; Wang, H.B.; Mao, F.Y.; Gong, Y.X.; Meng, J. Biostratigraphy and diversity of Paleogene perissodactyls from the Erlian Basin of Inner Mongolia, China. Am. Mus. Novit. 2018, 3914, 1–60. [Google Scholar] [CrossRef]
- Russell, D.E.; Zhai, R.J. The Paleogene of Asia: Mammals and stratigraphy. Mém. Mus. Natl. Hist. Nat. Série C Sci. Terre. 1987, 52, 1–488. [Google Scholar]
- Wang, Y.Q.; Meng, J.; Beard, K.C.; Li, Q.; Ni, X.; Gebo, D.L.; Bai, B.; Jin, X.; Li, P. Early Paleogene stratigraphic sequences, mammalian evolution and its response to environmental changes in Erlian Basin, Inner Mongolia, China. Sci. China Earth Sci. 2010, 53, 1918–1926. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Meng, J.; Jin, X. Comments on Paleogene localities and stratigraphy in the Erlian Basin, Nei Mongol, China. Vert. Palasiat. 2012, 50, 181–203. [Google Scholar]
- Meng, J.; Wang, Y.; Ni, X.; Beard, K.C.; Sun, C.; Li, Q.; Jin, X.; Bai, B. New stratigraphic data from the Erlian Basin: Implications for the division, correlation, and definition of Paleogene lithological units in Nei Mongol (Inner Mongolia). Am. Mus. Novit. 2007, 3570, 1–31. [Google Scholar] [CrossRef]
- Fostowicz-Frelik, Ł.; Li, C.K.; Mao, F.Y.; Meng, J.; Wang, Y.Q. A large mimotonid from the Middle Eocene of China sheds light on the evolution of lagomorphs and their kin. Sci. Rep. 2015, 5, 9394. [Google Scholar] [CrossRef]
- Meng, J.; Bowen, G.J.; Ye, J.; Koch, P.L.; Ting, S.Y.; Li, Q.; Jin, X. Gomphos elkema (Glires, Mammalia) from the Erlian Basin: Evidence for the Early Tertiary Bumbanian Land Mammal Age in Nei-Mongol, China. Am. Mus. Novit. 2004, 3425, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Fostowicz-Frelik, Ł.; Li, Q. A new genus of stem lagomorphs (Mammalia: Glires) from the Middle Eocene of the Erlian Basin, Nei Mongol, China. Acta Zool. Cracov. 2014, 57, 29–42. [Google Scholar] [CrossRef]
- Li, Q.; Meng, J. New ctenodactyloid rodents from the Erlian Basin, Nei Mongol, China and the phylogenic relationships of Eocene Asian ctenodactyloids. Am. Mus. Novit. 2015, 3828, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Li, Q. Eocene fossil rodent assemblages from the Erlian Basin (Inner Mongolia, China): Biochronological implications. Palaeoworld 2016, 25, 95–103. [Google Scholar] [CrossRef]
- Dashzeveg, D.; Russell, D.E. Palaeocene and Eocene Mixodontia (Mammalia, Glires) of Mongolia and China. Palaeontology 1988, 31, 129–164. [Google Scholar]
- Li, Q.; Wang, Y.Q.; Fostowicz-Frelik, Ł. Small mammal fauna from Wulanhuxiu (Nei Mongol, China) implies faunal turnover across the Irdinmanhan–Sharamurunian boundary. Acta Palaeontol. Pol. 2016, 61, 759–776. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; McKenna, M.C. Faunal turnovers of Palaeogene mammals from the Mongolian Plateau. Nature 1998, 394, 364–367. [Google Scholar] [CrossRef]
- Sun, J.; Ni, X.; Bi, S.; Wu, W.; Ye, J.; Meng, J.; Windley, B.F. Synchronous turnover of flora, fauna, and climate at the Eocene-Oligocene boundary in Asia. Sci. Rep. 2014, 4, 7463. [Google Scholar] [CrossRef] [Green Version]
- Li, J.X.; Yue, L.P.; Roberts, A.P.; Hirt, A.M.; Pan, F.; Guo, L.; Xu, Y.; Xi, R.G.; Guo, L.; Qiang, X.K. Global cooling and enhanced Eocene Asian mid-latitude interior aridity. Nat. Commun. 2018, 9, 3026. [Google Scholar] [CrossRef]
- Wasiljeff, J.; Kaakinen, A.; Salminen, J.N.; Zhang, Z.Q. Magnetostratigraphic constraints on the fossiliferous Ulantatal sequence in Inner Mongolia, China: Implications for Asian aridification and faunal turnover before the Eocene-Oligocene boundary. Earth Planet. Sci. Lett. 2020, 535, 116125. [Google Scholar] [CrossRef]
- Li, Q.; Meng, J. Eocene ischyromyids (Rodentia, Mammalia) from the Erlian Basin, Nei Mongol, China. Vert. Palasiat. 2013, 51, 289–304. [Google Scholar]
- Ni, X.; Li, Q.; Li, L.; Beard, K.C. Oligocene primates from China reveal divergence between African and Asian primate evolution. Science 2016, 352, 673–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Ni, X. An early Oligocene fossil demonstrates treeshrews are slowly evolving “living fossils”. Sci. Rep. 2016, 6, 18627. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Q.; Wang, J. On the geological age of mammalian fossils from Shanmacheng, Gansu Province. Vert. Palasiat. 2016, 54, 351–357. [Google Scholar]
- Seo, H.; Kim, J.; Hwang, J.J.; Jeong, H.G.; Han, S.S.; Park, W.; Ryu, K.; Seomun, H.; Kim, J.Y.; Cho, E.S.; et al. Regulation of root patterns in mammalian teeth. Sci. Rep. 2017, 7, 12714. [Google Scholar] [CrossRef] [Green Version]
- Fostowicz-Frelik, Ł.; Li, C.K.; Meng, J.; Wang, Y.Q. New Gobiolagus (Mammalia: Lagomorpha) material from the Middle Eocene of Erden Obo (Nei Mongol, China). Vert. Palasiat. 2012, 50, 219–236. [Google Scholar]
Figure 1.
Anagalid-bearing localities in Nei Mongol (gray area), China. (A). Schematic map of China with the approximate location of the Erlian Basin marked by red circle. (B). Localities in the vicinity of Erenhot, which yielded anagalid remains (yellow dots). ((B), after Fostowicz-Frelik et al. [24], modified).
Figure 1.
Anagalid-bearing localities in Nei Mongol (gray area), China. (A). Schematic map of China with the approximate location of the Erlian Basin marked by red circle. (B). Localities in the vicinity of Erenhot, which yielded anagalid remains (yellow dots). ((B), after Fostowicz-Frelik et al. [24], modified).

Figure 2.
Morphology of ultimate upper premolars in anagalids and large mimotonids. (A,B), left P4 (IVPP V26960), new specimen from the middle Eocene, Huheboerhe, Erlian Basin, Nei Mongol, China. (C), Gomphos elkema, left maxilla fragment with P3–M1 (IVPP NNG047) from the early Eocene, Nuhetingboerhe, Nei Mongol, China. (D), Mimolagus rodens, right P3–M1 (holotype specimen IVPP RV51002.1, mirror view) from the earliest Oligocene of Gansu, China. (E), Anagale gobiensis, right P4–M1 (cast of the holotype AMNH 26079, mirror view) from the early Oligocene of the Ulan Gochu Formation, Twin Obo, Nei Mongol, China. (F), Anagalopsis kansuensis, right P4–M1 (the holotype, IVPP RV51003, mirror view) from the earliest Oligocene of Gansu, China. (G), Hsiuannania tabiensis, right P4–M1 (IVPP V4274, in mirror view; a small hypocone at P4 is marked with yellow arrow) from the middle Paleocene of Anhui, China. (Photographs by ŁFF).
Figure 2.
Morphology of ultimate upper premolars in anagalids and large mimotonids. (A,B), left P4 (IVPP V26960), new specimen from the middle Eocene, Huheboerhe, Erlian Basin, Nei Mongol, China. (C), Gomphos elkema, left maxilla fragment with P3–M1 (IVPP NNG047) from the early Eocene, Nuhetingboerhe, Nei Mongol, China. (D), Mimolagus rodens, right P3–M1 (holotype specimen IVPP RV51002.1, mirror view) from the earliest Oligocene of Gansu, China. (E), Anagale gobiensis, right P4–M1 (cast of the holotype AMNH 26079, mirror view) from the early Oligocene of the Ulan Gochu Formation, Twin Obo, Nei Mongol, China. (F), Anagalopsis kansuensis, right P4–M1 (the holotype, IVPP RV51003, mirror view) from the earliest Oligocene of Gansu, China. (G), Hsiuannania tabiensis, right P4–M1 (IVPP V4274, in mirror view; a small hypocone at P4 is marked with yellow arrow) from the middle Paleocene of Anhui, China. (Photographs by ŁFF).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fostowicz-Frelik, Ł.; Li, Q.; Saha, A. A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads. Diversity 2020, 12, 420. https://doi.org/10.3390/d12110420
AMA Style
Fostowicz-Frelik Ł, Li Q, Saha A. A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads. Diversity. 2020; 12(11):420. https://doi.org/10.3390/d12110420
Chicago/Turabian StyleFostowicz-Frelik, Łucja, Qian Li, and Anwesha Saha. 2020. "A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads" Diversity 12, no. 11: 420. https://doi.org/10.3390/d12110420
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.