3.2. Systematics
Palaemonidae Rafinesque, 1815
Bruceclimenes gen. nov. (authored by Anker)
(urn:lsid:zoobank.org:act:898DB255-7A72-411B-A886-34B4AA5F4303)
Diagnosis: Medium-sized palaemonid shrimps (pocl 4.9–9.5 mm). Body subcylindrical, slightly depressed, smooth; lateral surface of carapace and pleon alveolate, sometimes less distinctly. Rostrum well developed, high, somewhat descending, exceeding antennular peduncle; dorsal margin convex, with 8–10 teeth, posterior-most tooth in postorbital position; ventral margin convex, with 3–6 teeth in distal half; rostral carina strong, continuing posteriorly as elevated mid-dorsal carina, lowering before reaching posterior margin of carapace. Inferior orbital angle strongly anteriorly produced, reaching beyond antennal tooth, blunt or subacute. Antennal and hepatic teeth well developed, acute; latter stouter and situated slightly below former, not reaching anterolateral margin of carapace. Epigastric and supraorbital teeth absent. Thoracic sternite 4 without median process. Pleonite 3 not carinate; posterior margin not posteriorly produced; pleura of pleonites 1–5 posteroventrally rounded; sixth pleonite not elongate. Telson with two pairs of minute, submarginal, dorsal spiniform setae in posterior half and three pairs of short spiniform setae on posterior margin, mesial ones with secondary setules; posterior margin with minute, central tooth between mesial spiniform setae. Antennule typical for family; stylocerite short, acute; distolateral tooth of first article of peduncle sharp; lateral flagellum biramous, with long fused basal part; secondary ramus well developed. Antenna typical for family; scaphocerite well developed, with broad blade and stout distolateral tooth falling short of anterior margin of blade; carpocerite short. Eyes well developed, with large globular cornea. Mandible without palp; molar and incisor processes normal. Maxillule with deeply bilobed palp (endopod), ventral lobe with minute, hooked seta. Maxilla with reduced coxal endite and bilobed basial endite; palp (endopod) small; scaphognathite broad. Maxilliped 1 with endites fused; palp (endopod) small, entire; exopod long, with well-developed caridean lobe; epipod moderately large, subtriangular. Maxilliped 2 with normal endopod and exopod; epipod rounded, without podobranch. Maxilliped 3 with coxa bearing broad, semicircular, lateral plate; endopod robust; antepenultimate article clearly separated from basis, with thick setae on ventral margin; ultimate article with stiff setae; exopod well developed; single arthrobranch moderately developed. Pereiopod 1 slender; carpo-propodal brush present; chela not elongated, not inflated, with fingers as long as palm; cutting edges of fingers unarmed. Pereiopods 2 moderately stout, similar in shape, subequal in size; merus and carpus unarmed, minutely tuberculate; carpus cup-shaped; chela not particularly swollen; palm subcylindrical, moderately elongate, densely covered with minute tubercles; fingers much shorter than palm; cutting edges of fingers each armed with two large teeth proximally, without tooth–fossa snapping mechanism. Pereiopods 3–5 moderately slender; propodus with small spiniform setae on ventral margin; dactylus subtly biunguiculate, with minute secondary unguis on stout corpus and slender, curved, terminal unguis. Male pleopod 1 with endopod simple, expanded distally; lateral margin with plumose setae; mesial margin with robust plumose setae and short spinules; distal margin with two feeble, simple setae. Male pleopod 2 with appendix masculina not overreaching appendix interna, with three longitudinal rows of stiff, simple setae. Uropod normal; protopod with two bluntly ending lobes; distolateral spiniform seta very small; endopod narrower and shorter than exopod.
Type species: Bruceclimenes foveolatus (Bruce, 1981) comb. nov., originally described as Periclimenes foveolatus Bruce, 1981, by present designation and monotypy.
Other species included: None.
Etymology: The name of the new genus combines the last name of Dr. Alexander J. Bruce (1929–2022), an eminent carcinologist, who described hundreds of Indo-Pacific palaemonid shrimps (including P. foveolatus, its type species, see above), and the second part of the word Periclimenes, one of the oldest and largest palaemonid genera. Gender masculine.
Hosts: Deep-water sea anemones (Actiniaria).
Bathymetric range: 187–471 m.
Distribution: Indo-West Pacific from the Red Sea to the Philippines and New Caledonia.
Remarks: The stepwise revision of the problematic genus
Periclimenes (e.g., [
8,
22,
38]) resulted in the description of new genera and revalidation of several previously synonymised genera over the last two decades (e.g., [
10,
20,
36,
39,
40,
41,
42,
43,
44]). The molecular analysis of selected, mainly symbiotic palaemonid taxa by Horká et al. [
8] showed multiple host switches and adaptive convergences, which impeded the genus-level taxonomy within this group.
Periclimenes was, not surprisingly, recovered as a complex, polyphyletic assemblage. Similarly, Frolová et al. [
45] did not recover monophyletic
Cuapetes Clark, 1919 and
Palaemonella Dana, 1852, with several symbiotic taxa belonging to other genera, such as
Vir Holthuis, 1952 and
Eupontonia Bruce, 1971, embedded within these genera, whereas some free-living taxa were found in different parts of the tree. These two studies, as well as molecular analyses by Gan et al. [
21] and Kou et al. [
22,
28,
29], using different datasets, show that numerous taxonomic changes, including redefinitions of larger genera and descriptions of new genera, are expected in the Palaemonidae.
The morphological distinctiveness of the species originally described as
Periclimenes foveolatus was recognised by Bruce [
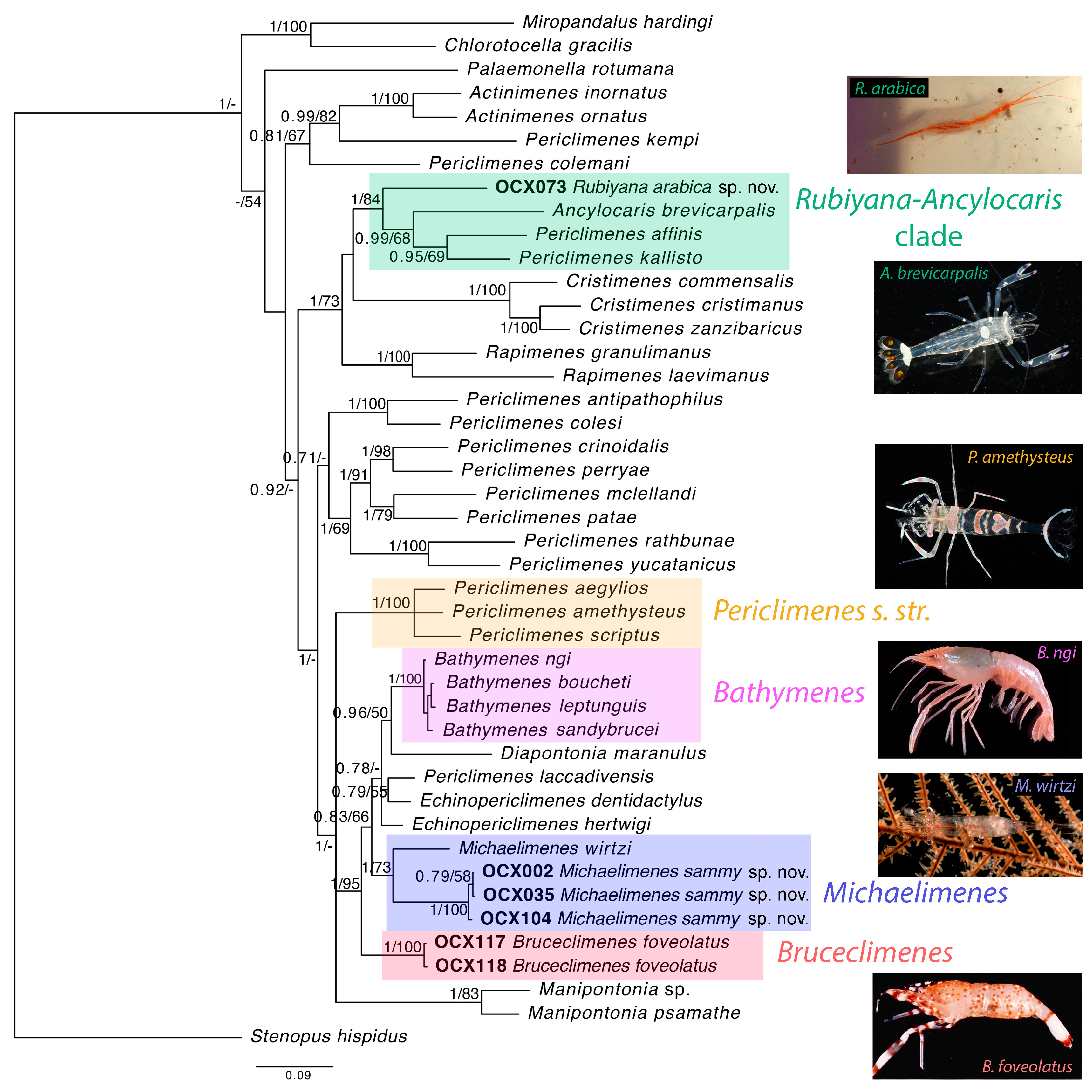
18] and is confirmed by our molecular analysis (
Figure 2), justifying the establishment of a new genus for this strikingly coloured and ecologically unique species associated with deep-water sea anemones (
Figure 3D).
Bruceclimenes can be separated from
Periclimenes s.
str. and related genera by the combination of the following characters: (1) carapace with strong, elevated, mid-dorsal carina, extending almost to its posterior margin; (2) hepatic tooth not greatly enlarged and not reaching or overreaching anterolateral margin of carapace; (3) lateral surface of carapace and pleon with a more or less pronounced “foveolation” (term used by Bruce [
37] for a surface characterised by the presence of numerous small depressions or pits); (4) third pleonite not forming a hump, i.e., not carinate or posteriorly produced; (5) dorsal surface of telson with two pairs of minute spiniform setae; (6) mandible without palp; (7) cutting edges of first pereiopod fingers blade-like, unarmed; (8) surface of second pereiopods finely tuberculate; and (9) dactylus of third to fifth pereiopods subtly biunguiculate without further specialised structures. This combination of characters separates
Bruceclimenes from all other genera allied to
Periclimenes, including
Bathymenes Kou, Li & Bruce, 2016 (see remarks below),
Echinopericlimenes Marin & Chan, 2014,
Michaelimenes Okuno, 2017,
Diapontonia Bruce, 1986 and
Zoukaris Anker & Corbari 2020, which are genera containing mainly deep-water species [
20,
36,
44,
46] as well as
Ancylocaris Schenkel, 1902, a well-known associate of shallow-water sea anemones [
10].
Bruce [
18], in his description of
Periclimenes foresti Bruce, 1981, another deep-water shrimp from the Philippines, stated that this species may be closely related to
P. foveolatus. Subsequently, four other deep-water species of
Periclimenes have been described and assigned to the
P. foresti complex/group [
47,
48,
49]. Two of these species were later transferred to
Bathymenes [
44,
50]. However, the remaining two species of the
P. foresti complex, as well as another deep-water species originally described as
Periclimenes crosnieri Li & Bruce, 2006, also conform to the new diagnosis of
Bathymenes in Ďuriš & Šobáňová [
50] and, therefore, must be transferred to that genus (see below). Based on overall morphology, colour patterns, available host data and preliminary molecular data (
Figure 2),
Bruceclimenes is rather distantly related to
Bathymenes and is also not particularly closely related to
Echinopericlimenes, another major lineage of deep-water shrimps associated with echinoderms. Several other Indo-West Pacific deep-water species of
Periclimenes s.
l., some of them with possible affinities to
Bruceclimenes, await generic reassignments (e.g., [
22,
37,
51]).
Bruceclimenes foveolatus (Bruce, 1981), comb. nov.
Periclimenes foveolatus Bruce [
18]: 196, Figures 6–9, 17a–b, 18b,e; Bruce [
37]: 312, Figures 8, 72; Chace & Bruce [
52]: 56.
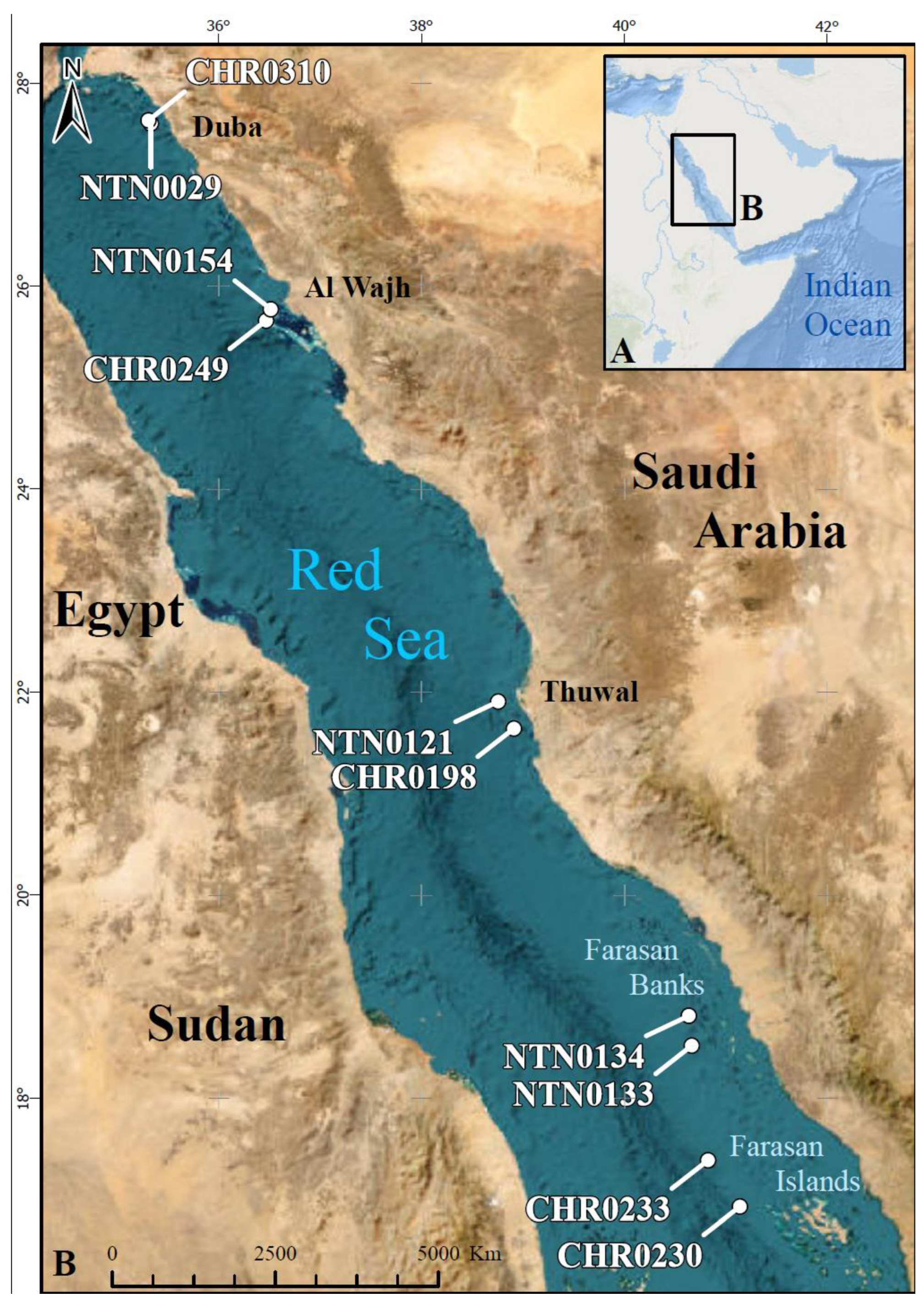
Material examined: Saudi Arabia, Red Sea. One ovig. female (pocl 7.8 mm, cl 11.2 mm), FLMNH UF 68695, west of Umm Urumah, Red Sea Decade Expedition, Leg 3 Phase 2, sta. NTN0154, 25.770922, 36.516308, depth: 471 m, possibly on sea anemone, coll. Neptune Dive Team/S. Vimercati, 14.05.2022 (NTN0154-Bio7/OCX-117*); one male (pocl 4.9 mm, cl 8.8 mm), KAUST (not deposited, see text), west of Mashabih, Red Sea Decade Expedition, Leg 3 Discovery, sta. CHR0249, 25.654252, 36.476551, depth 321 m, on sea anemone, coll. Chimera Dive Team/F. Marchese, 19.04.2022 (CHR0249-Bio8, Slurp2/OCX-118*). *Designates sequenced specimen.
Description: See Bruce [
18], with additional remarks and illustrations in Bruce [
37].
Colour pattern: Body partly translucent, with yellowish tinge, covered with numerous, relatively large, rounded or ovoid, rarely more irregular, bright red spots, and less numerous, less conspicuous, smaller white spots or incomplete transverse bands; red spots smaller on carapace mid-dorsal line and flanks and larger closer to posterior margin of carapace and on pleon; white spots smaller on carapace and forming short dorsal bands on first and third pleonite; rostrum and post-rostral mid-line of carapace with conspicuous, alternating white and red markings; fifth pleonite with red transverse band dorsally, in addition to few large spots; sixth pleonite with red transverse band only, this band continues to sternal surface and thus forms a continuous ring; antennular and antennal peduncles with some white and red blotches, flagella colourless; first pereiopods semi-transparent, with transverse red and white bands distally; second pereopods (main chelipeds) white with transverse, broad, bright red bands and greyish areas between them as following: one red–greyish–red band on merus, one red–greyish–red band on proximal two-thirds of chela palm and one red band across bases of fingers; proximal and distal areas of merus, carpus, distal palm and area adjacent to fingers tips white; third to fifth pereiopods (walking legs) semi-transparent, with four transverse, broad, bright red bands, one band per article (distal part of ischium, middle of merus and carpus and proximal part of propodus), and two or three white bands (on distal merus and carpus, and sometimes, ischium); telson translucent with bright red distal third; uropods translucent proximally, white on central part and red on distal thirds of exopod and endopod (
Figure 3A,B,E).
Distribution: Indo-West Pacific. Philippines: Ambil Island near Lubang Island (187–195 m) and southwest of Manila Bay, Luzon Island (188–191 m) [
18,
52]; Loyalty Islands east of New Caledonia (236 m) [
37]; Red Sea: off north-central coast of Saudi Arabia (321–471 m) (present study).
Ecology: In the Red Sea,
B. foveolatus associates with medium-sized (column in full extension ~15 cm, diameter with tentacles expanded ~20 cm), whitish, non-identified sea anemones (
Figure 3D) at depths of 321–471 m. The identification of the host, even if it had been collected, is extremely challenging as the deep-water actiniarian fauna of the Red Sea and Indian Ocean is generally poorly known [
53]. The hosts of the specimens from the Philippines and New Caledonia remain unknown.
Remarks:
Bruceclimenes foveolatus was described and illustrated in detail by Bruce [
18,
37] as
P. foveolatus. The male and female of
B. foveolatus from Saudi Arabia agree well with the original description of the species based on the material from the Philippines [
18]. The foveolation on the carapace and pleon in the present material is much less marked than in the type specimens (Bruce [
18]: Figure 6); however, this was also the case with the single male specimen from New Caledonia [
37]. The rostral dentitions of the Saudi Arabian male and female are 10/4 and 10/3, respectively, whereas Bruce [
18] gave 7–8/1–2 for the type series and 9/3 for the New Caledonian specimen; thus, the hitherto known rostral formula of
B. foveolatus is 7–10/1–3.
The colour pattern of the Red Sea specimens (
Figure 3A,B) is generally similar to that of the New Caledonian specimen, as illustrated by Bruce ([
37]: Figure 72), although in the former specimens, the spots are larger and less numerous, whilst the telson is mostly colourless rather than white, as in the latter specimen. In the original description of
B. foveolatus, Bruce ([
18]: 201) stated that the colour pattern of the type specimens from the Philippines is unknown. However, a colour slide of
B. foveolatus from the 1970s found by one of us (AA) in the slide collection of the Muséum National d’Histoire Naturelle in Paris (MNHN) presumably depicts one of the type specimens from the Philippines (
Figure 3E). Remarkably, the colour pattern of this specimen is almost identical to that of the two Red Sea specimens (
Figure 3A,B). Also noteworthy is the fact that in the Red Sea,
B. foveolatus was collected in slightly deeper water, at depths of 321 m and 471 m, whereas the specimens from the Philippines and New Caledonia came from depths ranging between 187 m and 236 m.
For the time being, the slight differences in morphology and colour patterns are treated as part of intraspecific variation in a single, widely distributed, deep-water species, apparently associated with sea anemones (see above), and ranging from the Red Sea to the Philippines and New Caledonia. However, a molecular comparison between the specimens of B. foveolatus from these three distant localities, which presently is impossible due to a lack of fresh material from the Philippines and New Caledonia, is highly desirable.
Michaelimenes Okuno, 2017
Michaelimenes Okuno [
20]: 2.
Diagnosis: See Okuno [
20].
Type species: Michaelimenes perlucidus (Bruce, 1969), originally described as Periclimenes perlucidus Bruce, 1969, by original designation (synonym: P. involens Bruce, 1996).
Other species included: Michaelimenes latipollex (Kemp, 1922); Michaelimenes platydactylus (Li, 2008); Michaelimenes sammy sp. nov.; Michaelimenes wirtzi (d’Udekem d’Acoz, 1996) comb. nov., originally described as Periclimenes wirtzi d’Udekem d’Acoz, 1996.
Hosts: Soft corals (Alcyonaria) and black corals (Antipatharia).
Bathymetric range: 15–494 m, but mainly in the depth range of 20–200 m.
Distribution: Indo-West Pacific from the Red Sea to Japan and French Polynesia [
16,
20,
54,
55], present study; East Atlantic from Madeira and Azores to the Cape Verde Islands [
56,
57].
Remarks: In the generic diagnosis of
Michaelimenes Okuno, 2017, [
20] stated that the pollex (=fixed finger) of the second pereiopod chela is proximally excavated, whereas the dactylus is characterised by a dorsal flange. All species currently assigned to
Periclimenes that present only the second condition, i.e., a flanged dactylus on the second pereiopod, were excluded from
Michaelimenes by Okuno [
20]. These species are
P. carinidactylus Bruce, 1969 from southern Australia [
58,
59];
P. compressus Borradaile, 1915 from the central Indian Ocean [
60,
61];
P. tenellus (Smith, 1882) from the western Atlantic [
62,
63,
64]; and
P. infraspinis (Rathbun, 1902) from the eastern Pacific [
64,
65]. Furthermore,
P. forcipulatus Bruce 1991 from the Loyalty Islands near New Caledonia [
37] was discussed by Okuno [
20] but excluded from
Michaelimenes because of the presence of minute teeth on the fingers of the first pereiopod in this species. Thus,
Michaelimenes hitherto included only three species, viz. the type species
M. perlucidus (Bruce, 1969),
M. latipollex (Kemp, 1922), both with several records from across the Indo-West Pacific; and
M. platydactylus (Li, 2008) from French Polynesia (see [
16,
20,
55,
59] and references therein).
Borradaile [
60] and Kemp [
16] considered
P. laccadivensis Alcock & Anderson, 1894 as possibly close to
M. latipollex (see also [
37,
66]). However, in
P. laccadivensis, the hepatic tooth is in a much lower position than in
M. latipollex ([
16,
67], see also Bruce [
37]: Figure 1a,b) and the below-described
M. sammy (
Figure 4B), whereas the dactylus of the second pereiopod is not flanged, although the pollex is shallowly excavated in its proximal part (Kemp [
16]: Figure 20b). The lateral flange on the dactylus of the second pereiopod is also present in many species of
Bathymenes, which differ from all the above-mentioned species by the presence of conical or subacute granules on the second pereiopods (at least on the chelae), and several other features [
44,
50]. In the molecular analyses,
P. laccadivensis was recovered either as a sister of
B. boucheti in Gan et al. [
21] or in an unresolved position within Clade 7 in Horká et al. [
8]. In our molecular analysis (
Figure 2),
P. laccadivensis clusters with two species of
Echinopericlimenes, although with relatively low branch support. Whatever its true affinities might be,
P. laccadivensis does not seem to be closely related to
Michaelimenes and is therefore not included in that genus.
On the other hand,
M. sammy and the eastern Atlantic
P. wirtzi have numerous morphological similarities, including the dactylar flange in the second pereiopod chela and what can be interpreted as a shallow proximal excavation on the pollex (cf. d’Udekem d’Acoz [
56]: Figure 7), share the same host group, i.e., Antipatharia ([
68]; see also insert in
Figure 2) and were recovered in sister position in our molecular analysis, although with a significant genetic divergence (
Figure 2). Therefore,
P. wirtzi is transferred to
Michaelimenes as
Michaelimenes wirtzi (d’Udekem d’Acoz, 1996) comb. nov. It must be noted, however, that the type species of
Michaelimenes,
M. perlucidus, lives on soft corals (Alcyonaria), but as pointed out by several previous studies, including Marin & Anker [
69] and Horká et al. [
8], host switches are very common in palaemonid shrimps. The only substantial deviation in
M. wirtzi from the generic diagnosis of
Michaelimenes in Okuno [
20] appears to be the shape of the rostrum, which is proximally arched, then descending and ascendant again towards the tip in
M. wirtzi (d’Udekem d’Acoz [
56]: Figure 2), whilst being more or less horizontal in the four Indo-West Pacific taxa (
Figure 3B; Okuno [
20]: Figure 3A).
Michaelimenes sammy sp. nov. (authored by Anker)
(urn:lsid:zoobank.org:act:8945277C-47BE-475E-B1E3-9E34ACB7D59C)
Type material: Saudi Arabia, Red Sea. Holotype: ovig. female (pocl 3.2 mm, cl 5.9 mm), FLMNH UF 68694, north of small islet ~25 km west of Jabal Alsabaya, Red Sea Decade Expedition, Leg 2, Phase 2, sta. NTN0133, 18.514100–18.510279, 40.666581–40.665597, max. depth 302 m, coll. Neptune Dive Team/F. Benzoni and F. Barreca, 01.04.2022 [OCX-037]. Paratypes: one female (pocl 2.1 mm, cl 3.9 mm), one ovig. female (pocl 2.8 mm, cl 5.6 mm), FLMNH UF 68692, same collection data as for the holotype (OCX-035*); one female (pocl 1.5 mm, cl 2.6 mm), KAUST, north of small islet 25 km west of Jabal Alsabaya, Red Sea Decade Expedition, Leg 2, Phase 2, sta. NTN0134, 18.812696, 40.637453, depth 191 m, coll. Neptune Dive Team/F. Benzoni and F. Barreca, 02.04.2022 (NTN0134-bycatch/OCX-104*); one ovig. female (pocl 2.9 mm, cl 5.3 mm), FLMNH UF 68693, south of Jazirt Abu Rashed, Neom, Deep Blue Expedition, sta. NTN0029, 27.600706, 35.334426, depth 494.1 m, coll. Neptune Dive Team/F. Marchese, 10.10.2020 (OCX-002*); one male (pocl 3.1 mm, cl 6.2 mm), RSRC (not deposited, see text), off Al Muwaileh, Neom, OceanX Red Sea Relationship Cultivation, sta. CHR0310, 27.632867, 35.311688, depth 407 m, coll. Chimaera Dive Team/F. Benzoni, F. Marchese and T.I. Terraneo, 23 June 2022 (CHR0310-8B/OCX-210); one ovig. female (pocl 2.3 mm, cl 4.4 mm), KAUST (not deposited, see text), north-west of Jeddah, 10 km west of Nelson Reef, Red Sea Decade Expedition, Leg 1, Phase 2, sta. NTN0121, 21.90876, 38.76103, depth 91.3 m, on black coral, coll. Neptune Dive Team/T.I. Terraneo, 22.02.2022 (NTN0121-Bio19B/OCX-202). *Designates sequenced specimen.
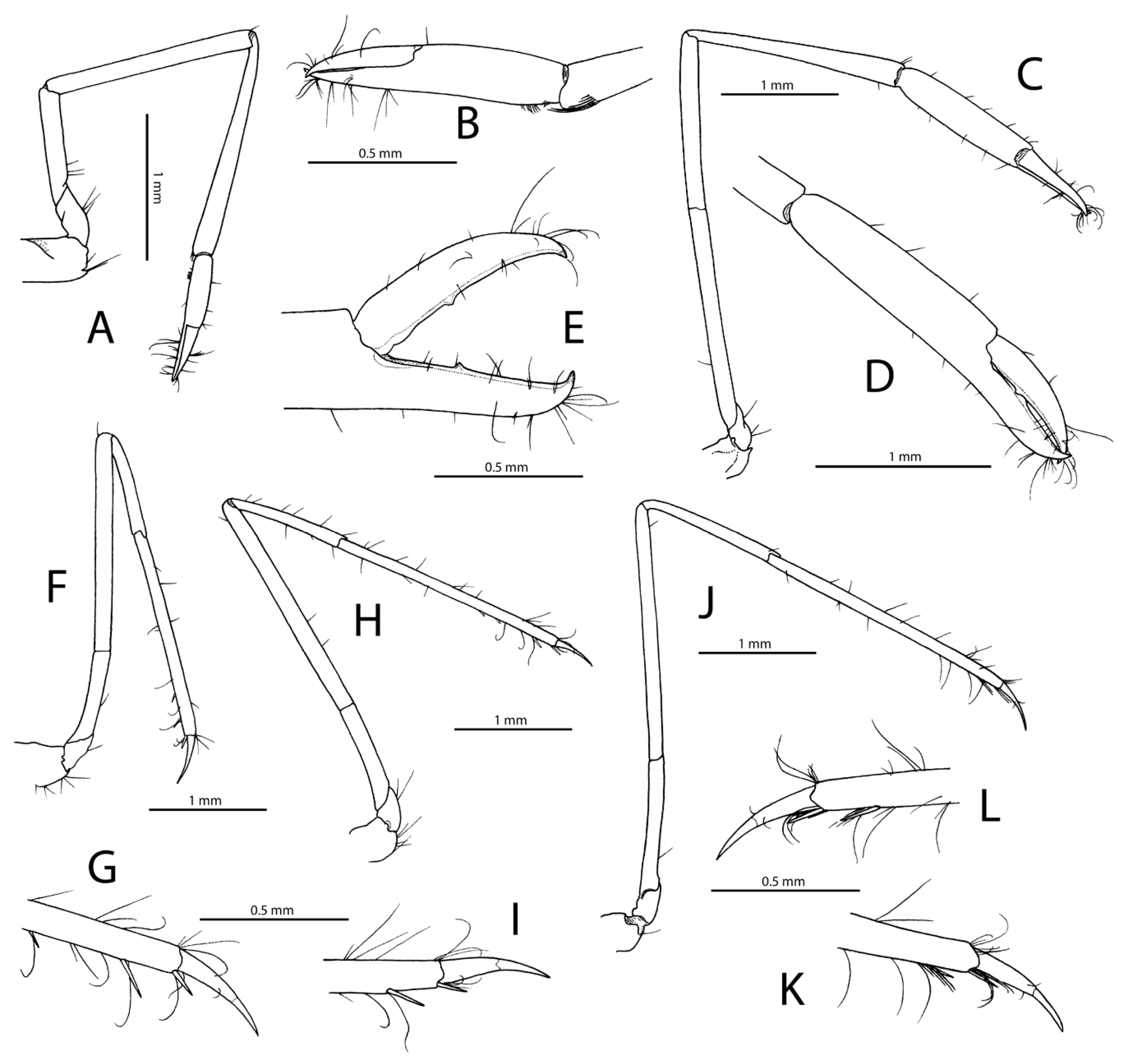
Description: Small-sized palaemonid shrimp (type material: pocl 1.5–3.2 mm) with cylindrical, not noticeably depressed or compressed body. Carapace smooth, glabrous. Rostrum (
Figure 4A,B,P–T; see also
Figure 6A,B) moderately high, straight, horizontal or shallowly concave near mid-length and ascendant towards tip, overreaching level of distal end of antennular peduncle, about as long or slightly shorter than carapace (postorbital length); dorsal margin armed with six to seven teeth, posterior-most tooth (rarely two teeth) situated posterior to or above posterior orbital margin; ventral margin straight or slightly convex, armed with two to three teeth on distal half; lateral carinae well marked; supraorbital eaves absent. Orbit (
Figure 4A,B) feebly developed; inferior orbital angle bluntly produced anteriorly. Antennal tooth (
Figure 4A,B) well developed, submarginal, sharp, overreaching level of distal end of inferior orbital angle. Hepatic tooth strong, sharp, situated posterior to and almost at same level as antennal tooth (
Figure 4A,B). Epigastric tooth (
Figure 4A,B,P–T) present, well-developed (missing, possibly broken off, in smallest specimen), somewhat removed from posterior-most tooth of rostral series, situated well posterior to the level of posterior orbital margin and slightly posterior to the level of hepatic tooth, semi-mobile, separated from carapace by well-demarcated suture. Supraorbital tooth absent. Ophthalmic somite without pronounced bec ocellaire (=interocular beak). Thoracic sternite 4 lacking median process. Epistome unarmed.
Pleon (
Figure 4C) smooth, glabrous, with all pleura rounded ventrally; pleonite 3 with posterior margin slightly produced, overhanging dorsal surface of pleonite 4, forming low hump; pleonite 6 not particularly elongate, 1.5 times as long as high, shorter than telson. Telson (
Figure 4D,E) narrow, gradually tapering towards posterior end; dorsal surface with two pairs of stout, submarginal spiniform setae located near mid-length and 0.8 of telson length, respectively; posterior margin subtriangular, with three pairs of spiniform setae, lateral ones stout, short, about 0.4 length of slender median ones and about 0.25 length of similarly stout intermediate ones.
Antennule (
Figure 4A,B) with first article of peduncle robust, much longer than the length of second and third articles combined; lateral margin somewhat convex, terminating distally in strong acute tooth; distal margin slightly convex mesial to distolateral tooth; stylocerite small, acute, overreaching 0.4 length of first article; ventromesial margin with small acute tooth; second and third articles subequal in length, much slenderer than first, unarmed; lateral flagellum biramous; accessory free ramus well developed, as long as fusion portion, but with poorly defined subdivisions, with numerous groups of aesthetascs. Antenna (
Figure 4A,B) with basicerite small, short, distally armed with sharp tooth; scaphocerite well developed; lateral margin straight, terminating in stout, sharp distolateral tooth, the latter not reaching bluntly angular distal margin of blade; carpocerite very short, not reaching 0.4 length of scaphocerite; flagellum normal, not thickened.
Mouthparts not especially modified, as illustrated (
Figure 4F–N and
Figure 5A). Mandible (
Figure 4F–H) without palp; molar process well developed; incisor process with four teeth distally. Maxillule, maxilla and maxillipeds 1 and 2 (
Figure 4I–N) without diagnostic features, as illustrated. Maxilliped 3 (
Figure 5A) with coxa bearing semicircular lateral plate; endopod moderately slender and setose; antepenultimate article clearly separated from basis, without spiniform setae; penultimate article about half as long as antepenultimate article, distally widening; ultimate article with stiff, non-spiniform setae; exopod short, reaching about 0.8 length of antepenultimate article; arthrobranch well developed.
Pereiopod 1 (
Figure 5B,C) slender, moderately elongate; ischium about half-length of merus; merus and carpus subequal in length; carpo-propodal grooming apparatus modestly developed; chela about 0.7 length of propodus; fingers about 0.7 length of propodus; dactylus not subspatulate; cutting edges of both fingers unarmed. Pereiopods 2 unequal in length, slightly dissimilar in shape and proportions, elongate, smooth (
Figure 5D–F,K–M and
Figure 6A,B). Major pereiopod 2 (
Figure 5D–F,K–M) with coxa lacking marked ventral process; basis very short; ischium slender, widening, markedly shorter than merus; merus more or less widening distally, unarmed; carpus variable in length and shape depending on age of individuals, short and cup-shaped in large specimens (
Figure 5K), more elongate in smaller specimens (
Figure 5L), always widening distally; chela feebly to moderately enlarged depending on age of individuals, particularly robust in larger specimens (
Figure 5K and
Figure 6A,B); palm cylindrical in cross-section, 4–5 times as long as high, smooth, sparsely setose; fingers 0.4–0.5 length of palm, slightly gaping distally, with crossing fingertips; dactylus with conspicuous lateral flange, especially well-marked in large specimens; pollex with shallow excavation proximally; cutting edge of each finger armed with two to three rounded or subacute teeth on proximal third of finger length. Minor pereiopod 2 variously smaller and weaker than major pereiopod 2 (
Figure 6A,B), generally similar in shape; dactylar flange much less obvious than in major pereiopod 2.
Ambulatory pereiopods (P3–5) similar, slender, elongate (
Figure 5G–J); ischium about 0.4 length of merus; merus about 14 (P3) to 15 (P5) times as long as wide, about twice as long as carpus; propodus subequal to merus in length; ventral margin of P3 and P4 with 3 spiniform setae on distal half and 1 pair of spiniform setae adjacent to dactylar base; ventromesial margin of P5 with 1 slender spiniform seta at about 0.9 length of propodus and one longer and stouter distal spiniform seta adjacent to dactylar base; propodal cleaning brush not distinct; dactylus less than 0.2 length of propodus, simple, moderately stout, subtly biunguiculate, with minute secondary unguis (sometimes notch) on stout corpus and slender, distinctly curved terminal unguis.
Male first pleopod without notable features. Male second pleopod (
Figure 5N,O) with endopod bearing typical appendices masculina and interna, former not reaching tip of latter, with stiff setae on apex and along distal half of mesial margin. Uropod (
Figure 4O) exceeding telson; protopod with two distally blunt lobes; exopod ovate-rectangular, rounded distally; distolateral tooth smooth; adjacent spiniform seta moderately strong; endopod narrowly ovoid, slightly shorter than exopod, its distal margin reaching to about 0.9 length of exopod.
Colour pattern: Translucent with pale orange tinge; ventral side of cephalothorax and pleon reddish; second pereiopods translucent with pale reddish fingers; walking legs with yellow spots; tail fan largely translucent (
Figure 6D).
Etymology: The new species is named after our colleague, Sammy De Grave (Oxford University Museum of Natural History), a well-known expert on caridean shrimps with a slight fondness for symbiotic palaemonid shrimps. Sammy’s first (given) name is used as a noun in apposition.
Distribution: Presently known only from northern and central Red Sea, off the coast of Saudi Arabia.
Ecology: The recorded depth range of the type series is 91.3–494.1 m; one of the specimens was found on a colony of bushy black corals of the family Myriopathidae (
Figure 6C) indicating a possible association between
M. sammy and this group of cnidarians.
Remarks: The overall combination of morphological characteristics strongly suggests that
M. sammy is a member of
Michaelimenes, displaying all essential diagnostic features of the genus (
Figure 4 and
Figure 5; Okuno [
20]). The proximal excavation on the cutting edge of the second pereiopod pollex is relatively shallow and best visible on the major chela of the largest specimens (
Figure 5K; see also below).
Michaelimenes sammy can be easily separated from both
M. perlucidus and
M. latipollex by the obscurely biunguiculate dactylus of the third to fifth pereiopods (
vs. strongly biunguiculate in
M. perlucidus and
M. latipollex); from
M. perlucidus by the noticeably larger distance between the epigastric tooth and the posterior-most rostral tooth; from
M. latipollex by the much more slenderer dactylus of the third to fifth pereiopods (in addition not being strongly biunguiculate, see above); and from
M. platydactylus by the posterior-most tooth of the dorsal rostral series in epigastric position, as in
M. latipollex (
vs. situated above the orbit in
M. platydactylus), and the noticeably slenderer third pereiopod (cf. Kemp [
16], Okuno [
20] and Li [
55]). In the position of the hepatic tooth,
M. sammy is most similar to
M. latipollex; in
M. perlucidus and
M. platydactylus, the hepatic tooth is closer to and slightly lower than the antennal tooth (
Figure 4B; cf. Kemp [
16], Okuno [
20] and Li [
55]). On the other hand, in the general shape of the dactylus of the walking legs,
M. sammy is closer to
M. platydactylus.
The separation of
M. sammy from the eastern Atlantic
M. wirtzi is relatively straightforward. The two species greatly diverge in the shape and length of the rostrum, which in the new species is usually straight (horizontal) or at most slightly concave and then ascendant, armed with two or three teeth ventrally, and with the tip only slightly overreaching the distal end of the antennular peduncle (
vs. proximally strongly arched and then concave all the way to the tip, armed with five to seven teeth on the ventral margin in the adults, and with the tip by far overreaching the distal end of the antennular peduncle in
M. wirtzi) (
Figure 4B,P–T; cf. d’Udekem d’Acoz [
56]: Figures 1 and 2A, D–H). They also differ in the proportions of the penultimate article of the third maxilliped and the degree of convexity of the distal margin of the antennal scaphocerite blade (
Figure 4 and
Figure 5A; cf. d’Udekem d’Acoz [
56]: Figures 3A and 4D).
The excavation of the pollex of the second pereiopod chela, which was considered by Okuno [
20] as one of the most important diagnostic characters of
Michaelimenes, is quite deep in
M. perlucidus, at least as illustrated by Okuno ([
20]: Figure 1B,E), but may be shallower and more subtle in other species, including
M. sammy (
Figure 5K). Kemp ([
16]: 151) described the cutting-edge area of the second pereiopod chela as “the single tooth on the dactylus fitting into a recess in the fixed finger”, whereas Li ([
55]: 233) stated that the “fixed finger length larger than dactylar teeth, with proximal excavation present to fit proximal part of dactyl cutting edge”, although he did not show it in his illustration of the opened chela fingers (cf. [
56]: Figure 19D). In fact, the presence or absence of a proximal excavation on the pollex of the second pereiopod chela alone should not exclude at least some species of
Periclimenes from
Michaelimenes. Indeed, as noted by Okuno [
20], the current taxonomic status of four species of
Periclimenes characterised by the flanged dactylus of the second pereiopod chela, but apparently without a proximal excavation on the pollex, viz.
P. carinidactylus,
P. compressus,
P. tenellus and
P. infraspinis, may need to be reassessed in the future. This is especially true for
P. tenellus, which possesses at least a shallow depression on the pollex of the second pereiopod chela [
63,
64] and may well represent a western Atlantic species of
Michaelimenes. In the absence of DNA data for these taxa, it is more prudent not to transfer
P. tenellus and the remaining three species from
Periclimenes to
Michaelimenes. Nevertheless, they need to be considered in the comparison with the herein-described new species of
Michaelimenes.
Michaelimenes sammy can be easily separated from
P. carinidactylus by the submarginal position of the dorsal spiniform setae of the telson (
vs. clearly marginal in
P. carinidactylus), the hepatic tooth situated at the same level as the antennal tooth (
vs. noticeably lower in
P. carinidactylus), the posterior-most tooth in the dorsal rostral series in epigastric position (
vs. much more anterior in
P. carinidactylus) and the slenderer dactylus of the third to fifth pereiopods (
vs. much stouter in
P. carinidactylus) (
Figure 5F–I; cf. Bruce [
59]: Figures 2A,F and 4K); from
P. compressus by two features on the third and fourth pereiopods, namely, their propodi armed with spiniform setae distally and their dactyli longer and less curved, and the first pereiopod carpus much longer than the chela (
vs. distinctly shorter than the chela in
P. compressus) (
Figure 5F–H; cf. Bruce [
61]: Figure 4G); from
P. tenellus by the hepatic tooth as large as and at the same level as the antennal tooth, also being in a more posterior position (
vs. much larger and lower than the antennal tooth and much closer to it in
P. tenellus), and by the obscurely biunguiculate dactylus of the third to fifth pereiopods (
vs. strongly biunguiculate in
P. tenellus) (
Figure 4B and
Figure 5F–H; cf. Anker et al. [
64]: Figure 3b, h); and finally, from
P. infraspinis by the hepatic tooth as large as and at the same level as the antennal tooth (
vs. much lower in
P. infraspinis), the dorsally non-elevated, more or less horizontal rostrum (
vs. dorsally strongly convex in
P. infraspinis), and the subtly biunguiculate dactyli on the third to fifth pereiopods (
vs. strongly biunguiculate in
P. infraspinis) (
Figure 4B and
Figure 5F,G; cf. Holthuis [
63]: pl. 13, figs. a, k).
Rubiyana gen. nov. (authored by Anker)
(urn:lsid:zoobank.org:act:85F38AF2-5452-4C32-A681-994245DB129C)
Diagnosis: Small-sized palaemonid shrimps (pocl 1.7–3.2 mm). Body subcylindrical, neither strongly compressed not depressed, smooth. Rostrum well developed, high, horizontal, reaching or almost reaching end of antennular peduncle; dorsal margin straight, with eight to nine teeth; ventral margin almost straight, with two to three teeth in distal half; rostral carina moderate, not extending beyond posterior-most tooth. Inferior orbital angle strongly anteriorly produced, not reaching beyond antennal tooth, blunt. Antennal and hepatic teeth well-developed, acute; latter stouter and situated slightly below former, not reaching anterolateral margin of carapace. Epigastric tooth present, with or without basal suture. Supraorbital teeth absent. Thoracic sternite 4 without median process. Pleonite 3 not carinate, slightly posteriorly produced, forming very low hump; pleura of pleonites 1–5 posteroventrally rounded; pleonite 6 elongate. Telson with two pairs of dorsal spiniform setae in posterior half, inserted at some distance from margin, and three pairs of short spiniform setae on posterior margin, mesial ones with secondary setules; posterior margin bluntly triangular. Antennule typical for family; stylocerite short, acute; distolateral tooth of first article of peduncle sharp; lateral flagellum biramous, with short fused basal part; secondary ramus poorly developed. Antenna typical for family; scaphocerite well developed, with broad blade and stout distolateral tooth falling short of anterior margin of blade; carpocerite short. Eyes well developed, with large globular cornea. Mandible without palp; molar and incisor processes normal. Maxillule with deeply bilobed palp (endopod), ventral lobe strongly hooked distally. Maxilla with reduced coxal endite and bilobed basial endite; palp (endopod) large; scaphognathite broad. Maxilliped 1 with endites fused; palp (endopod) small, entire; exopod long, with well-developed caridean lobe; epipod small, subtriangular. Maxilliped 2 with normal endopod and exopod; epipod small, ovate, without podobranch. Maxilliped 3 with coxa bearing broad, semicircular, lateral plate; endopod moderately slender; ischiomerus (antepenultimate article) not clearly separated from basis, without spiniform setae; ultimate article with robust, but non-spiniform setae; exopod short; arthrobranch absent. Pereiopod 1 slender; carpus more or less elongate; carpo-propodal brush present; chela not elongate, not inflated, with fingers as long as or shorter than palm; cutting edges of fingers unarmed. Pereiopods 2 moderately stout, similar in shape, slightly unequal in size, smooth; merus and carpus unarmed; carpus more or less elongate; chela not particularly swollen; palm subcylindrical, moderately elongate; fingers much shorter than palm; cutting edge of pollex and dactylus each armed with small tooth at proximal 0.3 of length; tooth–fossa snapping mechanism lacking. Pereiopods 3–5 slender; propodus with slender spiniform setae on ventral margin; dactylus simple. Male pleopod 1 and 2 unknown. Uropod with protopod with 2 bluntly ending lobes; exopod unarmed, except for slender, distolateral spiniform seta; endopod narrower and significantly shorter than exopod, not overreaching 0.75 of exopod length.
Type species: Rubiyana arabica sp. nov. (authored by Anker), by present designation and monotypy.
Other species included: None (but see remarks on Periclimenes kallisto Bruce, 2008 below).
Etymology: The name of the new genus derives from the Arabic word “rubiyan”, for shrimp or prawn. Gender feminine.
Hosts: Soft corals (Malacalcyonacea: Nephtheidae).
Remarks: Our molecular analysis recovered the type species of
Rubiyana very distantly from
Periclimenes s.
str., more precisely as a sister to a small clade composed of
Ancylocaris brevicarpalis,
Periclimenes affinis and
P. kallisto (
Figure 2). A phylogenetic affinity between
P. affinis and
A. brevicarpalis was first indicated by Gan et al. [
21], whose molecular analysis did not include
P. kallisto. Horká et al. [
8], using sequences of
P. affinis from Gan et al. [
21] and adding their sequences of
P. kallisto, recovered a clade composed of
A. brevicarpalis,
P. affinis and
P. kallisto, with the same topology as in our analysis (
Figure 2; [
8]: Figures 2 and 3). In the revalidation of
Ancylocaris, Ďuriš & Horká [
10] mentioned some morphological characters shared by
A. brevicarpalis,
P. affinis and
R. kallisto, and also between
A. brevicarpalis and
P. affinis, and between
P. affinis and
R. kallisto. With the addition of
Rubiyana, this lineage is hereafter called the
Rubiyana–
Ancylocaris clade (
Figure 2). In addition, Bruce [
19] stated that two further species of
Periclimenes may be part of the same species group as
P. affinis and
R. kallisto, more precisely,
P. canalinsulae Bruce & Coombes, 1997 and
P. jugalis Holthuis, 1952.
The type species of
Rubiyana is remarkably similar to
P. kallisto. In both species, the uropodal endopod is markedly reduced in size and length, reaching only 0.75 length of the exopod (
Figure 7O; Bruce [
19]: Figure 2I). In other words, in both species, the distal margin of the uropodal endopod does not exceed the level of the distolateral tooth of the uropodal exopod. The greatly shortened uropodal endopod is one of the most diagnostic features of
Rubiyana, and the presence of this feature in
P. kallisto, but not in
A. brevicarpalis (Lee & Koh [
70]: Figure 2L), is somewhat intriguing given the obtained topology (
Figure 2). The relative size and length of the uropodal endopod and exopod were neither described nor illustrated for
P. affinis [
59,
71] or
P. jugalis [
72]. Bruce [
59] only noted that in
P. affinis, the “uropods present no specific features”, whilst Holthuis [
72] stated that in
P. jugalis, the “uropods reach beyond the end of the telson, the endopod, however, fails to reach the end of the posterior spinules of the telson”, suggesting that the endopod is only slightly shorter than the exopod, as in
P. canalinsulae (Bruce & Coombes [
73]: Figure 3N). The present topology (
Figure 2) suggests that the unusual configuration of the uropod in
R. arabica and
P. kallisto may be the result of convergent evolution. Therefore,
P. kallisto,
P. affinis,
P. jugalis and
P. canalinsulae are not included in
Rubiyana, awaiting these and perhaps further species of the
Rubiyana–
Ancylocaris clade to be described in more detail and genetically analysed.
Hosts are known for
R. arabica (soft corals (Nephtheidae), see below),
P. kallisto (bushy black corals (Antipatharia), see Bruce [
19]),
P. affinis (several genera and species of feather stars (Crinoidea), see Holthuis [
59] and Bruce [
71]), and
A. brevicarpalis (several genera and species of sea anemones (Actiniaria), see Lee & Koh [
70], Anker & De Grave [
74] and Ďuriš & Horká [
10] and references therein), but not for
P. jugalis and
P. canalinsulae [
72,
73]. Thus, within the
Rubiyana–
Ancylocaris clade, the morphologically and ecologically most distinctive lineage is the monotypic genus
Ancylocaris.
Rubiyana can be separated from
Periclimenes s.
str. and all other species presently referred to
Periclimenes (except for
P. kallisto), as well as
Ancylocaris and all other satellite genera, by the above-mentioned, unique configuration of the uropod. The new genus also differs from
Periclimenes s.
str. as dactyli of the third to fifth pereiopods being simple (
Figure 8F–K) vs. strongly biunguiculate in
Pericilimenes s.
str. [
75,
76,
77]. Several other characteristics of
Rubiyana, such as the elongate sixth pleonite and the absence of arthrobranch on the third maxilliped, may be revealed as important for differentiation between the new genus and numerous clades of
Periclimenes s.
l. Male specimens of
Rubiyana are needed to verify the configuration of the appendix masculina of the second pleopod and the shape of the endopod of the first pleopod (simple or distally bilobed, as in
P. kallisto).
Rubiyana arabica sp. nov. (authored by Anker)
(urn:lsid:zoobank.org:act:96CA27B0-E5D7-4C3A-BC20-BC204EE1169A)
Type material: Saudi Arabia, Red Sea. Holotype: ovig. female (pocl 3.2 mm, cl 6.5 mm), RSRC (not deposited, see text), northwest of Farasan Islands, Red Sea Decade Expedition, Leg 2, Phase 2, sta. CHR0233, 17.387135–17.387619, 40.827209–40.828850, depth range 104–248.5 m, coll. Chimaera Dive Team/F. Benzoni and F. Barreca, 28.03.2022 (CHR0233-bycatch/OCX-073*). Paratype: ovig. female (pocl 2.9 mm, cl 6.0 mm), KAUST (not deposited, see text), about 15 km off Jeddah, Red Sea Decade Expedition, Leg 1, Phase 2, sta. CHR0198, 21.645553, 38.909941, depth 96 m, coll. Chimaera Dive Team/T.I. Terraneo, 23.02.2022 (CHR0198-Bio15A/OCX-198). *Designates sequenced specimen.
Description: Small-sized palaemonid shrimp (type material: pocl 2.9–3.2 mm) with cylindrical, not noticeably depressed or compressed body. Carapace smooth, glabrous. Rostrum (
Figure 7A–C,P) moderately high, straight, horizontal, not reaching level of distal end of antennular peduncle, about as long as carapace (postorbital length); dorsal margin armed with eight to nine teeth, all of them situated anterior to posterior orbital margin; ventral margin straight, armed with two to three teeth on distal third; lateral carinae obsolete; supraorbital eaves absent. Orbit (
Figure 7A,B) feebly developed; inferior orbital angle bluntly produced anteriorly. Antennal tooth (
Figure 7B) well developed, submarginal, sharp, reaching level of distal end of inferior orbital angle. Hepatic tooth strong, sharp, situated slightly ventrally to antennal tooth (
Figure 7A,B). Epigastric tooth (
Figure 7A–C,P) distinctly removed from posterior-most tooth of rostral series, situated well posterior to level of posterior orbital margin and hepatic tooth, separated from carapace by suture. Supraorbital tooth absent. Ophthalmic somite without pronounced bec ocellaire (=interocular beak). Thoracic sternite 4 lacking median process. Epistome unarmed.
Pleon (
Figure 7D) smooth, glabrous, with all pleura rounded ventrally; pleonite 3 with posterior margin slightly produced, overhanging dorsal surface of pleonite 4, forming low hump; pleonite 6 elongate, more than 2.5 times as long as high, about 1.2 times longer than telson (
Figure 9C). Telson (
Figure 7E) narrow, distinctly tapering from about 0.4 of length; dorsal surface with two pairs of spiniform setae, both located in posterior half and inserted at some distance from lateral margin; posterior margin triangular, with three pairs of spiniform setae, lateral ones very short, about 0.3 length of median ones and about 0.2 length of intermediate ones, latter much more robust.
Antennule (
Figure 7A,B) with first article of peduncle robust, longer than length of second and third articles combined; lateral margin somewhat convex, terminating distally in strong acute tooth; distal margin strongly convex mesial to distolateral tooth; stylocerite small, acute, reaching 0.4 length of first article; ventromesial margin with small acute tooth; second and third articles subequal in length, much slenderer than first, unarmed; lateral flagellum biramous; accessory free ramus composed of two to three poorly defined subdivisions, with groups of aesthetascs. Antenna (
Figure 7A,B,G) with basicerite short, distally armed with small sharp tooth; scaphocerite well developed; lateral margin straight, terminating in acute distolateral tooth, latter falling well short of bluntly angular distal margin of blade; carpocerite very short, not reaching 0.4 length of scaphocerite; flagellum normal, not thickened.
Mouthparts not especially modified, as illustrated (
Figure 7H–N). Mandible (
Figure 7H,I) without palp; molar process well-developed; incisor process with three teeth distally. Maxillule, maxilla and maxillipeds 1 and 2 (
Figure 7J–M) without specific features, as described for genus. Maxilliped 3 (
Figure 7N) with coxa bearing semicircular lateral plate; endopod moderately slender and setose; antepenultimate article not clearly separated from basis, without spiniform setae; penultimate article 0.6 times as long as antepenultimate article, much slenderer; ultimate article with stiff, non-spiniform setae; exopod short, not reaching 0.7 length of antepenultimate article; arthrobranch absent.
Pereiopod 1 (
Figure 8A,B) slender, elongate; ischium about 0.6 length of merus; merus 0.9 length of carpus, somewhat angular ventrodistally; carpo-propodal grooming apparatus normally developed; chela about half-length of propodus; fingers about 0.7 length of propodus; dactylus conical, not subspatulate; cutting edges of both fingers unarmed. Pereiopods 2 subequal in length, similar in shape, slender, elongate (
Figure 8C–E and
Figure 9C); coxa with stout ventromesial process; basis very short; ischium slender, almost 10 times as long as wide, 1.2 times as long as merus; merus about 0.8 length of carpus, unarmed; carpus subcylindrical, widening distally, about as long as chela; chela not inflated, elongate; palm cylindrical in cross-section, about 3.8 times as long as high, smooth, sparsely setose; fingers almost 0.6 length of palm, slightly gaping in distal half, with crossing fingertips; dactylus without lateral flange; cutting edge of each finger armed with small tooth at about 0.3 of finger length.
Ambulatory pereiopods (P3–5) similar, slender, elongate, somewhat increasing in length from third to fifth (
Figure 8F–L); ischium ranging from about half-length of merus (P3 and P4) to 0.6 length of merus (P5); merus ranging from 15.4 times (P3) to 18.4 times (P5) as long as wide, about twice as long as carpus; propodus as long as (P3 and P4) or slightly longer (P5) than merus; ventral margin of P3 and P4 with three to four spiniform setae on distal third or starting from 0.6 of propodal length, including distal one adjacent to dactylar base; ventromesial margin of P5 with two spiniform setae, including distal one adjacent to dactylar base; propodal cleaning brush reduced to two rows on distal ventrolateral surface; dactylus about 0.2–0.25 length of propodus, simple, moderately slender, with terminal unguis demarcated, slightly curved.
Male pleopods unknown. Uropod (
Figure 7O) exceeding telson; protopod with two distally blunt lobes; exopod ovate-rectangular, rounded distally; distolateral tooth smooth; adjacent spiniform seta moderately strong; endopod narrowly ovoid, much smaller and shorter than exopod, its distal margin reaching to about 0.75 length of exopod.
Colour pattern: Largely translucent (somewhat milky opaque when stressed), with dissipated red chromatophores on carapace and pleon; conspicuous, continuous, red, longitudinal line present on sternum of carapace and pleon; antennular peduncles and pleopods with red bands; second pereiopods with yellow-red tinge distally; eggs pale yellow-green (
Figure 9B,C).
Etymology: The specific epithet arabica (Latin adjective for Arabian/Arabic) refers to the Arabian region, which includes the Red Sea, where the new species occurs.
Distribution: Presently known only from the central and southern Red Sea, off the coast of Saudi Arabia.
Ecology: One of the specimens was found on a large soft coral of the genus
Dendronephthya (
Figure 9A,B), indicating a possible association between
R. arabica and this group of cnidarians.
Remarks: As mentioned above,
R. arabica is morphologically most similar to two species currently placed in
Periclimenes, viz.
P. kallisto and
P. affinis. These two species were excluded from
Rubiyana largely for phylogenetic reasons since their inclusion in the new genus would create a paraphyletic grouping (cf.
Figure 2). On the other hand, applying the generic name
Ancylocaris for the entire
Rubiyana–
Ancylocaris clade would result in a morphologically and ecologically wide, rather impractical genus. The inclusion of additional taxa with molecular data in the
Rubiyana–
Ancylocaris clade would certainly better resolve the relationships within this group. For the time being, both
P. kallisto and
P. affinis are retained in
Periclimenes (see also above).
Rubiyana arabica can be separated from the antipatharian-associated
P. kallisto by several morphological features, including the rostral formula with more numerous teeth on both margins, 9+1/2–3 (
vs. 6–7+1/1 in
P. kallisto); the epigastric tooth with distinct basal suture, i.e., semi-mobile (
vs. fixed in
P. kallisto); the much more elongate carpi of the first and second pereiopods; the second pereiopods subequal in size and with both chela fingers armed with a small tooth on their cutting edges (
vs. somewhat unequal and with only the cutting edge of the pollex armed with small tooth in
P. kallisto); and the ambulatory pereiopods terminating in simple, not subtly biunguiculate dactyli (
vs. with subtly biunguiculate dactyli in
P. kallisto) (
Figure 7 and
Figure 8; cf. Bruce [
19]: Figures 1–5).
Rubiyana arabica is also distinguishable from the crinoid-associated
P. affinis, for instance, by the carapace with a semi-mobile epigastric tooth distinctly removed from the posterior-most rostral tooth (
vs. without epigastric tooth in
P. affinis, in which all teeth on the mid-dorsal + rostral carina are equidistant); the ventral margin of the rostrum with two or three teeth (vs. with only one tooth in
P. affinis); the much slenderer, subequal second pereiopods, with elongate, cylindrical carpus (
vs. more robust, more unequal, and with cup-shaped carpus at least in the major cheliped, in
P. affinis); the ambulatory pereiopods terminating in slender, gradually tapering dactyli (
vs. with much stouter dactyli in
P. affinis); and the telson with the dorsal spiniform setae distinctly removed from the margin (
vs. submarginal in
P. affinis) (
Figure 7 and
Figure 8; cf. Holthuis [
71] and Bruce [
59]: Figure 2 and Figures 1–3, respectively). Finally, the new species can be easily separated from
P. jugalis and
P. canalinsulae, for example, from
P. jugalis by the almost straight rostrum (
vs. strongly arched in
P. jugalis), the cutting edge of the pollex with only one tooth (
vs. two in
P. jugalis), and the distinctly slenderer dactylus of the third pereiopod (
Figure 7B and
Figure 8E,G; cf. Holthuis [
72]: Figure 26); and from
P. canalinsulae by the very different armature of the rostrum, the different position of the hepatic tooth relative to the antennal tooth, and the much shorter uropodal endopod (
Figure 7B,O; cf. Bruce & Coombes [
73]: Figure 3B,N).
Apopontonia Bruce, 1976
Apopontonia falcirostris Bruce, 1976
Apopontonia falcirostris Bruce [
17]: 303, Figures 1–5; Bruce [
78]: 3, 26; De Grave [
79]: 121, Figure 1; Marin [
80]: 218, Figure 1; Mitsuhashi & Chan [
81]: 37 (key); Bruce [
82]: 495.
Material examined: Saudi Arabia, Red Sea. One male (pocl 1.4 mm, cl 2.2 mm), KAUST, east of Farasan Islands, Red Sea Decade Expedition, Leg 2 Phase 2, sta. CHR0230, 16.931091, 41.140585, depth 88 m, inside large white sponge, coll. Chimaera Dive Team/F. Benzoni and F. Barreca, 25.03.2022 (CHR0230-Bio18B/OCX-114).
Description: See Bruce [
17] for original description and De Grave [
79] and Marin [
80] for additional illustrations.
Colour pattern: Unknown.
Distribution: Indo-West Pacific: Red Sea, Madagascar, Maldives, Vietnam, Papua New Guinea, Australia [
17,
18,
79,
80,
82], present study.
Ecology: The Red Sea specimen was found inside a non-identified, white, large sponge collected at 88 m, which is the deepest record for the species. The Madagascan type specimen was collected by a bottom trawl at 72 m and may have been also associated with a sponge (Bruce [
17]: 311). All other specimens came from shallower water (i.e., scuba diving depths) and were associated with various sponges [
18,
79,
82].
Remarks: The single specimen of
A. falcirostris collected near the Farasan Islands represents the first record of this uncommon species and the monotypic genus
Apopontonia [
82] for the Red Sea and the Arabian region. The Red Sea specimen is a young male (with a short appendix masculina) that agrees rather well with the very detailed description of the female holotype of
A. falcirostris provided by Bruce [
17]. The few differences observed, e.g., in the proportions of the perfectly equal and symmetrical second pereiopods, are certainly due to the present specimen being male and of smaller size.
Bathymenes Kou, Li & Bruce, 2016
Bathymenes Kou et al. [
44]: 171; Ďuriš & Šobáňová [
50]: 383.
Diagnosis: See Kou et al. [
44] and Ďuriš & Šobáňová [
50].
Type species: Bathymenes alcocki (Kemp 1922), originally described as Periclimenes alcocki Kemp, 1922, by original designation.
Other species included: See Kou et al. [
44] and Ďuriš & Šobáňová [
50] for full list of species included prior to the present study. Additional species included herein:
Bathymenes foresti (Bruce, 1981) comb. nov.;
Bathymenes granuloides (Hayashi, 1986) comb. nov.;
Bathymenes crosnieri (Li & Bruce, 2006) comb. nov.; all three species originally described under
Periclimenes (see discussion below).
Hosts: Sea pens (Pennatulacea); sea anemones (Actiniaria) carried by pagurids; sea urchins (Echinoidea); unknown for most species [
44].
Bathymetric range: 187–824 m, but mainly in the depth range of 250–600 m.
Distribution: Indo-Pacific from East Africa to Chile; most species reported from the tropical western Pacific; not yet reported from the Red Sea.
Remarks: While comparing
Bruceclimenes with
Bathymenes (see above), some taxonomic incongruences were noted for the later genus. At least three species previously assigned to
Periclimenes but with clear morphological connection to
Bathymenes were not treated by previous authors [
44,
50]; for all three taxa, molecular data are currently lacking.
The species originally described as
Periclimenes crosnieri by Li & Bruce [
83] has all diagnostic characters of
Bathymenes, as originally defined by Kou et al. [
44], including the telson dorsally armed with four pairs of spiniform setae, the epigastric tooth clearly separated from the rostral series, and the second pereiopods finely granulated and with a lateral flange on the dactylus. Therefore, this species is herein transferred to
Bathymenes, as
Bathymenes crosnieri (Li & Bruce, 2006) comb. nov.
Along the same lines,
Periclimenes foresti Bruce, 1981 and
Periclimenes granuloides Hayashi, 1986 (perhaps a junior synonym of
P. foresti, see Li et al. [
48]) from the
P. foresti complex (see above) display all diagnostic characters of
Bathymenes sensu Ďuriš & Šobáňová [
50], and are presumably closely related to
B. boucheti (Li, Mitsuhashi & Chan, 2008) [
18,
47,
48]. Therefore, they too are transferred to
Bathymenes, as
Bathymenes foresti (Bruce, 1981) comb. nov. and
B. granuloides (Hayashi, 1986) comb. nov., respectively.
The generic diagnosis of
Bathymenes in Ďuriš & Šobáňová [
50], after inclusion of
B. boucheti, and now also
B. foresti and
B. granuloides, is lacking a single autapomorphic feature immediately distinguishing it from other palaemonid genera. Most importantly, the dorsal armature of the telson in
Bathymenes, as emended by Ďuriš & Šobáňová [
50], now ranges from two to seven pairs of spiniform setae [
44]. Variation also affects the dactylus of the ambulatory pereiopods, which can be simple or biunguiculate, the development of the cornea (usually reduced, but not always) and some other important features. The molecular analysis of two mitochondrial and two nuclear genes (
Figure 2) provides no support for the distinction between the more typical species of
Bathymenes, i.e., those with three or more pairs of dorsal spiniform setae on the telson, represented by
B. ngi (Li, Mitsuhashi & Chan, 2008),
B. leptunguis (Li, Mitsuhashi & Chan, 2008) and
B. sandybrucei (Mitsuhashi & Chan, 2009), and some species formerly assigned to the
P. foresti complex with two pairs of dorsal spiniform setae on the telson, such as
B. boucheti (see also Horká et al. [
8], and Ďuriš & Šobáňová [
50]). However, more species of
Bathymenes need to be included in future analyses to shed more light on the internal structure of the genus and perhaps propose a better classification scheme (two genera?), based on a combination of morphological and phylogenetic data.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








