
Diversity of the Pteridoflora of Montane Northwestern Mexico

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area

2.2. Distributional Data and Analysis
3. Results
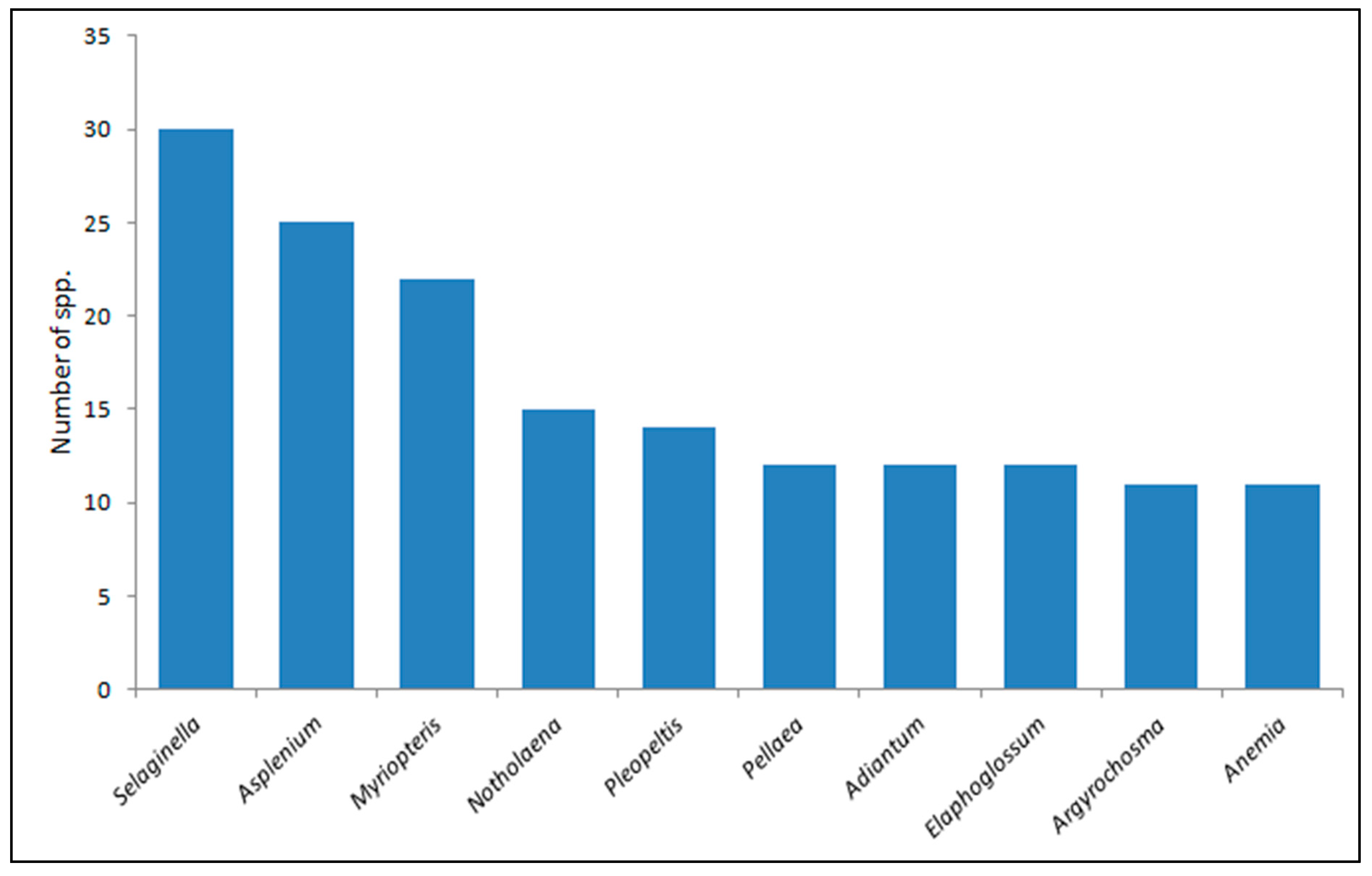
3.1. Floristics
3.2. Taxonomic Biodiversity Index
3.3. Threatened Species
3.4. Pteridodiverse States of the SMOc
3.5. Life Forms
3.6. Elevation
3.7. Vegetation Types Distribution
3.8. Geographic Distribution
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Species | LF | VT | GD | SD |
|---|---|---|---|---|---|
| LYCOPODIIDAE | |||||
| ISOETACEAE | |||||
| 1 | Isoetes mexicana Underw. | Hy | R, S | Mx | C, D |
| 2 | Isoetes montezumae A. A. Eaton | Hy | R | M | A, J, N, Z |
| 3 | Isoetes pringlei Underw. | Hy | R, G | Mx | J |
| LYCOPODIACEAE | |||||
| 4 | 1 Palhinhaea cernua (L.) Franco & Vasc. | T | M, BC, Q | Cos | J, N |
| 5 | Phlegmariurus cuernavacensis (Underw. & F. E. Lloyd) B. Øllg. | Ep/E | C, M, Q | M | I |
| 6 | Phlegmariurus taxifolia (Sw.) Á. Löve & D. Löve | E | S | Nt | I |
| SELAGINELLACEAE | |||||
| 7 | Selaginella arizonica Maxon | T/Ep | R, M, Q, X | Na | C, O |
| 8 | Selaginella arsenei Weath. | Ep | R, C, M, BC, T | Mx | O |
| 9 | Selaginella carnerosana T. Reeves | Ep | X, S | SM&ad | C |
| 10 | Selaginella delicatissima Linden ex A. Braun | T | C, Q | M | A, C, D, J, N, I, O |
| 11 | Selaginella eremophila Maxon | T/Ep | C, BC, X | Na | O |
| 12 | Selaginella extensa Underw. | Ep/E | R, M, BC, Q | Mx | J |
| 13 | Selaginella hoffmannii Hieron. | T/Ep | BC, Q, T | M | N, I |
| 14 | Selaginella landii Greenm. & N. Pfeiff. | T/Ep | C, BC | Mx | J, N, Z |
| 15 | Selaginella lepidophylla (Hook. & Grev.) Spring | T/Ep | Q | Na | C, D, J, I, O |
| 16 | Selaginella lineolata Mickel & Beitel | T/Ep | C, M, BC, T, S | M | J, N, I |
| 17 | Selaginella macrathera Weath. | T | C, BC, S | SM-e | O |
| 18 | Selaginella marginata (Humb. & Bonpl. ex Willd.) Spring | T | R, Q, T | Nt | N, I |
| 19 | Selaginella mutica D. C. Eaton ex Underw. | T/Ep | X | Na | C |
| 20 | Selaginella notohybrida Valdespino | T/Ep | T | Mx | C |
| 21 | Selaginella novoleonensis Hieron. | T/Ep | T, X | Mx | C, O |
| 22 | Selaginella pallescens (C. Presl) Spring | T/Ep | M, Q | Nt | A, C, D, J, N, I, O, Z |
| 23 | Selaginella parishii Underw. | T/Ep | C, X | Mx | Z |
| 24 | Selaginella peruviana (Milde) Hieron. | T/Ep | C, BC, Q, T, X | A | A, C, D, J, O, Z |
| 25 | Selaginella pilifera A. Braun | T/Ep | X, S | Na | C, D, O |
| 26 | 1Selaginella porphyrospora A. Braun | T/Ep | M, T | Nt | J, I, O |
| 27 | Selaginella reflexa Underw. | T/Ep | T | M | D |
| 28 | Selaginella rupincola Underw. | T/Ep | C, M, BC, Q, T, X | Na | A, C, D, J, O |
| 29 | Selaginella sartorii Hieron. | T/Ep | X, S | Nt | D, J, N, O |
| 30 | Selaginella schaffneri Hieron. | Ep | S | Mx | D, J, N |
| 31 | Selaginella sellowii Hieron. | T/Ep | S | Nt | D, J, I, O, Z |
| 32 | Selaginella sertata Spring | T | S | M | N |
| 33 | Selaginella tarda Mickel & Beitel | T/Ep | R, T, S | Mx | D, I |
| 34 | Selaginella tenella (P. Beauv.) Spring | Ep | R, T | Nt | I |
| 35 | Selaginella underwoodii Hieron. | Ep | X | Na | C, D, O |
| 36 | Selaginella wrightii Hieron. | Ep | T, X, S | Na | C, O, Z |
| POLYPODIIDAE | |||||
| ANEMIACEAE | |||||
| 37 | Anemia × recondita Mickel | T | BC, Q, T, S | Mx | J, N |
| 38 | Anemia affinis Hook. & Baker | T | Q, T | SM-e | D, J, N, I, O, Z |
| 39 | Anemia brandegeei Davenp. | T | T | SM-e | I |
| 40 | Anemia hirsuta (L.) Sw. | T | S | Nt | J, N, I |
| 41 | Anemia humilis (Cav.) Sw. | T | S | Nt | I |
| 42 | Anemia intermedia Copel. ex M.E. Jones | T | Q | SM-e | N |
| 43 | Anemia jaliscana Maxon | T | C, Q, T, S | Mx | D, J, N, I, O |
| 44 | Anemia karwinskyana (C. Presl) Prantl | T | T, S | Mx | J, N |
| 45 | Anemia multiplex Mickel | T | S | Mx | J, N, I, Z |
| 46 | Anemia phyllitidis (L.) Sw. | T | M | Nt | N |
| 47 | Anemia tomentosa (Sav.) Sw. | T | G | Nt | A, C, D, J, I, O, Z |
| ASPLENIACEAE | |||||
| 48 | Asplenium adiantum-nigrum L. | Ep | X | Cos | C, D |
| 49 | Asplenium arcanum A. R. Sm. | T | R, T | SM-e | D, I |
| 50 | Asplenium athyrioides Fée | T | M, T | M | I |
| 51 | Asplenium castaneum Schltdl. & Cham. | T/Ep | C | Nt | D, J |
| 52 | Asplenium cuspidatum Lam. | Ep/E | M, T | Nt | I |
| 53 | Asplenium dalhousiae Hook. | Ep | BC | Cos | C, I, O |
| 54 | Asplenium eatonii Davenp. | Ep/E | R, Q, T | Mx | I |
| 55 | Asplenium exiguum Bedd. | T/Ep | M, X | Cos | A, C, D, O |
| 56 | Asplenium fibrillosum Pringle & Davenp. ex Davenp. | T/Ep | T | Mx | A, D |
| 57 | 1Asplenium formosum Willd. | T/Ep | R, Q, T | Cos | D, J |
| 58 | Asplenium fragans Sw. | Ep/E | C, BC, Q | Nt | D, N, I |
| 59 | Asplenium gentryi A. R. Sm. | T/Ep | C, BC | Mx | A, C, D, I, O |
| 60 | Asplenium hallbergii Mickel & Beitel | T | X | Mx | A, C, D, J |
| 61 | Asplenium modestum Maxon | Ep | R, M, T | SM-e | C, D |
| 62 | 1Asplenium monanthes L. | T | M, Q | Cos | A, C, D, J, N, I, O, Z |
| 63 | Asplenium palmeri Maxon | Ep | C, X | A | A, C, D, J, I, O, Z |
| 64 | Asplenium potosinum Hieron. | T | M, T | M | N, I |
| 65 | Asplenium praemorsum Sw. | Ep/E | C | Nt | A, D, I |
| 66 | Asplenium pringlei Davenp. | Ep | M | Mx | C, D, J |
| 67 | Asplenium pumilum Sw. | Ep | R, T | Nt | J, N, I, O |
| 68 | Asplenium resiliens Kunze | T/Ep | X | A | C, O, Z |
| 69 | Asplenium sanchezii A. R. Sm. | Ep | BC, Q | Mx | C, D, I |
| 70 | Asplenium sessilifolium Desv. | T/Ep | M, Q, T | Mx | C, D, I, O |
| 71 | 1Asplenium trichomanes L. | Ep | C | Cos | C |
| 72 | Asplenium tryonii Correll | T | C | SM-e | C, D |
| ATHYRIACEAE | |||||
| 73 | Athyrium arcuatum Liebm. | T | R, C, BC, T | M | D |
| 74 | Athyrium bourgeaui E. Fourn. | T | R, C, M | M | C, D, I, O |
| 75 | Diplazium lonchophyllum Kunze | T | M | Nt | J, N |
| 76 | Ephemeropteris palmensis (Christ) R. C. Moran & Sundue | T | C, M | M | D, J, N |
| 77 | Ephemeropteris skinneri (Baker) R.C. Moran & Sundue | T | T | Nt | N, I |
| BLECHNACEAE | |||||
| 78 | Austroblechnum stoloniferum (Mett. ex E. Fourn.) Gasper & V.A.O. Dittrich | T | C, M, S | M | C, D, O |
| 79 | Blechnum appendiculatum Willd. | T | C, M, Q, S | Nt | J, N, O |
| 80 | Blechnum gracile Kaulf. | T | R, Q, T | Nt | J, N |
| 81 | Blechnum occidentale L. | T | R, M, T | A | J, N |
| 82 | Blechnum polypodioides Raddi | Ep | C, BC | Nt | N, I, D |
| 83 | Woodwardia fimbriata Sm. | T | R, M, T | Na | O |
| 84 | Woodwardia spinulosa M. Martens & Galeotti | T | M | M | C, D, J, N, I |
| CYATHEACEAE | |||||
| 85 | 1,3Cyathea costaricensis (Mett. ex Kuhn) Domin | T | M | M | J, N |
| CYSTOPTERIDACEAE | |||||
| 86 | Cystopteris diaphana (Bory) Blasdell | T | M, Q | Cos | A, D, N, I, O, Z |
| 87 | Cystopteris reevesiana Lellinger | T | M, BC | SM&ad | O |
| DENNSTAEDTIACEAE | |||||
| 88 | Dennstaedtia distenta (Kunze) T. Moore | T | C, Q, T | M | C, D, J, I, O |
| 89a | Pteridium aquilinum ssp. feei (W. Schaffn. ex Fée) Maxon | T | M, Q | M | A, D, J, N, I |
| 89b | 2Pteridium aquilinum ssp. latiusculum (Desv.) Hultén | T | C, S | Cos | C, D, I, O |
| 90 | Pteridium caudatum (L.) Maxon | T | Q, T | Nt | D, J, N, I |
| 91 | Pteridium esculentum ssp. arachnoideum (Kaulf.) J.A. Thomson | T | C, S | Nt | D |
| DRYOPTERIDACEAE | |||||
| 92 | Bolbitis portoricensis (Spreng.) Hennipman | T/Ep | R, M, Q, T | Nt | N |
| 93 | Ctenitis equestris (Kunze) Ching | T | M | M | D, N, I |
| 94 | Dryopteris cinnamomea (Cav.) C. Chr. | T/Ep | M, Q, T, X | Na | A, C, D, J, O, Z |
| 95 | Dryopteris karwinskyana (Mett.) Kuntze | T | R, M, T | M | N, I |
| 96 | Dryopteris knoblochii A. R. Sm. | T | C, Q | SM&ad | C, D, I, O |
| 97 | Dryopteris maxonii Underw. & C. Chr. | T/Ep | C | Mx | D, J, N, I |
| 98 | Dryopteris rossii C. Chr. | T | M | Mx | A, C, D, J, N, I, Z |
| 99 | Dryopteris wallichiana (Spreng.) Hyl. | T | C, M | Cos | C |
| 100 | Elaphoglossum glaucum T. Moore | T/E | C | M | N |
| 101 | Elaphoglossum gratum (Fée) T. Moore | T | M | M | D |
| 102 | Elaphoglossum hartwegii (Fée) T. Moore | T/Ep | M | Nt | D, J |
| 103 | Elaphoglossum monicae Mickel | T/Ep | BC | Mx | D, I |
| 104 | Elaphoglossum muelleri (E. Fourn.) C. Chr. | T/Ep | C, M | M | J, N, I |
| 105 | Elaphoglossum occidentale (Mickel) F.B. Matos | T | C, M | Mx | I |
| 106 | Elaphoglossum paleaceum (Hook. & Grev.) Sledge | E/Ep | BC | Cos | N |
| 107 | Elaphoglossum petiolatum (Sw.) Urb. | T | M, BC | Nt | D, J, N, I, Z |
| 108 | Elaphoglossum piloselloides (C. Presl) T. Moore | Ep | C, M, T | Nt | N, I |
| 109 | Elaphoglossum rzedowskii Mickel | Ep | M, Q | Mx | A, C, D, J, I, O, Z |
| 110 | Elaphoglossum sartorii (Liebm.) Mickel | T/E | M | Mx | D, N, I |
| 111 | Elaphoglossum tenuifolium (Liebm.) T. Moore | T/Ep | M, BC, Q | M | D |
| 112 | Phanerophlebia auriculata Underw. | T | M, T | Na | C, O |
| 113 | Phanerophlebia nobilis (Schltdl. & Cham.) C. Presl | T | M | Mx | C, J, I, O |
| 114 | Phanerophlebia umbonata Underw. | T | BC | Na | C, O |
| 115 | Polystichum hartwegii (Klotzsch) Hieron. | T | C, M, Q | Nt | N, O |
| 116 | Polystichum rachichlaena Fée | T | BC, Q | M | N, I |
| EQUISETACEAE | |||||
| 117 | Equisetum × ferrissii Clute | Hy | R, S | Na | C, D |
| 118 | Equisetum × haukeanum Mickel & A. R. Sm. | Hy | R, S | M | C, I |
| 119 | 2Equisetum hyemale L. | Hy | R, S | Cos | C, D, J, I, O |
| 120 | Equisetum laevigatum A. Braun | Hy | R, S | Na | C, D, O |
| 121 | Equisetum myriochaetum Schltdl. & Cham. | Hy | R, M, T | Nt | N, I |
| GLEICHENIACEAE | |||||
| 122 | Sticherus ferrugineus (Desv.) J. Gonzales | T | M, BC | Nt | J |
| HYMENOPHYLLACEAE | |||||
| 123 | 2Hymenophyllum tunbrigense (L.) Sm. | Ep/E | M, BC, Q | Cos | C |
| 124 | Vandenboschia radicans (Sw.) Copel | Ep/E | M, BC, Q, S | Cos | C, D |
| LYGODIACEAE | |||||
| 125 | Lygodium venustum Sw. | T | T | Nt | A, N, I |
| MARSILEACEAE | |||||
| 126 | Marsilea ancylopoda A. Braun | Hy | R | Nt | D |
| 127 | Marsilea mollis B. L. Rob. & Fernald | Hy | R, S | A | A, C, D, J, O, Z |
| 128 | Marsilea vestita Hook. & Grev. | Hy | R, S | A | D, J, I, O |
| 129 | Pilularia americana A. Braun | Hy | R, S | A | D |
| NEPHROLEPIDACEAE | |||||
| 130 | 2Nephrolepis undulata (Afzel. ex Sw.) J. Sm. | Ep | M, Q | Cos | D, J, N, I |
| OPHIOGLOSSACEAE | |||||
| 131 | Botrychium tolucaense W. H. Wagner & Mickel | T | C | Mx | D |
| 132 | Botrypus virginianum (L.) Michx. | T | Q | Cos | C, D, J, N, I, O |
| 133 | Ophioglossum crotalophoroides Walter | T | BC, Q, S | Nt | D, N, I |
| 134 | Ophioglossum engelmannii Prantl | T | Q, T | A | A, C, D, J, N, I, O |
| 135 | 2Ophioglossum nudicaule L.f. | T | T, S | Cos | A, C, D, J, N, O, Z |
| 136 | Ophioglossum polyphyllum A. Braun | T | C, BC, Q | Cos | C, O, Z |
| 137 | 2Ophioglossum reticulatum L. | T | C, Q, T | Cos | J, N, I |
| 138 | Sceptridium schaffneri (Underw.) Lyon | T | BC, Q, G, S | Nt | A, C, D, O |
| OSMUNDACEAE | |||||
| 139 | 2Osmunda regalis L. | Hy | R, C, M | Cos | A, J, N |
| PLAGIOGYRACEAE | |||||
| 140 | Plagiogyria pectinata (Liebm.) Lellinger | T | R, M | Nt | C, D, J, O |
| POLYPODIACEAE | |||||
| 141 | Campyloneurum angustifolium (Sw.) Fée | Ep/E | M, BC | Nt | D, J, I |
| 142 | Campyloneurum ensifolium (Willd.) J. Sm. | E | T | M | C, D, O |
| 143 | 1Campyloneurum phyllitidis (L.) C. Presl | Ep/E | R, M, BC, T | Nt | J, N |
| 144 | Campyloneurum xalapense Fée | Ep/E | R, M, BC, T | M | N |
| 145 | Melpomene moniliformis (Lag. Ex Sw.) A.R. Sm. | Ep/E | R, M | Nt | N |
| 146 | Pecluma alfredii (Rosenst.) M. G. Price | Ep/E | C, T | M | D, J, N, I |
| 147 | Pecluma ferruginea (M. Martens & Galeotti) M. G. Price | Ep | C, M, T | M | D, J, N, I |
| 148 | Pecluma hartwegiana (Hook.) F. C. Assis & Salino | Ep/E | M | M | C, D, N |
| 149 | Phlebodium pseudoaureum (Cav.) Lellinger | Ep/E | M, S | Nt | A, C, D, J, I, O, Z |
| 150 | Pleopeltis acicularis (Weath.) A. R. Sm. & T. Krömer | Ep/E | C, M, T, X | M | D, J, N, I, O, Z |
| 151 | Pleopeltis angusta Humb. & Bonpl. Ex Willd. | E | M, T, S | M | D, N |
| 152 | Pleopeltis astrolepis (Liebm.) E. Fourn. | Ep/E | C, BC | Nt | N |
| 153 | Pleopeltis furfuracea (Schltdl. & Cham.) A.R. Sm. & Tejero | E | T | Nt | D, J, N |
| 154 | Pleopeltis guttata (Maxon) E.G. Andrews & Windham | Ep | C, BC | Mx | A, C, D, Z |
| 155 | Pleopeltis madrensis (J. Sm.) A.R. Sm. & Tejero | Ep/E | C, M, T | Mx | A, C, D, J, N, I |
| 156 | Pleopeltis mexicana (Fée) Mickel & Beitel | Ep/E | M | M | D, J, N, I |
| 157 | Pleopeltis polylepis (Roemer ex Kunze) T. Moore | Ep/E | M, Q, X | Mx | A, C, D, J, I, Z |
| 158 | Pleopeltis polypodioides (L.) E.G. Andrews & Windham | Ep | C, BC, Q | A | C, J, N, I, O |
| 159 | Pleopeltis riograndensis (T. Wendt) E.G. Andrews & Windham | Ep | C | SM&ad | C, O |
| 160 | Pleopeltis rosei (Maxon) A.R. Sm. & Tejero | Ep/E | M, Q, T | Mx | D, J, N, I, Z |
| 161 | Pleopeltis rzedowskiana (Mickel) A.R. Sm. & Tejero | Ep/E | T | Mx | I |
| 162 | Pleopeltis sanctae-rosae (Maxon) A.R. Sm. & Tejero | E | C, T | M | N |
| 163 | Pleopeltis thyssanolepis E.G. Andrews & Windham | Ep/E | Q, T, X | Nt | A, C, D, J, I, O, Z |
| 164 | Polypodium arcanum Maxon | E | M, BC | Mx | D |
| 165 | Polypodium colpodes Kunze | Ep/E | T | Nt | N, I |
| 166 | Polypodium fraternum Schltdl. & Cham. | E | M, T | M | N |
| 167 | Polypodium hesperium Maxon | Ep | C | Na | C |
| 168 | Polypodium martensii Mett. | E | S | Mx | C, D, J |
| 169 | Polypodium plesiosorum Kunze | Ep/E | M, S | M | J, N, Z |
| 170 | Polypodium praetermissum Mickel & Tejero | Ep | R, M, BC, Q, T | SM-e | D, N, I, O |
| 171 | Polypodium subpetiolatum Hook. | Ep/E | T | M | A, C, D, N, I |
| PSILOTACEAE | |||||
| 172 | 1Psilotum × complanatum Sw. | Ep/E | Q, T | Cos | C, J, N, O |
| 173 | 2Psilotum nudum (L.) P. Beauv. | Ep/E | BC, Q, T | Cos | C, J, O, Z |
| PTERIDACEAE | |||||
| 174 | Adiantopsis seemannii (Hook.) Maxon | Ep | R, M, T | M | I |
| 175 | Adiantum aleuticum (Rupr.) C. A. Paris | Ep | C, S | Na | C |
| 176 | Adiantum amplum C. Presl. | T | R, T | Nt | N, I |
| 177 | Adiantum andicola Liebm. | T | C, M | Nt | C, D, J, I, O |
| 178 | Adiantum braunii Mett. Ex Kuhn | T | M | Nt | D, J, N, I, O |
| 179 | 2Adiantum capillus-veneris L. | Ep | X, S | Cos | A, C, D, J, O, Z |
| 180 | Adiantum concinnum Humb. & Bonpl. Ex Willd. | T/Ep | S | Nt | D, J, N, I, O |
| 181 | Adiantum lunulatum Burm. f. | T/Hy | R, T | Cos | I |
| 182 | Adiantum patens Willd. | T | T, S | Nt | D, J, N, I, O |
| 183 | Adiantum poiretii Wikstr. | T | S | Cos | A, C, D, J, N, I, O |
| 184 | Adiantum raddianum C. Presl | T | M | Cos | I |
| 185 | Adiantum trapeziforme L. | T | R, Q, T | Nt | N |
| 186 | Adiantum tricholepis Fée | T | R, T | M | C, D, J, N, I, O |
| 187 | Aleuritopteris farinosa (Forssk.) Fée | T/Ep | Q, T | Cos | A, C, D, J, I, Z |
| 188 | Ananthacorus angustifolius (Sw.) Underw. & Maxon | Ep/E | T, S | Nt | J, N |
| 189 | 2Anogramma leptophylla (L.) Link | T | S | Cos | C, D, J, N, I, O |
| 190 | Argyrochosma delicatula (Maxon & Weath.) Windham | Ep | Q, X | Mx | C, D |
| 191 | Argyrochosma fendleri (Kunze) Windham | Ep | Q | Na | O |
| 192 | Argyrochosma formosa (Liebm.) Windham | Ep | C, T | M | A |
| 193 | Argyrochosma incana (C. Presl) Windham | T/Ep | C, T | Na | A, C, D, J, N, I, O, Z |
| 194 | Argyrochosma jonesii (Maxon) Windham | Ep | R, Q, X | Na | O |
| 195 | Argyrochosma limitanea (Maxon) Windham | Ep | C, Q | Na | C, D, J, I, O |
| 196 | Argyrochosma lumholtzii (Maxon & Weath.) Windham | Ep | R, S | SM-e | O |
| 197 | Argyrochosma microphylla (Mett. ex Kuhn) Windham | T/Ep | C, BC, Q, X | Na | C, O, Z |
| 198 | Argyrochosma pallens (Weath. ex R. M. Tryon) Windham | Ep | C, M, BC, Q | Mx | A, C, D, J |
| 199 | Argyrochosma palmeri (Baker) Windham | Ep | Q | Mx | D, J |
| 200 | Argyrochosma pilifera (R.M. Tryon) Windham | Ep | C, X | Mx | C |
| 201 | Aspidotis meifolia (D. C. Eaton) Pic. Serm. | Ep | Q | Mx | C, D |
| 202 | Astrolepis cochisensis (Goodd.) D. M. Benham & Windham | Ep | C, X | Na | A, C, D, O, Z |
| 203 | Astrolepis crassifolia (T. Moore & Houlston) D. M. Benham & Windham | T/Ep | C, BC, Q, X | M | C, D |
| 204 | Astrolepis integerrima (Hook.) D. M. Benham & Windham | Ep | C | Na | A, C, D, O, Z |
| 205 | Astrolepis laevis (M. Martens & Galeotti) Mickel | Ep | X | M | A, C, D, J, Z |
| 206 | Astrolepis sinuata (Lag. ex Sw.) D. M. Benham & Windham | T/Ep | C, X | A | A, C, D, J, N, I, O, Z |
| 207 | Astrolepis windhamii D.M. Benham | Ep | C, X | SM&ad | O |
| 208 | Bommeria ehrenbergiana (Klotzsch) Underw. | T/Ep | C, BC, Q, T | Mx | D |
| 209 | Bommeria elegans (Davenp.) Ranker & Haufler | T | C, M, BC, Q | Mx | J, N, Z |
| 210 | Bommeria hispida (Mett. ex Kuhn) Underw. | T/Ep | BC, Q, T | Na | A, C, D, J, O, Z |
| 211 | Bommeria pedata (Sw.) E. Fourn. | T/Ep | M, Q | M | C, D, J, N, I, O, Z |
| 212 | Bommeria subpalacea Maxon | T/Ep | Q, T, X | Mx | C |
| 213 | Cheilanthes leucopoda Link | Ep | C, BC, Q, X | Na | C, D, O |
| 214 | Cheilanthes lozanoi (Maxon) R. M. Tryon & A. F. Tryon | T/Ep | BC | Mx | C, D, J, N, I, O, Z |
| 215 | Cheilanthes skinneri (Hook.) R. M. Tryon & A. F. Tryon | T | C, BC | Nt | C, D, J, N, O |
| 216 | Cheiloplecton rigidum (Sw.) Fée | Ep | T | M | C, J, N, O, Z |
| 217 | Gaga angustifolia (Kunth) Fay-Wei Li & Windham | T | T, X | M | A, C, D, J, N, I, O |
| 218 | Gaga arizonica (Maxon) Fay-Wei Li & Windham | T | T, X, S | Na | C, D, I, O, Z |
| 219 | Gaga chaerophylla (M. Martens & Galeotti) Fay-Wei Li & Windham | T | BC | Nt | D, J, I, O |
| 220 | Gaga cuneata (Kaulf. ex Link) Fay-Wei Li & Windham | Ep | C, M, Q | Mx | D, J |
| 221 | Gaga decomposita (M. Martens & Galeotti) Fay-Wei Li & Windham | T/Hy | S | Mx | D, J, N, I, Z |
| 222 | Gaga hirsuta (Link) Fay-Wei Li & Windham | T/Ep | C, T | M | A, C, D, J, N, I, O, Z |
| 223 | Gaga kaulfussii (Kunze) Fay-Wei Li & Windham | T/Ep | C, M, Q | A | A, C, D, J, N, I, O, Z |
| 224 | Gaga lerstenii (Mickel & Beitel) Fay-Wei Li & Windham | T | C, BC | Mx | J |
| 225 | Gaga marginata (Kunth) Fay-Wei Li & Windham | Ep | M | Nt | A, N |
| 226 | Gaga pellaeopsis (Mickel) Fay-Wei Li & Windham | T/Ep | C, BC | Mx | J |
| 227 | Gastoniella novogaliciana (Mickel) Li Bing Zhang & Liang Zhang | Ep | R, BC, T | Mx | J |
| 228 | xHemionanthes gryphus (Mickel) Mickel | T/Ep | R, T | Mx | N |
| 229 | Hemionitis palmata L. | T | R, T | Nt | I |
| 230 | Hemionitis subcordata (D.C. Eaton ex Davenp.) Mickel | T/Ep | R, Q, T | M | D, J, N, I |
| 231 | Llavea cordifolia Lag. | T/Ep | C, BC, Q, T | M | D |
| 232 | Mildella fallax (M. Martens & Galeotti) Nesom | Ep | C, T | M | C, J, I, O |
| 233 | Mildella intramarginalis (Kaulf. ex Link) Trevis. | T/Ep | C, T | M | I |
| 234 | Myriopteris aemula (Maxon) Grusz & Windham | Ep | T, X | Na | C |
| 235 | Myriopteris alabamensis (Buckley) Grusz & Windham | Ep | Q, X | Na | C, O |
| 236 | Myriopteris allosuroides (Mett.) Grusz & Windham | Ep | X | Mx | A, C, D, J, N, I, O, Z |
| 237 | Myriopteris aurea (Poir.) Grusz & Windham | Ep | M, Q | Nt | A, C, D, J, N, I, O, Z |
| 238 | Myriopteris cucullans (Fée) Grusz & Windham | Ep | C, M | M | A, C, D, J, O |
| 239 | Myriopteris fendleri (Hook.) E. Fourn. | Ep | C, BC | Na | O |
| 240 | Myriopteris gracilis Fée | T | X, G | Na | C, O |
| 241 | Myriopteris lendigera (Cav.) Fée | Ep | M, Q | A | C, D, J, I, O, Z |
| 242 | Myriopteris lindheimeri (Hook.) J. Sm. | T | C, BC, Q | Na | A, C, D, J, O, Z |
| 243 | Myriopteris longipila (Baker) Grusz & Windham | Ep | Q, T | Mx | J |
| 244 | Myriopteris mexicana (Davenp.) Grusz & Windham | Ep | C, BC, Q, X, S | M | C, D, Z |
| 245 | Myriopteris microphylla (Sw.) Grusz & Windham | Ep | T, X | Nt | C, D, O, Z |
| 246 | Myriopteris myriophylla (Desv.) Grusz & Windham | Ep | C, Q, S | Nt | A, C, D, J, I, O, Z |
| 247 | Myriopteris notholaenoides (Desv.) Grusz & Windham | Ep | C, Q, T, X, G | Nt | C, D, Z |
| 248 | Myriopteris pringlei (Davenp.) Grusz & Windham | Ep | C, BC, Q, X | Na | C, Z |
| 249 | Myriopteris rufa Fée | Ep | C, X | M | A, C, D, O, Z |
| 250 | Myriopteris scabra (C. Chr.) Grusz & Windham | T | C, BC, Q | Mx | C, D, Z |
| 251 | Myriopteris tomentosa (Link) Fée | Ep | C, BC, Q, X | Na | C, O |
| 252 | Myriopteris windhamii Grusz | Ep | X | Mx | C, D, J, O, Z |
| 253 | Myriopteris wootonii (Maxon) Grusz & Windham | Ep | C, BC, Q, G | Na | C, O |
| 254 | Myriopteris wrightii (Hook.) Grusz & Windham | Ep | G | Na | C, D, O |
| 255 | Myriopteris yatskievychiana (Mickel) Grusz & Windham | Ep | R | SM-e | O |
| 256 | Notholaena aliena Maxon | Ep | X | Na | C |
| 257 | Notholaena aschenborniana Klotzsch | Ep | X, G | Na | A, C, D, O, Z |
| 258 | Notholaena aurantiaca D.C. Eaton | Ep | S | SM-e | J, N |
| 259 | Notholaena brachypus (Kunze) Kunze | Ep | X | M | A, D, J, N, O, Z |
| 260 | Notholaena californica D.C. Eaton | Ep | X | Na | C, O |
| 261 | Notholaena candida (M. Martens & Galeotti) Hook. | T/Ep | T, S | M | A, C, D, J, N, I, O, Z |
| 262 | Notholaena galeottii Fée | Ep | C, S | M | D |
| 263 | Notholaena grayi Davenp. | Ep | R, BC, Q, T | Na | C, J, I, O, Z |
| 264 | Notholaena greggii (Mett. ex Kuhn) Maxon | T/Ep | BC, Q, G | Na | C, D |
| 265 | Notholaena jaliscana Yatsk. & Arbeláez | T/Ep | C | SM-e | J, N |
| 266 | Notholaena lemmonii D. C. Eaton | Ep | C, Q, X | Na | C, D, J, I, O |
| 267 | Notholaena neglecta Maxon | Ep | BC, Q, X | Na | C |
| 268 | Notholaena ochracea (Hooker) Yatsk. & Arbeláez | Ep | C, BC, Q, T | Mx | D, J, N |
| 269 | Notholaena schaffneri (E. Fourn.) Underw. ex Davenp. | Ep | BC, Q | M | D, J, Z |
| 270 | Notholaena standleyi Maxon | Ep | BC, Q | Na | C, D, I, O |
| 271 | Pellaea atropurpurea (L.) Link | T/Ep | C, Q, T, X | Na | C, D, O, Z |
| 272 | Pellaea cordifolia (Sessé & Moc.) A. R. Sm. | T/Ep | T, X | Na | A, C, D, J, Z |
| 273 | Pellaea intermedia Mett. ex Kuhn | Ep | C, Q, X | Na | C, D, O, Z |
| 274 | Pellaea mucronata (D. C. Eaton) D. C. Eaton | Ep | Q, X | Na | O |
| 275 | Pellaea oaxacana Mickel & Beitel | T/Ep | BC, T | Mx | J |
| 276 | Pellaea ovata (Desv.) Weath. | T/Ep | C, X | A | A, C, D, J, I, O, Z |
| 277 | Pellaea pringlei Davenp. | Ep | BC, Q, T, S | Mx | J, N, I |
| 278 | Pellaea sagittata (Cav.) Link | T/Ep | X | A | C, D, J, O, Z |
| 279 | Pellaea ternifolia (Cav.) Link | Ep | BC, Q, T, X | A | A, C, D, J, N, I, O, Z |
| 280 | Pellaea truncata Goodd. | Ep | M, BC, Q | Na | O |
| 281 | Pellaea villosa (Windham) Windham & Yatsk. | Ep | BC, Q, X | Mx | C, D, O |
| 282 | Pellaea wrightiana Hook. | Ep | C, BC, Q, X, S | Na | C, O |
| 283 | Pityrogramma calomelanos (L.) Link | T/Ep | R, M, T, S | Nt | D, N, I, O |
| 284 | Pityrogramma dealbata (C. Presl) Domin | T | R, BC, T | M | J, N |
| 285 | Pityrogramma tartarea (Cav.) Maxon | T | M, S | Nt | D, J, N, I |
| 286 | Pteris biaurita L. | T | R, T | Cos | N |
| 287 | 2Pteris cretica L. | T | C, M, Q | Cos | C, D, J, I, O |
| 288 | Pteris erosa Mickel & Beitel | T | M, T | M | J, N |
| 289 | Pteris orizabae M. Martens & Galeotti | T/Hy | S | M | D |
| 290 | Pteris podophylla Sw. | T/Hy | R, M | Nt | J |
| 291 | Pteris quadriaurita Retz. | T | S | Cos | J, N |
| SALVINIACEAE | |||||
| 292 | Azolla filiculoides Lam. | Hy | R, S | Cos | C |
| 293 | 2Azolla cristata Kaulf. | Hy | R, S | A | A, J, N, I |
| TECTARIACEAE | |||||
| 294 | Tectaria heracleifolia (Willd.) Underw. | T | R, Q, T, S | A | J, I |
| 295 | Tectaria mexicana (Fée) C.V. Morton | T | R, Q, T | Nt | N, I |
| THELYPTERIDACEAE | |||||
| 296 | Amauropelta cheilanthoides (Kunze) Á. Löve & D. Löve | T | C, M, S | Nt | J |
| 297 | Amauropelta oligocarpa (Humb. & Bonpl. ex Willd.) Pic. Serm. | T | M, Q, T | Nt | N |
| 298 | Amauropelta resinifera (Desv.) Pic. Serm. | T | S | Mx | C, J, N, I |
| 299 | Amauropelta rudis (Kunze) Pic. Serm. | T | S | Nt | C, D, J, N, I |
| 300 | Christella hispidula (Decne.) Holttum | T | T, S | Cos | J, N, I, O |
| 301 | 2Cyclosorus interruptus (Willd.) H. Itô | Hy | R, T, S | Cos | J, N |
| 302 | Goniopteris imbricata (Liebm.) A. & D. Löve | T/Hy | R, T | M | N |
| 303 | Leptogramma pilosa (M. Martens & Galeotti) Underw. | Ep | Q | M | C, D, J, N, O |
| 304 | Pelazoneuron ovatum (R.P. St. John) A.R.Sm. & S.E.Fawc. | Hy | R, S | M | O |
| 305 | Pelazoneuron puberulum (Baker) A.R.Sm. & S.E.Fawc. | T | C, M, S | M | A, C, D, J, N, I, O, Z |
| WOODSIACEAE | |||||
| 306 | Woodsia cochisensis Windham | Ep | C, BC, X, G | SM&ad | C, D, I, O |
| 307 | Woodsia cystopteroides Windham & Mickel | Ep | C, BC | SM-e | D, I, O |
| 308 | Woodsia mexicana Fée | T/Ep | C, X, S | Mx | C, Z |
| 309 | Woodsia mollis (Kaulf.) J. Sm. | T/Ep | X | M | A, C, D, J, N, I, O, Z |
| 310 | Woodsia neomexicana Windham | Ep | C, BC | Na | Z |
| 311 | Woodsia phillipsii Windham | T/Ep | BC | SM&ad | C, D, O, Z |
| 312 | Woodsia plummerae Lemmon | Ep | BC | SM&ad | C, O |
References
- Kessler, M. Biogeography of ferns. In Fern Ecology; Mehltreter, K., Walker, L.R., Sharpe, J.M., Eds.; Cambridge University Press: New York, NY, USA, 2010; pp. 22–60. [Google Scholar]
- Almeida, T.E.; Salino, A. State of the art and perspectives on Neotropical fern and lycophyte systematics. J. Syst. Evol. 2016, 54, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Suissa, J.S.; Sundue, M.A.; Testo, W.L. Mountains, climate and niche heterogeneity explain global patterns of fern diversity. J. Biogeogr. 2021, 48, 1296–1308. [Google Scholar] [CrossRef]
- Tejero-Díez, J.D.; Torres-Díaz, A.N.; Sánchez-González, A. Helechos de la Sierra Madre del Sur. In Biodiversidad de la Sierra Madre del Sur: Una Síntesis Preliminar; Luna-Vega, I., Espinosa, D., Contreras-Medina, R., Eds.; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2016; pp. 121–155. [Google Scholar]
- Contreras-Medina, R.; García-Martínez, A.I.; Ramírez-Martínez, J.C.; Espinosa, D.; Balam-Narváez, R.; Luna-Vega, I. Biogeographic analysis of ferns and lycophytes in Oaxaca: A Mexican beta-diverse area. Bot. Sci. 2022, 100, 204–222. [Google Scholar] [CrossRef]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Alvarado, A.F.; Tejero-Díez, J.D. Novelties and notes in Lindsaea (Lindsaeaceae) from Mexico and Central America. Phytotaxa 2017, 296, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Sundue, M.A. Three new species of grammitid ferns (Polypodiaceae) from the fern hunter’s paradise: Sierra Juárez, Oaxaca, Mexico. Syst. Bot. 2017, 42, 160–168. [Google Scholar] [CrossRef]
- Mickel, J.T.; Smith, A.R. The pteridophytes of Mexico. Mem. NY Bot. Gard. 2004, 88, 1–1054. [Google Scholar]
- Mickel, J.T.; Beitel, J.M. Pteridophyte Flora of Oaxaca, Mexico; The New York Botanical Garden: New York, NY, USA, 1988. [Google Scholar]
- Mickel, J.T. Pteridophytes. In Flora Novo-Galiciana: A Descriptive Account of the Vascular Plants of Western Mexico; McVaugh, R., Ed.; The University of Michigan Herbarium: Ann Arbor, MI, USA, 1992; pp. 120–431. [Google Scholar]
- Krömer, T.; Carvajal-Hernández, C.I.; Acebey, A.R.; Smith, A.R. A decade of new pteridophyte records for the state of Veracruz, Mexico. Am. Fern J. 2015, 105, 162–175. [Google Scholar] [CrossRef]
- Carvajal-Hernández, C.I.; Silva-Mijangos, L.; Kessler, M.; Lehnert, M. Adiciones a la pteridoflora de Tabasco, México: La importancia del bosque mesófilo de montaña. Acta Bot. Mex. 2018, 124, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, L.; Escareño-Mendoza, I.S.; Guzmán-Cornejo, L.; Márquez-Bautista, S.; Camargo, S.L.; Sánchez-Morales, A. New fern and lycophyte records from the states of Oaxaca, Zacatecas, Puebla, and San Luis Potosí, Mexico. Am. Fern J. 2021, 111, 46–50. [Google Scholar] [CrossRef]
- Sanginés-Franco, C.; Luna-Vega, I.; Alcántara-Ayala, O.; Contreras-Medina, R. Distributional patterns and biogeographic analysis of ferns in the Sierra Madre Oriental, Mexico. Am. Fern J. 2011, 101, 81–104. [Google Scholar] [CrossRef]
- Salinas-Rodríguez, M.M.; Hernández-Sandoval, L.; Carrillo-Reyes, P.; Castillo-Gómez, H.A.; Castro-Castro, A.; Estrada-Castillón, E.; Figueroa-Martínez, D.S.; Gómez-Escamilla, I.N.; González-Elizondo, S.; Gutiérrez-Ortega, J.S.; et al. Diversidad de plantas vasculares de la provincia fisiográfica de la Sierra Madre Oriental, México. Bot. Sci. 2022, 100, 469–492. [Google Scholar] [CrossRef]
- Hernández-Cárdenas, R.A.; Mendoza-Ruiz, A.; Arredondo-Amezcua, L.; Steinmann, V.W. The alpine ferns of the Trans-Mexican Volcanic Belt. Am. Fern J. 2019, 109, 11–25. [Google Scholar] [CrossRef]
- Smith, A.R. Pteridophytes. In Flora of Chiapas; Breedlove, D.E., Ed.; Academic of Sciences: San Francisco, CA, USA, 1981; pp. 1–370. [Google Scholar]
- Wiggins, I.L. Flora of Baja California; Stanford University Press: Stanford, CA, USA, 1980. [Google Scholar]
- León de la Luz, J.L.; Rebman, J.; Domínguez-León, M.; Domínguez-Cadena, R. The vascular flora and floristic relationships of the Sierra de La Giganta in Baja California Sur, Mexico. Rev. Mex. Biodivers. 2008, 79, 29–65. [Google Scholar]
- Tryon, R.M. Endemic areas and geographic speciation in tropical American ferns. Biotropica 1972, 4, 121–131. [Google Scholar] [CrossRef]
- Tryon, R.M. The biogeography of species, with special reference to ferns. Bot. Rev. 1986, 52, 117–156. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetación de México; Limusa: Mexico City, Mexico, 1978. [Google Scholar]
- Rzedowski, J. Diversidad y orígenes de la flora fanerogámica de México. Acta Bot. Mex. 1991, 14, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Rahbek, C.; Borregaard, M.K.; Antonelli, A.; Colwell, R.K.; Holt, B.G.; Nogués-Bravo, D.; Rasmussen, C.M.Ø.; Richardson, K.; Rosing, M.T.; Whittaker, R.J.; et al. Building mountain biodiversity: Geological and evolutionary processes. Science 2019, 365, 1114–1119. [Google Scholar] [CrossRef]
- Testo, W.L.; Sessa, E.; Barrington, D.S. The rise of the Andes promoted rapid diversification in Neotropical Phlegmariurus (Lycopodiaceae). New Phytol. 2019, 222, 604–613. [Google Scholar] [CrossRef]
- Perrigo, A.; Hoorn, C.; Antonelli, A. Why mountains matter for biodiversity. J. Biogeogr. 2020, 47, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Sanginés-Franco, C.; Luna-Vega, I.; Contreras-Medina, R.; Espinosa, D.; Tejero-Díez, J.D.; Rivas, G. Diversity, endemism and conservation of ferns (Polypodiales) in the Mexican Mountain Component. J. Mt. Sci. 2015, 12, 891–904. [Google Scholar] [CrossRef]
- Ferrari, L.; Valencia-Moreno, M.; Bryan, S. Magmatism and tectonics of the Sierra Madre Occidental and its relation with the evolution of the western margin of North America. Geol. Soc. Amer. Spec. Pap. 2007, 422, 1–39. [Google Scholar] [CrossRef] [Green Version]
- González-Elizondo, S.; González-Elizondo, M.; Tena-Flores, J.A.; Ruacho-González, L.; López-Enríquez, I.L. Vegetación de la Sierra Madre Occidental, México: Una síntesis. Acta Bot. Mex. 2012, 100, 351–403. [Google Scholar] [CrossRef] [Green Version]
- Kobelkowsky-Vidrio, T.; Ríos-Muñoz, C.A.; Navarro-Sigüenza, A.G. Biodiversity and biogeography of the avifauna of the Sierra Madre Occidental, Mexico. Biodivers. Conserv. 2014, 23, 2087–2105. [Google Scholar] [CrossRef]
- Morrone, J.J. Neotropical Biogeography: Regionalization and Evolution; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). Provincias Biogeográficas de México; Scale 1:4000000; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 1997. [Google Scholar]
- Ferrusquía-Villafranca, I. Geology of Mexico: A Synopsis. In Biological Diversity of Mexico: Origins and Distribution; Ramammoorthy, T.P., Bye, R.A., Lot, A., Fa, J., Eds.; Oxford University Press: New York, NY, USA, 1993; pp. 3–107. [Google Scholar]
- Cevallos-Ferriz, S.R.S.; González-Torres, E.A.; Calvillo-Canadell, L. Perspectiva paleobotánica y geológica de la biodiversidad en México. Acta Bot. Mex. 2012, 100, 317–350. [Google Scholar] [CrossRef] [Green Version]
- Douglas, M.W.; Maddox, R.A.; Howard, K.; Reyes, S. The Mexican Monsoon. J. Clim. 1993, 6, 1665–1677. [Google Scholar] [CrossRef]
- García, E. Modificaciones al Sistema de Clasificación climática de Köppen; Instituto de Geografía, Universidad Nacional Autónoma de México: Mexico City, Mexico, 2004. [Google Scholar]
- Instituto Nacional de Estadística y Geografía (INEGI). Conjunto de Datos Vectoriales de Uso de Suelo y Vegetación; Scale 1:250000; Serie VII; Instituto Nacional de Estadística y Geografía: Aguascalientes, Mexico, 2021. [Google Scholar]
- Silva-Flores, R.; Pérez-Verdín, G.; Wehenkel, C. Patterns of tree species diversity in relation to climatic factors on the Sierra Madre Occidental, Mexico. PLoS ONE 2014, 9, e105034. [Google Scholar] [CrossRef]
- Felger, R.; Wilson, M.F. Northern Sierra Madre Occidental and its Apachian outliers: A neglected center of biodiversity. In Biodiversity and Management of the Madrean Archipelago: The Sky Islands of Southwestern United States and Northwestern Mexico; DeBano, L.F., Ffolliott, P.F., Ortega-Rubio, A., Gottfried, G.J., Hamre, R.H., Edminster, C.B., Eds.; Department of Agriculture and Forest Service: Tucson, AZ, USA, 1995; pp. 36–59. [Google Scholar]
- Macías-Rodríguez, M.A.; Giménez de Azcárate-Cornide, J.; Gopar-Merino, L.F. Sistematización bioclimática de la Sierra Madre Occidental (México) y su relación con los pisos de vegetación. Polibotánica 2017, 43, 125–163. [Google Scholar] [CrossRef]
- Knobloch, I.W.; Correll, D.S. (Eds.) Ferns and Fern Allies of Chihuahua, Mexico; Contribution from Texas Research Foundation: Forth Worth, TX, USA, 1962. [Google Scholar]
- González-Elizondo, M.; González-Elizondo, S.; Herrera-Arrieta, Y. Listados florísticos de México: Flora de Durango; Instituto de Biología, Universidad Nacional Autónoma de México: Mexico City, Mexico, 1991. [Google Scholar]
- Spellenberg, R.; Lebgue, T.; Corral-Díaz, R. A Specimen Based, Annotated Checklist of the Vascular Plants of Parque Nacional Cascada de Basaseachi and Adjacent Areas, Chihuahua, México; Instituto de Biología, Universidad Nacional Autónoma de México: Mexico City, Mexico, 1996. [Google Scholar]
- Siqueiros-Delgado, M.E.; González-Adame, G. Checklist of the pteridophytes of Aguascalientes, Mexico. Aliso 2002, 21, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Castillón, A.E.; Jurado, E.; Návar, J.J.; Jiménez-Pérez, J.; Garza-Ocañas, F. Plant associations of Cumbres de Majalca National Park, Chihuahua, Mexico. Southwest. Nat. 2003, 48, 177–187. [Google Scholar] [CrossRef]
- Vázquez-García, J.A.; Cházaro, M.J.; Nieves, G.; Vargas-Rodríguez, Y.L.; Vázquez, M.; Flores, M.A. Flora del norte de Jalisco y etnobotánica huichola; Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara: Guadalajara, Mexico, 2004. [Google Scholar]
- Yatskievych, G.; Van Devender, T.R.; Reina-Guerrero, A.L. Pteridofitas. In Diversidad Biológica de Sonora; Molina-Freaner, F.E., Van Devender, T.R., Eds.; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2010; pp. 229–261. [Google Scholar]
- McDonald, J.A.; Martínez, J.; Nesom, G.L. Alpine flora of Cerro Mohinora, Chihuahua, Mexico. J. Bot. Res. Inst. Texas 2011, 5, 701–705. [Google Scholar]
- Ruacho-González, L.; González-Elizondo, S.; González-Elizondo, M.; López-González, C. Diversidad florística en cimas de la Sierra Madre Occidental, México, y su relación con variables ambientales. Bot. Sci. 2013, 91, 193–205. [Google Scholar] [CrossRef]
- Phylogeny Pteridophyte Group (PPG I). A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016, 54, 563–603. [Google Scholar] [CrossRef]
- Salino, A.; Almeida, T.E.; Smith, A.R. New combinations in Neotropical Thelypteridaceae. PhytoKeys 2015, 57, 11–50. [Google Scholar] [CrossRef] [Green Version]
- Gasper, A.L.; Almeida, T.E.; Dittrich, V.A.O.; Smith, A.R.; Salino, A. Molecular phylogeny of the fern family Blechnaceae (Polypodiales) with a revised genus-level treatment. Cladistics 2017, 33, 429–446. [Google Scholar] [CrossRef]
- Hauk, W.D.; Parks, C.R.; Chase, M.W. Phylogenetic studies of Ophioglossaceae: Evidence from rbcL and trnL-F plastid D.N.A. sequences and morphology. Mol. Phylogenet. Evol. 2003, 28, 131–151. [Google Scholar] [CrossRef]
- Fawcett, S.; Smith, A.R. A Generic Classification of the Thelypteridaceae; Botanical Research Institute of Texas Press: Forth Worth, TX, USA, 2021. [Google Scholar]
- Moran, R.C.; Garrison, J.; Sundue, M. Phylogenetic relationships of Neotropical lady ferns (Athyriaceae), with a description of Ephemeropteris, gen. nov. Taxon 2019, 68, 425–441. [Google Scholar] [CrossRef]
- Li, F.W.; Pryer, K.M.; Windham, M.D. Gaga, a new fern genus segregated from Cheilanthes (Pteridaceae). Syst. Bot. 2012, 37, 854–860. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, X.M.; Lu, N.T.; Zhang, L.B. Phylogeny of the fern subfamily Pteridoideae (Pteridaceae), with the description of a new genus: Gastoniella. Mol. Phylogenet. Evol. 2017, 109, 59–72. [Google Scholar] [CrossRef]
- Testo, W.; Øllgaard, B.; Field, A.; Almeida, T.; Kessler, M.; Barrington, D.S. Phylogenetic systematics, morphological evolution, and natural groups in Neotropical Phlegmariurus (Lycopodiaceae). Mol. Phylogenet. Evol. 2018, 125, 1–13. [Google Scholar] [CrossRef]
- Smith, A.R.; Tejero-Díez, J.D. Pleopeltis (Polypodiaceae), a redefinition of the genus and nomenclatural novelties. Bot. Sci. 2014, 92, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Grusz, A.L.; Windham, M.D. Toward a monophyletic Cheilanthes: The resurrection and recircumscription of Myriopteris (Pteridaceae). PhytoKeys 2013, 32, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, A.; Dubuisson, J.Y.; Iwatsuki, K.; Hennequin, S.; Ito, M. A taxonomic revision of Hymenophyllaceae. Blumea 2006, 51, 221–280. [Google Scholar] [CrossRef] [Green Version]
- Salazar, L.; Homeier, J.; Kessler, M.; Abrahamczyk, S.; Lehnert, M.; Krömer, T.; Kluge, J. Diversity patterns of ferns along elevational gradients in Andean tropical forests. Plant Ecol. Divers. 2013, 8, 13–24. [Google Scholar] [CrossRef]
- Carvajal-Hernández, C.I.; Krömer, T. Riqueza y distribución de helechos y licófitos en el gradiente altitudinal del Cofre de Perote, centro de Veracruz, México. Bot. Sci. 2015, 93, 601–614. [Google Scholar] [CrossRef] [Green Version]
- Murillo, J.; Murillo, M.T. Diversidad de los helechos y licófitos de Colombia. Acta Bot. Malacit. 2017, 42, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Squeo, F.A.; Cavieres, L.A.; Arancio, G.; Novoa, J.E.; Matthei, O.; Marticorena, C.; Rodríguez, R.; Arroyo, M.T.K.; Muñoz, M. Biodiversidad de la flora vascular en la región de Antofagasta, Chile. Rev. Chil. Hist. Nat. 1998, 71, 571–591. [Google Scholar]
- Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT). Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Protección Ambiental-Especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo. D. Of. Fed. México 2010, 30, 1–78. [Google Scholar]
- Sánchez-Salas, J.; Muro, G.; Estrada-Castillón, E.; Alba-Ávila, J.A. El MER: Un instrumento para evaluar el riesgo de extinción de especies en México. Rev. Chapingo Ser. Zonas Áridas 2013, 12, 30–35. [Google Scholar] [CrossRef] [Green Version]
- IUCN 2021 (International Union for Conservation of Nature). The IUCN Red List of Threatened Species. Version 2021-2. Available online: https://www.iucnredlist.org (accessed on 17 November 2022).
- CITES 2021. Convention on Internacional Trade in Endangered Species of wild fauna and flora. Apendix II. Available online: https://www.cites.org (accessed on 17 November 2022).
- Ceja-Romero, J.; Mendoza-Ruiz, A.; López-Ferrari, A.R.; Espejo-Serna, A.; Pérez-García, B.; García-Cruz, J. Las epífitas vasculares del estado de Hidalgo, México: Diversidad y distribución. Acta Bot. Mex. 2010, 93, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Miguel-Vázquez, M.I.; Espejo-Serna, M.A.; Ceja-Romero, J.; Cerros-Tlatilpa, R. Las angiospermas epífitas de Puebla, México: Riqueza y distribución. Bot. Sci. 2020, 98, 585–596. [Google Scholar] [CrossRef]
- Escalante, T.; Rodríguez, G.; Morrone, J.J. Las provincias biogeográficas del Componente Mexicano de Montaña desde la perspectiva de los mamíferos continentales. Rev. Mex. Biodivers. 2005, 76, 207–252. [Google Scholar] [CrossRef]
- Zhou, X.; Rothfels, C.J.; Zhang, L.; He, Z.R.; Le Pechón, T.; He, H.; Lu, N.T.; Knapp, R.; Lorence, D.; He, X.J.; et al. A large-scale phylogeny of the lycophyte genus Selaginella (Selaginellaceae: Lycopodiopsida) based on plastid and nuclear loci. Cladistics 2016, 32, 360–389. [Google Scholar] [CrossRef] [PubMed]
- Klaus, K.V.; Schulz, C.; Bauer, D.S.; Stützel, T. Historical biogeography of the ancient lycophyte genus Selaginella: Early adaptation to xeric habitats on Pangea. Cladistics 2017, 33, 469–480. [Google Scholar] [CrossRef]
- McHenry, M.A.; Barrington, D.S. Phylogeny and biogeography of exindusiate Andean Polystichum. Am. J. Bot. 2014, 101, 365–375. [Google Scholar] [CrossRef]
- Schuettpelz, E.; Schneider, H.; Huiet, L.; Windham, M.D.; Pryer, K.M. A molecular phylogeny of the fern family Pteridaceae: Assessing overall relationships and the affinities of previously unsampled genera. Mol. Phylogenet. Evol. 2007, 44, 1172–1185. [Google Scholar] [CrossRef]
- Kirkpatrick, R.E. Investigating the monophyly of Pellaea (Pteridaceae) in the context of a phylogenetic analysis of cheilanthoid ferns. Syst. Bot. 2007, 32, 504–518. [Google Scholar] [CrossRef]
- Pteridophytes and Gymnosperms. In Flora of North America; Morin, N.R. (Ed.) Oxford University Press: New York, NY, USA, 1993. [Google Scholar]
- Mehltreter, K. Fern conservation. In Fern Ecology; Mehltreter, K., Walker, L.R., Sharpe, M.J., Eds.; Cambridge University Press: New York, NY, USA, 2010; pp. 323–359. [Google Scholar]
- Guo, Q.; Thompson, D.B.; Valone, T.J.; Brown, J.H. The effect of vertebrate granivores and folivores on plant community structure in the Chihuahuan desert. Oikos 1995, 73, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Castellanos-Villegas, A.E.; Bravo, L.C.; Koch, G.W.; Llano, J.; López, D.; Méndez, R.; Rodríguez, J.C.; Romo, R.; Sisk, T.D.; Yanes-Arvayo, G. Impactos ecológicos por el uso del terreno en el funcionamiento de ecosistemas áridos y semiáridos. In Diversidad Biológica de Sonora; Molina-Freaner, F.E., Van Devender, T.R., Eds.; Universidad Nacional Autónoma de México y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 2010; pp. 157–186. [Google Scholar]
- Armenta-Montero, S.; Carvajal-Hernández, C.I.; Ellis, E.A.; Krömer, T. Distribution and conservation status of Phlegmariurus (Lycopodiaceae) in the state of Veracruz, Mexico. Trop. Conserv. Sci. 2015, 8, 114–137. [Google Scholar] [CrossRef] [Green Version]
- Kessler, M.; Betz, L.; Roedde, S. Red List of the Pteridophytes of Bolivia. 2006. Available online: https://www.fernsofbolivia.uni-goettingen.de (accessed on 17 November 2022).
- Hernández-Rojas, A.C.; Kluge, J.; Krömer, T.; Carvajal-Hernández, C.; Silva-Mijangos, L.; Miehe, G.; Lehnert, M.; Weigand, A.; Kessler, M. Latitudinal patterns of species richness and range size of ferns along elevational gradients at the transition from tropics to subtropics. J. Biogeogr. 2020, 47, 1383–1397. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, S.J. Vascular epiphytes in Argentinian Yungas: Distribution, diversity, and ecology. Bot. Rev. 2022. [Google Scholar] [CrossRef]
- Kessler, M.; Kluge, J.; Hemp, A.; Ohlemüller, R. A global comparative analysis of elevational species richness patterns of ferns. Glob. Ecol. Biogeogr. 2011, 20, 868–880. [Google Scholar] [CrossRef]
- Bhattarai, K.R.; Vetaas, O.R.; Grytnes, J.A. Fern species richness along a central Himalayan elevational gradient, Nepal. J. Biogeogr. 2004, 31, 389–400. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Holbrook, N.M.; Zwieniecki, M.A.; Palma, B. Leaf hydraulic capacity in ferns, conifers and angiosperms: Impacts on photosynthetic maxima. New Phytol. 2005, 165, 839–846. [Google Scholar] [CrossRef]
- Rzedowski, J. Análisis preliminar de la flora vascular de los bosques mesófilos de montaña de México. Acta Bot. Mex. 1996, 35, 25–44. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Jiménez, C.A.; Téllez-Valdés, O.; Luna-Vega, I. Clasificación de los bosques mesófilos de montaña de México: Afinidades de la flora. Rev. Mex. Biodivers. 2012, 83, 1110–1114. [Google Scholar] [CrossRef] [Green Version]
- Tejero-Díez, J.D.; Torres-Díaz, A.N.; Gual-Díaz, M. Licopodios y helechos en el bosque mesófilo de montaña en México. In Bosques Mesófilos de Montaña de México: Diversidad, Ecología y Manejo; Gual-Díaz, M., Ed.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 2014; pp. 197–220. [Google Scholar]
- Moran, R.C. A Natural History of Ferns; Timber Press: Portland, OR, USA, 2004. [Google Scholar]
- Kessler, M.; Kluge, J. Mountain ferns: What determines their elevational ranges and how will they respond to climate change? Am. Fern J. 2022, 112, 285–302. [Google Scholar] [CrossRef]
- Callaway, R.M.; Reinhart, K.O.; Moore, G.W.; Moore, D.J.; Pennings, S.C. Epiphyte host preferences and host traits: Mechanisms for species-specific interactions. Oecologia 2002, 132, 221–230. [Google Scholar] [CrossRef]
- Van Wesenbeeck, B.K.; Van Mourik, T.; Duivenvoorden, J.F.; Cleef, A.M. Strong effects of a plantation with Pinus patula on Andean subparamo vegetation: A case study from Colombia. Biol. Conserv. 2003, 114, 207–218. [Google Scholar] [CrossRef]
- Villers-Ruiz, L.; Trejo-Vázquez, I. Impacto del cambio climático en los bosques y áreas naturales protegidas de México. Interciencia 1998, 23, 10–19. [Google Scholar]
- Arriaga, L.; Gómez, L. Efectos del cambio climático en algunos componentes de la biodiversidad. In Cambio climático: Una visión desde México; Martínez, J., Fernández-Bremauntz, A., Eds.; Instituto Nacional de Ecología: Mexico City, Mexico, 2004; pp. 255–265. [Google Scholar]
- Villers-Ruiz, L.; Trejo-Vázquez, I. Evaluación de la vulnerabilidad en los sistemas forestales. In Cambio climático: Una visión desde México; Martínez, J., Fernández-Bremauntz, A., Eds.; Instituto Nacional de Ecología: Mexico City, Mexico, 2004; pp. 239–254. [Google Scholar]
- Novo-Fernández, A.; Franks, S.; Wehenkel, C.; López-Serrano, P.M.; Molinier, M.; López-Sánchez, C.A. Landsat time series analysis for temperate forest cover change detection in the Sierra Madre Occidental, Durango, Mexico. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 230–244. [Google Scholar] [CrossRef]
- Contreras-Medina, R.; Morrone, J.J.; Luna-Vega, I. Biogeographic methods identify gymnosperm biodiversity hotspots. Naturwissenschaften 2001, 88, 427–430. [Google Scholar] [CrossRef]




| Families | Species | Genera | Endemic Species | NOM-059 |
|---|---|---|---|---|
| Anemiaceae | 11 | 1 | 3 | |
| Aspleniaceae | 25 | 1 | 3 | |
| Athyriaceae | 5 | 3 | ||
| Blechnaceae | 7 | 3 | ||
| Cyatheaceae | 1 | 1 | Cyathea costaricensis | |
| Cystopteridaceae | 2 | 1 | ||
| Dennstaedtiaceae | 4 | 2 | ||
| Dryopteridaceae | 25 | 6 | ||
| Equisetaceae | 5 | 1 | ||
| Gleicheniaceae | 1 | 1 | ||
| Hymenophyllaceae | 2 | 2 | ||
| Isoetaceae | 3 | 1 | ||
| Lycopodiaceae | 3 | 2 | ||
| Lygodiaceae | 1 | 1 | ||
| Marsileaceae | 4 | 2 | ||
| Nephrolepidaceae | 1 | 1 | ||
| Ophioglossaceae | 8 | 4 | ||
| Osmundaceae | 1 | 1 | ||
| Plagiogyriaceae | 1 | 1 | ||
| Polypodiaceae | 31 | 6 | Polypodium 1 | Campyloneurum phyllitidis |
| Psilotaceae | 2 | 1 | Psilotum × complanatum P. nudum | |
| Pteridaceae | 118 | 23 | Argyrochosma 1, Myriopteris 1, Notholaena 2 | |
| Salviniaceae | 2 | 1 | ||
| Selaginellaceae | 30 | 1 | 1 | Selaginella porphyrospora |
| Tectariaceae | 2 | 1 | ||
| Thelypteridaceae | 10 | 6 | ||
| Woodsiaceae | 7 | 1 | 1 | |
| Total 27 | 312 | 75 | 13 | 4 |
| Study Area | Elevation Ranges | Area * (km2) | Number of Pteridophyte Species | TBI (S/InA) |
|---|---|---|---|---|
| SMOc (present study) | 300–3500 | 251,648 | 312 | 25.17 |
| SMOr [16] | 200–3694 | 220,192 | 424 | 34.47 |
| TMVB (Tejero-Díez pers. data) | 300–5636 | 163,015 | ~415 | 34.58 |
| SMS [4] | 300–3700 | ±143,447 | 614 | 51.71 |
| ST [18] | 300–3284 | ±59,161 | ~550 | 50.05 |
| Family | Species | NOM-059 | IUCN | CITES |
|---|---|---|---|---|
| Aspleniaceae | Asplenium formosum | LC | ||
| Aspleniaceae | Asplenium monanthes | LC | ||
| Aspleniaceae | Asplenium trichomanes | LC | ||
| Cyatheaceae | Cyathea costaricensis | P | Appendix II | |
| Dennstaedtiaceae | Pteridium latiusculum | LC | ||
| Equisetaceae | Equisetum hyemale | LC | ||
| Hymenophyllaceae | Hymenophyllum tunbrigense | LC | ||
| Lycopodiaceae | Palhinhaea cernua | LC | ||
| Nephrolepidaceae | Nephrolepis undulata | LC | ||
| Ophioglossaceae | Ophioglossum nudicaule | LC | ||
| Ophioglossaceae | Ophioglossum reticulatum | LC | ||
| Osmundaceae | Osmunda regalis | LC | ||
| Polypodiaceae | Campyloneurum phyllitidis | A | ||
| Psilotaceae | Psilotum × complanatum | A | ||
| Psilotaceae | Psilotum nudum | CR | ||
| Pteridaceae | Adiantum capillus-veneris | LC | ||
| Pteridaceae | Anogramma leptophylla | LC | ||
| Pteridaceae | Pteris cretica | LC | ||
| Salviniaceae | Azolla microphylla | LC | ||
| Selaginellaceae | Selaginella porphyrospora | P | ||
| Thelypteridaceae | Cyclosorus interruptus | LC | ||
| Total | 4 | 17 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejero-Díez, J.D.; Contreras-Medina, R.; Torres-Díaz, A.N.; González-Elizondo, M.S.; Sánchez-González, A.; Luna-Vega, I. Diversity of the Pteridoflora of Montane Northwestern Mexico. Diversity 2023, 15, 324. https://doi.org/10.3390/d15030324
Tejero-Díez JD, Contreras-Medina R, Torres-Díaz AN, González-Elizondo MS, Sánchez-González A, Luna-Vega I. Diversity of the Pteridoflora of Montane Northwestern Mexico. Diversity. 2023; 15(3):324. https://doi.org/10.3390/d15030324
Chicago/Turabian StyleTejero-Díez, J. Daniel, Raúl Contreras-Medina, Alin N. Torres-Díaz, M. Socorro González-Elizondo, Arturo Sánchez-González, and Isolda Luna-Vega. 2023. "Diversity of the Pteridoflora of Montane Northwestern Mexico" Diversity 15, no. 3: 324. https://doi.org/10.3390/d15030324