
Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species
by
 , , , ,
, , , ,
Qiuyi Zhong
1,2,
Min Ai
1,2,
Fiona Ruth Worthy
1,2,
Ancheng Yin
1,2,
Yi Jiang
3,
Lisong Wang
1,2 and
Xinyu Wang
1,2,* 1
Key Laboratory for Plant Diversity and Biogeography of East Asia, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
2
Yunnan Key Laboratory for Fungal Diversity and Green Development, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
3
Yunnan Institute of Microbiology, Chenggong Campus of Yunnan University, Kunming 650500, China
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(6), 705; https://doi.org/10.3390/d15060705
Submission received: 26 April 2023
/
Revised: 22 May 2023
/
Accepted: 23 May 2023
/
Published: 25 May 2023
(This article belongs to the Special Issue Phylogeny, Taxonomy and Ecosystems of Lichens)
Abstract
:Rinodina is a lichenized fungal genus belonging to the Physciaceae, with c. 300 species worldwide. Nearly a century ago, Zahlbruckner described five species of the genus Rinodina from Southwest China. The type collections were the only records for these species. In the present study, new records for four of these species: Rinodina cornutula, R. globulans, R. handelii, and R. setschwana, and a recently described species, R. pluriloculata, are documented based on specimens collected from the holotype localities. Furthermore, one new species was discovered: Rinodina hengduanensis, characterized by areolate to subsquamulose thallus, jigsaw-like areoles, lecanorine apothecium, and Dirinaria-type ascospores. Rinodina setschwana is transferred to the genus Buellia based on its morphology, chemistry, and phylogeny and proposed as Buellia setschwana. We provide detailed morphological descriptions, pictures, and molecular phylogenetic analyses.
1. Introduction
Among lichenized fungi with a Holarctic distribution, Rinodina (Ach.) Gray is one of the better understood genera. It has been studied in the Iberian Peninsula [1], Scandinavia [2], South Europe [3,4], the Old World [5], North America [6,7], and Northeast Asia [8]. However, molecular data have shown that Rinodina is a polyphyletic genus [9,10,11,12]. Species-level concepts within Rinodina are becoming better established, with clear outlines of species groups emerging in conjunction with a refined understanding of crustose microlichen transitions [8,12]. However, there remains a substantial knowledge gap for East Asia, especially for Southwest China, a region that has played an important role in the evolution of higher plants [8,13,14,15,16]. The inclusion of specimens and molecular data from East Asia is vital for the generation of testable hypotheses regarding lichen biogeography [8].
A total of 51 Rinodina species have been reported from China [5,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38]. In the early 20th century, five of these species were described from Southwest China by Zahlbruckner [19]. The chemistry of their lichen substances was not recorded. More recently, Aptroot and Sparrius described a new species, R. pluriloculata Aptroot and Sparrius, from the same area, including the chemistry of its lichen substances but not molecular data [29]. Since the initial publication of these species, no further studies have been conducted. Inter-specific comparisons of chemical compounds are lacking, and the phylogenetic position of these species remains unclear.
During the Second Tibetan Plateau Scientific Expedition and Research Program (STEP), we conducted a thorough survey of the lichen diversity of western China, with a particular focus on the Qinghai-Tibetan Plateau. With the addition of specimens that we previously collected in Yunnan Province, over 150 specimens that were confirmed as Rinodina species have been stored in the lichen Herbarium at the Kunming Institute of Botany (KUN). We surveyed the relevant literature, examined the morphology of type specimens, and compared these to our newly collected materials from the type localities. This confirmed that we had obtained fresh collections of the species, which were originally described from southwestern China nearly a century ago [19] and twenty years ago [29]. In addition, we discovered one new species of Rinodina. In the present paper, we characterize these species using morphological, chemical, and molecular evidence and provide a key to the species included in this study.
2. Materials and Methods
2.1. Morphological and Chemical Study
All the new materials from this study have been deposited in the Herbarium of the Kunming Institute of Botany (KUN). Observations of specimens were made using a Nikon SMZ 745T dissecting microscope (Nikon Corp., Tokyo, Japan). Vertical sections of apothecia and thalli were cut with a razor blade, then mounted in GAW (glycerol:ethanol:water = 1:1:1) and examined under a Nikon Eclipse 50i stereomicroscope. Measurements and descriptions of the sections and ascospores were conducted under the microscope and stereomicroscope at either ×10 or ×40 magnification to an accuracy of 10 μm and 2.5 μm, respectively. Photographs were all taken using a Nikon digital camera head, the DS-Fi2. Secondary metabolites were identified by spot testing and by thin-layer chromatography (TLC), following the methods of Culberson [39] and Orange et al. [40], using solvent system C (toluene:acetic acid = 85:15).
2.2. DNA Extraction, Purification, and Sequencing
Total genomic DNA was extracted from 26 specimens using DNAsecure Plant Kits (Tiangen Biotech, Beijing, China), following the manufacturer’s protocol. Three loci were amplified: the mitochondrial SSU gene with primer pairs mtSSU1 and mtSSU3R [41], the internal transcribed spacer region (nrITS) with ITS1F [42] and ITS4 [43], and the 5′ portion end of the 28S, including the D1–D2 domain [44], with LR0R [45] and LR5 [46]. PCR amplification was performed in 25 μL reactions containing 12.5 μL of 2 × MasterMix (0.1 units/μL TaqDNA polymerase, 4 mM MgCl2, and 0.4 nM dNTPs; Aidlab Biotechnologies Co. Ltd., Beijing, China), 0.5 μL of each primer, 9.5 μL of ddH2O, 2 μL of each primer, and 1 μL of DNA, using the PCR settings and primer profile issued by Zhao et al. [47]. Polymerase chain reaction (PCR) products were sequenced by TsingKe Biological Technology (Kunming, China).
2.3. Phylogenetic Analysis
The DNA sequences were aligned using MAFFT v. 7.107 in GENEIOUS v. 8.0.2 with the following parameters: algorithm = auto; scoring matrix = 200 PAM/k = 2; gap open penalty = 1.53; offset value = 0.123 [48]. A 3-locus (nrITS, mtSSU, and nrLSU) concatenated matrix was generated using GENEIOUS v. 8.0.2. This matrix contained both Rinodina species and species belonging to related genera in the Physciaceae and Caliciaceae. In addition to our newly generated sequences, other related sequences were downloaded from GenBank and added to the matrix (Table 1). Subsequently, a maximum likelihood (ML) tree and a Bayesian inference (BI) tree were constructed. Based on the lowest Bayesian information criterion (BIC), the partitioned analysis was selected using ModelFinder in IQ-TREE v.1.6.12 for the subsequent ML and BI analyses [49,50]: In ML analysis, GTR + F + I + G4 is for ITS1 and ITS2, TNe + I + G4 for 5.8S rRNA and nrLSU, and TPM3u + F + I + G4 for mtSSU; in BI analysis, GTR + F + I + G4 is for ITS1 and ITS2, SYM + I + G4 for 5.8S rRNA and nrLSU, and HKY + F + I + G4 for mtSSU. The ML analyses were performed by the IQ-TREE web server [51,52,53]. The BI analyses were performed by MrBayes v. 3.2.7 [54], using three Markov chains running for 10 million generations for the concatenated dataset. The trees were sampled every 100 generations, and the first 25% of the trees were discarded as burn-ins. Posterior probabilities (PPs) were obtained from the 95% majority rule consensus tree of all saved trees. The tree files were visualized with FigTree 1.4.2 [55].
3. Results and Discussion
By re-collecting these species from their holotype localities and conducting morphological and phylogenetic analyses, we confirmed the identities of Rinodina cornutula, R. globulans, R. handelii, Buellia setschwana (≡ R. setschwana), and R. pluriloculata. Detailed figures of morphology and spores, together with information regarding chemical compounds, have been provided. Furthermore, the phylogenetic positions of these species have been confirmed. An additional species has been described as new to science: R. hengduanensis, for which morphological descriptions and molecular data have been provided.
The sequence alignment comprised 105 terminals, of which 25 were newly generated sequences from this study (Table 1). Phylogenetic trees that were based on datasets generated from either ML or BI analyses had almost identical topologies, with minimal differences in the level of statistical support. The phylogenetic analysis showed that the families Physciaceae and Caliciaceae both formed monophyletic clades. Rinodina cornutula, R. globulans, R. handelii, and R. pluriloculata belong to the Physciaceae, while Buellia setschwana (syn. R. setschwana) belongs to the Caliciaceae.
Our results indicate that R. pluriloculata is phylogenetically close to R. conradii. Both species have 3–8-septate ascospores, but R. pluriloculata can be distinguished by its pale to olive green thallus, punctiform, discrete, orbicular, convex to flattened soralia, bluish, granular soredia, and submuriform-type ascospores. R. cornutula is phylogenetically closely related to R. oxydata. The morphological characters of these two species are largely similar: both are characterized by Mischoblastia-type spores and a thallus with atranorin in the cortex. They can be separated by their thallus color and habitat. The molecular data available for R. oxydata showed that this species is not monophyletic, so further research is required based on samples from its type locality. R. handelii appears close to R. orientalis and R. zeorina in the phylogenetic tree, but the three species can be separated by their ascospore morphology and their chemistry. R. handelii lacks secondary metabolites, and it has Milvina- to Mischoblastia-type ascospores (up to 27 μm), which are longer than the spores of the other two species (both less than 20 μm). R. orientalis has Physcia-type spores, whereas R. zeorina contains zeorin and has Dirinaria-type ascospores. R. globulans forms a well-supported clade (100/100/1.00), which is also supported by its morphological, chemical, and geographic characteristics. It differs from other species by its round, capitate, and scattered thallus areoles covered with soralia. Interestingly, Buellia setschwana (syn. R. setschwana) forms a monotypic lineage that is close to B. polita. These two species can be distinguished by the characters of their apothecia: B. polita has a reddish brown hypothecium and immersed lecideine apothecia, whereas B. setschwana has a hyaline hypothecium and cryptolecanorine apothecia. The apothecia characters of B. setschwana are identical to those of other species within Rinodina, which was probably the basis for its previous placement within the genus Rinodina.
The new species Rinodina hengduanensis formed a single clade, represented by a SH-aLRT support of 100, an ultrafast bootstrap support of 100, and a posterior probability of 1 for the branch (Figure 1). Its species status is further supported by its distinctive morphological, chemical, and geographic characteristics.
4. Taxonomy
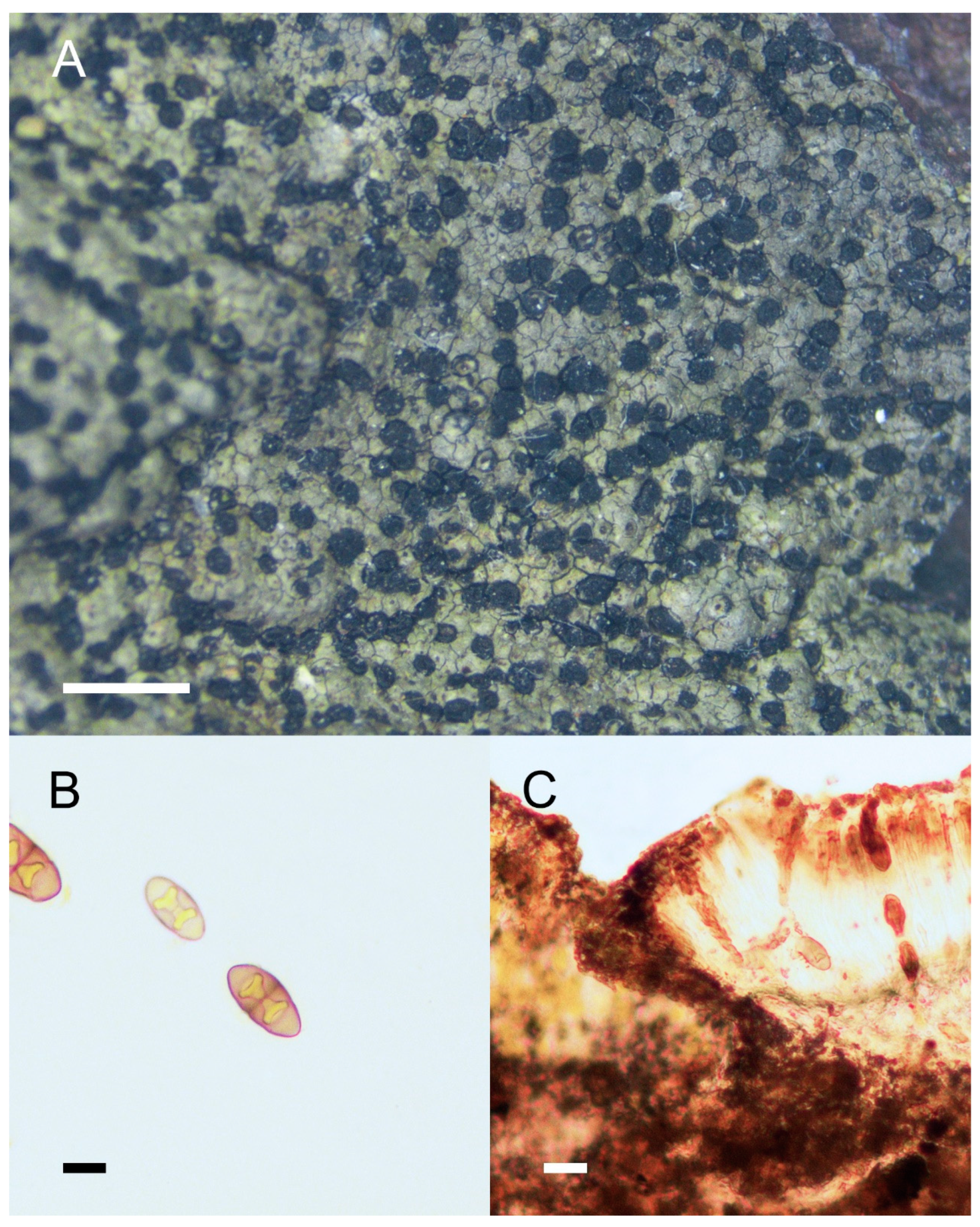
Rinodina cornutula Zahlbr., in Handel-Mazzetti, Symb. Sinic. 3: 233 (1930), Figure 2.
Type: China, Yunnan Prov., Manhao nahe der Grenze von Tonkin, Tonschieferfelsen am Flußufer in der tr. St., 200 m, 2 March 1915, Handel-Mazzetti no. 5858 (WU, holotype!).
Description:Thallus crustose, thin, areolate, continuous at the central part, scattered near the margins, margins not distinct; areoles irregular in shape, 0.2–0.6 mm wide; upper surface flat, brown to olive brown, mottled, dull, non-pruinose; prothallus absent or delimiting the thallus as a black outline; vegetative propagules absent. Apothecia common, usually abundant and contiguous, lecanorine (becoming lecideine), broadly attached to slightly innate, 0.2–0.6 mm in diam.; disc black, plane and roundish, non-pruinose; margin concolorous with thallus, entire, persistent, usually becoming carbonized and concolorous with the disc when mature; exciple hyaline to dark brown, with algal cells; hymenium hyaline, colorless, not inspersed, 75–125 μm high; paraphyses simple, conglutinate, apices expanded, light brown; epihymenium pale brown to dark brown; hypothecium hyaline, colorless, 35–75 μm deep; asci 8-spored, Lecanora-type; ascospores 1-septate, Type A development, Mischoblastia-type, pale brown, lumina triangle- or heart-shaped, walls thickened at both ends and the septum, 18–25 × 7.5–12.5 μm, torus indistinct. Pycnidia were not seen.
Chemistry: Cortex K+ yellow; KC+ yellow or orange; C−, P+ faint yellow; medulla I+ blue. Containing atranorin (confirmed by TLC).
Ecology and distribution: This species is distributed across subtropical to temperate regions, growing on siliceous rocks, rhyolite, or basalt, at elevations between 950 and 2500 m. Endemic to China, previously reported in Sichuan and Yunnan [19,36].
Notes: Rinodina cornutula is characterized by its flat, areolate, ochre to olive thallus, broadly attached to slightly innate apothecia, Mischoblastia-type ascospores, and containing atranorin. This species was originally collected from Manhao town, Yunnan province, China. The specimens that we collected from the same area were compared to type specimens and confirmed as identical to the holotype. This species is similar to Rinodina cana (Arnold) Arnold in morphology, but R. cana lacks atranorin (spot tests are all negative), and its apothecia are usually entirely innate, with Milvina-type ascospores. It might be confused with Rinodina moziana (Nyl.) Zahlbr., but the latter species can be distinguished by its verruciform or granular thallus and its constricted ascospores when mature. R. oxydata (A. Massal.) A. Massal differs from this species by its gray to ochraceous thallus and shorter ascospores (<18 μm long). R. cornutula would key out at couplet 9 in Lee and Hur’s paper, as it has a plane thallus, Mischoblastia-type, 18–25 × 7.5–12.5 μm ascospores, and is rarely swollen at the septum [76].
Specimens examined: China, Yunnan Prov.: Lijiang Co., Yufeng Temple, 2500 m, on rock, August 15, 1982, Wang Lisong 82-1099; Lijiang City, Yulong Naxi Autonomous Co., on the way from Lijiang to Ninglang, Jinsha River side, 1846–1883 m, on rock, April 9, 2019, Wang Lisong et al. 19-62703, 19-62705, 19-62706; Honghe prefecture, Lvchun Co., Niukong Vil., 972 m, on rock, December 17, 2020, Wang Xinyu et al. XY20-3571, XY20-3572.
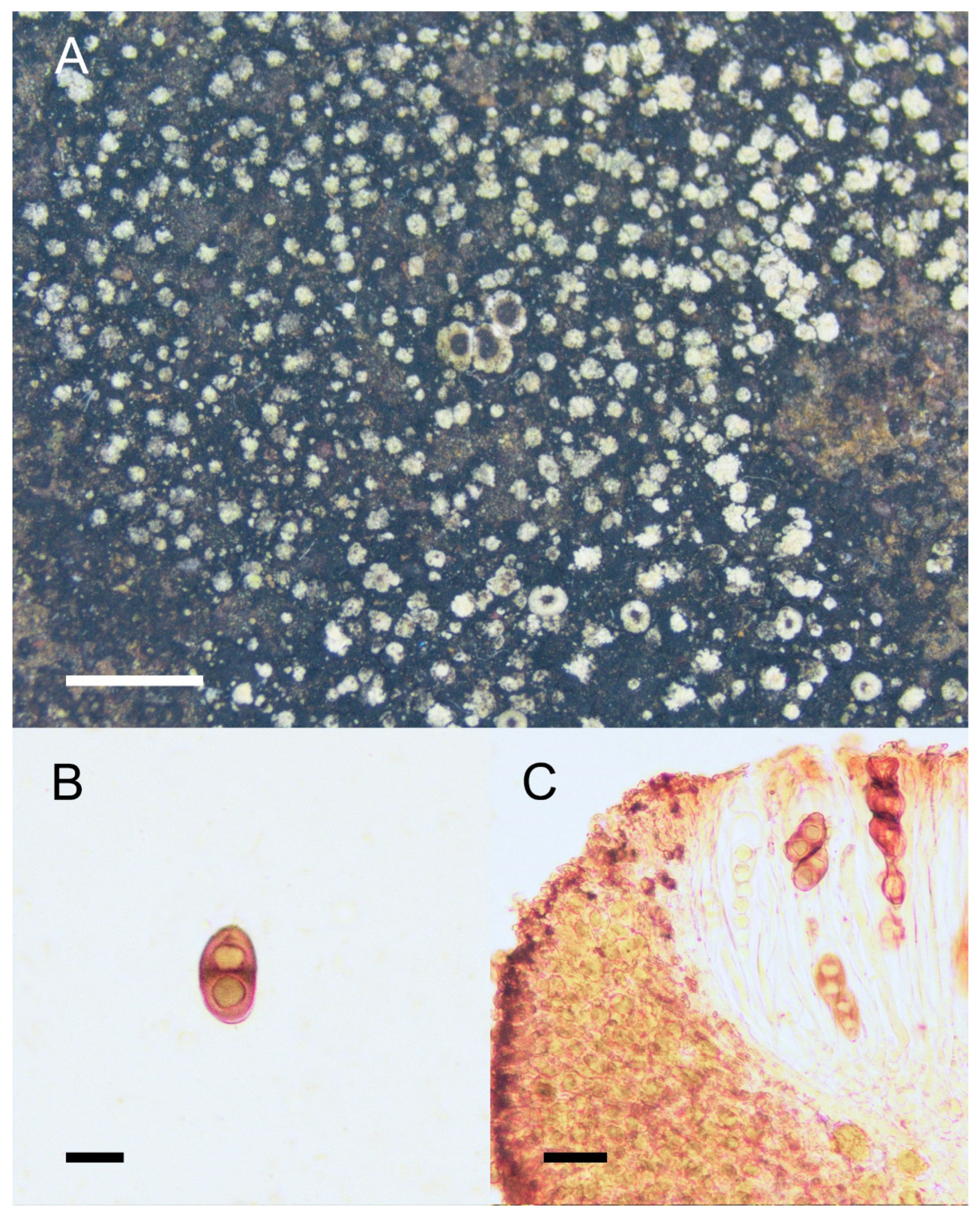
Rinodina globulans Zahlbr., in Handel-Mazzetii, Symb. Sinic. 3: 233 (1930), Figure 3.
Type: China, Sichuan Prov., Eisenschüssiger Sandstein in der tp. St. Um den Paß Dsiliba im Daliangschan e von Ningyüen, ±3000 m, 26 April 1914, Handel-Mazzetti no. 1771 (WU, holotype!).
Description: Thallus crustose, round, capitate to areolate, scattered; areoles 0.1–0.4 mm in diam.; surface pale green (greyish white to pale after storage in herbarium), with white to greyish white pruina, covered by abundant soralia, globular, pale green; prothallus prominent, black, persistent; Apothecia lecanorine, usually scattered, rarely contiguous, sessile, 0.6–0.8 mm in diam.; disc brown to dark brown, concave, round, non-pruinose; margin concolorous with thallus, thick, distinctly raised, entire, persistent; exciple hyaline to pale brown, with algal cells, persistent; hymenium hyaline, colorless, not inspersed, 80–130 μm high, I+ blue; paraphyses simple or merely branched at the apices; epihymenium olive to greyish brown; hypothecium hyaline, colorless, 40–65 μm deep; asci 8-spored, Lecanora-type; ascospores 1-septate, Type A development, Mischoblastia- to Pachysporaria-type, brown, lumina triangle- or trapezoid-shaped; walls thicken at both ends and at the septum, 15–25 × 7.5–12.5 μm; torus absent with oil droplets when over-mature. Pycnidia were not seen.
Chemistry: Cortex and medulla K+ yellow, KC−, C−, P−. Atranorin, zeorin, and stictic acid are detected by TLC.
Ecology and distribution: This species is saxicolous and often grows on sandstone at elevations between 2000 and 3600 m. It is endemic to China, previously reported in Sichuan province [19,36]. This is a new record for Yunnan Province, China.
Notes: R. globulans is a unique species with numerous, scattered, globular, pale green soralia and a prominent black prothallus, which can be reliably distinguished from other Rinodina species in China. Apothecia are rare in this species, and the ascospores are Mischoblastia-Pachysporaria-type. A recently published species, Rinodina punctosorediata Aptroot and Sparrius, is highly similar to this species, with identical external and anatomic morphology [29]. However, R. punctosorediata grows both on trees (Castanopsis sp.) and rocks, whereas the holotype of R. globulans and all our collections grew on rocks. Furthermore, R. punctosorediata has smaller ascospores (<18 μm long) than the Milvina-type. R. globulans would be keyed out at couplet 2 in Lee and Hur’s paper because it has scattered thallus areoles covered with abundant, globular, pale green soralia [76].
Specimens examined: China, Sichuan Prov.: Liangshan Yi Autonomous Prefecture, Yanyuan Co., Weicheng Town, 2816–2831 m, on sand rock, 28 2020, Wang Xinyu et al. XY20-280, XY20-279. Yunnan Prov.: Diqing Tibetan Autonomous Prefecture, Shangri-La City, Jiantang Town, Pudacuo National Park, Bita, 3496 m, on rock, 25 July 2020, Wang Xinyu et al. XY20-117; Lijiang City, Ninglang Yi Autonomous Co., Xichuan Vil., next to Lining Highway, 2509 m, on rock, 26 July 2020, Wang Xinyu et al. XY20-171; Dali Prefecture, Nanjian Co., Leqiu Vil., Awuhe Vil., Dayao Reservoir, 2095 m, on rock, 2 July 2015, Ye Xin and Wang Weicheng 15-47871; Luquan Co., Jiaozi Snow Mt., 3600 m, on rock, September 18, 2003, Wang Lisong 03-22628; Dali Co., Cangshan Mt., Zhonghe Temple, 2580 m, on rock, August 30, 2005, Wang Lisong et al. 05-25172.
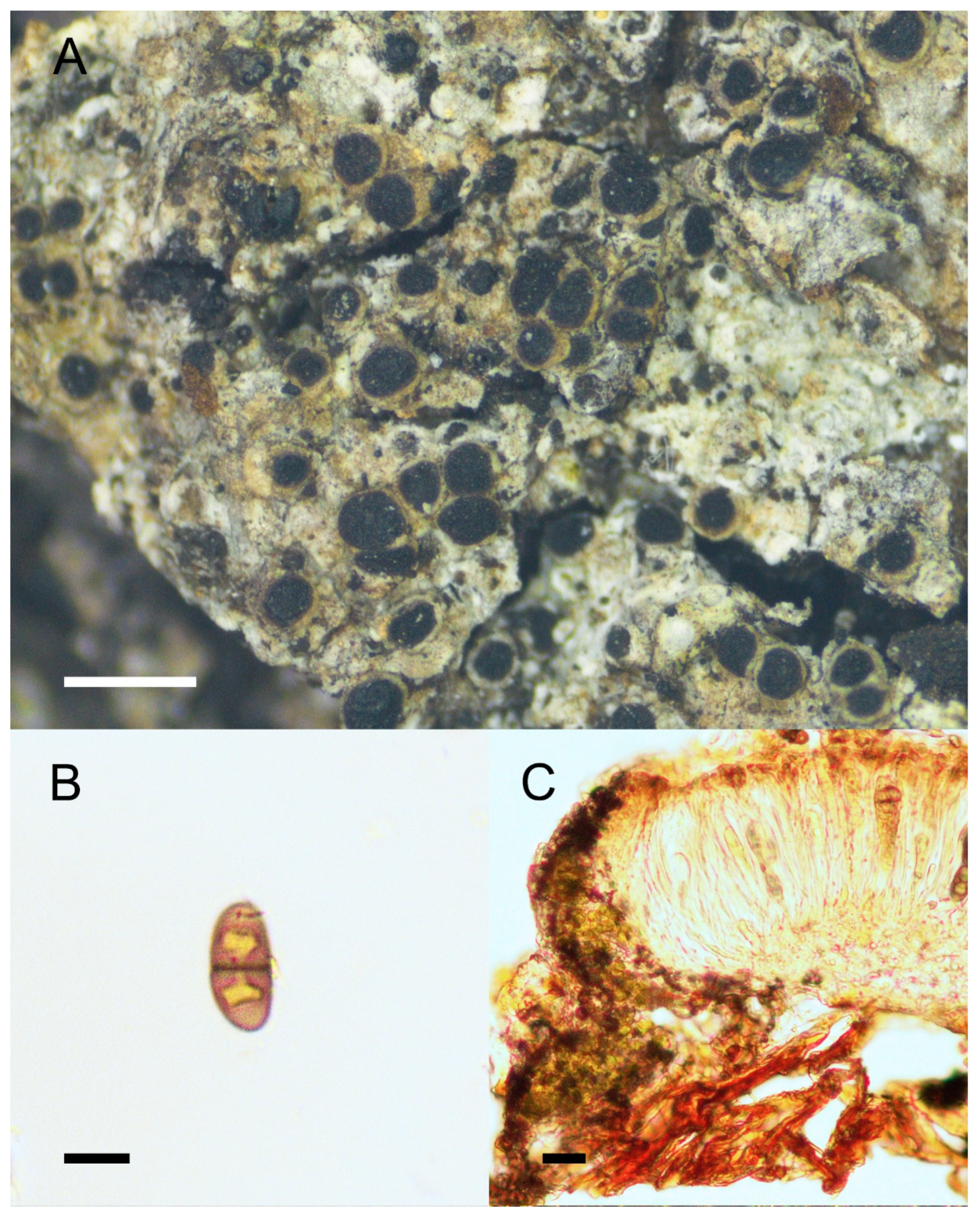
Rinodina handelii Zahlbr., in Handel-Mazzetti, Symb. Sinic. 3: 232 (1930), Figure 4.
Type: China, Yunnan Prov., Lebende Rinde von Pflaumenbäumen in Gärten der wtp. St. in Yünnanfu, 1920 m, February 24, 1914, Handel-Mazzetti no. 275 (WU, holotype; US, isotype).
Description: Thallus crustose, thin, rimose to continuous, somewhat verrucose; areoles 0.1–0.2 mm wide; upper surface rough to verrucous, grey, brown, or pale green, dull, non-pruinose; prothallus absent; vegetative propagules absent. Apothecia common, scattered or contiguous, lecanorine, broadly attached, 0.2–0.6 mm in diam.; disc ochre brown to dark brown, plane to convex, round, non-pruinose; margin concolorous with thallus, thin, entire, disappeared when mature; exciple hyaline, with abundant algal cells; hymenium hyaline, colorless, not inspersed, 50–100 μm high, I+ blue; paraphyses simple, somewhat conglutinate, apices expanded, dark pigmented; epihymenium dark brown; hypothecium hyaline, colorless or pale brown, 50–100 μm deep; asci 8-spored, Lecanora-type; ascospores 1-septate, Type A development, pale brown to brown, Milvina- to Mischoblastia-type, 20–27 × 7–12.5 μm, torus present, always inspersed with oil droplets when mature. Pycnidia were not seen.
Chemistry: Spot tests were all negative; no metabolites were detected by TLC.
Ecology and distribution: This species is corticolous, growing on bark or branches of Pinus, Quercus, Rhododendron, etc., in coniferous forest or mixed coniferous broad-leaved forest, at elevations between 2700 and 3600 m. It has only previously been reported from Yunnan province, China [19,36]. This is a new record for Sichuan province.
Notes: R. handelii is a corticolous species with a thin, rimose to continuous, rough thallus; sessile apothecia; a dark brown disc; characterized by ascospores with torus, always inspersed with oil droplets when mature; a size of 20–27 × 7–12.5 μm. It could be confused with other corticolous species such as Rinodina bolanderi and R. subminuta, but R. bolanderi can be distinguished by containing atranorin and zeorin, and Teichophila-type ascospores, and R. subminuta can be distinguished by containing zeorin and Physcia-type ascospores. R. handelii also resembles Rinodina subleprosula Jatta in external morphology, which is a corticolous species previously described by Jatta from China [17]. However, R. subleprosula has larger ascospores (27.5–30.5 × 12–15 μm). Another corticolous species, Rinodina cinereovirens (Vain.), has similar ascospores to R. handelii, but it can be distinguished by its medulla, which is UV + blue-white and contains sphaerophorin. R. handelii would key out at couplet 60 in Lee and Hur’s paper because the margins of the apothecia are not radially cracked and there are Milvina- to Mischoblastia-type ascospores [76].
Specimens examined: China, Sichuan Prov.: Liangshan Prefecture, Yanyuan Co., Weicheng Town, 3210 m, on Pinus densata, 28 July 2020, Wang Lisong et al. 20-66526. Yunnan Prov.: Diqing Prefecture, Shangri-La City, Jiantang Pudacuo National Park, Bita, 3540 m, on Quercus bark, Wang Xinyu et al. XY20-134; Gongshan Co., Yeniu Valley, 2700 m, on bush, 30 May 2000, Wang Lisong 00-19328; Lijiang City, Ninglang Co., Hongqiao Vil., c. 3100 m, on branch, 27 July 2020, Wang Xinyu et al. XY20-224, XY20-238-1.
Rinodina pluriloculata Aptroot and Sparrius, Fungal Diversity 14: 42 (2003), Figure 5.
Type: CHINA, Yunnan Prov., 5 km W of Kunming, just outside of the city, 1750 m alt., 48RTN604714, on bark of tree base of living Eucalyptus globulus, 16 October 2002, Aptroot no. 55505 (B, holotype; KUN isotype).
For a detailed description, see [29].
Notes: R. pluriloculata is characterized by its continuous, verrucous, pale green to olive thallus; punctiform, discrete, orbicular, convex to flattened soralia; bluish, granular soredia; lecanorine, sessile apothecia; and 3–8-septate, Submuriform-type, pale brown ascospores 27–35 × 15–17.5 μm. Spot tests: cortex and medulla K− or + yellow, KC−, C−, P−; containing skyrin and ± atranorin by TLC. This species was previously reported in Yunnan, growing on rock or bark at elevations around 2500 m. According to Aptroot and Sparrius, the holotype of this species grows on the tree trunk base of Eucalyptus globulus, with an additional cited specimen growing on soil and shale [29]. The specimens that we collected all grew on rocks. Similar to this species, Rinodina conradii Körb also has 3–8-septate ascospores, but it can be differentiated by its brown thallus, lack of vegetative propagules, and Type B development, Conradii-type ascospores. R. pluriloculata would key out at couplet 20 in Lee and Hur’s paper because it has Type A development ascospores (apical wall thickening after septum formation) and contains skyrin [76].
Specimens examined: China, Yunnan Prov.: Lijiang City, Ninglang Yi Autonomous Co., Xichuan Vil., beside Lining Highway, 2504–2520 m, on rock, 26 July 2020, Wang Xinyu et al. XY20-162, 20-66417.
Buellia setschwana (Zahlbr.) Q. Y. Zhong and Xin Y. Wang, comb. nov., Figure 6.
= Rinodina setschwana Zahlbr., in Handel-Mazzetti, Symb. Sinic. 3: 231 (1930)
Type: China, Sichuan Prov., Sandsteinfelsen in der str. St. bei Dötschang im Pjientschang, 1450 m, 4 April 1914, Handel-Mazzetti no. 1171 (WU, holotype; US, isotype).
MycoBank No. 848787
Description: Thallus crustose, thin, plane, closely attached to the substrate, rimose; upper surface pale gray to pale yellowish brown, dull, non-pruinose; prothallus black, prominent; vegetative propagules absent. Apothecia common, usually scattered or sometimes contiguous, cryptolecanorine to lecideine, innate, 0.3–0.5 (0.7) mm in diam.; disc black, plane, round, or somewhat irregular; margin concolorous with thallus, entire, distinct when young, sometimes becoming carbonized when mature; exciple prominent, brown; hymenium colorless, not inspersed, 80–90 μm high; paraphyses simple, contiguous; apices expanded and septate, brown pigmented; epihymenium brown; hypothecium hyaline, colorless or pale brown, 50–60 μm deep; asci 8-spored, Bacidia-type; ascospores 1-septate, Buellia-type, brown, 12.5–18 × 5–7.5 μm; mature spores with obtuse ends, slightly constricted at septum; torus absent. Pycnidia common, dark brown to black, immersed; ostioles brown pigmented; conidia hyaline, bacilliform, 4–6 × 1 μm.
Chemistry: Cortex K+ yellow; KC+ yellow or orange; C−, P−; medulla I+ blue. Atranorin, arthothelin, norstictic acid, and sometimes stictic acid are detected by TLC.
Ecology and distribution: This species grows on rocks at elevations between 1100 m and 2000 m. It has previously been reported from Sichuan Province, China [19]. This is a new record for Yunnan Province.
Notes: Buellia setschwana is characterized by its thin, rimose thallus; black and prominent prothallus; innate cryptolecanorine to lecideine; black apothecia; and containing atranorin and norstictic acid as main compounds. It resembles Buellia polita in its external morphology and chemical compounds but could be distinguished by the hyaline hypothecium and cryptolecanorine apothecia. Both B. setschwana and the globally distributed Buellia halonia contain the secondary metabolite arthothelin. This could lead to confusion between the two species. However, R. halonia has exciples with typical aeruginose pigments (HNO3+ purple), a reddish brown hypothecium, and adnate apothecia (innate in B. setschwana). Furthermore, R. halonia is always maritime, whereas B. setschwana is continental.
Rinodina setschwana was originally collected in Sichuan Province, Dechang County. It was placed in the genus Rinodina based on its cryptolecanorine apothecia and hyaline hypothecium [19]. After the collection of fresh specimens from the same locality and comparison with the type specimen, we confirmed that our collections were morphologically identical to the holotype. However, phylogenetic analyses demonstrated that this species should be placed within the genus Buellia rather than Rinodina. We consider that the type specimens were previously misassigned as species of Rinodina. Based on the morphology, chemistry, and phylogeny of specimens from the type locality, a new combination is proposed here, Buellia setschwana.
Specimens examined: China, Sichuan Prov.: Liangshan Prefecture, Huili Co., on the way from Dechang to Huili, beside Beijing-Kunming Railway, 1879 m, on rock, April 11, 2019, Wang Lisong et al. 19-62862, 19-62864; Jinyang Co., Dashaba Vil., 640 m, on rock, 12 May 2017, Wang Lisong et al. 17-55142. Yunnan Prov.: Xinping Co., Mopanshan Mt., 2540 m, on rock, 20 December 2008, Wang Lisong et al. 08-29962; Kunming Ci., Yinmin Town, 1100 m, on rock, 16 May 2014, Wang Lisong et al. 14-43770; Nanjian Co., Zhonghua Vil., Kongquedu Port, 2799 m, on rock, 4 July 2015, Ye Xin et al. 15-47743, 15-47744.
New species
Rinodina hengduanensis Q. Y. Zhong and Xin Y. Wang, sp. nov., Figure 7.
Type: China, Sichuan Prov., Liangshan Yi Autonomous Prefecture, Yanyuan Co., Weicheng Town, on sandstone, 2819 m elev., 27°30′ N, 101°41′ E, 28 July 2020, Wang Xinyu, Zhong Qiuyi, and Wang Luting no. XY20-290 (KUN, holotype).
MycoBank No. 847637
Characterized by saxicolous, subsquamulose, greyish brown to olivaceous thallus, lecanorine apothecium, and Dirinaria-type ascospores.
Etymology: The epithet ‘hengduanensis’ refers to its distribution, which is mainly in the Hengduan Mountains region.
Description: Thallus crustose and areolate in the center, areoles jigsaw-like, 0.2–0.4 mm wide, closely attached to the substrate, continuous to scattered, margins prominent with extending squamules, appearing subsquamulose; upper surface flat, greyish brown to olive green, slightly glossy, non-pruinose; prothallus sometimes present (when thallus scattered), black; vegetative propagules absent. Apothecia common, usually scattered, lecanorine, broadly attached, 0.5–0.8 mm in diam.; disc dark brown to black, plane or convex, irregular; margin concolorous with the thallus, crenulate, persistent; exciple hyaline, hyphae parallel, with algal cells; hymenium colorless, hyaline, not inspersed, 100–125 μm high; paraphyses simple, contiguous, apices expanded and septate, brown pigmented; epihymenium ochre brown; hypothecium hyaline, colorless, not inspersed, 80–100 μm deep; asci 8-spored, Lecanora-type; ascospores 1-septate, brown or dark, Type A development, Dirinaria-type, 20–24 × 7.5–12.5 μm, swollen at the septum. Swelling becomes more obvious upon treatment with KOH. Usually with oil inspersed when mature, torus present. Pycnidia were not seen.
Chemistry: Spot tests for cortex were all negative; medulla I− contained traces of gyrophoric acid and skyrin, confirmed by TLC.
Ecology and distribution: This species grows on rocks at approximately 2800 m elevation. It is reported from Sichuan and Yunnan provinces in China.
Notes: R. hengduanensis is characterized by subsquamulose, greyish-brown to olivaceous thallus, jigsaw-like areoles, and Dirinaria-type ascospores. Rinodina gennarii Bagl. is a saxicolous species with the same type of ascospores that has been reported from China [26,27,29,78]. It differs from this new species by having a whitish thallus, crowded brown apothecia, and smaller ascospores (length < 17.5 μm). Rinodina pycnocarpa H. Magn. was originally reported from China. It also has a subsquamulose thallus and saxicolous habit, but it can be differentiated by its Bicincta-type and smaller ascospores (<18 μm). Three other species of Rinodina have similar subsquamulose thalli: R. dolichospora Malme can be differentiated by its Pachysporaria-type I spores and lack of substances; R. intermedia Bagl. can be recognized by its 3-septate to submuriform ascospores with type A development; whereas R. willeyi Sheard and Giralt can be separated by having soredia on the areole margin (sublabriform soralia) and its pannarin and zeorin content. R. hengduanensis would key out at couplet 4 in Lee and Hur’s paper because it has a continental distribution and has Dirinaria-type, larger (length > 20 μm) ascospores [76].
Specimens examined: China, Sichuan Prov.: Liangshan Prefecture, Yanyuan Co., Weicheng Town, 2817–2865 m, on sandy rock, 28 July 2020, Wang Lisong et al. 20-66506, XY20-290, XY20-287. Yunnan Prov.: Diqing Prefecture, Shangri-La City, Luoji Vil., Niru Vil., 2774–2804 m, on rock, 21 July 2020, Wang Xinyu et al. XY20-44, XY20-41.
5. Conclusions
In the present study, using morphological, chemical, and phylogenetic methods, new records for the five endemic species Rinodina cornutula, R. globulans, R. handelii, R. setschwana, and R. pluriloculata from Southwest China were documented based on specimens collected from the holotype localities. Rinodina setschwana was transferred to the genus Buellia and proposed as Buellia setschwana. One new species was discovered: Rinodina hengduanensis. Photographs, chemical compounds, and sequences support the circumscription of these species. However, such details are lacking for many of the previously proposed species of Rinodina. The absence of genetic material means that no molecular phylogeny can yet be constructed that includes all species currently assigned to the genus. For those species lacking genetic material, further new collections are required from the type localities.
Currently available keys based on morphology and chemistry are either outdated, limited in their geographic scope, or do not include the recently described Rinodina spp. A full revision of Rinodina is required in order to produce a key that should incorporate all described species across its full global distribution.
Author Contributions
Conceptualization, X.W. and L.W.; methodology, X.W.; software, Q.Z. and M.A.; validation, F.R.W., A.Y. and Q.Z.; formal analysis, Q.Z. and M.A.; investigation, Q.Z., M.A. and X.W.; resources, M.A. and A.Y.; data curation, Q.Z. and M.A.; writing—original draft preparation, Q.Z.; writing—review and editing, F.R.W. and X.W.; visualization, Q.Z. and A.Y.; supervision, X.W. and L.W.; project administration, X.W. and L.W.; funding acquisition, X.W., Y.J. and L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Flora Lichenum Sinicorum, grant number 31750001; the Second Tibetan Plateau Scietific Expedition and Research Program (STEP), grant number 2019QZKK0503; Youth Innovation Promotion Association CAS, grant number 2020388, Yunnan Young and Elite Talents Project, National Natural Science Foundation of China, grant number 31970022,32060001; and the Government Project of Yunnan Province (YNWR-QNBJ-2018-085).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found from here: https://www.ncbi.nlm.nih.gov/ (accessed on 24 September 2022).
Acknowledgments
We would like to express our deep thanks to the herbaria WU and US for providing type specimens or digital images. We are grateful to Zhang Yanyun, Xie Congmiao, Wang Luting, Ye Xin, and Wang Weicheng for their help with field research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giralt, M. The lichen genera Rinodina and Rinodinella (lichenized Ascomycetes, Physciaceae) in the Iberian Peninsula. Bibl. Lichenol. 2001, 79, 1–160. [Google Scholar]
- Mayrhofer, H.; Moberg, R. Rinodina. In Nordic Lichen Flora; Ahti, T., Jørgensen, P.M., Kristinsson, H., Eds.; Nordic Lichen Society: Uddevalla, Sweden, 2002; Volume 2, pp. 41–69. [Google Scholar]
- Giralt, M.; Matzer, M. The corticolous species of the genus Rinodina with biatorine or lecideine apothecia in southern Europe and Macaronesia. Lichenologist 1994, 26, 319–332. Available online: https://www.researchgate.net/publication/231922212 (accessed on 11 April 2023). [CrossRef]
- Giralt, M.; Mayrhofer, H.; Sheard, J.W. The corticolous and lignicolous sorediate, blastidiate and isidiate species of the genus Rinodina in southern Europe. Lichenologist 1995, 27, 3–24. Available online: https://www.researchgate.net/publication/231957857 (accessed on 11 April 2023). [CrossRef]
- Mayrhofer, H. Die saxicolen Arten der Flechtengattungen Rinodina und Rinodinella in der Alten Welt. J. Hattori Bot. Lab. 1984, 55, 327–493. [Google Scholar]
- Sheard, J.W. The Lichen Genus Rinodina (Ach.) Gray (Lecanoromycetidae, Physciaceae) in North America, North of Mexico; NRC Research Press: Ottawa, EO, Canada, 2010; pp. 1–246. [Google Scholar]
- Sheard, J.W. A synopsis and new key to the species of Rinodina (Ach.) Gray (Physciaceae, lichenized Ascomycetes) presently recognized in North America. Herzogia 2018, 31, 395–423. [Google Scholar]
- Sheard, J.W.; Ezhkin, A.K.; Galanina, I.A.; Himelbrant, D.; Kuznetsova, E.; Shimizu, A.; Stepanchikova, I.; Thor, G.; Tønsberg, T.; Yakovchenko, L.S.; et al. The lichen genus Rinodina (Physciaceae, Caliciales) in north-eastern Asia. Lichenologist 2017, 49, 617–672. Available online: https://www.researchgate.net/publication/321054036 (accessed on 12 April 2023). [CrossRef]
- Helms, G.; Friedl, T.; Rambold, G. Phylogenetic relationships of the Physciaceae inferred from rDNA sequence data and selected phenotypic characters. Mycologia 2003, 95, 1078–1099. Available online: https://www.researchgate.net/publication/49674865 (accessed on 13 April 2023). [CrossRef]
- Nadyeina, O.; Grube, M.; Mayrhofer, H. A contribution to the taxonomy of the genus Rinodina (Physciaceae, lichenized Ascomycotina) using combined ITS and mtSSU rDNA data. Lichenologist 2010, 42, 521–531. Available online: http://europepmc.org/article/PMC/3223597 (accessed on 13 April 2023). [CrossRef]
- Gaya, E.; Högnabba, F.; Holguin, A.; Molnar, K.; Fernández-Brime, S.; Stenroos, S.; Arup, U.; Søchting, U.; den Boom, P.V.; Lücking, R.; et al. Implementing a cumulative supermatrix approach for a comprehensive phylogenetic study of the Teloschistales (Pezizomycotina, Ascomycota). Mol. Phylogenet. Evol. 2012, 63, 374–387. [Google Scholar] [CrossRef]
- Resl, P.; Mayrhofer, H.; Clayden, S.R.; Spribille, T.; Thor, G.; Tønsberg, T.; Sheard, J.W. Morphological, chemical and species delimitation analyses provide new taxonomic insights into two groups of Rinodina. Lichenologist 2016, 48, 469–488. Available online: https://www.researchgate.net/publication/308793339 (accessed on 12 April 2023). [CrossRef]
- Wen, J. Evolution of eastern Asian and eastern North American disjunct distributions of flowering plants. Annu. Rev. Eco. Evol. Syst. 1999, 30, 421–455. [Google Scholar] [CrossRef]
- Xiang, Q.-Y.; Soltis, D.E.; Soltis, P.S. The eastern Asian and eastern and western North American disjunction: Congruent phylogenetic patterns in seven diverse genera. Mol. Phylogenet. Evol. 1998, 10, 178–190. [Google Scholar] [CrossRef]
- Qian, H. Floristic relationships between eastern Asia and North America: Test of Gray’s hypothesis. Am. Nat. 2002, 160, 317–332. Available online: https://www.researchgate.net/publication/23174933 (accessed on 12 April 2023). [CrossRef]
- Ding, W.; Ree, R.; Spicer, R.; Xing, Y. Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora. Science 2020, 369, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Jatta, A. Licheni Cinesi raccolti allo Shen-si negli anni 1894-1898 dal. rev. Padre Missionario, G. Giraldi. Nuovo G. Bot. Ital. 1902, 9, 477. Available online: https://bibdigital.rjb.csic.es/viewer/12810/?offset=#page=490&viewer=picture&o=bookmark&n=0&q= (accessed on 11 April 2023).
- Paulson, R. Lichens of Mount Everest. Bot. J. Linn. Soc. Lond. 1925, 63, 189–193. [Google Scholar]
- Zahlbruckner, A. Lichenes (Übersicht über sämtliche bisher aus China bekannten Flechten). In Symbolae Sinicae: Botanische Ergebnisse der Expedition der Akademie der Wissenschaften in Wien Nach Südwest-China, 1914–1918; Handel-Mazzetti, H.F., Brotherus, V.F., Eds.; Springer: Wien, Austria, 1930; Volume 3, pp. 226–234. Available online: https://www.biodiversitylibrary.org/item/15292#page/1/mode/1up (accessed on 12 April 2023).
- Magnusson, A.H. Lichens from Central Asia, I. In Reports Scientific Expedition North-West Provinces of China (the Sino-Swedish Expedition) 13, XI, Botany, 1; Hedin, S., Ed.; Aktiebolaget Thule: Stockholm, Sweden, 1940; pp. 145–156. [Google Scholar]
- Magnusson, A.H. Lichens from Central Asia II. In Reports Scientific Expedition North-West Provinces of China (the Sino-Swedish Expedition) 22, XI, Botany, 2; Hedin, S., Ed.; Aktiebolaget Thule: Stockholm, Sweden, 1944; pp. 55–57. [Google Scholar]
- Moreau, F.; Moreau, M.F. Lichens de Chine. Rev. Bryol. Et Lichenol. 1951, 20, 183–199. [Google Scholar]
- Wang Yang, J.-R.; Lai, M.-J. A checklist of the lichens of Taiwan. Taiwania 1973, 18, 83–104. [Google Scholar]
- Wu, J.-N.; Xiang, T.A. Primary study on lichens from Yuntai Mountain, Lianyungang, Jiangsu Province. J. Nanjing Norm. Univ.-Nat. Sci. 1981, 3, 1–11. (In Chinese) [Google Scholar]
- Xu, B.S. Cryptogamic Flora of the Yangtze Delta and Adjacent Regions; Shanghai Scientific and Technical Publishers: Shanghai, China, 1989; pp. 254–255. (In Chinese) [Google Scholar]
- Abbas, A.; Jiang, Y.C.; Wu, J.N. Lichens from the Tianshan Mountains, Xinjiang, China. J. Nanjing Norm. Univ.-Nat. Sci. 1993, 16, 74–82. (In Chinese) [Google Scholar]
- Abbas, A.; Wu, J.N. Lichens of Xinjiang; Sci-Tec & Hygiene Publishing House of Xinjiang: Urumqi, China, 1998; pp. 1–178. (In Chinese) [Google Scholar]
- Aptroot, A.; Seaward, M.R.D. Annotated checklist of Hongkong Lichens. Trop. Bryol. 1999, 17, 57–101. Available online: https://www.researchgate.net/publication/265989726 (accessed on 10 April 2023).
- Aptroot, A.; Sparrius, L.B. New microlichens from Taiwan. Fungal Divers. 2003, 14, 1–50. Available online: https://www.researchgate.net/publication/216829813 (accessed on 10 April 2023).
- Obermayer, W.; Blaha, J.; Mayrhofer, H. Buellia centralis and chemotypes of Dimelaena oreina in Tibet and other Central-Asian regions. Symb. Bot. Ups. 2004, 34, 327–342. Available online: https://www.researchgate.net/publication/242552311 (accessed on 12 April 2023).
- Aptroot, A.; Saipunkaew, W.; Sipman, H.J.M.; Sparrius, L.B.; Wolseley, P.A. New lichens from Thailand, mainly microlichens from Chiang Mai. Fungal Divers. 2007, 24, 75–134. Available online: https://www.researchgate.net/publication/228515656 (accessed on 10 April 2023).
- Li, Y.; Chen, C.; Zhao, Z. A primary study on lichens from Mount Yi. J. Fungal Res. 2008, 6, 70–73. (In Chinese) [Google Scholar]
- Liu, M.; Wei, J.-C. Lichen diversity in Shapotou region of Tengger Desert, China. Mycosystema 2013, 32, 42–50. (In Chinese) [Google Scholar]
- Joshi, S.; Jayalal, U.; Oh, S.-O.; Li, X.-R.; Jia, R.-L.; Hur, J.-S. New records of lichens from Shapotou area in Ningxia of Northwest China. Mycosystema 2014, 33, 167–173. Available online: https://www.researchgate.net/publication/263655180 (accessed on 11 April 2023).
- Kondratyuk, S.Y.; Lökös, L.; Halda, J.P.; Moniri, M.H.; Farkas, E.; Park, J.S.; Lee, B.G.; Oh, S.-O.; Hur, J.-S. New and noteworthy lichen-forming and lichenicolous fungi: 5. Acta Bot. Hung. 2016, 58, 319–396. [Google Scholar] [CrossRef]
- Ren, Q.; Zheng, X. Notes on the genus Rinodina in mainland China. J. Liaocheng Univ. (Nat. Sci.) 2020, 33, 85–97. (In Chinese) [Google Scholar]
- Tursun, A.; Shahidin, H.; Tumur, A. Taxonomic study on Rinodina in Xinjiang, China. Acta Bot. Boreali-Occident. Sin. 2020, 40, 1978–1988. (In Chinese) [Google Scholar]
- Wei, J.-C. The Enumeration of Lichenized Fungi in China; China Forestry Publishing House: Beijing, China, 2020; pp. 450–454. [Google Scholar]
- Culberson, C.F. Improved conditions and new data for identification of lichen products by standardized thin-layer chromatographic method. J. Chromatogr. A 1972, 72, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens; British Lichen Society: London, UK, 2001; pp. 1–101. [Google Scholar]
- Zoller, S.; Scheidegger, C.; Sperisen, C. PCR primers for the amplification of mitochondrial small subunit ribosomal DNA of lichen-forming ascomycetes. Lichenologist 1999, 31, 511–516. Available online: https://www.researchgate.net/publication/229071472 (accessed on 12 April 2023). [CrossRef]
- Gardes, M.M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application for the identification of mycorrhizae and rust. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. Available online: https://www.researchgate.net/publication/223058289 (accessed on 12 April 2023).
- Hassouna, N.; Mithot, B.; Bachellerie, J.P. The complete nucleotide sequence of mouse 28S rRNA gene. Implications for the process of size increase of the large subunit rRNA in higher eukaryotes. Nucleic Acids Res. 1984, 12, 3563–3583. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. Available online: http://europepmc.org/article/PMC/213247 (accessed on 12 April 2023). [CrossRef]
- Zhao, X.; Zhang, L.L.; Zhao, Z.T.; Wang, W.C.; Leavitt, S.D.; Lumbsch, H.T. Molecular phylogeny of the lichen genus Lecidella focusing on species from mainland China. PLoS ONE 2015, 10, e0139405. Available online: https://www.researchgate.net/publication/283181184 (accessed on 12 April 2023). [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3603318/pdf/mst010.pdf (accessed on 11 April 2023). [CrossRef]
- Chernomor, O.A.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. Available online: http://europepmc.org/backend/ptpmcrender.fcgi?accid=PMC5066062&blobtype=pdf (accessed on 10 April 2023). [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5453245/pdf/emss-72237.pdf (accessed on 11 April 2023). [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. Available online: http://europepmc.org/backend/ptpmcrender.fcgi?accid=PMC4271533&blobtype=pdf (accessed on 11 April 2023). [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. Available online: http://europepmc.org/article/PMC/4987875 (accessed on 12 April 2023).
- Zhou, X.; Shen, X.-X.; Hittinger, C.T.; Rokas, A. Evaluating fast maximum likelihood-based phylogenetic programs using empirical phylogenomic data sets. Mol. Biol. Evol. 2017, 35, 486–530. Available online: https://academic.oup.com/mbe/article/35/2/486/4644721 (accessed on 12 April 2023). [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3329765/pdf/sys029.pdf (accessed on 12 April 2023). [CrossRef] [PubMed]
- Rambaut, A. FigTree v. 1.4.2. Institute of Evolutionary Biology, University of Edinburgh. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 12 April 2023).
- Prieto, M.; Wedin, M. Phylogeny, taxonomy and diversification events in the Caliciaceae. Fungal Divers. 2017, 82, 221–238. Available online: https://link.springer.com/article/10.1007/s13225-016-0372-y (accessed on 16 April 2023). [CrossRef]
- Tibell, L. Tholurna dissimilis and generic delimitations in Caliciaceae inferred from nuclear ITS and LSU rDNA phylogenies (Lecanorales, lichenized ascomycetes). Mycol. Res. 2003, 107, 1403–1418. [Google Scholar] [CrossRef]
- Ohmura, Y.; Sugimoto, M.; Yakovchenko, L.; Davydov, E.A. Additional species and ITS rDNA data for the lichen mycota of the Imperial Palace grounds, Tokyo, Japan. Bull. Natl. Mus. Nat. Sci. 2022, 48, 1–16. Available online: https://www.researchgate.net/publication/358952425 (accessed on 16 April 2023).
- Lohtander, K.; Ahti, T.; Stenroos, S.; Urbanavichus, G. Is Anaptychia monophyletic? A phylogenetic study based on nuclear and mitochondrial genes. Ann. Bot. Fenn. 2008, 45, 55–60. Available online: https://www.researchgate.net/publication/232273420 (accessed on 16 April 2023). [CrossRef]
- Molina, M.C.; Divakar, P.K.; González, N. Success in the isolation and axenic culture of Anaptychia ciliaris (Physciaceae, Lecanoromycetes) mycobiont. Mycoscience 2015, 56, 351–358. Available online: https://www.researchgate.net/publication/272891236 (accessed on 16 April 2023). [CrossRef]
- Wedin, M.; Baloch, E.; Grube, M. Parsimony analyses of mtSSU and nITS rDNA sequences reveal the natural relationships of the lichen families Physciaceae and Caliciaceae. Taxon 2002, 51, 655–660. Available online: https://nhm2.uio.no/botanisk/lav/RLL/PDF20/R24173.pdf (accessed on 16 April 2023). [CrossRef]
- Crespo, A.; Blanco, O.; Llimona, X.; Ferencová, Z.L.; Hawksworth, D.L. Coscinocladium, an overlooked endemic and monotypic Mediterranean lichen genus of Physciaceae, reinstated by molecular phylogenetic analysis. Taxon 2004, 53, 405–414. [Google Scholar] [CrossRef]
- Aptroot, A.; Maphangwa, K.; Zedda, L.; Tekere, M.; Alvarado, P.; Sipman, H. The phylogenetic position of Culbersonia is in the Caliciaceae (lichenized ascomycetes). Lichenologist 2019, 51, 187–191. [Google Scholar] [CrossRef]
- Schmull, M.; Miadlikowska, J.; Pelzer, M.; Stocker-Worgotter, E.; Hofstetter, V.; Fraker, E.; Hodkinson, B.P.; Reeb, V.; Kukwa, M.; Lumbsch, H.T.; et al. Phylogenetic affiliations of members of the heterogeneous lichen-forming fungi of the genus Lecidea sensu zahlbruckner (lecanoromycetes, ascomycota). Mycologia 2011, 103, 983–1003. Available online: https://www.researchgate.net/publication/51192614 (accessed on 16 April 2023). [CrossRef] [PubMed]
- Miadlikowska, J.; Kauff, F.; Hofstetter, V.; Fraker, E.; Grube, M.; Hafellner, J.; Reeb, V.; Hodkinson, B.P.; Kukwa, M.; Lücking, R.; et al. New insights into classification and evolution of the Lecanoromycetes (Pezizomycotina, Ascomycota) from phylogenetic analyses of three ribosomal RNA- and two protein-coding genes. Mycologia 2006, 98, 1088–1103. Available online: https://www.researchgate.net/publication/6344409 (accessed on 16 April 2023). [CrossRef] [PubMed]
- Grube, M.; Arup, U. Molecular and morphological evolution in the Physciaceae (Lecanorales, lichenized Ascomycotina), with special emphasis on the genus Rinodina. Lichenologist 2001, 33, 63–72. [Google Scholar] [CrossRef]
- Liu, D.; Hur, J.-S. Revision of the lichen genus Phaeophyscia and allied atranorin absent taxa (Physciaceae) in South Korea. Microorganisms 2019, 7, 242. Available online: https://www.mdpi.com/2076-2607/7/8/242 (accessed on 16 April 2023). [CrossRef]
- Kondratyuk, S.Y.; Lőkös, L.; Kim, J.A.; Jeong, M.-H.; Kondratiuk, A.; Oh, S.-O.; Hur, J.-S. Kashiwadia gen. nov. (Physciaceae, lichen-forming Ascomycota), proved by phylogenetic analysis of the Eastern Asian Physciaceae. Acta Bot. Hung. 2014, 56, 369–378. Available online: https://www.researchgate.net/publication/277924093 (accessed on 16 April 2023). [CrossRef]
- Lohtander, K.; Källersjö, M.; Moberg, R.; Tehler, A. The family Physciaceae in Fennoscandia: Phylogeny inferred from ITS sequences. Mycologia 2000, 92, 728–735. Available online: https://www.researchgate.net/publication/232273357 (accessed on 16 April 2023). [CrossRef]
- Kondratyuk, S.Y.; Lőkös, L.; Kim, J.A.; Jeong, M.-H.; Zarei-Darki, B.; Hur, J.-S. Oxnerella safavidiorum gen. et spec. nov. (Lecanoromycetidae, Ascomycota) from Iran (Asia) proved by phylogenetic analysis. Acta Bot. Hung. 2014, 56, 379–398. Available online: https://go.gale.com/ps/i.do?p=AONE&u=googlescholar&id=GALE|A458953398&v=2.1&it=r&asid=e883ca1e (accessed on 16 April 2023). [CrossRef]
- Marthinsen, G.; Rui, S.; Timdal, E. OLICH: A reference library of DNA barcodes for Nordic lichens. Biodivers. Data J. 2019, 7, e36252. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6711938/ (accessed on 16 April 2023). [CrossRef]
- Zhong, Q.; Zhang, Y.; Wang, X.; Timdal, E.; Gong, H.; Wang, Z.; Wang, L. Phaeorrhiza (Physciaceae), a new lichen genus record to China. Phytotaxa 2021, 510, 228–238. [Google Scholar] [CrossRef]
- Yang, M.-X.; Wang, X.-Y.; Liu, D.; Zhang, Y.-Y.; Li, L.-J.; Yin, A.-C.; Scheidegger, C.; Wang, L.-S. New species and records of Pyxine (Caliciaceae) in China. MycoKeys 2019, 45, 93–109. Available online: https://mycokeys.pensoft.net/article/29374/ (accessed on 16 April 2023). [CrossRef] [PubMed]
- Sipman, H.; Aptroot, A. Ikaeria serusiauxii, a new Caloplaca-like lichen from Macaronesia and mainland Portugal, with a lichen checklist for Porto Santo. Plant Fungal Syst. 2020, 65, 120–130. Available online: https://www.semanticscholar.org/paper/02f5c28f8a50f47d97994584d5283511e0c4c6d8 (accessed on 16 April 2023). [CrossRef]
- Kaschik, M. Taxonomic Studies on Saxicolous Species of the Genus Rinodina (Lichenized Ascomycetes, Physciaceae) in the Southern Hemisphere with Emphasis in Australia and New Zealand; Bibliotheca Lichenologica: Stuttgart, Germany, 2006; pp. 1–162. [Google Scholar]
- Lee, B.G.; Hur, J.-S. Two new Rinodina lichens from South Korea, with an updated key to the species of Rinodina in the far eastern Asia. Mycokeys 2022, 87, 159–182. Available online: https://europepmc.org/article/PMC/PMC8891229 (accessed on 16 April 2023). [CrossRef] [PubMed]
- Bhattacharya, D.; Friedl, T.; Helms, G. Vertical evolution and intragenic spread of lichen-fungal group I introns. J. Mol. Evol. 2002, 55, 74–84. [Google Scholar] [CrossRef]
- Abbas, A.; Wu, J.N. The lichens from Tielimaiti Pass Kuqa County, Xinjiang, China. Arid Zone Res. 1994, 11, 19–23. (In Chinese) [Google Scholar]
Figure 1.
Phylogenetic tree generated from maximum likelihood (ML) analysis based on combined nrITS, mtSSU, and nrLSU sequences. SH-aLRT support (%) for ML greater than 80%, ultrafast bootstrap support (%) greater than 95%, and Bayesian posterior probabilities (PPs) greater than 0.95 are given above the nodes. Newly generated sequences are indicated in bold.
Figure 1.
Phylogenetic tree generated from maximum likelihood (ML) analysis based on combined nrITS, mtSSU, and nrLSU sequences. SH-aLRT support (%) for ML greater than 80%, ultrafast bootstrap support (%) greater than 95%, and Bayesian posterior probabilities (PPs) greater than 0.95 are given above the nodes. Newly generated sequences are indicated in bold.

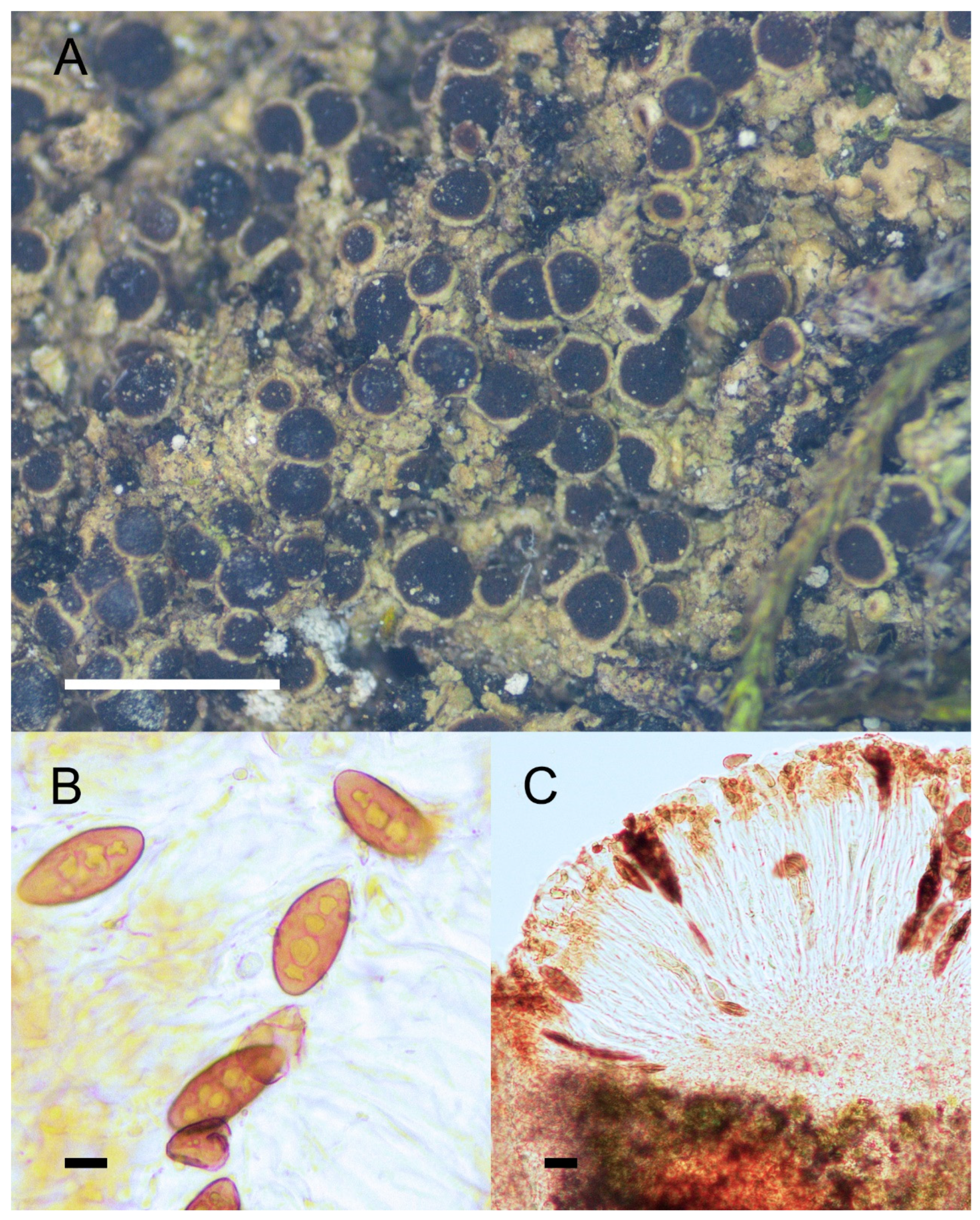
Figure 2.
Morphology and anatomy of Rinodina cornutula. (A) Thallus and apothecia. (B) Mischoblastia-type ascospores. (C) Section of apothecia. Scale bars: 2 mm (A); 10 μm (B); 20 μm (C).
Figure 2.
Morphology and anatomy of Rinodina cornutula. (A) Thallus and apothecia. (B) Mischoblastia-type ascospores. (C) Section of apothecia. Scale bars: 2 mm (A); 10 μm (B); 20 μm (C).

Figure 3.
Morphology and anatomy of Rinodina globulans. (A) Thallus and apothecia. (B) Pachysporaria-type ascospores. (C) Section of apothecia. Scale bars: 2 mm (A); 10 μm (B); 20 μm (C).
Figure 3.
Morphology and anatomy of Rinodina globulans. (A) Thallus and apothecia. (B) Pachysporaria-type ascospores. (C) Section of apothecia. Scale bars: 2 mm (A); 10 μm (B); 20 μm (C).

Figure 4.
Morphology and anatomy of Rinodina handelii. (A) Thallus and apothecia. (B) Mischoblastia-type ascospores. (C) Section of apothecia. Scale bars: 1 mm (A); 10 μm (B); 20 μm (C).
Figure 4.
Morphology and anatomy of Rinodina handelii. (A) Thallus and apothecia. (B) Mischoblastia-type ascospores. (C) Section of apothecia. Scale bars: 1 mm (A); 10 μm (B); 20 μm (C).

Figure 5.
Morphology and anatomy of Rinodina pluriloculata. (A) Thallus and apothecia. (B) Submuriform-type ascospores. (C) Section of apothecia. Scale bars: 2 mm (A); 10 μm (B); 20 μm (C).
Figure 5.
Morphology and anatomy of Rinodina pluriloculata. (A) Thallus and apothecia. (B) Submuriform-type ascospores. (C) Section of apothecia. Scale bars: 2 mm (A); 10 μm (B); 20 μm (C).

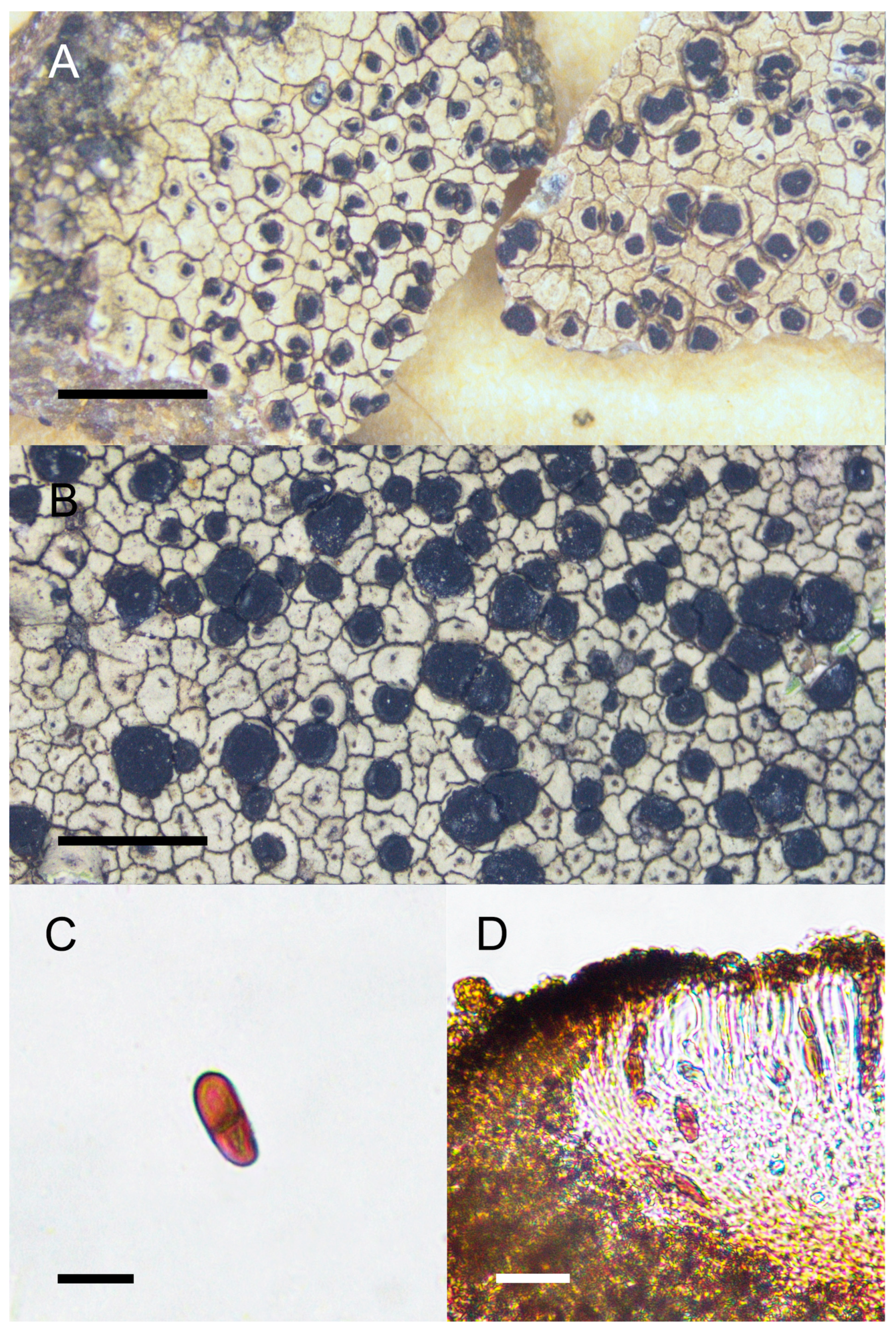
Figure 6.
Morphology and anatomy of Buellia setschwana. (A) Thallus and apothecia of the holotype (W-1171). (B–D) Specimen collected from the type locality (KUN 15-47744). (B) Thallus and apothecia. (C) Buellia-type of ascospores. (D) Section of apothecia. Scale bars: 2 mm (A,B); 10 μm (C); 20 μm (D).
Figure 6.
Morphology and anatomy of Buellia setschwana. (A) Thallus and apothecia of the holotype (W-1171). (B–D) Specimen collected from the type locality (KUN 15-47744). (B) Thallus and apothecia. (C) Buellia-type of ascospores. (D) Section of apothecia. Scale bars: 2 mm (A,B); 10 μm (C); 20 μm (D).

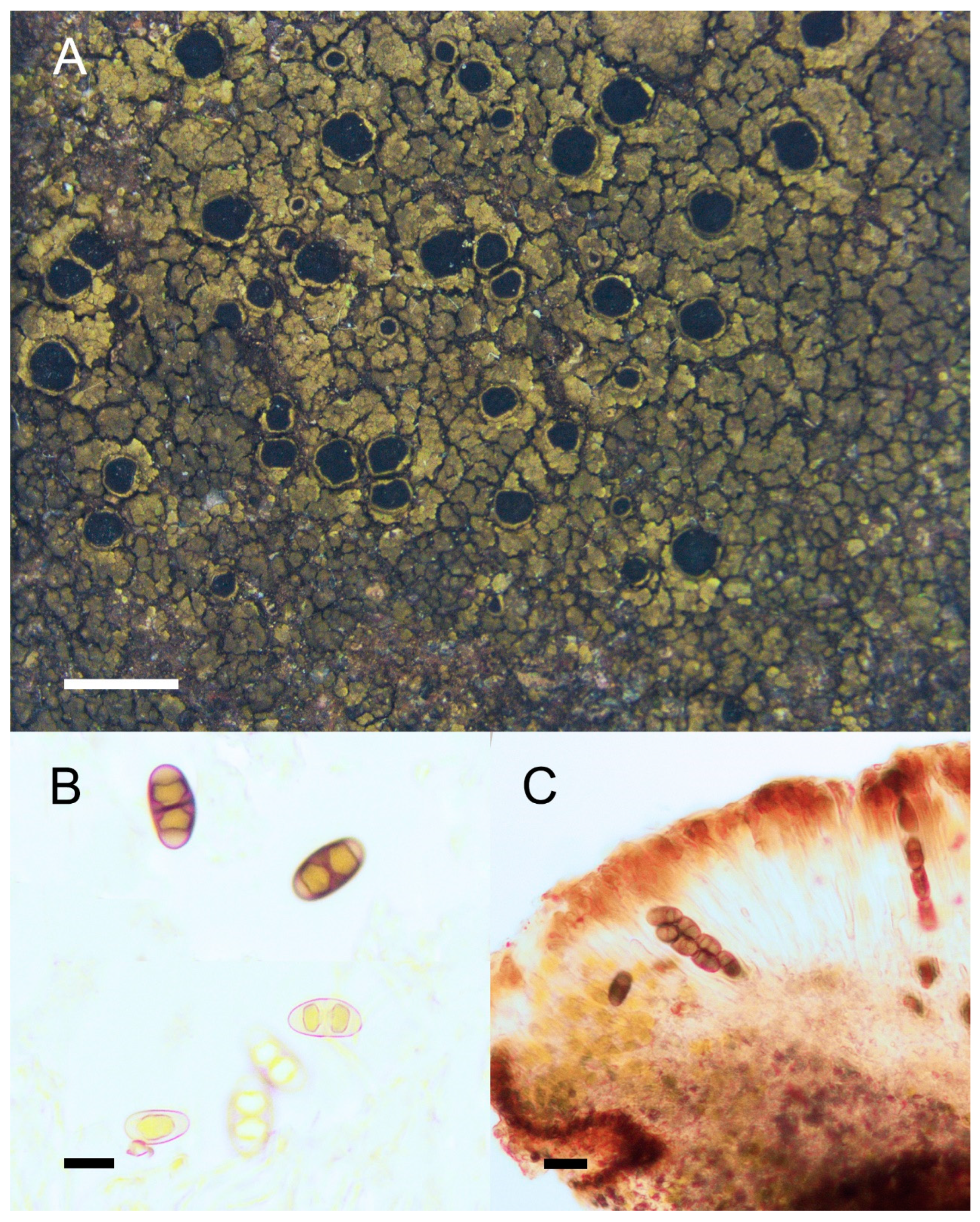
Figure 7.
Morphology and anatomy of Rinodina hengduanensis. (A) Thallus and apothecia. (B) Dirinaria-type ascospores. (C) Section of apothecia. Scale bars: 2 mm (A); 10 μm (B); 20 μm (C).
Figure 7.
Morphology and anatomy of Rinodina hengduanensis. (A) Thallus and apothecia. (B) Dirinaria-type ascospores. (C) Section of apothecia. Scale bars: 2 mm (A); 10 μm (B); 20 μm (C).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Specimens and sequences used for phylogenetic analyses. Newly generated sequences are in bold.
Table 1.
Specimens and sequences used for phylogenetic analyses. Newly generated sequences are in bold.
| Species | Locality | Voucher Specimens | GenBank Numbers | References | ||
|---|---|---|---|---|---|---|
| nrITS | mtSSU | nrLSU | ||||
| Acolium karelicum | unknown | UPS Hermansson 16472 | KX512897 | na | KX512879 | [56] |
| Acroscyphus sphaerophoroides 1 | unknown | UPS Obermayer 6077 | KX512898 | KX512984 | na | [56] |
| A. sphaerophoroides 2 | unknown | TNS Shimizu | AY450562 | na | na | [57] |
| Allocalicium adaequatum | UPS Spribille 14143 | KX512906 | KX512986 | KX512859 | [56] | |
| Amandinea punctata 1 | Japan, Tokyo | TNS YO6843 | LC669587 | na | na | [58] |
| A. punctata 2 | Japan, Tokyo | TNS YO6855 | LC669588 | na | na | [58] |
| Anaptychia bryorum | Bhutan, Thimphu | Soehting 8378 | EF582777 | EF582825 | na | [59] |
| A. ciliaris | Spain | MAF-Lich 17758 | KC559095 | na | na | [60] |
| Baculifera remensa | S Prieto | na | KX512962 | KX512881 | [56] | |
| Buellia disciformis 1 | Jämtland, Klaxåsen | Wedin 6155 | AY143392 | AY143401 | na | [61] |
| B. disciformis 2 | Sweden | UPS A. Nordin 4429 | AF540498 | na | na | [9] |
| B. elegans | unknown | S Hansen | KX512901 | KX512993 | na | [56] |
| B. erubescens | unknown | S Wetmore 95879 | KX512902 | KX512969 | KX512874 | [56] |
| B. frigida | unknown | S Westberg | KX512903 | KX512992 | KX512852 | [56] |
| B. polita 1 | China, Sichuan | KUN-L XY20-341 | OP526814 | OP526788 | OP526772 | |
| B. polita 2 | China, Yunnan | KUN-L 15-47741 | OP526792 | OP526774 | OP526763 | |
| B. polita 3 | China, Sichuan | KUN-L 19-62813 | OP526800 | OP526780 | OP526768 | |
| B. polita 4 | China | KUN-L 14-43546 | MN615679 | na | na | |
| B. polita 5 | China | KUN-L 15-48012 | MN615678 | na | na | |
| B. polita 6 | China | KUN-L 13-41328 | MN615677 | na | na | |
| B. setschwana 1 | China, Yunnan | KUN-L 15-48032 | OP526796 | na | na | |
| B. setschwana 2 | China, Yunnan | KUN-L 15-47744 | OP526794 | na | na | |
| B. setschwana 3 | China, Yunnan | KUN-L 15-47743 | OP526793 | OP526775 | OP526764 | |
| B. tesserata | unknown | S Tehler 7323 | KX512904 | na | KX512885 | [56] |
| Calicium abietinum | unknown | UPS Tibell 25061 | KX512905 | KX512971 | KX512872 | [56] |
| C. corynellum | unknown | S Prieto | KX512908 | KX512985 | KX512855 | [56] |
| Coscinocladium gaditanum 1 | Spain | MAF 9855 | AY449720 | AY464073 | na | [62] |
| C. gaditanum 2 | Spain | MAF 9856 | AY449721 | AY464074 | na | [62] |
| Culbersonia nubila 1 | South Africa | ALV14224 | MH121317 | na | MH121319 | [63] |
| C. nubila 2 | South Africa | ALV14225 | MH121318 | na | MH121320 | [63] |
| Dimelaena oreina | unknown | S Lendemer 4193 | KX512922 | KX512976 | KX512867 | [56] |
| Diplotomma alboatrum | unknown | S Prieto 3034 | KX512924 | KX512966 | KX512877 | [56] |
| Heterodermia diademata | Bolivia, Depto. Beni | B K. Bach, M. Kesseler and Portugal 389 | AF540518 | na | na | [9] |
| H. vulgaris | unknown | DUKE AFTOL-320 | HQ650704 | na | DQ883798 | [64,65] |
| Hyperphyscia adglutinata | unknown | BCN-Lich 17031 | AF250795 | GU247189 | na | [66] |
| H. crocata | South Korea | 120413 | MN150490 | na | na | [67] |
| Kashiwadia orientalis 1 | Eastern Asia | Hur 040044 | na | KM397366 | na | [68] |
| K. orientalis 2 | Eastern Asia | Hur 040164 | na | KM397365 | na | [68] |
| Mobergia calculiformis 1 | Mexico | Moberg 10412 | AF224359 | na | na | [69] |
| M. calculiformis 2 | unknown | M231 | AF250796 | na | na | [66] |
| Oxnerella safavidiorum | Iran | KW-L 70300 | KM410153 | KM410156 | na | [70] |
| Phaeophyscia endococcinodes | South Korea | 130163 | MN150503 | na | na | [67] |
| P. sciastra | Norway, Sor-Trondelag | O-L-196352 | MK812372 | na | na | [71] |
| Phaeorrhiza nimbosa | China, Xizang | KUN-L 19-65695 | MW133637 | MW133652 | MW133660 | [72] |
| P. sareptana | China, Gansu | KUN-L 18-59809 | MW133625 | MW133640 | MW133654 | [72] |
| Physcia adscendens | Norway, Vestfold | O-L-198947 | MK812201 | na | na | [71] |
| P. tenella 1 | Gästrikland | S Odelvik and Hellstrom 0827 | KX512932 | KX512974 | KX512869 | [56] |
| P. tenella 2 | Finland | Lohtander 650 | AF224425 | EF582800 | na | [59] |
| Physciella chloantha 1 | Iberian Peninsula | BCN-Lich 15525 | GU247166 | GU247200 | na | |
| P. chloantha 2 | Iberian Peninsula | BCN-Lich 17033 | GU247164 | GU247198 | na | |
| Physconia enteroxantha | Norway, Akershus | O-L-196328 | MK812142 | na | na | [71] |
| P. grisea | Morocco, Imli | Staro192 | LS483208 | na | na | [71] |
| Pseudothelomma ocellatum 1 | unknown | S Tehler 8063 | KX512934 | KX512957 | KX512862 | [56] |
| P. ocellatum 2 | unknown | UPS Hermansson 18662 | KX512935 | KX512952 | KX512891 | [56] |
| Pyxine himalayensis | China, Yunnan | KUN-L 12-36055 | KY611881 | KY751388 | na | [73] |
| P. subcinerea | China, Taiwan | KUN-L 15-49012 | KY611867 | KY751374 | na | [73] |
| Rinodina cana | unknown | Sipman 63008 | MN587029 | na | na | [74] |
| R. conradii | China, Yunnan | KUN-L 13-40531 | OP526791 | OP526773 | na | |
| R. cornutula 1 | China, Yunnan | KUN-L XY20-3571 | OP526815 | OP526789 | na | |
| R. cornutula 2 | China, Yunnan | KUN-L XY20-3572 | OP526816 | OP526790 | na | |
| R. gennarii | Netherlands, Utrecht | B H. Sipman 44435 | AJ544187 | na | na | [9] |
| R. globulans 1 | China, Yunnan | KUN-L 15-47871 | OP526795 | OP526776 | na | |
| R. globulans 2 | China, Yunnan | KUN-L XY20-171 | OP526807 | OP526785 | OP526770 | |
| R. globulans 3 | China, Yunnan | KUN-L XY20-117 | OP526805 | na | na | |
| R. globulans 4 | China, Sichuan | KUN-L XY20-279 | OP526810 | na | na | |
| R. globulans 5 | China, Sichuan | KUN-L XY20-280 | OP526811 | na | na | |
| R. handelii 1 | China, Yunnan | KUN-L XY20-224 | OP526808 | na | na | |
| R. handelii 2 | China, Yunnan | KUN-L XY20-238-1 | OP526809 | na | na | |
| R. mniaroea 1 | USA, Idaho | GZU Spribille 15242 | KX015687 | KX015706 | na | [12] |
| R. mniaroea 2 | USA, Montana | GZU Spribille 20391 | KX015692 | KX015711 | na | [12] |
| R. mniaroea 3 | Norway, Svalbard | TROM_L_565871 | MK812098 | na | na | [71] |
| R. mniaroea 4 | unknown | M249 | AF250811 | na | na | [66] |
| R. mniaroeiza | Canada | GZU V. Wagner 15.07.06/1 | KX015691 | KX015710 | na | [12] |
| R. moziana 1 | Australia, Queensland | GZU H. Mayrhofer 11742 | DQ849306 | na | na | [75] |
| R. moziana 2 | New Zealand, Nelson | GZU M. Lambauer 0214 | DQ849305 | na | na | [75] |
| R. oxydata 1 | Australia, Queensland | GZU H. Mayrhofer 11790 | DQ849311 | na | na | [75] |
| R. oxydata 2 | Australia, Queensland | GZU H. Mayrhofer 11406 | DQ849313 | na | na | [75] |
| R. oxydata 3 | New Zealand, Nelson | GZU M. Lambauer 0206 | DQ849310 | na | na | [75] |
| R. oxydata 4 | Austria, Steiermark | GZU H. Mayrhofer 13.930 | AF540548 | na | na | [9] |
| R. oxydata 5 | Austria, Styria | GZU H. Mayrhofer 15761 | DQ849312 | na | na | [75] |
| R. pluriloculata 1 | China, Yunnan | KUN-L XY20-162 | OP526806 | OP526784 | na | |
| R. pluriloculata 2 | China, Yunnan | KUN-L 20-66417 | OP526802 | OP526782 | na | |
| R. sophodes 1 | Austria, Styria | GZU 000272661 | GU553304 | GU553321 | na | [10] |
| R. sophodes 2 | Austria, Steiermark | GZU P. Bilovitz 968 | AF540550 | na | na | [9] |
| R. orientalis 1 | South Korea | KBA BDNA-L-0000284 | MW832807 | na | na | [76] |
| R. orientalis 2 | South Korea | KBA BDNA-L-0000653 | MW832808 | na | na | [76] |
| R. salicis 1 | South Korea | KBA BDNA-L-0000558 | MW832810 | na | na | [76] |
| R. salicis 2 | South Korea | KBA BDNA-L-0000560 | MW832811 | na | na | [76] |
| R. hengduanensis 1 | China, Sichuan | KUN-L 20-66506 | OP526803 | OP526783 | na | |
| R. hengduanensis 2 | China, Yunnan | KUN-L XY20-44 | OP526804 | na | na | |
| R. hengduanensis 3 | China, Sichuan | KUN-L XY20-290 | OP526813 | OP526787 | na | |
| R. hengduanensis 4 | China, Sichuan | KUN-L XY20-287 | OP526812 | OP526786 | OP526771 | |
| R. zeorina 1 | South Korea | KBA BDNA-L-0000642 | MW832812 | na | na | [76] |
| R. zeorina 2 | South Korea | KBA BDNA-L-0000646 | MW832813 | na | na | [76] |
| Rinodinella controversa 1 | unknown | M281 | AF250814 | na | na | [66] |
| R. controversa 2 | Greece, Kreta | GZU Mayrhofer and Ertl 13.74 | AJ421423 | na | na | [77] |
| Tetramelas pulverulentus | unknown | UPS Nordin 6368 | KX512940 | KX512983 | KX512860 | [56] |
| Texosporium sancti-jacobi | unknown | UPS Rosentreter and De Bolt 6514 | KX512941 | KX512981 | KX512863 | [56] |
| Thelomma mammosum | unknown | UPS Tibell 23775 | KX512942 | KX512954 | KX512888 | [56] |
| T. santessonii | unknown | UPS Nordin 4011 | KX512944 | KX512951 | KX512889 | [56] |
| Tholurna dissimilis | unknown | UPS Wedin 6330 | AY143397 | AY143407 | KX512893 | [56] |
| Tornabea scutellifera 1 | Spain | AFTOL-ID 1061 | JQ301698 | na | DQ973037 | [11,65] |
| T. scutellifera 2 | unknown | UPS Tibell 23833 | KX512946 | KX512970 | KX512873 | [56] |
| Xanthoria aureola | Sweden, Bohuslän | E. Gaya etc. (BCN) | JQ301690 | JQ301526 | JQ301585 | [12] |
| Xanthoria parietina | Sweden, Bohuslän | E. Gaya etc. (BCN) | JQ301691 | JQ301530 | JQ301589 | [12] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhong, Q.; Ai, M.; Worthy, F.R.; Yin, A.; Jiang, Y.; Wang, L.; Wang, X. Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species. Diversity 2023, 15, 705. https://doi.org/10.3390/d15060705
AMA Style
Zhong Q, Ai M, Worthy FR, Yin A, Jiang Y, Wang L, Wang X. Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species. Diversity. 2023; 15(6):705. https://doi.org/10.3390/d15060705
Chicago/Turabian StyleZhong, Qiuyi, Min Ai, Fiona Ruth Worthy, Ancheng Yin, Yi Jiang, Lisong Wang, and Xinyu Wang. 2023. "Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species" Diversity 15, no. 6: 705. https://doi.org/10.3390/d15060705
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.