
Subterranean Fauna of the Lukina Jama–Trojama Cave System in Croatia: The Deepest Cave in the Dinaric Karst
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Study Area
2.1. Northern Velebit
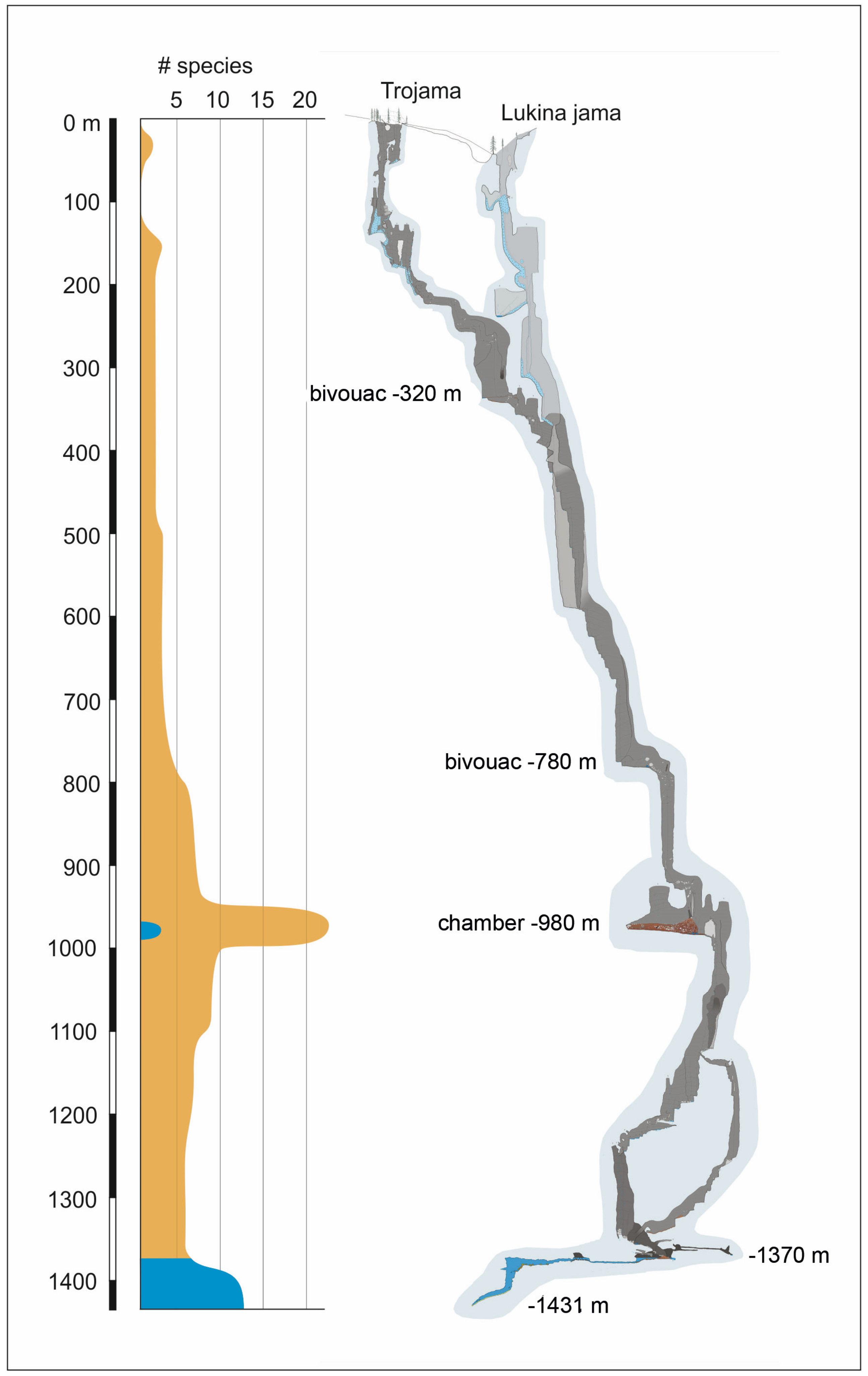
2.2. The Lukina Jama, Trojama Cave System
3. Materials and Methods
4. Results
4.1. Checklist of the Lukina Jama–Trojama Cave System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Group | Taxon | Ecology | Depth (m) | Distribution | Source |
|---|---|---|---|---|---|
| Porifera: Spongillidae | Eunapius subterraneus Sket and Velikonja, 1984 [39] | Sb | >1370 | NW Dinarides | [32] |
| Gastropoda: Hydrobiidae | Hauffenia sp. | Sb | >1370 | Velebit Mt. | [22] |
| Gastropoda: Hydrobiidae | Lanzaia sp. | Sb | >1370 | Velebit Mt. | [22] |
| Gastropoda: Hydrobiidae | Sadleriana sp. | Sb | >1370 | Velebit Mt. | RS coll. |
| Gastropoda: Acroloxidae | Acroloxus sp. | Sb? | >1370 | NA | RS coll. |
| Gastropoda: Carychiidae | Zospeum isselianum Pollonera, 1887 [40] | Tb | 800–1370 | NW Dinarides | RS coll. |
| Gastropoda: Carychiidae | Zospeum tholussum Weigand, 2013 [31] # | Tb | 980 | northern Velebit | [31] |
| Gastropoda: Carychiidae | Zospeum subobesum Bole, 1974 [41] | Tb | 800–1370 | Dinarides | [22] |
| Gastropoda: Carychiidae | Zospeum robustum Inäbnit, Jochum & Neubert, 2019 [42] | Tb | 800–1370 | NW Dinarides | [42] |
| Bivalvia: Dreissenidae | Congeria jalzici Morton and Bilandžija, 2013 [43] | Sb | >1370 | NW Dinarides | [43] |
| Polychaeta: Serpulidae | Marifugia cavatica Absolon and Hrabe, 1930 [44] | Sb | >1370 | Dinarides | [22] |
| Clitellata: Erpobdellidae | Croatobranchus mestrovi Kerovec, Kučinić and Jalžić, 1999 [30] # | Sb | 1370 | northern Velebit | [30] |
| Clitellata: Erpobdellidae | Dina. sp. * | Sb | 980 | northern Velebit | SubBio Lab coll. |
| Clitellata: Haplotaxidae | Haplotaxis cf. H. gordioides (Hartmann, in Oken 1819) [45] | Sp | 980 | Holarctic | CBSS coll. |
| Palpigradi: Eukoeneniidae | Eukoenenia sp. * | Tb | 980 | northern Velebit | RO coll. |
| Acari: Rhagidiidae | Rhagidia sp. * | Tb | 980 | northern Velebit | [32] |
| Acari: Labidostommatidae | Nicoletiella sp. * | Tb | 980 | northern Velebit | [32] |
| Araneae: Dysderidae | cf. Stalita sp. * | Tb | 980 | Velebit Mt. | CBSS coll. |
| Pseudoscorpiones: Neobisiidae | Neobisium sp. * | Tb | 980 | northern Velebit | [32] |
| Opiliones: Nemastomatidae | Hadzinia sp. * | Tb | 980 | northern Velebit | RO coll. |
| Opiliones: Sironidae | Cyphophthalmus sp. * | Tb | 980 | northern Velebit | RO coll. |
| Isopoda: Trichoniscidae | Androniscus sp. | Tp | 30 | NA | CBSS coll. |
| Isopoda: Trichoniscidae | Alpioniscus velebiticus Bedek and Taiti, 2019 [46] | Tb | 980–1370 | Velebit | [46] |
| Isopoda: Trichoniscidae | Gen. * sp. * | Tb | 800–980 | Velebit | CBSS coll. |
| Isopoda: Asellidae | Proasellus cf. P. slovenicus (Sket, 1957) [47] | Sb | >1370 | NW Dinarides | SubBio Lab coll. |
| Amphipoda: Niphargidae | Niphargus arbiter Karaman, 1984 [48] | Sb | >1370 | Velebit Mt. and Lika | [32] |
| Amphipoda: Niphargidae | Niphargus brevirostris Sket, 1971 [49] | Sb | >1370 | Velebit Mt. and Lika | [32] |
| Amphipoda: Niphargidae | Niphargus croaticus Jurinac, 1887 [50] | Sb | >1370 | NW Dinarides | [22] |
| Amphipoda: Niphargidae | Niphargus sp. * | Sb | >1370 | Lukina jama–Trojama | CBSS coll. & SubBio Lab coll. |
| Amphipoda: Niphargidae | Chaetoniphargus lubuskensis Karaman G.S. and Sket, 2019 [51] | Sb | 980 | northern Velebit | [51] |
| Decapoda: Atyidae | Troglocaris cf. T. kapelana Sket and Zakšek, 2009 [52] | Sb | >1370 | Velebit Mt. and Lika | SubBio Lab coll. |
| Diplopoda: Polydesmidae | Brachydesmus sp. | Tp | 30 | NA | [32] |
| Diplopoda: Anthogonidae | Haasia stenopodium (Strasser, 1966) [53] | Tb | 500–1370 | NW Dinarides | [22] |
| Chilopoda: Geophilidae | Geophilus hadesi Stoev, Akkari, Komerički, Edgecombe and Bonato 2015 [32] | Tb | 980–1100 | Velebit Mt. | [32] |
| Collembola: Onychiuridae | Gen. sp. * | Tb | 980–1370 | northern Velebit | CBSS coll. |
| Collembola: Oncopoduridae | Oncopodura sp. * | Tb | 980 | Velebit Mt. | CBSS coll. |
| Collembola: Isotomidae | Parisotoma sp. * | Tb | 980 | northern Velebit | CBSS coll. |
| Collembola: Isotomidae | Gen. * sp. * | Tb | 980 | Lukina jama–Trojama | [32] |
| Collembola: Sminthuridae | Disparrhopalites sp. * | Tb | 980 | Velebit Mt. | [32] |
| Diplura: Campodeidae | Plusiocampa sp. | Tb | 1370 | northern Velebit | CBSS coll. |
| Coleoptera: Cholevidae | Astagobius angustatus (Schmidt, 1852) [54] | Tb | 150–800 | NW Dinarides | [22] |
| Coleoptera: Cholevidae | Spelaeodromus pluto (Reitter, 1881) [55] | Tb | 150–800 | Velebit Mt. and Lika | [32] |
| Coleoptera: Cholevidae | Velebitodromus smidai Casale, Giachino and Jalžić 2004 [56] | Tb | 860–1200 | northern Velebit | [56] |
| Diptera: Chironomidae | Troglocladius hajdi Andersen, Baranov and Hagenlund, 2016 [33] # | Tb | 800–980 | Lukina jama–Trojama | [33] |
| Diptera: Mycetophilidae | Speolepta leptogaster (Winnertz, 1863) [57] | Tp | 980 | Europe | CBSS coll. |
4.2. The Fauna
5. Threats and Conservation
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats, 2nd ed.; Oxford University Press: Oxford, UK, 2019; p. 301. ISBN 978-0-19-882076-5. [Google Scholar]
- Ficetola, G.F.; Canedoli, C.; Stoch, F. The Racovitzan Impediment and the Hidden Biodiversity of Unexplored Environments. Conserv. Biol. 2019, 33, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Polak, S.; Pipan, T. The Subterranean Fauna of Križna jama, Slovenia. Diversity 2021, 13, 210. [Google Scholar] [CrossRef]
- Zagmajster, M.; Polak, S.; Fišer, C. Postojna-Planina Cave System in Slovenia, a Hotspot of Subterranean Biodiversity and a Cradle of Speleobiology. Diversity 2021, 13, 271. [Google Scholar] [CrossRef]
- Ozimec, R.; Baković, N.; Bakšić, D.; Basara, D.; Bevanda, L.; Brajković, H.; Brancelj, A.; Erhard, C.; Gašić, Z.; Grego, J.; et al. Vjetrenica—Cave Biodiversity Hotspot of the Dinarides; Ozimec, R., Ed.; Public Enterprise Vjetrenica: Ravno, Bosnia and Herzegovina; ADIPA: Zagreb, Croatia, 2021; p. 356. [Google Scholar]
- Bregović, P.; Zagmajster, M. Understanding Hotspots within a Global Hotspot—Identifying the Drivers of Regional Species Richness Patterns in Terrestrial Subterranean Habitats. Insect Conserv. Divers. 2016, 9, 268–281. [Google Scholar] [CrossRef]
- Reis-Venâncio, P.C.; Rabelo, L.M.; Pellegrini, T.G.; Ferreira, R.L. From Light to Darkness: The Duality of Influence of Habitat Heterogeneity on Neotropical Terrestrial Cave Invertebrate Communities. Stud. Neotrop. Fauna Environ. 2022, 1–10. [Google Scholar] [CrossRef]
- Trontelj, P.; Borko, Š.; Delić, T. Testing the Uniqueness of Deep Terrestrial Life. Sci. Rep. 2019, 9, 15188. [Google Scholar] [CrossRef]
- Mammola, S. Finding Answers in the Dark: Caves as Models in Ecology Fifty Years after Poulson and White. Ecography 2019, 42, 1331–1351. [Google Scholar] [CrossRef]
- Antić, D.Ž.; Reip, H.S. The Millipede Genus Leucogeorgia Verhoeff, 1930 in the Caucasus, with Descriptions of Eleven New Species, Erection of a New Monotypic Genus and Notes on the Tribe Leucogeorgiini (Diplopoda: Julida: Julidae). EJT 2020, 713, 1–106. [Google Scholar] [CrossRef]
- Sendra, A.; Reboleira, A.S.P.S. The World’s Deepest Subterranean Community—Krubera-Voronja Cave (Western Caucasus). Int. J. Speleol. 2012, 41, 221–230. [Google Scholar] [CrossRef]
- Zagmajster, M.; Eme, D.; Fišer, C.; Galassi, D.; Marmonier, P.; Stoch, F.; Cornu, J.-F.; Malard, F. Geographic Variation in Range Size and Beta Diversity of Groundwater Crustaceans: Insights from Habitats with Low Thermal Seasonality: Range Size and Beta Diversity in Non-Seasonal Habitats. Glob. Ecol. Biogeogr. 2014, 23, 1135–1145. [Google Scholar] [CrossRef]
- Culver, D.C.; Deharveng, L.; Bedos, A.; Lewis, J.J.; Madden, M.; Reddell, J.R.; Sket, B.; Trontelj, P.; White, D. The Mid-Latitude Biodiversity Ridge in Terrestrial Cave Fauna. Ecography 2006, 29, 120–128. [Google Scholar] [CrossRef]
- Sket, B.; Paragamian, K.; Trontelj, P. A Census of the Obligate Subterranean Fauna of the Balkan Peninsula. In Balkan Biodiversity—Pattern and Process in the European Hotspot; Griffiths, H.I., Kryštufek, B., Reed, J.M., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 309–322. ISBN 978-90-481-6732-6. [Google Scholar]
- Sket, B. Diversity Patterns in Dinaric Karst. In Encyclopedia of Caves, 2nd ed.; White, W.B., Culver, D.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 228–238. [Google Scholar]
- Bregović, P.; Fišer, C.; Zagmajster, M. Contribution of Rare and Common Species to Subterranean Species Richness Patterns. Ecol. Evol. 2019, 9, 11606–11618. [Google Scholar] [CrossRef] [PubMed]
- Zagmajster, M.; Culver, D.C.; Sket, B. Species Richness Patterns of Obligate Subterranean Beetles (Insecta: Coleoptera) in a Global Biodiversity Hotspot—Effect of Scale and Sampling Intensity. Divers. Distrib. 2008, 14, 95–105. [Google Scholar] [CrossRef]
- Borko, Š.; Altermatt, F.; Zagmajster, M.; Fišer, C. A Hotspot of Groundwater Amphipod Diversity on a Crossroad of Evolutionary Radiations. Divers. Distrib. 2022, 28, 2765–2777. [Google Scholar] [CrossRef]
- Culver, D.C.; Deharveng, L.; Pipan, T.; Bedos, A. An Overview of Subterranean Biodiversity Hotspots. Diversity 2021, 13, 487. [Google Scholar] [CrossRef]
- Fišer, C.; Borko, Š.; Delić, T.; Kos, A.; Premate, E.; Zagmajster, M.; Zakšek, V.; Altermatt, F. The European Green Deal Misses Europe’s Subterranean Biodiversity Hotspots. Nat. Ecol. Evol. 2022, 6, 1403–1404. [Google Scholar] [CrossRef]
- Ozimec, R.; Bedek, J.; Gottstein, S.; Jalžić, B.; Slapnik, R.; Štamol, V.; Bilandžija, H.; Dražina, T.; Kletečki, E.; Komerički, A.; et al. Crvena Knjiga Špiljske Faune Hrvatske. [Red Book of Croatian Cave Dwelling Fauna]; Ozimec, R., Katušić, L., Eds.; Ministarstvo Culture: Zagreb, Croatia; Državni Zavod za Zaštitu Prirode: Zagreb, Croatia, 2009; p. 371. ISBN 978-953-7169-73-2.
- Bakšić, D.; Barišić, T.; Božić, V.; Buzjak, S.; Čaplar, A.; Jalžić, B.; Lacković, D.; Stroj, A.; Šmida, B.; Vrbek, B.; et al. Jamski Sustav Lukina Jama—Trojama; Jalžić, B., Ed.; Hrvatski Planinarski Savez: Zagreb, Croatia, 2007; p. 143. [Google Scholar]
- Glavaš, V.; Grlj, A.; Hočevar, G.; Novljan, Ž.; Stepišnik, U.; Stojilković, B.; Trenchovska, A.; Žebre, M. Dinarski Kras; Stepišnik, U., Ed.; Znanstvena Založba Filozofske Fakultete Univerze v Ljubljani: Ljubljana, Slovenia, 2018; p. 159. ISBN 978-961-06-0147-0. [Google Scholar]
- Velić, I.; Velić, J. Od Morskih Plićaka do Planine: Geološki Vodič Kroz Nacionalni Park Sjeverni Velebit (From Marine Shallows to the Mountain: Geological Guidebook through the Northern Velebit National Park); Kljajo, D., Ed.; Nacionalni Park Sjeverni Velebit: Krasno, Croatia, 2009; p. 143. [Google Scholar]
- Perica, D.; Orešić, D. Klimatska Obilježja Velebita i Njihov Utjecaj na Oblikovanje Reljefa. Senj. Zb. 1999, 26, 1–50. [Google Scholar]
- Stroj, A.; Paar, D. Water and Air Dynamics within a Deep Vadose Zone of a Karst Massif: Observations from the Lukina jama–Trojama Cave System (−1431 m) in Dinaric Karst (Croatia). Hydrol. Process. 2019, 33, 551–561. [Google Scholar] [CrossRef]
- Bakšić, D.; Paar, D.; Stroj, A.; Lacković, D. Northern Velebit Deep Caves. In Proceedings of the 16th International Congress of Speleology, Brno, Czech Republic, 21–28 July 2013; Filippi, M., Bosak, P., Eds.; Czech Speleological Society and UIS: Brno, Czech Republic, 2013; Volume 2, pp. 24–29. [Google Scholar]
- Paar, D.; Buzjak, N.; Bakšić, D.; Radolić, V. Physical Research in Croatia’s Deepest Cave System: Lukina jama-Trojama, Mt. Velebit. In Proceedings of the 16th International Congress of Speleology, Brno, Czech Republic, 21–28 July 2013; Filippi, M., Bosak, P., Eds.; Czech Speleological Society and UIS: Brno, Czech Republic, 2013; Volume 2, pp. 442–446. [Google Scholar]
- Paar, D.; Mance, D.; Stroj, A.; Pavić, M. Northern Velebit (Croatia) Karst Hydrological System: Results of a Preliminary 2H and 18O Stable Isotope Study. Geol. Croat. 2019, 72, 205–213. [Google Scholar] [CrossRef]
- Kerovec, M.; Kučinić, M.; Jalžić, B. Croatobranchus mestrovi sp. n.—Predstavnik Nove Endemske Podzemne Vrste Pijavica (Hirudinea, Erpobdellidae). Speleolog 1999, 44, 35–36. [Google Scholar]
- Weigand, A. New Zospeum Species (Gastropoda, Ellobioidea, Carychiidae) from 980 m Depth in the Lukina jama–Trojama Cave System (Velebit Mts., Croatia). Subterr. Biol. 2013, 11, 45–53. [Google Scholar] [CrossRef]
- Stoev, P.; Akkari, N.; Komerički, A.; Edgecombe, G.; Bonato, L. At the End of the Rope: Geophilus hadesi sp. n.—The World’s Deepest Cave-Dwelling Centipede (Chilopoda, Geophilomorpha, Geophilidae). ZooKeys 2015, 510, 95–114. [Google Scholar] [CrossRef] [PubMed]
- Andersen, T.; Baranov, V.; Hagenlund, L.K.; Ivković, M.; Kvifte, G.M.; Pavlek, M. Blind Flight? A New Troglobiotic Orthoclad (Diptera, Chironomidae) from the Lukina jama—Trojama Cave in Croatia. PLoS ONE 2016, 11, e0152884. [Google Scholar] [CrossRef] [PubMed]
- Mihoci, T.; Kovač Konrad, P. Ekspedicija Lukina jama—Sifon 2013. Subterranea Croat. 2014, 12, 3–12. [Google Scholar]
- Stroj, A. Hidrogeološka Istraživanja Krških Vodonosnih Sustava. Primjer Istraživanja Podzemnih Tokova u Zaleđu Priobalnih Izvora Velebitskog Kanala. Vijesti Hrvat. Geol. Druš. 2011, 48, 20–34. [Google Scholar]
- Sket, B. The Cave Hygropetric—A Little Known Habitat and Its Inhabitants. Arch. Hydrobiol. 2004, 160, 413–425. [Google Scholar] [CrossRef]
- Calado, R.; Marschal, C.; Lejeusne, C.; Sket, B.; Chevaldonné, P. Improvements to the “Sket Bottle”: A Simple Manual Device for Sampling Small Crustaceans from Marine Caves and Other Cryptic Habitats. J. Crustac. Biol. 2008, 28, 185–188. [Google Scholar] [CrossRef]
- Sket, B. Can We Agree on an Ecological Classification of Subterranean Animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Sket, B.; Velikonja, M. Troglobitic Freshwater Sponges (Porifera, Spongillidae) Found in Yugoslavia. Stygologia 1986, 2, 254–266. [Google Scholar]
- Pollonera, C. Note Malacologiche. Bull. Della Soc. Malacol. Ital. 1887, 12, 204–223. [Google Scholar]
- Bole, J. Rod Zospeum Bourguignat 1856 (Gastropoda, Ellobiidae) v Jugoslaviji. Slov. Akad. Znan. Umet. 1974, 17, 249–291. [Google Scholar]
- Inäbnit, T.; Jochum, A.; Kampschulte, M.; Martels, G.; Ruthensteiner, B.; Slapnik, R.; Nesselhauf, C.; Neubert, E. An Integrative Taxonomic Study Reveals Carychiid Microsnails of the Troglobitic Genus Zospeum in the Eastern and Dinaric Alps (Gastropoda, Ellobioidea, Carychiinae). Org. Divers. Evol. 2019, 19, 135–177. [Google Scholar] [CrossRef]
- Bilandžija, H.; Morton, B.; Podnar, M.; Ćetković, H. Evolutionary History of Relict Congeria (Bivalvia: Dreissenidae): Unearthing the Subterranean Biodiversity of the Dinaric Karst. Front. Zool. 2013, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Absolon, K.; Hrabe, S. Über Einen Neuen Süsswasser-Polychaeten Aus Den Höhlengewässern Der Herzegowina. Zool. Anz. 1930, 88, 249–264. [Google Scholar]
- Oken, L. Isis, Oder Encyclopädische Zeitung von Oken; Expedition der Isis: Wetter, Germany, 1819; Volume 4. [Google Scholar]
- Bedek, J.; Taiti, S.; Bilandžija, H.; Ristori, E.; Baratti, M. Molecular and Taxonomic Analyses in Troglobiotic Alpioniscus (Illyrionethes) Species from the Dinaric Karst (Isopoda: Trichoniscidae). Zool. J. Linn. Soc. 2019, 187, 539–584. [Google Scholar] [CrossRef]
- Sket, B. Einige Neue Formen Der Malacostraca (Crust.) Aus Jugoslawien. Bull.Sci. Beogr. 1957, 3, 70–71. [Google Scholar]
- Karaman, G.S. Contribution to the Knowledge of the Amphipoda. Revision of the Niphargus Orcinus-Group, Part I. (Fam. Niphargidae). Montenegrin Acad. Sci. Arts Glas. Sect. Nat. Sci. 1984, 4, 7–79. [Google Scholar]
- Sket, B. Vier Neue Aberrante Niphargus—Arten (Amphipoda, Gammaridae) und Einige Bemerkungen zur Taxonomie der Niphargus—Ähnlichen Gruppen. Razpr. SAZU Diss. Acad. Sci. Artium Slov. Cl IV Hist. Nat. Med. 1971, 14, 1–25. [Google Scholar]
- Jurinac, A.E. Prilog Hrvatskoj Fauni Ogulinsko-Slunjske Okolice i Pećina. Rad Jugosl. Akad. Znan. Umjet. 1887, 83, 86–128. [Google Scholar]
- Karaman, G.S.; Sket, B. New Genus and Species of the Family Niphargidae (Crustacea: Amphipoda: Senticaudata), Chaetoniphargus lubuskensis gen. nov., sp. nov. from Croatia. Zootaxa 2019, 4545, 249. [Google Scholar] [CrossRef]
- Sket, B.; Zakšek, V. European Cave Shrimp Species (Decapoda: Caridea: Atyidae), Redefined after a Phylogenetic Study; Redefinition of Some Taxa, a New Genus and Four New Troglocaris Species. Zool. J. Linn. Soc. 2009, 155, 786–818. [Google Scholar] [CrossRef]
- Strasser, K. Neue Diplopoden Aus Höhlen Jugoslawiens. Senckenberg. Biol. 1966, 47, 379–398. [Google Scholar]
- Schmidt, F. Zwei Neue Arten von Leptoderus. Stettin. Entomol. Ztg. 1852, 13, 381–382. [Google Scholar]
- Reitter, E. Neue Und Seltene Coleopteren, Im Jahre 1880 in Süddalmatien Und Montenegro Gesammelt Und Beschrieben. Dtsch. Entomol. Z. 1881, 25, 177–230. [Google Scholar] [CrossRef]
- Casale, A.; Giachino, P.M.; Jalžić, B. Three New Species and One New Genus of Ultraspecialized Cave Dwelling Leptoderinae from Croatia (Coleoptera, Cholevidae). Nat. Croat. 2004, 13, 301–317. [Google Scholar]
- Winnertz, J. Beitrag Zu Einer Monografie Der Pilzmücken (Mycetophilidae). Verh. Der Zool. -Bot. Ges. Wien 1863, 13, 637–964. [Google Scholar]
- Bilandžija, H.; Bedek, J.; Jalžić, B.; Gottstein, S. The Morphological Variability, Distribution Patterns and Endangerment in the Ogulin Cave Sponge Eunapius subterraneus Sket & Velikonja, 1984 (Demospongiae, Spongillidae). Nat. Croat. 2007, 16, 1–17. [Google Scholar]
- Simone, L.R.L.; Ferreira, R.L. Eupera troglobia sp. nov.: The First Troglobitic Bivalve from the Americas (Mollusca, Bivalvia, Sphaeriidae). Subterr. Biol. 2022, 42, 165–184. [Google Scholar] [CrossRef]
- Jalžić, B.; Cukrov, M.; Bilandžija, H. Rasprostranjenost Dinarskog Špiljskog Cjevaša Marifugia cavatica Absolon i Hrabe, 1930. Subterranea Croat. 2021, 19, 38–46. [Google Scholar]
- Matjašič, J. Marifugijska Favnula (Faune Marifugiale). In Proceedings of the Treći Jugoslavenski Speleološki Kongres, Sarajevo, Yugoslavia, 21–27 June 1962; Kanaet, T., Ed.; Speleološki Savez Jugoslavije: Sarajevo, Yugoslavia, 1962; pp. 155–156. [Google Scholar]
- Sket, B.; Dovč, P.; Jalžić, B.; Kerovec, M.; Kučinić, M.; Trontelj, P. A Cave Leech (Hirudinea, Erpobdellidae) from Croatia with Unique Morphological Features. Zool. Scr. 2001, 30, 223–229. [Google Scholar] [CrossRef]
- Hlebec, D.; Podnar, M.; Kučinić, M.; Harms, D. Molecular Analyses of Pseudoscorpions in a Subterranean Biodiversity Hotspot Reveal Cryptic Diversity and Microendemism. Sci. Rep. 2023, 13, 430. [Google Scholar] [CrossRef]
- Bregović, P.; Rožman, T.; Bedek, J.; DražIna, T.; Jalžić, B.; Komerički, A.; Lukić, M.; Pavlek, M. Biospeleološka Istraživanja Špilja i Jama Nacionalnog Parka Sjeverni Velebit u 2018. Godini. Senj. Zb. 2019, 46, 37–60. [Google Scholar] [CrossRef]
- Delić, T.; Trontelj, P.; Zakšek, V.; Fišer, C. Biotic and Abiotic Determinants of Appendage Length Evolution in a Cave Amphipod. J. Zool. 2016, 299, 42–50. [Google Scholar] [CrossRef]
- Delić, T.; Švara, V.; Coleman Charles, O.; Trontelj, P.; Fišer, C. The Giant Cryptic Amphipod Species of the Subterranean Genus Niphargus (Crustacea, Amphipoda). Zool. Scr. 2017, 46, 740–752. [Google Scholar] [CrossRef]
- Zakšek, V.; Delić, T.; Fišer, C.; Jalžić, B.; Trontelj, P. Emergence of Sympatry in a Radiation of Subterranean Amphipods. J. Biogeogr. 2019, 46, 657–669. [Google Scholar] [CrossRef]
- Borko, Š.; Trontelj, P.; Seehausen, O.; Moškrič, A.; Fišer, C. A Subterranean Adaptive Radiation of Amphipods in Europe. Nat. Commun. 2021, 12, 3688. [Google Scholar] [CrossRef] [PubMed]
- Antić, D.; Dražina, T.; Raða, T.; Tomić, V.T.; Makarov, S.E. Review of the Family Anthogonidae (Diplopoda, Chordeumatida), with Descriptions of Three New Species from the Balkan Peninsula. Zootaxa 2015, 3948, 151. [Google Scholar] [CrossRef] [PubMed]
- Thibaud, J.; Massoud, Z. Un Nouveau Genre d’Insectes Collemboles Onychiuridae Cavernicoles Des Picos de Europa (Espagne). Bull. Mus. Hist. Nat. Paris 1986, 8, 327–331. [Google Scholar] [CrossRef]
- Deharveng, L.; Thibaud, J. Bessoniella procera n. g., n. sp., Nouvel Orchesellidae Cavernicole Relictues des Pyrenees. Bull. Mus. Natl. Hist. Nat. Sect. A 1989, 11, 397–405. [Google Scholar]
- Lukić, M.; Houssin, C.; Deharveng, L. A New Relictual and Highly Troglomorphic Species of Tomoceridae (Collembola) from a Deep Croatian Cave. ZooKeys 2010, 69, 1–16. [Google Scholar] [CrossRef]
- Vargovitsh, R.S. Cave Water Walker: An Extremely Troglomorphic Troglaphorura gladiator gen. et sp. nov. (Collembola, Onychiuridae) from Snezhnaya Cave in the Caucasus. Zootaxa 2019, 4619, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Lukić, M. Collembola. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Elsevier: Oxford, UK, 2019; pp. 308–319. [Google Scholar]
- Pretner, E. Koleopterološka Fauna Pećina i Jama Hrvatske s Historijskim Pregledom Istraživanja (Fauna Coleopterologica Subterranea Croatiae mit einer Geschichtlichen Übersicht der Forschungen). Krš. Jugosl. JAZU 1973, 8, 101–239. [Google Scholar]
- Jalžić, B. Über die Verbreitung der Hohlen-Gattung Astagobius Reitter (Col., Catopidae) in Velebit Gebirge (Kroatien, Jugoslawien) mit der Beschreibung von A. angustatus vukusici ssp. nov. Acta Entomol. Jugosl. 1982, 18, 15–20. [Google Scholar]
- Polak, S. Spelaeodromus sneznikensis sp. nov. from Slovenia (Coleoptera: Cholevidae: Leptodirinae). Acta Entomol. Slov. 2002, 10, 5–12. [Google Scholar]
- Armitage, P.D. Behaviour and Ecology of Adults. In The Chironomidae: The Biology and Ecology of Non-Biting Midges; Armitage, P.D., Cranston, P.S., Pinder, L.C.V., Eds.; Chapman & Hall: London, UK, 1995; pp. 194–224. [Google Scholar]
- Ševčík, J.; Kjærandsen, J.; Marshall, S.A. Revision of Speolepta (Diptera: Mycetophilidae), with Descriptions of New Nearctic and Oriental Species. Can. Entomol. 2012, 144, 93–107. [Google Scholar] [CrossRef]
- Dörge, D.; Zaenker, S.; Klussmann-Kolb, A.; Weigand, A. Traversing Worlds—Dispersal Potential and Ecological Classification of Speolepta leptogaster (Winnertz, 1863) (Diptera, Mycetophilidae). Subterr. Biol. 2014, 13, 1–16. [Google Scholar] [CrossRef]
- HEP Hidroenergetski Sustav Senj 2. Available online: https://www.hep.hr/projekti/hidroenergetski-sustav-senj-2/247 (accessed on 29 January 2023).
- Bilandžija, H.; Puljas, S.; Čuković, T. Nacionalni Programi za Praćenje Stanja Očuvanosti Vrsta i Staništa u Hrvatskoj; Congeria kusceri Bole, 1962 i Congeria jalzici Morton & Bilandžija, 2013; Državni Zavod za Zaštitu Prirode: Zagreb, Croatia, 2014; p. 31.
- Buzjak, N.; Bočić, N.; Paar, D.; Bakšić, D.; Dubovečak, V. Ice Caves in Croatia. In Ice Caves; Elsevier: Oxford, UK, 2018; pp. 335–369. ISBN 978-0-12-811739-2. [Google Scholar]
- Ford, D.; Williams, P. Karst Hydrogeology and Geomorphology; John Wiley & Sons Ltd.: West Sussex, UK, 2007; p. 562. ISBN 978-1-118-68498-6. [Google Scholar]
- Paoletti, M.G.; Mazzon, L.; Martinez-Sañudo, I.; Simonato, M.; Beggio, M.; Dreon, A.L.; Pamio, A.; Brilli, M.; Dorigo, L.; Summers Engel, A.; et al. A Unique Midgut-Associated Bacterial Community Hosted by the Cave Beetle Cansiliella servadeii (Coleoptera: Leptodirini) Reveals Parallel Phylogenetic Divergences from Universal Gut-Specific Ancestors. BMC Microbiol. 2013, 13, 129. [Google Scholar] [CrossRef]
- Giachino, P.M.; Vailati, D. Kircheria Beroni, a New Genus and New Species of Subterranean Hygropetricolous Leptodirinae from Albania (Coleoptera, Cholevidae). Subterr. Biol. 2006, 4, 103–116. [Google Scholar]
- Polak, S.; Delić, T.; Kostanjšek, R.; Trontelj, P. Molecular Phylogeny of the Cave Beetle Genus Hadesia (Coleoptera: Leiodidae: Cholevinae: Leptodirini), with a Description of a New Species from Montenegro. Arthropod Syst. Phylogeny 2016, 74, 241–254. [Google Scholar]
- Kupriyanova, E.K.; ten Hove, H.A.; Sket, B.; Zakšek, V.; Trontelj, P.; Rouse, G.W. Evolution of the Unique Freshwater Cave-dwelling Tube Worm Marifugia cavatica (Annelida: Serpulidae). Syst. Biodivers. 2009, 7, 389–401. [Google Scholar] [CrossRef]
- Harcet, M.; Bilandžija, H.; Bruvo-Mađarić, B.; Ćetković, H. Taxonomic Position of Eunapius subterraneus (Porifera, Spongillidae) Inferred from Molecular Data—A Revised Classification Needed? Mol. Phylogenet. Evol. 2010, 54, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Kováč, L. Caves as Oligotrophic Ecosystems. In Cave Ecology; Moldovan, O.T., Kováč, Ľ., Halse, S., Eds.; Ecological Studies; Springer International Publishing: Cham, Switzerland, 2018; Volume 235, pp. 297–307. ISBN 978-3-319-98850-4. [Google Scholar]
- Culver, D.C.; Trontelj, P.; Zagmajster, M.; Pipan, T. Paving the Way for Standardized and Comparable Subterranean Biodiversity Studies. Subterr. Biol. 2013, 10, 43–50. [Google Scholar] [CrossRef]
- Deeleman-Reinhold, C.L. Beitrag Zur Kenntnis Höhlenbewohnender Dysderidae (Araneida) Aus Jugoslawien. Slov. Akad. Znan.Umet. Razred Za Prirodosl. Vede Cl. IV Hist. Nat. 1971, 14, 95–120. [Google Scholar]
- Pavlek, M.; Gauthier, J.; Tonzo, V.; Bilat, J.; Arnedo, M.A.; Alvarez, N. Life-History Traits Drive Spatial Genetic Structuring in Dinaric Cave Spiders. Front. Ecol. Evol. 2022, 10, 910084. [Google Scholar] [CrossRef]
- Beier, M. Zur Kenntnis Der Troglobionten Neobisien (Pseudoscorp.). Eos Rev. Esp. Entomol. 1931, 7, 9–23. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukić, M.; Fišer, C.; Delić, T.; Bilandžija, H.; Pavlek, M.; Komerički, A.; Dražina, T.; Jalžić, B.; Ozimec, R.; Slapnik, R.; et al. Subterranean Fauna of the Lukina Jama–Trojama Cave System in Croatia: The Deepest Cave in the Dinaric Karst. Diversity 2023, 15, 726. https://doi.org/10.3390/d15060726
Lukić M, Fišer C, Delić T, Bilandžija H, Pavlek M, Komerički A, Dražina T, Jalžić B, Ozimec R, Slapnik R, et al. Subterranean Fauna of the Lukina Jama–Trojama Cave System in Croatia: The Deepest Cave in the Dinaric Karst. Diversity. 2023; 15(6):726. https://doi.org/10.3390/d15060726
Chicago/Turabian StyleLukić, Marko, Cene Fišer, Teo Delić, Helena Bilandžija, Martina Pavlek, Ana Komerički, Tvrtko Dražina, Branko Jalžić, Roman Ozimec, Rajko Slapnik, and et al. 2023. "Subterranean Fauna of the Lukina Jama–Trojama Cave System in Croatia: The Deepest Cave in the Dinaric Karst" Diversity 15, no. 6: 726. https://doi.org/10.3390/d15060726