
Histological Investigation of the Female Gonads of Chiropsalmus quadrumanus (Cubozoa: Cnidaria) Suggests Iteroparous Reproduction
 , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Material Samples
2.2. Maturity of Specimens
2.3. Histological Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
4.1. Seasonality
4.2. Fertilization Mode and Oocyte Nutrition
4.3. Individual Size and Maturity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, A.G.; Bentlage, B.; Gillan, W.; Lynn, T.H.; Morandini, A.C.; Marques, A.C. Naming the Bonaire Banded Box Jelly, Tamoya Ohboya, n. sp. (Cnidaria: Cubozoa: Carybdeida: Tamoyidae). Zootaxa 2011, 68, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Bentlage, B.; Cartwright, P.; Yanagihara, A.A.; Lewis, C.; Richards, G.S.; Collins, A.G. Evolution of Box Jellyfish (Cnidaria: Cubozoa), a Group of Highly Toxic Invertebrates. Proc. R. Soc. B Biol. Sci. 2010, 277, 493–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, C.; Bentlage, B.; Yanagihara, A.; Gillan, W.; van Blerk, J.; Keil, D.P.; Bely, A.E.; Collins, A.G. Redescription of Alatina alata (Reynaud, 1830) (Cnidaria: Cubozoa) from Bonaire, Dutch Caribbean. Zootaxa 2013, 3737, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Baxter, E.H.; Marr, A.G.M. Sea Wasp (Chironex Fleckeri) Venom: Lethal, Haemolytic and Dermonecrotic Properties. Toxicon 1969, 7, 195–210. [Google Scholar] [CrossRef]
- Werner, B.; Cutress, C.E.; Studebaker, J.P. Life Cycle of Tripedalia cystophora, Conant (Cubomedusae). Nature 1971, 232, 655–657. [Google Scholar] [CrossRef]
- Ceh, J.; Gonzalez, J.; Pacheco, A.S.; Riascos, J.M. The Elusive Life Cycle of Scyphozoan Jellyfish—Metagenesis Revisited. Sci. Rep. 2015, 5, 12037. [Google Scholar] [CrossRef] [Green Version]
- Morandini, A.C.; Schiariti, A.; Stampar, S.N.; Maronna, M.M.; Straehler-Pohl, I.; Marques, A.C. Succession of Generations Is Still the General Paradigm for Scyphozoan Life Cycles. Bull. Mar. Sci. 2016, 92, 343–351. [Google Scholar] [CrossRef]
- Picard, M.A.L.; Vicoso, B.; Bertrand, S.; Escriva, H. Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflic. Genes 2021, 12, 1136. [Google Scholar] [CrossRef]
- Siebert, S.; Juliano, C.E. Sex, Polyps, and Medusae: Determination and Maintenance of Sex in Cnidarians. Mol. Reprod. Dev. 2017, 84, 105–119. [Google Scholar] [CrossRef]
- Eckelbarger, K.J.; Hodgson, A.N. Invertebrate Oogenesis—A Review and Synthesis: Comparative Ovarian Morphology, Accessory Cell Function and the Origins of Yolk Precursors. Invertebr. Reprod. Dev. 2021, 65, 71–140. [Google Scholar] [CrossRef]
- Arneson, A.C.; Cutress, C.E. Life History of Carybdea alata Reynaud, 1830 (Cubomedusae). In Coelenterate Ecology and Behavior; Mackie, G.O., Ed.; Springer: New York, NY, USA, 1976; pp. 227–236. [Google Scholar]
- García-Rodríguez, J.; Lewis Ames, C.; Marian, J.E.A.R.; Marques, A.C. Gonadal Histology of Box Jellyfish (Cnidaria: Cubozoa) Reveals Variation between Internal Fertilizing Species Alatina alata (Alatinidae) and Copula sivickisi (Tripedaliidae). J Morphol 2018, 279, 841–856. [Google Scholar] [CrossRef]
- Avian, M.; Bonivento, P.; Montanari, G.; Rinaldi, A.; Rottini Sandrini, L. Carybdea marsupialis L. (Cubozoa), Istologia Delle Gonadi. Biol. Mar. Suppl. Notiz. SIBM 1993, 1, 75–78. [Google Scholar]
- Hartwick, R.F. Observations on the Anatomy, Behaviour, Reproduction and Life Cycle of the Cubozoan Carybdea sivickisi. Hydrobiologia 1991, 216–217, 171–179. [Google Scholar] [CrossRef]
- Lewis, C.; Long, T.A.F. Courtship and Reproduction in Carybdea sivickisi (Cnidaria: Cubozoa). Mar. Biol. 2005, 147, 477–483. [Google Scholar] [CrossRef]
- Lewis, C.; Kubota, S.; Migotto, A.E.; Collins, A.G. Sexually Dimorphic Cubomedusa Carybdea sivickisi in Seto, Wakayama, Japan. Publ. Seto Mar. Biol. Lab. 2008, 40, 1–8. Available online: https://repository.si.edu/handle/10088/6250 (accessed on 22 June 2023).
- Garm, A.; Lebouvier, M.; Tolunay, D. Mating in the Box Jellyfish Copula sivickisi—Novel Function of Cnidocytes. J. Morphol. 2015, 276, 1055–1064. [Google Scholar] [CrossRef]
- Toshino, S.; Hiroshi, M.; Ohtsuka, S.; Okuizumi, K.; Adachi, A.; Hamatsu, Y.; Urata, M.; Nakaguchi, K.; Yamaguchi, S. Development and Polyp Formation of the Giant Box Jelly Fish Morbakka virulenta (Kishinouye, 1910) (Cnidaria: Cubozoa) Collected from the Seto Inland Sea, Western Japan. Plankton Benthos Res. Benthos Res. 2013, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tiseo, G.R.; García-Rodríguez, J.; Zara, F.J.; Ames, C.L.; Marques, A.C.; Morandini, A.C. Spermatogenesis and Gonadal Cycle in Male Tamoya haplonema and Chiropsalmus quadrumanus (Cnidaria, Cubozoa). Zool. Anz. 2019, 279, 59–67. [Google Scholar] [CrossRef]
- García-Rodríguez, J.; Ames, C.L.; Marian, J.E.A.R.; Marques, A.C. Histomorphological Comparison of Testes in Species of Box Jellyfish (Cnidaria; Cubozoa): Does Morphology Differ with Mode of Reproduction and Fertilization? Org. Divers. Evol. 2020, 20, 25–36. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Hartwick, R. Early Life History of the Sea Wasp Chironex fleckeri (Class Cubozoa). In Development and Cellular Biology of Coelenterates; Elsevier: Amsterdam, The Netherlands, 1980; pp. 11–16. [Google Scholar]
- Hartwick, R.F. Distributional Ecology and Behaviour of the Early Life Stages of the Box-Jellyfish Chironex fleckeri. Hydrobiologia 1991, 216–217, 181–188. [Google Scholar] [CrossRef]
- Stewart, S. Field Behavior of Tripedalia cystophora (Class Cubozoa). Mar Freshw. Behav. Physiol. 1996, 27, 175–188. [Google Scholar] [CrossRef]
- Carrette, T.; Straehler-Pohl, I.; Seymour, J. Early Life History of Alatina Cf. moseri Populations from Australia and Hawaii with Implications for Taxonomy (Cubozoa: Carybdeida, Alatinidae). PLoS ONE 2014, 9, e84377. [Google Scholar] [CrossRef]
- Lewis Ames, C.; Ryan, J.F.; Bely, A.E.; Cartwright, P.; Collins, A.G. A New Transcriptome and Transcriptome Profiling of Adult and Larval Tissue in the Box Jellyfish Alatina alata: An Emerging Model for Studying Venom, Vision and Sex. BMC Genom. 2016, 17, 650. [Google Scholar] [CrossRef] [Green Version]
- Hamner, W.M.; Jones, M.S.; Hamner, P.P. Swimming, Feeding, Circulation and Vision in the Australian Box Jellyfish, Chironex fleckeri (Cnidaria: Cubozoa). Mar. Freshw. Res. 1995, 46, 985–990. [Google Scholar] [CrossRef]
- Garm, A.; O’Connor, M.; Parkefelt, L.; Nilsson, D.E. Visually Guided Obstacle Avoidance in the Box Jellyfish Tripedalia cystophora and Chiropsella bronzie. J. Exp. Biol. 2007, 210, 3616–3623. [Google Scholar] [CrossRef] [Green Version]
- Berger, E.W. Physiology and Histology of the Cubomedusae, including Dr. F. S. Conant’s Notes on the Physiology; Memoirs from the Biological Laboratory of the Johns Hopkins University; The Johns Hopkins Press: Baltimore, MD, USA, 1900; Volume 4, 84p. [Google Scholar]
- Coates, M.M. Visual Ecology and Functional Morphology of Cubozoa (Cnidaria). Integr. Comp. Biol. 2003, 43, 542–548. [Google Scholar] [CrossRef]
- Werner, B. Killermedusen und Ihr Liebesspiel. Umschau 1976, 76, 80–81. [Google Scholar]
- Subramoniam, T. Mode of Reproduction: Invertebrate Animals. In Encyclopedia of Reproduction; Elsevier: Amsterdam, the Netherlands, 2018; pp. 32–40. ISBN 9780128151457. [Google Scholar]
- Werner, B. Spermatozeugmen und Paarungsverhalten Bei Tripedalia cystophora (Cubomedusae). Mar. Biol. 1973, 18, 212–217. [Google Scholar] [CrossRef]
- Helmark, S.; Garm, A. Gonadal Cnidocytes in the Cubozoan Tripedalia cystophora Conant, 1897 (Cnidaria: Cubozoa). J. Morphol. 2019, 280, 1530–1536. [Google Scholar] [CrossRef] [PubMed]
- Bordehore, C.; Fuentes, V.L.; Atienza, D.; Barberá, C.; Fernandez-Jover, D.; Roig, M.; Acevedo-Dudley, M.J.; Canepa, A.J.; Gili, J.M. Detection of an Unusual Presence of the Cubozoan Carybdea marsupialis at Shallow Beaches Located near Denia, Spain (South-Western Mediterranean). Mar. Biodivers. Rec. 2011, 4, E69. [Google Scholar] [CrossRef] [Green Version]
- Keesing, J.K.; Strzelecki, J.; Stowar, M.; Wakeford, M.; Miller, K.J.; Gershwin, L.A.; Liu, D. Abundant Box Jellyfish, Chironex sp. (Cnidaria: Cubozoa: Chirodropidae), Discovered at Depths of over 50 m on Western Australian Coastal Reefs. Sci. Rep. 2016, 6, 22290. [Google Scholar] [CrossRef] [Green Version]
- Lawley, J.W.; Ames, C.L.; Bentlage, B.; Yanagihara, A.; Goodwill, R.; Kayal, E.; Hurwitz, K.; Collins, A.G. Box Jellyfish Alatina alata Has a Circumtropical Distribution. Biol. Bull. 2016, 231, 152–169. [Google Scholar] [CrossRef] [Green Version]
- Straehler-Pohl, I.; Garm, A.; Morandini, A.C. Sexual Dimorphism in Tripedaliidae (Conant 1897) (Cnidaria, Cubozoa, Carybdeida). Zootaxa 2014, 3785, 533–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toshino, S.; Miyake, H.; Iwanaga, S. Development of Copula sivickisi (Stiasny, 1926) (Cnidaria: Cubozoa: Carybdeidae: Tripedaliidae) Collected from the Ryukyu Archipelago, Southern Japan. Plankton Benthos Res. 2014, 9, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Campbell, R.D. Cnidaria. Reprod. Mar. Invertebr. 1974, 1, 133–199. [Google Scholar]
- Hyman, L.H. Metazoa of the Tissue Grade of Construction—The Radiate Phyla—Phylum Cnidaria. In The Invertebrates: Protozoa through Ctenophora; McGraw-Hill: New York, NY, USA; London, UK, 1940; Volume 1. [Google Scholar]
- Widersten, B. Genital Organs and Fertilization in Some Scyphozoa. Zool. Bidr. Fran Upps. 1965, 37, 45–58. [Google Scholar] [CrossRef]
- Miller, R.L. Cnidaria. Reprod. Biol. Invertebr. 1983, 2, 23–73. [Google Scholar]
- Marques, A.C.; Collins, A.G. Cladistic Analysis of Medusozoa and Cnidarian Evolution. Invertebr. Biol. 2004, 123, 23–42. [Google Scholar] [CrossRef]
- Southcott, R.V. Studies on Australian Cubomedusae, Including a New Genus and Species Apparently Harmful to Man. Mar. Freshw. Res. 1955, 7, 254–280. [Google Scholar] [CrossRef]
- Mohamed, R.; Skrypzeck, H.; Gibbons, M.J. Describing Gonad Development and Gametogenesis in Southern Africa’s Endemic Box Jellyfish Carybdea branchi (Cubozoa, Carybdeidae). Afr. J. Mar. Sci. 2019, 41, 83–91. [Google Scholar] [CrossRef]
- Mayer, A.G. Medusae of the World; Publication no. 109; Carnegie Institution of Washington: Washington, DC, USA, 1910; Volume 3. [Google Scholar]
- Migotto, A.E.; Marques, A.C.; Morandini, A.C.; da Silveira, F.L. Checklist of the Cnidaria Medusozoa of Brazil. Biota Neotrop. 2002, 2, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Morandini, A.C.; Ascher, D.; Stampar, S.N.; Ferreira, J.F.V. Cubozoa e Scyphozoa (Cnidaria: Medusozoa) de Águas Costeiras Do Brasil. Iheringia Ser. Zool. 2005, 95, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, M., Jr.; Haddad, M.A. Macromedusae (Cnidaria) from the Paraná coast, southern Brazil. J. Coast. Res. 2006, 2, 1161–1164. [Google Scholar]
- Gershwin, L.A. Comments on Chiropsalmus (Cnidaria: Cubozoa: Chirodropida): A Preliminary Revision of the Chiropsalmidae, with Descriptions of Two New Genera and Two New Species. Zootaxa 2006, 42, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Nogueira Júnior, M. Article Gelatinous Zooplankton Fauna (Cnidaria, Ctenophora and Thaliacea) from Baía da Babitonga (Southern Brazil). Zootaxa 2012, 3398, 1–21. [Google Scholar]
- Guest, W.C. The Occurence of the Jelllyfish Chiropsalmus quadrumanus in Matagorda Bay, Texas. Bull. Mar. Sci. 1959, 9, 79–83. [Google Scholar]
- Kraeuter, J.N.; Setzler, E.M. The Seasonal Cycle of Scyphozoa and Cubozoa. Bull. Mar. Sci. 1975, 25, 66–74. [Google Scholar]
- Phillips, P.J.; Burke, W.D. The Occurrence of Sea Waps (Cubomedusae) in Mississipi Sound and the Northern Gulf of Mexico. Bull. Mar. Sci. 1970, 20, 853–859. [Google Scholar]
- Straehler-Pohl, I. Critical Evaluation of Characters for Species Identification in the Cubomedusa Genus Malo (Cnidaria, Cubozoa, Carybdeida, Carukiidae). Plankton Benthos Res. 2014, 9, 83–98. [Google Scholar] [CrossRef] [Green Version]
- Pitt, K.A.; Kingsford, M.J. Reproductive Biology of the Edible Jellyfish Catostylus mosaicus (Rhizostomeae). Mar. Biol. 2000, 137, 791–799. [Google Scholar] [CrossRef]
- Humason, G.L. Animal Tissue Techniques; W.H. Freeman and Company: San Francisco, CA, USA, 1962. [Google Scholar]
- Behmer, O.A.; Tolosa, E.M.C.; Freitas Neto, A.G. Manual de Tecnicas Para Histologia Normal e Patologica; Livraria Editora: São Paulo, Brazil, 1976. [Google Scholar]
- Bancroft, J.; Stevens, A. Theory and Practice of Histological Techniques; Churchill Livingstone: London, UK, 1982. [Google Scholar]
- Pearse, A.G.E. Histochemistry Theoretical and Applied. In Analytical Tehnology; Churchill Livingstone: London, UK, 1985; Volume 2. [Google Scholar]
- Junqueira, L.C. Histology Revisited—Technical Improvement Promoted by the Use of Hydrophilic Resin Embedding. Cienc. Cult. 1995, 47, 92–95. [Google Scholar]
- Mendoza-Becerril, M.A.; Marian, J.E.A.R.; Migotto, A.E.; Marques, A.C. Exoskeletons of Bougainvilliidae and Other Hydroidolina (Cnidaria, Hydrozoa): Structure and Composition. PeerJ 2017, 5, e2964. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.; Rasband, W.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2010, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Marian, J.E.A.R. Spermatophoric Reaction Reappraised: Novel Insights into the Functioning of the Loliginid Spermatophore Based on Doryteuthis Plei (Mollusca: Cephalopoda). J. Morphol. 2012, 273, 248–278. [Google Scholar] [CrossRef]
- Eckelbarger, K.J.; Larson, R. Ultrastructure of the Ovary and Oogenesis in the Jellyfish Linuche unguiculata and Stomolophus meleagris, with a Review of Ovarian Structure in the Scyphozoa. Mar. Biol. 1992, 114, 633–643. [Google Scholar] [CrossRef]
- Mammone, M.; Bosch-Belmar, M.; Milisenda, G.; Castriota, L.; Sinopoli, M.; Allegra, A.; Falautano, M.; Maggio, T.; Rossi, S.; Piraino, S. Reproductive Cycle and Gonadal Output of the Lessepsian Jellyfish Cassiopea andromeda in NW Sicily (Central Mediterranean Sea). PLoS ONE 2023, 18, e0281787. [Google Scholar] [CrossRef] [PubMed]
- Bengston, K.; Nichols, M.M.; Schnadig, V.; Ellis, M.D. Sudden Death in a Child Following Jellyfish Envenomation by Chiropsalmus quadrumanus: Case Report and Autopsy Findings. JAMA J. Am. Med. Assoc. 1991, 266, 1404–1406. [Google Scholar] [CrossRef]
- Eckelbarger, K.J.; Larson, R.J. Ultrastructural Study of the Ovary of the Sessile Scyphozoan, Haliclystus octoradiatus (Cnidaria: Stauromedusae). J. Morphol. 1993, 218, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Toshino, S.; Miyake, H.; Srinui, K.; Luangoon, N.; Muthuwan, V.; Sawatpeera, S.; Honda, S.; Shibata, H. Development of Tripedalia binata Moore, 1988 (Cubozoa: Carybdeida: Tripedaliidae) Collected from the Eastern Gulf of Thailand with Implications for the Phylogeny of the Cubozoa. Hydrobiologia 2017, 792, 37–51. [Google Scholar] [CrossRef]
- Alexandrova, O.; Schade, M.; Böttger, A.; David, C.N. Oogenesis in Hydra: Nurse Cells Transfer Cytoplasm Directly to the Growing Oocyte. Dev. Biol. 2005, 281, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Honegger, T.G.; Zürrer, D.; Tardent, P. Oogenesis in Hydra carnea: A New Model Based on Light and Electron Microscopic Analyses of Oocyte and Nurse Cell Diferentiation. Tissue Cell 1989, 21, 381–393. [Google Scholar] [CrossRef]
- Toyokawa, M.; Shimizu, A.; Sugimoto, K.; Nishiuchi, K.; Yasuda, T. Seasonal Changes in Oocyte Size and Maturity of the Giant Jellyfish, Nemopilema nomurai. Fish. Sci. 2010, 76, 55–62. [Google Scholar] [CrossRef]
- Iguchi, N.; Lee, H.E.; Yoon, W.D.; Kim, S. Reproduction of the Giant Jellyfish, Nemopilema Nomurai (Scyphozoa: Rhizostomeae), in 2006–2008 as Peripherally-Transported Populations. Ocean. Sci. J. 2010, 45, 129–138. [Google Scholar] [CrossRef]
- Jarms, G.; Tiemann, H.; Båmstedt, U. Development and Biology of Periphylla Periphylla (Scyphozoa: Coronatae) in a Norwegian Fjord. Mar. Biol. 2002, 141, 647–657. [Google Scholar] [CrossRef]
- Schiariti, A.Í.; Christiansen, E.; Morandini, A.C.; da silveira, F.L.; Giberto, D.A.; Mianzan, H.W. Reproductive Biology of Lychnorhiza lucerna (Cnidaria: Scyphozoa: Rhizostomeae): Individual Traits Related to Sexual Reproduction. Mar. Biol. Res. 2012, 8, 255–264. [Google Scholar] [CrossRef]
- Hargitt, G.T. Germ Cells of Coelenterates. VI. General Considerations, Discussion, Conclusions. J. Morphol. 1919, 33, 1–59. [Google Scholar] [CrossRef] [Green Version]
- Jarms, G.; Båmstedt, U.; Tiemann, H.; Martinussen, M.B.; Fosså, J.H.; Høisœter, T. The Holopelagic Life Cycle of the Deep-Sea Medusa Periphylla periphylla (Scyphozoa, Coronatae). Sarsia 1999, 84, 55–65. [Google Scholar] [CrossRef]
- Berrill, N.J. Developmental Analysis of Scyphomedusae. Biol. Rev. 1949, 24, 393–409. [Google Scholar] [CrossRef]
- Russell, F.S. On the Scyphomedusa Poralia Rufescens Vanhöffen. J. Mar. Biol. Assoc. UK 1962, 42, 387–390. [Google Scholar] [CrossRef] [Green Version]
- Lucas, C.H.; Reed, A.J. Gonad Morphology and Gametogenesis in the Deep-Sea Jellyfish Atolla wyvillei and Periphylla periphylla (Scyphozoa: Coronatae) Collected from Cape Hatteras and the Gulf of Mexico. J. Mar. Biol. Assoc. UK 2010, 90, 1095–1104. [Google Scholar] [CrossRef]
- Toshino, S.; Miyake, H.; Shibata, H. Development of Carybdea brevipedalia Kishinouye, 1891 (Cnidaria: Cubozoa: Carybdeida: Carybdeidae) Collected from Northern Japan. Plankton Benthos Res. 2018, 13, 116–128. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
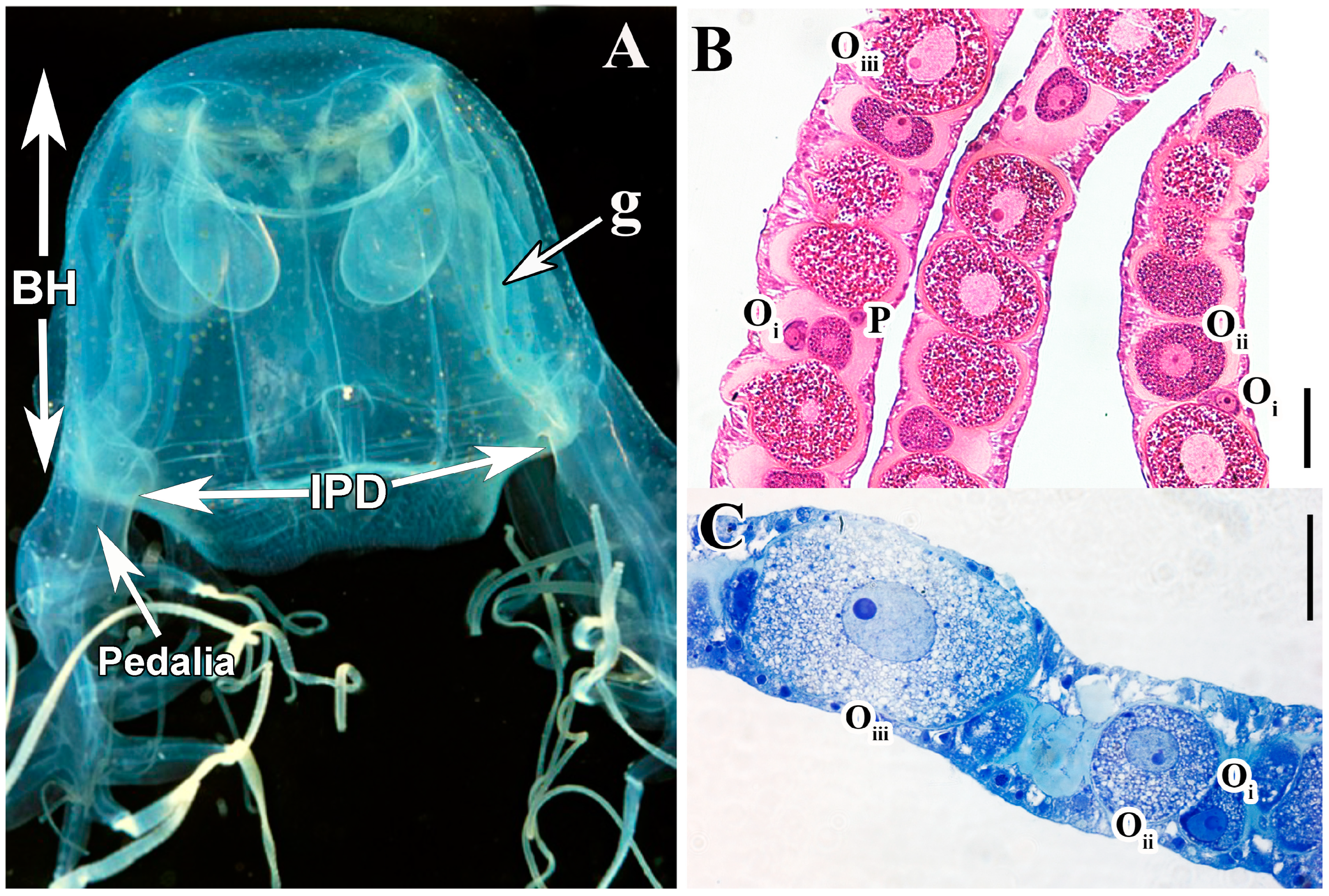{kind=link}
{kind=link}
{kind=link}
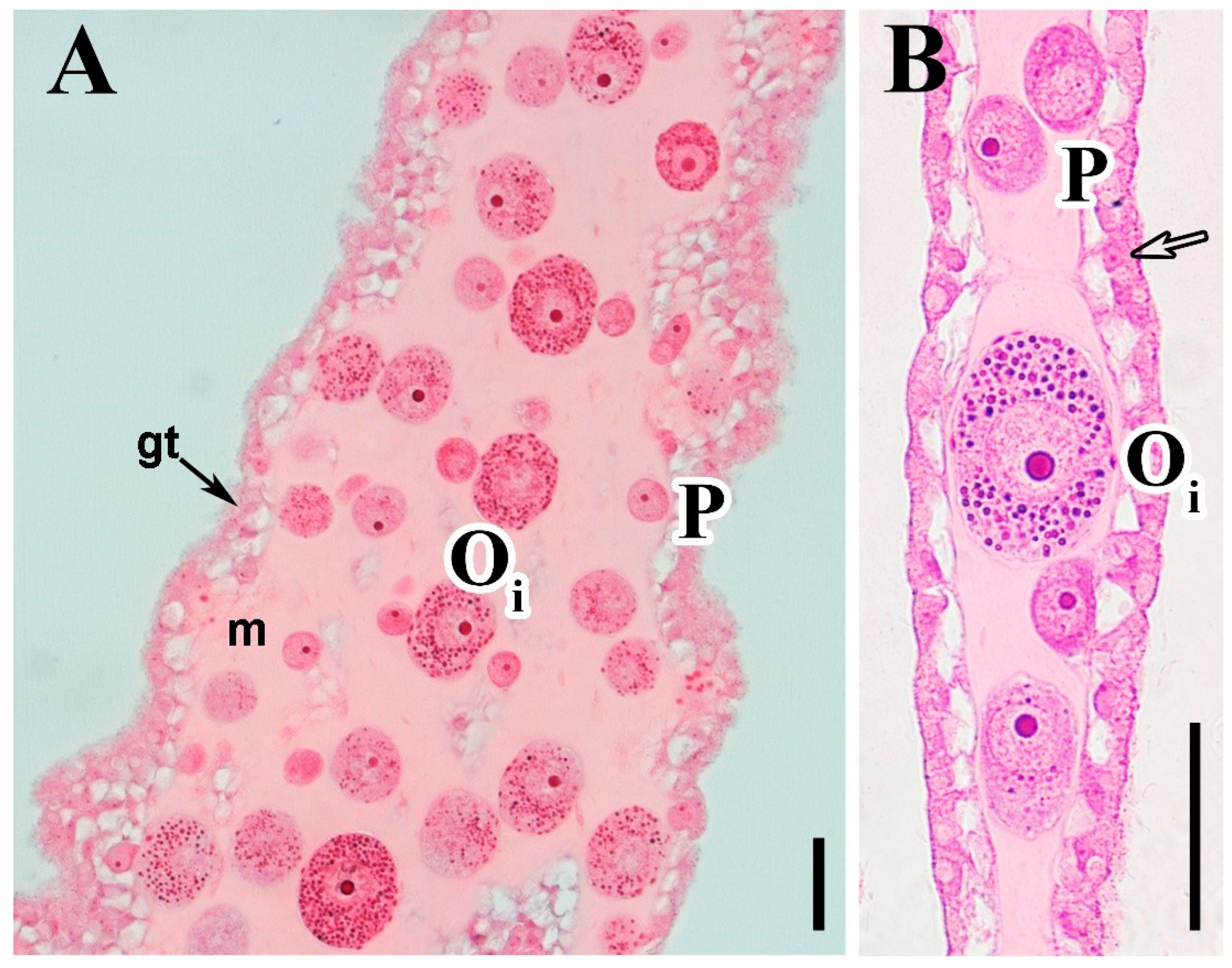{kind=link}
{kind=link}
| Specimen Number | Museum Number (MZUSP) and MEL Number | Bell Height (mm) | Interpedalial Distance (mm) | Locality | Depth (m) | Date (D/M/Y) | Collector |
|---|---|---|---|---|---|---|---|
| CH2 | MEL-CH2 | 4.5 | 3.8 | São Sebastião Channel (SP) | 10 | 10 June 2014 | JGR |
| CH6 | MEL-CH6 | 6.5 | 4.7 | São Sebastião Channel (SP) | 10 | 19 August 2014 | JGR |
| MA49 | MZUSP-1920 | 6.5 | 4.5 | Macaé (RJ) | 5–10 | 13 September 2008 | F.P.L. Marques |
| MA50 | MZUSP-1921 | 7.5 | 5 | Macaé (RJ) | 5–10 | 13 September 2008 | F.P.L. Marques |
| MA52 | MZUSP-1923 | 6.5 | 4.4 | Macaé (RJ) | 5–10 | 13 September 2008 | F.P.L. Marques |
| MA53 | MZUSP-1924 | 7 | 4.7 | Macaé (RJ) | 5–10 | 13 September 2008 | F.P.L. Marques |
| MA56 | MZUSP-1927 | 7 | 5 | Macaé (RJ) | 5–10 | 13 September 2008 | F.P.L. Marques |
| MA57 | MZUSP-1928 | 5.4 | 4 | Macaé (RJ) | 5–10 | 13 September 2008 | F.P.L. Marques |
| 53A | MEL-53A | 6.7 | 10.54 | Baía de Santos e São Vicente (SP) | 10–40 | April 2010 | Tiseo, G. R.; Zara, F. J. |
| 54C | MEL-54C | 5.6 | 8 | Baía de Santos e São Vicente (SP) | 10–40 | April 2010 | Tiseo, G. R.; Zara, F. J. |
| Specimen Number | Oiii Diameter (µm) | Oii Diameter (µm) | Oi Diameter (µm) | P Diameter (µm) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Average | Standard Deviation | Max | Min | Average | Standard Deviation | Max | Min | Average | Standard Deviation | Max | Min | Average | Standard Deviation | |
| CH2 | 140.014 | 76.292 | 110.796 | 22.54687 | 75.617 | 52.308 | 62.1977 | 6.2079 | 48.703 | 16.492 | 28.484 | 7.81417 | 31.68 | 11.544 | 20.5662 | 5.98748 |
| CH6 | 235.397 | 146.857 | 197.781 | 23.70315 | 170.486 | 99.398 | 132.068 | 22.297 | 92.352 | 49.656 | 69.34 | 12.3727 | 53.582 | 13.919 | 28.242 | 13.5341 |
| MA49 | 83.02 | 58.667 | 74.9411 | 7.3262 | 61.273 | 34.359 | 46.186 | 8.19608 | 26.104 | 16.271 | 20.8476 | 3.6763 | ||||
| MA50 | 98.593 | 70.107 | 85.8052 | 8.959 | 58.029 | 31.074 | 42.923 | 8.00621 | 37.838 | 14.411 | 24.7551 | 7.08516 | ||||
| MA52 | 131.602 | 96.736 | 114.187 | 10.50545 | 97.173 | 65.079 | 81.6818 | 11.168 | 59.958 | 26.203 | 38.339 | 10.4808 | 27.778 | 13.889 | 20.9081 | 4.47119 |
| MA53 | 113.525 | 95.89 | 103.493 | 5.407642 | 87.199 | 60.336 | 76.8158 | 8.5688 | 41.82 | 27.397 | 33.926 | 4.1092 | 34.013 | 13.784 | 20.6236 | 5.4142 |
| MA56 | 119.398 | 75.18 | 90.0476 | 13.532 | 63.774 | 26.698 | 43.078 | 10.8529 | 27.68 | 10.99 | 20.4653 | 5.60585 | ||||
| MA57 | 47.507 | 33.333 | 38.761 | 4.44204 | 23.292 | 10.623 | 16.8126 | 4.18917 | ||||||||
| 53A | 122.144 | 89.443 | 101.199 | 10.10663 | 91.952 | 56.063 | 79.3338 | 8.9946 | 57.007 | 25.298 | 37.208 | 10.46 | 24.828 | 14.717 | 19.9075 | 3.43497 |
| 54C | 131.966 | 82.262 | 99.711 | 15.20876 | 88.511 | 62.639 | 74.9315 | 8.3674 | 61.837 | 27.402 | 39.457 | 9.03138 | 29.829 | 14.583 | 20.7193 | 5.1459 |
| Specimen number | IPD | Oiii% | Oii% | Oi% | P% | Sampling Month |
|---|---|---|---|---|---|---|
| CH2 | 3.8 | 6.45 | 45.16 | 41.94 | 6.45 | June/2014 |
| CH6 * | 4.7 | 37.68116 | 27.53623 | 24.63768 | 10.14493 | August/2014 |
| MA49 | 4.5 | 0 | 8.474576 | 83.89831 | 7.627119 | September/2008 |
| MA50 | 5 | 0 | 38.70968 | 38.70968 | 22.58065 | September/2008 |
| MA52 * | 4.4 | 35.38462 | 47.69231 | 12.30769 | 4.615385 | September/2008 |
| MA53 * | 4.7 | 26.05042 | 35.29412 | 24.36975 | 14.28571 | September/2008 |
| MA56 | 5 | 0 | 62.63736 | 32.96703 | 4.395604 | September/2008 |
| MA57 | 4 | 0 | 0 | 85.65022 | 14.34978 | September/2008 |
| 53A * | 10.54 | 32.91139 | 44.3038 | 14.76793 | 8.016878 | April/2010 |
| 54C | 8 | 4.659498 | 60.9319 | 28.67384 | 5.734767 | April/2010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Rodríguez, J.; Ames, C.L.; Jaimes-Becerra, A.; Tiseo, G.R.; Morandini, A.C.; Cunha, A.F.; Marques, A.C. Histological Investigation of the Female Gonads of Chiropsalmus quadrumanus (Cubozoa: Cnidaria) Suggests Iteroparous Reproduction. Diversity 2023, 15, 816. https://doi.org/10.3390/d15070816
García-Rodríguez J, Ames CL, Jaimes-Becerra A, Tiseo GR, Morandini AC, Cunha AF, Marques AC. Histological Investigation of the Female Gonads of Chiropsalmus quadrumanus (Cubozoa: Cnidaria) Suggests Iteroparous Reproduction. Diversity. 2023; 15(7):816. https://doi.org/10.3390/d15070816
Chicago/Turabian StyleGarcía-Rodríguez, Jimena, Cheryl Lewis Ames, Adrian Jaimes-Becerra, Gisele Rodrigues Tiseo, André Carrara Morandini, Amanda Ferreira Cunha, and Antonio Carlos Marques. 2023. "Histological Investigation of the Female Gonads of Chiropsalmus quadrumanus (Cubozoa: Cnidaria) Suggests Iteroparous Reproduction" Diversity 15, no. 7: 816. https://doi.org/10.3390/d15070816