3.1.1. Genus Abyssaranea Wilson and Hessler, 1974
Abyssaranea Wilson and Hessler, 1974: 58.
Type species. Abyssaranea rupis Wilson and Hessler, 1974, by monotypy.
Species included. Abyssaranea minuta Golovan and Malyutina sp. nov., A. rupis Wilson and Hessler, 1974.
This species is registered in ZooBank under urn:lsid:zoobank.org:act:23F22647-F7C7-4EB5-9B6C-7659C802ACB2.
Material examined. Holotype: (SMF 61310) female (total length 1.1 mm), KB 223-8-9, designated here.
Etymology. The new species name is derived from the Latin adjective “minutus” that means “small, tiny”.
Diagnosis. Female. Dorsal surface lacking spines. Pereonite 1 length 0.3 width and 1.6 of pereonite 2 length. Pleotelson length 1.5 width, with about eight long UBS dorsally and one stout UBS on each lateral margin anteriorly to the place of uropod insertion. Pereopod I carpus is as long as the basis, with length 2.7 width; dactylus and propodus together are about as long as the carpus. Operculum length 1.2 width.
Description. Holotype, female:
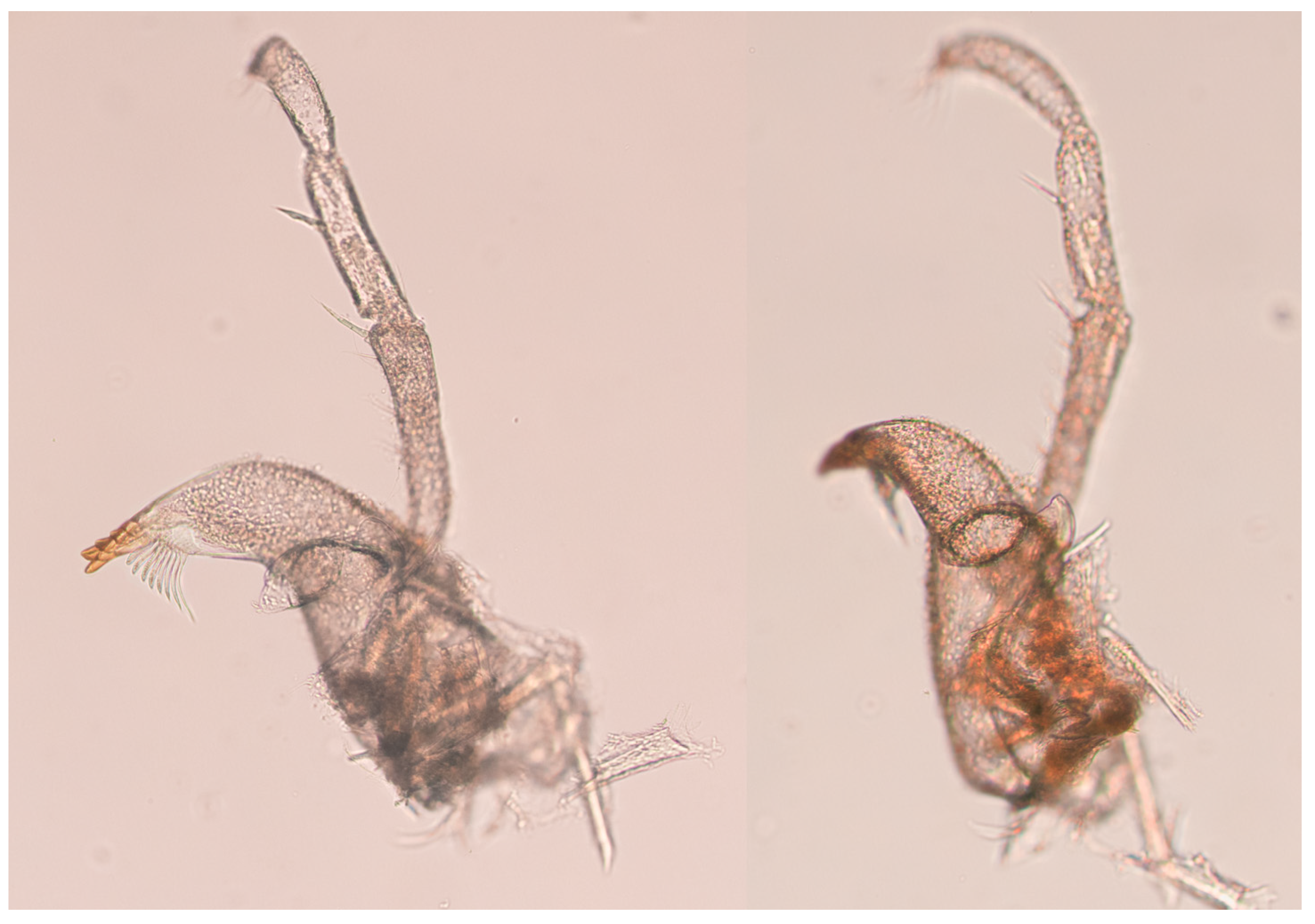
body (
Figure 2J and
Figure 3A,B) total length 2.0 width of pereonite 3; body thickest and deepest at pereonite 3; depth 0.38 total body length. Total length 1.1 mm, cephalothorax–pereon length 0.76 mm. Spines absent.
Cephalothorax: length 0.68 width and 0.2 total body length, width 0.7 width of pereonite 1; frons and clypeus rounded in dorsal view.
Pereonite 1 length 0.28 width and 0.12 body length; pereonite 6 visible in dorsal view as a thin belt; pereonites 2–6 lengths to pereonite 1 length ratios: 0.6; 0.7; 0.5; 0.3; 0.3; length to width ratios: 0.15; 0.16; 0.12; 0.09. Dorsal surface of pereonite 3 with pair of long submedial UBS.
Pleotelson length 1.5 width and 0.46 body length, oval in dorsal view, and posterior tip rounded. Dorsal surface with about eight long UBS; lateral margin with one stout UBS anteriorly to the place of uropod insertion.
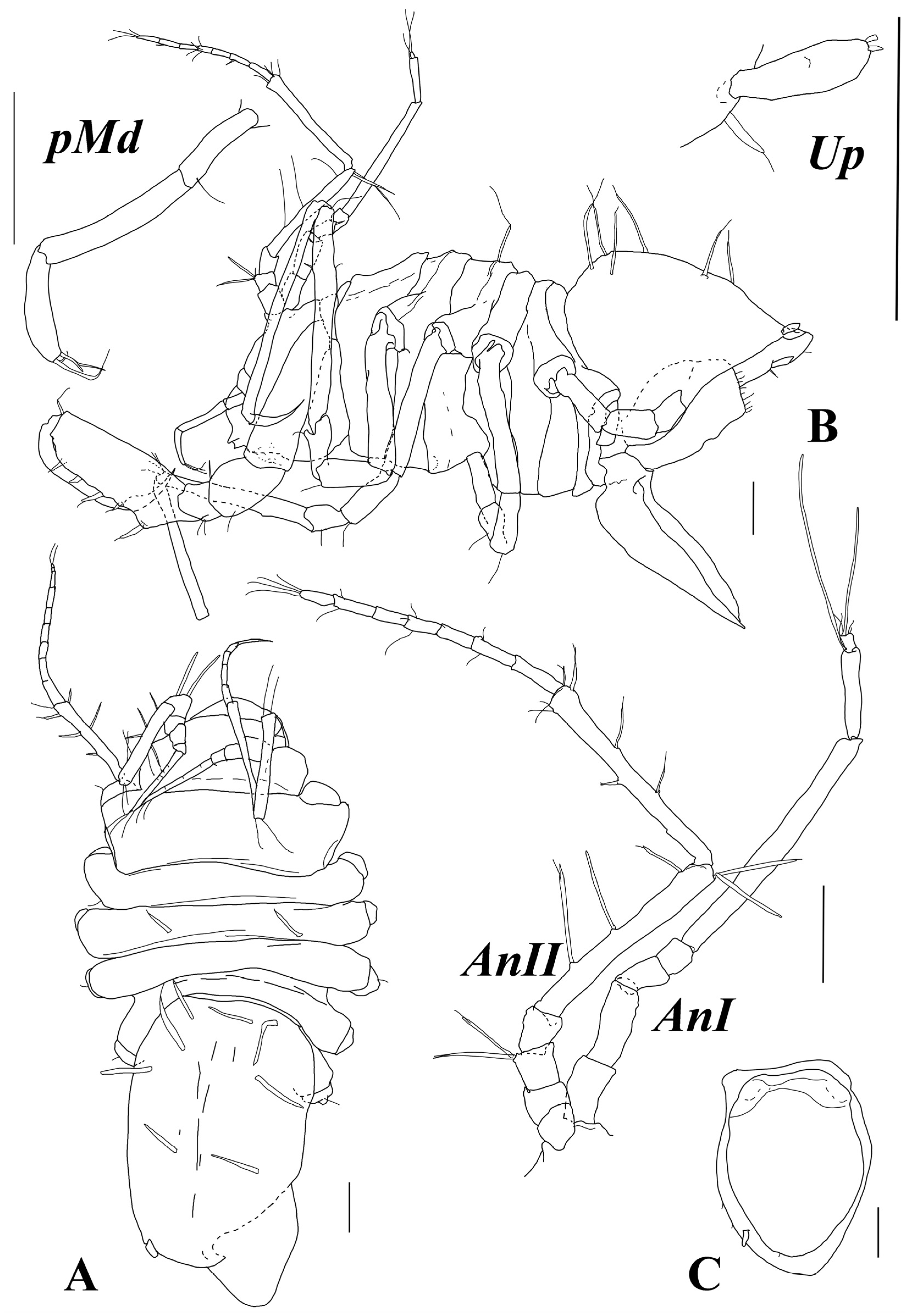
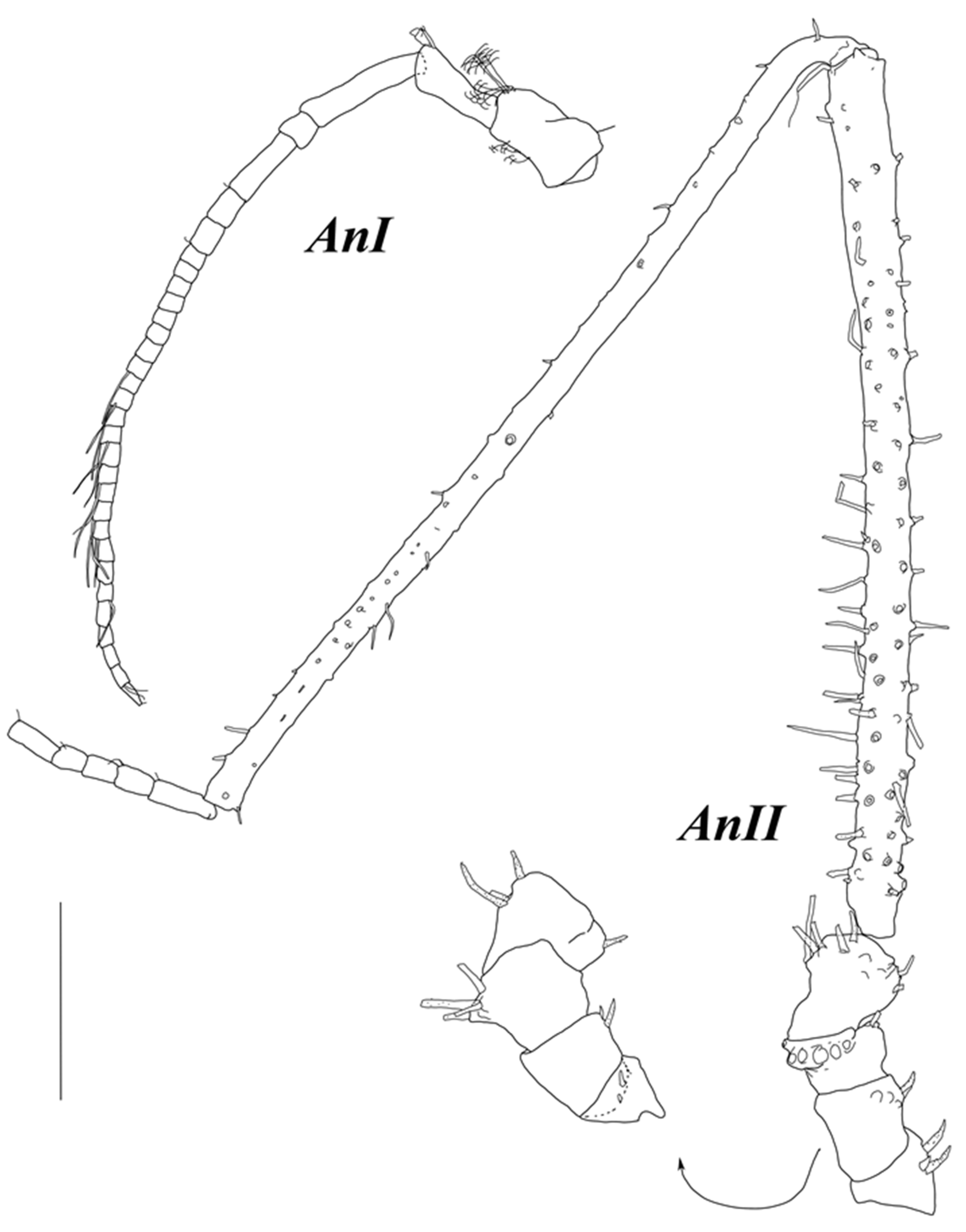
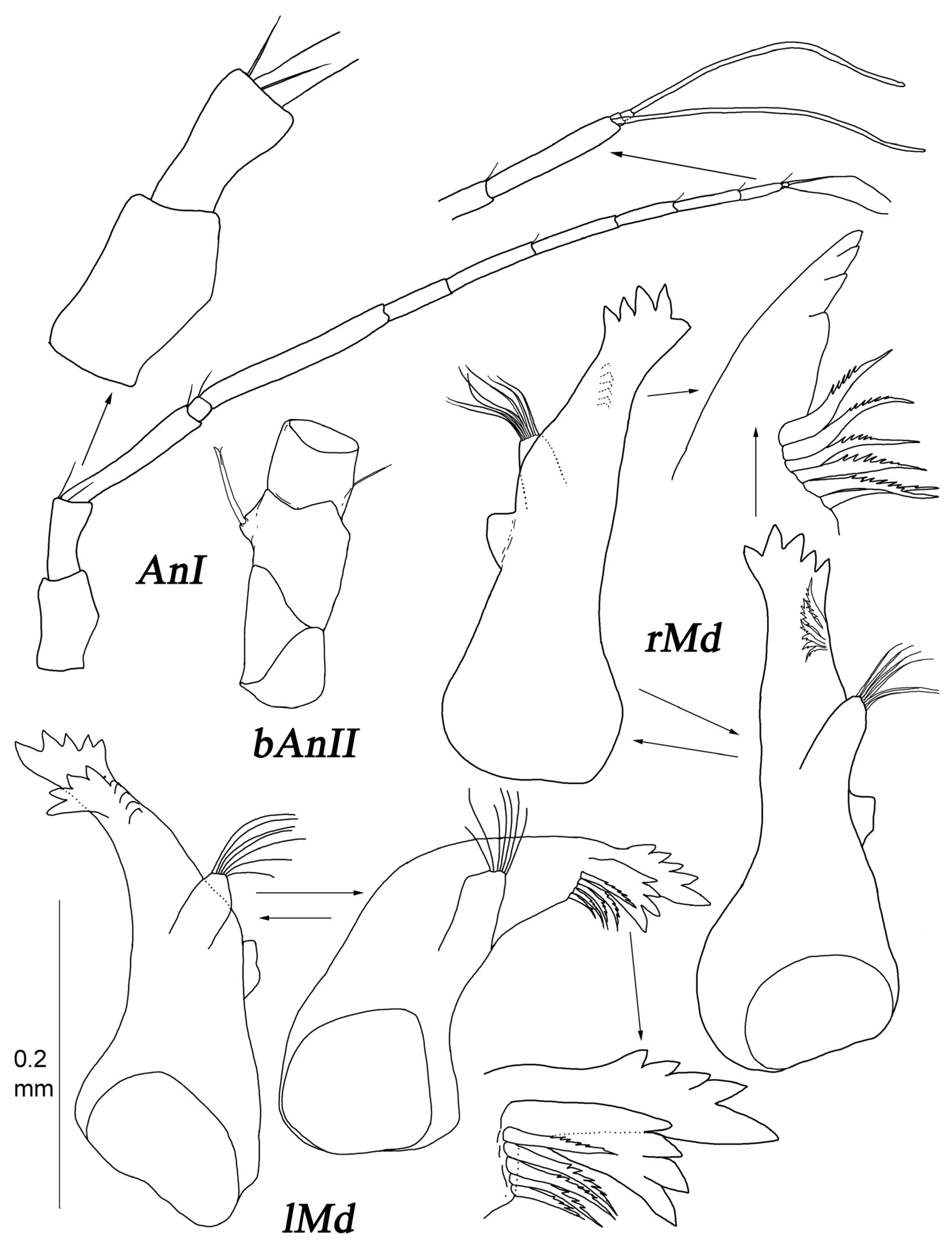
Antenna I (
Figure 3) with seven articles; articles 2–7 lengths to article 1 length ratios: 1.2; 0.64; 0.50; 3.9; 1.2; 0.23; articles 1–7 length to width ratios: 2.0; 2.4; 1.5; 1.2; 10; 4.9; 1.2. Articles 6 and 7, each with one aestetasc; article 7 with five fine setae terminally.
Antenna II (
Figure 3) articles 2–6 lengths to article 1 length ratios: 1.1; 1.1; 1.2; 6.4; 6.5; articles 1–6 length to width ratios: 0.86; 0.89; 0.92; 1.1; 7.6; 7.9. Article 3 with two long distoventral UBS; article 5 with three ventral and two distodorsal long UBS; article 6 with four UBS dorsally, one UBS, and one trace of seta insertion ventrally and few fine setae distoventrally. Flagellum length 1.2 length of article 6, with seven articles bearing fine SS.
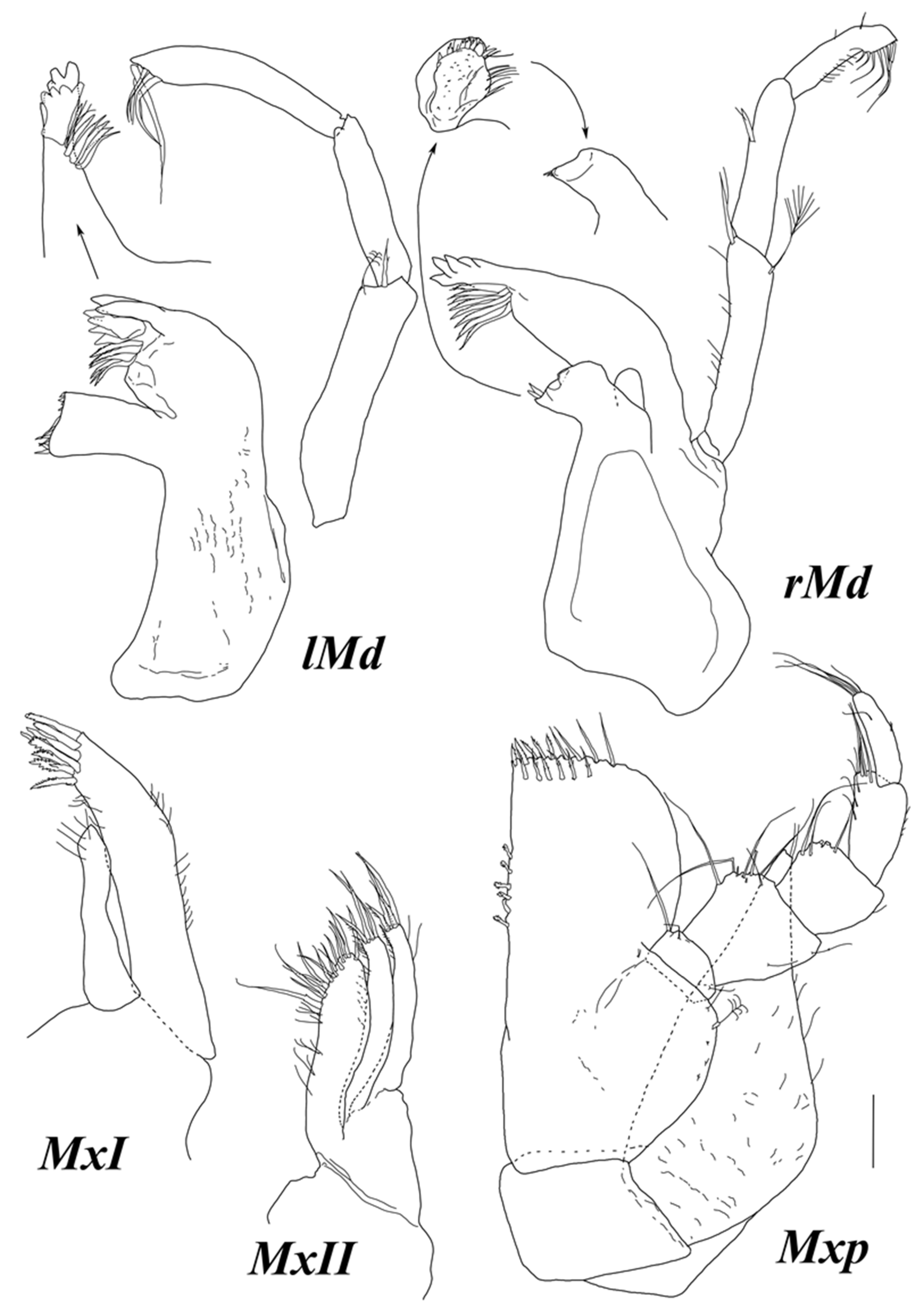
Mandibular palp (
Figure 3) about 0.2 of total body length. Article 1 length about 3.8 width, with one distoventral seta. Articles 2 and 3 lengths about 5 and 4 widths, and 1.6 and 1.1 article 1 length, respectively. Article 3 with a row of few setae.
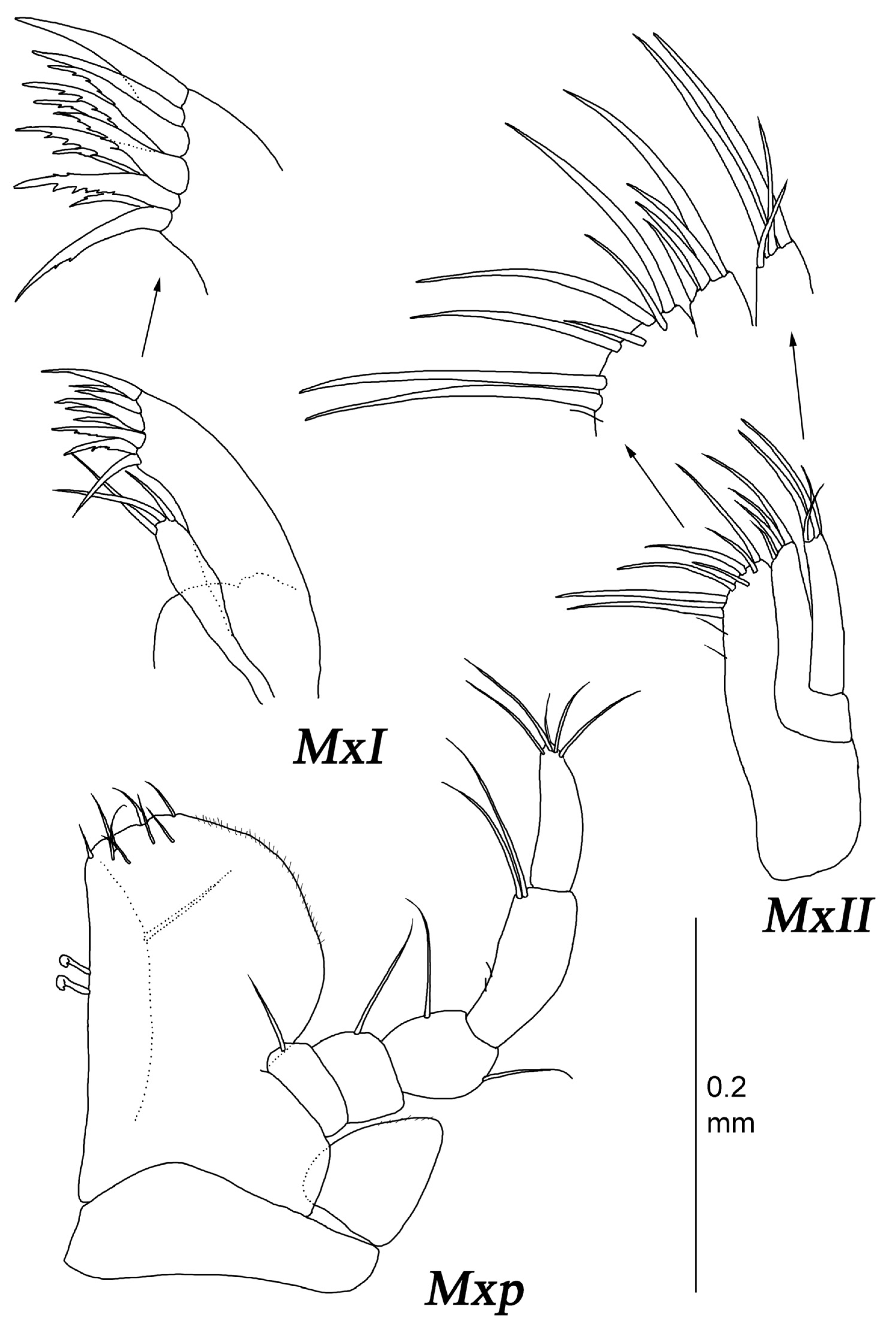
Maxilliped with two retinaculae.
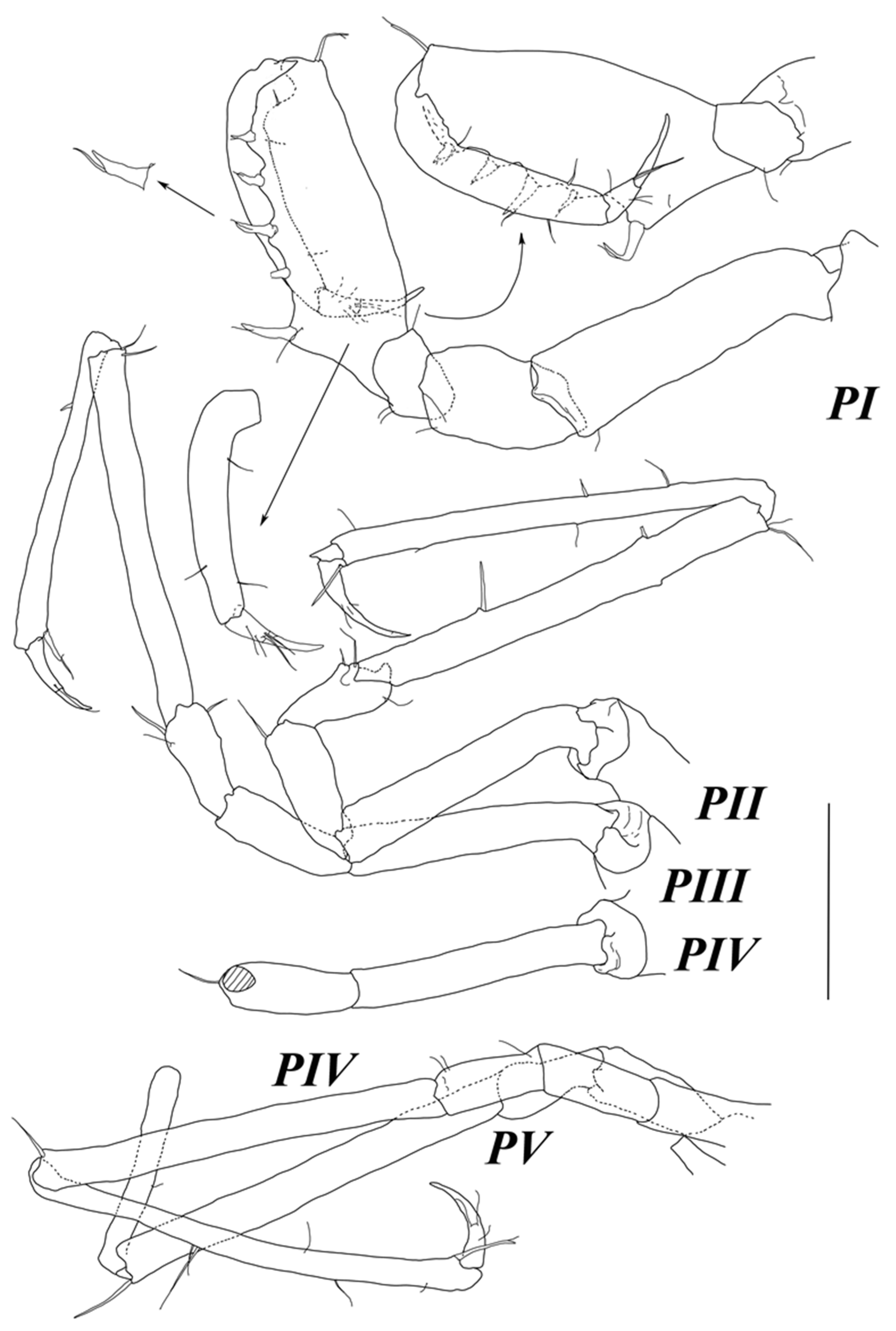
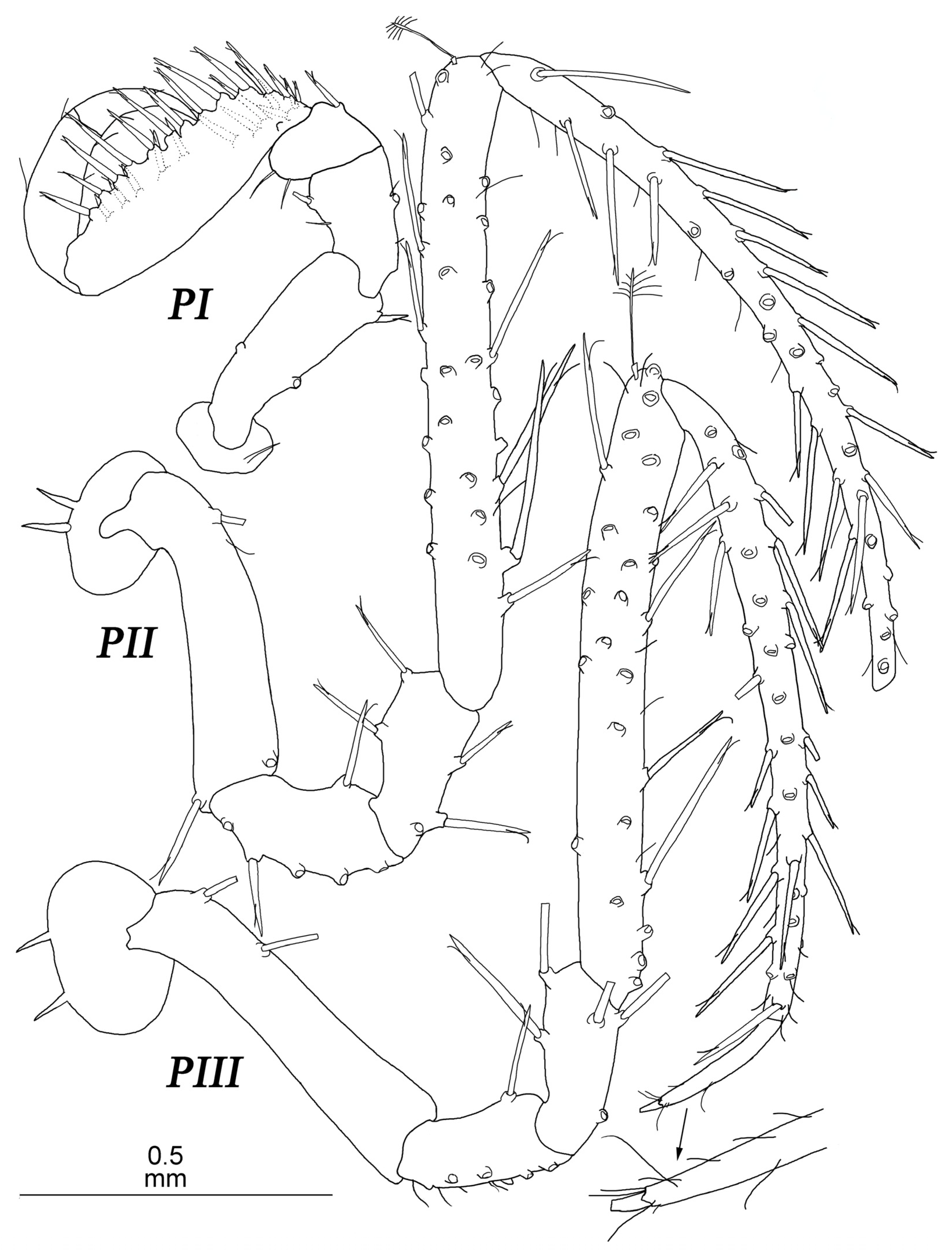
Pereopod I (
Figure 4) basis–dactylus length to width ratios: 3.4; 1.6; 1.4; 2.7; 6.1; 4.1; ischium–dactylus to basis length ratios: 0.47; 0.29; 1.0; 0.75; 0.32. Basis and ischium, each with one fine SS distoventrally. Merus with two setae ventrally and one SS distodorsally. Carpus ventral margin with a row of five strong UBS opposing propodus; proximal UBS largest, located on blunt projection and accompanied with one SS; distodorsal angle with one long UBS; few SS on lateral margin. Propodus with one dorsal and two ventral SS. Dactylus with few SS on lateral margin; claw 0.57 dactylus length, with two slender setae at base of claw.
Pereopods II–V (
Figure 2J,
Figure 3B and
Figure 4) slightly longer than body, similar in shape. Bases length about 5–6 width. Ischia length about 2.5–3 width and about half of bases length, with distoventral seta. Meri length about 2.3 width and 0.4 bases length, with few setae distally. Carpi length about 10 width and about 1.5 bases length, with 1–2 distodorsal setae; in pereopod 2 with two stout setae on ventral margin and one trace of seta insertion on dorsal margin. Propodi length about 14 width and about as long as carpi, with few fine setae, 1–2 UBS dorsally, and one long stout UBS distoventrally at the base of dactylus insertion. Dactyli length about 4–5 width and about 0.3–0.4 bases length; claw length about 0.4 dactylus length.
Pereopod VI broken off, except basis.
Operculum (
Figure 3B,C) length 1.4 width; convex ventrally.
Pleopods III–V not studied.
Uropod (
Figure 3) length about 3.0 width, with two setae distally (broken off).
Remarks. The new species can be distinguished from the only known species of the genus A. rupis by the shape and proportions of the body segments and appendages. In general, A. minuta sp. nov. has a slenderer and less vaulted body than A. rupis. Thus, the new species has a relatively short pereonite 1 (0.3 times longer than width, 1.6 times longer than pereonite 2) compared to that of A. rupis (0.4 times longer than width, 2 times longer than pereonite 2). This character is associated with a slenderer pereopod I in the new species, whose carpus is 2.7 times longer than its width and equal in length to the basis (vs. carpus 2.2 times longer than its width and 1.3 times longer than basis in A. rupis). The new species has more elongated pleotelson and operculum (each about 1.5 times longer than its width) compared to those in A. rupis (each 1.2 times longer than width). Unlike A. rupis, A. minuta sp. nov. has no spines on the dorsal surface and possesses only stout UBS on the pereonite 3 and pleotelson. Furthermore, the new species differs from A. rupis by 7-articled antenna I (vs. 11-articled in the females of A. rupis) with article 5 almost four times longer than article I (about 1.4 times longer in A. rupis). Nevertheless, this may be due to the immaturity stage of the holotype of the new species.
Wilson and Hessler [
9] reported a pronounced sexual dimorphism in
A. rupis. The females in
A. rupis are larger than males (2.1 mm vs. 1.5–1.7 mm) and possess more sturdy-looking bodies. The female holotype of
A. minuta sp. nov. (1.1 mm) is smaller than males of
A. rupis. Nevertheless, body size cannot be used as a diagnostic feature for the new species, since only one specimen of
A. minuta sp. nov. was studied.
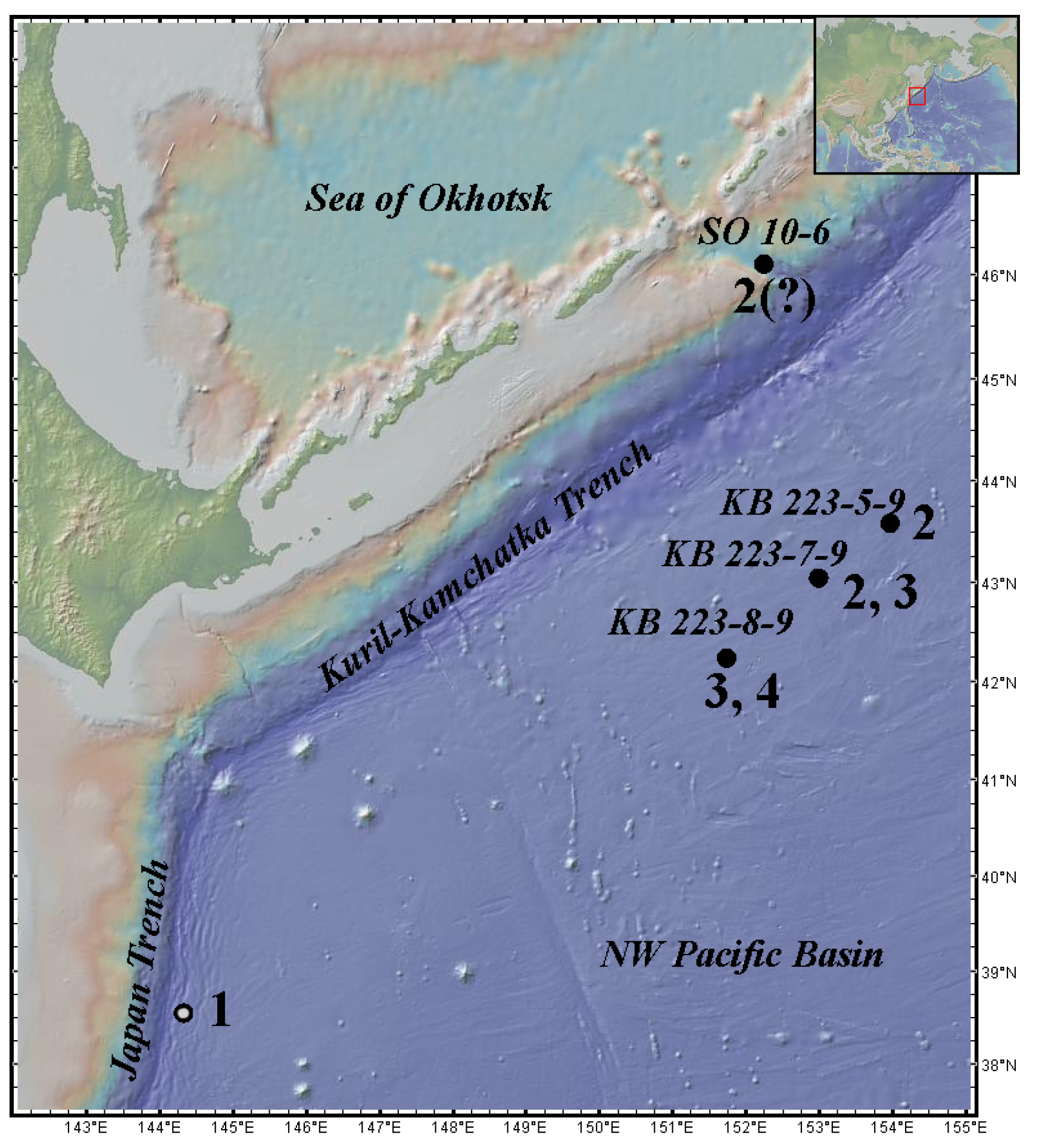
Distribution. The species is known only from the type locality in the NW Pacific Basin to the east of the Kuril–Kamchatka Trench, 42.2447° N 151.7351° E 42.2378° N 151.7082° E, at a depth of 5125–5126 m.
3.1.2. Genus Haplomunna Richardson, 1908
Munna Richardson, 1905: 483 (part).
Haplomunna Richardson, 1908: 79; Menzies, 1962: 32; Wilson, 1976: 574; Cunha and Wilson, 2003: 8.
Aryballurops Gamô, 1983: 13–16, figs 8 and 9.
Type species. Munna caeca Richardson, 1905, by monotypy.
Species included. Haplomunna caeca (Richardson, 1905); H. hubbsi Wilson and Hessler, 1976; H. japonica (Gamô, 1983); H. kurilensis Golovan and Malyutina sp. nov.
Haplomunna kurilensis Golovan and Malyutina sp. nov.
This species is registered in ZooBank under urn:lsid:zoobank.org:act:7050DA63-C582-4CC1-AB6C-C978D1D2E8AC.
Material examined. Holotype: (SMF 61311) male (total length 4.4 mm), KB 223-7-9, designated here.
Paratypes: (MIMB 46614) males 1 and 2 (total lengths 4.0 mm), used for dissection, two males (total lengths 3.4 mm), female (total length 3.5 mm), one damaged specimen, six mancas (total lengths 1.3–2.1 mm), the same data as holotype; (SMF 61312) six mancas, the same data as holotype; (SMF 61313) male, KB 223-5-9.
Etymology. The new species is named after the place where it was sampled, i.e., to the east of the Kuril Islands.
Diagnosis. Body surface with numerous thin solitary spines, covered with fine hairs; on dorsomedial parts of each of pereonites 1–3; these spines are grouped on two transverse ridges. Pleotelson inflated, about as long as wide, height 0.8 length, and length 0.6 cephalothorax–pereon length. Maxilliped with 5–6 retinaculae, epipod about as long as basis, narrowing distally. Mandibular palp article 1 with one UBS and one BS distally and about six SS. Pereopod I carpus length about 3 width. Male pleopod I posterior tip pointed, with pronounced distolateral angles.
Description. Holotype, male:
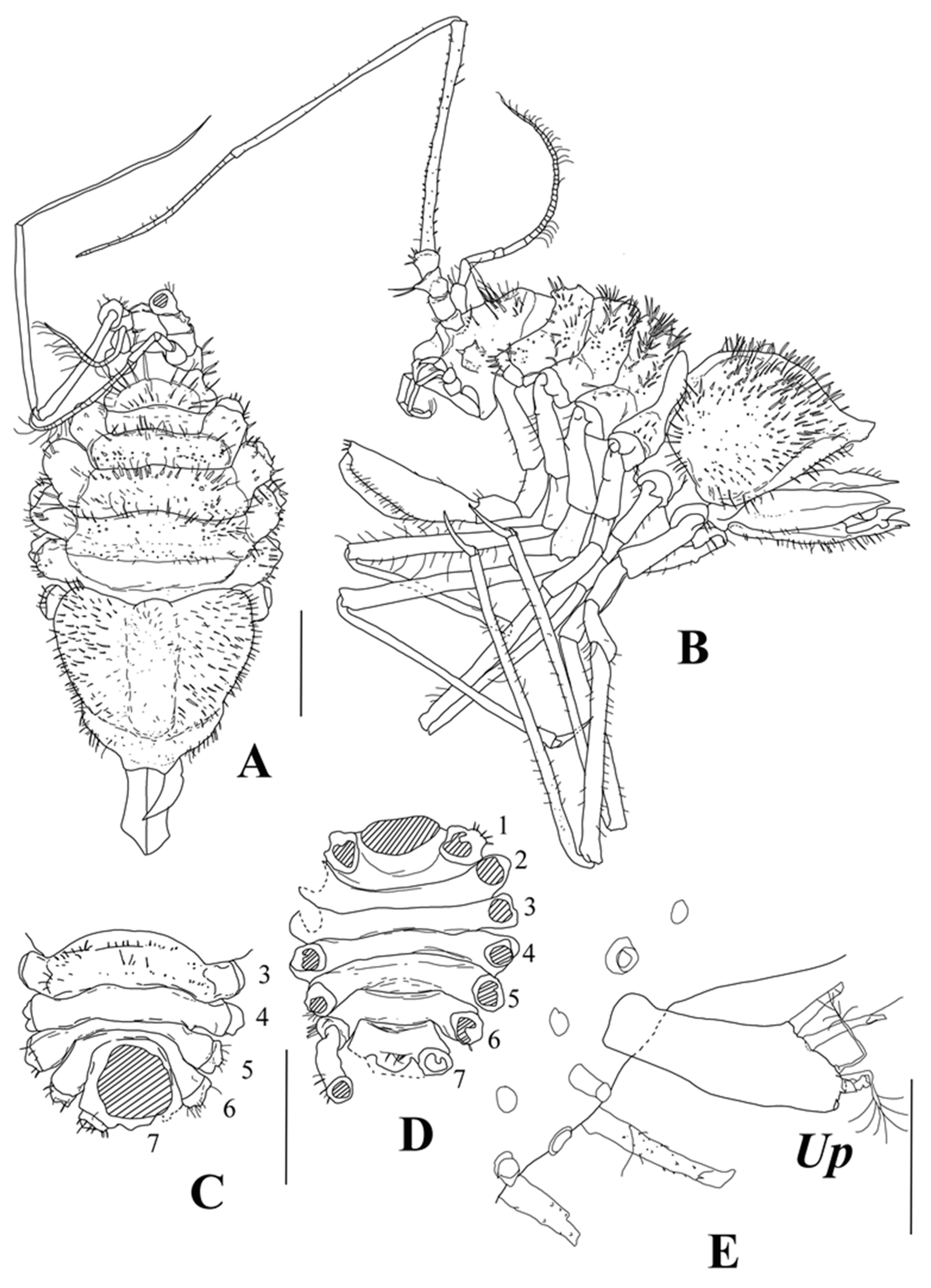
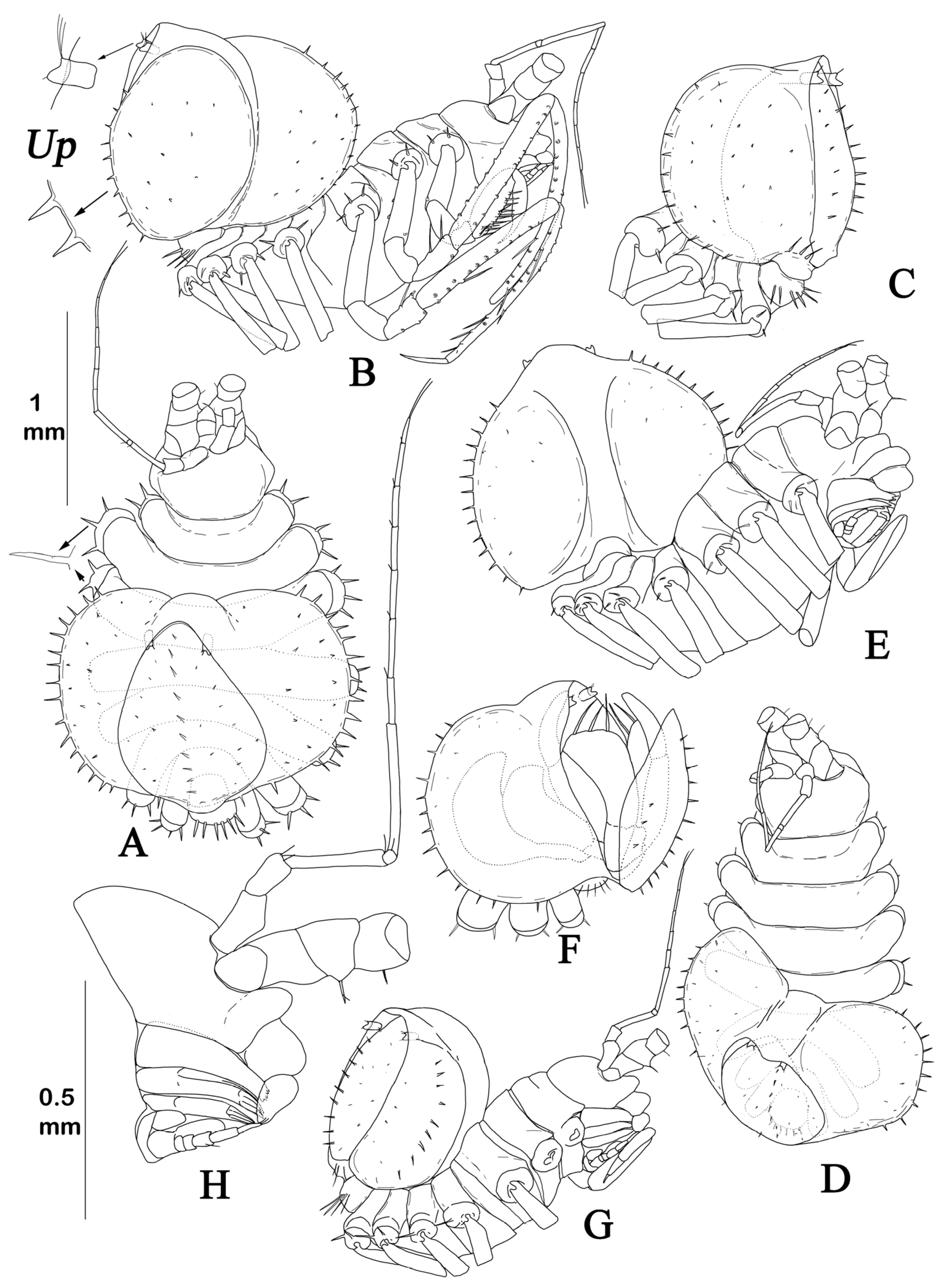
body (
Figure 2A,B and
Figure 5A,B) vaulted, thickest and deepest at pereonite 3, total length 1.9 width, cephalothorax–pereon length 1.4 width, and depth 0.48 cephalothorax–pereon length.
Cephalothorax, pereonites 1–3, and pleotelson dorsal surface with numerous thin spines (
Figure 5E), grouped in two transverse rows on medial parts of cephalothorax and pereonites, and evenly arranged on lateral expansions of pereonites and pleotelson. Surface spines of cephalothorax in anterior row are larger than in posterior row. Pereonite 4 with thin spines on dorsal surface of lateral expansions. Pereonites 5–7 with few thin spines laterally. Surface of body and spines with fine setulae, sitting on small tubercle (
Figure 5E). In dissected paratype male (
Figure 5D), pereonite 7 with a transverse row of small thin spines on ventral surface.
Cephalothorax of holotype length 0.69 width and 0.23 cephalothorax–pereon length, width 0.63 width of pereonite 1; with straight beveled frons.
Pereonites 1–3 lengths about 0.2 widths, pereonites 2 and 3 each 1.1 times longer than pereonite 1. In dissected paratype male with removed pleotelson on dorsal view (
Figure 5C),
pereonites 4–7 lengths pereonite 3 length ratios: 0.56; 0.14; 0.16; 0.09; on ventral view (
Figure 5D), pereonites 2–7 lengths in relation to pereonite 1 length ratios: 0.62; 0.59; 0.71; 1.16; 0.85; 1.2.
Pleotelson about as long as wide, in holotype length 0.61 cephalothorax–pereon length, height 0.80 length, swollen, heart-shaped, medial portion slightly depressed, distal margin flattened, and distal tip bilobed.
Antenna I about 0.84 cephalothorax–pereon length (
Figure 5B). In dissected paratype male (
Figure 6), articles 1–3 length/width ratios: 1.6; 2.5; 4.1; articles 2–3 lengths to article 1 length ratios: 0.82; 1.0. Article 1 with three small BS medially and five BS distolaterally; article 2 with one UBS and one broken large seta distally. Flagellum with 29 articles, articles 12–23 bearing aestetascs.
Antenna II 2.2 cephalothorax–pereon lengths (
Figure 5B). In dissected paratype male (
Figure 6), articles 1 and 2 about equal in length, article 3 slightly shorter, and article 4 slightly longer than article 1; articles 5 and 6 lengths 10.5 and 11.7 article 1 length, respectively. Articles 1–4 with thin setulated spines distally; articles 5 and 6 covered with stout setulated setae. Flagellum with more than 15 articles (
Figure 5B), in dissected specimen flagellum broken off.
Pereopod I (
Figure 5B) length in straightened state about 1.2 cephalothorax–pereon length;
pereopods 2–7 lengths about 2–2.3 cephalothorax–pereon length.
Male paratypes:
Mandibles (
Figure 7 and
Figure 8): incisor process with five cusps.
Lacinia mobilis of left mandible with four cusps; spine row with six spines and three SS. Spine row of right mandible with eight spines. Molar process thin, tubular, apex oval, and with sharp edges, producing two lobes bearing setae and scales. Palp length about 0.17 total body length; articles 1–3 lengths about 0.6, 0.4, 0.5 mandibular body length, respectively; article 1 with about six SS on anterior side and one UBS and one BS distally; article 2 with one UBS on anterior side; article 3 with about nine setae, first three of them much longer than other setae.
Maxilla I (
Figure 7) lateral endite with seven robust serrate setae distally, with few fine SS laterally and medially; mesial endite width 0.66 lateral endite width, distally with five fine SS.
Maxilla II (
Figure 7) lateral and middle endites similar in size, with four robust serrate setae distally and few fine SS laterally and medially; mesial endite width 1.8 lateral endite width, with 14 robust serrate setae distally, and fine SS medially.
Maxilliped (
Figure 7) basis length 2 width, with one SS and one BS near palp insertion; endite length 1.4 width, 0.6 basis length, with 5–6 retinaculae, and distal margin with 15 DSS. Palp almost as long as basis, article 1 width 0.36 basis width, lateral length 0.38 width; articles 2–5 to article 1 lateral length ratios: 3.9; 2.7; 3.6; 3.4; articles 1–5 lateral lengths to width ratios: 0.38; 0.99; 0.88; 1.9; 3.6; articles 1–4 with two, eight, five, and six distomedial SS, respectively; article 6 with four terminal SS. Epipod length 2.1 width and 0.91 basis length, narrowing distally, distal tip truncated, with two SS.
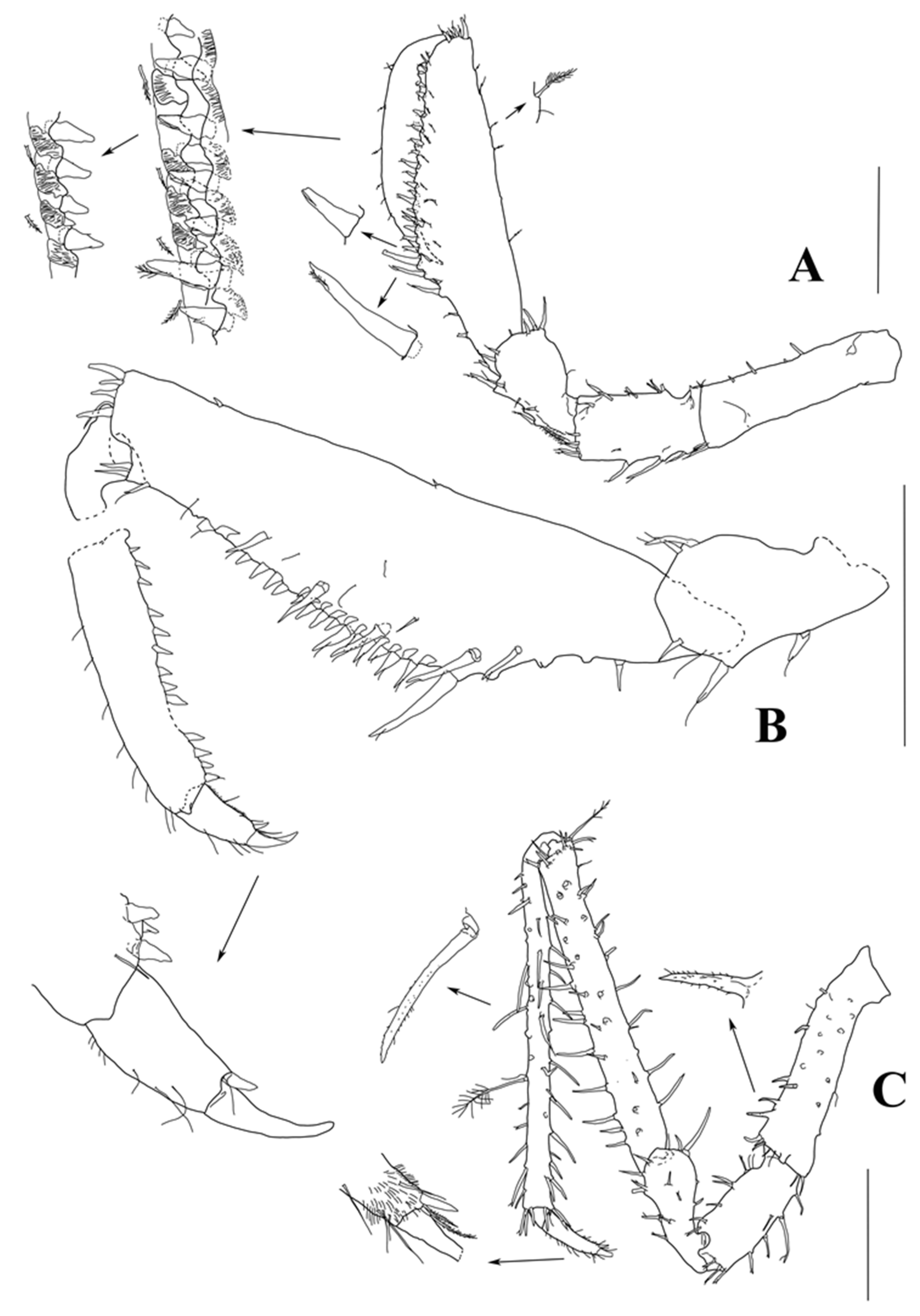
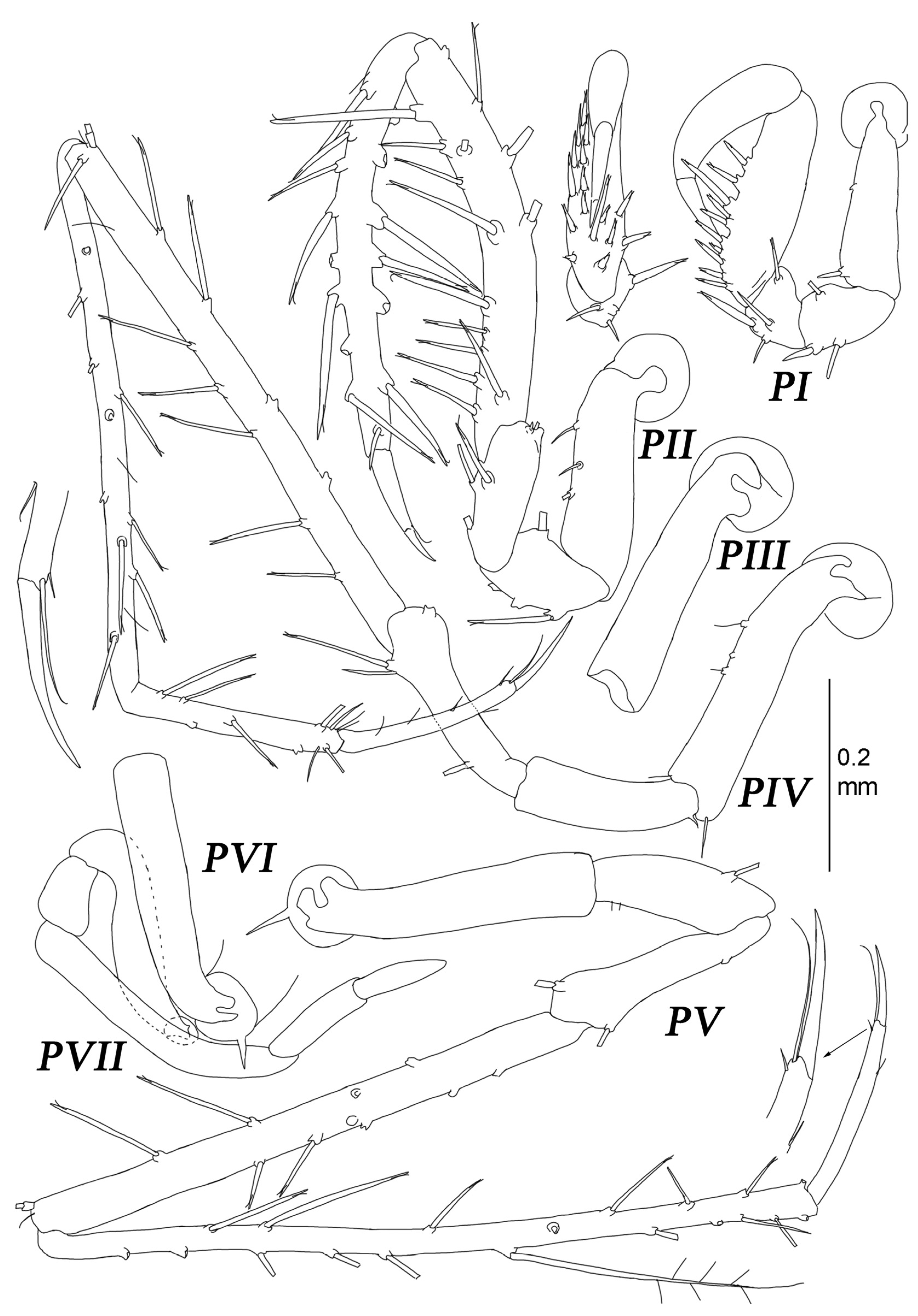
Pereopod I (
Figure 9) basis–dactylus length to width ratios: 3.5; 1.8; 1.5; 3.1; 4.2; 3.3; ischium–dactylus lengths to basis length ratios: 0.59; 0.52; 1.4; 0.82; 0.29. Basis, ischium, and merus bearing numerous stout-composed setae (UBS, UBDS). Carpus increasing in width towards midlength, with six stout UBS distodorsally and numerous fine DSS dorsally and subventrally; ventral margin convex, proximal part bearing 5–6 UBDS, increasing in size toward occlusal surface, with distal about as long as dactylus; occlusal surface occupying 0.6 of ventral margin, with about 15 strong tooth-like UBDS, which alternate with grooves (fit to same UBDS located on ventral margin of propodus); additionally, about five strong UBDS increasing in size toward occlusal surface, situated subventrally; ventral margin distal to occlusal surface with three small UBS. Propodus with numerous fine DSS on dorsal margin and along occlusal surface of ventral margin; occlusal surface similar to that of carpus, with about 15 tooth-like UBDS and grooves. Dactylus with few fine setae dorsally; dorsal claw length 0.49 dactylus length; ventral claw length 0.28 dorsal claw length; one seta inserted between claws.
Pereopods II–VII (
Figure 9C and
Figure 10) surface covered with numerous fine setulate; bases dorsal surface bearing few thin setulated spines; all articles except dactyli bearing numerous composed setae (setulated setae, UBS). Proximal articles (bases–meri) and dactyli similar in shape and proportions, while carpi and propodi increasing in length from anterior to posterior pereopods. Dactyli with two claws each; dorsal claw length about 0.4–0.5 dactylus length; ventral claw length about 0.3–0.4 dorsal claw length; two setae inserted between claws.
Pereopod II (
Figure 9C) basis–dactylus length to width ratios: 3.5; 2.1; 2.3; 7.7; 12.3; 4.0; ischium–dactylus to basis length ratios: 0.55; 0.58; 1.5; 1.7; 0.41; two setulated setae inserted between claws.
Pereopod III (
Figure 10) basis–dactylus length to width ratios: 3.5; 2.4; 2.2; 8.6; 19.8; 6.5; ischium–dactylus length to basis length ratios: 0.66; 0.69; 1.9; 2.8; 0.55.
Pereopod IV (
Figure 10) basis–dactylus length to width ratios: 4.3; 2.5; 2.5; 8.6; 18.3; 5.1; ischium–dactylus length to basis length ratios: 0.55; 0.61; 1.6; 2.3; 0.41.
Pereopod V (
Figure 10) basis–dactylus length to width ratios: 3.6; 2.4; 2.5; 9.3; 24.8; 6.2; ischium–dactylus length to basis length ratios: 0.63; 0.58; 1.8; 3.2. 0.56.
Pereopod VI (
Figure 10) basis–carpus length to width ratios: 3.1; 2.9; 2.6; 11.0; ischium–carpus length to basis length ratios: 0.80; 0.78; 2.3. Distal articles broken off in dissected specimens.
Pereopod VII (
Figure 10) basis–carpus length to width ratios: 3.1; 2.4; 2.7; 11.8; ischium–carpus length to basis length ratios: 0.68; 0.78; 2.53. Distal articles broken off in dissected specimens.
Pleopod I (
Figure 11) convex ventrally, flattened tip bent up; length 3.5 maximal width; widest basally, narrowest 0.2 length from base, becoming gradually wider distally toward pronounced rounded distolateral angles, located 0.1 length from apex, where pleopod width is 0.2 length; distal tip pointed. Ventral surface covered with fine setulate, proximally to lateral angles bearing stout setulate setae; distal tip with fine SS.
Pleopod II (
Figure 11) protopod length 3 width, narrowing distally, with about 10 stout UBS and numerous SS laterally. Endopod basal article length 1.9 width, and 0.19 protopod length; stylet 0.42 protopod length, distal third of stylet abruptly narrowing. Exopod length 1.2 width and 0.14 protopod length, with tuft of fine setae.
Pleopod III (
Figure 11) endopod length 2.3 width, with three plumose setae, setae length half of endopod length. Exopod length 5.3 width and 2.1 endopod length, lateral margin with fine SS; basal article length 3.3 width and 1.3 exopod length; distal article length 3.4 width, tapering distally, with straight lateral and convex medial margin.
Pleopod IV (
Figure 11) endopod length 1.3 width. Exopod length 3.7 width and 0.86 endopod length.
Pleopod V (
Figure 11) endopod length 1.3 width.
Uropod of holotype (
Figure 5) length 2.7 width, widest at 0.27 length from apex, distomedial angle oblique; distomedial margin with two stout UBS, distal margin with three BS and one trace of seta incretion.
Female, paratype:
Habitus generally similar with male (
Figure 2C–E).
Pleopod II (operculum) (
Figure 2E and
Figure 12) leaf-shaped, length 1.3 width; with numerous stout-composed setae ventrally; lateral margins with fine SS.
Manca (
Figure 2F) differing from adult individuals in less vaulted body, less inflated pleotelson, and fewer spines on body surface.
Remarks. Haplomunna kurilensis sp. nov. is similar to the second West Pacific species H. japonica (Gamô, 1983) from the Japan Trench. Both species significantly differ from the East Pacific species H. caeca (Richardson, 1905) and H. hubbsi Wilson, 1976 in having a large, much more inflated pleotelson.
Nevertheless,
H. kurilensis sp. nov. is readily distinguishable from
H. japonica (Gamô, 1983) in having the maxilliped with 5–6 retinaculae (in contrast to three retinaculae in
H. japonica), and the epipod which is about as long as basis and has abruptly narrowing distal part (in contrast to the epipod reaching about 2/3 of the basis length in
H. japonica). In
H. kurilensis sp. nov., the first article of the mandibular palp bears a stout UBS distally, while the original illustrations of
H. japonica provided by Gamô [
10] show no composed setae in this location. In addition, in
H. kurilensis sp. nov., the distal tip of the male pleopod I is pointed, with slightly pronounced distolateral angles, in contrast to the broadly rounded tip without angles in
H. japonica. Distribution. The species is known from the type locality in the NW Pacific Basin to the east of the KKT, from 43.0248° N 152.9727° E to 43.5913° N 153.9647° E at a depth of 5222–5380 m. It also possibly occurs to the west of the KKT from 46.10045° N 152.24065° E to 46.097117° N 152°242933 E at a depth of 4769–4798 m, but this information needs to be confirmed.
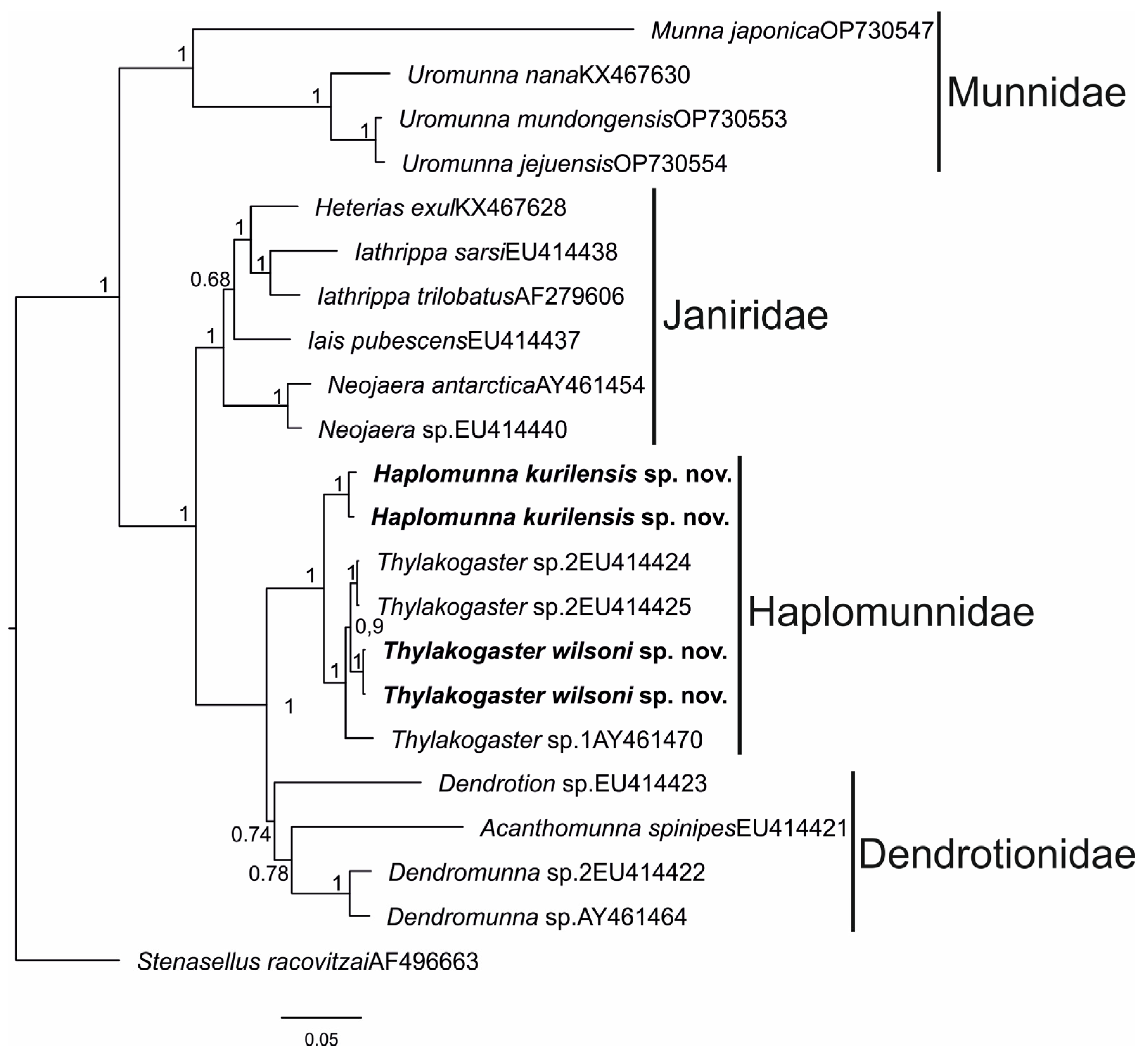
3.1.3. Genus Thylakogaster Wilson and Hessler, 1974
Type species. Thylakogaster peterpauli Wilson and Hessler, 1974.
Species included. Thylakogaster lobotourus Wilson and Hessler, 1974; T. majusculus Wilson and Hessler, 1974; T. namibiensis Brenke and Buschmann, 2009; T. peterpauli Wilson and Hessler, 1974; T. wilsoni Malyutina and Golovan sp. nov.
Thylakogaster wilsoni Malyutina and Golovan sp. nov.
This species is registered in ZooBank under urn:lsid:zoobank.org:act:2B54E2D4-0764-4B15-893B-557CB90AC529.
Material examined. Holotype: (SMF 61314), female (total length 3.0 mm, cephalothorax–pereon length 1.9 mm), KB 223-8-9.
Paratypes: (MIMB 46618) female 1 (total length 2.8 mm), female 2 (total length 2.6 mm), the same data as holotype; (SMF 61315) manca damaged, the same data as holotype; (MIMB 46619) manca (total length 1.7 mm), KB 223-7-9; (SMF 61316) manca damaged, KB 223-7-9.
Etymology. The new species is named after the American isopodologist Dr. George D.F. (Bus) Wilson, for all his contributions to the study of Asellota and, in particular, the genus Thylakogaster.
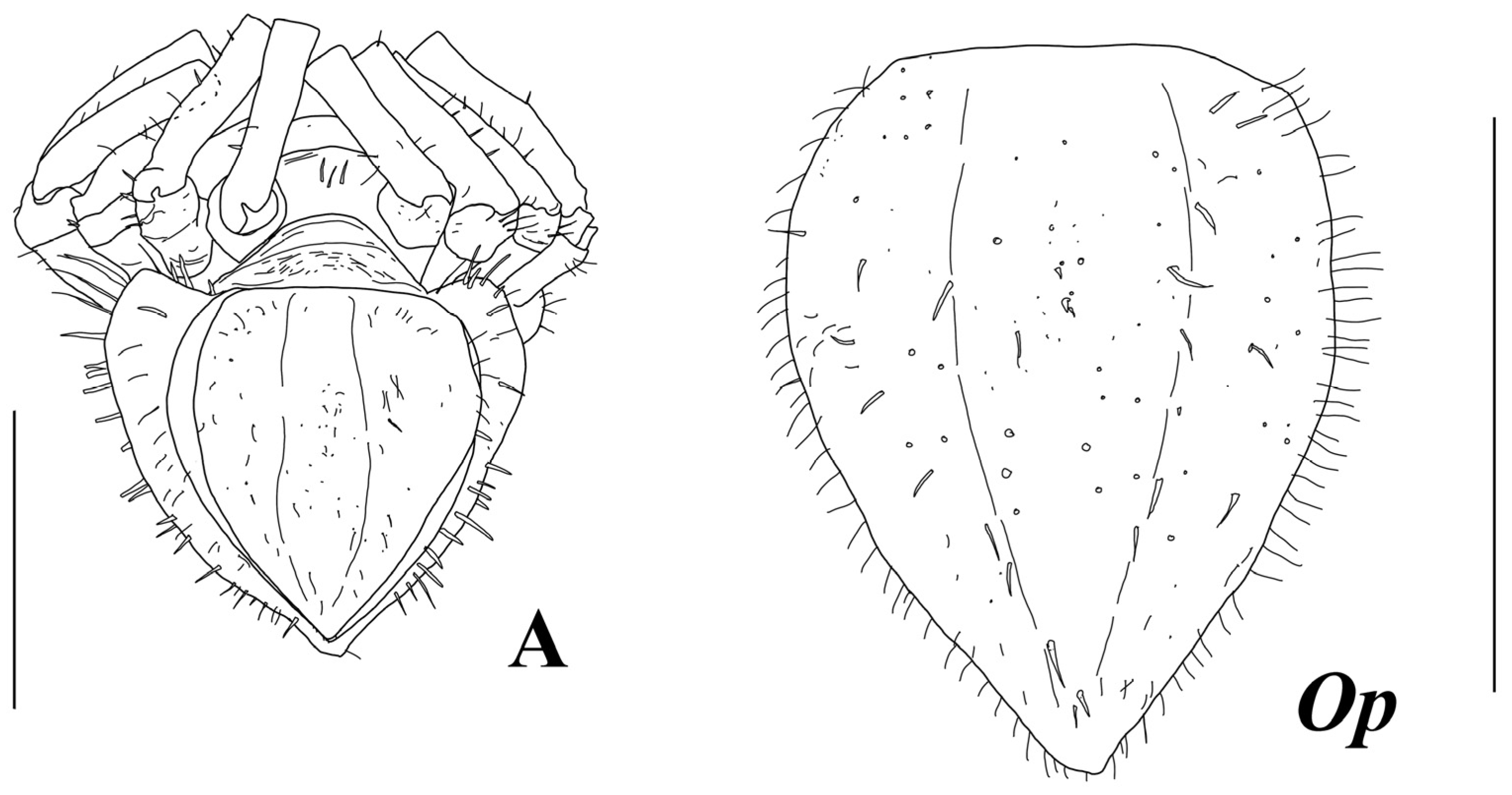
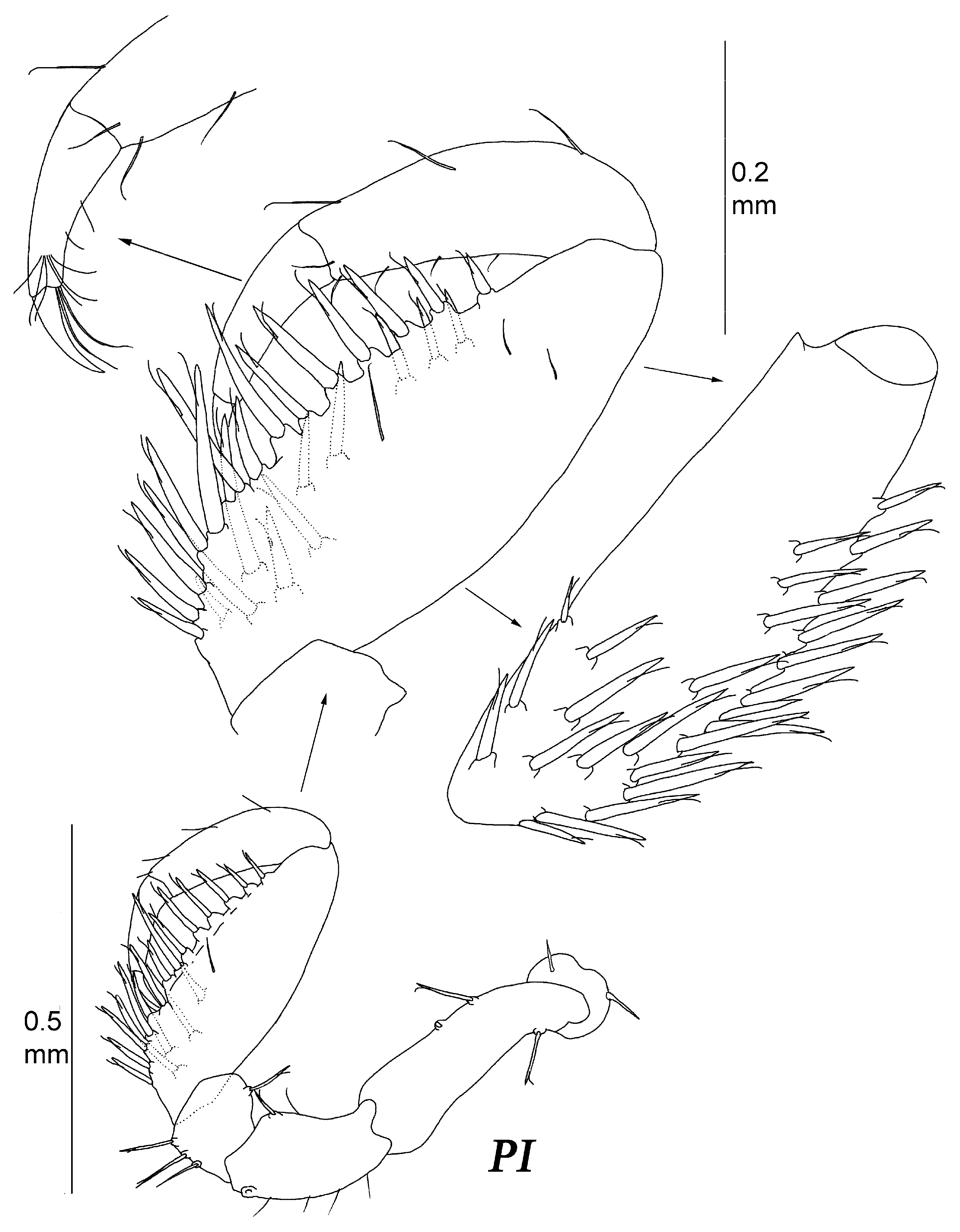
Diagnosis. Female. Body broadest and deepest at pleotelson, body total length about twice width, and cephalothorax–pereon length 1.5 pereonite 3 width. Cephalothorax weakly recessed into first pereonite. Cephalothorax–pereon dorsal surface without spines. Pleotelson length 1.5 width, 0.6 cephalothorax–pereon length, with fine cuticular spinules and lateral row of spines on dorsolateral convexities, and tip distinctly set off from lateral convexities. Antenna I about 0.5 of body total length, and article 3 about twice longer than article 2. Maxilliped endite length 1.1 width; palp article 3 length 1.1 width; epipod length 0.4 basis length. Pereopod I carpus length 3 width, about 1.3 basis length, with around 25 stout UB setae on ventral margin and medial surface. Propodus–dactylus length 0.85 carpus length. Pereopod III only slightly shorter than pereopod II. Operculum length 1.35 width.
Description. Holotype, female:
body (
Figure 13A–C) broadest and deepest at pleotelson, total length 2.0 pleotelson width, and cephalothorax–pereon length 1.5 pereonite 3 width.
Cephalothorax: weakly recessed into pereonite 1, length 0.8 width and 0.2 body length, width 0.7 width of pereonite 1, and lateral sides with angular “checks”. Frons and clypeus rounded in dorsal view and protruded forward.
Pereon: lateral expansions of pereonites 1–3 rather tightly packed, directed anteriorly; directed laterally on pereonite 4 and directed posteriorly on pereonites 5–7. Coxae rounded, visible on dorsal view, bearing 2–3 spines. Pereonite 1 length 0.5 cephalothorax length, width 0.86 cephalothorax width; length ratios of pereonites 2–7 to pereonite 1: 1.0, 1.0, 0.87, 0.75, 0.63, 0.56.
Pleon: with one pleonite, pleonite length subequal to pereonite 7 medial length, with a row of long ventral spines. Pleotelson length 0.75 width and 0.36 body total length, dorsolateral convexities highly vaulted, with longitudinal dorsal groove in between, apex rounded, and weakly produced. Pleotelson with fine spinules on dorsal and ventral surface and lateral row of rather long cuticular spines along dorsolateral convexities.
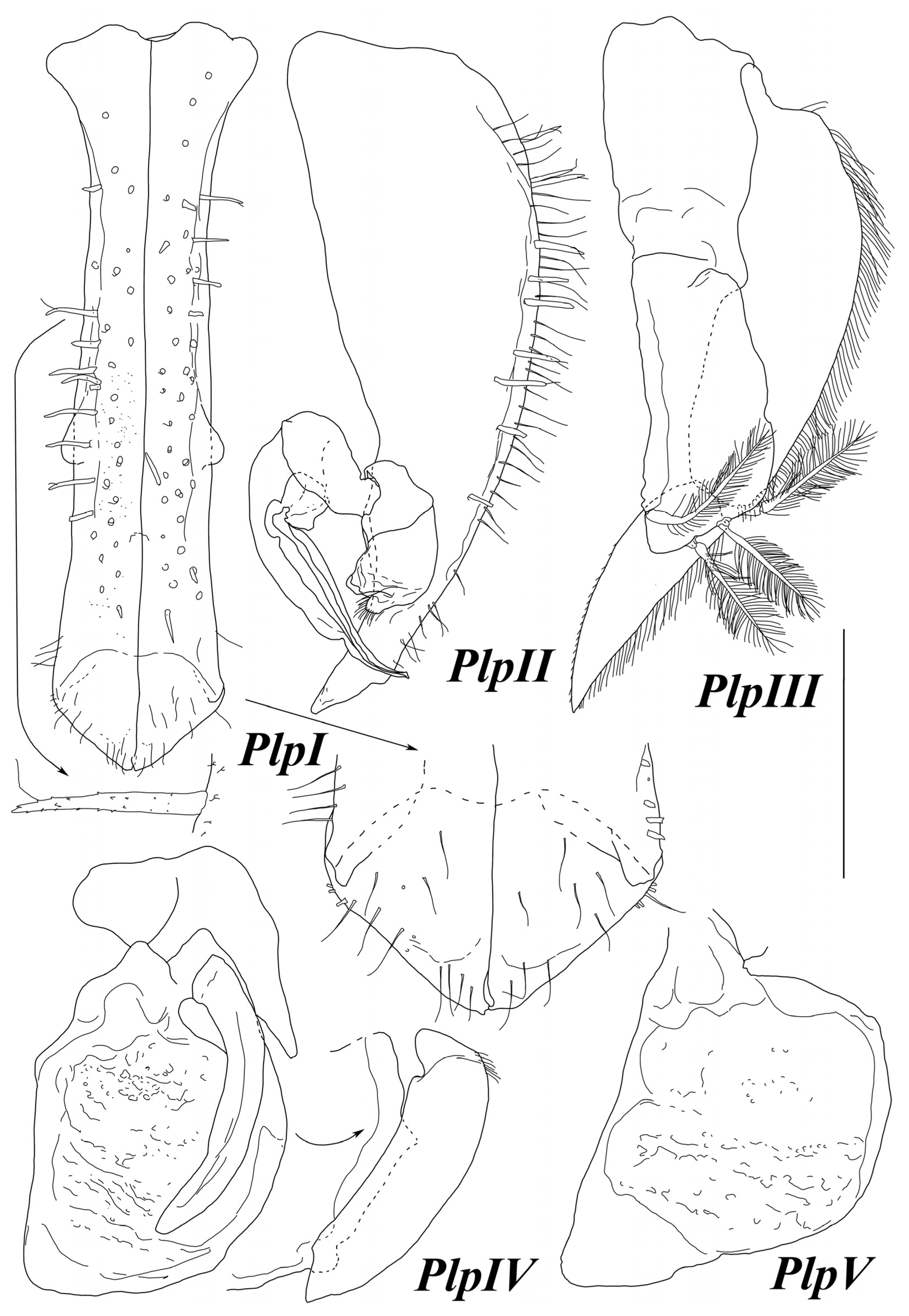
Antenna I (
Figure 14) length 0.53 body total length. Articles 1–5 length/width ratios: 1.7, 2.0, 5.7, 1.0, 8.5; length ratios of article 2–5 to article 1: 0.82, 1.41, 0.29, 2.0; following six flagellar articles subequal in length to article 2. Article 1 with ventral bump, article 2 with two distal SS, articles 3 and 4 each with one fine distal SS, some flagellar articles with fine distal seta, and the last article with two aestetascs twice longer than article.
Antenna II (
Figure 14) broken off after peduncle article 4. Articles 1–4 of similar length, and article 3 with ventromedial bump bearing one UBS. Article 4 with one small UBS medially.
Mouth parts of small female, paratype:
Mandibles (
Figure 14): Incisor process of both mandibles broadened distally, with five cusps. Spine row with five and six serrated setae on left and right mandibles, respectively.
Lacinia mobilis of left mandible broadened distally, with four cusps, length 0.83 incisor process length. Molar process almost as long as incisor, cylindrical, with seven long distal setae and condyle length about half of molar length.
Maxilla I (
Figure 15) lateral endite distally with 12 differently serrated setae; mesial endite distally with three strong SS.
Maxilla II (
Figure 15) lateral and middle endites together almost as broad as mesial endite; all endites with strong distal SS. Lateral endite longest, with two long and two short SS, and middle endite with one long and three short SS. Mesial endite shortest, with five long and two short SS distally and two fine SS medially.
Maxilliped (
Figure 15) basis length 1.54 width, endite as long as wide, 0.65 basis length, with two retinacula, and distal margin with six SS. Palp narrow, gradually tapering, length 1.37 basis length; article 1 width 0.42 basis width; article 5 0.4 of article 1 width; articles 1–5 length to width ratios: 0.41, 0.66, 1.12, 2.33, 3.57; articles 2–5 lengths to article 1 length ratios: 1.42, 2.42, 4.0, 3.57. Articles 1–3 with one long medial SS, and article 3 additionally with one distolateral SS; article 4 with two distomedial SS, and article 5 with 5 distal SS. Epipod triangular, width 1.1 length, 0.47 basis width, and length 0.38 basis length.
Pereopod I (
Figure 16) robust, subchelate, length 0.46 body length, and length ratios of ischium–dactylus to basis: 0.62, 0.32, 1.28, 0.72, 0.35. Basis to dactylus length/width ratios: 2.68, 1.69, 0.78, 3.05, 3.1, 2.1. Basis with three stout short UBS. Ischium with one ventral, one dorsal stout short UBS, and few SS. Merus with two SS on ventral margin and one UBS on dorsal margin. Carpus with 13–14 stout UBS on outer ventral margin; medial surface with longitudinal row of five UBS near outer ventral margin and six UBS proximally. Propodus with three ventral and two dorsal SS. Dactylus with two fine SS, claw length 0.7 dactylus length, two SS ventrally near claw, and tuft of six SS proximally to claw. Tip of claw reaching proximal quarter of carpus when folded against it.
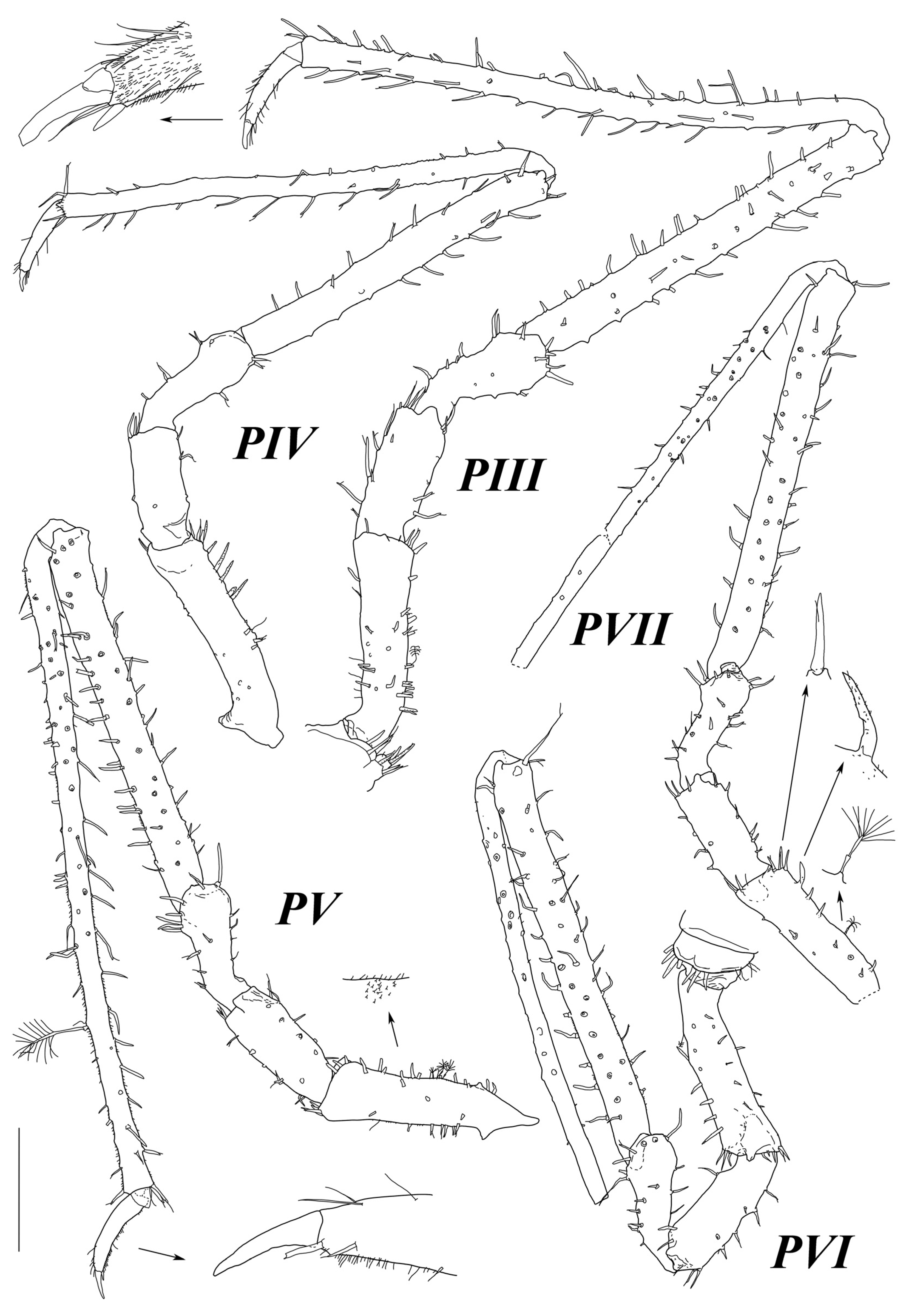
Pereopod II (
Figure 16) with broken dactylus. Length ratios of ischium–propodus to basis: 0.6, 0.55, 2.0, 2.52, 0.35. Basis length 1.44 basis of pereopod I length. Basis to dactylus length/width ratios: 3.87, 1.17, 2.12, 9.54, 17.5. Ischium with one dorsal and three ventral UBS. Merus with two ventral and two dorsal UBS. Carpus with around 25 long UBS. Propodus with >30 UBS.
Pereopod III (
Figure 16) length 2.2 pereopod I length, length without dactylus 0.95 P II length without dactylus. Length ratios of ischium–dactylus to basis: 0.52, 0.53, 1.82, 1.86, 0.42. Basis to dactylus length/width ratios: 3.9, 2.26, 2.33, 10.0, 13.7, 8.0. Ischium with five ventral UBS and few small SS. Merus with one ventral and two dorsal UBS. Carpus with about twenty UBS and two distodorsal SS. Propodus with 25 UBS, with about 10 SS distally. Dactylus with few fine SS; tip of claw broken off.
Pereopods IV–VII broken off; bases of subequal length and slightly narrower than basis of pereopod III.
Pereopod I of large female, paratype (
Figure 17) length 0.51 body length, length ratios of ischium–dactylus to basis: 0.67, 0.34, 1.28, 0.83, 0.39. Basis to dactylus length/width ratios: 2.68, 1.69, 0.78, 2.75, 3.1, 2.1. Basis length 3 width, with three UBS. Ischium with few SS and one ventral and one dorsal stout short UBS. Merus with two SS on ventral margin and one UBS on dorsal margin. Carpus with fourteen strong UBS on outer ventral margin; medial surface with longitudinal row of five UBS on outer ventral margin and six UBS proximally. Propodus with six ventral and three dorsal SS. Dactylus length approximately 0.5 propodus length. Dactylus with two fine SS ventrally, tuft of six SS proximally to claw, with claw about half of dactylus length and two long ventral SS.
Pereopods of manca, female, paratype (
Figure 18).
Pereopod I length 0.5 body length, length ratios of ischium–dactylus to basis: 0.55, 0.40, 1.25, 0.82, 0.42. Basis to dactylus length/width ratios: 3.1, 1.57, 1.06, 3.33, 3.0. Basis with two dorsal UBS. Ischium with two ventral and one dorsal stout UBS. Merus with three ventral and one dorsal UBS. Carpus with eight strong UBS on outer ventral margin; medial surface with longitudinal row of five UBS along outer ventral margin and six UBS proximally. Propodus with four ventral and three dorsal SS. Dactylus claw about 1/3 of dactylus length.
Pereopod II length 1.85 pereopod I length. Length ratios of ischium–dactylus to basis: 0.57, 0.72, 1.91, 2.0, 0.61. Basis to dactylus length/width ratios: 2.9, 1.68, 2.26, 7.5, 11.75, 7.25. Basis with three dorsal UBS. Ischium with five ventral UBS and few small SS. Merus with one ventral and two dorsal UBS. Carpus with about twenty UBS and two distodorsal SS. Propodus with 25 UBS, with about 10 SS distally. Dactylus with few fine SS.
Pereopod III broken off after basis. Basis length 1.27 basis II length and 4.0 width.
Pereopod IV length 2.95 pereopod I length and 1.63 pereopod II length. Length ratios of ischium–dactylus to basis: 0.62, 0.76, 1.97, 2.70, 0.90. Basis to dactylus length/width ratios: 4.7, 3.15, 3.12, 14.4, 29.66, 15.0. Basis with three dorsal UBS and two distal SS. Ischium without setae. Merus with one ventral, two distoventral UBS, and one distodorsal UBS. Carpus with about ten UBS and one strong distodorsal BS (broken in specimen). Propodus with about 20 UBS and few SS. Dactylus with few fine SS and one long SS distoventrally; claw slender, with length 0.5 dactylus length.
Pereopod V (
Figure 17) length subequal to pereopod IV length (0.99). Length ratios of ischium–dactylus to basis: 0.61, 0.76, 1.88, 2.52, 0.88. Basis to dactylus length/width ratios: 4.85, 3.0, 3.47, 14.22, 28.66, 15.0. Basis without setae. Ischium with one ventral UBS. Merus with one ventral, one distoventral, and one distodorsal UBS. Carpus with about ten UBS and one strong distodorsal BS (broken in specimen). Propodus with eighteen UBS and one long plumose seta dorsally. Dactylus with few fine SS and one long seta distoventrally; claw slender, with length 0.5 dactylus length.
Pereopod VI broken off after basis. Basis length equal to bases IV and V length 5.5 width.
Pereopod VII (
Figure 17) not developed, with six separated articles.
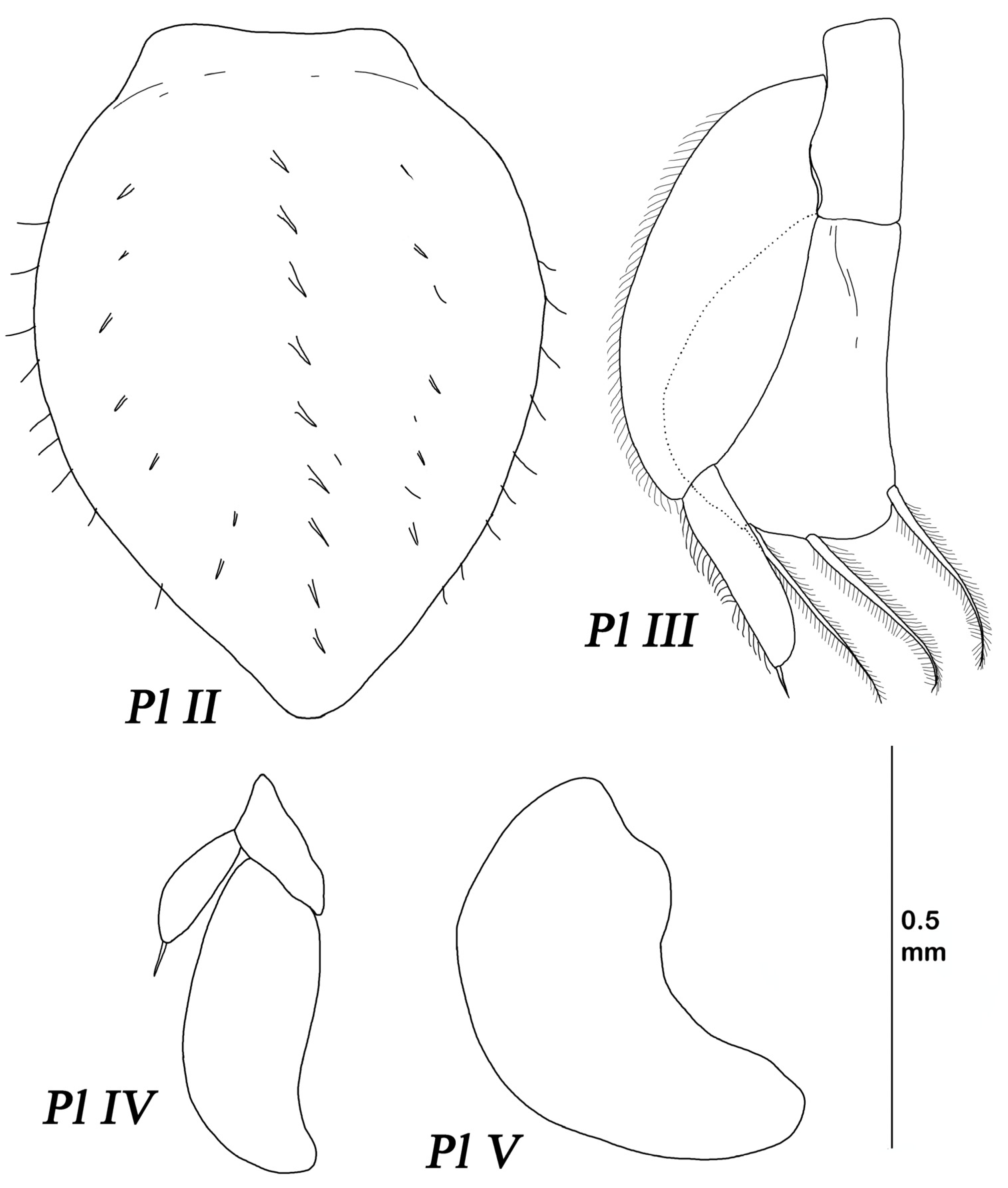
Pleopod II length 1.35 width, ventrally with medial row of spines like on lateral margins of pleotelson, with marginal small setae.
Pleopod III protopod length 2.36 width; endopod length 1.46 width, 1.58 protopod length, width 2.55 protopod width, and distal margin with three plumose setae, which is about 0.65 of endopod length. Exopod is twice longer than endopod, with fine setulae laterally, basal article length 2.71 width, 1.39 endopod length, distal article length 4.33 width, and 0.45 basal article length, with one short terminal SS.
Pleopod IV endopod length 2.87 width. Exopod length 3.2 width and 0.34 endopod length, with one short terminal SS.
Pleopod V endopod length 1.96 width.
Uropod of holotype (
Figure 13) length 2 width, distolateral lobe rounded, and twice longer and broader than tiny distomedial lobe, with one short terminal SS and distomedial lobe acute, with three terminal SS.
Remarks. The new species can be distinguished from four known species of the genus: T. lobotourus Wilson and Hessler, 1974, T. majusculus Wilson and Hessler, 1974, T. peterpauli Wilson and Hessler, 1974, and T. namibiensis Brenke and Buschmann, 2009 by the less spiny body, shape, and proportions of segments and appendage articles. In general, T. wilsoni sp. nov. has the broadest pleotelson of all known species from the Atlantic. In contrast to other species, pleotelson tip of the new species distinctly set off from the lateral convexities; by this character, T. wilsoni sp. nov. is close to T. namibiensis. Antenna I article 3 in T. wilsoni sp. nov. is about 2.5 times longer than article 2, while in females of T. namibiensis and T. lobotourus, these articles are subequal in length; in T. peterpauli and T. majusculus, this ratio is 1.6 and 1.8, respectively. In contrast to T. lobotourus, pereopod II of adult specimen of T. wilsoni sp. nov. is not distinctly longer than pereopod III, though T. wilsoni sp. nov. has carpus and propodus of pereopod II longer than these two articles of pereopod III (1.03 and 1.14, respectively). Unfortunately, we could not compare pereopod II with pereopod III in manca as it was broken. Pereopod I of the new species has relatively longer propodus–dactylus concerning carpus: 0.9–0.8, while in T. lobotourus, this ratio is 0.7-0.75; carpus is more elongated in T. wilsoni sp. nov.: length/width ratio is 2.75–3.05, while in T. lobotourus it is 2.1–2.5.
Maxilliped is an important structure for morphological comparison of species. In T. wilsoni sp. nov., the endite length/width ratio is 1.1, and the palp article 3 length/width ratio is also 1.1, while in T. peterpauli and T. majusculus, the ratio of endite is 0.8 and 0.9, respectively, and the ratio of palp article 3 1.9 and 1.7, respectively. By these two ratios, the new species is close to T. namibiensis, though differs from it by the short epipod (length/basis length is 0.4 in T. wilsoni sp. nov. and 0.7 in T. namibiensis).
Distribution. The species is known only from the type locality in the NW Pacific Basin to the east of the Kuril–Kamchatka Trench, 42.2447° N 151.7351° E 42.2378° N 151.7082° E at a depth of 5125–5126 m and 43.0473°N 152.9905° E to 43.0248° N 152.9727° E at a depth of 5222–5223 m.
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}