
Chaenothecopsis xishuiensis sp. nov. to Science and Lecanora pseudargentata Newly Reported from China
1
College of Life Sciences, Liaocheng University, Liaocheng 252059, China
2
Pharmacology Unit, Center for Neuroscience, School of Pharmacy, University of Camerino, 62032 Camerino, Italy
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(8), 893; https://doi.org/10.3390/d15080893
Submission received: 2 June 2023
/
Revised: 6 July 2023
/
Accepted: 25 July 2023
/
Published: 28 July 2023
(This article belongs to the Special Issue Phylogeny, Taxonomy and Ecosystems of Lichens)
Abstract
:In order to provide data for lichenologists studying taxonomy, Chaenothecopsis xishuiensis is supported and proposed as a new species from China based on phenotypic, molecular, and metabolite data. It is characterised by leprose thallus, single, conical to hemispherical apothecia, nonbranching stipe, cylindrical, eight-spored asci, and nonseptate and brown spores, and this lichenised fungus contains atranorin and zeorin in the thallus. In addition, Lecanora pseudargentata is reported for the first time as a new record from China. This species is characterised by red-brown to dark brown apothecial discs, eight-spored asci, nonseptate, hyaline spores, and the presence of atranorin and gangaleoidin. The biological activity of its lichen substances is discussed.
1. Introduction
Lichen is a mutually beneficial symbiotic complex of fungi and algae (or cyanobacteria) [1]. It is the pioneer of the Earth [2]. They can grow in extreme environments and adapt to their habitat mainly in two main ways: genes and metabolites. For example, some species that grow at high latitudes or altitudes have cold resistance genes, and some species that grow in deserts have drought tolerance genes, which is their genetic function. On the other hand, secondary metabolites and lichen polysaccharides have enormous medicinal properties, such as anticancer, antibacterial, antioxidant, human immunomodulatory properties, and so on. More and more values are being unlocked [3,4,5]. They are important resources for drug discovery and development. However, due to the characteristics of limited lichen resources and slow growth, scientists have rarely studied and exploited them, and many species are still unknown. There is a strong need for taxonomic studies on lichens to provide a basis for their further application. Therefore, we carried out taxonomic studies on some species belonging to Chaenothecopsis and Lecanora based on phenotypic, molecular, and metabolite data.
Lichenicolous fungi are highly parasitic organisms that develop on lichens [6,7]. They obtain fixed carbon from living lichens by forming haustoria [7,8]. To date, approximately 2000 obligate lichenicolous species and more than 60 facultative lichenicolous species have been identified [7]. These species are found in different classes and genera [7]. Although they have been studied for centuries, only 50 lichenicolous fungi have been reported from China, which means that many new species collected in China are waiting to be discovered [6].
Chaenothecopsis Vain., belonging to Sphinctrinaceae, Mycocaliciales, Mycocaliciomycetidae, Eurotiomycetes, Ascomycota, was established by Vainio in 1927 and has a cosmopolitan distribution [9,10,11,12,13]. The genus is characterised by the sessile or stalked ascomata, mostly obovoid to lenticular capitulum, cylindrical to subclavate, less than 60 μm long, and eight-spored asci with strongly thickened apex penetrated by a thin channel, and nonseptate or one-septate, nearly hyaline to brown spores [9,14]. A number of Chaenothecopsis species are lichenicolous fungi, growing as parasites or commensals on lichens, or lichenised fungi, while others are parasitic on algae, conifer resins, or exudates of vascular plants. And many species are highly specialised, producing ascomata only on specific substrates [3,9,14,15]. More than 50 species of Chaenothecopsis have been reported worldwide, 20 of which were found in China prior to this report [10,16,17].
Lecanora Ach. is a genus of lichenised fungi belonging to Lecanoraceae, Lecanorales, Lecanoromycetes, Ascomycota [10,18] with a worldwide distribution [19]. The genus is characterised by its crustose to scaly shell-shaped thallus, lecanorine or biatorine ascomata, lecanora-type and 8–32-spored asci, nonseptate and hyaline spores, the presence of atranorin and usnic acid, and green algae as photobionts [19,20,21,22], growing on bark, rock, wood, soil or detritus [19]. Lecanora comprises about 1000 species worldwide [18,19,23]. Previously, 119 species were known from China [24,25,26]. However, due to the large variation in morphological and chemical characteristics of the genus, the traditional concept of Lecanora corresponds to blurred species boundaries [22,27,28]. Since the 20th century, the concept of this genus has been somewhat stabilised by the introduction of molecular data [22,28,29,30,31,32,33]. The importance of combining morphological, chemical, and molecular data to delineate species within the genus has been highlighted [22,28,34]. In 2023, Santos et al. confirmed that the phylogenetic tree constructed from ITS sequences was a good method for classifying the species in the genus Lecanora [34].
2. Materials and Methods
2.1. Materials Specimens
Three Chaenothecopsis specimens were collected from Xishui National Nature Reserve, and one Lecanora specimen was collected from Sanqing Mountain. All the specimens are deposited in the Fungarium of the College of Life Sciences, Liaocheng University, China (LCUF).
2.2. Morphological Study
The external morphology of the thallus and apothecia was observed and measured under a dissecting microscope (OLYMPUS SZX16) while the characteristics and data were recorded. The observations included the growth type, colour and texture of the thallus, the shape, colour, size, mode of attachment, and disc situation of the apothecia.
2.3. Anatomical Study
Apothecia were cut longitudinally and filmed under a dissecting microscope (OLYMPUS SZX16), and their internal structures were later observed under a compound microscope (OLYMPUS BX53). The operation steps are as follows:
- Moisten the selected apothecia: Apply an appropriate amount of sterile water to the well-developed apothecia with a rubber-tipped dropper and remove the ascospores after the apothecia have absorbed the water and become soft.
- Slice: Using a single-sided blade, cut the selected apothecia longitudinally so that the slices are as thin and complete as possible.
- Production: Pick up the section with the tip of a needle on a slide moistened with sterile water, cover it with a coverslip and absorb excess water with absorbent paper.
- Observation and recording: Observe the internal structure of the apothecia under a compound microscope (OLYMPUS BX53), including the colour and thickness of exciple; the colour and thickness of epithecium, hymenium, and hypothecium; the shape, size of asci, and the number of ascospores contained in them; the colour, size, and type of ascospores. Take photographs and record the relevant information.
- Colour development: Stain the prepared mount with iodine solution and observe the amyloplastic reflection of the ascospores.
2.4. Methods for the Determination of Lichen Secondary Metabolites
The metabolites of lichens were determined using chemical chromogenic reaction assays (CT) and thin layer chromatography (TLC) assays [3,4,35,36,37].
For CT, aqueous solutions of 10–25% potassium hydroxide (K), saturated aqueous solution of calcium hypochlorite (C), and 4% ethanolic solution of p-phenylenediamine (P) reagents were added dropwise to the cortex of the thallus using capillary tubes, and the colour reactions were observed and recorded.
TLC was performed according to the method of Culberson and Kristinsson, slightly modified by Jia and Wei, using Lethariella cladonioides (Nyl.) Krog as the standard sample and the C-solvent system (toluene:acetic acid = 200:30 mL) as the spreading agent [3,4,35,36,37]. The operation steps are as follows:
- Prepare the glass silicone adhesive board. Use a precoated glass silicone backing sheet (20 cm long, 10 cm wide, 0.25 mm thick). Using a pencil, carefully draw a straight line 1.5 cm from the bottom of the glass silicone board. Mark a point every 1 cm on the straight line, which will be the sample point.
- Prepare the solvent. Mix 20 mL of toluene and 3 mL of acetic acid, add to a rectangular TLC developing tank, and place in a fume cupboard.
- Prepare the samples. Take an appropriate quality of the thallus to be examined and place them separately in small centrifuge tubes. Add a suitable volume of acetone to each small centrifuge tube until the acetone covers the sample. After 10 min, the samples can be placed in order.
- Spot sampling: Use microcapillary tubes to sample separately according to the position of the sampling points on the glass silicone board. The left, right, and centre sampling points are brushed with Lethariella cladonioides to facilitate the use of split standard samples, while the remaining sampling points are sampled sequentially for testing.
- Exposure layer: After sampling, place the silicone board in a chromatography cylinder and place it 1 cm below the solvent level so that the sample origin is approximately 0.5 cm from the solvent level and make the origin line parallel to the liquid level line. When the leading edge of the solvent moves from the origin to about 1.5 cm from the top of the silicone board, remove the silicone board and dry the solvent on the board surface with a hair dryer.
- Colour rendering: Spray the silicone board with 10% sulphuric acid and observe if there are any grease spots when it is wet. Then, heat it in an oven at 94 °C for about 10–15 min until the chromatography develops well. Observe and record the colour and position of the spots under white and ultraviolet light, respectively.
- Partition: Draw tangents at one point above and one point below the chromatographic origin for colour display. The area between the top and bottom tangents is the first zone. Using the same method, draw the fourth and seventh zones of the atranorin and norstictic acid stains, respectively. Then, draw a median line between the first and fourth zones, dividing them equally into the second and third zones; draw the fifth and sixth zones between the fourth and seventh zones using the same method.
2.5. DNA Extraction, PCR Amplification, and Sequencing
Collect some apothecia and thallus into 1.5 mL centrifuge tubes using a sterilised blade and tweezers. DNA was extracted using the Hi-DNAsecure Plant Kit according to the procedure in the slightly modified instructions, as follows:
- Collect apothecia and thallus in microcentrifuge tubes and add beads for thorough grinding. Add 400 μL FGA buffer and 6 μL RNaseA (10 mg/mL), vortex for 1 min, and leave for 10 min at room temperature.
- Add 130 μL LP2 buffer, mix thoroughly and vortex for 1 min.
- Centrifuge at 12,000 rpm (~13,400× g) for 5 min and transfer 300 μL of supernatant to a new tube.
- Add 1.5 times the volume of buffer LP3 and immediately vortex for 15 s.
- Add a total of 750 μL of the solution obtained in the previous step and the flocculated precipitate to an adsorbent column CB3, centrifuge at 12,000 rpm (~13,400× g) for 30 s, pour off the waste solution and place the column CB3 in a collection tube.
- Add 600 μL of rinsing solution PW to column CB3, centrifuge at 12,000 rpm (~13,400× g) for 30 s, pour off the waste solution, and place column CB3 in the collection tube. Repeat this step twice.
- Place the column CB3 back into the collection tube and centrifuge at 12,000 rpm (~13,400× g) for 2 min, discarding the waste solution. Leave the column CB3 at room temperature for a few minutes to thoroughly dry the residual rinse from the adsorbent material.
- Transfer the adsorbent column CB3 to a clean centrifuge tube, add 90 μL of elution buffer TB dropwise to the centre of the adsorbent membrane in suspension, allow to stand at room temperature for 2–5 min, centrifuge at 12,000 rpm (~13,400× g) for 2 min, and collect the solution in the centrifuge tube. Aspirate the solution from the centrifuge tube, drop it back onto the centre of the adsorbent membrane, leave at room temperature for 2–5 min, centrifuge at 12,000 rpm (~13,400× g) for 2 min, and collect the solution in a centrifuge tube. This solution contains the extracted genomic DNA.
The internal transcribed spacer (ITS) region of ribosomal DNA was amplified using ITS1F/ITS4 primers [38,39]. Polymerase chain reactions (PCR) were reacted in a volume of 50 µL mixture containing 25 µL 2 × PCR Master Mix, 17 µL ddH2O, 2 µL of each primer, and 4 µL DNA template. PCR thermal cycles were performed with the following reaction conditions: initial denaturation at 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 52 °C for 30 s, extension at 72 °C for 1.5 min, and a final extension at 72 °C for 10 min. The PCR products were purified and sequenced at Biosune Biotechnology Company (Jinan, China).
2.6. Alignment and Phylogenetic Tree Construction
ITS sequences were generated from the Chaenothecopsis specimen and Lecanora specimen. According to the results of BLAST on the NCBI website (https://www.ncbi.nlm.nih.gov/ (accessed on 27 April 2023)) and related research, other similar taxa sequences from different genera of Sphinctrinaceae were downloaded from GenBank (Table 1) [19,40,41]. Talaromyces acaricola Visagie, N. Yilmaz & K. Jacobs and Protoparmelia ochrococca (Nyl.) P.M. Jørg., Rambold & Hertel were used as outgroups, respectively. The selected ITS sequences, together with newly generated sequences, were aligned in BioEdit using the ClustalW method [42,43].
Phylogenetic relationships were inferred using maximum likelihood (ML) and Bayesian inference (BI) on the CIPRES Scientific Gateway portal (http://www.phylo.org/ (accessed on 24 April 2023)). ML analyses were performed using RAxML-HPC2 on XSEDE (8.2.12) with 1000 replicates as bootstrap analysis. BI analyses were performed using MrBayes on XSEDE (3.2.7a) based on the GTR+I+G model, with 2 independent analysis runs for 1 million generations. Each run included four chains, parameters were sampled every 1000 generations and 25% were discarded as burn-in. The remaining 75% were used to calculate the consensus tree [40,44,45,46]. Bootstrap support above 70% and posterior probabilities above 0.9 were considered significant support values. Trees were visualised in FigTree v.1.4.2 (http://tree.bio.ed.ac.uk/software/figtree (accessed on 27 April 2023)) and MEGA11 [47]. Phylogenetic trees are shown in Figure 1 and Figure 2.
3. Results
3.1. Phylogenetic Analysis
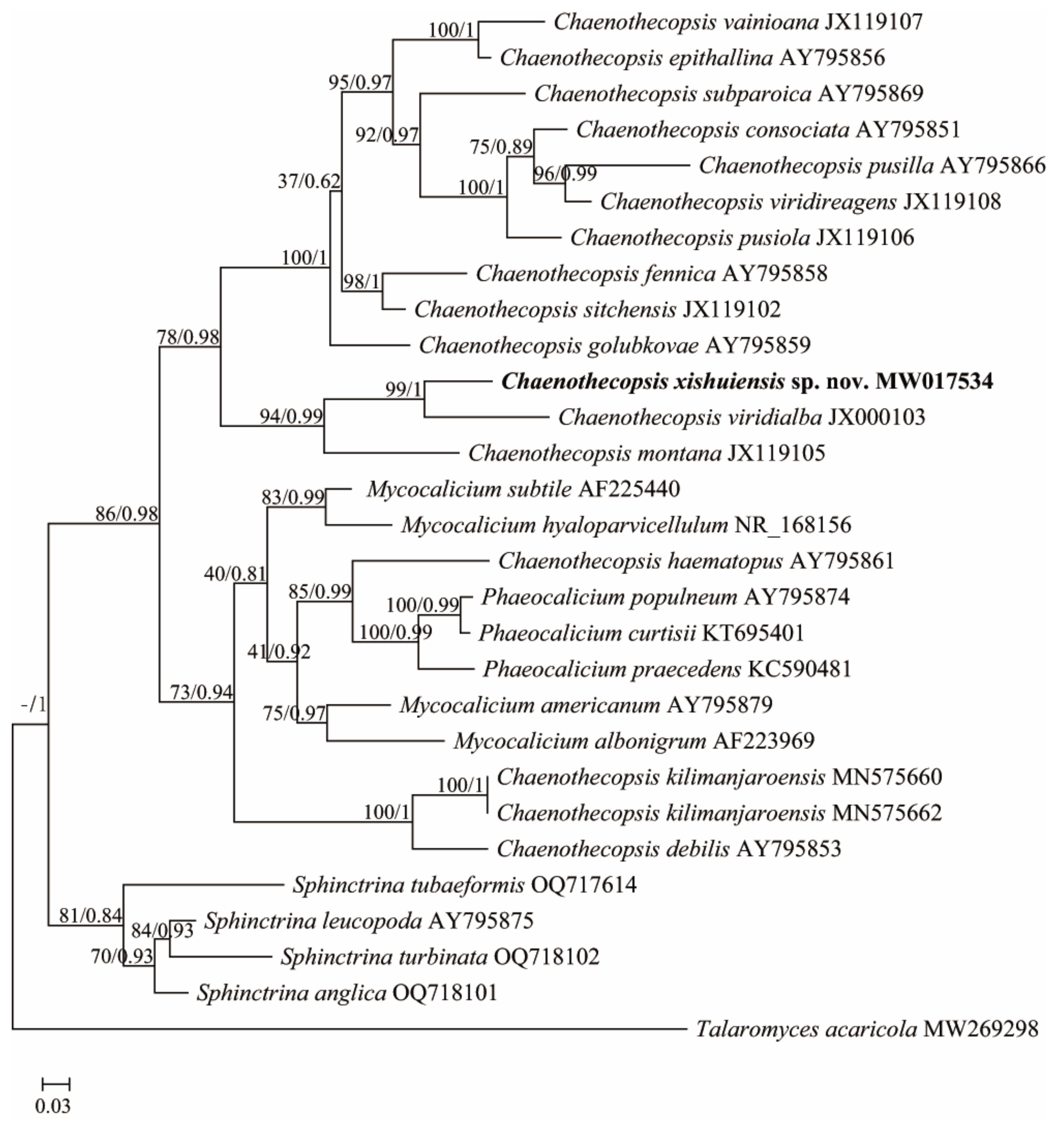
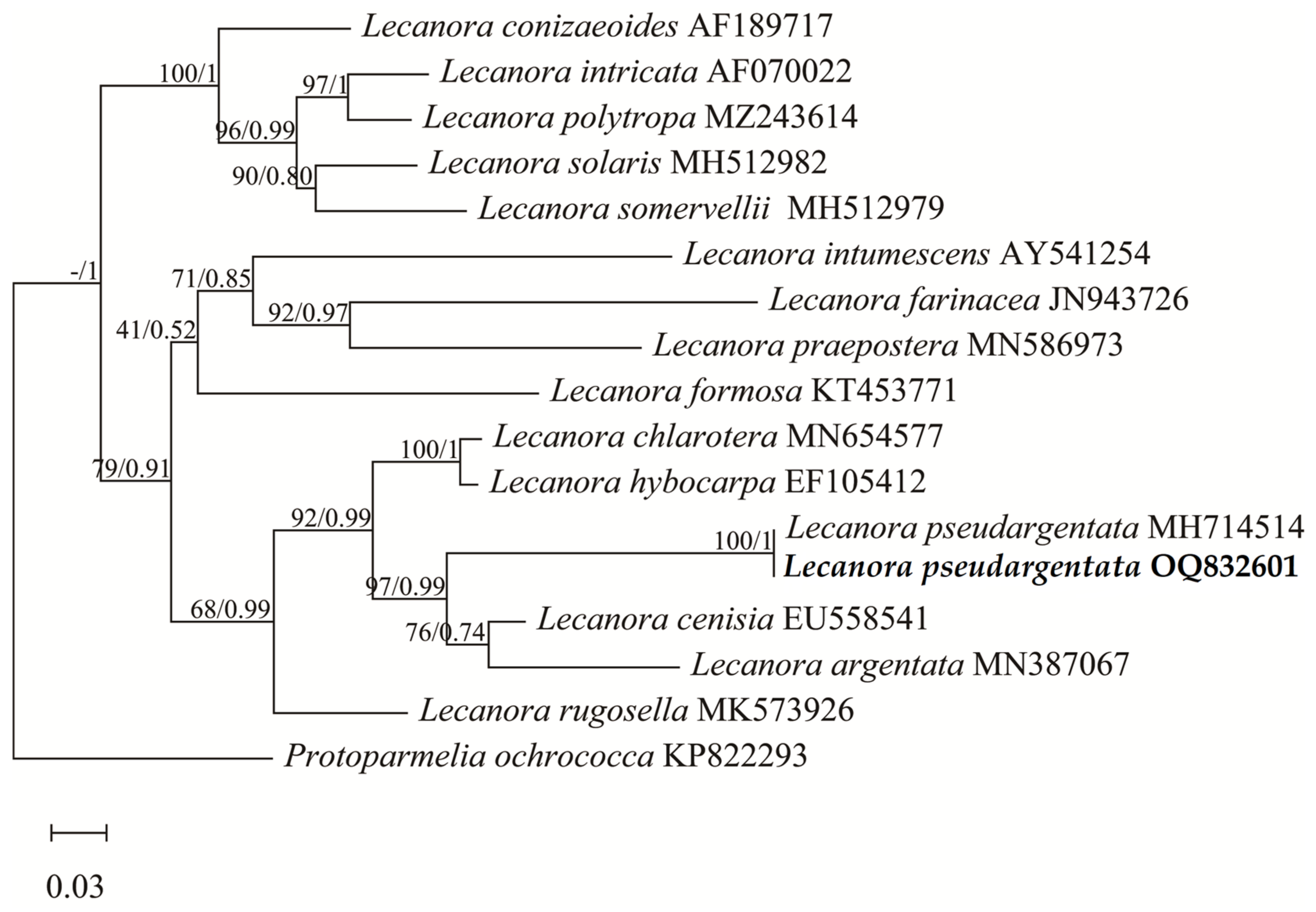
The Sphinctrinaceae s. str. phylogenetic tree dataset included 1 newly generated ITS sequence submitted to GenBank under accession number MW017534, plus 28 sequences downloaded from GenBank (Table 1). The Lecanora phylogenetic tree data set included 1 newly generated ITS sequence submitted to GenBank under accession number OQ832601, plus 16 sequences downloaded from GenBank (Table 1). The Sphinctrinaceae s. str. dataset and Lecanora dataset were used to construct phylogenetic trees, respectively. Since the topologies of the maximum likelihood tree and Bayesian Inference tree are congruent, maximum likelihood bootstrap probabilities (ML-BS) and Bayesian inference posterior probabilities (BI-PP) are combined and placed at the node of the maximum likelihood tree. ML-BS are on the left, and BI-PP are on the right. The results are shown in Figure 1 and Figure 2, respectively.
Within the Sphinctrinaceae s. str. phylogenetic tree, the newly offered Chaenothecopsis specimen forms a sister group with Chaenothecopsis viridialba (Kremp.) in an independent clade with other species (Figure 1). Based on the differences in phylogeny and morphology compared to other species, which are described in detail below, it is classified as a new species named Chaenothecopsis xishuiensis.
In Figure 2, the ITS sequences of the newly offered Lecanora specimen and Lecanora pseudargentata Lumbsch downloaded from GenBank are clustered together in the phylogenetic tree. Combined with the morphological similarity, we identified this specimen as Lecanora pseudargentata, which is a new record in China.
3.2. Taxonomy
Chaenothecopsis xishuiensis Z.F. Jia, sp. nov., Figure 3.
It differs from Chaenothecopsis kilimanjaroensis mainly by its single apothecia, wider and nonseptate ascospores.
TYPE: China. Guizhou: Zunyi City, Xishui County, Sanchahe Town, Chinese Danxia Valley, 28°33′ N 106°24′ E, alt. 1040 m, on bark, 17/XI/2019, M. L. Zhu GZ19513 (LCUF, holotype).
MycoBank No: 848ht5588
ETYMOLOGY: The specific epithet xishuiensis is derived from the type locality, xishui, and –ensis, Latin.
THALLUS: corticolous, leprose, irregular, and its surface is whitish-green to yellowish-green.
APOTHECIA: brownish black, single, stalked, conical to flattened flabelliform to hemispherical, 0.1–0.25 mm in diameter, epruinose; STIPE: brown, with pruina, nonbranching, 0.2–0.3 mm high, 31–77 µm wide, K−, in section the central part composed of irregularly intertwined hyphae; STIPE HYPHAE: intertwined, elongated, hyaline to lightly brown, 2.2–4.4 µm wide. EXCIPLE: dark brown, 20–23 µm thick; EPITHECIUM: dark brown, 10–12 µm tall; HYMENIUM: 52–65 µm tall, hyaline to brownish, oil droplets absent, K+ yellowish green, I–; PARAPHYSES: hyaline, filiform; HYPOTHECIUM: brown, convex, 33.5–43.5 µm tall. ASCI: cylindrical with a thickened apex penetrated by a fine canal, 39–45 × 5.6–7.3 µm, eight-spored. ASCOSPORES: uniseriate, brown, fusiform, nonseptate, 6.5–8 × 2.9–4.5 µm, I–. PYCNIDIA not observed.
CHEMISTRY: Thallus K+ yellow, C−, P+ blue-grey. Atranorin and zeorin were detected in the thallus by TLC.
ECOLOGY AND DISTRIBUTION: on bark in forests and are only known from the type locality.
ADDITIONAL SPECIMEN EXAMINED: China. Guizhou: Zunyi City, Xishui County, Sanchahe Town, Chinese Danxia Valley, alt. 1040 m, on bark, 17/XI/2019, M. L. Zhu GZ19514, GZ19515 (LCUF).
REMARKS: Chaenothecopsis xishuiensis is characterised by leprose thallus, single, conical to hemispherical apothecia, nonbranching stipe, cylindrical, eight-spored asci, nonseptate and brown spores, and it is a lichenised fungus containing atranorin and zeorin in the thallus.
The closest species phylogenetically are Chaenothecopsis viridialba and Chaenothecopsis montana (Figure 1). However, these species are morphologically clearly different. C. viridialba has whitish pruina or granules on longer-stalked (2–3 mm) apothecia and ellipsoid spores (6–10 × 2.5–3.5 µm) [9,48], whereas C. montana has narrower asci (3.5–5.5 mm), ellipsoid to ovoid ascospores, and all parts of apothecium K− [49].
Morphologically, the new species is similar to Chaenothecopsis kilimanjaroensis Temu & Tibell and Chaenothecopsis pusiola (Ach.) Vain., but the latter two are far from the new species in the phylogenetic tree. Moreover, C. kilimanjaroensis has 2–5 or single aggregated capitula, one-septate, and narrower ascospores (2.1–2.6 µm wide) [41]; C. pusiola has black and higher stalks (0.2–0.6 mm), one-septate spores, and lichenicolous on species of Chaenotheca (Th. Fr.) Th. Fr. [14].
As some species of Chaenothecopsis are described based on phenotype only, no sequence data are available. Therefore, we compare the new species with other available phenotypic information in the Chaenothecopsis genus species, with the result that the similar species are C. ochroleuca and C. ussuriensis, which share the short stalks and small, simple ascospores. But C. ochroleuca is distinguished from the new species by its larger apothecia (0.2–0.6 mm), longer asci (43–47 μm), black stalks with thick white pruina, and stalks are KOH+ red and green simultaneously [14,50,51,52]. C. ussuriensis is distinguished from the new species by its larger apothecia (0.34– 0.52 mm), smaller asci (30.8–33.7 × 3.0–3.9 μm), shorter ellipsoidal spores with rounded apices (4.0–4.8 × 2.3–3 µm) [53].
Lecanora pseudargentata Lumbsch, J. Hattori Bot. Lab. 77: 127 (1994)., Figure 4.
THALLUS: corticolous, crustose, thin, yellowish white to whitish grey surface, continuous to verrucose.
APOTHECIA: lecanorine, sessile, and constricted at base, 0.3–1.0 mm diameter, margins concolorous with thallus, entire, verrucose; DISCS: red-brown to dark brown, epruinose to slightly pruinose; EXCIPLE: 50–98 µm thick; EPITHECIUM: red-brown, 15–29 µm tall, with granules; HYMENIUM: hyaline, 80–132 µm tall, I+ blue; HYPOTHECIUM: hyaline to pale grey, 27–62 µm tall. ASCI: cylindrical with amyloid apex, 35–55 × 10–15 µm, eight-spored. ASCOSPORES: ellipsoid, 7.5–10.5 × 4.5–7 µm, I–.
CHEMISTRY: Thallus K+ yellow, C−, P−. It contains atranorin, gangaleoidin, chloroatranorin, and norgangaleoidin.
In addition to Lecanora pseudoargentata, atranorin, gangaleoidin, chloroatranorin, and norgongaleoidin have also been isolated from various other lichen species and have been shown to have different biological activities [3,54]. For example, atranorin has been reported to have antimicrobial, antioxidant, anti-inflammatory, and anticancer properties [3]. It is also used in traditional medicine to treat skin conditions, wounds, and inflammation and is being investigated for its potential as a natural food and cosmetic preservative [55]. Gangaleoidin is a scavenger of hydroxyl and superoxide anion radicals. It has therefore been investigated for its potential as an anti-inflammatory and antioxidant agent [56]. It has also been shown to have antifungal and antibacterial activities. Chloroatranorin has been shown to have antibacterial activity against various bacteria and fungi [57]. It has also been investigated for its potential as a natural food preservative and anti-inflammatory agent. However, there are no clear reports on the specific bioactive function of norgangaleoidin. And it is worth noting that although some compounds have shown promising results in some studies, more research is needed to fully understand the potential uses and limitations of the lichen metabolites found and being discovered.
SUBSTRATE: Bark.
ECOLOGY AND DISTRIBUTION: Pantropical area [21] including Thailand, Argentina, Australia, Brazil, Colombia, Costa Rica, Paraguay, Puerto Rico, America, Uruguay, and Venezuela [21,58]. New record to China.
SPECIMEN EXAMINED: China. Jiangxi: Shangrao City, Yushan county, Entrance to Sanqing Mountain, alt. 395 m, on bark, 17/III/2019, M. Li JX19079 (LCUF).
REMARKS: Lecanora pseudargentata is characterised by its crustose, yellowish white to whitish grey thallus, lecanorine ascomata, red-brown to dark brown discs, eight-spored asci, nonseptate and hyaline spores, and the presence of atranorin and gangaleoidin. The species is similar to Lecanora argentata (Ach.) Röhl. and Lecanora subrugosa Nyl., but L. argentata differs in having nongranular epithecium, larger ascospores (11–15 × 6.5–9 μm), and the absence of atranorin [21,58,59], and L. subrugosa has epruinose discs, nongranular epithecium, and atranorin is a constant component in Lecanora s. str. and also present in L. argentata, which does not contain gangaleoidin, chloroatranorin, and norgangaleoidin [60].
4. Discussion
In Figure 1, the phylogenetic analysis dataset includes 29 ITS sequences representing four relative genera and an outgroup of Talaromyces, which is based on 28 species from 29 specimens. The phylogenetic results showed that two individuals of Chaenothecopsis kilimanjaroensis were strongly supported as monophyletic (ML = 100% PP = 1); the other species were not grouped with any other species in the tree. The genus of Chaenothecopsis was resolved as polyphyletic, and the genera of Phaeocalicium and Sphinctrina were resolved as monophyletic clades, which is consistent with the previous findings of Thiyagaraja et al. [13]. In addition, the results showed that the genus Mycocalicium is not a monophyletic lineage, which is consistent with the results of Tuovila et al. and Rikkinen et al. [17,40]. The ITS phylogeny revealed the new species located in a well-supported clade of Chaenothecopsis s. str. (ML = 78% PP = 0.98) (C. vainioana, C. epithallina, C. subparoica, C. consociate, C. pusilla, C. viridireagens, C. pusiola, C. fennica, C. sitchensis, C. golubkovae, C. xishuiensis, C. viridialba, and C. montana). Although blast searches of ITS sequences indicated that C. xishuiensis has close affinities with C. montana (83% identity) and C. viridialba (87% identity), the phylogenetic analyses strongly support that C. xishuiensis is not grouped with these two species in the phylogenetic tree. Although C. xishuiensis occupies a close position to C. viridialba and C. montana, which three together form a supported phylogenetic clade (BS = 94%, PP = 0.99), the phylogenetic analyses strongly support that C. xishuiensis is separated from two species in the tree, appearing as sister to C. viridialba with a high support value (BS = 99%, PP = 1) and in a completely different clade from C. montana (Figure 1). Moreover, morphologically C. viridialba and C. montana differ from C. xishuiensis in that C. viridialba has whitish pruina or granules on longer-stalked (2–3 mm) apothecia and ellipsoid spores (6–10 × 2. 5–3.5 µm) [9,48], whereas C. montana has narrower asci (3.5–5.5 mm), ellipsoid to ovoid ascospores, and all parts of the apothecium K− [49]. We, therefore, describe it as a new species of C. xishuiensis, which is characterised by leprose thallus, single, conical to hemispherical apothecia, nonbranching stipe, cylindrical, eight-spored asci, nonseptate and brown spores, and it is a lichenised fungus containing atranorin and zeorin in the thallus.
Within the Lecanora phylogenetic tree (Figure 2), the molecular phylogeny based on the 16 ITS of Lecanora shows a well-supported monophyletic lineage with Protoparmelia ochrococca as an outgroup. The tree shows that the specimens collected in China and South Africa are monophyletic with a high support value (BS = 100%, PP = 1.0). This result confirms the hypothesis that the specimen belongs to L. pseudargentata on the basis of obvious phenotypic and metabolic characteristics, where L. pseudargentata is characterised by its crustose, yellowish white to whitish grey thallus, lecanorine ascomata, red-brown to dark brown discs, eight-spored asci, nonseptate and hyaline spores, and the presence of atranorin and gangaleoidin [21,58]. Phylogenetically L. pseudargentata is closely related to the species L. cenisia and L. argentata, together with L. cenisi and L. argentata forming a lineage with good support (BS = 97%, PP = 0.99), in which L. pseudargentata is revealed as sister to the clade consisting of L. cenisia and L. argentata. In addition, L. cenisia differs in epruinose thallus, 0.5–2 (2.5) mm apothecia, and the absence of gangaleoidin. L. argentata differs in having nongranular epithecium, larger ascospores (11–15 × 6.5–9 μm), and the absence of atranorin [21,58,59].
In this paper, our results suggest that Chaenothecopsis xishuiensis is a new species to science, and Lecanora pseudargentata is a new record for China based on phenotypic, molecular, and metabolite data. We have provided a detailed description and discussion of the Chaenothecopsis xishuiensis and Lecanora pseudargentata, together with photos of external morphology and internal anatomical features, and provided phylogenetic trees constructed from ITS sequences of some species. We also provided data for lichen classification research. However, we have only roughly identified lichen secondary metabolites and have not further investigated their medicinal value. Next, we plan to conduct a meta-analysis of reported articles on the medicinal value of lichens, screen compounds with high reliability of application value, and conduct preclinical studies to provide data to support their clinical applications.
Author Contributions
Conceptualization, Z.J. and M.L.; methodology, Z.J., M.L. and Y.J.; software, M.L. and Y.J.; validation, Z.J., M.L. and Y.J.; formal analysis, M.L. and Y.J.; investigation, M.L.; resources, Z.J.; data curation, Z.J.; writing—original draft preparation, M.L.; writing—review and editing, Z.J. and Y.J.; visualization, M.L. and Y.J.; supervision, Z.J.; project administration, Z.J.; funding acquisition, Z.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 31750001.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All newly generated sequences were deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 27 April 2023)). Publicly available datasets were used in this study can be obtained from NCBI website (https://www.ncbi.nlm.nih.gov/ 24 April 2023).
Acknowledgments
The authors express their gratitude to Xinli Wei (State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing) and Lulu Zhang (College of Life Sciences, Shandong Normal University, China) for giving us previous suggestions on the present paper, and thank the reviewers for improving the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wei, J.C. A review on the present situation of lichenology in China. Mycosystema 2018, 37, 812–818. [Google Scholar]
- Chen, J.; Blume, H.; Beyer, L. Weathering of rocks induced by lichen colonization—A review. Catena 2000, 39, 121–146. [Google Scholar] [CrossRef]
- Ranković, B. Lichen Secondary Metabolites; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–195. [Google Scholar]
- Jia, Z.F.; Wei, J.C. Flora Lichenum Sinicorum, Volume 13: Ostropales (I), Graphidaceae (1); Science Press: Beijing, China, 2016. [Google Scholar]
- Wei, J.C. Biocarpet Engineering Using Microbiotic Crust for Controlling Sand. Arid Zone Res. 2005, 22, 287–288. [Google Scholar]
- Chang, R.; Wang, Y.; Liu, Y.; Wang, Y.; Li, S.; Zhao, G.; Zhang, S.; Dai, M.; Zheng, X.; Bose, T.; et al. Nine new species of black lichenicolous fungi from the genus Cladophialophora (Chaetothyriales) from two different climatic zones of China. Front. Microbiol. 2023, 14, 1191818. [Google Scholar] [CrossRef]
- Diederich, P.; Lawrey, J.D.; Ertz, D. The 2018 classification and checklist of lichenicolous fungi, with 2000 non lichenized, obligately lichenicolous taxa. Bryologist 2018, 121, 340–425. [Google Scholar] [CrossRef]
- Lawrey, J.D.; Diederich, P. Lichenicolous Fungi: Interactions, Evolution, and Biodiversity. Bryologist 2003, 106, 80–120. [Google Scholar] [CrossRef]
- Groner, U. The genus Chaenothecopsis (Mycocaliciaceae) in Switzerland, and a key to the European species. Lichenologist 2006, 38, 395–406. [Google Scholar] [CrossRef]
- Jaklitsch, W.; Baral, H.O.; Lücking, R.; Lumbsch, H.T. Ascomycota. In Syllabus of Plant Families - Adolf Engler’s Syllabus der pflanzenfamilien, 13th ed.; Gebr. Borntraeger Verlagsbuchhandlung: Stuttgart, Germany, 2016; pp. 1–150. [Google Scholar]
- Lücking, R.; Hodkinson, B.P.; Leavitt, S.D. Corrections and amendments to the 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota. Bryologist 2017, 120, 58–69. [Google Scholar] [CrossRef]
- Falswal, A.; Bhandari, B.S. A New Lichenicolous Fungus from Garhwal Himalayan Region of Uttarakhand. Acta Bot. Hung. 2021, 63, 297–302. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Ertz, D.; Lücking, R.; Wanasinghe, D.N.; Aptroot, A.; Cáceres, M.E.d.S.; Hyde, K.D.; Tapingkae, W.; Cheewangkoon, R. Taxonomic and Phylogenetic Reassessment of Pyrgidium (Mycocaliciales) and Investigation of Ascospore Morphology. J. Fungi 2022, 8, 966. [Google Scholar] [CrossRef]
- Selva, S.B. The calicioid lichens and fungi of the Acadian Forest ecoregion of northeastern North America, II. The rest of the story. Bryologist 2014, 117, 336–367. [Google Scholar] [CrossRef]
- Selva, S.B.; Tuovila, H. Two new resinicolous mycocalicioid fungi from the Acadian Forest: One new to science, the other new to North America. Bryologist 2016, 119, 417–422. [Google Scholar] [CrossRef]
- Titov, A. Notes on Calicioid lichens and fungi from the Gongga Mountains (Sichuan, China). Lichenologist 2000, 32, 553–569. [Google Scholar] [CrossRef]
- Tuovila, H.; Davey, M.L.; Yan, L.; Huhtinen, S.; Rikkinen, J. New resinicolous Chaenothecopsis species from China. Mycologia 2014, 106, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar]
- Yakovchenko, L.S.; Davydov, E.A.; Ohmura, Y.; Printzen, C. The phylogenetic position of species of Lecanora s. l. containingcalycin and usnic acid, with the description of Lecanora solaris Yakovchenko & Davydov sp. nov. Lichenologist 2019, 51, 147–156. [Google Scholar]
- Printzen, C. Corticolous and lignicolous species of Lecanora (Lecanoraceae, Lecanorales) with usnicor isousnic acid in the Sonoran Desert Region. Bryologist 2001, 104, 382–409. [Google Scholar] [CrossRef]
- Papong, K.; Lumbsch, H.T. A taxonomic survey of Lecanora sensu stricto in Thailand (Lecanoraceae; Ascomycota). Lichenologist 2011, 43, 299–320. [Google Scholar] [CrossRef]
- Sliwa, L.; Miadlikowska, J.; Redelings, B.D.; Molnar, K.; Lutzoni, F. Are widespread morphospecies from the ´ Lecanora dispersa group (lichen-forming Ascomycota) monophyletic? Bryologist 2012, 115, 265–277. [Google Scholar] [CrossRef]
- Ivanovich, C.; Dolnik, C.; Otte, V.; Palice, Z.; Sohrabi, M.; Printzen, C. A preliminary phylogeny of the Lecanora saligna-group, with notes on species delimitation. Lichenologist 2021, 53, 63–79. [Google Scholar] [CrossRef]
- Lü, L.; Zhag, L.L.; Yang, M.Z.; Zhao, Z.T. Studies on lichen of the genus Lecanora (Lecanoraceae) in China. In Proceedings of the 2019 Annual Meeting of Mycological Society of China, Xian, China, 5 August 2019. [Google Scholar]
- Lü, L.; Zhao, Z.T. Lecanora shangrilaensis sp. nov., on pinecones from China. Mycotaxon 2017, 132, 441–444. [Google Scholar] [CrossRef]
- Lü, L.; Zhao, Z.T. Lecanora subloekoesii sp. nov. and four other species of the L. subfusca group new to China. Mycotaxon 2017, 132, 539–546. [Google Scholar] [CrossRef]
- Papong, K.; Boonpragob, K.; Parnmen, S.; Lumbsch, H.T. Molecular phylogenetic studies on tropical species of Lecanora sensu stricto (Lecanoraceae, Ascomycota). Nova Hedwig 2013, 96, 1–13. [Google Scholar] [CrossRef]
- Zhao, X.; Leavitt, S.D.; Zhao, Z.T.; Zhang, L.L.; Arup, U.; Grube, M.; Pérez-Ortega, S.; Printzen, C.; Sliwa, L.; Kraichak, E.; et al. Towards a revised generic classification of lecanoroid lichens (Lecanoraceae, Ascomycota) based on molecular, morphological and chemical evidence. Fungal Divers. 2016, 78, 293–304. [Google Scholar] [CrossRef]
- Arup, U.; Grube, M. Molecular systematics of Lecanora subgenus Placodium. Lichenologist 1998, 30, 415–425. [Google Scholar] [CrossRef]
- Arup, U.; Grube, M. Is Rhizoplaca (Lecanorales, lichenized Ascomycota) a monophyletic genus? Can. J. Bot. 2000, 78, 318–327. [Google Scholar] [CrossRef]
- Grube, M.; Baloch, E.; Arup, U. A phylogenetic study of the Lecanora rupicola group (Lecanoraceae, Ascomycota). Mycol. Res. 2004, 108, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, J.G.; dos Santos, L.A.; Aptroot, A.; Lücking, R.; Caceres, M.E.S. A new species of Lecanora (Ascomycota: Lecanoraceae) from mangrove in northeast Brazil identified using DNA barcoding and phenotypical characters. Bryologist 2019, 122, 553–558. [Google Scholar] [CrossRef]
- Zhang, Y.; Clancy, J.; Jensen, J.; McMullin, R.T.; Wang, L.; Leavitt, S.D. Providing scale to a known taxonomic unknown—At least a 70-fold increase in species diversity in a cosmopolitan nominal taxon of lichen-forming fungi. J. Fungi 2022, 8, 490. [Google Scholar] [CrossRef]
- Santos, L.A.d.; Aptroot, A.; Lücking, R.; Cáceres, M.E.d.S. Lecanora s.lat. (Ascomycota, Lecanoraceae) in Brazil: DNA Barcoding Coupled with Phenotype Characters Reveals Numerous Novel Species. J. Fungi 2023, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Culberson, C.F.; Kristinsson, H.D. A standardized method for the identification of lichen products. J. Chromatogr. 1970, 46, 85–93. [Google Scholar] [CrossRef]
- Culberson, C.F. Improved conditions and new data for identification of lichen products by standardized thin-layer chromatographic method. J. Chromatogr. A 1972, 72, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens; British Lichen Society: London, UK, 2001. [Google Scholar]
- Larena, I.; Salazar, O.; Gonzalez, V.; Julian, M.C.; Rubio, V. Design of a primer for ribosomal DNA internal transcribed spacer with enhanced specificity for ascomycetes. J. Biotechnol. 1999, 75, 187–194. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Rikkinen, J.; Tuovila, H.; Beimforde, C.; Seyfullah, L.J.; Perrichot, V.; Schmidt, A.R. Chaenothecopsis neocaledonica sp. nov.: The first resinicolous mycocalicioid fungus from an araucarian conifer. Phytotaxa 2014, 173, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Temu, S.G.; Tibell, S.; Tibuhwa, D.D.; Tibell, L. Crustose Calicioid Lichens and Fungi in Mountain Cloud Forests of Tanzania. Microorganisms 2019, 7, 491. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.; Higgins, D.; Gibson, T. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Beimforde, C.; Tuovila, H.; Schmidt, A.R.; Lee, W.; Gube, M.; Rikkinen, J. Chaenothecopsis schefflerae (Ascomycota: Mycocaliciales): A widespread fungus on semi-hardened exudates of endemic New Zealand Araliaceae. N. Z. J. Bot. 2017, 55, 387–406. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Peterson, E.B.; Rikkinen, J. Range extensions of selected pin-lichens and allied fungi in the Pacific Northwest. Bryologist 1999, 370–376. [Google Scholar] [CrossRef]
- Tuovila, H.; Larsson, P.; Rikkinen, J. Three resinicolous North American species of Mycocaliciales in Europe with a re-evaluation of Chaenothecopsis oregana Rikkinen. Karstenia 2011, 51, 37–49. [Google Scholar] [CrossRef]
- Selva, S.B.; Tibell, L. Lichenized and non-lichenized calicioid fungi from North America. Bryologist 1999, 102, 377–397. [Google Scholar] [CrossRef]
- Stordeur, R.; Braun, U.; Tkach, N. Titov: Mycocaliciale Pilze der Holarktis—Übersetzung der Bestimmungsschlüssel und Beschreibungen neuer Arten. Herzogia 2010, 23, 19–67. [Google Scholar] [CrossRef]
- Paquette, H.A.; Gates, K.; McMullin, R.T. Chaenothecopsis ochroleuca, Haematomma ochroleucum, and Multiclavula vernalis reported for the first time from Maine. Northeast. Nat. 2020, 27, N34–N39. [Google Scholar] [CrossRef]
- Titov, A.; Tibell, L. Chaenothecopsis in the Russian Far East. Nord. J. Bot. 2008, 13, 313–329. [Google Scholar] [CrossRef]
- Mahandru, M.M.; Gilbert, O.L. Norgangaleoidin, a Dichlorodepsidone from Lecanora chlarotera. Bryologist 1979, 82, 292–295. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Galanty, A.; Bylka1, W. Atranorin—An Interesting Lichen Secondary Metabolite. Mini-Rev. Med. Chem. 2017, 17, 1633–1645. [Google Scholar] [CrossRef]
- Bay, M.V.; Nam, P.C.; Quang, D.T.; Mechler, A.; Hien, N.K.; Hoa, N.T.; Vo, Q.V. Theoretical study on the antioxidant activity of natural depsidones. ACS Omega 2020, 5, 7895–7902. [Google Scholar] [CrossRef]
- Türk, H.; Yilmaz, M.; Tay, T.; Türk, A.O.; Kivanç, M. Antimicrobial Activity of Extracts of Chemical Races of the Lichen Pseudevernia furfuracea and their Physodic Acid, Chloroatranorin, Atranorin, and Olivetoric Acid Constituents. Z. Naturforsch. C J. Biosci. 2006, 61, 499–507. [Google Scholar] [CrossRef] [PubMed]
- LaGreca, S.; Lumbsch, H.T. Three species of Lecanora new to North America, with notes on other Poorly known lecanoroid lichens. Bryologist 2001, 104, 204–211. [Google Scholar] [CrossRef]
- Malíček, J. A revision of the epiphytic species of the Lecanora subfusca group (Lecanoraceae, Ascomycota) in the Czech Republic. Lichenologist 2014, 46, 489–513. [Google Scholar] [CrossRef]
- Lü, L.; Wang, H.Y.; Zhao, Z.T. Five lichens of the genus Lecanora new to China. Mycotaxon 2009, 107, 391–396. [Google Scholar] [CrossRef]
Figure 1.
Maximum likelihood tree of Sphinctrinaceae s. str. based ITS sequences with Talaromyces acaricola as outgroup. ML-BS > 70% (left) and BI-PP > 0.9 (right) are considered to be strongly supported. Terminal in bold indicates newly generated sequence.
Figure 1.
Maximum likelihood tree of Sphinctrinaceae s. str. based ITS sequences with Talaromyces acaricola as outgroup. ML-BS > 70% (left) and BI-PP > 0.9 (right) are considered to be strongly supported. Terminal in bold indicates newly generated sequence.

Figure 2.
Maximum likelihood tree of Lecanora based on ITS sequences with Protoparmelia ochrococca as outgroup. ML-BS > 70% (left) and BI-PP > 0.9 (right) are considered to be strongly supported. Terminal in bold indicates newly generated sequence.
Figure 2.
Maximum likelihood tree of Lecanora based on ITS sequences with Protoparmelia ochrococca as outgroup. ML-BS > 70% (left) and BI-PP > 0.9 (right) are considered to be strongly supported. Terminal in bold indicates newly generated sequence.

Figure 3.
Chaenothecopsis xishuiensis (holotype, M.L. Zhu GZ19513 LCUF). (A) Thallus and ascomata; (B) thallus and ascomata; (C) mycelia and algal cells of thallus; (D) cross section of ascomata; (E) cross section of ascomata and stalk; (F) an ascus containing ascospores. Scales: (A) = 1.5 mm; (B) = 0.5 mm; (C) = 50 µm; (D) = 50 µm; (E) = 100 µm; (F) = 20 µm.
Figure 3.
Chaenothecopsis xishuiensis (holotype, M.L. Zhu GZ19513 LCUF). (A) Thallus and ascomata; (B) thallus and ascomata; (C) mycelia and algal cells of thallus; (D) cross section of ascomata; (E) cross section of ascomata and stalk; (F) an ascus containing ascospores. Scales: (A) = 1.5 mm; (B) = 0.5 mm; (C) = 50 µm; (D) = 50 µm; (E) = 100 µm; (F) = 20 µm.

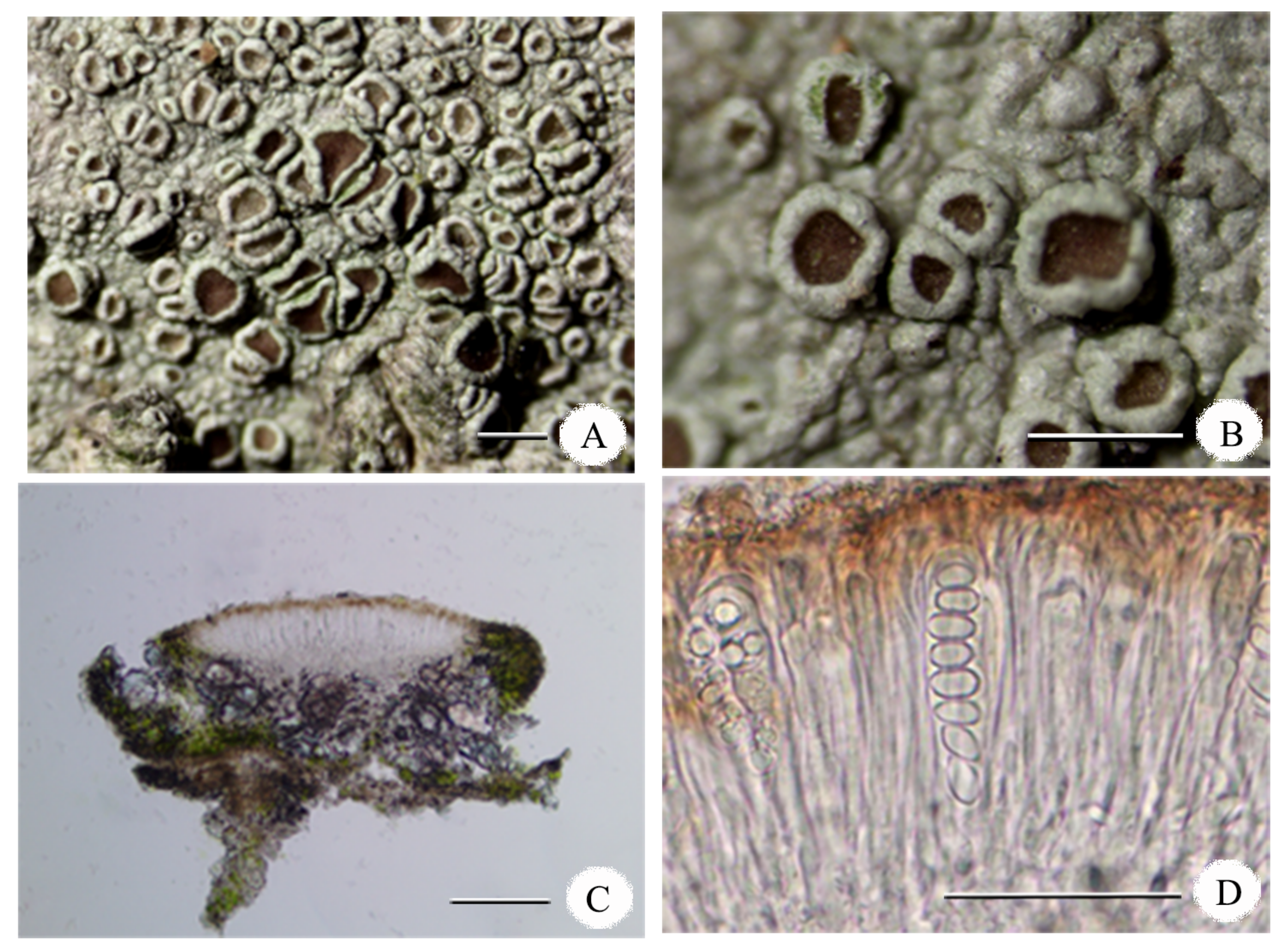
Figure 4.
Lecanora pseudargentata (M. Li JX19079 LCUF). (A) Thallus with apothecia; (B) apothecia; (C) cross section of apothecium; (D) an ascus containing ascospores. Scales: (A) = 1 mm; (B) = 1 mm; (C) = 50 µm; (D) = 100 µm.
Figure 4.
Lecanora pseudargentata (M. Li JX19079 LCUF). (A) Thallus with apothecia; (B) apothecia; (C) cross section of apothecium; (D) an ascus containing ascospores. Scales: (A) = 1 mm; (B) = 1 mm; (C) = 50 µm; (D) = 100 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Taxa and their GenBank accession numbers used to construct phylogenetic trees. Information on new species and records are indicated in bold.
Table 1.
Taxa and their GenBank accession numbers used to construct phylogenetic trees. Information on new species and records are indicated in bold.
| Species | GenBank Accession Numbers |
|---|---|
| Chaenothecopsis consociata (Nádv.) A.F.W. Schmidt | AY795851 |
| Chaenothecopsis debilis (Sm.) Tibell | AY795853 |
| Chaenothecopsis epithallina Tibell | AY795856 |
| Chaenothecopsis fennica (Laurila) Tibell | AY795858 |
| Chaenothecopsis golubkovae Tibell & Titov | AY795859 |
| Chaenothecopsis haematopus Tibell | AY795861 |
| Chaenothecopsis kilimanjaroensis Temu & Tibell | MN575660 |
| Chaenothecopsis kilimanjaroensis Temu & Tibell | MN575662 |
| Chaenothecopsis montana Rikkinen | JX119105 |
| Chaenothecopsis pusilla (Ach.) A.F.W. Schmidt | AY795866 |
| Chaenothecopsis pusiola (Ach.) Vain. | JX119106 |
| Chaenothecopsis sitchensis Rikkinen | JX119102 |
| Chaenothecopsis subparoica (Nyl.) Tibell | AY795869 |
| Chaenothecopsis vainioana (Nádv.) Tibell | JX119107 |
| Chaenothecopsis viridialba (Kremp.) A.F.W. Schmidt | JX000103 |
| Chaenothecopsis viridireagens (Nádv.) A.F.W. Schmidt | JX119108 |
| Mycocalicium albonigrum (Nyl.) Fink | AF223969 |
| Mycocalicium americanum (R. Sant.) Tibell | AY795879 |
| Mycocalicium hyaloparvicellulum Daranag. & K.D. Hyde | NR_168156 |
| Mycocalicium subtile (Pers.) Szatala | AF225440 |
| Phaeocalicium curtisii (Tuck.) Tibell | KT695401 |
| Phaeocalicium populneum (Brond. ex Duby) A.F.W. Schmidt | AY795874 |
| Phaeocalicium praecedens (Nyl.) A.F.W. Schmidt | KC590481 |
| Sphinctrina anglica Nyl. | OQ718101 |
| Sphinctrina leucopoda Nyl. | AY795875 |
| Sphinctrina tubaeformis A. Massal. | OQ717614 |
| Sphinctrina turbinata (Pers.) De Not. | OQ718102 |
| Chaenothecopsis xishuiensis Z.F. Jia | MW017534 |
| Talaromyces acaricola Visagie, N. Yilmaz & K. Jacobs | MW269298 |
| Lecanora argentata (Ach.) Malme | MN387067 |
| Lecanora cenisia Ach. | EU558541 |
| Lecanora chlarotera Nyl. | MN654577 |
| Lecanora conizaeoides Nyl. ex Cromb. | AF189717 |
| Lecanora farinacea Fée | JN943726 |
| Lecanora formosa (Bagl. & Carestia) Knoph & Leuckert | KT453771 |
| Lecanora hybocarpa (Tuck.) Brodo | EF105412 |
| Lecanora intricata (Ach.) Ach. | AF070022 |
| Lecanora intumescens (Rebent.) Rabenh. | AY541254 |
| Lecanora somervellii Paulson | MH512979 |
| Lecanora praepostera Nyl. | MN586973 |
| Lecanora pseudargentata Lumbsch | OQ832601 |
| Lecanora pseudargentata Lumbsch | MH714514 |
| Lecanora rugosella Zahlbr. | MK573926 |
| Lecanora solaris L.S. Yakovchenko & E.A. Davydov | MH512982 |
| Lecanora polytropa (Ehrh.) Rabenh. | MZ243614 |
| Protoparmelia ochrococca (Nyl.) P.M. Jørg., Rambold & Hertel | KP822293 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, M.; Ju, Y.; Jia, Z. Chaenothecopsis xishuiensis sp. nov. to Science and Lecanora pseudargentata Newly Reported from China. Diversity 2023, 15, 893. https://doi.org/10.3390/d15080893
AMA Style
Li M, Ju Y, Jia Z. Chaenothecopsis xishuiensis sp. nov. to Science and Lecanora pseudargentata Newly Reported from China. Diversity. 2023; 15(8):893. https://doi.org/10.3390/d15080893
Chicago/Turabian StyleLi, Min, Yanhu Ju, and Zefeng Jia. 2023. "Chaenothecopsis xishuiensis sp. nov. to Science and Lecanora pseudargentata Newly Reported from China" Diversity 15, no. 8: 893. https://doi.org/10.3390/d15080893
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.