
Dynamics of Zooplankton along the Romanian Black Sea Coastline: Temporal Variation, Community Structure, and Environmental Drivers


1
Ecology and Marine Biology Department, National Institute for Marine Research and Development “Grigore Antipa”, 300 Mamaia Blvd., 900581 Constanta, Romania
2
Chemical Oceanography and Marine Pollution Department, National Institute for Marine Research and Development “Grigore Antipa”, 300 Mamaia Blvd., 900581 Constanta, Romania
3
National Institute for Marine Research and Development “Grigore Antipa”, 300 Mamaia Blvd., 900581 Constanta, Romania
*
Authors to whom correspondence should be addressed.
Diversity 2023, 15(9), 1024; https://doi.org/10.3390/d15091024
Submission received: 30 August 2023
/
Revised: 15 September 2023
/
Accepted: 18 September 2023
/
Published: 20 September 2023
(This article belongs to the Special Issue Advances in the Diversity and Ecology of Zooplankton)
Abstract
:Between 2013 and 2020, a total of 27 distinct zooplankton taxa, encompassing both holoplanktonic and meroplanktonic organisms spanning different classes and orders, were successfully identified along the Romanian Black Sea coastline. The seasonal dispersion patterns of these zooplankton taxa revealed a notable augmentation in species identification during the warmer periods, contrasting with reduced zooplankton diversity during the colder seasons. This phenomenon was underscored by the outcomes of SIMPER analysis, which highlighted that copepods, meroplankton, and cladocerans predominantly contributed to the community structure during warmer seasons. In contrast, during colder seasons, cladocerans were scarcely represented, and the density levels of copepods and meroplankton were diminished. From a quantitative perspective, the dinoflagellate Noctiluca scintillans, exhibited its highest density and biomass during the warm season, with a relatively lower representation during colder periods. The variation in the abundance of zooplankton was closely linked to changes in salinity and temperature, culminating in noticeable fluctuations, which were particularly evident during the cold season. During this period, the values of zooplankton abundance were notably lower compared to those observed in the warmer season. The comprehension of zooplankton dynamics stands as a critical facet for assessing biodiversity, monitoring ecosystem well-being, and guiding conservation endeavours.
Keywords:
zooplankton density; analysis; taxa; season; salinity; temperature; biodiversity assessment; Black Sea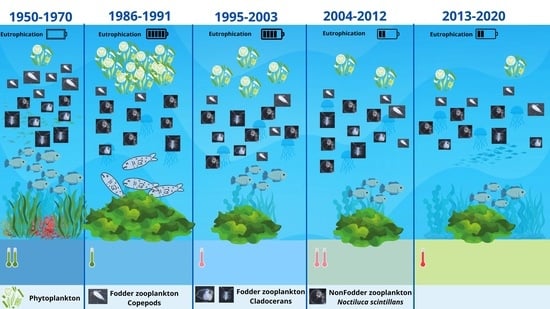
1. Introduction
Zooplankton help to detect the extent of climate change by fixing carbon through the biological pump; but, at the same time, they are very susceptible to climate change [1]. Thus, zooplankton significantly mitigate climate change by aiding in carbon sequestration through the biological pump; yet, they face high vulnerability to climate change impacts, including rising temperatures, ocean acidification, and shifts in their food sources, which could disrupt their crucial role in carbon cycling and exacerbate climate change [2]. Different ecosystem components (trophic groups) respond phenologically to changes in environmental conditions (eutrophication and temperature rise), which can lead to mismatches between successive trophic levels and disrupt the synchrony between primary, secondary, and tertiary consumers. Pelagic fish are also directly and indirectly influenced by climate change. As rising water temperatures cause changes in fish migrations and lead to changes in the presence and development of their zooplankton prey, any variation in zooplankton dynamics has implications for fishing and ecosystem services [1]. Unfortunately, all these have been observed in recent years in the Black Sea [3,4,5,6].
The dynamics of zooplankton populations, their reproductive cycles, growth, and survival rate are key factors influencing the recruitment of fish stocks. The latter role has made zooplankton ecology of interest for research.
Several marine organisms feed on plankton either at some point in their life cycle or throughout their lives. Therefore, the importance of plankton in the transfer of energy to higher trophic levels through different food chains cannot be underestimated, as it is the basis of the marine food web and, therefore, plays a key role in fishing. Primary productivity and plankton growth are closely related to the physicochemical parameters of seawater [7].
In the Black Sea, zooplankton have the role of a “key industry” by converting into organic matter not only a large amount of primary production but also dissolved organic substances. In this way, the matter and energy generated in the ecosystem and particulate or solvated organic substances from the water mass become accessible for organisms that are not planktivorous or do not have mechanisms for capturing organo-bacterial aggregates. Thus, zooplankton become the main food source for juvenile and adult pelagic fishes, species that in the Black Sea, are essential elements not only in the trophic base of cetaceans but also in the qualitative composition of industrial fishing [8].
Zooplankton play a significant role in the production of any aquatic ecosystem. To a confirmed extent, fishing failure and success, especially in the case of pelagic species, are intricately linked to plankton availability, with large fish stocks being found in regions with abundant planktonic biomass [9]. Owing to the key role of zooplankton in the early stages of fish development, changes in species composition and abundance have significant implications for fish recruitment and dynamics. Therefore, understanding the abundance, species composition, and distribution of zooplankton is valuable in supporting research on fish production and changes in the ecosystem [9,10].
Assessment of zooplankton biodiversity is a crucial element in monitoring marine ecosystem processes and community responses to changes that may occur in the marine ecosystem [11]. The biodiversity of zooplankton is one of the most important ecological indicators of the environment, with each species having a specific role in the ecosystem [12]. Biological diversity plays a crucial role in the functioning of systems and in the many services they provide [13].
Zooplankton communities are sensitive to anthropogenic impacts, and environmental fluctuations and their study can be useful in predicting long-term changes [14,15,16,17]. Species diversity and zooplankton community composition may indicate environmental change or disturbance; it has been reported by several researchers that zooplankton can serve as an indicator of studies of trophic dynamics and the ecological status of aquatic ecosystems [16,18]. This holds significant relevance for the Romanian Black Sea, which experiences human impacts from factors like river input (Danube, Dniester, and Dnieper), ports, and industrial operations [19,20].
This study aims to comprehensively investigate the seasonal dynamics and composition of zooplankton taxa along the Romanian Black Sea coast from 2013 to 2020. Through taxonomic identification and analysis, the research seeks to elucidate how the distribution and diversity of zooplankton vary between warm and cold seasons. The study uses statistical analysis to identify key factors affecting the community structure in various seasons, with a particular focus on copepods, meroplankton, cladocerans, and the limited presence of cladocerans during the cold season. All the non-gelatinous and non-opportunistic species groups, such as Copepoda, Cladocera, meroplankton, and other groups that act as the primary prey for larger predators, are referred to as fodder zooplankton [3,5] in this paper and within the Black Sea riparian countries [3,5]. Furthermore, the research aims to quantify the density and biomass and explore between warm and cold seasons the variation in the N. scintillans dinoflagellate, which is an opportunistic species [3]; therefore, it is analysed as the non-fodder zooplanktonic component. By investigating the relationships between fodder zooplankton, salinity, temperature, and nutrients, the study aims to reveal distinct abundance patterns. This investigation also underscores the significance of understanding zooplankton dynamics in the context of biodiversity assessment, ecosystem health monitoring, and the formulation of effective conservation strategies. This holds important significance, especially when considering that N. scintillans serves as the indicator for assessing blooms in the Romanian Black Sea as a part of the assessment of Descriptor 5—Eutrophication—under the Marine Strategy Framework Directive (Directive 2008/56/EC) implementation.
2. Materials and Methods
2.1. Description of the Study Area
Zooplankton samples were collected from stations located along the Romanian Black Sea monitoring network (Figure 1) in 2013–2020 in the warm (May–October) and cold (November–April) seasons. The warm season was characterised by a higher number of samples; while in the cold season, expeditions were only carried out in 2014, 2016, and 2017 (Table S2). The monitoring network, consisting of 45 sampling stations, extends over the entire continental platform of the Romanian Black Sea coast and consists of 13 profiles perpendicular to the coastline: Sulina, Mila 9, Sf. Gheorghe, Portița, Gura Buhaz, Cazino Mamaia, Constanța North, Constanța East, Constanța South, Eforie, Costinești, Mangalia, and Vama Veche. A total of 533 zooplankton samples and 387 water samples for evaluating physicochemical parameters were collected from 45 monitoring stations and analysed in NIMRD laboratories.
2.2. Zooplankton Sampling and Analysis
Zooplankton samples were collected according to the methodology [21] using a Juday net (0.1 m2 mouth opening area and 150 μm mesh size) equipped with a flow meter for estimating the volume of water filtered by vertically towing the net in the water column. In the warm season, zooplankton samples were collected from standard depths (10–0 m, 25–10 m, 50–25 m, and 100–50 m); while in the cold season, sampling was performed from the entire water column.
The quantitative and qualitative processing of the zooplankton samples were performed under an OlympusSZX10 stereomicroscope. When specifying the taxonomic affiliation of zooplankton and for species determination, we used proper manuals and guides [22,23]; classification for identified taxa was conducted according to the World Register of Marine Species (WoRMS).
Not all the organisms could be determined at the species level; the meroplanktonic elements were identified only at the group level. In the subsample(s) examined in the Bogorov chamber, all the organisms were counted until at least 100 individuals for each of the three dominant taxonomic groups were recorded. For the estimation of large animal numbers, the whole sample was examined in a Petri dish. The number of individuals and mean individual weights were used for estimating the density in ind/m3 and the biomass in mg/m3 of wet weight, respectively, based on the tables of constant weight of the Black Sea zooplankters [21,24].
Additional data (qualitative and quantitative) from the scientific literature was used to analyse the long-term trend [8].
Seawater samples were collected at the same time as the biological ones from the same stations using Niskin bottles. A CastAway CTD multiparameter was properly calibrated and used for in situ temperature and salinity measurements. Dissolved nutrients (phosphate, silicate, inorganic forms of nitrogen-nitrite, nitrate, and ammonium) were analysed in the NIMRD laboratory using spectrophotometric methods according to Methods of Seawater Analysis [25].
2.3. Data Analysis
Multivariate and univariate analyses were conducted using PRIMER v. 7.0 [26], and statistical analysis was performed via XLSTAT [27] and Statistica® 14.0.1.25 (TIBCO Software Inc., Palo Alto, CA, USA) [28].
Data analysed with PRIMER v. 7.0 were square-root transformed. Multivariate analyses are characterised based on comparisons between two (or more) samples at the extent to which these samples share certain species at comparable levels of abundance. These facilitate a classification of samples into groups that are then mutually similar or an ordination diagram in which, for example, samples are “mapped” (usually in two or three dimensions) to reflect their relative differences in specific composition [26]. In this research, as a multivariate analysis, the Bray–Curtis similarity was used to evaluate the similarity of the zooplankton community between the analysed years and seasons.
As univariate analyses, diversity indices, shade plots, boxplots, and SIMPER analysis were used. Different diversity indices emphasise the species richness or equitability components of the diversity, the most common being the Shannon–Wiener diversity index (H′), species richness, and Pielou’s evenness index (J′) [26].
where Pi is the proportion of each species in the sample, and å is the abundance of the ith species.
where H′max is the maximum possible value of the Shannon diversity, i.e., that which would be achieved if all the species were equally abundant (namely, log S).
Shade plots are visual displays in the form of the data matrix itself [26], which were used for the identified zooplankton taxa. Boxplots provide a good graphical representation of the concentration of data, showing how far extreme values are from much information. This type of diagram was used to compare the distribution of the zooplankton community according to the taxonomic groups, taking into consideration the frequency of the identified taxa. SIMPER (similarity percentages) analysis was used for assessing the percentage contributions of species to the similarity and dissimilarity between groups, by season.
Statistica® was used for analysing the descriptive statistics of the environmental variables, including correlations. By default, the spreadsheet shows all the correlation coefficients that are significant at p < 0.05 (two-tailed) in a different colour (highlighted in red). Additionally, t-tests were used to compare the environmental characteristics between seasons by evaluating the differences in means between two groups (warm and cold seasons). The null hypothesis assumes that the two means are equal. Using the formulas, values are calculated and compared against the standard values. The assumed null hypothesis is accepted or rejected accordingly. The test returned the p values to check for statistical significance.
XLSTAT [27] was used to perform the Canonical Correspondence Analysis (CCA) to relate the abundance of zooplankton species to environmental variables. The included permutation test concludes that the species data either are or are not linearly related to the variable data at a 5% significance level. CCA uses linear combinations of environmental (explanatory) variables to optimally explain the species (response) variables.
ArcGIS Desktop 10.7 software (ESRI, 2019) [29] was used for creating distribution maps. The data interpolation was carried out using the IDW (inverse distance weighted) method, which determines the cell values using a linearly weighted combination of a set of sampling points. This method assumes that the variable decreases in influence with increasing distance from the sampled location. Therefore, for the maps of distribution, the untransformed data of the studied components were used.
3. Results
3.1. The Environmental Parameters
The Danube’s mouths areas exhibited the lowest temperatures and salinities during different seasons. In the colder season, temperatures decreased; whereas in the warmer season, the salinity dropped to extremely low levels due to the increased flow of the Danube, resulting in freshwater conditions (as shown in Tables S3–S5).
The oxygen saturation reached its minimum point within the marine water-stratified water column (0–30 m), accompanied by elevated nutrient levels. Additionally, the region influenced by the Danube displayed the highest nutrient levels.
Overall, the data indicate distinct statistical differences between the cold and warm seasons across all the parameters except for phosphate and nitrate concentrations, where the levels are similar. The warm season is characterised by higher temperatures, lower salinities, lower dissolved oxygen levels, and higher concentrations of nitrite and ammonium. The cold season, on the other hand, features lower temperatures, higher salinities, higher dissolved oxygen levels, and higher concentrations of silicate (Table S6).
3.2. Zooplankton Composition
For analysing the zooplankton community from the Romanian Black Sea coastline, we distinguished between non-fodder zooplankton, represented by N. scintillans, and the fodder component, made up of copepods, cladocerans, meroplankton, and other groups.
Long-term data on the zooplankton community structure in the Romanian coastal waters showed fluctuations in species diversity (Figure 2). Upon examining the list of zooplanktonic species dating back to 1986 (Table S1), the most considerable count (45) was documented during 1995–2003, while a relatively consistent pattern is noted from 2004 to 2020.
During 2013–2020, 27 zooplanktonic taxa, of holoplanktonic and meroplanktonic natures, belonging to different classes and orders, were identified on the Romanian coastline (Table S2). Throughout the analysed timeframe, a reduction in the count of identified species is noticeable because, over the past few decades, certain species have vanished or grown scarce within the pelagic ecosystem [30].
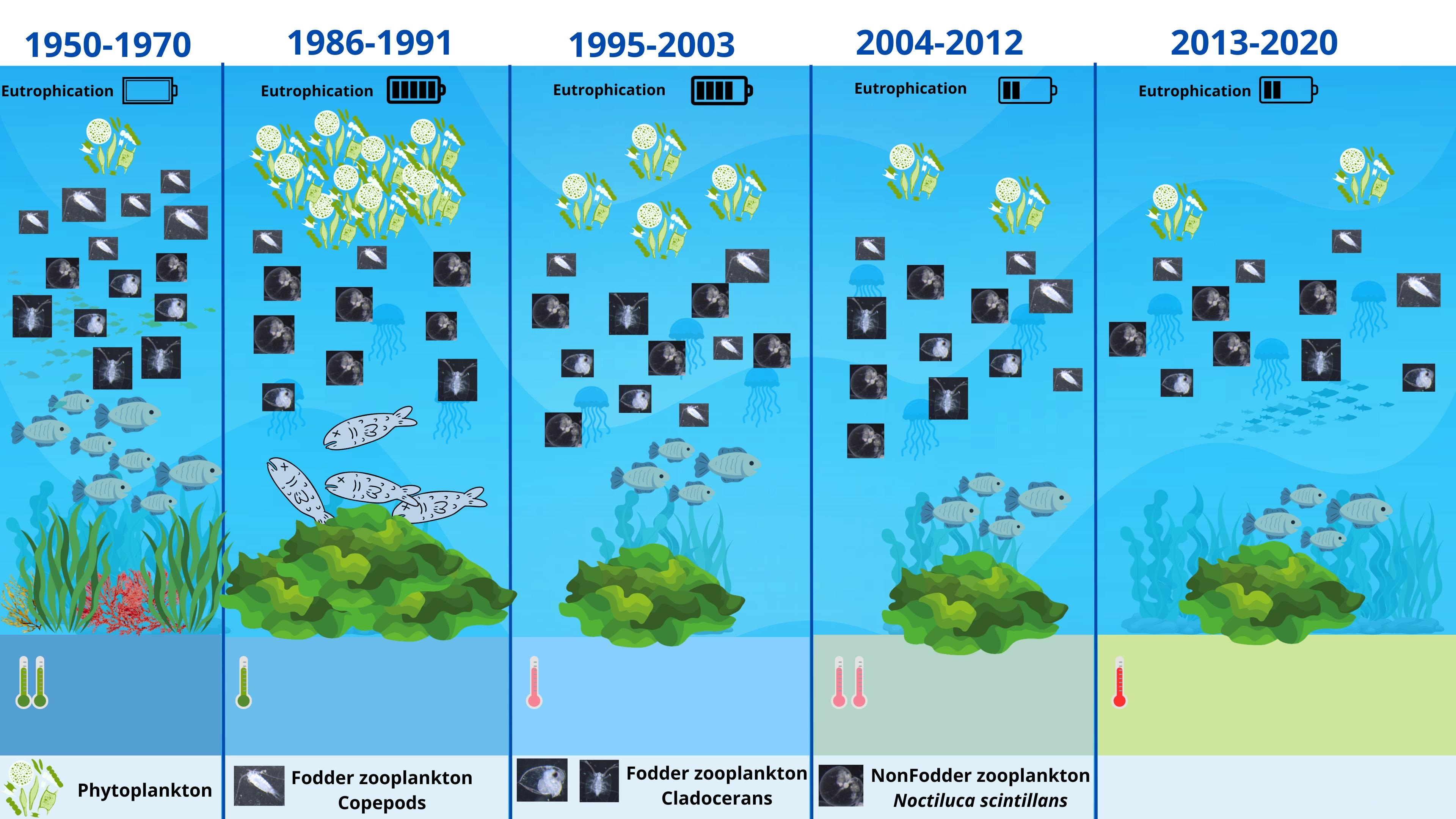
The non-fodder component (N. scintillans) was identified over the entire period, in both seasons (Figure 3). The group of copepods showed variations in the number of species, being best represented in 2019 and 2020, with a maximum of nine species (Figure 3). The core of the copepods (2013–2020) was represented by the species Acartia clausi, Pseudocalanus elongatus, Oithona similis, Paracalanus parvus, Calanus euxinus, and Centropages ponticus, which were identified in all the analysed samples. At the same time, a total of nine species of cladocerans was identified on the Romanian coast (Figure 3); their appearance in such a large number in the plankton was quite rare and closely related to the influence of the freshwater input that the Danube exerts on marine waters. The high number of Cladocera is given by freshwater elements, such as Chydorus sphaericus, Bosmina longirostris, Daphnia longispina, and Diaphanosoma brachyurum, that occasionally appear in the marine waters near the Danube. B. longirostris was identified in 2013, 2015, and 2020; C. sphaericus and D. longispina appeared in samples only in 2015; and the species D. brachyurum was present only in samples from 2019 (Table S2). It is noted that Pleopis polyphemoides, Evadne spinifera, Penilia avirostris, and Pseudevadne tergestina are the main representatives of the cladocerans; these species were constantly identified throughout the analysed period, especially in the warm season. The meroplankton did not show large variations in the number of taxa, recording a maximum of five taxa (Figure 3), namely, the larval stages of Cirripedia, Decapoda, Gastropoda, Bivalvia, and Polychaeta (Table S2). The other group category reached a maximum of three species (Figure 3) and is represented by the chaetognath Parasagitta setosa, the appendicular Oikopleura dioica, and the mysid Mesopodopsis slabberi (Table S2). Chaetognaths play an important role in zooplankton trophodynamics, as their diet includes small copepods (Oithona sp.) [31]; the species P. setosa was identified every year (2013–2020). O. dioica, a planktonic chordate with an exceptional filtering capacity, fast generation time, conserved early development, and a compact genome [32], is mainly distributed in coastal waters [33,34], occurs often in high abundance during phytoplankton blooms [26], and was identified throughout the analysed period. M. slabberi usually occurs in coastal ecosystems; and, on the Romanian coast, it was only identified in five years of the eight analysed (Table S2). The seasonal distribution of zooplankton species reveals that in the warm season, the number of identified species increases, while the diversity is lower in the cold season (Figure 3).
Following the discriminant analysis (DA) for significant differences among the groups by seasonal variation (Table 1), it was observed that of the 27 identified zooplankton taxa in 2013–2020, the most frequent were copepods A. clausi, O. similis, P. parvus, and P. elongatus; the cladoceran P. polyphemoides; meroplanktonic elements, such as Balanus, Bivalvia, Gastropoda, and Polychaeta; the non-fodder component represented by N. scintillans; the appendicular O. dioica; and the chaetognath P. setosa, representing the bulk of the zooplankton community and forming high-similarity clusters (Figure 4). Notably, it should be highlighted that the warm season exhibited the highest relative frequency (81.9%), attributed to the larger number of samples collected during 2013–2020 (Table 1).
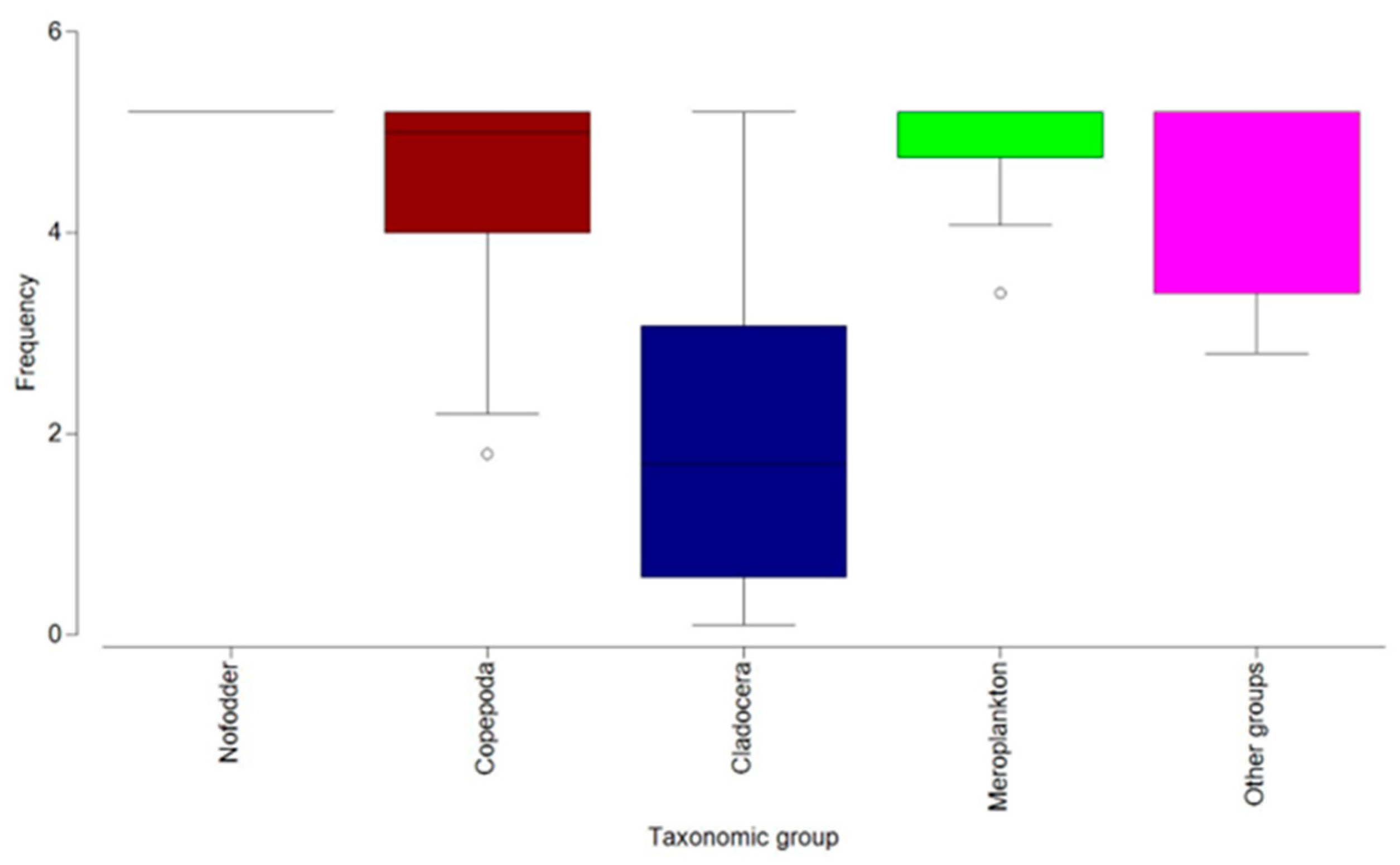
Distinct distributions were observed in the relative frequencies among taxonomic groups, with cladocerans displaying the lowest values. Conversely, the other zooplankton components were more prominently represented (Figure 5).
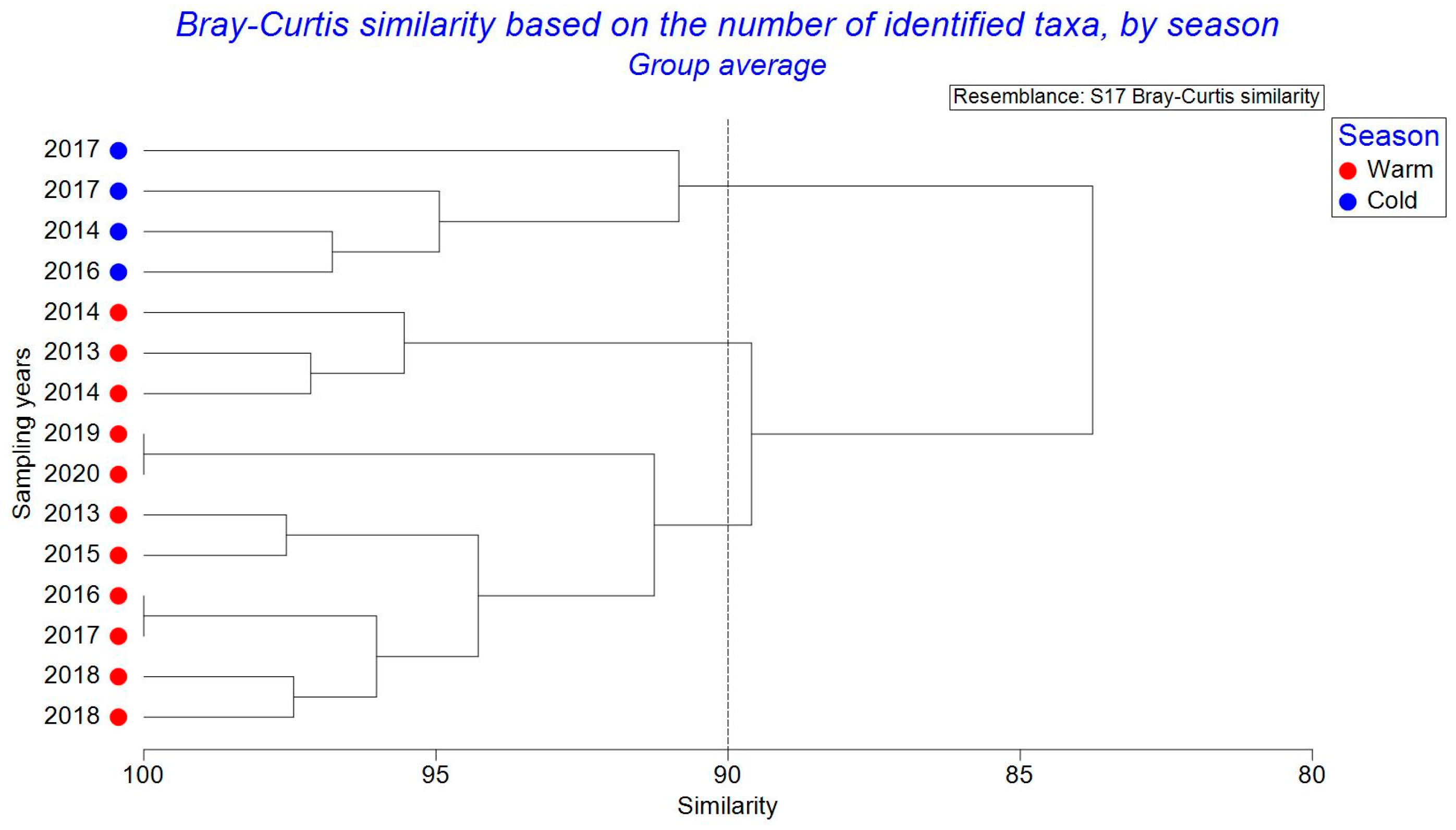
The constancy of zooplankton species is evident, with the copepod category maintaining dominance over the studied interval and displaying yearly fluctuations in the count of identified species. Similarly, the group of cladocerans, well-represented in qualitative terms, exhibits yearly fluctuations in species count, followed by the meroplanktonic segment. The release timing of meroplankton larvae varies among species, with temperature, nutrient availability, and meteorological conditions influencing egg production and larval emergence [35,36]. Other groups were less-well qualitatively represented throughout the study period (Figure 3). The Bray–Curtis analysis generated based on the number of zooplankton taxa indicates a very significant degree of similarity between the years, given the specific uniformity of the zooplankton (Figure 6). A very high degree of similarity was also recorded between the years 2019 and 2020 because the zooplankton component reached the highest number of species and had a similar structure in terms of the dominance of the identified species (Figure 6). The same situation was recorded for 2016 and 2017 owing to the same qualitative structure identified for the groups of copepods.
SIMPER analysis revealed that the main zooplanktonic groups that contributed to the community structure in the warm season are represented by copepods, meroplankton, and cladocerans (Table 2). Copepods were the most abundant in both seasons, while the taxa primarily responsible for the differences between the seasons were cladocerans. In the cold season, they were poorly represented and replaced by the presence of other groups (Table 2).
Throughout the study, a clear fluctuation in species diversity was observed. The Shannon–Wiener diversity index (H′) varied from 1 in 2015 to 2.3 in 2018, in the warm season (Table 3). Pielou’s evenness index values were the highest for the warm season of 2018 (0.8) and the lowest for the warm season of 2015 (0.4) (Table 3).
3.3. Zooplankton Structure–Seasonal Variations
The mean total zooplankton density and biomass values in the Romanian Black Sea waters were the highest in the warm season and significantly declined in the cold one (Figure 7). The non-fodder component, represented by N. scintillans, was best represented in terms of density and biomass, recording the highest mean values (6929 ind/m3 and 610 mg/m3, respectively) in the warm season, followed by copepods, at 4991 ind/m3 in the same season (Figure 7). The meroplanktonic component and cladocerans were also better represented in the warm season; while in the cold season, the mean average density and biomass values for these two taxonomic groups were lower. In the cold season, copepods dominated only in terms of density, which indicates that smaller organisms dominated. On the other hand, the N. scintillans biomass dominated at lower densities (Figure 7).
3.4. Zooplankton Structure in Relation to Physicochemical Parameters
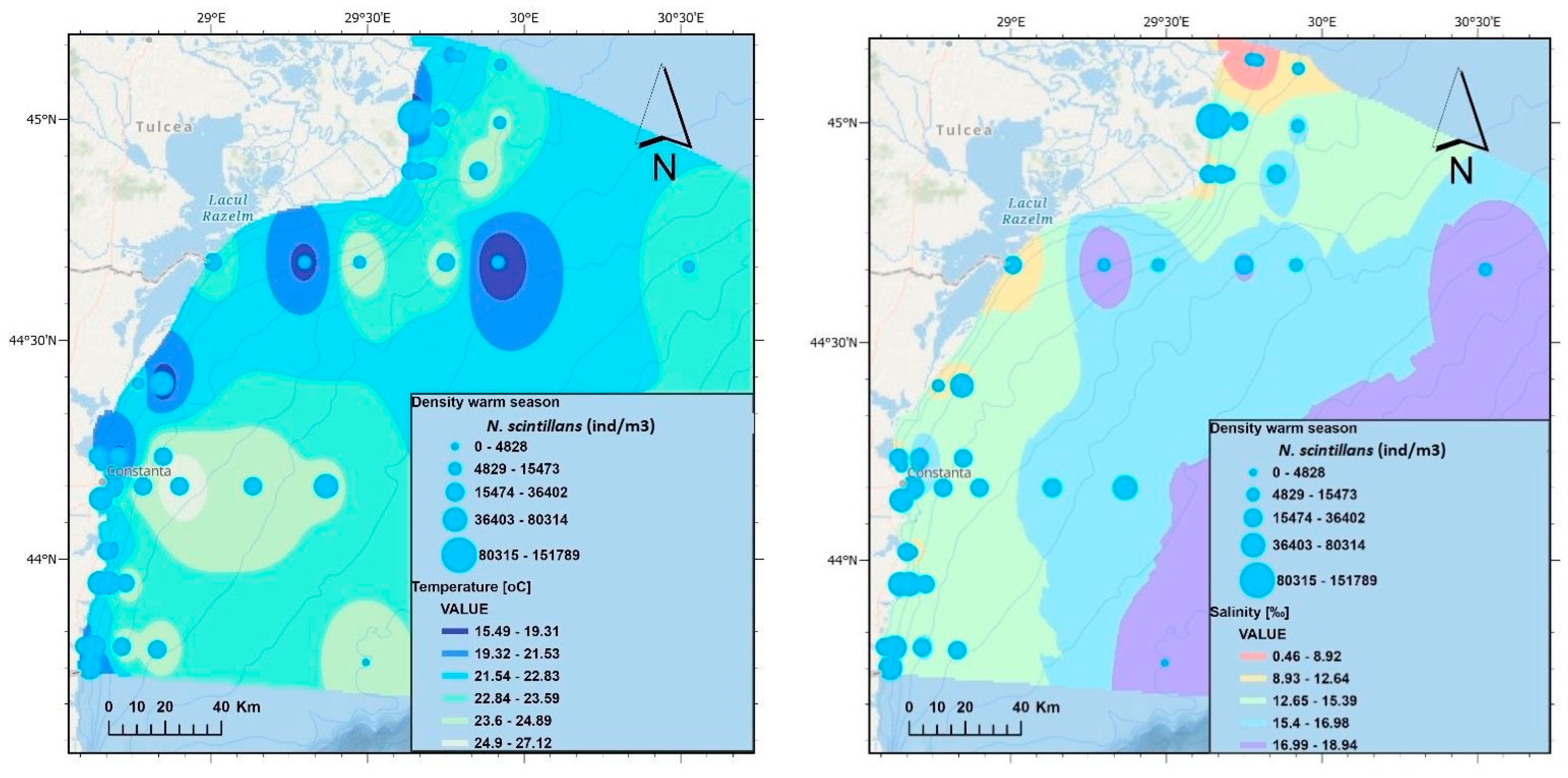
By analysing the non-fodder zooplankton component, represented by the dinoflagellate N. scintillans, in relation to the temperature in the warm seasons of 2013–2020, it is observed that the highest densities (80, 315–15, 1789 ind/m3) were recorded in the northern sector at temperatures between 19 °C and 20 °C (Figure 8). N. scintillans recorded similar values in all three sectors at temperatures between 23.6 °C and 24.9 °C. Regarding the salinity in the warm season, it is observed that at salinities between 16.99‰and 18.94‰, N. scintillans recorded the lowest density values, with the dinoflagellate being much better represented quantitatively at salinities between 8.93‰ and 16.98‰ and the maximum density being reached in the northern sector at a salinity of 12.65–15.39 PSU (Figure 8).
The fodder zooplankton of the warm season recorded high densities. It is noted that the highest values (60.893–114,494 ind/m3) were recorded in shallow waters (Figure 9). The density of the fodder zooplankton in the warm season showed variations in relation to salinity, with the highest density being recorded in the southern and central sectors at salinities of 12.65–15.39‰ (Figure 9). It is observed that the lowest densities were reached in the offshore area, where the salinity was higher, between 16.99‰ and 18.94‰ (Figure 9).
Although it had much lower density values than those in the warm seasons, the dinoflagellate N. scintillans was also present in the cold seasons of 2013–2020. It is noteworthy that the species developed in all three sectors and in all the temperature ranges (Figure 10). The same pattern was observed for the salinity, where the species was found in all the salinity ranges in the cold season, even at 0.59–8.92‰, a range specific to the northern sector of the Romanian Black Sea coast under the direct influence of the Danube River (Figure 10).
Variations in the zooplankton abundance are very clear in the cold seasons of the period 2013–2020, with the maximum density values being much lower compared to those in the warm seasons. It is observed that in the cold season, the zooplankton community recorded density values between 5164–25,028 ind/m3, regardless of the temperature and salinity values (Figure 11).
4. Discussion
4.1. Zooplankton Composition
Significant long-term changes in the zooplankton community occurred in the Black Sea between 1950 and 1970 and in 1995 owing to a remarkable increase in the anthropogenic impact on the ecosystem of the Black Sea, which noticeably affected the species composition of zooplankton assemblages [37]. The most important feature of the zooplankton community after the 1970s was the change in species composition; some species almost disappeared, whereas others (N. scintillans) increased in their abundance. These events affected mostly the northwestern part of the Black Sea, where the nutrient enrichment supplied by Danube, Dniester, and Dnieper runoffs was higher. The first signs of ecosystem rehabilitation appeared after 1994 because of pollution and eutrophication reductions and changes in the hydro-climatic regime in the warm climatic cycle [38].
Thus, according to Porumb (1977, 1994–1995) [39,40] the core of the holoplankton from the Romanian Black Sea consisted mainly of N. scintillans; calanoid copepods represented by C. euxinus (ponticus), P. elongatus, P. parvus, C. ponticus, and A. clausi; to which are also added cyclopoid copepods—O. nana and O. similis; Cladocera species, such as P. avirostris, P. polyphemoides, E. spinifera, and P. tergestina; the chaetognath P. setosa (S. setosa); and the appendicular O. dioica.
In the 1970s–1980s, the abundance of common species greatly decreased [41]. This change was further pronounced after the invasion of the ctenophore Mnemiopsis leidyi, which caused major changes in the structure and dynamics of pelagic communities in the Black Sea after 1988, reflecting the stressed state of the pelagic habitat dominated by the influence of the ctenophore [3]. M. leidyi flourished in the food web along the western coastal waters during 1989–1992 [42] owing to its wide prey base (e.g., ciliates, copepods of various sizes, nauplii, cirripeds larvae, bivalves, eggs, and fish larvae) [3]. The previously dominant cyclopoid O. nana and the calanoid Labidocera brunescens no longer occur in zooplankton samples [43]. In the northwestern Black Sea, the abundance of hyponeustonic copepods (Pontellidae) decreased between 1960 and 1980. In the same region, P. parvus, C. ponticus, P. elongatus, C. euxinus (helgolandicus), P. setosa (S. setosa), P. avirostris, and E. spinifera decreased in the same period [44] All these changes observed in the pelagic system of the Black Sea seem to be due to pollution and heavy predation by M. leidyi on zooplanktonic organisms. The zooplankton community was drastically affected by the population outburst of the alien ctenophore species M. leidyi after 1988, causing changes in the water’s physical properties by reducing transparency and, more significantly, the biological properties by causing a cascade effect on all the trophic levels, favouring phytoplankton growth and reducing food resources for planktivorous and predatory fishes. [45]. The introduction of its predator Beroe ovata, which came from either the Mediterranean Sea or the eastern coast of the North Atlantic through ballast waters in 1997, helped the later recovery of the ecosystem [45]. B. ovata was first encountered in the western shelf [46] and the northeastern basin in the summer of 1997 [47]. In addition, the entire planktonic component was affected by the severe climatic cooling regime in the 1980s, followed by a strong warming regime in the 1990s [48].
The physical and chemical environment regulates the size, structure, and taxonomic composition of plankton communities [10]. Most of the zooplankton in marine pelagic ecosystems usually consists of copepods, and these small crustaceans often constitute up to 80% of the zooplankton biomass [49]. In 2010, a new zooplanktonic species was identified for the Romanian waters of the Black Sea—the cyclopoid copepod Oithona brevicornis, a species reported for the first time in the waters of the Black Sea in 2001, in the Gulf of Sevastopol, and in the form of two isolated specimens in 2005 and 2006, which were observed in the same location but with much more abundant populations and in all the developmental stages [30]. This species was most likely brought to the Black Sea in the ballast water of ships [50], occupying the niche of O. nana [51], a species that was a valuable food source for fish larvae [52]. This species was also reported on the Romanian coastline, being first identified as O. brevicornis [53]. For the period 2013–2020, the copepod was identified and considered as O. davisae, according to studies concluding that the cyclopoid species recently recorded as new to the Black Sea was not O. brevicornis but O. davisae [51]. Species of a holoplanktonic nature that were very sensitive to environmental quality and eutrophication, such as C. ponticus, P. avirostris, and P. setosa (S. setosa), began to recover the populations [38]. As emphasised by this study, the mentioned species were prominently represented over the examined period.
The Shannon–Wiener diversity index (H′) is a suitable indicator of water quality; high values of this index indicate greater species diversity or even the distribution of the species, while lower values indicate poor species composition. This index also determines the pollution status of a water body [54].
In the ongoing study, the diversity indices indicate that the environment exhibited non-uniformity throughout the research period. Consequently, the population density of the zooplankton has consistently fluctuated in response to the varying environmental conditions.
The pelagic environments of the Eastern Mediterranean and the Black Sea present some common features due to their connection through the Dardanelles and Bosphorus Straits. Similarities are recorded in flora and fauna compositions [54], the zooplanktonic communities in both regions are mainly represented by species of Atlantic origin. However, noticeable differences in hydrology, temperature, salinity, and biological production are found in these two distinct areas, driven by geographical and geo-morphological characteristics [55].
Limited species exchange and the lack of adaptation of Mediterranean species to the Black Sea even occur now under climate warming, habitat niche vacancy, and eutrophication. Similar peculiarities are also found in the Baltic Sea owing to biogeographical barriers to species expansions [56]. Therefore, both the Black and Marmara Seas have recorded low plankton and fish species biodiversities since the beginning of the post-glacial phase [55].
The low species biodiversity of the Black Sea (Table 3) is a sign of the ineffective bio-invasion process of Mediterranean species. This phenomenon is explained by strong differences in physical gradients and some physiological limitations along the colonization route of the Turkish Straits System (TSS), which make Mediterranean plankton species vulnerable to long-distance natural expansions. In conclusion, the Mediterranization of the Black Sea is an ongoing process under the prevailing physical dispersal and physiological limitations driven by peculiar horizontal and vertical variations in temperature, salinity, and dissolved oxygen concentration along the TSS. Climate change may have been a facilitating factor for a recent increase in new species observations in the TSS and the Black Sea [57].
4.2. Zooplankton Structure–Seasonal Variations
The tendency of zooplankton evolution, both in terms of qualitative and quantitative structures, depends on the levels of nutrient input to the sea [58]. With the increase in water temperature, species of warm waters appear in the water mass, while those of cold waters migrate to the colder water layers [59]. The qualitative variety of zooplankton increases in the warm period, with copepods and Cladocera being the best represented both qualitatively and quantitatively [60], as shown in the present paper (Figure 3).
Abiotic factors, like water temperature, and biotic factors, such as food availability and predator-induced stress, influence the distribution of zooplankton [61]. Studies on marine copepods showed that across the temperature range from −1.7 °C to 30 °C, the body size decreased by 43.9% [62]. This suggests that under the phenomenon of global warming, smaller copepod species are likely to dominate, with effects on fisheries and carbon sequestration [62,63,64]. The energetic demands of zooplankton are higher under warming conditions; therefore, nutrient limitation also increases and leads to smaller zooplankton body sizes, with negative effects on the community and carbon export [63,65]. Regional zooplankton communities present seasonal shifts between a cold and a warm season as a result of changes in environmental variables [66]. Seasonal changes in the zooplankton structure are also recorded in the central region of the Mediterranean Sea and the Sea of Azov, with variations within similar limits. The range of seasonal changes in the quantitative indices of zooplankton in the central regions increases from the Mediterranean Sea to the Sea of Azov, corresponding to the increase in the range of seasonal variations in the temperature and quantity of phytoplankton [55].
4.3. Zooplankton Structure in Relation to Physicochemical Parameters
The distribution of the zooplankton community depends on several factors, such as climatic conditions, water physical and chemical parameters, and biological interactions [67]. Zooplankton changes in response to changes in water quality and environmental parameters influencing the changes in zooplankton abundance [68].
Seasonality, with significant changes in the temperature and salinity regime, influences the food and nutrients available to zooplankton and is an important driver of community distribution [69]. We observed that the fodder component records high abundances in the warm season. N. scintillans, the non-fodder component of the zooplankton community, is a dominant species in the Black Sea and is a voracious predator of phytoplankton, bacteria, detritus, microzooplankton, eggs, and naupliar stages of copepods [70]. It has almost no predator (except for P. setosa and a limited number of copepod species) owing to its low carbon and high ammonia contents [3]. Although the increase in the abundance of N. scintillans is affected by temperature and salinity, it is known to be an eurythermal and euryhaline organism [71]. Many previous research results have shown that the optimal temperature and salinity demands of N. scintillans are different in each ecosystem [72,73,74]. Therefore, it is difficult to discover only the effect of temperature and salinity on N. scintillans bloom formation [75]. Moreover, zooplankton, like all aquatic organisms, require oxygen for respiration. Adequate dissolved oxygen levels are essential for their survival and metabolic functions. Low dissolved oxygen levels (hypoxia) can lead to stress or mortality in zooplankton populations. Some species are more tolerant of hypoxic conditions than others. Hypoxic events can also disrupt zooplankton vertical migration patterns, impacting their feeding and predator avoidance behaviours [76,77]. Nutrient availability, especially nitrogen and phosphorus, can influence primary productivity in the water column, which subsequently affects zooplankton food sources (phytoplankton) [78]. High nutrient concentrations can lead to phytoplankton blooms, providing a surplus of food for zooplankton. This can result in population explosions known as “bloom and bust” cycles. Nutrient availability affects the nutritional quality of phytoplankton, which, in turn, influences the growth and reproduction of zooplankton that feed on them [79].
Thus, the dynamics of zooplankton populations in marine ecosystems are significantly influenced by a combination of environmental factors, including the temperature, salinity, and dissolved oxygen and nutrient concentrations [76,80,81].
The analysis of the most pronounced significant correlations, as presented in Tables S7–S9, underscores the substantial impact of temperature fluctuations across different seasons on the overall growth of the zooplankton population, encompassing both fodder and non-fodder species. Furthermore, the interplay of temperature and salinity variations emerges as a major influence on the development of “other groups” and meroplankton. Notably, “other groups” show a positive correlation with salinity, while meroplankton densities exhibit a negative one. The examination of correlations within individual seasons reveals that temperature and salinity continue to hold importance, with the warm season exhibiting stronger relationships. This is particularly pronounced given the heightened environmental parameter variability during this period, as indicated by the raised standard deviation (as detailed in Tables S3–S5). Climate change has led to significant alterations in seawater temperature and salinity patterns. Rising global temperatures are causing an increase in sea surface temperatures, impacting marine ecosystems. This warming is often accompanied by changes in salinity due to various factors, including altered precipitation and river flow patterns and changes in sea circulation. These shifts in temperature and salinity can affect zooplankton species/group distributions and influence the functioning of marine ecosystems, underscoring the complex interplay between climate change and sea dynamics.
Additionally, an intriguing correlation emerged exclusively during the cold season, highlighting the connection between non-fodder zooplankton densities and nitrite concentrations (Figure 12). Notably, the presence of N. scintillans blooms serves as an indicator of eutrophication in this specific area of the Black Sea, aligning with the criteria set forth by the Marine Strategy Framework Directive implementation in Romania.
Canonical Correspondence Analysis (CCA) examined the connection between the presence of five zooplankton species or groups and various environmental factors, such as temperature, salinity, dissolved oxygen, dissolved oxygen saturation, and nutrient levels (phosphate, silicate, nitrate, nitrite, and ammonium), considering the two seasons—warm and cold. The null hypothesis, which stated that there was no linear relationship between the species data and the environmental variables, was decisively rejected (p < 0.0001) (Figure 13).
The significance of each environmental variable in influencing the factors is evident, with salinity and nitrate (F1) making a noteworthy contribution and salinity, nitrate, and temperature (F2) also playing a substantial role (Table 4). When considered together, F1 and F2 account for 86.45% of the variation in zooplankton abundance (Figure 14). Among these variables, copepods are predominantly affected by salinity and nitrate concentrations, while Cladocera and N. scintillans exhibit a stronger association with temperature and ammonium levels (Figure 14).
The results of our investigation highlight the dynamic nature of the environment during the study period, leading to fluctuations in zooplankton population density. Notably, salinity and nitrate (F1) emerge as key influencers; and, when combined with temperature in F2, they collectively account for a significant 86.45% of the variability in zooplankton abundance (Figure 14). This underscores the importance of monitoring and understanding the interplay of these environmental factors in aquatic ecosystems. Furthermore, our findings reveal specific associations, such as the influence of salinity and nitrate on copepods and the impact of temperature and ammonium levels on Cladocera and N. scintillans (Figure 14). The findings are significant in the context of climate changes, which can result in altered precipitation patterns, leading to increased freshwater and nutrients input into coastal areas and in marine circulation changes, which can affect the water column stratification, leading to variations in temperature, salinity, and nutrient levels. Coastal areas stretching from the Mediterranean Sea to the Sea of Azov experience seasonal fluctuations in both zooplankton abundance and biomass. These variations are influenced by diverse factors, including temperature and the availability of food resources for zooplankton, which can vary under different conditions [5].
The seasonal shifts in the coastal areas of the Mediterranean Sea are a result of variations in both regional and local environmental conditions. To these changes, the influence of human activities on the ecosystem is further added [82]. In the Baltic Sea, salinity has been established as a crucial regulating factor for large neritic copepods, as demonstrated by Vuorinen et al. in 1998 and related studies. However, Hänninen et al., in 2000, revealed that there exists a significant time delay of approximately one year between freshwater runoff and salinity levels at a depth of 200 m [83].
5. Conclusions
The zooplankton taxa showed strong variability in species composition starting in the early 1970s; this fact was identified over the entire Black Sea basin and was mainly generated by eutrophication and anthropogenic impacts, to which the invasion of the ctenophore M. leidyi was added. Starting in 1995, signs of improvement in the specific diversity of the zooplankton were observed, with species that were very sensitive to environmental conditions being frequently recorded. M. leidyi invasion in the Black Sea occurred in an already disturbed ecosystem, with eutrophication leading to a reduction in zooplankton diversity and quantity values, especially for the fodder component, which has a trophic value for planktivorous fishes.
However, after 1995, the zooplankton community had a stable trend, with species that were scarcely represented becoming abundant. The community structure of the zooplankton in 2013–2020 had a significant seasonal pattern, with the occurrence of cladocerans being negligible in the cold season but appreciable in the warm one. The share of copepods that play a leading role in fisheries was quite similar in both seasons, with the highest values in the warm one.
The results of our investigation highlight the dynamic nature of the environment during the study period, leading to fluctuations in zooplankton population density. Notably, salinity and nitrate emerge as key influencers; and, when combined with temperature, they collectively account for a significant 86.45% of the variability in the zooplankton abundance. These underline the importance of monitoring and understanding the interplay of these environmental factors in aquatic ecosystems. Furthermore, our findings reveal specific associations, such as the influence of salinity and nitrate on copepods and the impact of temperature and ammonium levels on Cladocera and N. scintillans. This knowledge contributes valuable insights into the intricate relationships between environmental variables and zooplankton populations, which can inform future research and conservation efforts in aquatic ecosystems and the ecological implications of climate change for effective marine management strategies.
Understanding the distribution and abundance of zooplankton under the influence of physical and chemical processes may shape the effects of natural and anthropogenic pressures on the ecosystem structure and hold significant importance for the environment. As future research, a comprehensive understanding of how climate change affects zooplankton development and its broader ecological implications is of the utmost importance. Such knowledge is essential for informed decision-making, conservation efforts, and the development of strategies to mitigate and adapt to the challenges posed by a changing climate.
To increase our current understanding of the changes in the Black Sea biodiversity through natural and human-mediated processes, future observational studies necessitate the development of a cross-disciplinary collaboration [57].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15091024/s1. Table S1: List of zooplankton taxa identified until 2020 along the Romanian Black Sea coastline (after Timofte, F., 2017 and personal observations); Table S2: List of zooplankton taxa identified along the Romanian Black Sea coastline in 2013–2020; Table S3: Descriptive statistics of physicochemical parameters under study, both seasons; Table S4: Descriptive statistics of physicochemical parameters under study, cold season; Table S5: Descriptive statistics of physicochemical parameters under study, warm season; Table S6: Statistical differences of physicochemical parameters under study, cold and warm seasons; Table S7: Correlations (all) between biological and environmental parameters; Table S8: Correlations (warm season) between biological and environmental parameters; Table S9: Correlations (cold season) between biological and environmental parameters.
Author Contributions
Conceptualization, E.B. and L.L.; methodology, E.B. and L.L.; software, E.B. and L.L.; validation, E.B., L.L. and F.T.; formal analysis, E.B., L.L. and F.T.; investigation, E.B., L.L. and F.T.; resources, E.B., L.L. and F.T.; data curation, E.B., L.L. and F.T.; writing—original draft preparation, E.B. and L.L.; writing—review and editing, L.L. and F.T.; visualization, L.L. and E.B.; supervision L.L. and F.T.; funding acquisition, E.B., L.L. and F.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Nucleu Programme -CEMAR 2012–2015 Contract no. 32N/2009, PROMARE 2016–2017 Contract no. 35N/2016, SIMAR 2018 Contract no. 40N/2018 INTELMAR 2019–2022 Contract no. 45N/2019) and by the Integrated monitoring program of the marine ecosystem at the Black Sea according to the requirements of art. 11 of the Marine Strategy Framework Directive (2008/56/EC). The APC was funded by the Ministry of Environment, Water and Forests, Monitoring Study, Contract no. 50/21.04.2023.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are belonging to National Institute for Marine Research and Development “Grigore Antipa”—NIMRD and can be accessed by the requirement to http://www.nodc.ro/data_policy_nimrd.php (accessed on 29 August 2023).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analysis, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Caroppo, C.; Buttino, I.; Camatti, E.; Caruso, G.; De Angelis, R.; Facca, C.; Magaletti, E. State of the Art and Perspectives on the Use of Planktonic Communities as Indicators of Environmental Status in Relation to the EU Marine Strategy Framework Directive. Biol. Mar. Mediterr. 2013, 20, 65–73. [Google Scholar]
- Kiko, R.; Bianchi, D.; Grenz, C.; Hauss, H.; Iversen, M.; Kumar, S.; Maas, A.; Robinson, C. Editorial: Zooplankton and Nekton: Gatekeepers of the Biological Pump. Front. Mar. Sci. 2020, 7, 545. [Google Scholar] [CrossRef]
- Oguz, T.; Velikova, V. Abrupt Transition of the Northwestern Black Sea Shelf Ecosystem from a Eutrophic to an Alternative Pristine State. Mar. Ecol. Prog. Ser. 2010, 405, 231–242. [Google Scholar] [CrossRef]
- Daskalov, G.M. Overfishing Drives Atrophic Cascade in the Black Sea. Mar. Ecol. Prog. Ser. 2002, 225, 53–63. [Google Scholar] [CrossRef]
- Kideys, A.E.; Kovalev, A.V.; Shulman, G.; Gordina, A.; Bingel, F. A review of zooplankton investigations of the Black Sea over the last decade. J. Mar. Syst. 2000, 24, 355–371. [Google Scholar] [CrossRef]
- Erdogan, N.; Duzgunes, E.; Ogut, H. Black Sea fisheries and climate change. In CIESM Workshop Monographs; CIESM: Monaco, 2009. [Google Scholar]
- Drira, Z.; Kmiha-Megdiche, S.; Sahnoun, H.; Pagano, M.; Tedetti, M.; Ayadi, H. Water Quality Affects the Structure of Copepod Assemblages along the Sfax Southern Coast (Tunisia, Southern Mediterranean Sea). Mar. Freshw. Res. 2018, 69, 220–231. [Google Scholar] [CrossRef]
- Onciu, T.M.; Skolka, M.; Gomoiu, M.T. Ecologia Comunităților Zooplanctonice de la Litoralul Românesc al Mării Negre; Ovidius University Press: Constanța, Romania, 2006. [Google Scholar]
- Liu, H.; Zhang, X.; Yang, Q.; Zuo, T.; Quigg, A. Mesozooplankton Dynamics in Relation to Environmental Factors and Juvenile Fish in a Subtropical Estuary of the Gulf of Mexico. J. Coast. Res. 2017, 335, 1038–1050. [Google Scholar] [CrossRef]
- Richardson, A.J. In Hot Water: Zooplankton and Climate Change. ICES J. Mar. Sci. 2008, 65, 279–295. [Google Scholar] [CrossRef]
- Schroeder, A.; Stanković, D.; Pallavicini, A.; Gionechetti, F.; Pansera, M.; Camatti, E. DNA Metabarcoding and Morphological Analysis—Assessment of Zooplankton Biodiversity in Transitional Waters. Mar. Environ. Res. 2020, 160, 104946. [Google Scholar] [CrossRef]
- Jeelani, M.; Kaur, H.; Kumar, R. Impact of Climate Warming on the Biodiversity of Freshwater Ecosystem of Kashmir, India. In Proceedings of the Taal 2007: The 12th World Lake Conference, Jaipur, Rajasthan, India, 28 October–2 November 2007; Volume 1103, p. 1109. [Google Scholar]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef]
- Ferrara, O.; Vagaggini, D.; Margaritora, F.G. Zooplankton Abundance and Diversity in Lake Bracciano, Latium, Italy. J. Limnol. 2002, 61, 169–175. [Google Scholar] [CrossRef]
- Jeppesen, E.; Nõges, P.; Davidson, T.A.; Haberman, J.; Nõges, T.; Blank, K.; Lauridsen, T.L.; Søndergaard, M.; Sayer, C.; Laugaste, R.; et al. Zooplankton as Indicators in Lakes: A Scientific-Based Plea for Including Zooplankton in the Ecological Quality Assessment of Lakes According to the European Water Framework Directive (WFD). Hydrobiologia 2011, 676, 279–297. [Google Scholar] [CrossRef]
- Kehayias, G.; Chalkia, E.; Doulka, E. Zooplankton Variation in Five Greek Lakes. In Zooplankton; Nova Science Publisher, Inc.: Hauppauge, NY, USA, 2014. [Google Scholar]
- Preston, N.D.; Rusak, J.A. Homage to Hutchinson: Does Inter-Annual Climate Variability Affect Zooplankton Density and Diversity? Hydrobiologia 2010, 653, 165–177. [Google Scholar] [CrossRef]
- Caroni, R.; Irvine, K. The Potential of Zooplankton Communities for Ecological Assessment of Lakes: Redundant Concept or Political Oversight? Biol. Environ. Proc. R. Ir. Acad. 2010, 110, 35–53. [Google Scholar] [CrossRef]
- Lazăr, L.; Boicenco, L.; Moncheva, S.; Denga, Y.; Atabay, H.; Abaza, V.; Bişinicu, E.; Coatu, V.; Filimon, A.; Harcotă, G.; et al. Impact of the Rivers on the Black Sea Ecosystem; CD Press: Bucharest, Romania, 2021. [Google Scholar]
- Damir, N.A.; Coatu, V.; Pantea, E.D.; Galațchi, M.; Botez, E.; Birghilă, S. Assessment of Polycyclic Aromatic Hydrocarbons Content in Marine Organisms of Commercial Interest from the Romanian Black Sea Coast. Polycycl. Aromat. Compd. 2022, 42, 7595–7606. [Google Scholar] [CrossRef]
- Alexandrov, B.; Arashkevich, E.; Gubanova, A.; Korshenko, A. Manual for Mesozooplankton Sampling and Analysis in the BlackSea Monitoring; Black Sea Commission: Istanbul, Turkey, 2014. [Google Scholar]
- Mordukhay-Boltovskoy, F.D. (Ed.) . Guide of the Black Sea and the Sea of Azov Fauna; Naukova Dumka: Kiev, Ukraine, 1972. [Google Scholar]
- Mordukhay-Boltovskoy, F.D. (Ed.) . Identification Manual on the Fauna of the Black and Azov Seas; Naukova Dumka: Kiev, Ukraine, 1968. [Google Scholar]
- Petipa, T.S. On the Mean Weight of the Principle Forms of Zooplankton in the Black Sea. Sevast. Biol. Stn. 1957, 9, 39–57. [Google Scholar]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis, 3rd ed.; Grasshoff, K., Kremling, K., Ehrhardt, M., Eds.; Willey-VCH: Weinheim, Germany, 1999. [Google Scholar]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J. Change in Marine Communities: An Approach to Statistical Analysis, 3rd ed.; PRIMER-E: Plymouth, UK, 2014. [Google Scholar]
- Addinsoft, Inc. Addinsoft XLSTAT Software; Version 2021.2.1; Addinsoft, Inc.: New York, NY, USA, 2021. [Google Scholar]
- TIBCO Software, Inc. TIBCO Statistica; Version 14.0.1.25; TIBCO Software, Inc.: Palo Alto, CA, USA, 2023. [Google Scholar]
- ESRI. ArcGIS Desktop; Version 10.7; Environmental Systems Research Institute: Redlands, CA, USA, 2019. [Google Scholar]
- Gubanova, A.; Altukhov, D. Establishment of Oithona brevicornis Giesbrecht, 1892 (Copepoda: Cyclopoida) in the Black Sea. Aquat. Invasions 2007, 2, 407–410. [Google Scholar] [CrossRef]
- Niette, V. Huliselan. The Role of Zooplankton Predator, Chaetognaths (Sagitta Spp.) in Baguala Bay Waters, Ambon Island. J. Coast. Dev. 2002, 6, 9–21. [Google Scholar]
- Masunaga, A.; Liu, A.W.; Tan, Y.; Scott, A.; Luscombe, N.M. Streamlined Sampling and Cultivation of the Pelagic Cosmopolitan Larvacean. J. Vis. Exp. 2020, 160, e61279. [Google Scholar] [CrossRef]
- Sato, R.; Tanaka, Y.; Ishimaru, T. House Production by Oikopleura dioica (Tunicata, Appendicularia) Under Laboratory Conditions. J. Plankton Res. 2001, 23, 415–423. [Google Scholar] [CrossRef]
- Tiselius, P.; Petersen, J.; Nielsen, T.; Maar, M.; Møller, E.; Satapoomin, S.; Tönnesson, K.; Zervoudaki, T.; Christou, E.; Giannakourou, A.; et al. Functional Response of Oikopleura dioica to House Clogging Due to Exposure to Algae of Different Sizes. Mar. Biol. 2003, 142, 253–261. [Google Scholar] [CrossRef]
- Byrne, P. Seasonal Composition of Meroplankton in the Dunkellin Estuary, Galway Bay. Biol. Environ. Proc. R. Ir. Acad. 1995, 95, 35–48. [Google Scholar]
- Üstün, F.; Bat, L.; Mutlu, E. Seasonal Variation and Taxonomic Composition of Mesozooplankton in the Southern Black Sea (off Sinop) between 2005 and 2009. Turk. J. Zool. 2018, 42, 541–556. [Google Scholar] [CrossRef]
- Kıdeyş, A.E.; Bingel, F.; Niermann, U. The Effect of Environmental conditions on the Distribution of Eggs and Larvae of Anchovy (Engraulis encrasicolus L.) in the Black Sea. ICES J. Mar. Sci. 1999, 24, 58–64. [Google Scholar] [CrossRef]
- Shiganova, T.; Musaeva, E.; Arashkevich, E.; Shirshov, P.P.; Kamburska, L.; Stefanova, K.; Mihneva, V.; Polishchuk, L.; Timofte, F.; Ustun, F.; et al. The State of Zooplankton. In State of the Environment of the Black Sea (2001–2006/7); Oguz, T., Ed.; The Commission on the Protection of the Black Sea Against Pollution Publication: Istanbul, Turkey, 2007; pp. 201–246. [Google Scholar]
- Porumb, F. Le Zooplancton de La Mer Noire. Biologie Des Eaux Saumâtres de La Mer Noire. IRCM 1977, 1, 99–108. [Google Scholar]
- Porumb, F. L’Histoire Des Recherches Marines Roumaines En Mer Noire; Inst. Roman de Cercetari Marine: Constanta, Romania, 1995; pp. 32–33. [Google Scholar]
- Zaitsev, Y. Recent Changes in the Trophic Structure of the Black Sea. Fish. Oceanogr. 1992, 1, 180–189. [Google Scholar] [CrossRef]
- Kamburska, L.; Schrimpf, W.; Djavidnia, S.; Shiganova, T.; Stefanova, K. Adressing the Ecological Issue of the Invasive Species Special Focus on the Ctenophore Mnemiopsis Leidy (Agassiz, 1865) in the Black Sea; Office for Official Publications of the European Communities: Luxembourg, 2006. [Google Scholar]
- Kovalev, A.V.; Finenko, Z.Z. Macrozooplankton: Plankton of the Black Sea; Naukova Dumka: Kiev, Ukraine, 1993; pp. 183–193. [Google Scholar]
- Polischuk, L.N.; Nastenko, E.V.; Garkavaya, G.P. Some Peculiarities of Modern State of Pelagic and Neustonic Zoocenosis of the Black Sea. Ecol. Morya 1984, 18, 25–34. [Google Scholar]
- Shiganova, T.A.; Shirshov, P.P. Invasion of the Black Sea by the Ctenophore Mnemiopsis leidyi and Recent Changes in Pelagic Community Structure. Fish. Oceanogr. 1998, 7, 305–310. [Google Scholar] [CrossRef]
- Konsutov, A.S.; Kamburska, L.T. Ecological Determination of the New Ctenophora-Beroe ovata invasion in the Black Sea. Oceanology 1998, 2, 195–198. [Google Scholar]
- Shiganova, T.A.; Dumont, H.J.; Mikaelyan, A.; Glazov, D.M.; Bulgakova, Y.V.; Musaeva, E.I.; Studenikina, E. Interaction between the Invading Ctenophores Mnemiopsis Leidyi (A. Agassiz) and Beroe ovata Mayer 1912, and Their Influence on the Pelagic Ecosystem of the Northeastern Black Sea. In Aquatic Invasions in the Black, Caspian, and Mediterranean Seas; Dumont, H., Shiganova, T.A., Niermann, U., Eds.; Kluwer Academic Publishers: Alphen aan den Rijn, The Netherlands, 2004; Volume 35, pp. 33–70. [Google Scholar]
- Oguz, T.; Dippner, J.W.; Kaymaz, Z. Climatic Regulation of the Black Sea Hydro-Meteorological and Ecological Properties at Interannual-to-Decadal Time Scales. J. Mar. Syst. 2006, 60, 235–254. [Google Scholar] [CrossRef]
- Vargas, C.A.; Martínez, R.A.; Escribano, R.; Lagos, N.A. Seasonal Relative Influence of Food Quantity, Quality, and Feeding Behaviour on Zooplankton Growth Regulation in Coastal Food Webs. J. Mar. Biol. Assoc. U. K. 2010, 90, 1189–1201. [Google Scholar] [CrossRef]
- Petran, A.; Moldoveanu, M. Post-Invasion Ecological Impact of The Atlantic Ctenophore Mnemiopsis leidyi Agassiz, 1865 On the Zooplankton from The Romanian Black Sea Waters. Cercet. Mar. 1995, 27–28, 135–157. [Google Scholar]
- Shiganova, T.; Stupnikova, A.; Stefanova, K. Genetic Analyses of Non-Native Species Oithona davisae Ferrari, F.D. & Orsi, 1984 in the Black Sea. Bioinvasions Rec. 2015, 4, 91–95. [Google Scholar] [CrossRef]
- Tkach, A.V. Changes in the Larvae Nutrition of the Black Sea Fishes with Respect to Plankton. NATO Sci. Ser. 2 Environ. Secur. 1997, 47, 235–248. [Google Scholar]
- Timofte, F.; Tabarcea, C. Oithona brevicornis Giesbrecht, 1892 (Copepoda: Cyclopoida)—First Record in the Romanian Black Sea Waters. J. Environ. Prot. Ecol. 2012, 13, 1683–1687. [Google Scholar]
- Islam, M.S.; Azadi, M.A.; Nasiruddin, M.; Sarker, M.M. Plankton Species Composition, Abundance and Diversity Indices In Three Ponds Of Chittagong University Campus, Bangladesh. Fish. Aquac. J. 2022, 14, 321. [Google Scholar]
- Kovalev, A.V.; Mazzocchi, M.G.; Siokou-Frangou, I.; Kideys, A.E. Zooplankton of the Black Sea and the Eastern Mediterranean: Similarities and Dissimilarities. Mediterr. Mar. Sci. 2001, 2, 69–77. [Google Scholar] [CrossRef]
- Leppäkoski, E.; Gollasch, S.; Gruszka, P.; Ojaveer, H.; Olenin, S.; Panov, V. The Baltic—A Sea of Invaders. Can. J. Fish. Aquat. Sci. 2002, 7, 1175–1188. [Google Scholar] [CrossRef]
- Oğuz, T.; Öztürk, B. Mechanisms Impeding Natural Mediterranization Process of Black Sea Fauna. J. Black Sea Mediterr. Environ. 2011, 17, 234–253. [Google Scholar]
- Moldoveanu, M.; Timofte, F. Signs of Marine Ecosystem Rehabilitation Along the Romanian Black Sea Littoral Identified by Zooplankton Indicator after Cercet. Mar. 2004, 35, 87–108. [Google Scholar]
- Timofte, F. Contribuții La Evaluarea Stării Ecologice Actuale a Populațiilor Zooplanctonice Din Apele Românești Ale Mării Negre; Ex Ponto: Constanta, Romania, 2017. [Google Scholar]
- Băcescu, M.; Gomoiu, M.T.; Bodeanu, N.; Petran, A.; Muller, G.I.; Chirilă, V. Dinamica Populațiilor Animale Și Vegetale Din Zona Nisipurilor Fine de La. Nord de Constanța În Condițiile Anilor 1962–1965; Academiei Republicii SocialIste România: Bucharest, Romania, 1967. [Google Scholar]
- Hembre, L.K.; Megard, R.O. Seasonal and Diel Patchiness of a Daphnia Population: An Acoustic Analysis. Limnol. Ocean. 2003, 48, 2221–2233. [Google Scholar] [CrossRef]
- Campbell, M.D.; Schoeman, D.S.; Venables, W.; Abu-Alhaija, R.; Batten, S.D.; Chiba, S.; Coman, F.; Davies, C.H.; Edwards, M.; Eriksen, R.S.; et al. Testing Bergmann’s Rule in Marine Copepods. Ecography 2021, 44, 1283–1295. [Google Scholar] [CrossRef]
- Ratnarajah, L.; Abu-Alhaija, R.; Atkinson, A.; Batten, S.; Bax, N.J.; Bernard, K.S.; Canonico, G.; Cornils, A.; Everett, J.D.; Grigoratou, M.; et al. Monitoring and Modelling Marine Zooplankton in a Changing Climate. Nat. Commun. 2023, 14, 564. [Google Scholar] [CrossRef]
- Brandão, M.C.; Benedetti, F.; Martini, S.; Soviadan, Y.D.; Irisson, J.-O.; Romagnan, J.-B.; Elineau, A.; Desnos, C.; Jalabert, L.; Freire, A.S.; et al. Macroscale Patterns of Oceanic Zooplankton Composition and Size Structure. Sci. Rep. 2021, 11, 15714. [Google Scholar] [CrossRef]
- Brun, P.; Stamieszkin, K.; Visser, A.W.; Licandro, P.; Payne, M.R.; Kiørboe, T. Climate Change Has Altered Zooplankton-Fuelled Carbon Export in the North Atlantic. Nat. Ecol. Evol. 2019, 3, 416–423. [Google Scholar] [CrossRef]
- Figueroa, D.F. Environmental Forcing on Zooplankton Distribution in the Coastal Waters of the Galápagos Islands: Spatial and Seasonal Patterns in the Copepod Community Structure. Mar. Ecol. Prog. Ser. 2021, 661, 49–69. [Google Scholar] [CrossRef]
- Abdulwahab, S.; Rabee, A.M. Ecological Factors Affecting the Distribution of the Zooplankton Community in the Tigris River at Baghdad Region, Iraq. Egypt. J. Aquat. Res. 2015, 41, 187–196. [Google Scholar] [CrossRef]
- Sinu, J.V.; Ajimila, B. Quantitative Composition, Distribution and Abundance of Zooplankton Communities in Relation to Physico-Chemical Parameters from Selected Beaches of Alappuzha in Arabian Sea, Southwest Coast of India. Total Environ. Res. 2023, 7, 100054. [Google Scholar] [CrossRef]
- Evans, L.E.; Hirst, A.G.; Kratina, P.; Beaugrand, G. Temperature-mediated Changes in Zooplankton Body Size: Large Scale Temporal and Spatial Analysis. Ecography 2020, 43, 581–590. [Google Scholar] [CrossRef]
- Fonda Umani, S. Noctiluca Scintillans MACARTNEY in the Northern Adriatic Sea: Long-Term Dynamics, Relationships with Temperature and Eutrophication, and Role in the Food Web. J. Plankton Res. 2004, 26, 545–561. [Google Scholar] [CrossRef]
- Elbrächter, M. and Q. Z. Aspects of Noctiluca (Dinophyceae) Population Dynamics, Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Huang, C.; Qi, Y. The Abundance Cycle and Influence Factors on Red Tide Phenomena of Noctiluca scintillans (Dinophyceae) in Dapeng Bay, the South China Sea. J. Plankton Res. 1997, 19, 303–318. [Google Scholar] [CrossRef]
- Tada1, K.; Plthakpol1, S.; Montani, S. Seasonal Variation in the Abundance of Noctiluca scintillans in the Seto Inland Sea, Japan. Plankton Biol. Ecol. 2004, 51, 7–14. [Google Scholar]
- Miyaguchi, H.; Fujiki, T.; Kikuchi, T.; Kuwahara, V.S.; Toda, T. Relationship between the Bloom of Noctiluca Scintillans and Environmental Factors in the Coastal Waters of Sagami Bay, Japan. J. Plankton Res. 2006, 28, 313–324. [Google Scholar] [CrossRef]
- Turkoglu, M. Red Tides of the Dinoflagellate Noctiluca scintillans Associated with Eutrophication in the Sea of Marmara (the Dardanelles, Turkey). Oceanologia 2013, 55, 709–732. [Google Scholar] [CrossRef]
- Weinstock, J.B.; Vargas, L.; Collin, R. Zooplankton Abundance Reflects Oxygen Concentration and Dissolved Organic Matter in a Seasonally Hypoxic Estuary. J. Mar. Sci. Eng. 2022, 10, 427. [Google Scholar] [CrossRef]
- Kiko, R.; Hauss, H. On the Estimation of Zooplankton-Mediated Active Fluxes in Oxygen Minimum Zone Regions. Front. Mar. Sci. 2019, 6, 741. [Google Scholar] [CrossRef]
- Glibert, P.M. Harmful Algae at the Complex Nexus of Eutrophication and Climate Change. Harmful Algae 2020, 91, 101583. [Google Scholar] [CrossRef] [PubMed]
- Bairagi, N.; Saha, S.; Chaudhuri, S.; Dana, S.K. Zooplankton Selectivity and Nutritional Value of Phytoplankton Influences a Rich Variety of Dynamics in a Plankton Population Model. Phys. Rev. E 2019, 99, 012406. [Google Scholar] [CrossRef]
- Wei, Y.; Ding, D.; Gu, T.; Jiang, T.; Qu, K.; Sun, J.; Cui, Z. Different Responses of Phytoplankton and Zooplankton Communities to Current Changing Coastal Environments. Environ. Res. 2022, 215, 114426. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, H.; Wang, Z.; Huang, T.; Tian, W.; Huang, H. Responses of Zooplankton Community Pattern to Environmental Factors along the Salinity Gradient in a Seagoing River in Tianjin, China. Microorganisms 2023, 11, 1638. [Google Scholar] [CrossRef]
- Kovalev, A.V.; Mazzocchi, M.G. Seasonal Changes in the Composition and Abundance of Zooplankton in the Seas of the Mediterranean Basin. Turk. J. Zool. 2003, 27, 205–219. [Google Scholar]
- Dippner, J.W.; Hänninen, J.; Kuosa, H.; Vuorinen, I. The Influence of Climate Variability on Zooplankton Abundance in the Northern Baltic Archipelago Sea (SW Finland). ICES J. Mar. Sci. 2001, 58, 569–578. [Google Scholar] [CrossRef]
Figure 1.
Map of the study area. The square shows the location of the sampling sites on the Romanian coastline (A) and the position in the Black Sea region (B).
Figure 1.
Map of the study area. The square shows the location of the sampling sites on the Romanian coastline (A) and the position in the Black Sea region (B).

Figure 2.
Number of zooplankton taxa identified on the Romanian Black Sea coastline (1986–2020).

Figure 3.
Matrix of identified zooplankton taxa (taxonomic groups) in the Romanian Black Sea (2013–2020) by years and seasons.
Figure 3.
Matrix of identified zooplankton taxa (taxonomic groups) in the Romanian Black Sea (2013–2020) by years and seasons.

Figure 4.
Dendrogram of cluster analysis using Bray–Curtis similarity, based on zooplankton taxa frequency in the Romanian Black Sea area, by taxonomic groups (2013–2020).
Figure 4.
Dendrogram of cluster analysis using Bray–Curtis similarity, based on zooplankton taxa frequency in the Romanian Black Sea area, by taxonomic groups (2013–2020).

Figure 5.
Boxplot of zooplankton frequency, by groups (2013–2020).

Figure 6.
Dendrogram of cluster analysis using Bray–Curtis similarity, based on the number of zooplankton taxa identified in the Romanian Black Sea area, by seasons (2013–2020).
Figure 6.
Dendrogram of cluster analysis using Bray–Curtis similarity, based on the number of zooplankton taxa identified in the Romanian Black Sea area, by seasons (2013–2020).

Figure 7.
The bar plot of the zooplankton mean density and biomass values, by taxonomic groups and seasons (2013–2020).
Figure 7.
The bar plot of the zooplankton mean density and biomass values, by taxonomic groups and seasons (2013–2020).

Figure 8.
Spatial distributions of non-fodder zooplankton density, water temperature, and salinity on the Romanian coast—warm seasons, 2013–2020.
Figure 8.
Spatial distributions of non-fodder zooplankton density, water temperature, and salinity on the Romanian coast—warm seasons, 2013–2020.

Figure 9.
Spatial distributions of fodder zooplankton density, water temperature, and salinity on the Romanian coast—warm seasons, 2013–2020.
Figure 9.
Spatial distributions of fodder zooplankton density, water temperature, and salinity on the Romanian coast—warm seasons, 2013–2020.

Figure 10.
Spatial distributions of non-fodder zooplankton density, water temperature, and salinity on the Romanian coast—cold seasons of 2014, 2016, and 2017.
Figure 10.
Spatial distributions of non-fodder zooplankton density, water temperature, and salinity on the Romanian coast—cold seasons of 2014, 2016, and 2017.

Figure 11.
Spatial distributions of fodder zooplankton density, water temperature, and salinity on the Romanian coast—cold seasons of 2014, 2016, and 2017.
Figure 11.
Spatial distributions of fodder zooplankton density, water temperature, and salinity on the Romanian coast—cold seasons of 2014, 2016, and 2017.

Figure 12.
Scatterplot of NO2 against Noctiluca scintillans densities in the cold seasons of 2014, 2016, and 2017—Black Sea.
Figure 12.
Scatterplot of NO2 against Noctiluca scintillans densities in the cold seasons of 2014, 2016, and 2017—Black Sea.

Figure 13.
Permutation test (CCA) for zooplankton species groups related to environmental variables.
Figure 13.
Permutation test (CCA) for zooplankton species groups related to environmental variables.

Figure 14.
The Canonical Correspondence Analysis (CCA) map of zooplankton groups and environmental variables.
Figure 14.
The Canonical Correspondence Analysis (CCA) map of zooplankton groups and environmental variables.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary statistics (Qualitative data) by season (2013–2020).
| Variables | Categories | Counts | Frequencies | Rel. Frequency per Category (%) | Taxonomic Groups |
|---|---|---|---|---|---|
| Season | Cold | 1314 | 1314 | 18.1 | |
| Warm | 5940 | 5940 | 81.9 | ||
| Taxa | Acartia clausi | 379 | 379 | 5.2 | Copepoda |
| Balanus | 378 | 378 | 5.2 | Meroplankton | |
| Bivalvia | 378 | 378 | 5.2 | Meroplankton | |
| Bosmina longirostris | 123 | 123 | 1.7 | Cladocera | |
| Calanus euxinus | 364 | 364 | 5.0 | Copepoda | |
| Centropages ponticus | 334 | 334 | 4.6 | Copepoda | |
| Chydorus sphaericus | 47 | 47 | 0.6 | Cladocera | |
| Cyclops sp. | 127 | 127 | 1.8 | Copepoda | |
| Daphnia longispina | 42 | 42 | 0.6 | Cladocera | |
| Decapoda | 249 | 249 | 3.4 | Meroplankton | |
| Diaphanosoma brachyurum | 36 | 36 | 0.5 | Cladocera | |
| Evadne spinifera | 179 | 179 | 2.5 | Cladocera | |
| Gastropoda | 377 | 377 | 5.2 | Meroplankton | |
| Harpacticoida | 350 | 350 | 4.8 | Copepoda | |
| Mesopodopsis slabberi | 202 | 202 | 2.8 | Other groups | |
| Noctiluca scintillans | 379 | 379 | 5.2 | Non-fodder | |
| Oikopleura dioica | 378 | 378 | 5.2 | Other groups | |
| Oithona davisae | 158 | 158 | 2.2 | Copepoda | |
| Oithona similis | 377 | 377 | 5.2 | Copepoda | |
| Paracalanus parvus | 378 | 378 | 5.2 | Copepoda | |
| Parasagitta setosa | 377 | 377 | 5.2 | Other groups | |
| Penilia avirostris | 305 | 305 | 4.2 | Cladocera | |
| Pleopis polyphemoides | 378 | 378 | 5.2 | Cladocera | |
| Podon sp. | 4 | 4 | 0.1 | Cladocera | |
| Polychaeta | 378 | 378 | 5.2 | Meroplankton | |
| Pseudevadne tergestina | 199 | 199 | 2.7 | Cladocera | |
| Pseudocalanus elongatus | 378 | 378 | 5.2 | Copepoda |
Table 2.
Summary of the one-way crossed SIMPER results for the comparison of zooplanktonic groups between seasons (2013–2020).
Table 2.
Summary of the one-way crossed SIMPER results for the comparison of zooplanktonic groups between seasons (2013–2020).
| Warm | ||||||
| Average similarity: 96.14 | ||||||
| Taxonomic group | Av. Abund. | Av. Sim. | Sim./SD | Contrib. % | Cum. % | |
| Copepoda | 2.69 | 27.36 | 27.5 | 28.46 | 28.46 | |
| Meroplankton | 2.16 | 22.32 | 19.19 | 23.21 | 51.67 | |
| Cladocera | 2.05 | 20.49 | 11.8 | 21.32 | 72.99 | |
| Cold | ||||||
| Average similarity: 96.83 | ||||||
| Taxonomic group | Av. Abund. | Av. Sim. | Sim./SD | Contrib. % | Cum. % | |
| Copepoda | 2.54 | 28.35 | 18.77 | 29.27 | 29.27 | |
| Meroplankton | 2 | 23.9 | 35.29 | 24.68 | 53.95 | |
| Other groups | 1.73 | 20.69 | 35.29 | 21.37 | 75.32 | |
| Groups Warm and Cold | ||||||
| Average dissimilarity = 8.85 | ||||||
| Warm | Cold | |||||
| Taxonomic group | Av. Abund. | Av. Abund. | Av. Diss. | Diss./SD | Contrib. % | Cum. % |
| Cladocera | 2.05 | 1.1 | 5.33 | 3.45 | 60.16 | 60.16 |
| Copepoda | 2.69 | 2.54 | 1.44 | 1.29 | 16.24 | 76.4 |
Table 3.
Zooplankton diversity indices, by years and seasons (Shannon–Wiener legislation on the aquatic environment classified water quality as >4 = very good quality, 3–4 = good quality, 2–3 = moderate quality, 1–2 = poor quality, and <1 = very poor quality.).
Table 3.
Zooplankton diversity indices, by years and seasons (Shannon–Wiener legislation on the aquatic environment classified water quality as >4 = very good quality, 3–4 = good quality, 2–3 = moderate quality, 1–2 = poor quality, and <1 = very poor quality.).
| Year | Species Richness (S) | Shannon–Weiner Index (H′) | Pielou’s Evenness Index (J′) | Season |
|---|---|---|---|---|
| 2013 | 25 | 1.8 | 0.6 | Warm |
| 2014 | 18 | 1.4 | 0.5 | Warm |
| 2015 | 18 | 1.0 | 0.4 | Warm |
| 2016 | 19 | 1.9 | 0.7 | Warm |
| 2017 | 18 | 1.2 | 0.4 | Warm |
| 2018 | 21 | 2.3 | 0.8 | Warm |
| 2019 | 25 | 2.0 | 0.6 | Warm |
| 2020 | 23 | 1.6 | 0.5 | Warm |
| 2014 | 16 | 1.5 | 0.5 | Cold |
| 2016 | 15 | 1.3 | 0.5 | Cold |
| 2017 | 18 | 1.7 | 0.6 | Cold |
Table 4.
CCA Regression coefficients.
| F1 | F2 | F3 | F4 | |
|---|---|---|---|---|
| T (°C) | −0.017 | 0.512 | 0.506 | −0.378 |
| S (‰) | −0.704 | 0.887 | −0.415 | −0.400 |
| PO4 (µM) | −0.180 | −0.311 | 0.236 | −0.372 |
| SiO4 (µM) | −0.452 | 0.096 | −0.394 | −0.150 |
| NO2 (µM) | 0.333 | 0.455 | 0.126 | 0.071 |
| NO3 (µM) | −0.506 | −0.649 | 0.262 | −0.207 |
| NH4 (µM) | 0.094 | 0.156 | −0.161 | −0.046 |
| O2 (µM) | 0.258 | 0.363 | −0.459 | −0.887 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bișinicu, E.; Lazăr, L.; Timofte, F. Dynamics of Zooplankton along the Romanian Black Sea Coastline: Temporal Variation, Community Structure, and Environmental Drivers. Diversity 2023, 15, 1024. https://doi.org/10.3390/d15091024
AMA Style
Bișinicu E, Lazăr L, Timofte F. Dynamics of Zooplankton along the Romanian Black Sea Coastline: Temporal Variation, Community Structure, and Environmental Drivers. Diversity. 2023; 15(9):1024. https://doi.org/10.3390/d15091024
Chicago/Turabian StyleBișinicu, Elena, Luminița Lazăr, and Florin Timofte. 2023. "Dynamics of Zooplankton along the Romanian Black Sea Coastline: Temporal Variation, Community Structure, and Environmental Drivers" Diversity 15, no. 9: 1024. https://doi.org/10.3390/d15091024
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.