
Millepora spp. as Substrates of Their Hydrozoan Counterparts Stylaster sp. in the Pacific Ocean

 ,
,  , ,
, , 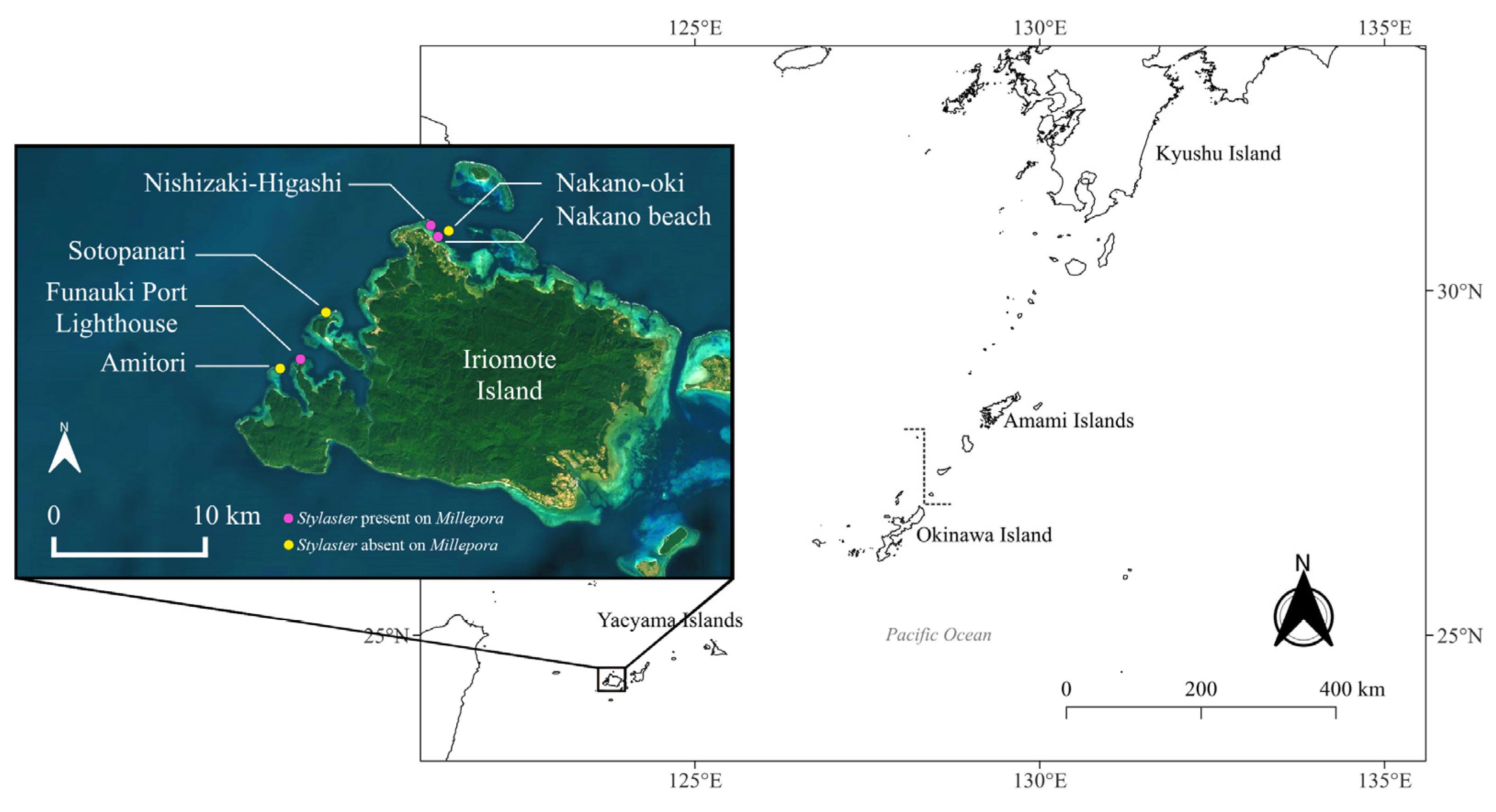{kind=link}
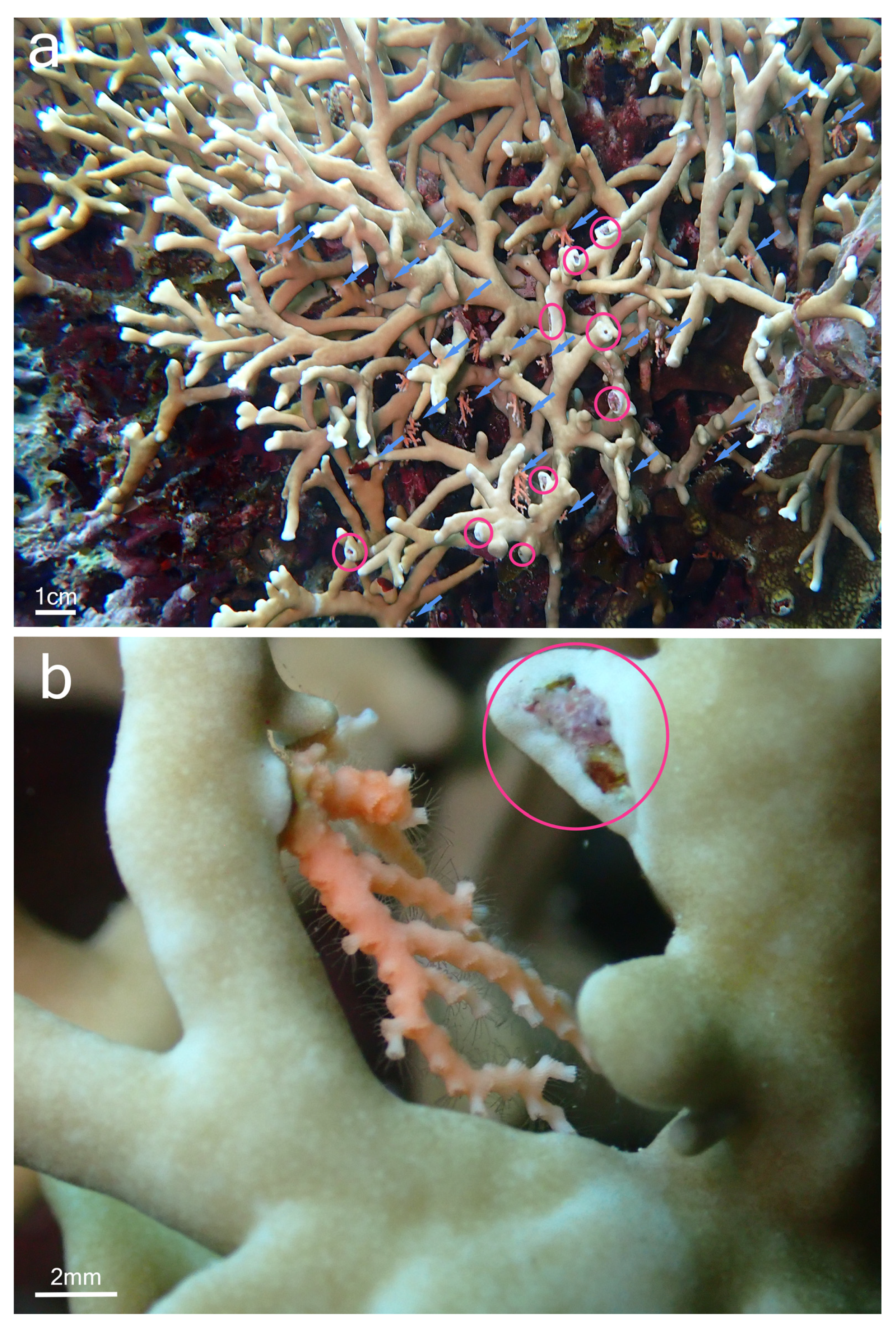{kind=link}
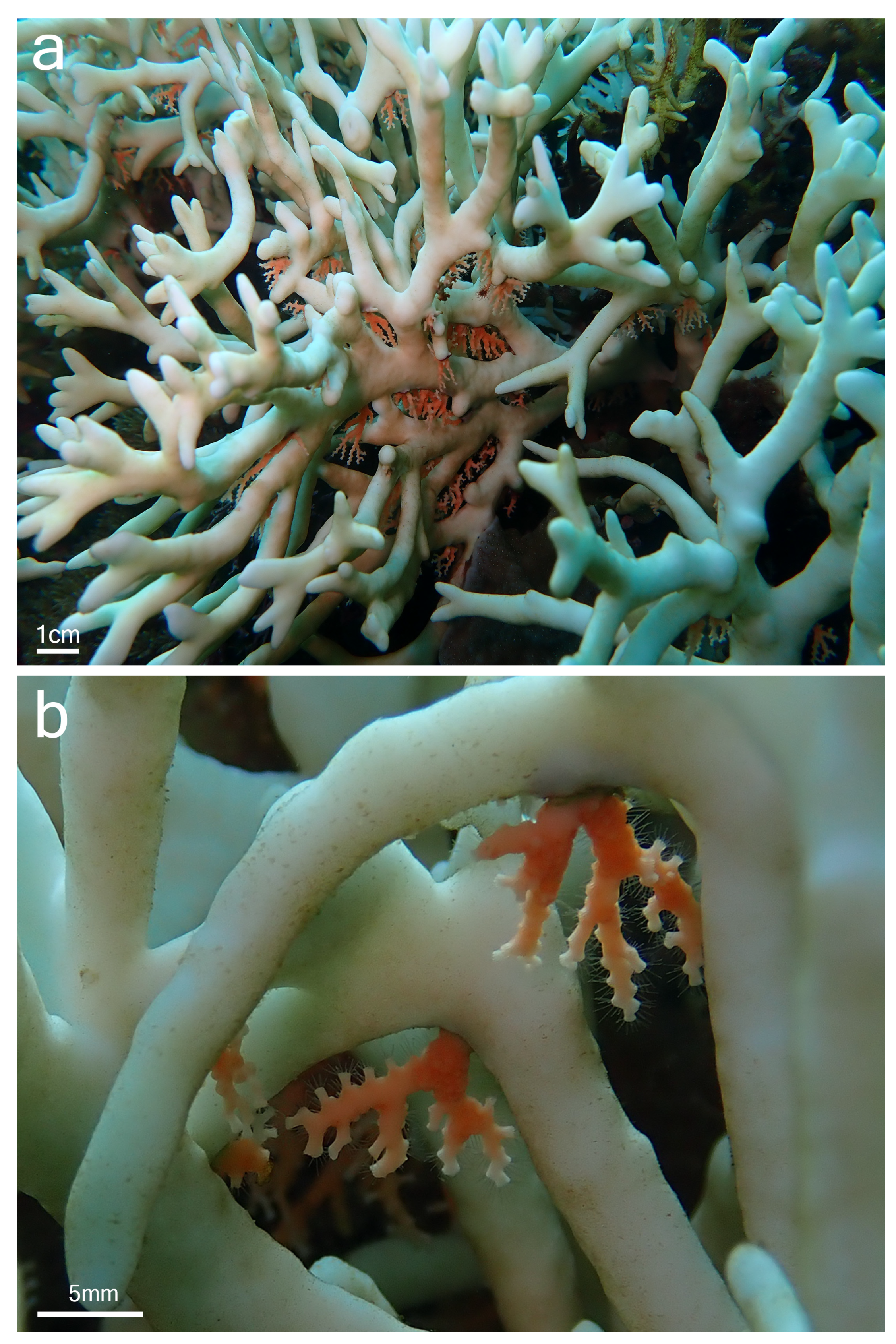{kind=link}
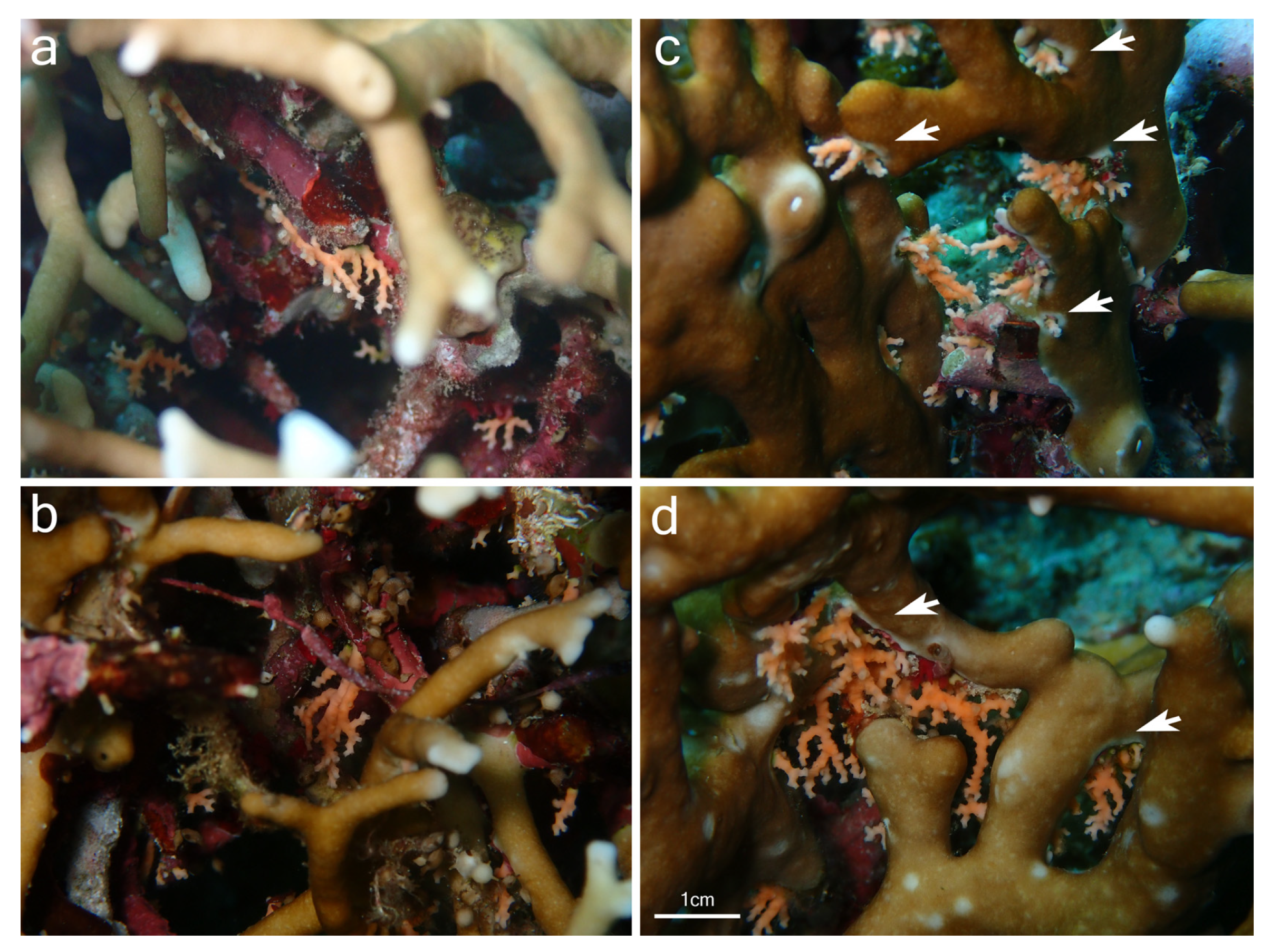{kind=link}
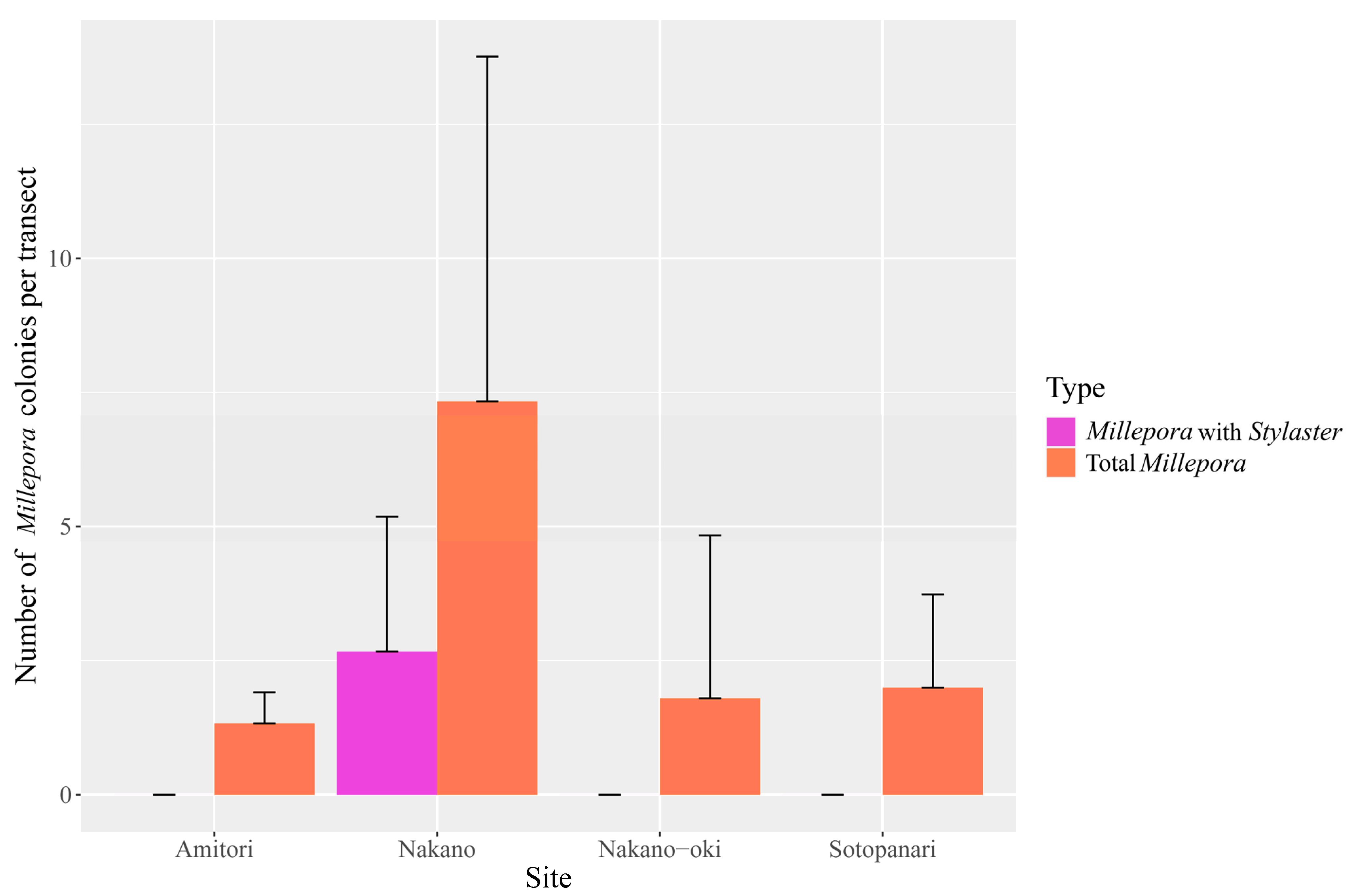{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stella, J.; Pratchett, M.; Hutchings, P.; Jones, G. Coral-associated invertebrates: Diversity, ecological importance and vulnerability to disturbance. Oceanogr. Mar. Biol. Annu. Rev. 2011, 49, 43–104. [Google Scholar]
- Garcia, T.; Matthews-Cascon, H.; Franklin-Júnior, W. Macrofauna associated with the branching fire coral Millepora alcicornis (Cnidaria: Hydrozoa). Thalassas 2009, 24, 11–19. [Google Scholar]
- Pereira, P.H.C.; Leal, I.C.S.; de Araújo, M.E.; Souza, A.T. Feeding association between reef fishes and the fire coral Millepora spp. (Cnidaria: Hydrozoa). Marine Biodiver Rec. 2012, 5, e42. [Google Scholar] [CrossRef]
- Sanchez, J.; Navas, S.G. Notes on the distribution, habitat and morphology of Stylaster roseus (Pallas, 1766) (Hydrozoa: Stylasterina) in the Colombian Caribbean. Bol. Investig. Mar. Costeras—INVEMAR 1994, 23, 193–197. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; van Moorsel, G.W.N.M. Stony corals of St. Eustatius. In Marine Biodiversity Survey of St. Eustatius; Hoeksema, B.W., Ed.; Dutch Caribbean, Naturalis Biodiversity Center: Leiden, The Netherlands, 2016; pp. 32–37. [Google Scholar]
- Montano, S.; Reimer, J.D.; Ivanenko, V.N.; García-Hernández, J.E.; van Moorsel, G.W.N.M.; Galli, P.; Hoeksema, B.W. Widespread occurrence of a rarely known association between the hydrocorals Stylaster roseus and Millepora alcicornis at Bonaire, Southern Caribbean. Diversity 2020, 12, 218. [Google Scholar] [CrossRef]
- Cairns, S.D. Global diversity of the Stylasteridae (Cnidaria: Hydrozoa: Athecatae). PLoS ONE 2011, 6, e21670. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Jimi, N.; Goto, R. Widening the host range of the ectosymbiotic scale-worm Asterophilia culcitae (Annelida: Polynoidae) to three echinoderm classes, with data on its body color variation. Plankton Benthos Res. 2020, 15, 289–295. [Google Scholar] [CrossRef]
- Hamamoto, K.; Reimer, J. Ascidians observed associating epizoically on holothurians in waters in the Kerama Islands, Okinawa, Japan. Plankton Benthos Res. 2022, 17, 338–342. [Google Scholar] [CrossRef]
- Kushida, Y.; Kunihiro, S.; Reimer, J.D. First observation of Waminoa sp. on Dendronephthya aff. rigida. Plankton Benthos Res. 2023, 18, 52–54. [Google Scholar] [CrossRef]
- Reimer, J.D.; Fourreau, C.J.L.; Fujii, T.; Ise, Y.; Kushida, Y.; Mizukami, I.; Nakano, M. Records and distribution of the coral-killing sponge Chalinula nematifera in the Ryukyu Islands, Japan. Plankton Benthos Res. 2022, 17, 249–254. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 19th February 2023).
- Yap, F.-C.; Høeg, J.T.; Chan, B.K.K. Living on fire: Deactivating fire coral polyps for larval settlement and symbiosis in the fire coral-associated barnacle Wanella milleporae (Thoracicalcarea: Wanellinae). Ecol. Evol. 2022, 12, e9057. [Google Scholar] [CrossRef] [PubMed]
- Takama, O.; Fernandez-Silva, I.; López, C.; Reimer, J.D. Molecular phylogeny demonstrates the need for taxonomic reconsideration of species diversity of the hydrocoral genus Millepora (Cnidaria: Hydrozoa) in the Pacific. Zool. Sci. 2018, 35, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Razak, T.B.; Hoeksema, B.W. The hydrocoral genus Millepora (Hydrozoa: Capitata: Milleporidae) in Indonesia. Zool. Verh. 2003, 345, 313–336. [Google Scholar]
- Dubé, C.E.; Bourmaud, C.A.F.; Mercière, A.; Planes, S.; Boissin, E. Ecology, Biology and Genetics of Millepora Hydrocorals on Coral Reefs; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Gnecco, M.; Nunes, F.L.D.; González-Zapata, F.L.; Dueñas, L.; Zilberberg, C.; Lindner, A.; Sánchez, J.A. Remarkable population structure in the tropical Atlantic lace corals Stylaster roseus (Pallas, 1766) and Stylaster blatteus (Boschma, 1961). Coral Reefs 2023, 42, 181–194. [Google Scholar] [CrossRef]
- Yu, M.C.; Gnmanee, M.; Tsao, Y.F.; Chan, B.K.K. Genetic differentiation and host usage of coral and fire coral-associated barnacles (Cirripedia: Pyrgomatinae and Wanellininae) across the Indian and Pacific Oceans. Zool. J. Linn. Soc. 2023, 199, 871–888. [Google Scholar] [CrossRef]
- Casartelli, M.; Terraneo, T.I.; Anker, A.; Vimercati, S.; Maggioni, D.; Paulay, G.; Benzoni, F. New ecological and phylogenetic insights in the boring barnacle Berndtia Utinomi, 1950 (Acrothoracica: Lithoglyptidae) reveal higher diversity, new hosts, and range extension to the Western Indian Ocean. Mar. Bioidivers. 2023, 53, 74. [Google Scholar] [CrossRef]
- Cartwright, P.; Nawrocki, A.M. Character evolution in Hydrozoa (phylum Cnidaria). Integr. Comp. Bio. 2010, 50, 456–472. [Google Scholar] [CrossRef] [PubMed]
- Lindner, A.; Cairns, S.D.; Cunningham, C.W. From offshore to onshore: Multiple origins of shallow-water corals from deep-sea ancestors. PLoS ONE 2008, 3, e2429. [Google Scholar] [CrossRef] [PubMed]
- Cairns, S.D. Worldwide distribution of the Stylasteridae (Cnidaria: Hydrozoa). Sci. Marina 1992, 56, 125–130. [Google Scholar]
- Eguchi, M. Notes on coral genera of the Yaeyama insland group, with descriptions of a new species, Cladocora kabiraensis n. sp. Proc. JPN. Soci. Syst. Zool. 1975, 11, 1–4. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fourreau, C.J.L.; Pica, D.; Jamodiong, E.A.; Mironenko Castelló, G.; Mizukami, I.; Reimer, J.D. Millepora spp. as Substrates of Their Hydrozoan Counterparts Stylaster sp. in the Pacific Ocean. Diversity 2024, 16, 142. https://doi.org/10.3390/d16030142
Fourreau CJL, Pica D, Jamodiong EA, Mironenko Castelló G, Mizukami I, Reimer JD. Millepora spp. as Substrates of Their Hydrozoan Counterparts Stylaster sp. in the Pacific Ocean. Diversity. 2024; 16(3):142. https://doi.org/10.3390/d16030142
Chicago/Turabian StyleFourreau, Chloé Julie Loïs, Daniela Pica, Emmeline A. Jamodiong, Guillermo Mironenko Castelló, Iori Mizukami, and James Davis Reimer. 2024. "Millepora spp. as Substrates of Their Hydrozoan Counterparts Stylaster sp. in the Pacific Ocean" Diversity 16, no. 3: 142. https://doi.org/10.3390/d16030142