
The Angiotensin II Type 2 Receptor, a Target for Protection and Regeneration of the Peripheral Nervous System?

 , , , ,
, , , ,
Abstract
:1. Introduction
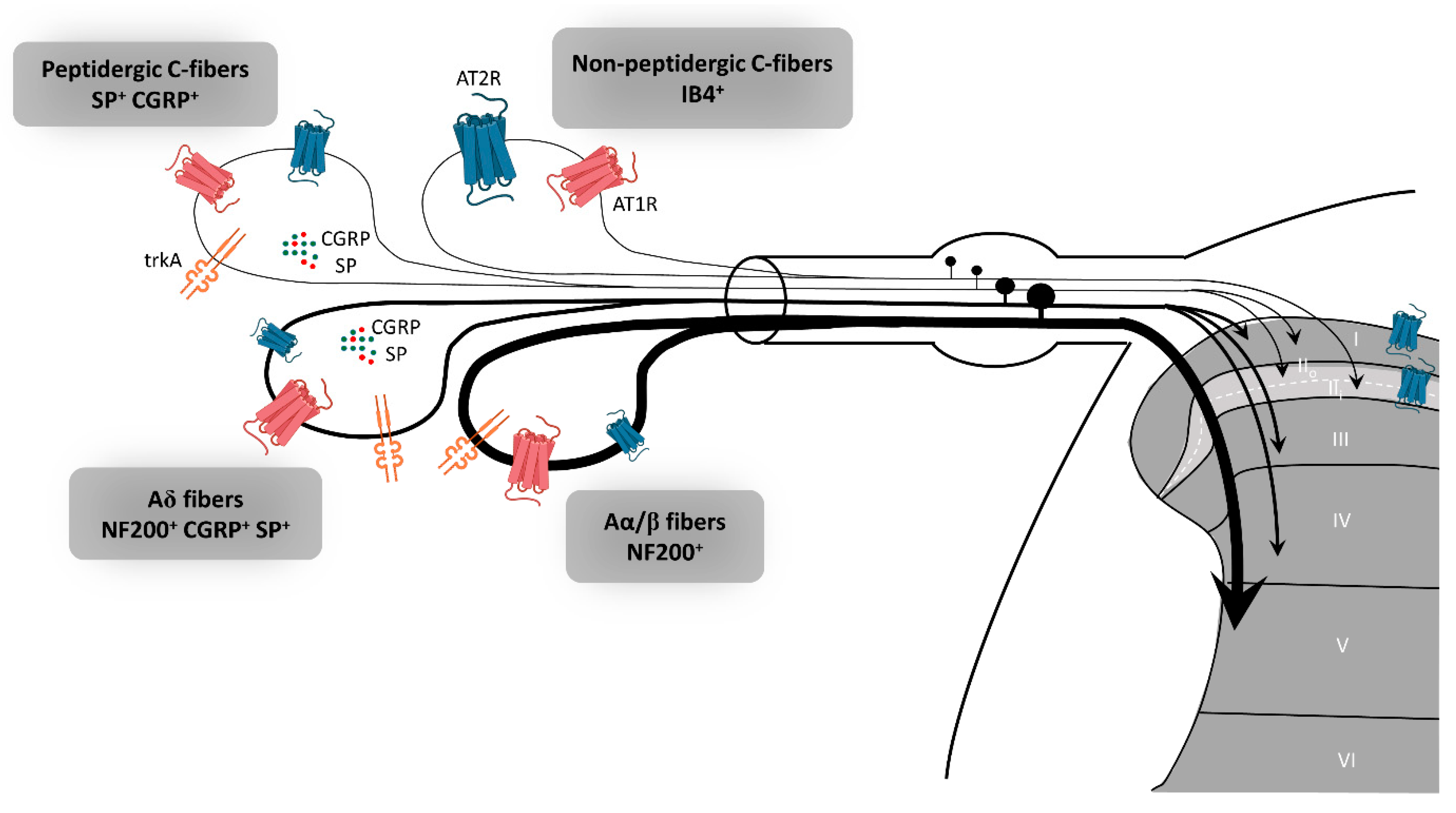
2. Evidence for AT2R Expression in the Peripheral Nervous System
3. AT2R Signaling
4. Neuroprotection
5. Neuroregeneration
5.1. In Vitro
5.2. In Vivo
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rettig, R.; Healy, D.P.; Printz, M.P. Cardiovascular Effects of Microinjections of Angiotensin II into the Nucleus Tractus Solitarii. Brain Res. 1986, 364, 233–240. [Google Scholar] [CrossRef]
- Bosnyak, S.; Jones, E.S.; Christopoulos, A.; Aguilar, M.I.; Thomas, W.G.; Widdop, R.E. Relative Affinity of Angiotensin Peptides and Novel Ligands at AT1 and AT2 Receptors. Clin. Sci. (Lond. Engl. 1979) 2011, 121, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Fei, D.T.; Coghlan, J.P.; Fernley, R.T.; Scoggins, B.A.; Tregear, G.W. Peripheral Production of Angiotensin II and III in Sheep. Circ Res 1980, 46, I135–I137. [Google Scholar] [PubMed]
- Karlsson, C.; Lindell, K.; Ottosson, M.; Carlsson, B.R.; Carlsson, L.M.S. Human Adipose Tissue Expresses Angiotensinogen and Enzymes Required for Its Conversion to Angiotensin II. J. Clin. Endocrinol. Metab. 1998, 83, 3925–3929. [Google Scholar] [CrossRef]
- van Kats, J.P.; Danser, A.H.; van Meegen, J.R.; Sassen, L.M.; Verdouw, P.D.; Schalekamp, M.A. Angiotensin Production by the Heart: A Quantitative Study in Pigs with the Use of Radiolabeled Angiotensin Infusions. Circulation 1998, 98, 73–81. [Google Scholar] [CrossRef] [Green Version]
- van Kats, J.P.; van Meegen, J.R.; Verdouw, P.D.; Duncker, D.J.; Schalekamp, M.A.; Danser, A.H. Subcellular Localization of Angiotensin II in Kidney and Adrenal. J. Hypertens. 2001, 19, 583–589. [Google Scholar] [CrossRef]
- van Thiel, B.S.; Góes Martini, A.; Te Riet, L.; Severs, D.; Uijl, E.; Garrelds, I.M.; Leijten, F.P.; van der Pluijm, I.; Essers, J.; Qadri, F.; et al. Brain Renin–Angiotensin System. Hypertension 2017, 69, 1136–1144. [Google Scholar] [CrossRef]
- Jurewicz, M.; McDermott, D.H.; Sechler, J.M.; Tinckam, K.; Takakura, A.; Carpenter, C.B.; Milford, E.; Abdi, R. Human T and Natural Killer Cells Possess a Functional Renin-Angiotensin System: Further Mechanisms of Angiotensin II-Induced Inflammation. J. Am. Soc. Nephrol. 2007, 18, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Steckelings, U.M.; Wollschläger, T.; Peters, J.; Henz, B.M.; Hermes, B.; Artuc, M. Human Skin: Source of and Target Organ for Angiotensin II. Exp. Dermatol. 2004, 13, 148–154. [Google Scholar] [CrossRef]
- Nehme, A.; Zouein, F.A.; Zayeri, Z.D.; Zibara, K. An Update on the Tissue Renin Angiotensin System and Its Role in Physiology and Pathology. JCDD 2019, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.; Poyan Mehr, A.; Kreutz, R. Physiology of Local Renin-Angiotensin Systems. Physiol. Rev. 2006, 86, 747–803. [Google Scholar] [CrossRef]
- Nouet, S.; Nahmias, C. Signal Transduction from the Angiotensin II AT2 Receptor. Trends Endocrinol. Metab. 2000, 11, 1–6. [Google Scholar] [CrossRef]
- Grady, E.F.; Sechi, L.A.; Griffin, C.A.; Schambelan, M.; Kalinyak, J.E. Expression of AT2 Receptors in the Developing Rat Fetus. J. Clin. Investig. 1991, 88, 921–933. [Google Scholar] [CrossRef] [Green Version]
- Lenkei, Z.; Palkovits, M.; Corvol, P.; Llorens-Cortès, C. Expression of Angiotensin Type-1 (AT1) and Type-2 (AT2) Receptor MRNAs in the Adult Rat Brain: A Functional Neuroanatomical Review. Front. Neuroendocrinol. 1997, 18, 383–439. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chao, J.; Parbhu, K.-J.K.; Yu, L.; Xiao, L.; Gao, F.; Gao, L. Ontogeny of Angiotensin Type 2 and Type 1 Receptor Expression in Mice. J. Renin Angiotensin Aldosterone Syst. 2012, 13, 341–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Shao, C.; Gao, L. Developmental Expression Patterns for Angiotensin Receptors in Mouse Skin and Brain. J. Renin-Angiotensin-Aldosterone Syst. JRAAS 2014, 15, 139–149. [Google Scholar] [CrossRef]
- AbdAlla, S.; Lother, H.; Abdel-tawab, A.M.; Quitterer, U. The Angiotensin II AT2 Receptor Is an AT1 Receptor Antagonist. J. Biol. Chem. 2001, 276, 39721–39726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benitez, S.G.; Seltzer, A.M.; Messina, D.N.; Foscolo, M.R.; Patterson, S.I.; Acosta, C.G. Cutaneous Inflammation Differentially Regulates the Expression and Function of Angiotensin-II Types 1 and 2 Receptors in Rat Primary Sensory Neurons. J. Neurochem. 2020, 152, 675–696. [Google Scholar] [CrossRef]
- Bessaguet, F.; Danigo, A.; Bouchenaki, H.; Duchesne, M.; Magy, L.; Richard, L.; Sturtz, F.; Desmoulière, A.; Demiot, C. Neuroprotective Effect of Angiotensin II Type 2 Receptor Stimulation in Vincristine-Induced Mechanical Allodynia. PAIN 2018, 159, 2538–2546. [Google Scholar] [CrossRef]
- Lucius, R.; Gallinat, S.; Rosenstiel, P.; Herdegen, T.; Sievers, J.; Unger, T. The Angiotensin II Type 2 (AT2) Receptor Promotes Axonal Regeneration in the Optic Nerve of Adult Rats. J. Exp. Med. 1998, 188, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Guimond, M.-O.; Gallo-Payet, N. How Does Angiotensin AT2 Receptor Activation Help Neuronal Differentiation and Improve Neuronal Pathological Situations? Front. Endocrinol. 2012, 3, 164. [Google Scholar] [CrossRef] [Green Version]
- Namsolleck, P.; Recarti, C.; Foulquier, S.; Steckelings, U.M.; Unger, T. AT(2) Receptor and Tissue Injury: Therapeutic Implications. Curr. Hypertens. Rep. 2014, 16, 416. [Google Scholar] [CrossRef] [Green Version]
- Wilms, H.; Rosenstiel, P.; Unger, T.; Deuschl, G.; Lucius, R. Neuroprotection with Angiotensin Receptor Antagonists: A Review of the Evidence and Potential Mechanisms. Am. J. Cardiovasc. Drugs 2005, 5, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, A.J.; Copits, B.A.; Mickle, A.D.; Karlsson, P.; Kadunganattil, S.; Haroutounian, S.; Tadinada, S.M.; de Kloet, A.D.; Valtcheva, M.V.; McIlvried, L.A.; et al. Angiotensin II Triggers Peripheral Macrophage-to-Sensory Neuron Redox Crosstalk to Elicit Pain. J. Neurosci. 2018, 38, 7032–7057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, A.J.; Mickle, A.D.; Golden, J.P.; Mack, M.R.; Halabi, C.M.; de Kloet, A.D.; Samineni, V.K.; Kim, B.S.; Krause, E.G.; Gereau, R.W.; et al. Macrophage Angiotensin II Type 2 Receptor Triggers Neuropathic Pain. Proc. Natl. Acad. Sci. USA 2018, 115, E8057–E8066. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.K.; O’Hara, C.L.; Stucky, C.L. Mechanical Sensitization of Cutaneous Sensory Fibers in the Spared Nerve Injury Mouse Model. Mol. Pain 2013, 9, 1744–8069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, U.; Yiangou, Y.; Sinisi, M.; Fox, M.; MacQuillan, A.; Quick, T.; Korchev, Y.E.; Bountra, C.; McCarthy, T.; Anand, P. Mechanisms Underlying Clinical Efficacy of Angiotensin II Type 2 Receptor (AT2R) Antagonist EMA401 in Neuropathic Pain: Clinical Tissue and in Vitro Studies. Mol. Pain 2015, 11, 38. [Google Scholar] [CrossRef] [Green Version]
- Rice, A.S.C.; Dworkin, R.H.; McCarthy, T.D.; Anand, P.; Bountra, C.; McCloud, P.I.; Hill, J.; Cutter, G.; Kitson, G.; Desem, N.; et al. EMA401, an Orally Administered Highly Selective Angiotensin II Type 2 Receptor Antagonist, as a Novel Treatment for Postherpetic Neuralgia: A Randomised, Double-Blind, Placebo-Controlled Phase 2 Clinical Trial. Lancet 2014, 383, 1637–1647. [Google Scholar] [CrossRef]
- Marion, E.; Song, O.R.; Christophe, T.; Babonneau, J.; Fenistein, D.; Eyer, J.; Letournel, F.; Henrion, D.; Clere, N.; Paille, V.; et al. Mycobacterial Toxin Induces Analgesia in Buruli Ulcer by Targeting the Angiotensin Pathways. Cell 2014, 157, 1565–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, O.-R.; Kim, H.-B.; Jouny, S.; Ricard, I.; Vandeputte, A.; Deboosere, N.; Marion, E.; Queval, C.J.; Lesport, P.; Bourinet, E.; et al. A Bacterial Toxin with Analgesic Properties: Hyperpolarization of DRG Neurons by Mycolactone. Toxins 2017, 9, 227. [Google Scholar] [CrossRef] [Green Version]
- Bessaguet, F.; Danigo, A.; Magy, L.; Sturtz, F.; Desmoulière, A.; Demiot, C. Candesartan Prevents Resiniferatoxin-Induced Sensory Small-Fiber Neuropathy in Mice by Promoting Angiotensin II-Mediated AT2 Receptor Stimulation. Neuropharmacology 2017, 126, 142–150. [Google Scholar] [CrossRef]
- Hashikawa-Hobara, N.; Hashikawa, N.; Inoue, Y.; Sanda, H.; Zamami, Y.; Takatori, S.; Kawasaki, H. Candesartan Cilexetil Improves Angiotensin II Type 2 Receptor-Mediated Neurite Outgrowth via the PI3K-Akt Pathway in Fructose-Induced Insulin-Resistant Rats. Diabetes 2012, 61, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Hobara, N.; Goda, M.; Yoshida, N.; Takatori, S.; Kitamura, Y.; Mio, M.; Kawasaki, H. Angiotensin II Type 2 Receptors Facilitate Reinnervation of Phenol-Lesioned Vascular Calcitonin Gene-Related Peptide-Containing Nerves in Rat Mesenteric Arteries. Neuroscience 2007, 150, 730–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinecke, K.; Lucius, R.; Reinecke, A.; Rickert, U.; Herdegen, T.; Unger, T. Angiotensin II Accelerates Functional Recovery in the Rat Sciatic Nerve in Vivo: Role of the AT2 Receptor and the Transcription Factor NF-KappaB. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 2094–2096. [Google Scholar] [CrossRef]
- Pavel, J.; Tang, H.; Brimijoin, S.; Moughamian, A.; Nishioku, T.; Benicky, J.; Saavedra, J.M. Expression and Transport of Angiotensin II AT1 Receptors in Spinal Cord, Dorsal Root Ganglia and Sciatic Nerve of the Rat. Brain Res. 2008, 1246, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Patil, J.; Schwab, A.; Nussberger, J.; Schaffner, T.; Saavedra, J.M.; Imboden, H. Intraneuronal Angiotensinergic System in Rat and Human Dorsal Root Ganglia. Regul. Pept. 2010, 162, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarty, A.; Blacklock, A.; Svojanovsky, S.; Smith, P.G. Estrogen Elicits Dorsal Root Ganglion Axon Sprouting via a Renin-Angiotensin System. Endocrinology 2008, 149, 3452–3460. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Chakrabarty, A.; Mu, Y.; Bhattacherjee, A.; Goestch, M.; Leclair, C.M.; Smith, P.G. A Local Inflammatory Renin-Angiotensin System Drives Sensory Axon Sprouting in Provoked Vestibulodynia. J. Pain 2017, 18, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Benitez, S.; Seltzer, A.; Acosta, C. Nociceptor-like Rat Dorsal Root Ganglion Neurons Express the Angiotensin-II AT2 Receptor throughout Development. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2017, 56, 10–17. [Google Scholar] [CrossRef]
- Hafko, R.; Villapol, S.; Nostramo, R.; Symes, A.; Sabban, E.L. Commercially Available Angiotensin II At 2 Receptor Antibodies Are Nonspecific. PLoS ONE 2013, 8, 69234. [Google Scholar] [CrossRef]
- Khan, N.; Muralidharan, A.; Smith, M.T. Attenuation of the Infiltration of Angiotensin II Expressing CD3+ T-Cells and the Modulation of Nerve Growth Factor in Lumbar Dorsal Root Ganglia—A Possible Mechanism Underpinning Analgesia Produced by EMA300, An Angiotensin II Type 2 (AT2) Receptor Ant. Front. Mol. Neurosci. 2017, 10. [Google Scholar] [CrossRef]
- Nio, Y.; Matsubara, H.; Murasawa, S.; Kanasaki, M.; Inada, M. Regulation of Gene Transcription of Angiotensin II Receptor Subtypes in Myocardial Infarction. J. Clin. Investig. 1995, 95, 46–54. [Google Scholar] [CrossRef]
- Yoshida, T.; Huq, T.S.; Delafontaine, P. Angiotensin Type 2 Receptor Signaling in Satellite Cells Potentiates Skeletal Muscle Regeneration. J. Biol. Chem. 2014, 289, 26239–26248. [Google Scholar] [CrossRef] [Green Version]
- Gallinat, S.; Yu, M.; Dorst, A.; Unger, T.; Herdegen, T. Sciatic Nerve Transection Evokes Lasting Up-Regulation of Angiotensin AT2 and AT1 Receptor MRNA in Adult Rat Dorsal Root Ganglia and Sciatic Nerves. Mol. Brain Res. 1998, 57, 111–122. [Google Scholar] [CrossRef]
- Bleuel, A.; de Gasparo, M.; Whitebread, S.; Püttner, I.; Monard, D. Regulation of Protease Nexin-1 Expression in Cultured Schwann Cells Is Mediated by Angiotensin II Receptors. J. Neurosci. 1995, 15, 750–761. [Google Scholar] [CrossRef]
- Hu, P.; Bembrick, A.L.; Keay, K.A.; McLachlan, E.M. Immune Cell Involvement in Dorsal Root Ganglia and Spinal Cord after Chronic Constriction or Transection of the Rat Sciatic Nerve. Brain Behav. Immun. 2007, 21, 599–616. [Google Scholar] [CrossRef]
- Tsutsumi, K.; Saavedra, J.M. Heterogeneity of Angiotensin II AT2 Receptors in the Rat Brain. Mol. Pharmacol. 1992, 41, 290–297. [Google Scholar]
- Hein, L.; Barsh, G.S.; Pratt, R.E.; Dzau, V.J.; Kobilka, B.K. Behavioural and Cardiovascular Effects of Disrupting the Angiotensin II Type-2 Receptor in Mice. Nature 1995, 377, 744–747. [Google Scholar] [CrossRef]
- Ichiki, T.; Labosky, P.A.; Shiota, C.; Okuyama, S.; Imagawa, Y.; Fogo, A.; Niimura, F.; Ichikawa, I.; Hogan, B.L.; Inagami, T. Effects on Blood Pressure and Exploratory Behaviour of Mice Lacking Angiotensin II Type-2 Receptor. Nature 1995, 377, 748–750. [Google Scholar] [CrossRef]
- Wong, H.H.W.; Lin, J.Q.; Ströhl, F.; Roque, C.G.; Cioni, J.M.; Cagnetta, R.; Turner-Bridger, B.; Laine, R.F.; Harris, W.A.; Kaminski, C.F.; et al. RNA Docking and Local Translation Regulate Site-Specific Axon Remodeling In Vivo. Neuron 2017, 95, 852–868.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, M.; Mukoyama, M.; Pratt, R.E.; Horiuchi, M.; Dzau, V.J. Cloning of CDNA and Analysis of the Gene for Mouse Angiotensin II Type 2 Receptor. Biochem. Biophys. Res. Commun. 1993, 197, 393–399. [Google Scholar] [CrossRef]
- Mukoyama, M.; Nakajima, M.; Horiuchi, M.; Sasamura, H.; Pratt, R.E.; Dzau, V.J. Expression Cloning of Type 2 Angiotensin II Receptor Reveals a Unique Class of Seven-Transmembrane Receptors. J. Biol. Chem. 1993, 268, 24539–24542. [Google Scholar] [CrossRef]
- Asada, H.; Horita, S.; Hirata, K.; Shiroishi, M.; Shiimura, Y.; Iwanari, H.; Hamakubo, T.; Shimamura, T.; Nomura, N.; Kusano-Arai, O.; et al. Crystal Structure of the Human Angiotensin II Type 2 Receptor Bound to an Angiotensin II Analog. Nat. Struct. Mol. Biol. 2018, 25, 570–576. [Google Scholar] [CrossRef]
- Asada, H.; Inoue, A.; Ngako Kadji, F.M.; Hirata, K.; Shiimura, Y.; Im, D.; Shimamura, T.; Nomura, N.; Iwanari, H.; Hamakubo, T.; et al. The Crystal Structure of Angiotensin II Type 2 Receptor with Endogenous Peptide Hormone. Structure 2020, 28, 418–425.e4. [Google Scholar] [CrossRef]
- Sadybekov, A.; Katritch, V. Breaking the Enigma Code of Angiotensin II Type 2 Receptor Signaling. Structure 2020, 28, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Horiuchi, M.; Dzau, V.J. Angiotensin II Type 2 Receptor Mediates Programmed Cell Death. Proc. Natl. Acad. Sci. USA 1996, 93, 156–160. [Google Scholar] [CrossRef] [Green Version]
- Horiuchi, M.; Hayashida, W.; Kambe, T.; Yamada, T.; Dzau, V.J. Angiotensin Type 2 Receptor Dephosphorylates Bcl-2 by Activating Mitogen-Activated Protein Kinase Phosphatase-1 and Induces Apoptosis. J. Biol. Chem. 1997, 272, 19022–19026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.; Wallinder, C.; Plouffe, B.; Beaudry, H.; Mahalingam, A.K.; Wu, X.; Johansson, B.; Holm, M.; Botoros, M.; Karlén, A.; et al. Design, Synthesis, and Biological Evaluation, of the First Selective Nonpeptide AT2 Receptor Agonist. J. Med. Chem. 2004, 47, 5995–6008. [Google Scholar] [CrossRef]
- Namsolleck, P.; Boato, F.; Schwengel, K.; Paulis, L.; Matho, K.S.; Geurts, N.; Thöne-Reineke, C.; Lucht, K.; Seidel, K.; Hallberg, A.; et al. AT2-Receptor Stimulation Enhances Axonal Plasticity after Spinal Cord Injury by Upregulating BDNF Expression. Neurobiol. Dis. 2013, 51, 177–191. [Google Scholar] [CrossRef]
- Stroth, U.; Meffert, S.; Gallinat, S.; Unger, T. Angiotensin II and NGF Differentially Influence Microtubule Proteins in PC12W Cells: Role of the AT2 Receptor. Mol. Brain Res. 1998, 53, 187–195. [Google Scholar] [CrossRef]
- Laflamme, L.; de Gasparo, M.; Gallo, J.-M.; Payet, M.D.; Gallo-Payet, N. Angiotensin II Induction of Neurite Outgrowth by AT 2 Receptors in NG108-15 Cells. J. Biol. Chem. 1996, 271, 22729–22735. [Google Scholar] [CrossRef] [Green Version]
- Bouquet, C.; Soares, S.; von Boxberg, Y.; Ravaille-Veron, M.; Propst, F.; Nothias, F. Microtubule-Associated Protein 1B Controls Directionality of Growth Cone Migration and Axonal Branching in Regeneration of Adult Dorsal Root Ganglia Neurons. J. Neurosci. 2004, 24, 7204–7213. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Ji, F.; Liu, L.; Li, F. Expression Changes of Parvalbumin and Microtubule- Associated Protein 2 Induced by Chronic Constriction Injury in Rat Dorsal Root Ganglia. Chin. Med. J. 2011, 124, 2184–2190. [Google Scholar] [CrossRef] [PubMed]
- Gallinat, S.; Csikos, T.; Meffert, S.; Herdegen, T.; Stoll, M.; Unger, T. The Angiotensin AT2 Receptor Down-Regulates Neurofilament M in PC12W Cells. Neurosci. Lett. 1997, 227, 29–32. [Google Scholar] [CrossRef]
- Gendron, L.; Laflamme, L.; Rivard, N.; Asselin, C.; Payet, M.D.; Gallo-Payet, N. Signals from the AT2 (Angiotensin Type 2) Receptor of Angiotensin II Inhibit P21ras and Activate MAPK (Mitogen-Activated Protein Kinase) to Induce Morphological Neuronal Differentiation in NG108-15 Cells. Mol. Endocrinol. 1999, 13, 1615–1626. [Google Scholar] [CrossRef] [Green Version]
- Plouffe, B.; Guimond, M.O.; Beaudry, H.; Gallo-Payet, N. Role of Tyrosine Kinase Receptors in Angiotensin II AT2 Receptor Signaling: Involvement in Neurite Outgrowth and in P42/P44mapk Activation in NG108-15 Cells. Endocrinology 2006, 147, 4646–4654. [Google Scholar] [CrossRef] [Green Version]
- Anand, U.; Facer, P.; Yiangou, Y.; Sinisi, M.; Fox, M.; McCarthy, T.; Bountra, C.; Korchev, Y.E.; Anand, P. Angiotensin II Type 2 Receptor (AT2R) Localization and Antagonist-Mediated Inhibition of Capsaicin Responses and Neurite Outgrowth in Human and Rat Sensory Neurons. Eur. J. Pain 2013, 17, 1012–1026. [Google Scholar] [CrossRef] [Green Version]
- Côté, F.; Laflamme, L.; Payet, M.D.; Gallo-Payet, N. Nitric Oxide, a New Second Messenger Involved in the Action of Angiotensin II on Neuronal Differentiation of NG108-15 Cells. Endocr. Res. 1998, 24, 403–407. [Google Scholar] [CrossRef]
- Thippeswamy, T.; Morris, R. Nerve Growth Factor Inhibits the Expression of Nitric Oxide Synthase in Neurones in Dissociated Cultures of Rat Dorsal Root Ganglia. Neurosci. Lett. 1997, 230, 9–12. [Google Scholar] [CrossRef]
- Gendron, L.; Côté, F.; Payet, M.D.; Gallo-Payet, N. Nitric Oxide and Cyclic GMP Are Involved in Angiotensin II AT(2) Receptor Effects on Neurite Outgrowth in NG108-15 Cells. Neuroendocrinology 2002, 75, 70–81. [Google Scholar] [CrossRef]
- Thippeswamy, T.; Jain, R.K.; Mumtaz, N.; Morris, R. Inhibition of Neuronal Nitric Oxide Synthase Results in Neurodegenerative Changes in the Axotomised Dorsal Root Ganglion Neurons: Evidence for a Neuroprotective Role of Nitric Oxide in Vivo. Neurosci Res. 2001, 40, 37–44. [Google Scholar] [CrossRef]
- Contestabile, A.; Ciani, E. Role of Nitric Oxide in the Regulation of Neuronal Proliferation, Survival and Differentiation. Neurochem. Int. 2004, 45, 903–914. [Google Scholar] [CrossRef]
- Abadir, P.M.; Foster, D.B.; Crow, M.; Cooke, C.A.; Rucker, J.J.; Jain, A.; Smith, B.J.; Burks, T.N.; Cohn, R.D.; Fedarko, N.S.; et al. Identification and Characterization of a Functional Mitochondrial Angiotensin System. Cell Biol. 2011, 108, 6. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, R.; Costa-Besada, M.A.; Iglesias-Gonzalez, J.; Perez-Costas, E.; Villar-Cheda, B.; Garrido-Gil, P.; Melendez-Ferro, M.; Soto-Otero, R.; Lanciego, J.L.; Henrion, D.; et al. Mitochondrial Angiotensin Receptors in Dopaminergic Neurons. Role in Cell Protection and Aging-Related Vulnerability to Neurodegeneration. Cell Death Dis 2016, 7, e2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.-M.; Mogi, M.; Tsukuda, K.; Tomochika, H.; Iwanami, J.; Min, L.-J.; Nahmias, C.; Iwai, M.; Horiuchi, M. Angiotensin II-Induced Neural Differentiation via Angiotensin II Type 2 (AT 2 ) Receptor-MMS2 Cascade Involving Interaction between AT 2 Receptor-Interacting Protein and Src Homology 2 Domain-Containing Protein-Tyrosine Phosphatase 1. Mol. Endocrinol. 2007, 21, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Benedetto, M.; Bièche, I.; Deshayes, F.; Vacher, S.; Nouet, S.; Collura, V.; Seitz, I.; Louis, S.; Pineau, P.; Amsellem-Ouazana, D.; et al. Structural Organization and Expression of Human MTUS1, a Candidate 8p22 Tumor Suppressor Gene Encoding a Family of Angiotensin II AT2 Receptor-Interacting Proteins, ATIP. Gene 2006, 380, 127–136. [Google Scholar] [CrossRef]
- Nouet, S.; Amzallag, N.; Li, J.-M.; Louis, S.; Seitz, I.; Cui, T.-X.; Alleaume, A.-M.; Di Benedetto, M.; Boden, C.; Masson, M.; et al. Trans-Inactivation of Receptor Tyrosine Kinases by Novel Angiotensin II AT2 Receptor-Interacting Protein, ATIP. J. Biol. Chem. 2004, 279, 28989–28997. [Google Scholar] [CrossRef] [Green Version]
- Wruck, C.J.; Funke-Kaiser, H.; Pufe, T.; Kusserow, H.; Menk, M.; Schefe, J.H.; Kruse, M.L.; Stoll, M.; Unger, T. Regulation of Transport of the Angiotensin AT2 Receptor by a Novel Membrane-Associated Golgi Protein. Arter. Thromb. Vasc. Biol. 2005, 25, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrester, S.J.; Booz, G.W.; Sigmund, C.D.; Coffman, T.M.; Kawai, T.; Rizzo, V.; Scalia, R.; Eguchi, S. Angiotensin II Signal Transduction: An Update on Mechanisms of Physiology and Pathophysiology. Physiol Rev. 2018, 98, 1627–1738. [Google Scholar] [CrossRef] [PubMed]
- Porrello, E.R.; Delbridge, L.M.D.; Thomas, W.G. The Angiotensin II Type 2 (AT2) Receptor: An Enigmatic Seven Transmembrane Receptor. Front. Biosci. (Landmark Ed.) 2009, 14, 958–972. [Google Scholar] [CrossRef] [Green Version]
- Sumners, C.; Fleegal, M.A.; Zhu, M. Angiotensin AT1 Receptor Signalling Pathways in Neurons. Clin. Exp. Pharmacol. Physiol. 2002, 29, 483–490. [Google Scholar] [CrossRef]
- Fujita, T.; Hirooka, K.; Nakamura, T.; Itano, T.; Nishiyama, A.; Nagai, Y.; Shiraga, F. Neuroprotective Effects of Angiotensin II Type 1 Receptor (AT1-R) Blocker via Modulating AT1-R Signaling and Decreased Extracellular Glutamate Levels. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4099–4110. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Zhu, Y.-Z.; Wong, P.T.-H. Neuroprotective Effects of Candesartan against Cerebral Ischemia in Spontaneously Hypertensive Rats. Neuroreport 2005, 16, 1963–1967. [Google Scholar] [CrossRef] [PubMed]
- Iwai, M.; Liu, H.-W.; Chen, R.; Ide, A.; Okamoto, S.; Hata, R.; Sakanaka, M.; Shiuchi, T.; Horiuchi, M. Possible Inhibition of Focal Cerebral Ischemia by Angiotensin II Type 2 Receptor Stimulation. Circulation 2004, 110, 843–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.-Y.; Yin, L. Neuroprotective Effect of Angiotensin II Type 2 Receptor during Cerebral Ischemia/Reperfusion. Neural Regen Res. 2016, 11, 1102–1107. [Google Scholar] [CrossRef]
- Mateos, L.; Perez-Alvarez, M.J.; Wandosell, F. Angiotensin II Type-2 Receptor Stimulation Induces Neuronal VEGF Synthesis after Cerebral Ischemia. Biochim Biophys Acta 2016, 1862, 1297–1308. [Google Scholar] [CrossRef]
- Schwengel, K.; Namsolleck, P.; Lucht, K.; Clausen, B.H.; Lambertsen, K.L.; Valero-esquitino, V.; Thöne-reineke, C.; Müller, S.; Widdop, R.E.; Denton, K.M.; et al. Angiotensin AT2-Receptor Stimulation Improves Survival and Neurological Outcome after Experimental Stroke in Mice. J. Mol. Med. 2016, 94, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Matavelli, L.C.; Siragy, H.M. AT2 Receptor Activities and Pathophysiological Implications. J. Cardiovasc. Pharm. 2015, 65, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Sumners, C.; Horiuchi, M.; Widdop, R.E.; McCarthy, C.; Unger, T.; Steckelings, U.M. Protective Arms of the Renin-Angiotensin-System in Neurological Disease. Clin. Exp. Pharmacol. Physiol. 2013, 40, 580–588. [Google Scholar] [CrossRef]
- Guimond, M.-O.; Wallinder, C.; Alterman, M.; Hallberg, A.; Gallo-Payet, N. Comparative Functional Properties of Two Structurally Similar Selective Nonpeptide Drug-like Ligands for the Angiotensin II Type-2 (AT(2)) Receptor. Effects on Neurite Outgrowth in NG108-15 Cells. Eur J. Pharm. 2013, 699, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Meffert, S.; Stolf, M.; Steckelings, U.M.; Bottari, S.P.; Unger, T. The Angiotensin II AT2 Receptor Inhibits Proliferation and Promotes Differentiation in PC12W Cells. Mol. Cell. Endocrinol. 1996, 122, 59–67. [Google Scholar] [CrossRef]
- Hashikawa-Hobara, N.; Hashikawa, N. Angiotensin II AT2receptors Regulate NGF-Mediated Neurite Outgrowth via the NO-CGMP Pathway. Biochem. Biophys. Res. Commun. 2016, 478, 970–975. [Google Scholar] [CrossRef]
- Chakrabarty, A.; Liao, Z.; Smith, P.G. Angiotensin II Receptor Type 2 Activation Is Required for Cutaneous Sensory Hyperinnervation and Hypersensitivity in a Rat Hind Paw Model of Inflammatory Pain. J. Pain 2013, 14, 1053–1065. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarty, A.; Liao, Z.; Mu, Y.; Smith, P.G. Inflammatory Renin-Angiotensin System Disruption Attenuates Sensory Hyperinnervation and Mechanical Hypersensitivity in a Rat Model of Provoked Vestibulodynia. J. Pain 2018, 19, 264–277. [Google Scholar] [CrossRef]
- Bohm-Starke, N.; Hilliges, M.; Falconer, C.; Rylander, E. Increased Intraepithelial Innervation in Women with Vulvar Vestibulitis Syndrome. GOI 1998, 46, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Bohm-Starke, N.; Hilliges, M.; Falconer, C.; Rylander, E. Neurochemical Characterization of the Vestibular Nerves in Women with Vulvar Vestibulitis Syndrome. Gynecol. Obstet. Investig. 1999, 48, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Chadha, S.; Gianotten, W.L.; Drogendijk, A.C.; Weijmar Schultz, W.C.; Blindeman, L.A.; van der Meijden, W.I. Histopathologic Features of Vulvar Vestibulitis. Int. J. Gynecol. Pathol. 1998, 17, 7–11. [Google Scholar] [CrossRef]
- Pulakat, L.; Sumners, C. Angiotensin Type 2 Receptors: Painful, or Not? Front. Pharmacol. 2020, 11, 571994. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Model | Species | Effect of Neuropathy | Treatment | Administration | Effect of Treatment | Antagonism | AT2R Expression | Ref. |
|---|---|---|---|---|---|---|---|---|
| Topical phenol treatment | Wistar rat (6 week-old) | Decrease of CGRP nerve fibers in mesenteric arteries | Losartan + Ang II | Osmotic pump | Increase of CGRP nerve fibers | Effect suppressed by PD123319 | DRG neuron (WB) | [33] |
| RTX-induced sensory neuropathy | Male Swiss mouse (25–30 g) | Depletion of CGRP and SP in IENFs and in DRG neurons | Candesartan | I.P.. | Improvement of the neuropeptide depletion | Effect counteracted by PD123319 and in AT2R-KO mice | / | [31] |
| Vincristine-induced neuropathy | Male Swiss mouse (25–30 g) | Nonpeptidergic IENFs loss, loss of myelinated fibers and enlargement of axonal diameter in sciatic nerve | Candesartan and C21 | I.P. | Prevention of IENF loss by candesartan and C21. Prevention of myelinated nerve fiber impairment by C21 | Effects counteracted by PD123319 | / | [19] |
| Sciatic nerve crush | Male Sprague-Dawley rats (200–250 g) | Axonal degeneration and demyelination | Ang II | Osmotic pump | Enhancement of axonal regeneration and myelination | Effect lost with PD123319 | SCs | [34] |
| Hind paw inflammation (CFA injection) | Female Sprague-Dawley rats (190–200 g) | Increase of PGP9.5+ nerve fibers in dermis and epidermis. Increase of CGRP+ nerve fibers in dermis | PD123319 | Osmotic pump | Decrease of CFA-induced hyper-innervation in dermis and epidermis. | / | / | [93] |
| Rat DRG cells | Adult female Wistar rat | / | Ang II | / | Ang II increased neurite length and density | Effects counteracted by EMA401 | DRG neurons (ICC, IHC) | [67] |
| Avulsed Human cervical DRG cells | From surgical nerve repair procedure | / | EMA401 | / | Mean neurite density was reduced by EMA401 | / | DRG cells | [67] |
| Adult mouse DRG cells | Male C57BL6J mice (25–30 g) | / | NGF + PD123319 or AT2R siRNA | / | AT2R inhibition is associated with decrease of NGF-induced neurite outgrowth | / | DRG cells | [92] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danigo, A.; Rovini, A.; Bessaguet, F.; Bouchenaki, H.; Bernard, A.; Sturtz, F.; Bourthoumieu, S.; Desmoulière, A.; Magy, L.; Demiot, C. The Angiotensin II Type 2 Receptor, a Target for Protection and Regeneration of the Peripheral Nervous System? Pharmaceuticals 2021, 14, 175. https://doi.org/10.3390/ph14030175
Danigo A, Rovini A, Bessaguet F, Bouchenaki H, Bernard A, Sturtz F, Bourthoumieu S, Desmoulière A, Magy L, Demiot C. The Angiotensin II Type 2 Receptor, a Target for Protection and Regeneration of the Peripheral Nervous System? Pharmaceuticals. 2021; 14(3):175. https://doi.org/10.3390/ph14030175
Chicago/Turabian StyleDanigo, Aurore, Amandine Rovini, Flavien Bessaguet, Hichem Bouchenaki, Amandine Bernard, Franck Sturtz, Sylvie Bourthoumieu, Alexis Desmoulière, Laurent Magy, and Claire Demiot. 2021. "The Angiotensin II Type 2 Receptor, a Target for Protection and Regeneration of the Peripheral Nervous System?" Pharmaceuticals 14, no. 3: 175. https://doi.org/10.3390/ph14030175