Natural Products of Marine Macroalgae from South Eastern Australia, with Emphasis on the Port Phillip Bay and Heads Regions of Victoria



Abstract
:
1. Introduction
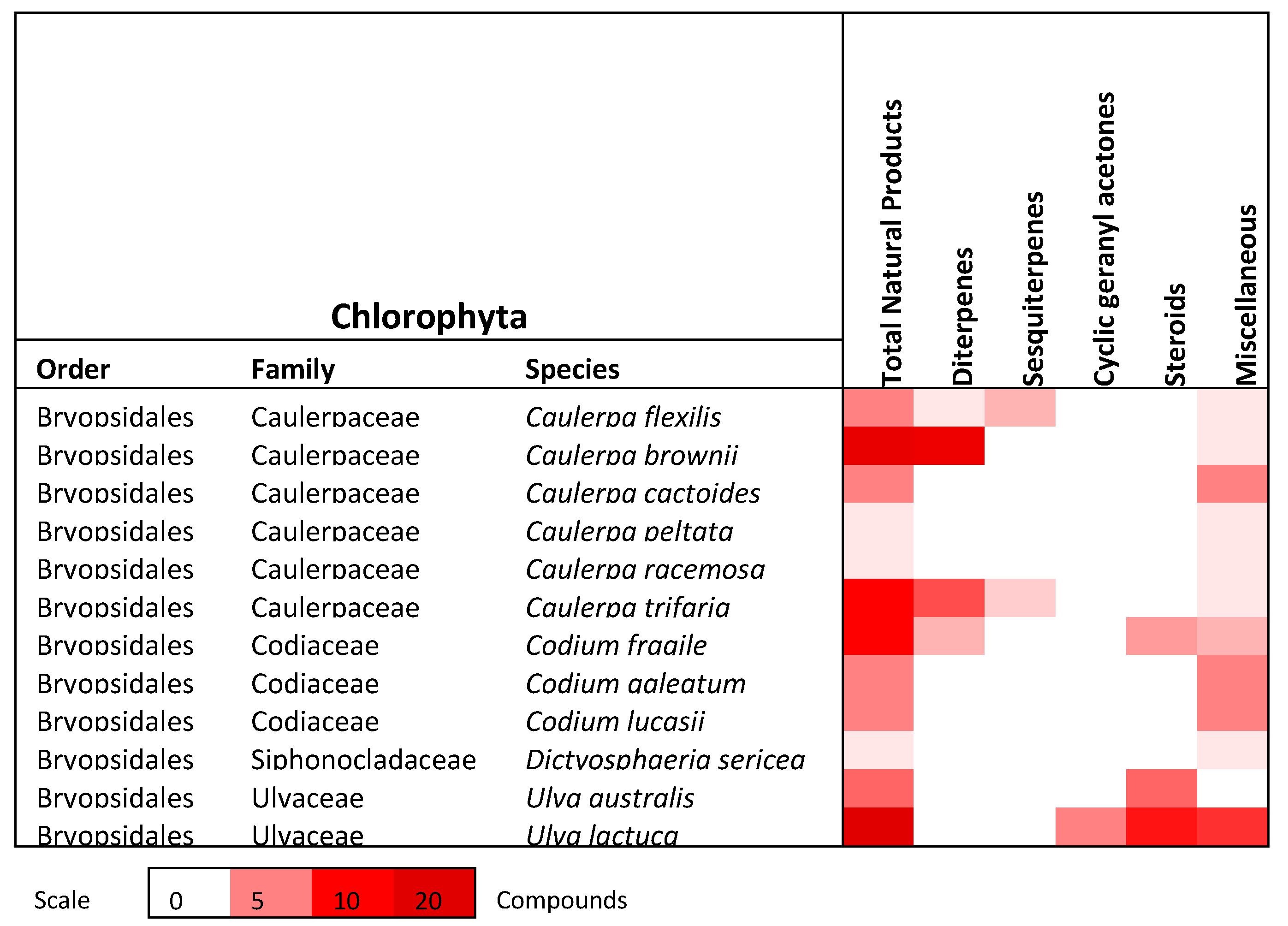
2. Chlorophyta (Green Algae)
2.1. Terpenoids
2.1.1. Diterpenes
2.1.2. Sesquiterpenes
2.1.3. Cyclic geranylacetone
2.2. Steroids/Sterols
2.3. Miscellaneous
3. Ochrophyta (Brown Algae)
3.1. Terpenoids
3.1.1. Tocotrienols
3.1.2. Monoterpenes
3.1.3. Prenylated Phenols
3.1.4. Meroditerpenoids
3.1.5. Sesquiterpenes and Monoterpenes
3.1.6. Diterpenoids
3.2. Steroids/Sterols
3.3. Lipids
3.3.1. Polyenes
3.3.2. Oxy/Epoxy lipids
3.4. Phenols
3.4.1. Phloroglucinols
3.4.2. Phenols/Phenolic acids/Resorcinols
3.5. Miscellaneous
4. Rhodophyta (Red Algae) of Port Phillip Bay
4.1. Terpenoids
4.1.1. Halogenated Monoterpenes
4.1.2. Parguerenes
4.1.3. Chamigrenes
4.1.4. Laurenes
4.1.5. Sesquiterpenes
4.1.6. Lauroxocanes (C15 acetogenins)
4.1.7. Polyhalogenated Indoles
4.1.8. Polyhalogenated Hydrocarbons
4.1.9. Halogenated Furanones
4.2. Steroids
4.3. Miscellaneous
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2017, 34, 235–294. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Hu, B.; Wang, J.; Zhu, F.; Kang, Y.; Li, D.; Sun, H.; Kong, D.X.; Hou, T. Cheminformatic Insight into the Differences between Terrestrial and Marine Originated Natural Products. J. Chem. Inf. Model. 2018, 58, 1182–1193. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M. Marine pharmacology in 2009-2011: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar]
- Cheung, R.C.; Wong, J.H.; Pan, W.L.; Chan, Y.S.; Yin, C.M.; Dan, X.L.; Wang, H.X.; Fang, E.F.; Lam, S.K.; Ngai, P.H.; et al. Antifungal and antiviral products of marine organisms. Appl. Microbiol. Biotechnol. 2014, 98, 3475–3494. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Control. 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Zhou, Z.-F.; Guo, Y.-W. Bioactive natural products from Chinese marine flora and fauna. Acta Pharmacol. Sin. 2012, 33, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.-F.; Menna, M.; Cai, Y.-S.; Guo, Y.-W. Polyacetylenes of Marine Origin: Chemistry and Bioactivity. Chem. Rev. 2014, 115, 1543–1596. [Google Scholar] [CrossRef]
- Scieszka, S.; Klewicka, E. Algae in food: A general review. Crit Rev. Food Sci Nutr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bouissil, S.; Pierre, G.; Alaoui-Talibi, Z.E.; Michaud, P.; El Modafar, C.; Dalattre, C. Applications of Algal Polysaccharides and Derivatives in Therapeutic and Agricultural Fields. Curr. Pharm. 2019, 25, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sust. Energ. Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Perez, M.J.; Falque, E.; Dominguez, H. Antimicrobial Action of Compounds from Marine Seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.; Botana, L.M.; Pedrosa, R. From Marine origin to therapeutics: The antitumor potential of marine algae-derived compounds. Front. Pharmacol. 2018, 9, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Tchokouaha Yamthe, L.R.; Appiah-Opong, R.; Tsouh Fokou, P.V.; Tsabang, N.; Fekam Boyom, F.; Kwadwo Nyarko, A.; Wilson, M. Marine algae as a source of novel antileishmanial drugs: A review. Mar. Drugs 2017, 15, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, I.S.; Nah, J.W.; Jeon, Y.J. Potential anti-inflammatory natural products from marine algae. Environ. Toxicol. Pharmacol. 2016, 48, 22–30. [Google Scholar] [CrossRef]
- Dahms, H.U.; Dobretsov, S. Antifouling compounds from marine macroalgae. Mar. Drugs 2017, 15, 265. [Google Scholar] [CrossRef]
- Torres, F.A.E.; Passalacqua, T.G.; Velasquez, A.M.A.; de Souza, R.A.; Colepicolo, P.; Graminha, M.A.S. New drugs with antiprotozoal activity from marine algae: A review. Rev. Bras. Farmacogn. 2014, 24, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Leal, M.C.; Munro, M.H.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef]
- Womersley, H.B.S. Port Phillip Survey 1957–1963. Mem. Nat. Mus. Vict. 1966, 27, 133–153. [Google Scholar] [CrossRef]
- Edgar, G. Australian Marine Life: The Plants and Animals of Temperate Waters; Reed New Holland: Sydney, Australia, 2000. [Google Scholar]
- VictorianStateGovernment. Victorian Biodiversity Atlas. Available online: https://www.environment.vic.gov.au/biodiversity/victorian-biodiversity-atlas (accessed on 10 October 2018).
- Paul, V.J.; Fenical, W. Diterpenoid metabolites from the pacific marine algae of the order Caulerpales (Chlorophyta). Phytochemistry 1985, 24, 2239–2243. [Google Scholar] [CrossRef]
- Handley, J.T.; Blackman, A.J. Monocyclic Diterpenes from the Marine Alga Caulerpa trifaria (Chlorophyta). Aust. J. Chem. 2000, 53, 67–71. [Google Scholar] [CrossRef]
- Masaki, Y.; Hashimoto, K.; Iwai, H.; Kaji, K. Synthesis of optically active natural caulerpol. Chem. Lett. 1978, 1203–1204. [Google Scholar] [CrossRef]
- Handley, J.T.; Blackman, A.J. Secondary Metabolites from the Marine Alga Caulerpa brownii (Chlorophyta). Aust. J. Chem. 2005, 58, 39–46. [Google Scholar] [CrossRef]
- Yin, S.-W.; Wang, C.-Y.; Li, X.-M.; Wang, B.-G. A new clerosterol derivative, trans-phytol, and related metabolites from marine green alga Codium fragile (Codiaceae) and their chemotaxonomic significance. Biochem. Syst. Ecol. 2005, 33, 1288–1292. [Google Scholar] [CrossRef]
- Sun, H.H.; Fenical, W. Rhipocephalin and rhipocephenal; Toxic feeding deterrents from the tropical marine alga Rhipocephalus Phoenix. Tetrahedron Lett. 1979, 8, 685–688. [Google Scholar] [CrossRef]
- Maric, P.; Ahel, M.; Senta, I.; Terzic, S.; Mikac, I.; Zuljevic, A.; Smital, T. Effect-directed analysis reveals inhibition of zebrafish uptake transporter Oatp1d1 by caulerpenyne, a major secondary metabolite from the invasive marine alga. Caulerpa Taxifolia Chemosphere 2017, 174, 643–654. [Google Scholar] [CrossRef] [Green Version]
- Paul, V.J.; Fenical, W. Bioactive terpenoids from caribbean marine algae of the genera penicillus and udotea (Chlorophyta). Tetrahedron 1984, 40, 2913–2918. [Google Scholar] [CrossRef]
- D’Abrosca, B.; DellaGreca, M.; Fiorentino, A.; Monaco, P.; Oriano, P.; Temussi, F. Structure elucidation and phytotoxicity of C13 nor-isoprenoids from Cestrum Parqui. Phytochemistry 2004, 65, 497–505. [Google Scholar] [CrossRef]
- DellaGreca, M.; Marino, C.D.; Zarrelli, A.; D’Abrosca, B. Isolation and Phytotoxicity of Apocarotenoids from Chenopodium album. J. Nat. Prod. 2004, 67, 1492–1495. [Google Scholar] [CrossRef] [PubMed]
- Dollmann, G.F.; Schreier, P.; Winterhalter, P.; Guntert, M.; Sommer, H. Synthesis and Enantiodifferentiation of Riesling Acetals. Phytochem. Anal. 1995, 6, 106–111. [Google Scholar] [CrossRef]
- Strauss, C.R.; Dimitriadis, E.; Wilson, B.; Williams, P.J. Studies on the Hydrolysis of Two Megastigma-3, 6, 9-triols Rationlizing the Origins of Some Volatile C13 Norisoprenoids of Vitis vinifera Grapes. J. Agric. Food Chem. 1986, 34, 145–149. [Google Scholar] [CrossRef]
- Capon, R.J.; Ghisalberti Emilio, L.; Jefferies Phillip, R. New Sesquiterpenes from Caulerpa flexilis var. muelleri. Aust. J. Chem. 1981, 34, 1775–1778. [Google Scholar] [CrossRef]
- Blackman, A.J.; Wells, R.J. Flexilin and trifarin, Terpene 1, 4-diacetoxybuta-1, 3-dienes from two caulerpa species. Tetrahedron Lett. 1978, 33, 3063–3064. [Google Scholar] [CrossRef]
- Sun, Y.; Zhan, Y.C.; Sha, Y.; Pei, Y.H. Norisoprenoids from Ulva lactuca. J. Asian Nat. Prod. Res. 2007, 9, 321–325. [Google Scholar] [CrossRef]
- Li, G.-L.; Guo, W.-J.; Wang, G.-B.; Wang, R.-R.; Hou, Y.-X.; Liu, K.; Liu, Y.; Wang, W. Sterols from the Green Alga Ulva australis. Mar. Drugs 2017, 15, 299. [Google Scholar] [CrossRef] [Green Version]
- Kapetanovic, R.; Sladic, D.; Popov, S.; Zlatovic, M.; Kljajic, Z.; Gasic, M. Sterol composition of the Adriatic sea algae Ulva lactuca, Codium dichotomum, Cystoseira adriatica and Fucus virsoides. J. Serb. Chem. Soc. 2005, 70, 1395–1400. [Google Scholar] [CrossRef]
- Awad, N.E. Biologically active steroid from the green Alga Ulva lactuca. Phytother. Res. 2000, 14, 641–643. [Google Scholar] [CrossRef]
- Govindan, M.; Hodge, J.D.; Brown, K.A.; Nunez-Smith, M. Distribution of cholesterol in Caribbean marine algae. Steroids 1993, 58, 178–180. [Google Scholar] [CrossRef]
- Lopes, G.; Sousa, C.; Bernardo, J.; Andrade, P.B.; Valentão, P.; Ferreres, F.; Mouga, T. Sterol Profiles in 18 Macroalgae of the Portuguese Coast1. J. Phycol. 2011, 47, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, I.; Goad, L.J. Sterols of the siphonous marine alga Codium Fragile. Phytochemistry 1974, 13, 481–484. [Google Scholar] [CrossRef]
- Romeo, G.; Toscano, M.A. The isolation and characterization of (24S)-24-Methylcholesta-5, 25-dien-3B-ol from the siphonous marine alga Codium bursa. J. Nat. Prod. 1983, 46, 187–189. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Mabinya, L.V.; Olaniran, A.O.; Okoh, A.I. Chemical characterization of sulfated polysaccharides from Gracilaria gracilis and Ulva lactuca and their radical scavenging, metal chelating, and cholinesterase inhibitory activities. Int. J. Food Prop. 2019, 22, 100–110. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Xu, Y.; Chen, H.; Sun, P. Extraction, Structural Characterization, and Potential Antioxidant Activity of the Polysaccharides from Four Seaweeds. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef]
- Kammoun, I.; Bkhairia, I.; Ben Abdallah, F.; Jaballi, I.; Ktari, N.; Boudawara, O.; Nasri, M.; Gharsallah, N.; Hakim, A.; Ben Amara, I. Potential protective effects of polysaccharide extracted from Ulva lactuca against male reprotoxicity induced by thiacloprid. Arch. Physiol. Biochem. 2017, 123, 334–343. [Google Scholar] [CrossRef]
- MAhmed, O. Anti-Proliferative and Apoptotic Efficacies of Ulvan Polysaccharides against Different Types of Carcinoma Cells In Vitro and In Vivo. J. Canc. Sci. Ther. 2014, 6, 202–208. [Google Scholar]
- Capon, R.J.; Ghisalberti Emilio, L.; Jefferies Phillip, R. Metabolites of the green algae, Caulerpa species. Phytochemistry 1983, 22, 1465–1467. [Google Scholar] [CrossRef]
- Rochfort, S.J.; Watson, R.; Capon Robert, J. Dictyosphaerin: A Novel bicyclic lipid from a Southern Australian marine green algae, Dictyosphaeria sericea. J. Nat. Prod. 1996, 59, 1154–1156. [Google Scholar] [CrossRef]
- El Sayed, H.; Choudhary, M.I.; Kandil, S.H.; El Nemr, A.; Gulzar, T.; Shobier, A.H. Studies on the constituents of the green alga Ulva lactuca. Chem. Nat. Comp. 2011, 47, 335–338. [Google Scholar]
- Flodin, C.; Whitfield Frank, B. 4-Hydroxybenzoic acid: A likely precursor of 2, 4, 6-tribromophenol in Ulva lactuca. Phytochemistry 1999, 51, 249–255. [Google Scholar] [CrossRef]
- Whitfield, F.B.; Helidoniotis, F.; Shaw, K.J.; Svoronos, D. Distribution of bromophenols in species of marine algae from Eastern Australia. J. Agric. Food Chem. 1999, 47, 2367–2373. [Google Scholar] [CrossRef]
- Gregson, R.P.; Kazlauskas, R.; Murphy, P.T.; Wells, R.J. New Metabolites from the Brown Alga Cystophora torulosa. Aust. J. Chem. 1977, 30, 2527–2532. [Google Scholar] [CrossRef]
- Bian, B.; Van Altena, I.A. Four new compounds from the Australian brown alga Cystophora brownii. Aust. J. Chem. 1998, 51, 1157–1165. [Google Scholar] [CrossRef]
- Brkljača, R.; Urban, S. HPLC–NMR and HPLC–MS investigation of antimicrobial constituents in Cystophora monilifera and Cystophora subfarcinata. Phytochemistry 2015, 117, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Laird, D.W.; Bennett, S.; Bian, B.; Suer, B.; Wright, K.; Hughes, V.; Van Altena, I.A. Chemical Investigation of seven Australasian Cystophora species: New chemistry and taxanomic insights. Biochem. Syst. Ecol. 2010, 38, 187–194. [Google Scholar] [CrossRef]
- Kimura, J.; Maki, N. New Loliolide derivatives from the brown alga Undaria pinnatifida. J. Nat. Prod. 2002, 65, 57–58. [Google Scholar] [CrossRef]
- Peng, Y.; Huang, R.M.; Lin, X.P.; Liu, Y.H. Norisoprenoids from the Brown Alga Sargassum naozhouense Tseng et Lu. Molecules 2018, 23, 348. [Google Scholar] [CrossRef] [Green Version]
- Masum, S.; Hossain, M.A.; Akamine, H.; Sakagami, J.-I.; Ishii, T.; Gima, S.; Kensaku, T.; Bhowmik, P. Isolation and characterization of allelopathic compounds from the indigenous rice variety ‘Boterswar’ and their biological activity against Echinochloa crus-galli L. Allelopath. J. 2018, 43, 31–42. [Google Scholar] [CrossRef]
- Brkljača, R.; Urban, S. Chemical Profiling (HPLC-NMR & HPLC-MS), Isolation, and Identification of Bioactive Meroditerpenoids from the Southern Australian Marine Brown Alga Sargassum paradoxum. Mar. Drugs 2014, 13, 102–127. [Google Scholar]
- Reddy, P.; Urban, S. Meroditerpenoids from the southern Australian marine brown alga Sargassum fallax. Phytochemistry 2009, 70, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Ovenden, S.P.; Nielson, J.L.; Liptrot, C.H.; Willis, R.H.; Wright, A.D.; Motti, C.A.; Tapiolas, D.M. Comosusols A-D and comosone A: Cytotoxic compounds from the brown alga Sporochnus comosus. J. Nat. Prod. 2011, 74, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, L.S.; Wright, A.E.; Gunasekera, S.P.; McCarthy, P.; Reed, J. Antimicrobial Constituent of the Brown Alga Sporochnus pedunculatus. Int. J. Pharmacogn. 1995, 33, 253–255. [Google Scholar] [CrossRef]
- Rochfort, S.J.; Capon, R.J. A new sesquiterpene/phenol from the Australian marine brown alga Perithalia caudata. J. Nat. Prod. 1994, 6, 849–851. [Google Scholar] [CrossRef]
- Blackman, A.J.; Rogers, G.I.; Volkman, J.K. A phenol from the brown alga Perithalia caudata. Phytochemistry 1988, 27, 3686–3687. [Google Scholar] [CrossRef]
- Ravi, B.N.; Murphy, P.T.; Lidgard, R.O.; Warren, R.G.; Wells, R.J. C18 Terpenoid metabolites of the brown alga Cystophora moniliformis. Aust. J. Chem. 1982, 35, 171–182. [Google Scholar] [CrossRef]
- El Hattab, M.; Culioli, G.; Valls, R.; Richou, M.; Piovetti, L. Apo-fucoxanthinoids and loliolide from the brown alga Cladostephus spongiosus f. verticillatus (Heterokonta, Sphacelariales). Biochem. Syst. Ecol. 2008, 36, 447–451. [Google Scholar] [CrossRef]
- Van Altena, I.A. Terpenoids from the brown alga Cystophora moniliformis. Aust. J. Chem. 1988, 41, 49–56. [Google Scholar] [CrossRef]
- Li, Y.; Li, W.; Li, Y. An Alternative Short Synthesis of 6, 10, 14-Trimethyl-5E, 9E-Pentadecadiene-2, 13-dione. Synth. Commun. 1994, 24, 117–121. [Google Scholar] [CrossRef]
- Reddy, P.; Urban, S. Linear and cyclic C18 terpenoids from the Southern Australian marine brown alga Cystophora moniliformis. J. Nat. Prod. 2008, 71, 1441–1446. [Google Scholar] [CrossRef]
- Chen, J.; Li, H.; Zhao, Z.; Xia, X.; Li, B.; Zhang, J.; Yan, X. Diterpenes from the marine algae of the genus Dictyota. Mar. Drugs 2018, 16, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlop, R.W.; Ghisalberti, E.L.; Jefferies, P.R.; Skeltoni, B.W.; White, A.H. Structure of a new dolastane diterpene from Dictyota furcellata. Aust. J. Chem. 1989, 42, 315–319. [Google Scholar] [CrossRef]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Technol. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Aknin, M.; Dogbevi, K.; Samb, A.; Kornprobst, J.M.; Gaydou, E.M.; Miralles, J. Fatty acid and sterol composition of eight brown algae from the senegales coast. Comp. Biochem. Physiol. 1992, 102B, 841–843. [Google Scholar]
- Castro-Silva, E.S.; Bello, M.; Hernandez-Rodriguez, M.; Correa-Basurto, J.; Murillo-Alvarez, J.I.; Rosales-Hernandez, M.C.; Munoz-Ochoa, M. In vitro and in silico evaluation of fucosterol from Sargassum horridum as potential human acetylcholinesterase inhibitor. J. Biomol. Struct. Dyn. 2018. [Google Scholar] [CrossRef]
- Perumal, P.; Sowmiya, R.; Prasanna Kumar, S.; Ravikumar, S.; Deepak, P.; Balasubramani, G. Isolation, structural elucidation and antiplasmodial activity of fucosterol compound from brown seaweed, Sargassum linearifolium against malarial parasite Plasmodium falciparum. Nat. Prod. Res. 2017, 32, 1316–1319. [Google Scholar] [CrossRef]
- Narkowicz, C.K.; Blackman, A.J. Further acetogenins from Tasmanian collections of Caulocystis cephalornithos demonstrating chemical variability. Biochem. Syst. Ecol. 2006, 34, 635–641. [Google Scholar] [CrossRef]
- Gutiérrez-Cepeda, A.; Fernandez, J.J.; Norte, M.; Montalvao, S.; Tammela, P.; Souto, M.L. Acetate-Derived Metabolites from the Brown Alga Lobophora variegata. J. Nat. Prod. 2015, 78, 1716–1722. [Google Scholar] [CrossRef]
- Basha, S.; Jaiswar, S.; Jha, B. On the biosorption, by brown seaweed. Lobophora variegata, of Ni(II) from aqueous solutions: Equilibrium and thermodynamic studies. Biodegradation 2010, 21, 661–680. [Google Scholar]
- Capon, R.J.; Barrow, R.A.; Skene, C.; Rochfort, S. The biomimetic synthesis of marine epoxylipids: Bisepoxides to tetrahydrofurans. Tetrahedron Lett. 1997, 38, 7609–7612. [Google Scholar] [CrossRef]
- Kurihara, H.; Kagawa, Y.; Konno, R.; Kim, S.M.; Takashi, K. Lipoxygenase inhibitors derived from marine macroalgae. Bioorg. Med. Chem. Lett. 2014, 24, 1383–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capon, R.J.; Barrow, R.A.; Rochfort, S.; Jobling, M.; Skene, C. Marine nematocides: Tetrahydrofurans from a Southern Australian brown alga, Notheia Anomala. Tetrahedron 1998, 54, 2227–2242. [Google Scholar] [CrossRef]
- Roy, S.; Spilling, C.D. An expeditious total synthesis of both diastereoisomeric lipid dihydroxytetrahydrofurans from Notheia anomala. Org. Lett. 2012, 14, 2230–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrow, R.A.; Capon, R.J. Epoxy lipids from the Australian epiphytic brown alga Notheia anomala. Aust. J. Chem. 1990, 43, 895–911. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Mulder, J.; Murphy, P.T.; Wells, R.J. New metabolites from the brown alga Caulocystis cephalornithos. Aust. J. Chem. 1980, 33, 2097–2101. [Google Scholar] [CrossRef]
- Rochfort, S.; Murray, L.; Capon, R.J. The chemistry of notheia anomala, III: Two new methylene-interrupted trisepoxylipids. J. Nat. Prod. 1992, 55, 1332–1335. [Google Scholar] [CrossRef]
- Murray, L.M.; Barrow, R.A.; Capon, R.J. Epoxy lipids from the Australian epiphytic brown alga Notheia anomala. II. Aust. J. Chem. 1991, 44, 843–854. [Google Scholar] [CrossRef]
- Womersley, H.B.S. The morphology and taxonomy of Cystophora and related genera (Phaeophyta). Aust. J. Bot. 1964, 12, 53–110. [Google Scholar] [CrossRef]
- Brkljaca, R.; Gker, E.S.; Urban, S. Dereplication and chemotaxonomical studies of marine algae of the Ochrophyta and Rhodophyta phyla. Mar. Drugs 2015, 13, 2714–2731. [Google Scholar] [CrossRef] [Green Version]
- Kazlauskas, R.; King, L.; Murphy, P.T.; Warren, R.G.; Wells, R.J. New metabolites from the brown algal genus Cystophora. Aust. J. Chem. 1981, 34, 439–447. [Google Scholar] [CrossRef]
- Zhang, H.; Xiao, X.; Conte, M.M.; Khalil, Z.; Capon, R.J. Spiralisones A-D: Acylphloroglucinol hemiketals from an Australian marine brown alga, Zonaria spiralis. Org. Biomol. Chem. 2012, 10, 9671–9676. [Google Scholar] [CrossRef] [PubMed]
- Blackman, A.J.; Rogers, G.I. Phloroglucinol derivatives from three Australian marine algae of the genus Zonaria. J. Nat. Prod. 1988, 51, 158–160. [Google Scholar] [CrossRef]
- Gerwick, W.; Fenical, W. Phenolic lipids from related marine algae of the order Dictyotales. Phytochemistry 1982, 21, 633–637. [Google Scholar] [CrossRef]
- Choukchou-Braham, N.; Asakawa, Y.; Lepoittevin, J.P. Isolation, Structure determination and synthesis of new dihydroisocoumarins from Ginkgo balboa L. Tetrahedron Lett. 1994, 35, 3949–3952. [Google Scholar] [CrossRef]
- Zhu, Y.; Conklin, D.R.; Chen, H.; Wang, L.; Sang, S. 5-alk(en)ylresorcinols as the major active components in wheat bran inhibit human colon cancer cell growth. Bioorg. Med. Chem. 2011, 19, 3973–3982. [Google Scholar] [CrossRef]
- Wang, K.-W.; Zhang, T.-T.; Zhang, L. Chemical Constituents and Biological Activities of Grevillea robusta. Chem. Nat. Compd. 2018, 54, 153–155. [Google Scholar] [CrossRef]
- Guan, Y.F.; Song, X.; Qiu, M.H.; Luo, S.H.; Wang, B.J.; Van Hung, N.; Cuong, N.M.; Soejarto, D.D.; Fong, H.H.; Franzblau, S.G.; et al. Bioassay-Guided Isolation and Structural Modification of the Anti-TB Resorcinols from Ardisia gigantifolia. Chem. Biol. Drug Des. 2016, 88, 293–301. [Google Scholar] [CrossRef]
- Usov, A.I.; Smirnova, G.P.; Kamenarska, Z.; Dimitrova-Konaklieva, S.; Stefanov, K.L.; Popov, S.S. Polar constituents of brown seaweed Colpomenia peregrina (Sauv.) Hamel from the Black Sea. Russ. J. Bioorg. Chem. 2004, 30, 161–167. [Google Scholar] [CrossRef]
- Dasyam, N.; Munkacsi, A.B.; Fadzilah, N.H.; Senanayake, D.S.; O’Toole, R.F.; Keyzers, R.A. Identification and bioactivity of 3-epi-xestoaminol C isolated from the New Zealand brown alga Xiphophora chondrophylla. J. Nat. Prod. 2014, 77, 1519–1523. [Google Scholar] [CrossRef]
- Wirth, D.; Fischer-Liu, I.; Boland, W.; Icheln, D.; Runge, T.; Konig, W.A.; Philips, J.; Clayton, M. Unusual and novel C11H16 Hydrocarbons from the Southern Australian brown alga Dictyopteris acrostichoides (Phaeophyceae). Helv. Chim. Acta. 1992, 75, 734–744. [Google Scholar] [CrossRef]
- Boland, W. The chemistry of gamete attraction: Chemical structures, biosynthesis, and (a)biotic degradation of algal pheromones. Proc. Natl. Acad. Sci. USA 1995, 92, 37–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boland, W.; Marner, F.J.; Jaenicke, L.; Muller, D.G.; Folster, E. Comparative receptor study in gamete chemotaxis of the seaweeds Ectocarpus siliculosis and Cutleria multifida. Eur. J. Biochem. 1983, 134, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Schotten, T.; Boland, W.; Jaenicke, L. Synthesis of enantiomerically pure pheromones of south-pacific brown algae: Hormosirene and dictyopterene A. Helv. Chim. Acta. 1985, 68, 1186–1192. [Google Scholar] [CrossRef]
- Muller, D.G. Cystophorene and Hormosirene, Sperm Attractants in Australian Brown Algae. Naturwissenschaften 1985, 72, 97–99. [Google Scholar] [CrossRef]
- Green, D.; Kashman, Y. Colpol, a new cytotoxic C6-C4-C6 metabolite from the alga Colpomenia sinuosa. J. Nat. Prod. 1993, 56, 1201–1202. [Google Scholar] [CrossRef]
- Martin, L.J. Fucoxanthin and Its Metabolite Fucoxanthinol in Cancer Prevention and Treatment. Mar. Drugs 2015, 13, 4784–4798. [Google Scholar] [CrossRef] [Green Version]
- Zorofchian Moghadamtousi, S.; Karimian, H.; Khanabdali, R.; Razavi, M.; Firoozinia, M.; Zandi, K.; Abdul Kadir, H. Anticancer and antitumor potential of fucoidan and fucoxanthin, two main metabolites isolated from brown algae. Sci. World J. 2014, 2014, 768323. [Google Scholar] [CrossRef]
- Hussain, E.; Wang, L.-J.; Jiang, B.; Riaz, S.; Butt, G.Y.; Shi, D.-Y. A review of the components of brown seaweeds as potential candidates in cancer therapy. RSC Adv. 2016, 6, 12592–12610. [Google Scholar] [CrossRef]
- Mori, K.; Ooi, T.; Hiraoka, M.; Oka, N.; Hamada, H.; Tamura, M.; Kusumi, T. Fucoxanthin and its metabolites in edible brown algae cultivated in deep seawater. Mar. Drugs 2004, 2, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; Hogg, M.; Ulms, K.; Vetter, W. Concentrations, stability, and Isolation of the furan fatty acid 9-(3-methyl-5-pentylfuran-2-yl)-nonanoic acid from disposable latex gloves. J. Agric. Food Chem. 2017, 65, 7919–7925. [Google Scholar] [CrossRef]
- Kubanek, J.; Jensen, P.R.; Keifer, P.A.; Sullards, M.C.; Collins, D.O.; Fenical, W. Seaweed resistance to microbial attack: A targeted chemical defense against marine fungi. Proc. Natl. Acad. Sci. USA 2003, 100, 6916–6921. [Google Scholar] [CrossRef] [Green Version]
- Wirth, D.; Boland, W. Absolute configuration and synthesis of (+)-Caudoxirene, the gamete-releasing and gamete-attracting pheromone of the brown alga Perithalia caudata (Phaeophyceae). Helv. Chim. Acta. 1992, 75, 751–758. [Google Scholar] [CrossRef]
- Muller, D.G. Sexual pheromones in Cladostephus (Sphacelariale, Phaeophyceae). Naturwissenschaften 1986, 73, 99–100. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Murphy, P.T.; Wells, R.J.; Gregson, P. Two new furans from the brown alga Acrocarpia paniculata: The Use of 4-Phenyl-4H-1, 2, 4-triazolina-3, 5-dione to Determine the Substitution pattern of a Furan. Aust. J. Chem. 1982, 35, 165–170. [Google Scholar] [CrossRef]
- Norton, R.S.; Warren, R.G.; Wells, R.J. Three new polyhalogenated monoterpenes from Plocamium species. Tetrahedron Lett. 1977, 44, 3905–3908. [Google Scholar] [CrossRef]
- Dias, D.; Urban, S. Phytochemical analysis of the Southern Australian marine alga, Plocamium mertensii using HPLC-NMR. Phytochem. Anal. 2008, 19, 453–470. [Google Scholar] [CrossRef]
- San-martin, A.; Negrette, R.; Rovirosa, J. Insecticide and acaricide activities of polyhalogenated monoterpenes from chilean Plocamium cartilagineum. Phytochemistry 1991, 30, 2165–2169. [Google Scholar] [CrossRef]
- Argandona, V.H.; Rovirosa, J.; San-Martin, A.; Riquelme, A.; Diaz-Marrero, A.R.; Cueto, M.; Darias, J.; Santana, O.; Guadano, A.; Gonzalez-Coloma, A. Antifeedant effects of marine halogenated monoterpenes. J. Agric. Food Chem. 2002, 50, 7029–7033. [Google Scholar] [CrossRef]
- Konig, G.M.; Wright, A.D.; Linden, A. Plocamium hamatum and its monoterpenes: Chemical and biological investigations of the tropical marine red alga. Phytochemistry 1999, 52, 1047–1053. [Google Scholar] [CrossRef]
- Timmers, M.A.; Dias, D.A.; Urban, S. Application of HPLC-NMR in the Identification of Plocamenone and Isoplocamenone from the marine red alga Plocamium angustum. Mar. Drugs 2012, 10, 2089–2102. [Google Scholar] [CrossRef] [Green Version]
- Stierle, D.B.; Wing, R.M.; Sims, J.J. Marine natural products XI Costatone and Costatolide, new halogenated monoterpenes from the red seaweed, Plocamium costatum. Tetrahedron Lett. 1976, 49, 4455–4458. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Murphy, P.T.; Quinn, R.J.; Wells, R.J.; Schönhlzer, P.; F-Konzern, J.S.; Hoffmann-La Roche, F. Two polyhalogenated monoterpenes from the red alga Plocamium costatum. Tetrahedron Lett. 1976, 49, 4451–4454. [Google Scholar] [CrossRef]
- Motti, C.A.; Thomas-Hall, P.; Hagiwara, K.A.; Simmons, C.J.; Willis, R.; Wright, A.D. Accelerated identification of halogenated monoterpenes from Australian specimens of the red algae Plocamium hamatum and Plocamium costatum. J. Nat. Prod. 2014, 77, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Sakata, K.; Iwase, Y.; Ina, K.; Fujita, D. Halogenated terpenes isolated from the red alga Plocamium leptophyllum as feeding inhibitors for marine herbivores. Nippon Suisan Gakk. 1991, 57, 743–746. [Google Scholar] [CrossRef] [Green Version]
- Rochfort, S.J.; Capon, R.J. Parguerenes revisited: New brominated diterpenes from the Southern Australian marine red alga Laurencia filifromis. Aust. J. Chem. 1996, 49, 19–26. [Google Scholar] [CrossRef]
- Huang, X.-C.; Sun, Y.-L.; Salim, A.A.; Chen, Z.-S.; Capon, R.J. Parguerenes: Marine red alga bromoditerpenes as inhibitors of P-glycoprotein (ABCB1) in multidrug resistant human cancer cells. Biochem. Pharmacol. 2013, 85, 1257–1268. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Kurosawa, E.; Komiyama, K.; Suzuki, T. The structures of cytotoxic diterpenes containing bromine from the marine red alga Laurencia obtusa (Hudson) Lamouroux. Bull. Chem. Soc. Jpn. 1990, 63, 3066–3072. [Google Scholar] [CrossRef] [Green Version]
- Kurata, K.; Taniguchi, K.; Agatsuma, Y.; Suzuki, M. Diterpenoid feeding-deterrents from Laurencia saitoi. Phytochemistry 1998, 47, 363–369. [Google Scholar] [CrossRef]
- Awad, N.E. Bioactive brominated diterpenes from the marine red alga Jania rubens (L.) Lamx. Phytother. Res. 2004, 18, 275–279. [Google Scholar] [CrossRef]
- Sims, J.J.; Lin, G.H.Y.; Wing, M. Marine natural products elatol, a halogenated sesquiterpene alcohol from the red alga Laurencia elata. Tetrahedron Lett. 1974, 39, 3487–3490. [Google Scholar] [CrossRef]
- Dias, D.A.; Urban, S. Phytochemical studies of the southern Australian marine alga, Laurencia elata. Phytochemistry 2011, 72, 2081–2089. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.; Brito, I.; Moujir, L.; Paiz, N.; Darias, J.; Cueto, M. Cytotoxic sesquiterpenes from Aplysia dactylomela. J. Nat. Prod. 2005, 68, 1677–1679. [Google Scholar] [CrossRef] [PubMed]
- Davyt, D.; Fernandez, R.; Suescun, L.; Mombru, A.W.; Saldana, J.; Dominguez, L.; Coll, J.; Fujii, M.T.; Manta, E. New Sesquiterpene derivatives from the red alga Laurencia scoparia. Isolation, structure determination, and anthelintic activity. J. Nat. Prod. 2001, 64, 1552–1555. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Anangdan, S.P.; Tan, K.L.; Matsunaga, S. Role of secondary metabolites as defense chemicals against ice-ice disease bacteria in biofouler at carrageenophyte farms. J. Appl. Phycol. 2009, 22, 305–311. [Google Scholar] [CrossRef]
- Nocchi, N.; Soares, A.R.; Souto, M.L.; Fernandez, J.J.; Martin, M.N.; Pereira, R.C. Detection of a chemical cue from the host seaweed Laurencia dendroidea by the associated mollusc Aplysia brasiliana. PLoS ONE 2017, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Jongaramruong, J.; Blackman, A.J.; Skelton, B.W.; White, A.H. Chemical relationships between the sea hare Aplysia parvula and the red seaweed Laurencia filiformis from Tasmania. Aust. J. Chem. 2002, 55, 275–280. [Google Scholar] [CrossRef]
- Thomas, S.G.; Beveridge, A.A. The novel marine natural product plocamadiene A causes histamine release from mast cells of the guinea-pig and rat in vitro. Clin. Exp. Pharmacol. 1993, 20, 223–229. [Google Scholar] [CrossRef]
- Dunlop, R.W.; Murphy, P.T.; Wells, R.J. A new polyhalogenated monoterpene from the red alga Plocamium angustum. Aust. J. Chem. 1979, 32, 2735–2739. [Google Scholar] [CrossRef]
- Brownlee, R.T.C.; Hall, J.G.; Reiss, J.A. An application of the INEPT pulse sequence to the NMR assignment of halogenated marine natural products. Org. Magn. Reson. 1983, 21, 544–547. [Google Scholar] [CrossRef]
- Konig, G.M.; Wright, A.D.; de Nys, R. Halogenated Monoterpenes from Plocamium costatum and their biological activity. J. Nat. Prod. 1999, 62, 383–385. [Google Scholar] [CrossRef]
- Antunes, B.L.; Fleury, B.G.; Fujii, M.T.; Teixeira, V.L. Sesquiterpenes of the Brazilian marine red alga Laurencia filiformis (Rhodophyta, Ceramiales). Nat. Prod. Commun. 2008, 3, 1653–1654. [Google Scholar] [CrossRef] [Green Version]
- Kazlauskas, R.; Murphy, P.T.; Quinn, R.J.; Wells, R.J. New Laurene derivatives from Laurencia filiformis. Aust. J. Chem. 1976, 29, 2533–2539. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Murphy, P.T.; Wells, R.J.; Daly, J.J.; Oberhansli, W.E. Heterocladol, a halogenated selinana sesquiterpene of biosynthetic significance from the red alga Laurencia filiformis: Its isolation, crystal structure and absolute configuration. Aust. J. Chem. 1977, 30, 2679–2687. [Google Scholar] [CrossRef]
- Dias, D.A.; White, J.M.; Urban, S. Laurencia filiformis: Phytochemical profiling by conventional and HPLC-NMR approaches. Nat. Prod. Commun. 2009, 4, 157–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kladi, M.; Xenaki, H.; Vagias, C.; Papazafiri, P.; Roussis, V. New cytotoxic sesquiterpenes from the red algae Laurencia obtusa and Laurencia microcladia. Tetrahedron 2006, 62, 182–189. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Kawamoto, T.; Miwa, H.; Suzuki, M. Potent antibacterial activity of halogenated compounds against antibiotic resistant bacteria. Planta Med. 2004, 70, 1087–1090. [Google Scholar] [CrossRef] [Green Version]
- Konig, G.M.; Wright, A.D.; Franzblau, S.G. Assessment of antimycobacterial activity of a series of mainly marine derived natural products. Planta Med. 2000, 66, 337–342. [Google Scholar] [CrossRef]
- Brennan, M.R.; Erickson, K.L. Austradiol acetate and austradiol diacetate, 4, 6- dihydroxy-(+)-selinane derivatives from an Australian Laurencia sp. J. Org. Chem. 1982, 47, 3917–3921. [Google Scholar] [CrossRef]
- Capon, R.J.; Ghisalberti, E.L.; Jefferies, P.R.; Skelton, B.W.; White, A.H. Sesquiterpene metabolites from Laurencia filiformis. Tetrahedron 1981, 37, 1613–1621. [Google Scholar] [CrossRef]
- Barbero, H.; Díez-Poza, C.; Barbero, A. The Oxepane Motif in Marine Drugs. Mar. Drugs 2017, 15, 361. [Google Scholar] [CrossRef] [Green Version]
- Pettit, G.R.; Herald, C.L.; Allen, M.S.; Von Dreele, R.B.; Vanell, L.D.; Kao, J.P.Y.; Blake, W. Antineoplastic agents. 48. The isolation and structure of aplysistatin. J. Am. Chem. Soc. 1977, 99, 262–263. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Kamada, T.; Lee, W.-W.; Jeon, Y.-J. Anti-inflammatory activity of halogenated secondary metabolites of Laurencia snackeyi (Weber-van Bosse) Masuda in LPS-stimulated RAW 264.7 macrophages. J. Appl. Phycol. 2013, 25, 1805–1813. [Google Scholar] [CrossRef]
- Couladouros, E.A.; Vidali, V.P. Novel stereocontrolled approach to syn- and anti-oxepene-cyclogeranyl trans-fused polycyclic systems: Asymmetric total synthesis of (-)-Aplysistatin, (+)-Palisadin A, (+)-Palisadin B, (+)-12-hydroxy-palisadin B, and the AB ring system of adociasulfate-2 and toxicol A. Chemistry 2004, 10, 3822–3835. [Google Scholar] [PubMed]
- Hall, J.G.; Reiss, J.A. Elatenyne- a Pyrano[3, 2-b]pyranyl vinyl acetylene from the red alga Laurencia elata. Aust. J. Chem. 1986, 39, 1401–1409. [Google Scholar] [CrossRef]
- Brkljaca, R.; Urban, S. Relative configuration of the marine natural products Elatenyne using NMR spectroscopic and chemical derivatization methodologies. Nat. Prod. Commun. 2013, 8, 729–732. [Google Scholar] [CrossRef] [Green Version]
- Urban, S.; Brkljaca, R.; Hoshino, M.; Lee, S.; Fujita, M. Determination of the Absolute Configuration of the Pseudo-Symmetric Natural Product Elatenyne by the Crystalline Sponge Method. Angew. Chem. Int. Ed. Engl. 2016, 55, 2678–2682. [Google Scholar] [CrossRef]
- Dyson, B.S.; Burton, J.W.; Sohn, T.-I.; Kim, B.; Bae, H.; Kim, D. Total Synthesis and Structure Confirmation of Elatenyne: Success of Computational Methods for NMR Prediction with Highly Flexible Diastereomers. J. Am. Chem. Soc. 2012, 134, 11781–11790. [Google Scholar] [CrossRef]
- Snyder, S.A.; Brucks, A.P.; Treitler, D.S.; Moga, I. Concise synthetic approaches for the Laurencia family: Formal total syntheses of (+/-)-laurefucin and (+/-)-E- and (+/-)-Z-pinnatifidenyne. J. Am. Chem. Soc. 2012, 134, 17714–17721. [Google Scholar] [CrossRef]
- Iliopoulou, D.; Vagias, C.; Harvala, C.; Roussis, V. C15 Acetogenins from the red alga Laurencia obtusa. Phytochemistry 2002, 59, 111–116. [Google Scholar] [CrossRef]
- Abdel-Mageed, W.M.; Ebdel, R.; Valeriote, F.A.; Jaspars, M. Laurefurenynes A–F, new Cyclic Ether Acetogenins from a Marine Red Alga, Laurencia sp. Tetrahedron 2010, 66, 2855–2862. [Google Scholar] [CrossRef]
- Kinnel, R.B.; Dieter, R.K.; Meinwald, J.; Van engen, D.; Clardy, J.; Eisner, T.; Stallard, M.O.; Fenical, W. Brasilenyne and cis-dihydrorhodophytin: Antifeedant medium-ring haloethers from a sea hare (Aplysia brasiliana). Proc. Natl. Acad. Sci. USA 1979, 76, 3576–3579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capon, R.J.; Ghisalberti, E.L.; Mori, T.A.; Jefferies, P.R. Sesquiterpenes from Laurencia spp. J. Nat. Prod. 1988, 51, 1302–1304. [Google Scholar] [CrossRef]
- Woolner, V.H.; Jones, C.M.; Field, J.J.; Fadzilah, N.H.; Munkacsi, A.B.; Miller, J.H.; Keyzers, R.A.; Northcote, P.T. Polyhalogenated Indoles from the Red alga Rhodophyllis membranacea: The first Isolation of Bromo-Chloro-Iodo Secondary Metabolites. J. Nat. Prod. 2016, 79, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.R.; Erickson, K.L. Polyhalogenated indoles from the marine alga Rhodophyllis membranacea harvey. Tetrahedron Lett. 1978, 19, 1637–1640. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Lidgard, R.O.; Wells, R.J. New Polybrominated metabolites from the red alga Ptilonia australasica. Tetrahedron Lett. 1978, 34, 3165–3168. [Google Scholar] [CrossRef]
- Tran, T.D.; Pham, N.B.; Quinn, R.J. Unique Polybrominated Hydrocarbons from the Australian Endemic Red Alga Ptilonia australasica. J. Nat. Prod. 2016, 79, 570–577. [Google Scholar] [CrossRef]
- de Nys, R.; Coll, J.C.; Bowden, B.F. Delisea pulchra (cf. fimbriata) Revisited. The structural determination of two new metabolites from the red alga Delisea pulchra. Aust. J. Chem. 1992, 45, 1625–1632. [Google Scholar] [CrossRef]
- de Nys, R.; Wright, A.D.; Konig, G.M.; Sticher, O. New Halogenated Furanones from the marine alga Delisea pulchra (cf. fimbriata). Tetrahedron 1993, 49, 11213–11220. [Google Scholar] [CrossRef]
- Wright, A.D.; de Nys, R.; Angerhofer, C.K.; Pezzuto, J.M.; Gurrath, M. Biological activities and 3D QSAR studies of a series of Delisea pulchra (cf. fimbriata) derived natural products. J. Nat. Prod. 2006, 69, 1180–1187. [Google Scholar] [CrossRef]
- Ankisetty, S.; Nandiraju, S.; Win, H.; Park, Y.C.; Amsler, C.D.; McClintock, J.B.; Baker, J.A.; Diyabalanage, T.K.; Pasaribu, A.; Singh, M.P.; et al. Chemical investigation of predator-deterred macroalgae from the Antarctic peninsula. J. Nat. Prod. 2004, 67, 1295–1302. [Google Scholar] [CrossRef]
- Patterson, G.W. The Distribution of Sterols in Algae. Lipids 1971, 6, 120–127. [Google Scholar] [CrossRef]
- de Nys, R.; Steinberg, P.D.; Rogers, C.N.; Charlton, T.S.; Duncan, M.W. Quantitative variation of secondary metabolites in the sea hare Aplysia parvula and its host plant, Delisea pulchra. Mar. Ecol. Prog. Ser. 1996, 130, 135–146. [Google Scholar] [CrossRef]
- Konig, G.M.; Wright, A.D. Determination of the absolute configuration of a series of halogenated furanones from the marine alga Delisea pulchra. Helv. Chim. Acta. 1995, 78, 758–764. [Google Scholar] [CrossRef]
- Kabore, S.A.; Combaut, G.; Vidal, J.P.; Codomier, L.; Passet, J.; Girard, J.P.; Rossi, J.C. Sterols of the red alga Rissoella verruculosa. Phytochem. 1983, 22, 1239–1240. [Google Scholar] [CrossRef]
- Andrade, P.B.; Barbosa, M.; Matos, R.P.; Lopes, G.; Vinholes, J.; Mouga, T.; Valentao, P. Valuable compounds in macroalgae extracts. Food Chem. 2013, 138, 1819–1828. [Google Scholar] [CrossRef]
- Murray, L.; Currie, G.; Capon, R.J. A new Macrocyclic pyrone from a Southern Australian marine red alga. Aust. J. Chem. 1995, 48, 1485–1489. [Google Scholar] [CrossRef]
- Shin, J.; Paul, V.J.; Fenical, W. New macrocyclic alpha- and gamma- pyrones from the marine red alga phacelocarpus labillardieri. Tetrahedron Lett. 1986, 27, 5189–5192. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Murphy, P.T.; Wells, R.J.; Blackman, A.J. Macrocyclic Enol-Ethers containing an acetylic group from the red alga Phacelocarpus labillardieri. Aust. J. Chem. 1982, 35, 113–120. [Google Scholar] [CrossRef]
- Nozaki, H.; Minohara, K.; Miyazaki, I.; Kondo, H.; Shirane, F.; Nakayama, M. Two new pyrogallols from the marine alga, Grateloupia filicina (Wulfen) J. Agardh. Agric. Biol. Chem. 1988, 52, 3229–3230. [Google Scholar]
- Ohira, S.; Nozaki, H.; Ando, M.; Shirane, F.; Kondo, H.; Nakayama, M. Absolute structure of (-)-4, 5-trans-dihydroxy-3-methyl-2-cyclopentenone isolated from the marine alga, Grateloupia filicina (Wulfen) J. Agardh by the synthesis of both enantiomers. Agric. Biol. Chem. 1991, 55, 2437–2438. [Google Scholar]
- Watanabe, M.; Fuda, H.; Jin, S.; Sakurai, T.; Ohkawa, F.; Hui, S.-P.; Takeda, S.; Watanabe, T.; Koike, T.; Chiba, H. Isolation and Characterisation of a phenolic antioxidant from the Pacific Oyster (Crassostrea gigas). J. Agric. Food Chem. 2012, 60, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Fuda, H.; Jin, S.; Sakurai, T.; Ohkawa, F.; Hui, S.-P.; Takeda, S.; Watanabe, T.; Koike, T.; Chiba, H. A phenolic antioxidant fom the pacific oyster (Crassostrea gigas) inhibits oxidation of cultured human heptocytes mediated by diphenyl-1-prnylphosphine. Food Chem. 2012, 134, 2086–2089. [Google Scholar] [CrossRef] [PubMed]
- Arnesen, U.; Hallenstvet, M.; Liaaen-jensen, S. More about the carotenoids of red algae. Biochem. Syst. Ecol. 1979, 7, 87–89. [Google Scholar] [CrossRef]
- Schubert, N.; García-Mendoza, E.; Pacheco-Ruiz, I. Carotenoid Composition of Marine Red Algae. J. Phycol. 2006, 42, 1208–1216. [Google Scholar] [CrossRef]
- Impellizzeri, G.; Mangiafico, S.; Oriente, G.; Piatelli, M.; Sciuto, S.; Fattorusso, E.; Magno, S.; Santacroce, C.; Sica, D. Amino acids and low-molecular-weight carbohydrates of some marine red algae. Phytochemistry 1975, 14, 1549–1557. [Google Scholar] [CrossRef]
- Czeczuga, B.; Taylor, F.J. Carotenoid content in some species of the brown and red algae from the coastal area of New Zealand. Biochem. Syst. Ecol. 1987, 15, 5–8. [Google Scholar] [CrossRef]
- Urban, S.; Timmers, M. HPLC-NMR Chemical profiling and dereplication studies of the marine brown alga, Cystophora torulosa. Nat. Prod. Commun. 2013, 8, 715–719. [Google Scholar] [CrossRef] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CHLOROPHYTA | ■Carpoglossum confluens | ■Sargassum fallax | ■Dasyphloea insignis | ■Plocamium angustum |
| ▲Apjohnia laetevirens | ■Chlanidophora microphylla | ■Sargassopsis heteromorphum | ▲Delisea hypneoides | ■Plocamium cirrhosum |
| ▲Bryopsis vestita | ■Cladostephus spongiosus | ■Sargassum linearifolium | ■Dictyomenia harveyana | ▲Plocamium dilatatum |
| ■Caulerpa alternans | ■Colpomenia sinuosa | ■Sargassum paradoxum | ■Dictyomenia tridens | ■Plocamium mertensii |
| ■Caulerpa brownie | ■Cystophora brownii | ■Sargassum sonderi | ■Diplocladia patersonis | ▲Plocamium patagiatum |
| ■Caulerpa cactoides | ■Cystophora expansa | ■Sargassum spinuligerum | ▲Dudresnaya australis | ■Plocamium preissianum |
| ▲Caulerpa cliftonii | ■Cystophora grevillei | ■Sargassum vestitum | ■Echinothamnion hystrix | ■Pollexfenia lobata |
| ■Caulerpa flexilis | ■Cystophora moniliformis | ■Scytosiphon lomentaria | ■Echinothamnion mallardiae | ■Pollexfenia pedicellata |
| ■Caulerpa geminata | ■Cystophora monilifera | ■Seirococcus axillaris | ▲Erythroclonium sonderi | ■Polysiphonia decipiens |
| ▲Caulerpa longifolia | ■Cystophora platylobium | ■Sirophysalis trinodis | ▲Erythroclonium muelleri | ■Polyopes constrictus |
| ■Caulerpa obscura | ■Cystophora retorta | ■Suringariella harveyana | ▲Gelidium australe | ■Pterocladia lucida |
| ▲Caulerpa papillosa | ■Cystophora retroflexa | ■Undaria pinnatifida | ■Gelidium asperum | ■Rhodoglossum gigartinoides |
| ▲Caulerpa remotifolia | ▲Cystophora siliquosa | ■Xiphophora chondrophylla | ■Gelinaria ulvoidea | ■Rhodymenia leptophylla |
| ■Caulerpa scalpelliformis | ■Cystophora subfarcinata | ■Zonaria angustata | ▲Gigartina sonderi | ▲Rhodymenia novaehollandica |
| ■Caulerpa sedoides | ■Cystophora torulosa | ■Zonaria crenata | ■Gracilaria cliftonii | ■Rhodymenia obtusa |
| ■Caulerpa simpliciuscula | ■Caulocystis uvifera | ■Zonaria spiralis | ♦Grateloupia filicina | ▲Rhodymenia prolificans |
| ▲Caulerpa trifaria | ■Dictyota dichotoma | ▲Zonaria turneriana | ■Halopeltis australis | ■Sarcothalia crassifolia |
| ■Caulerpa vesiculifera | ■Dictyota furcellata | RHODOPHYTA | ▲Halymenia plana | ■Sonderophycus capensis |
| ■Chaetomorpha coliformis | ■Dictyota gunniana | ■Acrosorium ciliolatum | ■Hemineura frondosa | ■Sonderophycus coriaceus |
| ■Chaetomorpha linum | ▲Dictyota paniculata | ■Acrotylus australis | ■Heterosiphonia gunniana | ■Stenogramma interruptum |
| ■Chaetomorpha valida | ■Dictyopteris acrostichoides | ■Agarophyton chilense | ■Heterosiphonia muelleri | ■Thuretia quercifolia |
| ■Chlorodesmis baculifera | ■Dictyopteris muelleri | ■Ahnfeltiopsis fastigiata | ■Hypnea ramentacea | ■Wrangelia nobilis |
| ■Cladophora prolifera | ▲Distromium flabellatum | ■Areschougia congesta | ■Jania rosea | ■Wrangelia plumosa |
| ▲Cladophora rhizoclonioidea | ■Durvillaea potatorum | ■Ballia callitricha | ■Jania sagittata | |
| ▲Codium australicum | ■Ecklonia radiata | ▲Botryocladia sonderi | ■Laurencia botryoides | |
| ▲Codium duthieae | ■Exallosorus olsenii | ■Callophycus laxus | ♦Laurencia elata | |
| ■Codium fragile | ■Halopteris pseudospicata | ■Callophyllis rangiferina | ■Laurencia filiformis | |
| ■Codium galeatum | ■Homoeostrichus sinclairii | ■Callophyllis lambertii | ▲Lenormandia marginata | |
| ▲Codium harveyi | ■Hormosira banksii | ■Camontagnea oxyclada | ■Lenormandia muelleri | |
| ■Codium lucasii | ■Leathesia marina | ▲Capreolia implexa | ■Lophurella periclados | |
| ▲Codium pomoides | ■Lobospira bicuspidata | ■Centroceras clavulatum | ■Martensia australis | |
| ▲Dictyosphaeria sericea | ■Lobophora variegata | ■Cephalocystis furcellata | ■Melanthalia obtusata | |
| ■Ulva australis | ■Macrocystis pyrifera | ■Champia viridis | ■Meredithia nana | |
| ■Ulva compressa | ▲Notheia anomala | ■Chrysymenia brownii | ■Metagoniolithon radiatum | |
| ■Ulva lactuca | ■Padina fraseri | ■Corallina officinale | ■Metamastophora flabellata | |
| ■Ulva linza | ■Perithalia caudata | ■Corynecladia clavata | ■Nizymenia australis | |
| ■Ulva rigida | ■Petalonia fascia | ■Crassiphycus secundatus | ■Pachymenia orbicularis | |
| ■Ulva taeniata | ■Phyllospora comosa | ▲Cryptonemia undulata | ■Palisada tumida | |
| OCHROPHYTA | ■Phyllotricha decipiens | ■Curdiea angustata | ■Perbella minuta | |
| ■Acrocarpia paniculata | ■Phyllotricha varians | ■Dasya ceramioides | ▲Phacelocarpus alatus | |
| ▲Bellotia eriophorum | ■Phyllotricha verruculosa | ■Dasya naccarioides | ▲Phacelocarpus complanatus | |
| ■Caulocystis cephalornithos | ♦Sargassum decipiens | ■Dasya wilsonis | ■Phacelocarpus peperocarpos |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 1, 2 | Diterpene | C. brownii | Spring Beach, East Tasmania | [27] |
| 2a–f | Diterpene | C. brownii | Spring Beach, East Tasmania | [27] |
| 3, 4 | Diterpene | C. trifaria | Taroona Beach, Hobart, Tasmania | [25] |
| 5 | Diterpene | C. trifaria | Taroona Beach, Hobart, Tasmania | [25,27] |
| C. brownii | Spring Beach, East Tasmania | |||
| 6 | Diterpene | C. brownii | Spring Beach, East Tasmania | [27] |
| 7 | Diterpene | C. brownii | Flinders Reef, Victoria | [27] |
| 8 | Diterpene | C. brownii | Spring Beach, East Tasmania | [27] |
| 9 | Diterpene | C. trifaria | Taroona Beach, Hobart, Tasmania | [25] |
| 10 | Diterpene | C. brownii | Spring Beach, East Tasmania | [27] |
| 11 | Diterpene | C. trifaria | Taroona Beach, Hobart, Tasmania | [25] |
| 12 | Diterpene | C. brownii | Spring Beach, East Tasmania | [27] |
| 13–15 | Diterpene | C. fragile | Qingdao Coastline, Shangdong, China | [28] |
| 16 | Diterpene | C. brownii | Spring Beach, East Tasmania | [27] |
| 17 | Diterpene | C. trifaria | Taroona Beach, Hobart, Tasmania | [25,36] |
| C. flexilis | Cosy corner, Western Australia | |||
| 18 | Diterpene | C. trifaria | Taroona Beach, Hobart, Tasmania | [25,27] |
| C. brownii | Spring Beach, East Tasmania | |||
| 19, 20 | Diterpene | C. brownii | Spring Beach, East Tasmania | [27] |
| 21, 22 | Sesquiterpene | C. flexilis | Cosy corner, Western Australia | [36] |
| 23 | Sesquiterpene | C. flexilis | Cosy corner, Western Australia | [36,37] |
| C. trifaria | - | |||
| 24 | Sesquiterpene | C. trifaria | Taroona Beach, Hobart, Tasmania | [25] |
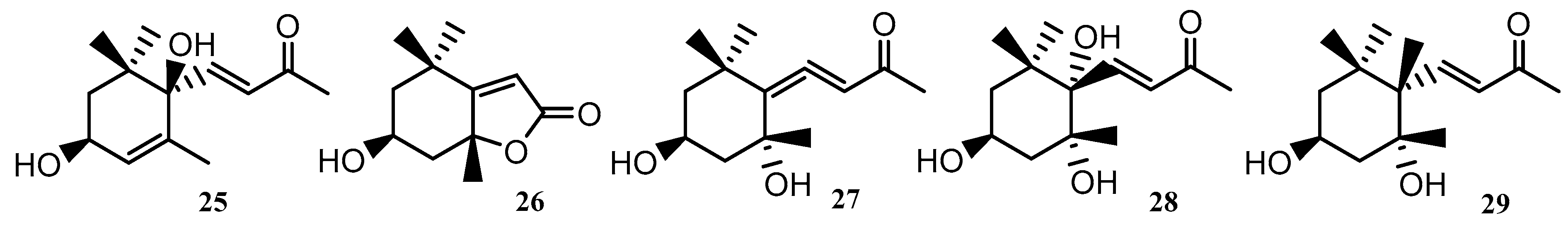
| 25–29 | Cyclic geranylacetone | U. lactuca | BoHai Coastline, China | [38] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 30–36 | Steroid/Sterol | U. lactuca | Bay of Kotor, Southern Adriatic Sea | [40] |
| 37 | Steroid/Sterol | C. fragile | - | [45] |
| 38, 39 | Steroid/Sterol | C. fragile | Qingdao Coastline, Shangdong, China | [28] |
| 40 | Steroid/Sterol | U. lactuca | Abu Qir Bay, Alexandria, Egypt | [40,52] |
| Bay of Kotor, Southern Adriatic Sea | ||||
| 41–46 | Steroid/Sterol | U. australis | Dalian Coast, China | [39] |
| 47 | Steroid/Sterol | U. lactuca | Abu Qir Bay, Alexandria, Egypt | [41] |
| 48 | Steroid/Sterol | C. fragile | Qingdao Coastline, Shangdong, China | [28] |
| 49 | Lipid | D. sericea | Cape Schank and Point Lonsdale, Victoria | [51] |
| 50–52 | Lipid | U. lactuca | Abu Qir Bay, Alexandria, Egypt | [41] |
| 53 | Di indolo pigment | C. trifaria | Point Peron, WA | [50] |
| C. brownii | Augusta, WA | |||
| C. flexilis | Augusta, WA | |||
| C. peltata | Big Nook Island, WA | |||
| C. racemosa | Big Nook Island, WA | |||
| 54 | Bromophenolic | U. lactuca | Bateau Bay, NSW | [54] |
| C. lucasii | Bateau Bay, NSW | |||
| C. galeatum | Bateau Bay, NSW | |||
| C. cactoides | Bateau Bay, NSW | |||
| 55 | Bromophenolic | U. lactuca | Bateau Bay, NSW | [54] |
| C. lucasii | Bateau Bay, NSW | |||
| C. galeatum | Bateau Bay, NSW | |||
| C. cactoides | Bateau Bay, NSW | |||
| 56 | Bromophenolic | U. lactuca | Bateau Bay, NSW | [54] |
| C. lucasii | Bateau Bay, NSW | |||
| C. galeatum | Bateau Bay, NSW | |||
| C fragile | Bateau Bay, NSW | |||
| C. cactoides | Bateau Bay, NSW | |||
| 57 | Bromophenolic | U. lactuca | Bateau Bay, NSW | [54] |
| C. lucasii | Bateau Bay, NSW | |||
| C. galeatum | Bateau Bay, NSW | |||
| C. fragile | Bateau Bay, NSW | |||
| C. cactoides | Bateau Bay, NSW | |||
| 58 | Bromophenolic | U. lactuca | Bateau Bay, NSW | [54] |
| C. lucasii | Bateau Bay, NSW | |||
| C. galeatum | Bateau Bay, NSW | |||
| C fragile | Bateau Bay, NSW | |||
| C. cactoides | Bateau Bay, NSW |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
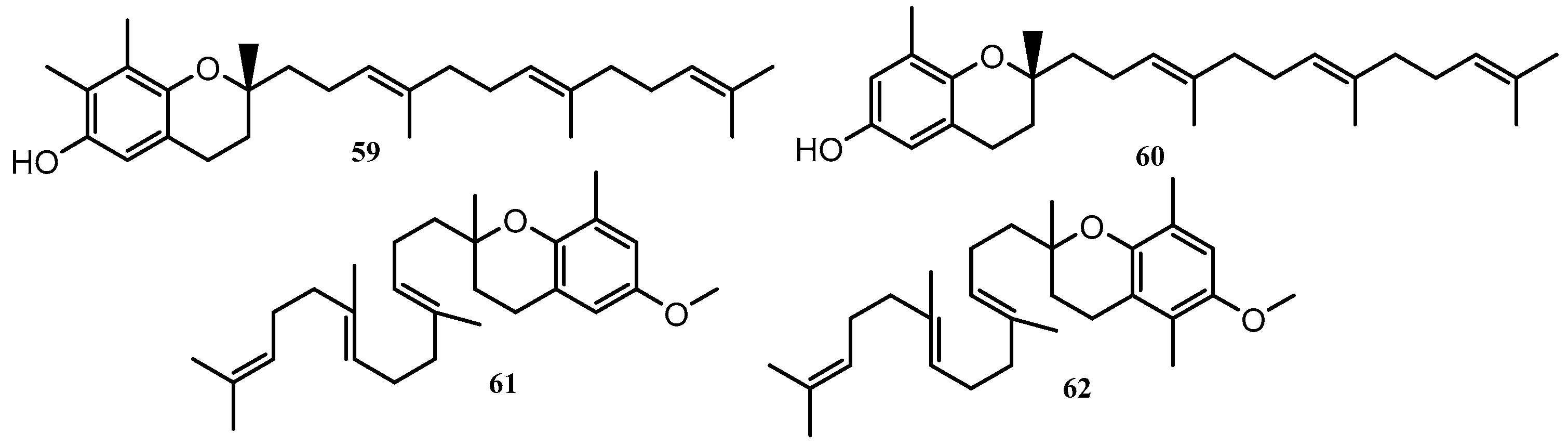
| 59 | Tocotrienols | C. monilifera | Governor Reef, Indented Head, Victoria | [57] |
| 60 | Tocotrienols | C. subfarcinata | Queenscliffe, Victoria | [55,56,57,58] |
| C. platylobium | - | |||
| C. monilifera | Governor Reef, Indented Head, Victoria | |||
| C. siliquosa | Sorrento Back Beach, Victoria | |||
| C. retorta | Cowaramup Bay, WA | |||
| 61, 62 | Tocotrienols | C. torulosa | Torquay, Victoria | [55] |
| 63–66 | Monoterpenes | U. pinnatifida | Miura Peninsula, Japan | [59] |
| 67 | Monoterpenes | U. pinnatifida | Miura Peninsula, Japan | [59,68,69] |
| C. moniliformis | - | |||
| C. spongiosus | Tipaza, Algerian Mediterranean Coast | |||
| 68, 69 | Prenylated Phenols | S. paradoxum | Governor Reef, Indented Head, Victoria | [62,63] |
| S. fallax | Governor Reef, Indented Head, Victoria | |||
| 70, 71 | Prenylated Phenols | S. paradoxum | Governor Reef, Indented Head, Victoria | [57] |
| 72 | Prenylated Phenols | P. caudata | Flinders Reef, Victoria | [66] |
| 73 | Prenylated Phenols | P. caudata | Ninepin point, D’Entrecasteaux Channel, Tasmania | [67] |
| 74 | Prenylated Phenols | P. caudata | Flinders Reef, Victoria | [66] |
| 75, 76 | Prenylated Phenols | C. brownii | Victor Harbour, SA | [56] |
| 77 | Prenylated Phenols | S. paradoxum | Governor Reef, Indented Head, Victoria | [57] |
| 78, 79 | Prenylated Phenols | C. torulosa | Cook Straight, Wellington, New Zealand | [58] |
| 80 | Prenylated Phenols | P. caudata | Flinders Reef, Victoria | [66] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 81 | Meroditerpenoids | C. torulosa | Cook Straight, Wellington, New Zealand | [58] |
| 82 | Meroditerpenoids | S. paradoxum | Governor Reef, Indented Head, Victoria | [57] |
| 83 | Meroditerpenoids | S. paradoxum | Governor Reef, Indented Head, Victoria | [57,63] |
| S. fallax | Governor Reef, Indented Head, Victoria | |||
| 84, 85 | Meroditerpenoids | S. paradoxum | Governor Reef, Indented Head, Victoria | [57] |
| 86, 87 | Meroditerpenoids | S. paradoxum | Governor Reef, Indented Head, Victoria | [57,63] |
| S. fallax | Governor Reef, Indented Head, Victoria | |||
| 88 | Meroditerpenoids | S. paradoxum | Governor Reef, Indented Head, Victoria | [57] |
| 89 | Meroditerpenoids | S. fallax | Governor Reef, Indented Head, Victoria | [63] |
| 90 | Meroditerpenoids | S. paradoxum | Governor Reef, Indented Head, Victoria | [57] |
| 91 | Meroditerpenoids | S. fallax | Governor Reef, Indented Head, Victoria | [63] |
| 92–94 | Farnesylacetone epoxide | C. moniliformis | Sarge Bay, Cape Leeuwin, WA | [58] |
| 95–97 | Farnesylacetone | C. moniliformis | Port Phillip Bay, Victoria | [72] |
| 98–100 | Geranylacetone, Geranylgeranal epoxide | C. moniliformis | North East side of West Island, SA | [70] |
| 101 | Farnesylacetone | C. moniliformis | - | [71] |
| 102, 103 | Cyclic farnesylacetone | C. moniliformis | Port Phillip Bay, Victoria | [72] |
| 104, 105 | Cyclic farnesylacetone | C. moniliformis | - | [68] |
| 106, 107 | Cyclic farnesylacetone | C. moniliformis | Port Phillip Bay, Victoria | [72] |
| 108 | Cyclic farnesylacetone | C. moniliformis | - | [68] |
| 109 | Cyclic farnesylacetone | C. moniliformis | North East side of West Island, SA | [70] |
| 110 | Cyclic farnesylacetone | C. moniliformis | Port Phillip Bay, Victoria | [72] |
| 111 | Aromadendrene | C. moniliformis | - | [68] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 112 | Diterpene | D. dichotoma | Northern Adriatic Sea | [73] |
| 113 | Diterpene | D. dichotoma | Red Sea | [73] |
| 114 | Diterpene | D. dichotoma | Saronicos Gulf, Greece | [73] |
| 115–117 | Diterpene | D. dichotoma | Tyrrhenian Sea | [73] |
| 118 | Diterpene | D. dichotoma | Puerto Madryn | [73] |
| 119 | Diterpene | D. dichotoma | Red Sea | [73] |
| 120 | Diterpene | D. dichotoma | Northern Adriatic Sea | [73] |
| 121 | Diterpene | D. dichotoma | Tyrrhenian Sea | [73] |
| 122–124 | Diterpene | D. dichotoma | Red Sea, Egypt | [73] |
| 125 | Diterpene | D. dichotoma | Troitsa Bay, Russian Far East | [73] |
| 126–128 | Diterpene | D. dichotoma | Red Sea, Egypt | [73] |
| 129 | Diterpene | D. dichotoma | Japan | [73] |
| 130 | Diterpene | D. dichotoma | Red Sea, Egypt | [73] |
| 131, 132 | Diterpene | D. dichotoma | Patagonia | [73] |
| 133 | Diterpene | D. dichotoma | Tyrrhenian Sea | [73] |
| 134 | Diterpene | D. dichotoma | - | [73] |
| 135 | Diterpene | D. dichotoma | Acicastello, Italy | [73] |
| 136, 137 | Diterpene | D. dichotoma | Russian Far East | [73] |
| 138, 140 | Diterpene | D. dichotoma | - | [73] |
| 141, 142 | Diterpene | D. dichotoma | Acicastello, Italy | [73] |
| 143 | Diterpene | D. furcellata | Cape Peron, Shark Bay, WA | [73] |
| 144–151 | Diterpene | D. dichotoma | Indian Ocean | [73] |
| 152–157 | Diterpene | D. dichotoma | Acicastello, Italy | [73] |
| 158–164 | Diterpene | D. dichotoma | Indian Ocean | [73] |
| 165, 166 | Diterpene | D. dichotoma | Red Sea | [73] |
| 167, 168 | Diterpene | D. dichotoma | - | [73] |
| 169, 171 | Diterpene | D. dichotoma | Karachi Coast, Arabian Sea | [73] |
| 172, 173 | Diterpene | D. dichotoma | Red Sea | [73] |
| 174 | Diterpene | D. dichotoma | Indian Ocean | [73] |
| 175 | Diterpene | D. dichotoma | - | [73] |
| 176 | Diterpene | D. dichotoma | Oshoro Bay, Hokkaido, Japan | [73] |
| 177–179 | Diterpene | D. dichotoma | Yagachi, Okinawa, Japan | [73] |
| 180 | Diterpene | D. dichotoma | - | [73] |
| 181–183 | Diterpene | D. dichotoma | Oshoro Bay, Hokkaido, Japan | [73] |
| 184–186 | Diterpene | D. dichotoma | - | [73] |
| 187 | Diterpene | D. dichotoma | Yagachi, Okinawa, Japan | [73] |
| 188 | Diterpene | D. dichotoma | - | [73] |
| 189 | Diterpene | D. dichotoma | Nagahama Beach, Ehime, Japan | [73] |
| 190, 191 | Diterpene | D. dichotoma | Troitsa Bay, Russian Far East | [73] |
| 192 | Diterpene | D. dichotoma | - | [73] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 193–195 | Steroids/Sterols | C. sinuosa | Cap Vert, Dakar | [76] |
| D. dichotoma | - | |||
| 196 | Steroids/Sterols | C. sinuosa | Cap Vert, Dakar | [43,76] |
| C. spongiosus | Praia do quebrado, Portugal | |||
| D. dichotoma | - | |||
| 197 | Steroids/Sterols | C. sinuosa | Cap Vert, Dakar | [76] |
| D, dichotoma | ||||
| 198, 199 | Steroids/Sterols | C. sinuosa | Cap Vert, Dakar | [42,43,76] |
| C. spongiosus | Praia do quebrado, Portugal | |||
| L. variegata | St Thomas, Virgin Islands | |||
| D. dichotoma | Cap Vert, Dakar | |||
| 200 | Steroids/Sterols | S. linearfolium | Bateau Bay, NSW | [43,75,76] |
| C. sinuosa | Cap Vert, Dakar | |||
| D. dichotoma | Cap Vert, Dakar | |||
| C. spongiosus | Praia do quebrado, Portugal | |||
| 201 | Steroids/Sterols | C. brownii | Victor Harbour, SA | [56] |
| 202 | Steroids/Sterols | C. brownii | Victor Harbour, SA | [56,76] |
| C. sinuosa | Cap Vert, Dakar | |||
| 203–221 | Lipid | C. cephalornithos | Southern and South Eastern Tasmania | [79] |
| 222 | Lipid | C. cephalornithos | Victorian Coastline | [87] |
| 223 | Lipid | C. cephalornithos | Southern and South Eastern Tasmania | [79] |
| 224 | Lipid | C. cephalornithos | Victorian Coastline | [87] |
| 225–227 | Lipid | L. variegata | Tenerife, Canary Islands | [80] |
| 228 | Polyene | N. anomala | Bells Beach, Victoria | [55,58,86] |
| C. torulosa | Torquay, Victoria | |||
| C. retorta | Cowaramup Bay, WA | |||
| 229, 230 | Polyene | N. anomala | Bells Beach, Victoria | [86] |
| 231 | Polyene | N. anomala | Bells Beach, Victoria | [58,86] |
| C. torulosa | Cook Straight, Wellington, New Zealand | |||
| C. retorta | Cowaramup Bay, WA | |||
| 232, 233 | Oxylipid | N. anomala | Bells Beach, Victoria | [84] |
| 234 | Epoxylipid | N. anomala | Torquay, Victoria | [82] |
| 235 | Epoxylipid | N. anomala | Southern and South Eastern Tasmania | [88] |
| 236–245 | Epoxylipid | N. anomala | Bells Beach, Victoria | [86,89] |
| 246–254 | Oxylipid | N. anomala | Bells Beach, Victoria | [86,89] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 255 | Phloroglucinol | C. subfarcinata | Queenscliffe, Victoria | [57,58,91] |
| C. monilifera | Governor Reef, Indented Head, Victoria | |||
| C. retroflexa | Governor Reef, Indented Head, Victoria | |||
| C. retorta | Cowaramup Bay, WA | |||
| 256 | Phloroglucinol | C. torulosa | Torquay, Victoria | [55,57,58] |
| C. subfarcinata | Queenscliffe, Victoria | |||
| C. siliquosa | Sorrento Back Beach | |||
| C. retorta | Cowaramup Bay, WA | |||
| C. monilifera | Governor Reef, Indented Head, Victoria | |||
| 257, 258 | Phloroglucinol | Z. spiralis | North Walkerville, Victoria | [93] |
| 259 | Phloroglucinol | C. subfarcinata | Queenscliffe, Victoria | [57] |
| C. monilifera | Governor Reef, Indented Head, Victoria | |||
| 260 | Phloroglucinol | C. subfarcinata | Queenscliffe, Victoria | [57] |
| 261 | Phloroglucinol | C. subfarcinata | North Eastern West Island, SA | [58,92] |
| C. monilifera | - | |||
| 262 | Phloroglucinol | C. retroflexa | Governor Reef, Indented Head, Victoria | [57,91] |
| C. monilifera | Governor Reef, Indented Head, Victoria | |||
| 263 | Phloroglucinol | C. retroflexa | Governor Reef, Indented Head, Victoria | [91] |
| 264 | Phloroglucinol | Z. spiralis | North Walkerville, Victoria | [93] |
| 265 | Phloroglucinol | C. subfarcinata | Queenscliffe, Victoria | [57,91] |
| C. monilifera | Governor Reef, Indented Head, Victoria | |||
| C. retroflexa | Governor Reef, Indented Head, Victoria | |||
| 266 | Phloroglucinol | C. monilifera | Governor Reef, Indented Head, Victoria | [57] |
| 267, 268 | Phloroglucinol | Z. turneriana | Tinderbox, Tasmania | [94] |
| Z. crenata | Tinderbox, Tasmania | |||
| Z angustata | Sisters Beach, Tasmania | |||
| 269, 270 | Phloroglucinol | C. subfarcinata | Queenscliffe, Victoria | [57,91] |
| C. monilifera | Governor Reef, Indented Head, Victoria | |||
| C. retroflexa | Governor Reef, Indented Head, Victoria | |||
| 271 | Phloroglucinol | C. subfarcinata | Queenscliffe, Victoria | [57,91] |
| C. retroflexa | Governor Reef, Indented Head, Victoria | |||
| 272 | Phloroglucinol | C. monilifera | Governor Reef, Indented Head, Victoria | [57] |
| 273–275 | Phloroglucinol | Z. spiralis | North Walkerville, Victoria | [93] |
| 276, 277 | Benzopyranones | C. cephalornithos | Southern and South Eastern Tasmania | [79] |
| 278 | Phenolic Acid | C. cephalornithos | Southern and South Eastern Tasmania | [79,91] |
| S. decipiens | Governor Reef, Indented Head, Victoria | |||
| 279 | Phenolic Acid | C. cephalornithos | Southern and South Eastern Tasmania | [79] |
| 280 | Phenolic Acid | C. cephalornithos | Southern and South Eastern Tasmania | [79,91] |
| S. decipiens | Governor Reef, Indented Head, Victoria | |||
| 281, 282 | Phenolic Acid | C. cephalornithos | Southern and South Eastern Tasmania | [79] |
| 283 | Phenol | C. cephalornithos | Southern and South Eastern Tasmania | [79,91] |
| S. decipiens | Governor Reef, Indented Head, Victoria | |||
| 284 | Phenol | C. cephalornithos | Southern and South Eastern Tasmania | [79] |
| 285 | Resorcinol | C. cephalornithos | Southern and South Eastern Tasmania | [79,91] |
| S. decipiens | Governor Reef, Indented Head, Victoria | |||
| 286 | Resorcinol | C. cephalornithos | Southern and South Eastern Tasmania | [55,79] |
| C. torulosa | Torquay, Victoria | |||
| 287 | Resorcinol | C. torulosa | Torquay, Victoria | [55] |
| 288, 289 | Phenolic Acid | C. cephalornithos | Southern and South Eastern Tasmania | [79] |
| 290 | Resorcinol | C. torulosa | Cook Straight, Wellington, New Zealand | [58] |
| 291 | Resorcinol | C. torulosa | Torquay, Victoria | [55] |
| 292–295 | Phenolic Acid | C. peregrina | Bulgarian Coast | [100] |
| 296, 297 | Phenolic Acid | L. variegata | Tenerife, Canary Islands | [80] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 298 | Xestoaminol | X. chondrophylla | Hen and Chicken Islands, New Zealand | [101] |
| 299–303 | Pheromone | D. acrostichoides | Point Lonsdale and Sorrento, Victoria | [102] |
| 304, 305 | Pheromone | D. acrostichoides | Point Lonsdale and Sorrento, Victoria | [102] |
| C. peregrina | Flinders, Victoria | [106] | ||
| 306 | Pheromone | D. acrostichoides | Point Lonsdale and Sorrento, Victoria | [102] |
| 307 | Pheromone | X. chondrophylla | - | [105] |
| S. lomentaria | - | |||
| 308 | Pheromone | H. banksii | Flinders Reef, Victoria | [103] |
| 309 | Pheromone | X. chondrophylla | - | [105] |
| S. lomentaria | - | |||
| 310–312 | Pheromone | P. caudata | - | [114] |
| 313 | Pheromone | C. spongiosus | Flinders Reef, Victoria | [102,115] |
| D. acrostichoides | Point Lonsdale and Sorrento, Victoria | |||
| 314 | Pheromone | M. pyrifera | - | [103] |
| U. pinnatifida | - | |||
| 315 | Pheromone | C. spongiosus | Flinders Reef, Victoria | [115] |
| 316, 317 | Pheromone | D. acrostichoides | Point Lonsdale and Sorrento, Victoria | [102] |
| 318–322 | Bromophenolic | C. spongiosus | Bateau Bay, NSW | [54] |
| C. sinuosa | Bateau Bay, NSW | |||
| E. radiata | Bateau Bay, NSW | |||
| H. sinclairii | Bateau Bay, NSW | |||
| H. banksii | Bateau Bay, NSW | |||
| P. comosa | Bateau Bay, NSW | |||
| L. variegata | Bateau Bay, NSW | |||
| 323 | Bromophenolic | C. sinuosa | Gulf of Eilat, Israel | [107] |
| 324 | Xanthophyll | S. lomentaria | Aikappu, Akkeshi, Hokkaido | [111] |
| 325 | Xanthophyll | S. lomentaria | Aikappu, Akkeshi, Hokkaido | [69,111] |
| C. spongiosus | Algerian Mediterranean Coast, Tipaza | |||
| 326 | Xanthophyll | S. lomentaria | Aikappu, Akkeshi, Hokkaido | [69,93,111] |
| C. spongiosus | Algerian Mediterranean Coast, Tipaza | |||
| Z. spiralis | North Walkerville, Victoria | |||
| 327, 328 | Xanthophyll | U. pinnatifida | - | [108] |
| 329 | Xanthophyll | U. pinnatifida | - | [69,91,108,111] |
| S. lomentaria | Aikappu, Akkeshi, Hokkaido | |||
| C. spongiosus | Algerian Mediterranean Coast, Tipaza | |||
| H. pseudospicata | Queenscliffe, Victoria | |||
| S. vestitum | Queenscliffe, Victoria | |||
| 330, 331 | Furans | A. paniculata | Port MacDonnell | [116] |
| 332 | Pyridine | C. peregrina | Bulgarian Coast | [100] |
| 333 | Amine | C. peregrina | Bulgarian Coast | [100] |
| 334 | Polyketide Macrolide | L. variegata | Cay Lobos, Bahamas | [113] |
| 335–339 | Polyketides | L. variegata | Tenerife, Canary Islands | [80] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 340–342 | Halogenated Monoterpene | P. mertensii | Queenscliffe, Victoria | [118] |
| 343 | Halogenated Monoterpene | P. mertensii | Carnac Island, WA | [139] |
| 344, 345 | Halogenated Monoterpene | P. mertensii | Queenscliffe, Victoria | [118] |
| 346 | Halogenated Monoterpene | P. mertensii | - | [117] |
| 347 | Halogenated Monoterpene | P. mertensii | Queenscliffe, Victoria | [118] |
| 348 | Halogenated Monoterpene | P. angustum | Cape Northumberland, SA | [140] |
| 349, 350 | Halogenated Monoterpene | P. angustum | Rocky Point, Torquay, Victoria | [141] |
| 351 | Halogenated Monoterpene | P. angustum | Queenscliffe, Victoria | [122,123] |
| P. costatum | Robe, South Australia | |||
| 352, 353 | Halogenated Monoterpene | P. angustum | Queenscliffe, Victoria | [122] |
| 354 | Halogenated Monoterpene | P. costatum | Robe, South Australia | [123] |
| 355 | Halogenated Monoterpene | P. costatum | Port MacDonnell, South Australia | [124] |
| 356 | Halogenated Monoterpene | P. costatum | Deep Glen Bay, Tasmania | [142] |
| 357 | Halogenated Monoterpene | P. costatum | Pandalowie Bay, South Australia | [125] |
| 358 | Halogenated Monoterpene | P. costatum | Pandalowie Bay, South Australia | [125] |
| 359, 360 | Halogenated Monoterpene | P. costatum | Deep Glen Bay, Tasmania | [142] |
| 361–364 | Halogenated Monoterpene | P. costatum | Pandalowie Bay, South Australia | [125] |
| 365 | Halogenated Monoterpene | P. leptophyllum | Toyama Bay, Japan | [126] |
| 366–374 | Parguerene | L. filiformis | South Australia | [127,128] |
| 375, 376 | Chamigrene | L. elata | St. Pauls Beach, Sorrento, Victoria | [133] |
| 377 | Chamigrene | L. filiformis | Taroona Beach, Hobart, Tasmania | [133,138] |
| L. elata | St. Pauls Beach, Sorrento, Victoria | |||
| 378 | Chamigrene | L. elata | New South Wales Coast | [132] |
| 379, 380 | Chamigrene | L. filiformis | Taroona Beach, Hobart, Tasmania | [138] |
| 381 | Chamigrene | L. filiformis | Taroona Beach, Hobart, Tasmania | [132,138] |
| L. elata | New South Wales Coast | |||
| 382–385 | Chamigrene | L. filiformis | Stella Maris Beach, Salvador, Brazil | [143] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 386 | Laurene | L. filiformis | Hamelin Bay, Perth, WA | [164] |
| Shoalwater Bay, Perth, WA | ||||
| Cottesloe Beach, Perth, WA | ||||
| Lancelin, Perth, WA | ||||
| 387, 388 | Laurene | L. filiformis | Shoalwater Bay, Perth, WA | [164] |
| Cottesloe Beach, Perth, WA | ||||
| Lancelin, Perth, WA | ||||
| 389 | Laurene | L. filiformis | South Australian Coast | [144] |
| 390 | Laurene | L. filiformis | St. Pauls Beach, Sorrento, Victoria | [146] |
| 391 | Laurene | L. filiformis | Shoalwater Bay, Perth, WA | [164] |
| 392 | Laurene | L. filiformis | Port MacDonnell Beach, South Australia | [145] |
| 393, 394 | Laurene | L. filiformis | Shoalwater Bay, Perth, WA | [164] |
| 395, 396 | Laurene | L. filiformis | St. Pauls Beach, Sorrento, Australia | [146] |
| 397 | Laurene | L. filiformis | Port MacDonnell Beach, South Australia | [145] |
| 398, 399 | Sesquiterpenoids | L. filiformis | Western Australia | [127,150] |
| South Australia | ||||
| 400 | Sesquiterpenoids | L. filiformis | Port MacDonnell Beach, South Australia | [145,146] |
| St. Pauls Beach, Sorrento, Australia | ||||
| 401, 402 | Sesquiterpenoids | L. filiformis | Point Peron, WA | [151] |
| 403, 404 | Sesquiterpenoids | L. elata | Batemans Bay, New South Wales | [133] |
| St. Pauls Beach, Sorrento, Australia | ||||
| 405, 406 | Lauroxocane | L. filiformis | Western Australia | [150] |
| 407, 408 | Lauroxocane | L. filiformis | St. Pauls Beach, Sorrento, Australia | [133] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 409–424 | Polyhalogenated Indole | R. membranacea | Moa Point, New Zealand | [165,166] |
| 425–426 | Polyhalogenated Hydrocarbon | P. australasica | Pearsons Point, Tasmania | [168] |
| 427 | Polyhalogenated Hydrocarbon | D. pulchra | Cape Banks, New South Wales | [167,169,170] |
| P. australasica | Tasmania | |||
| 428–430 | Polyhalogenated Hydrocarbon | P. australasica | Tasmania | [167] |
| 431 | Polyhalogenated Hydrocarbon | D. pulchra | Cape Banks, New South Wales | [169,170] |
| 432 | Polyhalogenated Hydrocarbon | P. australasica | Pearsons Point, Tasmania | [168] |
| 433 | Polyhalogenated Hydrocarbon | P. australasica | Tasmania | [167] |
| 434 | Polyhalogenated Hydrocarbon | D. pulchra | Cape Banks, New South Wales | [169] |
| 435 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [169] |
| 436 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [170,172] |
| Palmer Station, Antarctica | ||||
| 437, 438 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [169,170] |
| 439–442 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [170,171] |
| 443 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [170,171,172] |
| Palmer Station, Antarctica | ||||
| 444–448 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [170,171] |
| 449 | Polyhalogenated Furanones | D. pulchra | New South Wales | [174] |
| 450 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [169] |
| 451–462 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [171,175] |
| 463–465 | Polyhalogenated Furanones | D. pulchra | Palmer Station, Antarctica | [172] |
| 466 | Polyhalogenated Furanones | D. pulchra | Cape Banks, New South Wales | [169] |
| 467, 468 | Steroid | A. armata | - | [176] |
| 469–472 | Steroid | A. armata | Praia do Quebrado, Portugal | [43] |
| 473 | Steroid | A. armata | Portugal | [177] |
| No. | Compound Type | Species | Origin | Ref |
|---|---|---|---|---|
| 474 | Bromophenolic | C. officinale | Bateau Bay, NSW | [54] |
| P. lucida | Bateau Bay, NSW | |||
| G. secundada | Batemans Bay, NSW | |||
| A. anceps | Bateau Bay, NSW | |||
| J. sagittata | Bateau Bay, NSW | |||
| D. pulchra | Botany Bay, NSW | |||
| S. robusta | Batemans Bay, NSW | |||
| 475–478 | Bromophenolic | C. officinale | Bateau Bay, NSW | [54] |
| P. angustum | Bateau Bay, NSW | |||
| P. lucida | Batemans Bay, NSW | |||
| G. secundada | Batemans Bay, NSW | |||
| A. anceps | Bateau Bay, NSW | |||
| J. sagittata | Bateau Bay, NSW | |||
| D. pulchra | Botany Bay, NSW | |||
| S. robusta | Batemans Bay, NSW | |||
| 479 | Xanthophyll | L. filiformis | Australia | [185] |
| L. botryoides | Australia | |||
| 480 | Xanthophyll | L. botryoides | Australia | [185] |
| M. abscissa | Leigh, New Zealand | |||
| 481, 482 | Xanthophyll | A. ciliolatum | Ensenada, Baja, California | [186] |
| C. clavulatum | Ensenada, Baja, California | |||
| P. capillacea | Ensenada, Baja, California | |||
| 483 | Xanthophyll | L. botryoides | Australia | [185] |
| 484 | Xanthophyll | L. botryoides | Australia | [186] |
| A. ciliolatum | Ensenada, Baja, California | |||
| C. clavulatum | Ensenada, Baja, California | |||
| M. abscissa | Leigh, New Zealand | |||
| P. capillacea | Ensenada, Baja, California | |||
| 485 | Aromadendrene | L. filiformis | South Australia | [127] |
| 486, 488 | γ−Pyrones | P. peperocarpos | South Australia | [178] |
| 489 | Lipid | L. filiformis | Taroona Beach, Hobart, Tasmania | [138] |
| 490 | γ−Pyrones | P. australasica | Pearsons Point, Tasmania | [168] |
| 491, 492 | γ−Pyrones | P. australasica | - | [167] |
| 493, 494 | Pyrogallols | G. filicina | Bay of Hiroshima, Japan | [181] |
| 495 | Cyclic lipid | G. filicina | Bay of Hiroshima, Japan | [182] |
| 496 | Xanthophyll | C. clavulatum | Ensenada, Baja, California | [186] |
| M. abscissa | Leigh, New Zealand | |||
| P. capillacea | Ensenada, Baja, California | |||
| 497 | Cyclic lipid | C. clavulatum | East Coast of Sicily, Italy | [187] |
| 498 | Xanthophyll | C. clavulatum | Ensenada, Baja, California | [186,188] |
| M. abscissa | Leigh, New Zealand | |||
| 499 | Xanthophyll | M. abscissa | Leigh, New Zealand | [188] |
| 500 | Diterpene | P. costatum | Deep Glen Bay, Tasmanina | [142] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lever, J.; Brkljača, R.; Kraft, G.; Urban, S. Natural Products of Marine Macroalgae from South Eastern Australia, with Emphasis on the Port Phillip Bay and Heads Regions of Victoria. Mar. Drugs 2020, 18, 142. https://doi.org/10.3390/md18030142
Lever J, Brkljača R, Kraft G, Urban S. Natural Products of Marine Macroalgae from South Eastern Australia, with Emphasis on the Port Phillip Bay and Heads Regions of Victoria. Marine Drugs. 2020; 18(3):142. https://doi.org/10.3390/md18030142
Chicago/Turabian StyleLever, James, Robert Brkljača, Gerald Kraft, and Sylvia Urban. 2020. "Natural Products of Marine Macroalgae from South Eastern Australia, with Emphasis on the Port Phillip Bay and Heads Regions of Victoria" Marine Drugs 18, no. 3: 142. https://doi.org/10.3390/md18030142