Neurotoxins Acting at Synaptic Sites: A Brief Review on Mechanisms and Clinical Applications

1
Department of Pharmacology and Laboratory of Aging and Nervous Diseases, College of Pharmaceutical Sciences, Suzhou Medical College of Soochow University, Suzhou 215123, China
2
Department of Neurology and Clinical Research Center of Neurological Disease, The Second Affiliated Hospital of Soochow University, Suzhou 215004, China
3
Institute of Pain Medicine and Special Environmental Medicine, Nantong University, Nantong 226019, China
*
Author to whom correspondence should be addressed.
Toxins 2023, 15(1), 18; https://doi.org/10.3390/toxins15010018
Submission received: 30 October 2022
/
Revised: 20 December 2022
/
Accepted: 21 December 2022
/
Published: 27 December 2022
(This article belongs to the Special Issue Animal Venom: Challenges and Perspectives in Drug Discovery)
Abstract
:Neurotoxins generally inhibit or promote the release of neurotransmitters or bind to receptors that are located in the pre- or post-synaptic membranes, thereby affecting physiological functions of synapses and affecting biological processes. With more and more research on the toxins of various origins, many neurotoxins are now widely used in clinical treatment and have demonstrated good therapeutic outcomes. This review summarizes the structural properties and potential pharmacological effects of neurotoxins acting on different components of the synapse, as well as their important clinical applications, thus could be a useful reference for researchers and clinicians in the study of neurotoxins.
Key Contribution: Summarized basic biological characteristics of neurotoxins, new developments in understanding the mechanisms of their pharmacological actions, and their current clinical applications with critical comments on their unique benefits and limitations.
1. Introduction
There are thousands of biological species that can produce toxins, and most of them are neurotoxins [1]. Over millions of years of evolution, biological toxins have acquired specific selectivity to interfere with some physiological functions and disrupt a large number of basic neurobiological processes such as synaptic transmission [2].
At the neuromuscular junction (NMJ), nerve signals from spinal motor neurons are transmitted to the muscles via the release of synaptic acetylcholine (Ach), which causes muscle contraction. Structurally, there are three main components of the NMJ: the presynaptic nerve terminals, the synaptic cleft, and the postsynaptic receptors, mainly the dense cluster of nicotinic acetylcholine receptors (nAChRs) [3]. Ach-containing synaptic vesicles at presynaptic nerve terminals initiate the process of coalescing with the presynaptic membrane; then, the nerve terminal depolarizes, leading Ca2+ to flow in through voltage-gated channels. As a result of the influx of Ca2+, the vesicles begin to fuse with the membrane surface. Subsequently, Ach is released to the synaptic cleft from vesicles [4]. The Ca2+-mediated exocytosis of Ach described above primarily depends on the SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor) complex assembly [5], which generally comprises syntaxin, VAMP, and SNAP-25 homologs. The AChR is a type of ligand-gated ion channel [6] formed by five subunits, and it has several subtypes and these subtypes are assembed of 17 different subunits (α1–α10, β1–β4, γ, δ, and ε) [7]. The different AchRs are expressed in specific regions of the brain and peripheral tissues of mammals [8]. The AchR was first identified as a classic neurotransmitter receptor [9], and recently there is also evidence showing that the nAChR exists in non-neuronal cells, including certain types of tumor cells. When it is activated, it promotes the development of tumors by inducing the release of autocrine growth factors [10].
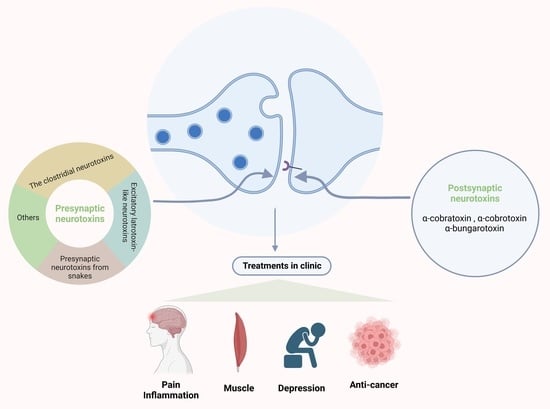
Generally, the release of ACh and the process of Ach binding to the AchR are two of the main targets of neurotoxins [4]. Many neurotoxins are presynaptic neurotoxins that act at the presynapse and specifically bind to ion channels to regulate neurotransmitter release or to block the neuronal transmission of signals [1]. According to this feature, presynaptic neurotoxins can be roughly divided into four categories: (1) clostridial neurotoxins that block the neurotransmitter function by their specific metalloproteolytic activity aiming at SNARE proteins; (2) presynaptic neurotoxins in snake venoms with phospholipase A2 activity; (3) excitatory latrotoxin-like neurotoxins [11]; and (4) neurotoxins that function via various ion channels (such as calcium channels, potassium channels, etc.) [12]. There are also a variety of neurotoxins that have postsynaptic effects. They bind to acetylcholine receptors, acetylcholinesterase, or ion channels, thereby altering cholinergic functions [1]. There are a large class of snake neurotoxins defined as postsynaptic neurotoxins, collectively called “α-neurotoxins” [13]. All members of the α-neurotoxin family show high similarity in the mechanism of neuromuscular paralysis, in which they primarily target and then bind the postsynaptic nAChRs, specifically at the NMJ [14,15,16].
This review aims to discuss the structures and mechanisms of action of the presynaptic and postsynaptic neurotoxins and to provide a summary of recent developments in clinical applications of these neurotoxins. This review might be considered as a first-hand reference for experts and clinicians interested in translating these neurotoxins into clinical therapies and uncovering the underlying mechanisms associated with clinical efficacy. Critical comments on the prospects for clinical application of these neurotoxins in diverse diseases are also provided.
2. Presynaptic Neurotoxins
2.1. The Clostridial Neurotoxins
The anaerobic bacteria that belong to the clostridia class are a vital threat to human health, causing a variety of disorders such as food poisoning and, even worse, gas gangrene [17]. Above all, two main categories of clostridial neurotoxins are considered as the most potent toxins that we have known: botulinum neurotoxins (BoNTs) and tetanus toxins (TeNTs) [18]. These two types of toxins have a similar molecular structure: 150 kDa molecular size with three main functional domains [18], the light chain (LC, 50 kDa) and the heavy chain (HC, ~100 kDa) with two domains. The LC and HC are connected by a disulfide bond. Both BoNT and TeNT act in a similar way: (i) binding to the presynaptic membrane; (ii) internalization; (iii) membrane translocation and release of the LC into the cytosol; and (iv) cleavage of SNAREs driven by the LC [19]. The HC will form a channel to help translocation of the LC into the cytosol [20]. The C-terminal part of the HC mediates the interaction of the toxins with neurons [21] via a neurospecific double binding to the polysialoganglioside and the glycosylated lumenal domain of a synaptic vesicle protein, which then leads to the internalization of the toxin [18,19]. The function of the N-terminal part of the HC is not fully understood, although some studies revealed that it protects the LC from the cutting of non-specific substrates until the LC is localized within the cytosol [22,23]. The LC is a metalloprotease [24], which has selectivity for SNARE proteins and can cut SNARE at different peptide bonds [25]. This specific structure is the basis for the action of BoNT and TeNT [26]. Both toxins can target and enter the nerve terminals at NMJs, then change the conformation to enable translocation of the LC into the cytosol. In there, the release of the neurotransmitters is disturbed by the LC via cleaving SNARE proteins [27,28,29] (Figure 1).
The neuroselectivity of BoNT and TeNT is probably due to the following aspects: (1) The C-terminal part of the HC mediates the interaction of the toxins with their receptors, and the receptors are mainly enriched in the neuronal terminals. (2) The receptor-mediated endocytosis makes them enter the neuronal cells, but BoNT and TeNT enter in different endocytic vesicles. (3) SNAREs, the target molecules of BoNT and TeNT, are expressed in the neuronal cells of almost all vertebrate phylla [30,31,32]. However, the neuroselectivity of BoNT is not absolute and it can act on the non-neuronal cells to exert some functions, such as glial cells [33].
Up to now, there are eight BoNT serotypes that have been identified and named as A–G and X. They are classified due to lack of cross-neutralization by different antisera against each toxin type [34,35]. Over 40 subtypes have been identified [36]. They bind to different receptors to drive the process of internalization [19]. The flaccid paralysis induced by BoNTs occurs primarily due to the blockade of peripheral cholinergic nerve endings [21,37,38,39], whereas TeNT, produced by a Gram-positive bacillus, Clostridium tetani [40], has only one type [18]. The release of neurotransmitters, such as GABA and glycine, is blocked by TeNT which can lead to spastic paralysis [41,42,43]. With prolonged action, it causes death when muscular hypertonus occurs in the respiratory muscle and leads to breathing failure [44].
The two clostridium neurotoxins act in a similar way, but they cause very different diseases. The reason for this is that TeNT travels retroaxonally and is transferred via a trans-synaptic movement to inhibitory interneurons in the CNS to block the release of neurotransmitters, which results in motor neuron hyperactivity and spastic paralysis. However, BoNT mainly acts on the NMJs to inhibit the release of acetylcholine and then induce flaccid paralysis [18,41,45]. Specifically, TeNT moves retroaxonally along the axons of motor neurons into the cell body, releases and thereby enters the connecting inhibitor neurons, and then the LC exerts the function of blocking neurotransmitter release [46]. TeNT can bind to not only the connecting inhibitor neurons but also the dendrites of sensory and adrenergic neurons [41].
2.2. Excitatory Latrotoxin-like Neurotoxins
There are high-molecular neurotoxins extracted from the venom of black widow spiders called latrotoxin-like neurotoxins (LaTXs). They consist of various specific types: one vertebrate-specific toxin (α-latrotoxin (α-LTX)) [47], five highly specific insecticidal toxins (α-, β-, γ-, δ-, and ε-latroinsectotoxin (LITs)) [48], and one crustacean-specific toxin (α-latrocrustatoxin (α-LCT)) [49]. LaTXs are secreted into the gland lumen as 160 kDa inactive precursor polypeptides. In the gland lumen, the N-terminal signal peptide and a C-terminal inhibitory domain are cleaved and proteolyzed, which produces the final mature 130 kDa toxin [39,40].
Among the above-mentioned toxins, there are a number of studies on α-LTX. α-LTX causes a syndrome called lactrodectism in the clinic, which has the feature of serious muscle spasm and lots of other effects, for example, hypertension, sweating, and vomiting [50,51]. The α-LTX also affects the process of exocytosis and has a high affinity for three types of receptors: cell adhesion protein neurexin (NRX) [52,53,54]; G-protein-coupled receptor latrophilin (LPHN or CIRL) [55,56]; and the receptor-like protein tyrosine phosphatase σ (PTPσ) [57]. The α-LTX initiates the release of neurotransmitters by two distinct mechanisms, both of them relying on the binding of the toxin to three types of receptors [58,59,60]: (1) in a Ca2+-dependent manner: α-LTX binds to the cell adhesion protein neurexin in the presence of Ca2+ and then inserts into the plasma membrane to form the pore and thereby induces the influx of Ca2+ [61], and (2) in a Ca2+-independent manner: it binds to the other two receptors without Ca2+. Furthermore, LPHN may mediate the process of stimulating PLC, producing IP3 and diacyl glycerol, releasing the stored Ca2+, and activating PKC. This cascade promotes the release of neurotransmitters form vesicles [60,62] (Figure 1).
2.3. Presynaptic Neurotoxins from Snakes
Most snake venoms contain both pre- and postsynaptic neurotoxins [63], whereas some snake venoms contain only presynaptic neurotoxins [64]. These presynaptic neurotoxins belong to phospholipases A2 (PLA2), which are Ca2+-dependent enzymes [63]. They can hydrolyze the sn-2 ester bond of 1,2-diacyl-3-sn-phosphoglycerides to produce fatty acids and lysophospholipids [11,65]. Various snake presynaptic PLA2 neurotoxins have a similar secondary structure with three larger α-helices and a short two-stranded β-sheet [66,67]. The pharmacological effects of these neurotoxins are also attributed to their PLA2 enzymatic activity, including antibacterial, cardiotoxic, and neurotoxic actions [68,69,70,71,72,73,74,75].
The studies on PLA2 snake neurotoxins are extensive, and the neurotoxicity induced by them can be attributed to the inhibition of presynaptic neuromuscular transmission [76]. Snake presynaptic neurotoxins exert a function that blocks the release of ACh in the nerve terminals [63] because they can strongly reduce the frequency of spontaneous exocytosis that releases ACh from small synaptic vesicles (SSVs) but does not affect the amount of ACh in one single SSV [65]. The electron microscopic studies of NMJ affected by snake presynaptic neurotoxins revealed: (1) swollen and enlarged axon terminals; and (2) the emergence of lots of Ω-shaped plasma membrane invaginations [77,78,79,80,81]. Recently, a PLA2 neurotoxin MiDCA1 extracted from the venom of Micrurus dumerilii carinicauda coral snake [82,83] has been found to affect the process of the release of neurotransmitters [82] through targeting the Kv2 channels [84]. Thus, it was proposed that MiDCA1 blocks the Kv2.1 channels and thereby decreases the release of Ach [84].
2.4. Other Presynaptic Neurotoxins Acting on Ion Channels
The release of neurotransmitters, which happens in a Ca2+-dependent manner [85], relies on the voltage-gated Ca2+ (Cav) channels (VGCCs) that have selective permeability to Ca2+ and can drive calcium influx [86,87]. Among all types of VGCCs, Cav2 channels mainly regulate exocytosis. Cav2 channels contain Cav2.1(P/Q-type), Cav2.2 (N-type), and Cav2.3 (R-type). Furthermore, the release of neurotransmitters at ribbon synapses in the retina and inner ear mainly relies on Cav1 channels (L-type) [88]. In addition to the above-mentioned types of presynaptic neurotoxins, there are some presynaptic neurotoxins that act on ion channels, such as agatoxins, conotoxins, and dendrotoxins.
Agatoxins are a class of toxins extracted from the American funnel web spider A. aperta that can target different types of ion channels. They have various components that are divided into three classes: α-Agatoxins, µ-Agatoxins, and ω-Agatoxins, according to their functions on different channels [89,90]. Among these, ω-Agatoxins are particularly special because they are selective for different subtypes of calcium channels [89,91]. The ω-Agatoxin IA, which has five disulfide bonds [92], appears to be selective for insect calcium channels; the ω-Agatoxin IIA with three disulfide bonds disturbs both mammalian and invertebrate calcium channels [93]; ω-Agatoxin IIIA with six disulfide bonds [93] acts as a pore blocker due to the poor selectivity among the family of high-voltage-activated channels [94,95]; and ω-Agatoxin IVA exerts functions as a gating modulator via interfering with the domains of the voltage sensor [96,97,98].
Conotoxins are peptides with distinct cysteine frameworks, and they mainly target various ion channels and receptors [99], such as G-protein-coupled receptors (GPCRs), transporters, and enzymes [100,101]. Among the various subgroups of conotoxins, ω-conotoxins, consisting of 24–30 amino acids [102], physically block the pore of the channel to prevent calcium influx [98,103,104]. The two most characterized ω-conotoxins are GVIA [105] and MVIIA [106], which can selectively inhibit the N-type VGCC (neuron-type voltage-gated calcium channel). Furthermore, the selectivity of MVIIA on the channel is weaker than that of GVIA, but it can dissociate quicker due to the weaker selectivity [107,108], and therefore it was developed as a therapeutic peptide ziconotide (Prialt®) for pain treatment [90].
3. Postsynaptic Neurotoxins
The postsynaptic neurotoxins have postsynaptic blocking functions at the skeletal muscle end-plate and neurons, named α-neurotoxins [15]. They act as antagonists of nAChRs in the NMJs and neurons [109]. The α-neurotoxin family has a similar mechanism of the function that causes muscular paralysis due to blockade of the postsynaptic nAChRs at the NMJ [110,111,112].
The α-neurotoxins in snake venoms belong to the three-finger toxins [113]. These kinds of toxins are flat molecules, and the reason for the name of “three-finger” is that the flat molecules have a small globular hydrophobic core, and this core can form three adjacent loops, with the shape of three fingers [111,114,115]. The three-finger toxins are mainly generated form Elapid snakes, including α-cobratoxin (α-Cbtx), α-cobrotoxin (α-Cotx), and α-bungarotoxin (α-Bgtx) [116]. In addition, based on their molecular structures, α-neurotoxins are roughly divided into three categories: short-chain toxins with 60–62 amino acids residues and four disulphide bridges (α-Cotx) [117]; long-chain toxins with 66–75 amino acid residues and five disulphide bridges (α-Cbtx) [118]; and weak toxins, which have the feature of hypotoxicity (LD50 ~ 5–80 mg/kg as compared to LD50 ~ 0.04–0.3 mg/kg for other toxins) [103]. What they have in common is that they all target muscle α1 nAChRs, but only long-chain α-neurotoxins can bind to the α7 nAChR with high affinity [109,119,120], while only the dimmer of cobrotoxin binds to it. As we know, the α7-type nAChR is considered as a potentially essential target for the treatment of many diseases in the clinic [121], such as cognitive disorders [122,123], inflammatory diseases, and chronic pain [124]. Alzheimer‘s disease, Parkinson’s disease, Lewy body dementia, and schizophrenia all have the feature of neuroinflammation [125,126,127,128,129,130,131,132,133]. At present, some researchers have started to explore the effect of α-neurotoxins on central nervous system diseases [134,135,136,137,138]. Among these toxins, the α-bungarotoxin (αBgt) is the most popular one as a pharmacological tool used for studying α7 nAChR [121] because of the practically irreversible binding to the Torpedo acetylcholine receptors [139].
In addition to snake neurotoxins, α-conotoxins generated from marine organisms of the genus Colocasia also target nAChRs [140]. There are many subgroups of α-conotoxins that are selective for different subtypes of AChRs, such as α3/5-conotoxins that selectively block the muscle nAChR, and α4/3-, α4/4-, α4/5-, α4/6-, and α4/7-conotoxins that selectively block the neuronal nAChRs [140,141,142] (Figure 2).
All the above-mentioned neurotoxins that are classified into presynaptic and postsynaptic neurotoxins, even though they are from different sources, have many types, and their complex subtypes and structures make it difficult to generalize with a symbolic model that they have a similar principle of function in the intervention of the neuromuscular signal transmission process, in which they affect the transmission of neurotransmitters at synaptic sites by regulating ions, receptors, and membrane fusion proteins and in other ways.
Besides those mentioned above, there are still some toxins not marked as “neurotoxins” that exert the same action on the nervous system. For example, the epsilon toxin (ET), produced by clostridium perfringens types B and D, can cause enterotoxaemia in sheep, goats, and cattle. When it enters the brain, it targets the neural cells and also induces the release of glutamate and other transmitters; but the specific mechanism needs more exploration [143]. Some bacterial enterotoxins, such as toxin B (TcdB) and cholera toxin (CT), can attack the enteric nervous/endocrine system. There is evidence that CT activates a secretomotor neural reflex to further enhance the secretory responses; and TcdB can catalyze the glucosylation of Rho proteins (Rho, Rac, and Cdc42) to block neurotransmitter exocytosis [30,144,145,146,147].
4. Clinical Applications of Neurotoxins
For the clinical applications of neurotoxins, besides their pharmacological effects, we also need to consider their half-life in vivo and their distributions in our body to exert their clinical effects. For most neurotoxins, they are limited to the injection compartment and cannot cross the barrier due to their peptide structure [148]. However, there are some special toxins that can break through this limitation, such as apamin (a peptide found in bee venom), which has the capacity to cross the blood–brain barrier [149]. Moreover, (1) toxins impairing the blood–brain barrier, such as clostridium perfringens epsilon toxin, can cross the barrier [143]; and (2) neurotoxins with retrograde axonal transport can reach the target, such as TeNT, mentioned above [46]. Because of this feature, they can be a more effective treatment in pain management [150].
4.1. Clostridial Neurotoxins
As we can see from Table 1, only BoNT/A and BoNT/B have been officially approved. The use of BoNT/B is limited due to its higher incidence of adverse effects than that of BoNT/A [151,152]. In addition, it does not have such long-lasting effects as BoNT/A [153]. The current treatments with BoNT are mainly used in the area of dystonic muscle contractions, which depends on the inhibition of the excessive release of the ACh neurotransmitter [154]. Recently, several articles also revealed that for some other diseases, such as chronic pain that is associated with a variety of neurological disorders (trigeminal neuralgia), neuroinflammation [155,156], depression [157], and skin diseases [158], BoNT can also play a therapeutic role. However, the therapeutic effects of BoNT mentioned above cannot be simply explained by the mechanism of NMJ blockade and needs more exploration. It has a long half-life in vivo, and one of the factors responsible for this phenomenon is that the catalytic light chain escapes from the proteasomal degradation by binding to the deubiquitinating enzyme, VCIP135/VCPIP1, and therefore remains enzymatically active for months [159,160]. This explains the clinical phenomenon that after the injection of BoNT in the detrusor muscle, the patients with neurogenic urinary incontinence do not need additional injections for 36 weeks and side effects are not observed [161,162]. For its distribution, sometimes it needs to cross barriers to reach the target area, and there is evidence that it can travel in a way similar to TeNT [18,163]. Therefore, it can influence the ascending pain processing pathway [150].
Owing to the technical advances in recombinant DNA technology and purification techniques of recombinant proteins, more and more engineered BoNTs are produced for clinical uses [184]. The indications of them are classified into the following categories based on the inhibition of different transmitters or other unclear mechanisms. For the inhibition of Ach release, here are the indications:
4.1.1. Dystonic Muscle Contractions
From a translational perspective, when the nerve–muscle impulse is inhibited, NMJ function can be restored after BoNT treatment, which provides a scientific basis for BoNT in the treatment of various human diseases characterized by hyperfunction of cholinergic activity [1]. The initial clinical application was the treatment of benign essential blepharospasm by Scott and his coworkers [185]. Therefore, BoNT was first applied in the clinic for patients with dystonia and had a remarkable benefit, and it is now still considered as a choice of treatment for patients with problems of focal or segmental dystonia, including blepharospasm, oromandibular dystonia, spastic dysphonia, and so on. Furthermore, the treatment of hemifacial spasm and primary dystonia, such as cervical dystonia, is another application of BoNT in the clinic. What is more, BoNT can also be used for the treatment of some occupational dystonias, such as writer’s and musician’s cramps and stroke-related hemiplegia [186]. For laryngeal dystonia, characterized by spasmodic dysphonia, which manifests as either a laborious and tense sound that is interrupted by frequent articulatory interruptions and silent pauses, or a respiratory murmur, BoNT is also considered the treatment choice [187]. As for gastrointestinal, genitourinary, and sphincter disorders (such as achalasia, spasm of the sphincter of Oddi, and anal fissure [186]), the therapeutic efficacy of BoNT also acts on the spasm muscles in these organs [162,188,189,190,191]. Generally speaking, BoNT mainly leads to the alleviation of dystonic muscle contractions.
BoNT also interferes with transmission located at the cholinergic autonomic parasympathetic and postganglionic sympathetic nervous system, and therefore this toxin has been widely used for the diseases of the autonomic nervous system [192], such as essential focal hyperhidrosis, which is characterized by excessive sweating of the palms, feet, or axillae, and it is related to secretomotor hyperactivity [192]. After the treatment with BoNT, the patients’ quality of life is significantly improved due to a significant improvement in the symptoms [193,194,195]. The current clinical applications of botulinum toxin are mainly concentrated in the field of aesthetic medicine, such as the glabellar frown lines [196] and aging neck (hypertrophic platysma muscle bands) [197].
4.1.2. Skin Diseases
For skin diseases, BoNTs have some label and off-label applications [158]. In sweat gland disorders (idiopathic hyperhidrosis, chromhidrosis, and bromhidrosis), the patients mainly suffer from excessive sweating in one or more parts of the body. The BoNT injection can decrease sweat secretion [198] by preventing the release of Ach and some other neurotransmitters from presynaptic vesicles [199]. In alopecia (alopecia areata and androgenetic alopecia), a study showed that BoNT downregulated the expression of TGF-β, thus suppressed follicular epithelial cell growth [200].
4.1.3. Neuropathic Pain and Neuroinflammation
BoNT can inhibit not only the release of Ach but also other neurotransmitters [32]. For several types of neuropathic pain such as trigeminal, posttraumatic, or postherpetic neuralgia, significant analgesic effects have been observed after administration of botulinum toxin A (BoNT/A) [165,201,202]. It is speculated that BoNT/A exerts its therapeutic effect by inhibiting the process of the secretion of some pain mediators (substance P, glutamate, and calcitonin-gene-related protein (CGRP)) and other pain transmitters released from the nerve terminals and dorsal root ganglions (DRGs) and trigeminal sensory neurons [165,203,204,205]. There is evidence that after peripheral administration of BoNT/A, the antinociceptive action is not primarily mediated by the direct prevention of central CGRP release. This indicates that it might depend on the toxin’s axonal transport [206].
Research revealed that intraplantar administration of BoNT/A alleviated mechanical and thermal hypersensitivity and the activation of microglia induced by chronic constriction nerve injury in the ipsilateral lumbar spinal cord in rats [207], decreased the amount of the pro-inflammatory cytokines IL-1β and IL-18, and increased the level of IL-1 receptor antagonist and IL-10 in the spinal cord and/or the DRG [167]. Recently, an in vitro study indicated that in primary rat microglia the expression of pro-inflammatory IL-1β, IL-18, IL-6, and nitric oxide synthase 2 (NOS2) was inhibited by BoNT/A, and the intracellular signaling pathways mediated by p38, ERK1/2, and NF-κB were also inhibited. Additionally, the expression of SNAP-23 was decreased, whereas the expression of TLR2 was increased [208].
4.1.4. Depression
The first article that reported the antidepressant effect of BoNT was in 2006. It was found that the self-rated depression scores on the Beck Depression Inventory (BDI) II were significantly improved after 8 weeks in ten middle-aged women that had moderate-to-severe, partly chronic, and treatment-resistant depression when they received one single application of BoNT administration in the glabella [209]. Subsequently, several randomized controlled trials were conducted, and they confirmed the efficacy of BoNT for the treatment of depression [210,211,212,213,214]. With more and more evidence showing that BoNT can be used for the treatment of depression, it is important to find the underlying mechanisms of this action. So far, several possible mechanisms have been proposed. Firstly, there is a feedback loop from the face to the brain, called “emotional proprioception”, that can reinforce and maintain the negative emotions. When BoNT/A disturbs it, depression can be ameliorated [215]; Secondly, in rat depression models, there is evidence that after the administration of BoNT in the face, the metabolism of monoaminergic neurotransmitters is changed in the limbic brain regions [216]. Thirdly, some experimental evidence shows that high-dose [217], locally injected BoNT can enter the central nervous system (CNS), possibly through retrotransportation, which might be another mechanism for BoNT in regulating mood [218]. Moreover, some substances might be related to the mechanism of BoNT for antidepression, such as the RAS-related C3 botulinum toxin substrate 1 (Rac1). This is the central nervous substrate of BoNT [219]. Emerging evidence has also shown that the expression of BDNF diminishes in animal models of depression and depressed patients [220,221,222]. BDNF is essential for neurogenesis and the reduction in depression-like behaviors [223]. It promotes the phosphorylation of CREB through ERK activation [224]. A recent article has shown that the expression of BDNF at both the mRNA and protein level was up-regulated in the hippocampus by BoNT/A, and therefore the downstream ERK-CREB signaling pathway was activated in depression mice models [168].
4.1.5. Headache
Headache is a common neurological disorder. According to the international classification of headache disorder (ICHD-3), it is classified into primary headaches and secondary headaches. Primary headaches include migraine, trigeminal autonomic cephalalgia (TAC), and tension-type headache (TTH). Secondary headaches include neuropathies, facial pains, and other forms of headache [225]. They all share the common pathophysiology of the abnormal activation of the trigeminovascular system [226], which leads to vasodilation and neurogenic inflammation and pain sensitization in the peripheral and central nervous system, resulting in the persistent headache [166]. The onabotulinum toxin A, as a BoNT formulation, has been used for the treatment of chronic migraine [169]. Several articles suggested that BoNT/A could suppress the release of neuropeptides and neurotransmitters that contain CGRP [227], substance P [228], and glutamate from sensory peptidergic sensory neurons [229]. In addition, BoNT/A can also interfere with the pain-sensing receptors expressed on the plasma membrane, including transient receptor potential cation channel vanilloid subfamily member 1 (TRPV1), transient receptor potential cation channel ankyrin subfamily member 1 (TRPA1), ATP-gated P2X receptor cation channel family 3 (P2X3) [230], and AMPA receptor [231,232,233,234]. All mechanisms mentioned above may contribute to the antinociceptive action of BoNT/A.
As for TeNT, it is the only known toxin that has the potential for selective improvement of motor functions, so the flabby and weak muscles induced by the injury of the brain and spinal cord can be treated with TeNT [170,235,236]. Due to the presence of the blood–brain barrier [2], it makes it difficult for some drugs to enter the brain. Another clinically relevant area of TeNT has generated great interest, which is its potential as a fusion protein or carrier to deliver drugs into the CNS. TTFC (a segment with 50 kDa from the carboxy-terminal HC of TeNT) can exert the function of delivery by interacting with neural gangliosides and specific proteins linked with lipid microdomains of the neuronal surface [171,237,238,239,240,241] and then moving retroaxonally to the CNS [242,243,244]. It can bind to polysialylgangliosides GD1b and GT1b and then is internalized by motor neurons at the NMJ. Finally, it reaches and influences the CNS via traveling retroaxonally [245,246]. Some potential therapeutic molecules, such as cardiotrophin-1 (CT1) and Bcl-xL, have been successfully transported into neurons by coupling with TTFC [245]. Furthermore, when coupling with these molecules, TTFC still keeps its capacity of neuronal binding [246]. However, their effects as therapeutic agents will need more studies in vivo.
Moreover, TTFC has also been demonstrated to have neuroprotective activities [246]. In the amyotrophic lateral sclerosis mice model, TIFC can modulate the levels of NLRP3 and caspase-1 in the spinal cord and reduce the level of IL-6 in tissues [247]. In the models of Parkinson’s disease (PD) and Alzheimer’s disease (AD), it can be used for neuronal dysfunction, learning impairment, and memory impairment [248,249]. TTFC can protect against apoptosis via activating the MAPK/ERK pathway to inhibit the disruption of the dopaminergic neurons caused by MPP+ [245].
There are several engineered BoNTs; here are some of them: (1) the Erythrina cristagalli lectin replaces the C-terminal of the HC of BoNT, and thereby forms a novel conjugate. It binds to eDRG neuronal cell types and inhibits the neurotransmitter release [250]. After injecting it into the intrathecal space of a mouse, it was demonstrated to have a long-lasting analgesic effect [251]; (2) the botulinum construct (Bitox), synthesized by “stapling” the recombinant LC/translocation domain of BoNT/A and receptor-binding domain has the potential to treat pain and does not result in muscle paralysis [252,253]. The recombinant LC/translocation domain of BoNT/A can bind to the TeNT receptor-binding domain [254]; (3) the chimeras of BoNT/A and BoNT/E can significantly reduce the acute nociception induced by capsaicin [255].
4.2. LaTXs
α-latrotoxin is an inducer of Ca2+ influx and is expected to ameliorate dysfunctions and diseases that are associated with the reduction in the release of transmitters and hormones. It is also expected to treat metabolic diseases, such as type I diabetes [172,256]. The structure of α-latrotoxin is homologous to glycogen-like peptide-1 (GLP1) an endogenous peptide, and it can control the level of blood glucose by binding to GLP1 receptors in the cell membrane of pancreatic β-cells to increase the release of insulin. Therefore, α-latrotoxin has been used as a potential drug for the treatment of obesity, diabetes, and other related metabolic disorders [257,258,259].
4.3. Snake Presynaptic Neurotoxins
4.3.1. Anticancer
Snake presynaptic neurotoxins have PLA2 activity. PLA2 has been reported to have anticancer properties, which involve inhibiting angiogenesis, migration, and adhesion of various kinds of cancer cells [260]. For example, crotoxin extracted from the venom of South American rattlesnake has been found to have potential anticancer effects in several types of cancers [173]. Several experiments demonstrated that crotoxin arrested the cell cycle at the G2/M phase [261]. Furthermore, recently it has been reported that crotoxin is also a potential regulator of the signaling cascade involved in epithelial–mesenchymal transition (EMT) [262]. Crotoxin may have a high affinity for EGFR [263], which suggests that crotoxin might modulate the EGFR signaling pathway to exert its anti-tumor activity in SPCA-1 cells [264]. The research on lung cancer has attracted people’s attention because it is the leading cause of cancer-related mortality worldwide [265]. In human lung squamous carcinoma SK-MES-1 cells, crotoxin can induce apoptosis and autophagy through the p38MAPK signaling pathway in vitro [266]. Furthermore, in the process of treating cancer with crotoxin, a phase I clinical trial revealed that besides its anticancer effect, analgesic effects were also observed in terminal cancer patients [267]. In some other animal models, such as acute pain models, studies have demonstrated that crotoxin also showed the effect of antinociception [268] without the involvement of opioid receptors [175,269], instead of the central cholinergic, serotonergic, and noradrenergic systems [270].
4.3.2. Antibacterial
In addition to the anticancer and analgesia effects, crotoxin also exerts an antibacterial effect because of the change in membrane permeability induced by PLA2, which destroys the integrity of the bacterial cell membrane [271].
4.4. Other Presynaptic Neurotoxins Acting on Ion Channels
4.4.1. Analgesia
For presynaptic neurotoxins targeting calcium and potassium channels, the clinical applications mainly depend on their ability to block or activate the channels. For example, voltage-gated calcium channels are important for the process that conducts pain signals from the periphery into the dorsal horn of the spinal cord. Some conditions of pain that are difficult to treat with clinically available drugs, such as cancer and neuropathic pain, can be treated effectively with crotoxin [272].
Due to the selective blockage of the Ca2+ channel by ω-agatoxin, it may be potentially useful in clinical applications to treat a variety of disorders. Evidence from behavioral and electrophysiological reports suggests that ω-agatoxin IVA-sensitive P-type channels significantly modulate spinal nociceptive processes [176,273]. A study reported that intrathecal administration of ω-agatoxin IVA reduced late nociceptive behaviors induced by formalin [274].
Conotoxins have been widely used in basic neuroscience research to analyze neuronal voltage-gated Ca2+ currents, as well as in treatments in the clinic because of their powerful and pervasive ability to block Ca2+ channels. Conotoxins have the potential to become popular drugs [275,276] because ω-conotoxins specifically target presynaptic voltage-gated Ca2+ channels (VGCC), particularly N-type VGCCs, that have been proved to be involved in pain pathways, making ω-conotoxins potential analgesics [273]. Prialt™ (a form of the ω-conotoxin MVIIA) is a drug that has been approved by the FDA to treat the chronic pain that results from cancer- or AIDS-related neuropathy [177]. However, it is difficult for Prialt™ to cross the blood–brain barrier due to its inherent large molecular size and hydrophilicity [277], so its treatments are confined to intrathecal administration. Besides, there are various side effects (such as cognitive and neuropsychiatric adverse reactions [278]) that arise in the clinic and which need to be further investigated [273]. It is well known that the release of glutamate is essential in the process of spinal nociception, and the inhibition of the glutamate transporter GLT-1 can significantly reduce nociception behaviors [279]. Prialt™ is usually administered intrathecally to relieve pain by blocking VGCCs and abolishing capsaicin-evoked glutamate release in the spinal cord synaptosomes [220].
4.4.2. Neuroprotection
Studies have reported that Prialt™ has neuroprotective potential after spinal cord injury (SCI), which suggests it may be a good alternative treatment for acute SCI [178]. After SCI, secondary neuronal death happens due to the glutamate-mediated excitotoxicity, leading to excessive intracellular calcium, mitochondrial dysfunction, acidosis, and the overproduction of free radicals [280,281,282,283,284,285,286]. This condition can be prevented by Prialt™ via inhibiting the release of glutamate [287,288] and calcium influx [276] and protecting mitochondria from traumatic brain injury [289,290,291]. Moreover, Prialt™ can reduce the expression of nNOS to inhibit apoptosis [178]. Furthermore, a recent article has reported a novel ω-conotoxin Bu8 with five amino acid residues and three disulfide bonds synthesized by Conus Bullatus. The potency in inhibiting N-type voltage-gated calcium (CaV2.2) channels is twice that of MVIIA, and the inhibition of CaV2.2 channels is highly selective, so there are fewer side effects [292].
Because dendrotoxins (DTXs) selectively bind to the Kv1.1 channel, they can be used to study the basic biology of K+ channels and the mechanisms of synaptic transmission. DTX plays an essential role in neuronal degeneration and seizures in nonclinical models. Deletion of Kcna1/Kv1.1 or Kcna2/Kv1.2 has been reported to cause epilepsy in rodents [179,180]. DTXs can bind to presynaptic KV channels with a high affinity, which reveals their great potential to quantify the density of synapse in the CNS, and this potential usage might be applied to the diagnosis of neurodegenerative diseases that affect the integrity of the brain’s connectomes. For example, the loss of synapses in hippocampal tissue has been detected by α-DTX [2].
4.5. Postsynaptic Neurotoxins
Although there is currently no drug derived from α-neurotoxins on the market for muscle-associated diseases, there is data suggesting that some short-chain and long-chain α-neurotoxins that target the NMJ and other nAchRs have the potential therapeutic ability of treating disorders linked to NMJ dysfunction and others [181].
4.5.1. Anticancer
One characteristic of many tumors is the increased expression of nAChRs. The enhancement of tumor cell proliferation and the acceleration of tumor growth are associated with nAChRs [293]. Ach, an autocrine growth factor of human lung cancer, can bind to nAChRs in lung cancer cells to accelerate their proliferation, migration, and invasion [293]. Studies have shown that the nicotinic and/or muscarinic receptors mediate the growth and apoptosis signals in mesothelioma and non-small-cell lung cancer (NSCLC) [294,295,296,297]. Furthermore, the activation of nAChRs stimulates lung cancer growth [294,298,299]. These effects suggest that α-neurotoxins have potential anticancer therapeutic prospects because they antagonize the receptors of the nAchRs. Moreover, α-cobratoxin has shown potential anticancer effects for NSCLC [182].
4.5.2. Analgesia and Anti-Inflammation
We mainly discuss the long-chain toxin (cobratoxin) and short-chain toxin (cobrotoxin) in this review, focusing on their analgesic effects. Cobratoxin is purified from Naja naja kaouthia, and it has been suggested to have several essential functions including modulation of the nerves, suppression of the immune system, and anti-tumor, anti-inflammatory, and analgesic effects [183,300]. The possible analgesic mechanism is mediated by M4 mAChR, which is activated by cobratoxin and then triggers the CaMKII/CREB signaling pathway and inhibits low-potential Ca2+ channels [301]. Its effects of antinociception and anti-inflammation are also due to binding to the α7 subtype of nAChR with high affinity, which is followed by the decreased production of inflammatory cytokines such as TNF-α, IL-1, and IL-2 [302].
Cobrotoxin is purified from Naja atra venom (NNAV). It shows great inhibition of the inflammatory response in rat models of rheumatoid arthritis induced by adjuvant [303], as well as analgesic effects in rodent models of inflammatory pain induced by formalin and acetate [183]. It has been widely used for the treatment of chronic arthralgia, sciatica, and neuropathic headache in the clinic. Cobrotoxin is an approved drug by the CFDA [304] and has been called “Cobratide” in Chinese clinics. Recently, it has been reported that cobrotoxin in a higher dose has potential for the treatment of trigeminal neuralgia [305]. Cobrotoxin has similar analgesic action and underlying mechanisms of action as cobratoxin. It might have dual pain regulation, and there is a hypothesis to explain this phenomenon, namely, that cobrotoxin might activate the adenosine A1Rs and then inhibit the mitogen-activated protein kinases/extracellular signal-regulated protein kinase pathway to exert analgesic effects. However, with the increasing dose of cobrotoxin, the adenosine A2ARs are activated to produce sensitization to pain [306]. Besides the analgesic effects, cobrotoxin can also exert the effect of anti-inflammation by interacting with the NF-κB signaling pathway [307]. The binding between cobrotoxin and the kinase domain of IKK can inhibit the phosphorylation of IKK and then prevent the release of free NF-κB from IκB, thereby reducing the transcription of inflammatory genes [308]. It is quite interesting that initial animal and clinical studies showed that cobrotoxin (Cobratide) can treat acute gout and relieve motor and some non-motor symptoms of PD (unpublished observations from Dr Qin’s lab).
5. Summary
Neurotoxins targeting the synapse structure have attracted attention for treating diseases that do not have a very effective therapy in the clinic. Up to now, natural neurotoxins have been proven to be highly useful for the treatment of many kinds of diseases, and the list of diseases being treated with neurotoxins keeps expanding. The neurotoxins that have been approved for clinical use by the FDA and CFDA are Botox, Xeomin, Dysport, Myobloc/Neurobloc, Cobratide, and Prialt™; the rest are currently under preclinical investigation for a variety of diseases.
5.1. Clinical Applications of FDA-Approved Neurotoxins
As we can see from Table 2, the clinical applications of neurotoxins, mainly BoNTs, on the market are still limited, and most of them focus on muscle-related diseases and pain. Therefore, other indications for these neurotoxins are now being studied to expand their range of applications. BoNT has been found to treat neuropathic pain, neuroinflammation, depression, and skin diseases, but the mechanisms involved have not been confirmed yet, which limits their use in the clinic.
5.2. Neurotoxins in Preclinical Studies
The other neurotoxins are mostly studied in the preclinical period. More information about their pharmacological properties and toxicity is needed. For example, our understanding of the Ca2+-independent effects of α-latrotoxin is still insufficient.
5.3. Expectations
In biological therapy, neurotoxic peptides offer great therapeutic potential, but they still have many functions that need to be discovered and validated. In this review, we briefly summarized the biological and clinical functions of neurotoxins aiming at synapses. There are methodological challenges, such as the fact that these neurotoxins are peptides and cannot cross the BBB or that they are unstable, which limits the wider use of neurotoxins and is a major challenge. However, these neurotoxins aiming at synapses have passed the test of time as biological therapeutics and have also made positive contributions in animal models. In the future, more and more detailed studies are needed, and the structure of these neurotoxins should be continuously improved to make them suitable for clinical applications and benefit human beings.
Author Contributions
K.Z. contributed to writing the manuscript; W.L., T.L., Y.N., and Z.Q. conceived and supervised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (No. 81730092) and the Priority Academic Program Development of the Jiangsu Higher Education Institutes (PAPD).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schiavo, G.; Matteoli, M.; Montecucco, C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar] [CrossRef] [Green Version]
- Ovsepian, S.V.; O’Leary, V.B.; Ayvazyan, N.M.; Al-Sabi, A.; Ntziachristos, V.; Dolly, J.O. Neurobiology and therapeutic applications of neurotoxins targeting transmitter release. Pharmacol. Ther. 2019, 193, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B.W.; Kusner, L.L.; Kaminski, H.J. Molecular architecture of the neuromuscular junction. Muscle Nerve 2006, 33, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Ayvazyan, N.M.; O’Leary, V.B.; Dolly, J.O.; Ovsepian, S.V. Neurobiology and therapeutic utility of neurotoxins targeting postsynaptic mechanisms of neuromuscular transmission. Drug Discov. Today 2019, 24, 1968–1984. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Few, A.P. Calcium channel regulation and presynaptic plasticity. Neuron 2008, 59, 882–901. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.A.; Frølund, B.; Liljefors, T.; Krogsgaard-Larsen, P. Neuronal nicotinic acetylcholine receptors: Structural revelations, target identifications, and therapeutic inspirations. J. Med. Chem. 2005, 48, 4705–4745. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, E.X.; Pereira, E.F.R.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [Green Version]
- Wonnacott, S. Presynaptic nicotinic ACh receptors. Trends Neurosci. 1997, 20, 92–98. [Google Scholar] [CrossRef]
- Chen, J.; Cheuk, I.W.Y.; Shin, V.Y.; Kwong, A. Acetylcholine receptors: Key players in cancer development. Surg. Oncol. 2019, 31, 46–53. [Google Scholar] [CrossRef]
- Schuller, H.M. Is cancer triggered by altered signalling of nicotinic acetylcholine receptors? Nat. Rev. Cancer 2009, 9, 195–205. [Google Scholar] [CrossRef]
- Rossetto, O.; Rigoni, M.; Montecucco, C. Different mechanism of blockade of neuroexocytosis by presynaptic neurotoxins. Toxicol. Lett. 2004, 149, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Atlas, D. The voltage-gated calcium channel functions as the molecular switch of synaptic transmission. Annu. Rev. Biochem. 2013, 82, 607–635. [Google Scholar] [CrossRef] [PubMed]
- Tsetlin, V.I. Three-finger snake neurotoxins and Ly6 proteins targeting nicotinic acetylcholine receptors: Pharmacological tools and endogenous modulators. Trends Pharmacol. Sci. 2015, 36, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Nicke, A.; Tsetlin, V.I. Nicotinic acetylcholine receptor inhibitors derived from snake and snail venoms. Neuropharmacology 2017, 127, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon Off. J. Int. Soc. Toxinology 2013, 66, 47–58. [Google Scholar] [CrossRef]
- Nirthanan, S. Snake three-finger α-neurotoxins and nicotinic acetylcholine receptors: Molecules, mechanisms and medicine. Biochem. Pharmacol. 2020, 181, 114168. [Google Scholar] [CrossRef]
- Lalli, G.; Bohnert, S.; Deinhardt, K.; Verastegui, C.; Schiavo, G. The journey of tetanus and botulinum neurotoxins in neurons. Trends Microbiol. 2003, 11, 431–437. [Google Scholar] [CrossRef]
- Dong, M.; Masuyer, G.; Stenmark, P. Botulinum and Tetanus Neurotoxins. Annu. Rev. Biochem. 2019, 88, 811–837. [Google Scholar] [CrossRef]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum Neurotoxins: Biology, Pharmacology, and Toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef] [Green Version]
- Montal, M. Translocation of botulinum neurotoxin light chain protease by the heavy chain protein-conducting channel. Toxicon Off. J. Int. Soc. Toxinology 2009, 54, 565–569. [Google Scholar] [CrossRef]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gardberg, A.S.; Edwards, T.E.; Sankaran, B.; Robinson, H.; Varnum, S.M.; Buchko, G.W. Structural insights into the functional role of the Hcn sub-domain of the receptor-binding domain of the botulinum neurotoxin mosaic serotype C/D. Biochimie 2013, 95, 1379–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, A.; Montal, M. Molecular dissection of botulinum neurotoxin reveals interdomain chaperone function. Toxicon Off. J. Int. Soc. Toxinology 2013, 75, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavo, G.; Poulain, B.; Rossetto, O.; Benfenati, F.; Tauc, L.; Montecucco, C. Tetanus toxin is a zinc protein and its inhibition of neurotransmitter release and protease activity depend on zinc. EMBO J. 1992, 11, 3577–3583. [Google Scholar] [CrossRef] [PubMed]
- Breidenbach, M.A.; Brunger, A.T. Substrate recognition strategy for botulinum neurotoxin serotype A. Nature 2004, 432, 925–929. [Google Scholar] [CrossRef]
- Montal, M. Botulinum neurotoxin: A marvel of protein design. Annu. Rev. Biochem. 2010, 79, 591–617. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Zhou, P.; Wang, A.L.; Wu, D.; Zhao, M.; Südhof, T.C.; Brunger, A.T. The primed SNARE-complexin-synaptotagmin complex for neuronal exocytosis. Nature 2017, 548, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Südhof, T.C.; Rothman, J.E. Membrane fusion: Grappling with SNARE and SM proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [Green Version]
- Jahn, R.; Scheller, R.H. SNAREs--Engines for membrane fusion. Nat Rev Mol Cell Biol 2006, 7, 631–643. [Google Scholar] [CrossRef]
- Popoff, M.R.; Poulain, B. Bacterial toxins and the nervous system: Neurotoxins and multipotential toxins interacting with neuronal cells. Toxins 2010, 2, 683–737. [Google Scholar] [CrossRef]
- Poulain, B.; Popoff, M.R. Why Are Botulinum Neurotoxin-Producing Bacteria So Diverse and Botulinum Neurotoxins So Toxic? Toxins 2019, 11, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulain, B.; Lemichez, E.; Popoff, M.R. Neuronal selectivity of botulinum neurotoxins. Toxicon Off. J. Int. Soc. Toxinology 2020, 178, 20–32. [Google Scholar] [CrossRef]
- Luvisetto, S. Botulinum Neurotoxins beyond Neurons: Interplay with Glial Cells. Toxins 2022, 14, 704. [Google Scholar] [CrossRef]
- Zhang, S.; Masuyer, G.; Zhang, J.; Shen, Y.; Lundin, D.; Henriksson, L.; Miyashita, S.-I.; Martínez-Carranza, M.; Dong, M.; Stenmark, P. Identification and characterization of a novel botulinum neurotoxin. Nat. Commun. 2017, 8, 14130. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.L. Identification of the major steps in botulinum toxin action. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 167–193. [Google Scholar] [CrossRef]
- Peck, M.W.; Smith, T.J.; Anniballi, F.; Austin, J.W.; Bano, L.; Bradshaw, M.; Cuervo, P.; Cheng, L.W.; Derman, Y.; Dorner, B.G.; et al. Historical Perspectives and Guidelines for Botulinum Neurotoxin Subtype Nomenclature. Toxins 2017, 9, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgen, A.S.V.; Dickens, F.; Zatman, L.J. The action of botulinum toxin on the neuro-muscular junction. J. Physiol. 1949, 109, 10–24. [Google Scholar] [CrossRef]
- Van der Kloot, W.; Molgó, J. Quantal acetylcholine release at the vertebrate neuromuscular junction. Physiol. Rev. 1994, 74, 899–991. [Google Scholar] [CrossRef]
- Poulain, B.; Molgó, J.; Thesleff, S. Quantal neurotransmitter release and the clostridial neurotoxins’ targets. Curr. Top. Microbiol. Immunol. 1995, 195, 243–255. [Google Scholar]
- Cook, T.M.; Protheroe, R.T.; Handel, J.M. Tetanus: A review of the literature. Br. J. Anaesth. 2001, 87, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Megighian, A.; Pirazzini, M.; Fabris, F.; Rossetto, O.; Montecucco, C. Tetanus and tetanus neurotoxin: From peripheral uptake to central nervous tissue targets. J. Neurochem. 2021, 158, 1244–1253. [Google Scholar] [CrossRef] [PubMed]
- Brooks, V.B.; Curtis, D.R.; Eccles, J.C. Mode of action of tetanus toxin. Nature 1955, 175, 120–121. [Google Scholar] [CrossRef] [PubMed]
- Yen, L.M.; Thwaites, C.L. Tetanus. Lancet 2019, 393, 1657–1668. [Google Scholar] [CrossRef]
- Alfery, D.D.; Rauscher, L.A. Tetanus: A review. Crit. Care Med. 1979, 7, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Pellizzari, R.; Rossetto, O.; Schiavo, G.; Montecucco, C. Tetanus and botulinum neurotoxins: Mechanism of action and therapeutic uses. Philos. Trans. R Soc. Lond. B Biol. Sci. 1999, 354, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, M.; Stenmark, P. The Structure and Classification of Botulinum Toxins. Handb. Exp. Pharmacol. 2021, 263, 11–33. [Google Scholar] [CrossRef]
- Tzeng, M.C.; Siekevitz, P. The effect of the purified major protein factor (alpha-latrotoxin) of black widow spider venom on the release of acetylcholine and norepinephrine from mouse cerebral cortex slices. Brain Res. 1978, 139, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Grishin, E.V. Black widow spider toxins: The present and the future. Toxicon Off. J. Int. Soc. Toxinology 1998, 36, 1693–1701. [Google Scholar] [CrossRef]
- Krasnoperov, V.G.; Shamotienko, O.G.; Grishin, E.V. A crustacean-specific neurotoxin from the venom of the black widow spider Latrodectus mactans tredecimguttatus. Bioorg. Khim. 1990, 16, 1567–1569. [Google Scholar]
- Müller, G.J. Black and brown widow spider bites in South Africa. A series of 45 cases. S. Afr. Med. J. 1993, 83, 399–405. [Google Scholar]
- Zukowski, C.W. Black widow spider bite. J. Am. Board Fam. Pract. 1993, 6, 279–281. [Google Scholar] [PubMed]
- Petrenko, A.G.; Kovalenko, V.A.; Shamotienko, O.G.; Surkova, I.N.; Tarasyuk, T.A.; Ushkaryov Yu, A.; Grishin, E.V. Isolation and properties of the alpha-latrotoxin receptor. EMBO J. 1990, 9, 2023–2027. [Google Scholar] [CrossRef] [PubMed]
- Ushkaryov, Y.A.; Petrenko, A.G.; Geppert, M.; Südhof, T.C. Neurexins: Synaptic cell surface proteins related to the alpha-latrotoxin receptor and laminin. Science 1992, 257, 50–56. [Google Scholar] [CrossRef]
- Davletov, B.A.; Krasnoperov, V.; Hata, Y.; Petrenko, A.G.; Südhof, T.C. High affinity binding of alpha-latrotoxin to recombinant neurexin I alpha. J. Biol. Chem. 1995, 270, 23903–23905. [Google Scholar] [CrossRef] [Green Version]
- Davletov, B.A.; Shamotienko, O.G.; Lelianova, V.G.; Grishin, E.V.; Ushkaryov, Y.A. Isolation and biochemical characterization of a Ca2+-independent alpha-latrotoxin-binding protein. J. Biol. Chem. 1996, 271, 23239–23245. [Google Scholar] [CrossRef] [Green Version]
- Krasnoperov, V.G.; Beavis, R.; Chepurny, O.G.; Little, A.R.; Plotnikov, A.N.; Petrenko, A.G. The calcium-independent receptor of alpha-latrotoxin is not a neurexin. Biochem. Biophys. Res. Commun. 1996, 227, 868–875. [Google Scholar] [CrossRef]
- Krasnoperov, V.; Bittner, M.A.; Mo, W.; Buryanovsky, L.; Neubert, T.A.; Holz, R.W.; Ichtchenko, K.; Petrenko, A.G. Protein-tyrosine phosphatase-sigma is a novel member of the functional family of alpha-latrotoxin receptors. J. Biol. Chem. 2002, 277, 35887–35895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, H.T.; Rosenthal, L.; Meldolesi, J.; Nicholls, D.G. Alpha-latrotoxin releases both vesicular and cytoplasmic glutamate from isolated nerve terminals. J. Neurochem. 1990, 55, 2039–2047. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, L.; Zacchetti, D.; Madeddu, L.; Meldolesi, J. Mode of action of alpha-latrotoxin: Role of divalent cations in Ca2(+)-dependent and Ca2(+)-independent effects mediated by the toxin. Mol. Pharmacol. 1990, 38, 917–923. [Google Scholar] [PubMed]
- Ushkaryov, Y.A.; Rohou, A.; Sugita, S. α-Latrotoxin and its receptors. In Pharmacology of Neurotransmitter Release; Handbook of Experimental Pharmacology book series; Springer: Berlin/Heidelberg, Germany, 2008; pp. 171–206. [Google Scholar]
- Grasso, A.; Alemà, S.; Rufini, S.; Senni, M.I. Black widow spider toxin-induced calcium fluxes and transmitter release in a neurosecretory cell line. Nature 1980, 283, 774–776. [Google Scholar] [CrossRef]
- Ashton, A.C.; Volynski, K.E.; Lelianova, V.G.; Orlova, E.V.; Van Renterghem, C.; Canepari, M.; Seagar, M.; Ushkaryov, Y.A. α-Latrotoxin, acting via two Ca2+-dependent pathways, triggers exocytosis of two pools of synaptic vesicles. J. Biol. Chem. 2001, 276, 44695–44703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.; Hodgson, W.C.; Isbister, G.K. Antivenom for Neuromuscular Paralysis Resulting from Snake Envenoming. Toxins 2017, 9, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.; Kuruppu, S.; Othman, I.; Goode, R.J.A.; Hodgson, W.C.; Isbister, G.K. Neurotoxicity in Sri Lankan Russell’s Viper (Daboia russelii) Envenoming is Primarily due to U1-viperitoxin-Dr1a, a Pre-Synaptic Neurotoxin. Neurotox. Res. 2017, 31, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Rossetto, O. How do presynaptic PLA2 neurotoxins block nerve terminals? Trends Biochem. Sci. 2000, 25, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Carredano, E.; Westerlund, B.; Persson, B.; Saarinen, M.; Ramaswamy, S.; Eaker, D.; Eklund, H. The three-dimensional structures of two toxins from snake venom throw light on the anticoagulant and neurotoxic sites of phospholipase A2. Toxicon Off. J. Int. Soc. Toxinology 1998, 36, 75–92. [Google Scholar] [CrossRef]
- Heinrikson, R.L.; Krueger, E.T.; Keim, P.S. Amino acid sequence of phospholipase A2-alpha from the venom of Crotalus adamanteus. A new classification of phospholipases A2 based upon structural determinants. J. Biol. Chem. 1977, 252, 4913–4921. [Google Scholar] [CrossRef]
- Andrião-Escarso, S.H.; Soares, A.M.; Rodrigues, V.M.; Angulo, Y.; Díaz, C.; Lomonte, B.; Gutiérrez, J.M.; Giglio, J.R. Myotoxic phospholipases A(2) in bothrops snake venoms: Effect of chemical modifications on the enzymatic and pharmacological properties of bothropstoxins from Bothrops jararacussu. Biochimie 2000, 82, 755–763. [Google Scholar] [CrossRef]
- Barbosa, P.S.F.; Martins, A.M.C.; Havt, A.; Toyama, D.O.; Evangelista, J.S.A.M.; Ferreira, D.P.P.; Joazeiro, P.P.; Beriam, L.O.S.; Toyama, M.H.; Fonteles, M.C.; et al. Renal and antibacterial effects induced by myotoxin I and II isolated from Bothrops jararacussu venom. Toxicon Off. J. Int. Soc. Toxinology 2005, 46, 376–386. [Google Scholar] [CrossRef]
- Costa Torres, A.F.; Dantas, R.T.; Toyama, M.H.; Diz Filho, E.; Zara, F.J.; Rodrigues de Queiroz, M.G.; Pinto Nogueira, N.A.; Rosa de Oliveira, M.; de Oliveira Toyama, D.; Monteiro, H.S.A.; et al. Antibacterial and antiparasitic effects of Bothrops marajoensis venom and its fractions: Phospholipase A2 and L-amino acid oxidase. Toxicon Off. J. Int. Soc. Toxinology 2010, 55, 795–804. [Google Scholar] [CrossRef]
- Evangelista, I.L.; Martins, A.M.C.; Nascimento, N.R.F.; Havt, A.; Evangelista, J.S.A.M.; de Norões, T.B.S.; Toyama, M.H.; Diz-Filho, E.B.; Toyama, D.d.O.; Fonteles, M.C.; et al. Renal and cardiovascular effects of Bothrops marajoensis venom and phospholipase A2. Toxicon Off. J. Int. Soc. Toxinology 2010, 55, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.B.; Grubb, B.D.; Maltin, C.A.; Dixon, R. The neurotoxicity of the venom phospholipases A(2), notexin and taipoxin. Exp. Neurol. 2000, 161, 517–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kini, R.M.; Evans, H.J. Structure-function relationships of phospholipases. The anticoagulant region of phospholipases A2. J. Biol. Chem. 1987, 262, 14402–14407. [Google Scholar] [CrossRef] [PubMed]
- Landucci, E.C.; de Castro, R.C.; Toyama, M.; Giglio, J.R.; Marangoni, S.; De Nucci, G.; Antunes, E. Inflammatory oedema induced by the lys-49 phospholipase A(2) homologue piratoxin-i in the rat and rabbit. Effect of polyanions and p-bromophenacyl bromide. Biochem. Pharmacol. 2000, 59, 1289–1294. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Ferreira, A.L.; Menaldo, D.L.; Sartim, M.A.; Riul, T.B.; Dias-Baruffi, M.; Sampaio, S.V. Antithrombotic activity of Batroxase, a metalloprotease from Bothrops atrox venom, in a model of venous thrombosis. Int. J. Biol. Macromol. 2017, 95, 263–267. [Google Scholar] [CrossRef]
- Hiu, J.J.; Yap, M.K.K. Cytotoxicity of snake venom enzymatic toxins: Phospholipase A2 and l-amino acid oxidase. Biochem. Soc. Trans. 2020, 48, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.W.; Harris, J.B. Nerve terminal damage by beta-bungarotoxin: Its clinical significance. Am. J. Pathol. 1999, 154, 447–455. [Google Scholar] [CrossRef]
- Gopalakrishnakone, P.; Hawgood, B.J. Morphological changes induced by crotoxin in murine nerve and neuromuscular junction. Toxicon Off. J. Int. Soc. Toxinology 1984, 22, 791–804. [Google Scholar] [CrossRef]
- Cull-Candy, S.G.; Fohlman, J.; Gustavsson, D.; Lüllmann-Rauch, R.; Thesleff, S. The effects of taipoxin and notexin on the function and fine structure of the murine neuromuscular junction. Neuroscience 1976, 1, 175–180. [Google Scholar] [CrossRef]
- Chen, I.L.; Lee, C.Y. Ultrastructural changes in the motor nerve terminals caused by beta-bungarotoxin. Virchows Arch. B Cell Pathol 1970, 6, 318–325. [Google Scholar] [CrossRef]
- Lee, C.Y.; Tsai, M.C.; Chen, Y.M.; Ritonja, A.; Gubensek, F. Mode of neuromuscular blocking action of toxic phospholipases A2 from Vipera ammodytes venom. Arch. Int. Pharmacodyn. Ther. 1984, 268, 313–324. [Google Scholar]
- Belo, C.A.D.; Leite, G.B.; Toyama, M.H.; Marangoni, S.; Corrado, A.P.; Fontana, M.D.; Southan, A.; Rowan, E.G.; Hyslop, S.; Rodrigues-Simioni, L. Pharmacological and structural characterization of a novel phospholipase A2 from Micrurus dumerilii carinicauda venom. Toxicon Off. J. Int. Soc. Toxinology 2005, 46, 736–750. [Google Scholar] [CrossRef] [PubMed]
- Belo, C.A.D.; Toyama, M.H.; Toyama, D.d.O.; Marangoni, S.; Moreno, F.B.; Cavada, B.S.; Fontana, M.D.; Hyslop, S.; Carneiro, E.M.; Boschero, A.C. Determination of the amino acid sequence of a new phospholipase A(2) (MIDCA1) isolated from Micrurus dumerilii carinicauda venom. Protein J. 2005, 24, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Schütter, N.; Barreto, Y.C.; Vardanyan, V.; Hornig, S.; Hyslop, S.; Marangoni, S.; Rodrigues-Simioni, L.; Pongs, O.; Dal Belo, C.A. Inhibition of Kv2.1 Potassium Channels by MiDCA1, A Pre-Synaptically Active PLA2-Type Toxin from Micrurus dumerilii carinicauda Coral Snake Venom. Toxins 2019, 11, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, D.T. The dawn of high-resolution structure for the queen of ion channels. Neuron 2004, 42, 357–359. [Google Scholar] [CrossRef]
- Pitt, G.S.; Matsui, M.; Cao, C. Voltage-Gated Calcium Channels in Nonexcitable Tissues. Annu. Rev. Physiol. 2021, 83, 183–203. [Google Scholar] [CrossRef]
- Nimmrich, V.; Gross, G. P/Q-type calcium channel modulators. Br. J. Pharmacol. 2012, 167, 741–759. [Google Scholar] [CrossRef] [Green Version]
- Dolphin, A.C.; Lee, A. Presynaptic calcium channels: Specialized control of synaptic neurotransmitter release. Nat. Rev. Neurosci. 2020, 21, 213–229. [Google Scholar] [CrossRef]
- Adams, M.E. Agatoxins: Ion channel specific toxins from the American funnel web spider, Agelenopsis aperta. Toxicon Off. J. Int. Soc. Toxinology 2004, 43, 509–525. [Google Scholar] [CrossRef]
- Pringos, E.; Vignes, M.; Martinez, J.; Rolland, V. Peptide neurotoxins that affect voltage-gated calcium channels: A close-up on ω-agatoxins. Toxins 2011, 3, 17–42. [Google Scholar] [CrossRef] [Green Version]
- Bindokas, V.P.; Venema, V.J.; Adams, M.E. Differential antagonism of transmitter release by subtypes of omega-agatoxins. J. Neurophysiol. 1991, 66, 590–601. [Google Scholar] [CrossRef]
- Santos, A.D.; Imperial, J.S.; Chaudhary, T.; Beavis, R.C.; Chait, B.T.; Hunsperger, J.P.; Olivera, B.M.; Adams, M.E.; Hillyard, D.R. Heterodimeric structure of the spider toxin omega-agatoxin IA revealed by precursor analysis and mass spectrometry. J. Biol. Chem. 1992, 267, 20701–20705. [Google Scholar] [CrossRef]
- Olivera, B.M.; Miljanich, G.P.; Ramachandran, J.; Adams, M.E. Calcium channel diversity and neurotransmitter release: The omega-conotoxins and omega-agatoxins. Annu. Rev. Biochem. 1994, 63, 823–867. [Google Scholar] [CrossRef] [PubMed]
- Mintz, I.M.; Venema, V.J.; Adams, M.E.; Bean, B.P. Inhibition of N- and L-type Ca2+ channels by the spider venom toxin omega-Aga-IIIA. Proc. Natl. Acad. Sci. USA 1991, 88, 6628–6631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mintz, I.M. Block of Ca channels in rat central neurons by the spider toxin omega-Aga-IIIA. J. Neurosci. 1994, 14, 2844–2853. [Google Scholar] [CrossRef] [PubMed]
- Bourinet, E.; Soong, T.W.; Sutton, K.; Slaymaker, S.; Mathews, E.; Monteil, A.; Zamponi, G.W.; Nargeot, J.; Snutch, T.P. Splicing of alpha 1A subunit gene generates phenotypic variants of P- and Q-type calcium channels. Nat. Neurosci. 1999, 2, 407–415. [Google Scholar] [CrossRef] [PubMed]
- McDonough, S.I.; Mintz, I.M.; Bean, B.P. Alteration of P-type calcium channel gating by the spider toxin omega-Aga-IVA. Biophys. J. 1997, 72, 2117–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mintz, I.M.; Venema, V.J.; Swiderek, K.M.; Lee, T.D.; Bean, B.P.; Adams, M.E. P-type calcium channels blocked by the spider toxin omega-Aga-IVA. Nature 1992, 355, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.-H.; Dekan, Z.; Smout, M.J.; Wilson, D.; Dutertre, S.; Vetter, I.; Lewis, R.J.; Loukas, A.; Daly, N.L.; Alewood, P.F. Conotoxin Φ-MiXXVIIA from the Superfamily G2 Employs a Novel Cysteine Framework that Mimics Granulin and Displays Anti-Apoptotic Activity. Angew. Chem. Int. Ed. Engl. 2017, 56, 14973–14976. [Google Scholar] [CrossRef]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, synthesis, and structure-activity relationships of conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef]
- Vetter, I.; Lewis, R.J. Therapeutic potential of cone snail venom peptides (conopeptides). Curr. Top. Med. Chem. 2012, 12, 1546–1552. [Google Scholar] [CrossRef]
- Adams, D.J.; Callaghan, B.; Berecki, G. Analgesic conotoxins: Block and G protein-coupled receptor modulation of N-type (Ca(V) 2.2) calcium channels. Br. J. Pharmacol. 2012, 166, 486–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boland, L.M.; Morrill, J.A.; Bean, B.P. omega-Conotoxin block of N-type calcium channels in frog and rat sympathetic neurons. J. Neurosci. 1994, 14, 5011–5027. [Google Scholar] [CrossRef] [PubMed]
- Ellinor, P.T.; Zhang, J.F.; Horne, W.A.; Tsien, R.W. Structural determinants of the blockade of N-type calcium channels by a peptide neurotoxin. Nature 1994, 372, 272–275. [Google Scholar] [CrossRef]
- Kerr, L.M.; Yoshikami, D. A venom peptide with a novel presynaptic blocking action. Nature 1984, 308, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Cruz, L.J.; de Santos, V.; LeCheminant, G.W.; Griffin, D.; Zeikus, R.; McIntosh, J.M.; Galyean, R.; Varga, J.; Gray, W.R. Neuronal calcium channel antagonists. Discrimination between calcium channel subtypes using omega-conotoxin from Conus magus venom. Biochemistry 1987, 26, 2086–2090. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Lin, Z.; Haus, S.; Edgerton, J.; Lipscombe, D. Identification of functionally distinct isoforms of the N-type Ca2+ channel in rat sympathetic ganglia and brain. Neuron 1997, 18, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Servent, D.; Winckler-Dietrich, V.; Hu, H.Y.; Kessler, P.; Drevet, P.; Bertrand, D.; Ménez, A. Only snake curaremimetic toxins with a fifth disulfide bond have high affinity for the neuronal alpha7 nicotinic receptor. J. Biol. Chem. 1997, 272, 24279–24286. [Google Scholar] [CrossRef] [Green Version]
- Changeux, J.P. The TiPS lecture. The nicotinic acetylcholine receptor: An allosteric protein prototype of ligand-gated ion channels. Trends Pharmacol. Sci. 1990, 11, 485–492. [Google Scholar] [CrossRef]
- Fruchart-Gaillard, C.; Gilquin, B.; Antil-Delbeke, S.; Le Novère, N.; Tamiya, T.; Corringer, P.-J.; Changeux, J.-P.; Ménez, A.; Servent, D. Experimentally based model of a complex between a snake toxin and the alpha 7 nicotinic receptor. Proc. Natl. Acad. Sci. USA 2002, 99, 3216–3221. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C. Looking back on the discovery of alpha-bungarotoxin. J. Biomed. Sci. 1999, 6, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Nirthanan, S.; Gwee, M.C.E. Three-finger alpha-neurotoxins and the nicotinic acetylcholine receptor, forty years on. J. Pharmacol. Sci. 2004, 94, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirthanan, S.; Gopalakrishnakone, P.; Gwee, M.C.E.; Khoo, H.E.; Kini, R.M. Non-conventional toxins from Elapid venoms. Toxicon Off. J. Int. Soc. Toxinology 2003, 41, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Tsetlin, V. Snake venom alpha-neurotoxins and other ‘three-finger’ proteins. Eur. J. Biochem. 1999, 264, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Modahl, C.M.; Mukherjee, A.K.; Mackessy, S.P. An analysis of venom ontogeny and prey-specific toxicity in the Monocled Cobra (Naja kaouthia). Toxicon Off. J. Int. Soc. Toxinology 2016, 119, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Tamiya, N. Current view on the structure-function relationship of postsynaptic neurotoxins from snake venoms. Pharmacol. Ther. 1987, 34, 403–451. [Google Scholar] [CrossRef]
- Tsetlin, V.; Utkin, Y.; Kasheverov, I. Polypeptide and peptide toxins, magnifying lenses for binding sites in nicotinic acetylcholine receptors. Biochem. Pharmacol. 2009, 78, 720–731. [Google Scholar] [CrossRef]
- Servent, D.; Antil-Delbeke, S.; Gaillard, C.; Corringer, P.J.; Changeux, J.P.; Ménez, A. Molecular characterization of the specificity of interactions of various neurotoxins on two distinct nicotinic acetylcholine receptors. Eur. J. Pharmacol. 2000, 393, 197–204. [Google Scholar] [CrossRef]
- Antil-Delbeke, S.; Gaillard, C.; Tamiya, T.; Corringer, P.J.; Changeux, J.P.; Servent, D.; Ménez, A. Molecular determinants by which a long chain toxin from snake venom interacts with the neuronal alpha 7-nicotinic acetylcholine receptor. J. Biol. Chem. 2000, 275, 29594–29601. [Google Scholar] [CrossRef] [Green Version]
- Papke, R.L.; Horenstein, N.A. Therapeutic Targeting of 7 Nicotinic Acetylcholine Receptors. Pharmacol. Rev. 2021, 73, 1118–1149. [Google Scholar] [CrossRef]
- Briggs, C.A.; Grønlien, J.H.; Curzon, P.; Timmermann, D.B.; Ween, H.; Thorin-Hagene, K.; Kerr, P.; Anderson, D.J.; Malysz, J.; Dyhring, T.; et al. Role of channel activation in cognitive enhancement mediated by alpha7 nicotinic acetylcholine receptors. Br. J. Pharmacol. 2009, 158, 1486–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieschl, R.L.; Miller, R.; Jones, K.M.; Post-Munson, D.J.; Chen, P.; Newberry, K.; Benitex, Y.; Molski, T.; Morgan, D.; McDonald, I.M.; et al. Effects of BMS-902483, an α7 nicotinic acetylcholine receptor partial agonist, on cognition and sensory gating in relation to receptor occupancy in rodents. Eur. J. Pharmacol. 2017, 807, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, H.; Ulloa, L.; et al. Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Schröder, H.; Giacobini, E.; Struble, R.G.; Zilles, K.; Maelicke, A. Nicotinic cholinoceptive neurons of the frontal cortex are reduced in Alzheimer’s disease. Neurobiol. Aging 1991, 12, 259–262. [Google Scholar] [CrossRef]
- Schröder, H.; Giacobini, E.; Struble, R.G.; Zilles, K.; Maelicke, A.; Luiten, P.G.; Strosberg, A.D. Cellular distribution and expression of cortical acetylcholine receptors in aging and Alzheimer’s disease. Ann. New York Acad. Sci. 1991, 640, 189–192. [Google Scholar] [CrossRef]
- Lange, K.W.; Wells, F.R.; Jenner, P.; Marsden, C.D. Altered muscarinic and nicotinic receptor densities in cortical and subcortical brain regions in Parkinson’s disease. J. Neurochem. 1993, 60, 197–203. [Google Scholar] [CrossRef]
- Freedman, R.; Hall, M.; Adler, L.E.; Leonard, S. Evidence in postmortem brain tissue for decreased numbers of hippocampal nicotinic receptors in schizophrenia. Biol. Psychiatry 1995, 38, 22–33. [Google Scholar] [CrossRef]
- James, J.R.; Nordberg, A. Genetic and environmental aspects of the role of nicotinic receptors in neurodegenerative disorders: Emphasis on Alzheimer’s disease and Parkinson’s disease. Behav. Genet. 1995, 25, 149–159. [Google Scholar] [CrossRef]
- Perry, E.K.; Morris, C.M.; Court, J.A.; Cheng, A.; Fairbairn, A.F.; McKeith, I.G.; Irving, D.; Brown, A.; Perry, R.H. Alteration in nicotine binding sites in Parkinson’s disease, Lewy body dementia and Alzheimer’s disease: Possible index of early neuropathology. Neuroscience 1995, 64, 385–395. [Google Scholar] [CrossRef]
- Nordberg, A.; Lundqvist, H.; Hartvig, P.; Andersson, J.; Johansson, M.; Hellstrŏm-Lindahi, E.; Långström, B. Imaging of nicotinic and muscarinic receptors in Alzheimer’s disease: Effect of tacrine treatment. Dement. Geriatr. Cogn. Disord. 1997, 8, 78–84. [Google Scholar] [CrossRef]
- Spurden, D.P.; Court, J.A.; Lloyd, S.; Oakley, A.; Perry, R.; Pearson, C.; Pullen, R.G.; Perry, E.K. Nicotinic receptor distribution in the human thalamus: Autoradiographical localization of [3H]nicotine and [125I] alpha-bungarotoxin binding. J. Chem. Neuroanat. 1997, 13, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Gotti, C.; Moretti, M.; Bohr, I.; Ziabreva, I.; Vailati, S.; Longhi, R.; Riganti, L.; Gaimarri, A.; McKeith, I.G.; Perry, R.H.; et al. Selective nicotinic acetylcholine receptor subunit deficits identified in Alzheimer’s disease, Parkinson’s disease and dementia with Lewy bodies by immunoprecipitation. Neurobiol. Dis. 2006, 23, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef] [PubMed]
- van Westerloo, D.J.; Giebelen, I.A.; Florquin, S.; Bruno, M.J.; Larosa, G.J.; Ulloa, L.; Tracey, K.J.; van der Poll, T. The vagus nerve and nicotinic receptors modulate experimental pancreatitis severity in mice. Gastroenterology 2006, 130, 1822–1830. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Ochani, M.; Yang, L.-H.; Gallowitsch-Puerta, M.; Ochani, K.; Lin, X.; Levi, J.; Parrish, W.R.; Rosas-Ballina, M.; Czura, C.J.; et al. Selective alpha7-nicotinic acetylcholine receptor agonist GTS-21 improves survival in murine endotoxemia and severe sepsis. Crit. Care Med. 2007, 35, 1139–1144. [Google Scholar] [CrossRef]
- Rosas-Ballina, M.; Goldstein, R.S.; Gallowitsch-Puerta, M.; Yang, L.; Valdés-Ferrer, S.I.; Patel, N.B.; Chavan, S.; Al-Abed, Y.; Yang, H.; Tracey, K.J. The selective alpha7 agonist GTS-21 attenuates cytokine production in human whole blood and human monocytes activated by ligands for TLR2, TLR3, TLR4, TLR9, and RAGE. Mol. Med. 2009, 15, 195–202. [Google Scholar] [CrossRef]
- Rosas-Ballina, M.; Tracey, K.J. Cholinergic control of inflammation. J. Intern. Med. 2009, 265, 663–679. [Google Scholar] [CrossRef]
- Wang, G.K.; Schmidt, J. Primary structure and binding properties of iodinated derivatives of alpha-bungarotoxin. J. Biol. Chem. 1980, 255, 11156–11162. [Google Scholar] [CrossRef]
- Azam, L.; McIntosh, J.M. Alpha-conotoxins as pharmacological probes of nicotinic acetylcholine receptors. Acta Pharmacol. Sin. 2009, 30, 771–783. [Google Scholar] [CrossRef] [Green Version]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [Green Version]
- Janes, R.W. α-Conotoxins as selective probes for nicotinic acetylcholine receptor subclasses. Curr. Opin. Pharmacol. 2005, 5, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Wioland, L.; Dupont, J.-L.; Bossu, J.-L.; Popoff, M.R.; Poulain, B. Attack of the nervous system by Clostridium perfringens Epsilon toxin: From disease to mode of action on neural cells. Toxicon Off. J. Int. Soc. Toxinology 2013, 75, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Castagliuolo, I.; LaMont, J.T.; Letourneau, R.; Kelly, C.; O’Keane, J.C.; Jaffer, A.; Theoharides, T.C.; Pothoulakis, C. Neuronal involvement in the intestinal effects of Clostridium difficile toxin A and Vibrio cholerae enterotoxin in rat ileum. Gastroenterology 1994, 107, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Burleigh, D.E.; Banks, M.R. Stimulation of intestinal secretion by vasoactive intestinal peptide and cholera toxin. Auton. Neurosci. 2007, 133, 64–75. [Google Scholar] [CrossRef]
- Aktories, K. Bacterial protein toxins that modify host regulatory GTPases. Nat. Rev. Microbiol. 2011, 9, 487–498. [Google Scholar] [CrossRef]
- Aktories, K.; Schwan, C.; Papatheodorou, P.; Lang, A.E. Bidirectional attack on the actin cytoskeleton. Bacterial protein toxins causing polymerization or depolymerization of actin. Toxicon Off. J. Int. Soc. Toxinology 2012, 60, 572–581. [Google Scholar] [CrossRef]
- Blanchfield, J.T.; Gallagher, O.P.; Cros, C.; Lewis, R.J.; Alewood, P.F.; Toth, I. Oral absorption and in vivo biodistribution of alpha-conotoxin MII and a lipidic analogue. Biochem. Biophys. Res. Commun. 2007, 361, 97–102. [Google Scholar] [CrossRef]
- Oller-Salvia, B.; Teixidó, M.; Giralt, E. From venoms to BBB shuttles: Synthesis and blood-brain barrier transport assessment of apamin and a nontoxic analog. Biopolymers 2013, 100, 675–686. [Google Scholar] [CrossRef]
- Matak, I.; Bölcskei, K.; Bach-Rojecky, L.; Helyes, Z. Mechanisms of Botulinum Toxin Type A Action on Pain. Toxins 2019, 11, 459. [Google Scholar] [CrossRef] [Green Version]
- Hallett, M.; Benecke, R.; Blitzer, A.; Comella, C.L. Treatment of focal dystonias with botulinum neurotoxin. Toxicon Off. J. Int. Soc. Toxinology 2009, 54, 628–633. [Google Scholar] [CrossRef] [Green Version]
- Dressler, D. Botulinum toxin for treatment of dystonia. Eur. J. Neurol. 2010, 17 (Suppl. 1), 88–96. [Google Scholar] [CrossRef] [PubMed]
- Sloop, R.R.; Cole, B.A.; Escutin, R.O. Human response to botulinum toxin injection: Type B compared with type A. Neurology 1997, 49, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Meunier, F.A.; Schiavo, G.; Molgó, J. Botulinum neurotoxins: From paralysis to recovery of functional neuromuscular transmission. J. Physiol. Paris 2002, 96, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.R. Evidence for antinociceptive activity of botulinum toxin type A in pain management. Headache 2003, 43 (Suppl. 1), S9–S15. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-M.; Chung, M.E. Botulinum Toxin for Neuropathic Pain: A Review of the Literature. Toxins 2015, 7, 3127–3154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollmer, M.A.; Magid, M.; Kruger, T.H.C.; Finzi, E. The Use of Botulinum Toxin for Treatment of Depression. Handb. Exp. Pharmacol. 2021, 263, 265–278. [Google Scholar] [CrossRef]
- Martina, E.; Diotallevi, F.; Radi, G.; Campanati, A.; Offidani, A. Therapeutic Use of Botulinum Neurotoxins in Dermatology: Systematic Review. Toxins 2021, 13, 120. [Google Scholar] [CrossRef]
- Simpson, L. The life history of a botulinum toxin molecule. Toxicon Off. J. Int. Soc. Toxinology 2013, 68, 40–59. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Kotiya, A.; Kiris, E.; Yang, M.; Bavari, S.; Tessarollo, L.; Oyler, G.A.; Weissman, A.M. Deubiquitinating enzyme VCIP135 dictates the duration of botulinum neurotoxin type A intoxication. Proc. Natl. Acad. Sci. USA 2017, 114, E5158–E5166. [Google Scholar] [CrossRef] [Green Version]
- Schurch, B.; Schmid, D.M.; Stöhrer, M. Treatment of neurogenic incontinence with botulinum toxin A. N. Engl. J. Med. 2000, 342, 665. [Google Scholar] [CrossRef]
- Schurch, B.; Stöhrer, M.; Kramer, G.; Schmid, D.M.; Gaul, G.; Hauri, D. Botulinum-A toxin for treating detrusor hyperreflexia in spinal cord injured patients: A new alternative to anticholinergic drugs? Preliminary results. J. Urol. 2000, 164, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Restani, L.; Novelli, E.; Bottari, D.; Leone, P.; Barone, I.; Galli-Resta, L.; Strettoi, E.; Caleo, M. Botulinum neurotoxin A impairs neurotransmission following retrograde transynaptic transport. Traffic 2012, 13, 1083–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anandan, C.; Jankovic, J. Botulinum Toxin in Movement Disorders: An Update. Toxins 2021, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Safarpour, Y.; Jabbari, B. Botulinum toxin treatment of pain syndromes—An evidence based review. Toxicon Off. J. Int. Soc. Toxinology 2018, 147, 120–128. [Google Scholar] [CrossRef]
- Goadsby, P.J.; Holland, P.R.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S. Pathophysiology of Migraine: A Disorder of Sensory Processing. Physiol. Rev. 2017, 97, 553–622. [Google Scholar] [CrossRef]
- Zychowska, M.; Rojewska, E.; Makuch, W.; Luvisetto, S.; Pavone, F.; Marinelli, S.; Przewlocka, B.; Mika, J. Participation of pro- and anti-nociceptive interleukins in botulinum toxin A-induced analgesia in a rat model of neuropathic pain. Eur. J. Pharmacol. 2016, 791, 377–388. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Liu, X.; Su, C.-J.; Zhang, Q.-L.; Wang, Z.-H.; Cao, L.-F.; Guo, X.-Y.; Huang, Y.; Luo, W.; et al. Antidepressant-Like Action of Single Facial Injection of Botulinum Neurotoxin A is Associated with Augmented 5-HT Levels and BDNF/ERK/CREB Pathways in Mouse Brain. Neurosci. Bull. 2019, 35, 661–672. [Google Scholar] [CrossRef]
- Yuan, H.; Silberstein, S.D. The Use of Botulinum Toxin in the Management of Headache Disorders. Handb. Exp. Pharmacol. 2021, 263, 227–249. [Google Scholar] [CrossRef]
- Brooks, V.B.; Curtis, D.R.; Eccles, J.C. The action of tetanus toxin on the inhibition of motoneurones. J. Physiol. 1957, 135, 655–672. [Google Scholar] [CrossRef] [Green Version]
- Bizzini, B.; Stoeckel, K.; Schwab, M. An antigenic polypeptide fragment isolated from tetanus toxin: Chemical characterization, binding to gangliosides and retrograde axonal transport in various neuron systems. J. Neurochem. 1977, 28, 529–542. [Google Scholar] [CrossRef]
- Holz, G.G.; Habener, J.F. Black widow spider alpha-latrotoxin: A presynaptic neurotoxin that shares structural homology with the glucagon-like peptide-1 family of insulin secretagogic hormones. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1998, 121, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.P.; Silva, V.A.O.; Silvestrini, A.V.P.; de Macedo, L.H.; Caetano, G.F.; Reis, R.M.; Mazzi, M.V. Crotoxin from Crotalus durissus terrificus venom: In vitro cytotoxic activity of a heterodimeric phospholipase A on human cancer-derived cell lines. Toxicon Off. J. Int. Soc. Toxinology 2018, 156, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, J.R.; Lancellotti, M.; Soares, A.M.; Calderon, L.A.; Ramírez, D.; González, W.; Marangoni, S.; Da Silva, S.L. CoaTx-II, a new dimeric Lys49 phospholipase A2 from Crotalus oreganus abyssus snake venom with bactericidal potential: Insights into its structure and biological roles. Toxicon Off. J. Int. Soc. Toxinology 2016, 120, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-L.; Han, R.; Gu, Z.-L.; Chen, Z.-X.; Chen, B.-W.; Reid, P.F.; Raymond, L.N.; Qin, Z.-H. A short-chain alpha-neurotoxin from Naja Naja atra produces potent cholinergic-dependent analgesia. Neurosci. Bull. 2006, 22, 103–109. [Google Scholar] [PubMed]
- Diaz, A.; Dickenson, A.H. Blockade of spinal N- and P-type, but not L-type, calcium channels inhibits the excitability of rat dorsal horn neurones produced by subcutaneous formalin inflammation. Pain 1997, 69, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Montagut-Bordas, C.; Dickenson, A.H. Calcium channel modulation as a target in chronic pain control. Br. J. Pharmacol. 2018, 175, 2173–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, K.M.; Binda, N.S.; Lavor, M.S.L.; Silva, C.M.O.; Rosado, I.R.; Gabellini, E.L.A.; Da Silva, J.F.; Oliveira, C.M.; Melo, M.M.; Gomez, M.V.; et al. Conotoxin MVIIA improves cell viability and antioxidant system after spinal cord injury in rats. PLoS ONE 2018, 13, e0204948. [Google Scholar] [CrossRef]
- Ovsepian, S.V.; LeBerre, M.; Steuber, V.; O’Leary, V.B.; Leibold, C.; Oliver Dolly, J. Distinctive role of KV1.1 subunit in the biology and functions of low threshold K(+) channels with implications for neurological disease. Pharmacol. Ther. 2016, 159, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Robbins, C.A.; Tempel, B.L. Kv1.1 and Kv1.2: Similar channels, different seizure models. Epilepsia 2012, 53 (Suppl. 1), 134–141. [Google Scholar] [CrossRef]
- Blacklow, B.; Kornhauser, R.; Hains, P.G.; Loiacono, R.; Escoubas, P.; Graudins, A.; Nicholson, G.M. α-Elapitoxin-Aa2a, a long-chain snake α-neurotoxin with potent actions on muscle (α1)(2)βγδ nicotinic receptors, lacks the classical high affinity for neuronal α7 nicotinic receptors. Biochem. Pharmacol. 2011, 81, 314–325. [Google Scholar] [CrossRef] [Green Version]
- Grozio, A.; Paleari, L.; Catassi, A.; Servent, D.; Cilli, M.; Piccardi, F.; Paganuzzi, M.; Cesario, A.; Granone, P.; Mourier, G.; et al. Natural agents targeting the alpha7-nicotinic-receptor in NSCLC: A promising prospective in anti-cancer drug development. Int. J. Cancer 2008, 122, 1911–1915. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.-n.; Liu, Y.-l.; Lin, H.-m.; Yang, S.-l.; Feng, Y.-l.; Reid, P.F.; Qin, Z.-h. Involvement of cholinergic system in suppression of formalin-induced inflammatory pain by cobratoxin. Acta Pharmacol. Sin. 2011, 32, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Webb, R.P. Engineering of Botulinum Neurotoxins for Biomedical Applications. Toxins 2018, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Scott, A.B. Botulinum toxin injection into extraocular muscles as an alternative to strabismus surgery. J. Pediatr. Ophthalmol. Strabismus 1980, 17, 21–25. [Google Scholar] [CrossRef]
- Jankovic, J. Botulinum toxin in clinical practice. J. Neurol. Neurosurg. Psychiatry 2004, 75, 951–957. [Google Scholar] [CrossRef] [PubMed]
- O’Flynn, L.C.; Simonyan, K. Short- and Long-term Central Action of Botulinum Neurotoxin Treatment in Laryngeal Dystonia. Neurology 2022, 99, e1178–e1190. [Google Scholar] [CrossRef]
- Eaker, E.Y.; Gordon, J.M.; Vogel, S.B. Untoward effects of esophageal botulinum toxin injection in the treatment of achalasia. Dig. Dis. Sci. 1997, 42, 724–727. [Google Scholar] [CrossRef]
- Pasricha, P.J.; Miskovsky, E.P.; Kalloo, A.N. Intrasphincteric injection of botulinum toxin for suspected sphincter of Oddi dysfunction. Gut 1994, 35, 1319–1321. [Google Scholar] [CrossRef] [Green Version]
- Joo, J.S.; Agachan, F.; Wolff, B.; Nogueras, J.J.; Wexner, S.D. Initial North American experience with botulinum toxin type A for treatment of anismus. Dis. Colon Rectum 1996, 39, 1107–1111. [Google Scholar] [CrossRef]
- Zermann, D.h.; Ishigooka, M.; Schubert, J.; Schmidt, R.A. Perisphincteric injection of botulinum toxin type A. A treatment option for patients with chronic prostatic pain? Eur. Urol. 2000, 38, 393–399. [Google Scholar] [CrossRef]
- Naumann, M.; Jost, W.H.; Toyka, K.V. Botulinum toxin in the treatment of neurological disorders of the autonomic nervous system. Arch. Neurol. 1999, 56, 914–916. [Google Scholar] [CrossRef] [Green Version]
- Saadia, D.; Voustianiouk, A.; Wang, A.K.; Kaufmann, H. Botulinum toxin type A in primary palmar hyperhidrosis: Randomized, single-blind, two-dose study. Neurology 2001, 57, 2095–2099. [Google Scholar] [CrossRef]
- Naumann, M.K.; Hamm, H.; Lowe, N.J. Effect of botulinum toxin type A on quality of life measures in patients with excessive axillary sweating: A randomized controlled trial. Br. J. Dermatol. 2002, 147, 1218–1226. [Google Scholar] [CrossRef]
- Heckmann, M.; Ceballos-Baumann, A.O.; Plewig, G. Botulinum toxin A for axillary hyperhidrosis (excessive sweating). N. Engl. J. Med. 2001, 344, 488–493. [Google Scholar] [CrossRef]
- Frampton, J.E.; Easthope, S.E. Botulinum toxin A (Botox Cosmetic): A review of its use in the treatment of glabellar frown lines. Am. J. Clin. Dermatol. 2003, 4, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Matarasso, A.; Matarasso, S.L.; Brandt, F.S.; Bellman, B. Botulinum A exotoxin for the management of platysma bands. Plast. Reconstr. Surg. 1999, 103, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Hoorens, I.; Ongenae, K. Primary focal hyperhidrosis: Current treatment options and a step-by-step approach. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 1–8. [Google Scholar] [CrossRef]
- Lakraj, A.-A.D.; Moghimi, N.; Jabbari, B. Hyperhidrosis: Anatomy, pathophysiology and treatment with emphasis on the role of botulinum toxins. Toxins 2013, 5, 821–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shon, U.; Kim, M.H.; Lee, D.Y.; Kim, S.H.; Park, B.C. The effect of intradermal botulinum toxin on androgenetic alopecia and its possible mechanism. J. Am. Acad. Dermatol. 2020, 83, 1838–1839. [Google Scholar] [CrossRef]
- Shackleton, T.; Ram, S.; Black, M.; Ryder, J.; Clark, G.T.; Enciso, R. The efficacy of botulinum toxin for the treatment of trigeminal and postherpetic neuralgia: A systematic review with meta-analyses. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2016, 122, 61–71. [Google Scholar] [CrossRef]
- Meng, F.; Peng, K.; Yang, J.-P.; Ji, F.-H.; Xia, F.; Meng, X.-W. Botulinum toxin-A for the treatment of neuralgia: A systematic review and meta-analysis. J. Pain Res. 2018, 11, 2343–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Park, H.J. Botulinum Toxin for the Treatment of Neuropathic Pain. Toxins 2017, 9, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, Y.; Matsuka, Y.; Spigelman, I.; Ishihara, Y.; Yamamoto, Y.; Sonoyama, W.; Kamioka, H.; Yamashiro, T.; Kuboki, T.; Oguma, K. Botulinum toxin type a (150 kDa) decreases exaggerated neurotransmitter release from trigeminal ganglion neurons and relieves neuropathy behaviors induced by infraorbital nerve constriction. Neuroscience 2009, 159, 1422–1429. [Google Scholar] [CrossRef]
- Meng, J.; Dolly, J.O.; Wang, J. Selective cleavage of SNAREs in sensory neurons unveils protein complexes mediating peptide exocytosis triggered by different stimuli. Mol. Neurobiol. 2014, 50, 574–588. [Google Scholar] [CrossRef]
- Matak, I.; Rossetto, O.; Lacković, Z. Botulinum toxin type A selectivity for certain types of pain is associated with capsaicin-sensitive neurons. Pain 2014, 155, 1516–1526. [Google Scholar] [CrossRef] [Green Version]
- Mika, J.; Rojewska, E.; Makuch, W.; Korostynski, M.; Luvisetto, S.; Marinelli, S.; Pavone, F.; Przewlocka, B. The effect of botulinum neurotoxin A on sciatic nerve injury-induced neuroimmunological changes in rat dorsal root ganglia and spinal cord. Neuroscience 2011, 175, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, A.; Popiolek-Barczyk, K.; Pavone, F.; Mika, J. Comparison of the Expression Changes after Botulinum Toxin Type A and Minocycline Administration in Lipopolysaccharide-Stimulated Rat Microglial and Astroglial Cultures. Front. Cell. Infect. Microbiol. 2017, 7, 141. [Google Scholar] [CrossRef] [Green Version]
- Finzi, E.; Wasserman, E. Treatment of depression with botulinum toxin A: A case series. Dermatol. Surg. 2006, 32, 645–650. [Google Scholar] [CrossRef]
- Wollmer, M.A.; de Boer, C.; Kalak, N.; Beck, J.; Götz, T.; Schmidt, T.; Hodzic, M.; Bayer, U.; Kollmann, T.; Kollewe, K.; et al. Facing depression with botulinum toxin: A randomized controlled trial. J. Psychiatr. Res. 2012, 46, 574–581. [Google Scholar] [CrossRef] [Green Version]
- Wollmer, M.A.; Kalak, N.; Jung, S.; de Boer, C.; Magid, M.; Reichenberg, J.S.; Brand, S.; Holsboer-Trachsler, E.; Kruger, T.H.C. Agitation predicts response of depression to botulinum toxin treatment in a randomized controlled trial. Front. Psychiatry 2014, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Finzi, E.; Rosenthal, N.E. Treatment of depression with onabotulinumtoxinA: A randomized, double-blind, placebo controlled trial. J. Psychiatr. Res. 2014, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Magid, M.; Reichenberg, J.S.; Poth, P.E.; Robertson, H.T.; LaViolette, A.K.; Kruger, T.H.C.; Wollmer, M.A. Treatment of major depressive disorder using botulinum toxin A: A 24-week randomized, double-blind, placebo-controlled study. J. Clin. Psychiatry 2014, 75, 837–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanian, A.; Ghanbari Jolfaei, A.; Mehran, G.; Azizian, Z. Efficacy of Botox versus Placebo for Treatment of Patients with Major Depression. Iran J. Public Health 2017, 46, 982–984. [Google Scholar] [PubMed]
- Finzi, E.; Rosenthal, N.E. Emotional proprioception: Treatment of depression with afferent facial feedback. J. Psychiatr. Res. 2016, 80, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Ibragić, S.; Matak, I.; Dračić, A.; Smajlović, A.; Muminović, M.; Proft, F.; Sofić, E.; Lacković, Z.; Riederer, P. Effects of botulinum toxin type A facial injection on monoamines and their metabolites in sensory, limbic and motor brain regions in rats. Neurosci. Lett. 2016, 617, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Caleo, M.; Schiavo, G. Central effects of tetanus and botulinum neurotoxins. Toxicon Off. J. Int. Soc. Toxinology 2009, 54, 593–599. [Google Scholar] [CrossRef]
- Marchand-Pauvert, V.; Aymard, C.; Giboin, L.-S.; Dominici, F.; Rossi, A.; Mazzocchio, R. Beyond muscular effects: Depression of spinal recurrent inhibition after botulinum neurotoxin A. J. Physiol. 2013, 591, 1017–1029. [Google Scholar] [CrossRef] [Green Version]
- Golden, S.A.; Christoffel, D.J.; Heshmati, M.; Hodes, G.E.; Magida, J.; Davis, K.; Cahill, M.E.; Dias, C.; Ribeiro, E.; Ables, J.L.; et al. Epigenetic regulation of RAC1 induces synaptic remodeling in stress disorders and depression. Nat. Med. 2013, 19, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Molendijk, M.L.; Spinhoven, P.; Polak, M.; Bus, B.A.A.; Penninx, B.W.J.H.; Elzinga, B.M. Serum BDNF concentrations as peripheral manifestations of depression: Evidence from a systematic review and meta-analyses on 179 associations (N = 9484). Mol. Psychiatry 2014, 19, 791–800. [Google Scholar] [CrossRef]
- Guo, F.; Zhang, Q.; Zhang, B.; Fu, Z.; Wu, B.; Huang, C.; Li, Y. Burst-firing patterns in the prefrontal cortex underlying the neuronal mechanisms of depression probed by antidepressants. Eur. J. Neurosci. 2014, 40, 3538–3547. [Google Scholar] [CrossRef]
- Pilar-Cuéllar, F.; Vidal, R.; Díaz, A.; Castro, E.; dos Anjos, S.; Pascual-Brazo, J.; Linge, R.; Vargas, V.; Blanco, H.; Martínez-Villayandre, B.; et al. Neural plasticity and proliferation in the generation of antidepressant effects: Hippocampal implication. Neural Plast. 2013, 2013, 537265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gascón, S.; Ortega, F.; Götz, M. Transient CREB-mediated transcription is key in direct neuronal reprogramming. Neurogenesis 2017, 4, e1285383. [Google Scholar] [CrossRef] [PubMed]
- Einoch, R.; Weinreb, O.; Mandiuk, N.; Youdim, M.B.H.; Bilker, W.; Silver, H. The involvement of BDNF-CREB signaling pathways in the pharmacological mechanism of combined SSRI- antipsychotic treatment in schizophrenia. Eur. Neuropsychopharmacol. 2017, 27, 470–483. [Google Scholar] [CrossRef] [PubMed]
- GBD 2016 DALYs and HALE Collaborators. Global, regional, and national disability-adjusted life-years (DALYs) for 333 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1260–1344. [Google Scholar] [CrossRef] [PubMed]
- Vollesen, A.L.; Benemei, S.; Cortese, F.; Labastida-Ramírez, A.; Marchese, F.; Pellesi, L.; Romoli, M.; Ashina, M.; Lampl, C. Migraine and cluster headache—The common link. J. Headache Pain 2018, 19, 89. [Google Scholar] [CrossRef] [Green Version]
- Durham, P.L.; Cady, R.; Cady, R. Regulation of calcitonin gene-related peptide secretion from trigeminal nerve cells by botulinum toxin type A: Implications for migraine therapy. Headache 2004, 44, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Welch, M.J.; Purkiss, J.R.; Foster, K.A. Sensitivity of embryonic rat dorsal root ganglia neurons to Clostridium botulinum neurotoxins. Toxicon Off. J. Int. Soc. Toxinology 2000, 38, 245–258. [Google Scholar] [CrossRef]
- Cui, M.; Khanijou, S.; Rubino, J.; Aoki, K.R. Subcutaneous administration of botulinum toxin A reduces formalin-induced pain. Pain 2004, 107, 125–133. [Google Scholar] [CrossRef]
- Xiao, L.; Cheng, J.; Dai, J.; Zhang, D. Botulinum toxin decreases hyperalgesia and inhibits P2X3 receptor over-expression in sensory neurons induced by ventral root transection in rats. Pain Med. 2011, 12, 1385–1394. [Google Scholar] [CrossRef] [Green Version]
- Hong, B.; Yao, L.; Ni, L.; Wang, L.; Hu, X. Antinociceptive effect of botulinum toxin A involves alterations in AMPA receptor expression and glutamate release in spinal dorsal horn neurons. Neuroscience 2017, 357, 197–207. [Google Scholar] [CrossRef]
- Shimizu, T.; Shibata, M.; Toriumi, H.; Iwashita, T.; Funakubo, M.; Sato, H.; Kuroi, T.; Ebine, T.; Koizumi, K.; Suzuki, N. Reduction of TRPV1 expression in the trigeminal system by botulinum neurotoxin type-A. Neurobiol. Dis. 2012, 48, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Strassman, A.M.; Novack, V.; Brin, M.F.; Burstein, R. Extracranial injections of botulinum neurotoxin type A inhibit intracranial meningeal nociceptors’ responses to stimulation of TRPV1 and TRPA1 channels: Are we getting closer to solving this puzzle? Cephalalgia 2016, 36, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Do, T.P.; Hvedstrup, J.; Schytz, H.W. Botulinum toxin: A review of the mode of action in migraine. Acta Neurol. Scand. 2018, 137, 442–451. [Google Scholar] [CrossRef]
- Dillingham, T.R. Musculoskeletal rehabilitation: Current understandings and future directions. Am. J. Phys. Med. Rehabil. 2007, 86, S19–S28. [Google Scholar] [CrossRef] [PubMed]
- Sasse, A.; Conduit, R.; Ryan, D.; Woods, W.; Tucker, A.P. A pharmacotherapy for obstructive sleep apnea. Sleep 2005, 28, 1015–1016. [Google Scholar] [CrossRef]
- Chen, C.; Baldwin, M.R.; Barbieri, J.T. Molecular basis for tetanus toxin coreceptor interactions. Biochemistry 2008, 47, 7179–7186. [Google Scholar] [CrossRef]
- Dimpfel, W.; Huang, R.T.; Habermann, E. Gangliosides in nervous tissue cultures and binding of 125I-labelled tetanus toxin, a neuronal marker. J. Neurochem. 1977, 29, 329–334. [Google Scholar] [CrossRef]
- Herreros, J.; Ng, T.; Schiavo, G. Lipid rafts act as specialized domains for tetanus toxin binding and internalization into neurons. Mol. Biol. Cell. 2001, 12, 2947–2960. [Google Scholar] [CrossRef] [Green Version]
- Rogers, T.B.; Snyder, S.H. High affinity binding of tetanus toxin to mammalian brain membranes. J. Biol. Chem. 1981, 256, 2402–2407. [Google Scholar] [CrossRef]
- Walton, K.M.; Sandberg, K.; Rogers, T.B.; Schnaar, R.L. Complex ganglioside expression and tetanus toxin binding by PC12 pheochromocytoma cells. J. Biol. Chem. 1988, 263, 2055–2063. [Google Scholar] [CrossRef]
- Lalli, G.; Gschmeissner, S.; Schiavo, G. Myosin Va and microtubule-based motors are required for fast axonal retrograde transport of tetanus toxin in motor neurons. J. Cell. Sci. 2003, 116, 4639–4650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deinhardt, K.; Salinas, S.; Verastegui, C.; Watson, R.; Worth, D.; Hanrahan, S.; Bucci, C.; Schiavo, G. Rab5 and Rab7 control endocytic sorting along the axonal retrograde transport pathway. Neuron 2006, 52, 293–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafezparast, M.; Klocke, R.; Ruhrberg, C.; Marquardt, A.; Ahmad-Annuar, A.; Bowen, S.; Lalli, G.; Witherden, A.S.; Hummerich, H.; Nicholson, S.; et al. Mutations in dynein link motor neuron degeneration to defects in retrograde transport. Science 2003, 300, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, J.M.; Oliván, S.; Osta, R. Tetanus toxin C-fragment: The courier and the cure? Toxins 2010, 2, 2622–2644. [Google Scholar] [CrossRef]
- Bayart, C.; Mularoni, A.; Hemmani, N.; Kerachni, S.; Jose, J.; Gouet, P.; Paladino, J.; Le Borgne, M. Tetanus Toxin Fragment C: Structure, Drug Discovery Research and Production. Pharmaceuticals 2022, 15, 756. [Google Scholar] [CrossRef]
- Moreno-Martinez, L.; de la Torre, M.; Muñoz, M.J.; Zaragoza, P.; Aguilera, J.; Calvo, A.C.; Osta, R. Neuroprotective Fragment C of Tetanus Toxin Modulates IL-6 in an ALS Mouse Model. Toxins 2020, 12, 330. [Google Scholar] [CrossRef]
- Moreno-Galarza, N.; Mendieta, L.; Palafox-Sánchez, V.; Herrando-Grabulosa, M.; Gil, C.; Limón, D.I.; Aguilera, J. Peripheral Administration of Tetanus Toxin Hc Fragment Prevents MPP Toxicity In Vivo. Neurotox. Res. 2018, 34, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Patricio-Martínez, A.; Mendieta, L.; Martínez, I.; Aguilera, J.; Limón, I.D. The recombinant C-terminal fragment of tetanus toxin protects against cholinotoxicity by intraseptal injection of β-amyloid peptide (25–35) in rats. Neuroscience 2016, 315, 18–30. [Google Scholar] [CrossRef]
- Duggan, M.J.; Quinn, C.P.; Chaddock, J.A.; Purkiss, J.R.; Alexander, F.C.G.; Doward, S.; Fooks, S.J.; Friis, L.M.; Hall, Y.H.J.; Kirby, E.R.; et al. Inhibition of release of neurotransmitters from rat dorsal root ganglia by a novel conjugate of a Clostridium botulinum toxin A endopeptidase fragment and Erythrina cristagalli lectin. J. Biol. Chem. 2002, 277, 34846–34852. [Google Scholar] [CrossRef] [Green Version]
- Chaddock, J.A.; Purkiss, J.R.; Alexander, F.C.G.; Doward, S.; Fooks, S.J.; Friis, L.M.; Hall, Y.H.J.; Kirby, E.R.; Leeds, N.; Moulsdale, H.J.; et al. Retargeted clostridial endopeptidases: Inhibition of nociceptive neurotransmitter release in vitro, and antinociceptive activity in in vivo models of pain. Mov. Disord. 2004, 19 (Suppl. 8), S42–S47. [Google Scholar] [CrossRef]
- Mangione, A.S.; Obara, I.; Maiarú, M.; Geranton, S.M.; Tassorelli, C.; Ferrari, E.; Leese, C.; Davletov, B.; Hunt, S.P. Nonparalytic botulinum molecules for the control of pain. Pain 2016, 157, 1045–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsenault, J.; Ferrari, E.; Niranjan, D.; Cuijpers, S.A.G.; Gu, C.; Vallis, Y.; O’Brien, J.; Davletov, B. Stapling of the botulinum type A protease to growth factors and neuropeptides allows selective targeting of neuroendocrine cells. J. Neurochem. 2013, 126, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zurawski, T.H.; Meng, J.; Lawrence, G.W.; Aoki, K.R.; Wheeler, L.; Dolly, J.O. Novel chimeras of botulinum and tetanus neurotoxins yield insights into their distinct sites of neuroparalysis. FASEB J. 2012, 26, 5035–5048. [Google Scholar] [CrossRef] [PubMed]
- Antoniazzi, C.; Belinskaia, M.; Zurawski, T.; Kaza, S.K.; Dolly, J.O.; Lawrence, G.W. Botulinum Neurotoxin Chimeras Suppress Stimulation by Capsaicin of Rat Trigeminal Sensory Neurons In Vivo and In Vitro. Toxins 2022, 14, 116. [Google Scholar] [CrossRef]
- Saez, N.J.; Senff, S.; Jensen, J.E.; Er, S.Y.; Herzig, V.; Rash, L.D.; King, G.F. Spider-venom peptides as therapeutics. Toxins 2010, 2, 2851–2871. [Google Scholar] [CrossRef] [Green Version]
- Gejl, K.D.; Ørtenblad, N.; Andersson, E.; Plomgaard, P.; Holmberg, H.-C.; Nielsen, J. Local depletion of glycogen with supramaximal exercise in human skeletal muscle fibres. J. Physiol. 2017, 595, 2809–2821. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.J.; Garcia, M.L. Therapeutic potential of venom peptides. Nat. Rev. Drug Discov. 2003, 2, 790–802. [Google Scholar] [CrossRef]
- Perry, T.; Greig, N.H. The glucagon-like peptides: A double-edged therapeutic sword? Trends Pharmacol. Sci. 2003, 24, 377–383. [Google Scholar] [CrossRef]
- Sobrinho, J.C.; Simões-Silva, R.; Holanda, R.J.; Alfonso, J.; Gomez, A.F.; Zanchi, F.B.; Moreira-Dill, L.S.; Grabner, A.N.; Zuliani, J.P.; Calderon, L.A.; et al. Antitumoral Potential of Snake Venom Phospholipases A2 and Synthetic Peptides. Curr. Pharm. Biotechnol. 2016, 17, 1201–1212. [Google Scholar] [CrossRef]
- DiPaola, R.S. To arrest or not to G(2)-M Cell-cycle arrest: Commentary re: A. K. Tyagi et al., Silibinin strongly synergizes human prostate carcinoma DU145 cells to doxorubicin-induced growth inhibition, G(2)-M arrest, and apoptosis. Clin. Cancer Res. 2002, 8, 3512–3519. [Google Scholar]
- Kato, E.E.; Sampaio, S.C. Crotoxin Modulates Events Involved in Epithelial-Mesenchymal Transition in 3D Spheroid Model. Toxins 2021, 13, 830. [Google Scholar] [CrossRef] [PubMed]
- Donato, N.J.; Martin, C.A.; Perez, M.; Newman, R.A.; Vidal, J.C.; Etcheverry, M. Regulation of epidermal growth factor receptor activity by crotoxin, a snake venom phospholipase A2 toxin. A novel growth inhibitory mechanism. Biochem. Pharmacol. 1996, 51, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qin, X.; Zhang, Z.; Chen, M.; Wang, Y.; Gao, B. Crotoxin suppresses the tumorigenic properties and enhances the antitumor activity of Iressa® (gefinitib) in human lung adenocarcinoma SPCA-1 cells. Mol. Med. Rep. 2014, 10, 3009–3014. [Google Scholar] [CrossRef] [Green Version]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics, 2010. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Liang, H.; Qin, Z.-h.; Liu, C.-y. Crotoxin induces apoptosis and autophagy in human lung carcinoma cells in vitro via activation of the p38MAPK signaling pathway. Acta Pharmacol. Sin. 2014, 35, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Cura, J.E.; Blanzaco, D.P.; Brisson, C.; Cura, M.A.; Cabrol, R.; Larrateguy, L.; Mendez, C.; Sechi, J.C.; Silveira, J.S.; Theiller, E.; et al. Phase I and pharmacokinetics study of crotoxin (cytotoxic PLA(2), NSC-624244) in patients with advanced cancer. Clin. Cancer Res. 2002, 8, 1033–1041. [Google Scholar] [PubMed]
- Zhang, H.-L.; Han, R.; Chen, Z.-X.; Chen, B.-W.; Gu, Z.-L.; Reid, P.F.; Raymond, L.N.; Qin, Z.-H. Opiate and acetylcholine-independent analgesic actions of crotoxin isolated from Crotalus durissus terrificus venom. Toxicon Off. J. Int. Soc. Toxinology 2006, 48, 175–182. [Google Scholar] [CrossRef]
- Zhu, Q.; Wu, D.-C.; Zhou, X.-P.; Gong, S.; Cheng, B.-C.; Qin, Z.-H.; Reid, P.F.; Yin, Q.-Z.; Jiang, X.-H. Inhibitory effect of crotoxin on the pain-evoked discharge of neurons in thalamic parafascicular nucleus in rats. Toxicon Off. J. Int. Soc. Toxinology 2008, 51, 102–111. [Google Scholar] [CrossRef]
- Millan, M.J. N-methyl-D-aspartate receptor-coupled glycine B receptors in the pathogenesis and treatment of schizophrenia: A critical review. Curr. Drug Targets CNS Neurol. Disord. 2002, 1, 191–213. [Google Scholar] [CrossRef]
- Santos, I.A.; Shimizu, J.F.; de Oliveira, D.M.; Martins, D.O.S.; Cardoso-Sousa, L.; Cintra, A.C.O.; Aquino, V.H.; Sampaio, S.V.; Nicolau-Junior, N.; Sabino-Silva, R.; et al. Chikungunya virus entry is strongly inhibited by phospholipase A2 isolated from the venom of Crotalus durissus terrificus. Sci. Rep. 2021, 11, 8717. [Google Scholar] [CrossRef]
- Trevisan, G.; Oliveira, S.M. Animal Venom Peptides Cause Antinociceptive Effects by Voltage-gated Calcium Channels Activity Blockage. Curr. Neuropharmacol. 2022, 20, 1579–1599. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Luo, Z.D. Calcium channel functions in pain processing. Channels 2010, 4, 510–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmberg, A.B.; Yaksh, T.L. Antinociception produced by spinal delivery of the S and R enantiomers of flurbiprofen in the formalin test. Eur. J. Pharmacol. 1994, 256, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Livett, B.G.; Gayler, K.R.; Khalil, Z. Drugs from the sea: Conopeptides as potential therapeutics. Curr. Med. Chem. 2004, 11, 1715–1723. [Google Scholar] [CrossRef]
- Twede, V.D.; Miljanich, G.; Olivera, B.M.; Bulaj, G. Neuroprotective and cardioprotective conopeptides: An emerging class of drug leads. Curr. Opin. Drug Discov. Devel. 2009, 12, 231–239. [Google Scholar]
- Schroeder, C.I.; Smythe, M.L.; Lewis, R.J. Development of small molecules that mimic the binding of omega-conotoxins at the N-type voltage-gated calcium channel. Mol. Divers 2004, 8, 127–134. [Google Scholar] [CrossRef]
- Schmidtko, A.; Lötsch, J.; Freynhagen, R.; Geisslinger, G. Ziconotide for treatment of severe chronic pain. Lancet 2010, 375, 1569–1577. [Google Scholar] [CrossRef]
- Niederberger, E.; Schmidtko, A.; Rothstein, J.D.; Geisslinger, G.; Tegeder, I. Modulation of spinal nociceptive processing through the glutamate transporter GLT-1. Neuroscience 2003, 116, 81–87. [Google Scholar] [CrossRef]
- Carlson, G.D.; Gorden, C. Current developments in spinal cord injury research. Spine J. 2002, 2, 116–128. [Google Scholar] [CrossRef]
- Hall, E.D.; Springer, J.E. Neuroprotection and acute spinal cord injury: A reappraisal. NeuroRx 2004, 1, 80–100. [Google Scholar] [CrossRef]
- Lu, J.; Ashwell, K.W.; Waite, P. Advances in secondary spinal cord injury: Role of apoptosis. Spine 2000, 25, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Isaac, L.; Pejic, L. Secondary mechanisms of spinal cord injury. Surg. Neurol. 1995, 43, 484–485. [Google Scholar] [CrossRef] [PubMed]
- Amar, A.P.; Levy, M.L. Pathogenesis and pharmacological strategies for mitigating secondary damage in acute spinal cord injury. Neurosurgery 1999, 44, 1027–1039. [Google Scholar] [CrossRef]
- Liu, W.-M.; Wu, J.-Y.; Li, F.-C.; Chen, Q.-X. Ion channel blockers and spinal cord injury. J. Neurosci. Res 2011, 89, 791–801. [Google Scholar] [CrossRef]
- Naderi, A.; Asgari, A.R.; Zahed, R.; Ghanbari, A.; Samandari, R.; Jorjani, M. Estradiol attenuates spinal cord injury-related central pain by decreasing glutamate levels in thalamic VPL nucleus in male rats. Metab. Brain Dis. 2014, 29, 763–770. [Google Scholar] [CrossRef]
- Gonçaves, J.M.; Ferreira, J.; Prado, M.A.M.; Cordeiro, M.N.; Richardson, M.; Pinheiro, A.C.d.N.; Silva, M.A.R.; Junior, C.J.d.C.; Souza, A.H.; Gomez, M.V. The effect of spider toxin PhTx3-4, ω-conotoxins MVIIA and MVIIC on glutamate uptake and on capsaicin-induced glutamate release and [Ca2+]i in spinal cord synaptosomes. Cell Mol. Neurobiol. 2011, 31, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, S.; Matsushima, K.; Fujita, H.; Nanri, K.; Ogawa, S.; Shinohara, Y. A selective N-type calcium channel antagonist reduces extracellular glutamate release and infarct volume in focal cerebral ischemia. J. Cereb. Blood Flow. Metab. 1995, 15, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Verweij, B.H.; Muizelaar, J.P.; Vinas, F.C.; Peterson, P.L.; Xiong, Y.; Lee, C.P. Mitochondrial dysfunction after experimental and human brain injury and its possible reversal with a selective N-type calcium channel antagonist (SNX-111). Neurol. Res. 1997, 19, 334–339. [Google Scholar] [CrossRef]
- Verweij, B.H.; Muizelaar, J.P.; Vinas, F.C.; Peterson, P.L.; Xiong, Y.; Lee, C.P. Improvement in mitochondrial dysfunction as a new surrogate efficiency measure for preclinical trials: Dose-response and time-window profiles for administration of the calcium channel blocker Ziconotide in experimental brain injury. J. Neurosurg. 2000, 93, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Peterson, P.L.; Verweij, B.H.; Vinas, F.C.; Muizelaar, J.P.; Lee, C.P. Mitochondrial dysfunction after experimental traumatic brain injury: Combined efficacy of SNX-111 and U-101033E. J. Neurotrauma 1998, 15, 531–544. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Yu, S.; Liu, J.; Chen, R.; Zhang, Y.; Jiang, L.; Dai, Q. A novel -conotoxin Bu8 inhibiting N-type voltage-gated calcium channels displays potent analgesic activity. Acta Pharm. Sin. B 2021, 11, 2685–2693. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.R.; Richbart, S.D.; Merritt, J.C.; Brown, K.C.; Nolan, N.A.; Akers, A.T.; Lau, J.K.; Robateau, Z.R.; Miles, S.L.; Dasgupta, P. Acetylcholine signaling system in progression of lung cancers. Pharmacol. Ther. 2019, 194, 222–254. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Catassi, A.; Cesario, A.; Servent, D. Development of novel therapeutic strategies for lung cancer: Targeting the cholinergic system. Curr. Med. Chem. 2006, 13, 3493–3512. [Google Scholar] [CrossRef] [PubMed]
- Trombino, S.; Cesario, A.; Margaritora, S.; Granone, P.; Motta, G.; Falugi, C.; Russo, P. Alpha7-nicotinic acetylcholine receptors affect growth regulation of human mesothelioma cells: Role of mitogen-activated protein kinase pathway. Cancer Res. 2004, 64, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesario, A.; Russo, P.; Viaggi, S.; Trombino, S.; Imperatori, A.; Margaritora, S.; Dominioni, L.; Festi, L.; Porziella, V.; Granone, P. Malignant pleural mesothelioma: Time for translational research. Lancet Oncol. 2004, 5, 591. [Google Scholar] [CrossRef] [PubMed]
- Trombino, S.; Bisio, A.; Catassi, A.; Cesario, A.; Falugi, C.; Russo, P. Role of the non-neuronal human cholinergic system in lung cancer and mesothelioma: Possibility of new therapeutic strategies. Curr. Med. Chem. Anticancer. Agents 2004, 4, 535–542. [Google Scholar] [CrossRef] [PubMed]
- West, K.A.; Brognard, J.; Clark, A.S.; Linnoila, I.R.; Yang, X.; Swain, S.M.; Harris, C.; Belinsky, S.; Dennis, P.A. Rapid Akt activation by nicotine and a tobacco carcinogen modulates the phenotype of normal human airway epithelial cells. J. Clin. Invest. 2003, 111, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, P.; Kinkade, R.; Joshi, B.; Decook, C.; Haura, E.; Chellappan, S. Nicotine inhibits apoptosis induced by chemotherapeutic drugs by up-regulating XIAP and survivin. Proc. Natl. Acad. Sci. USA 2006, 103, 6332–6337. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, Y.; Jiang, D.; Reid, P.F.; Jiang, X.; Qin, Z.; Tao, J. Alpha-cobratoxin inhibits T-type calcium currents through muscarinic M4 receptor and Gο-protein βγ subunits-dependent protein kinase A pathway in dorsal root ganglion neurons. Neuropharmacology 2012, 62, 1062–1072. [Google Scholar] [CrossRef]
- Xu, L.; Wang, S.; Zhang, L.; Liu, B.; Zheng, S.; Yao, M. Cobratoxin Alleviates Cancer-Induced Bone Pain in Rats via Inhibiting CaMKII Signaling Pathway after Acting on M4 Muscarinic Cholinergic Receptors. ACS Chem. Neurosci. 2022, 13, 1422–1432. [Google Scholar] [CrossRef]
- Gazerani, P.; Cairns, B.E. Venom-based biotoxins as potential analgesics. Expert Rev. Neurother. 2014, 14, 1261–1274. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Huang, J.; Wang, S.-Z.; Qin, Z.-H.; Lin, F. Cobrotoxin extracted from Naja atra venom relieves arthritis symptoms through anti-inflammation and immunosuppression effects in rat arthritis model. J. Ethnopharmacol. 2016, 194, 1087–1095. [Google Scholar] [CrossRef]
- Bordon, K.d.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.-Q.; Qian, X.-Y.; An, J.-X.; Liu, C.-C.; Fang, Q.-W.; Wang, Y.; Jiang, Y.-D.; Cope, D.K.; Williams, J.P. Rat Model of Trigeminal Neuralgia Using Cobra Venom Mimics the Electron Microscopy, Behavioral, and Anticonvulsant Drug Responses Seen in Patients. Pain Physician 2015, 18, E1083–E1090. [Google Scholar] [PubMed]
- Zhao, C.; Zhao, J.; Yang, Q.; Ye, Y. Cobra neurotoxin produces central analgesic and hyperalgesic actions via adenosine A and A receptors. Mol. Pain 2017, 13, 1744806917720336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, F.; Reid, P.F.; Qin, Z.-H. Cobrotoxin could be an effective therapeutic for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1258–1260. [Google Scholar] [CrossRef]
- Park, M.H.; Song, H.S.; Kim, K.H.; Son, D.J.; Lee, S.H.; Yoon, D.Y.; Kim, Y.; Park, I.Y.; Song, S.; Hwang, B.Y.; et al. Cobrotoxin inhibits NF-kappa B activation and target gene expression through reaction with NF-kappa B signal molecules. Biochemistry 2005, 44, 8326–8336. [Google Scholar] [CrossRef]
Figure 1.
This is a synaptic structure. The top left is the normal release of neurotransmitters from synapses in response to calcium stimulation; in the middle, botulinum toxin and α-LTX affect neurotransmitter release by acting on synapses (created with BioRender.com, accessed on 20 December 2022). SNARE: soluble N-ethylmaleimide-sensitive factor attachment protein receptor.
Figure 1.
This is a synaptic structure. The top left is the normal release of neurotransmitters from synapses in response to calcium stimulation; in the middle, botulinum toxin and α-LTX affect neurotransmitter release by acting on synapses (created with BioRender.com, accessed on 20 December 2022). SNARE: soluble N-ethylmaleimide-sensitive factor attachment protein receptor.

Figure 2.
Different subgroups of α-conotoxins are selective for different subtypes of AChRs. (Created with BioRender.com, accessed on 29 October 2022).
Figure 2.
Different subgroups of α-conotoxins are selective for different subtypes of AChRs. (Created with BioRender.com, accessed on 29 October 2022).


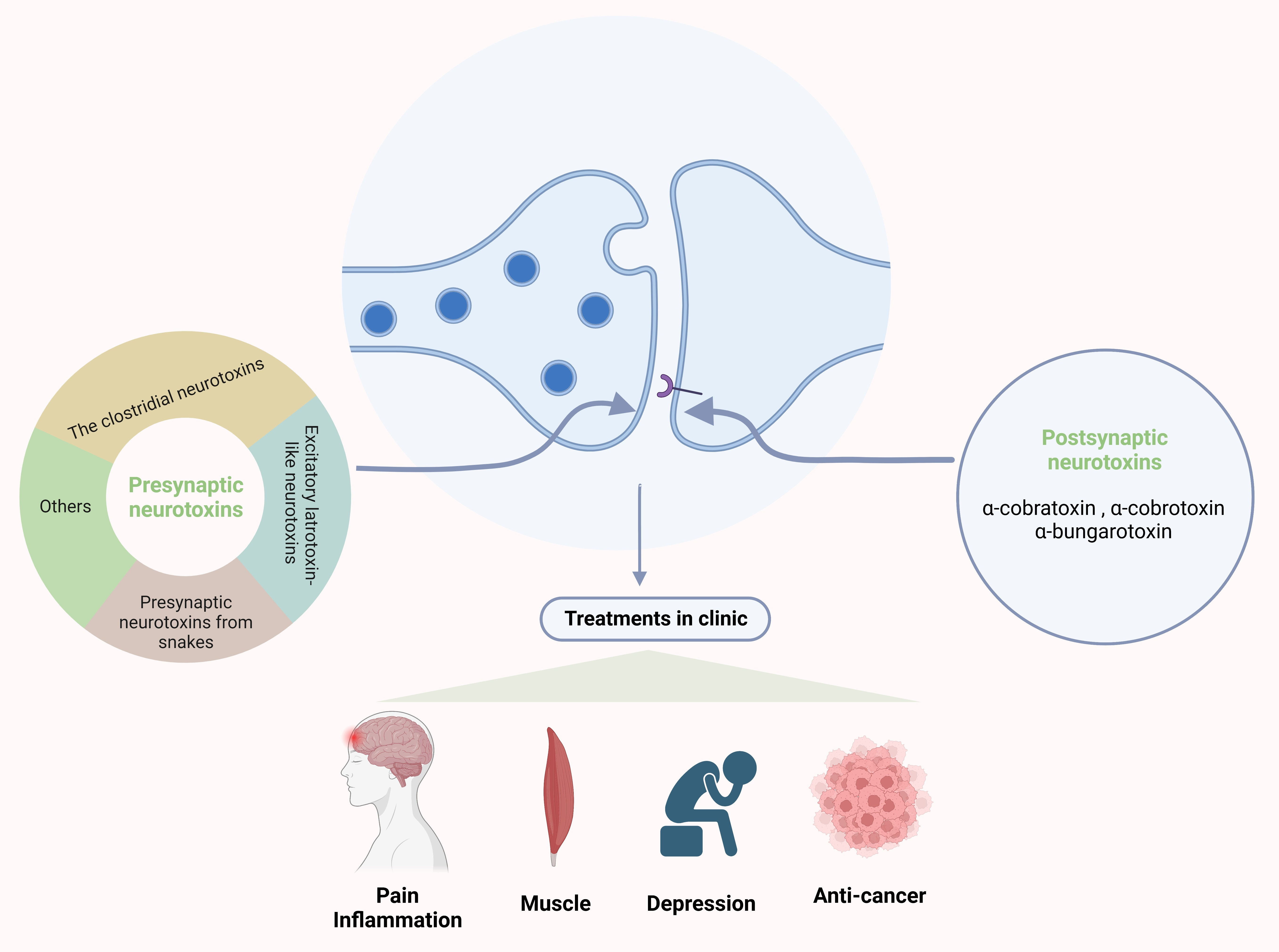{kind=link}
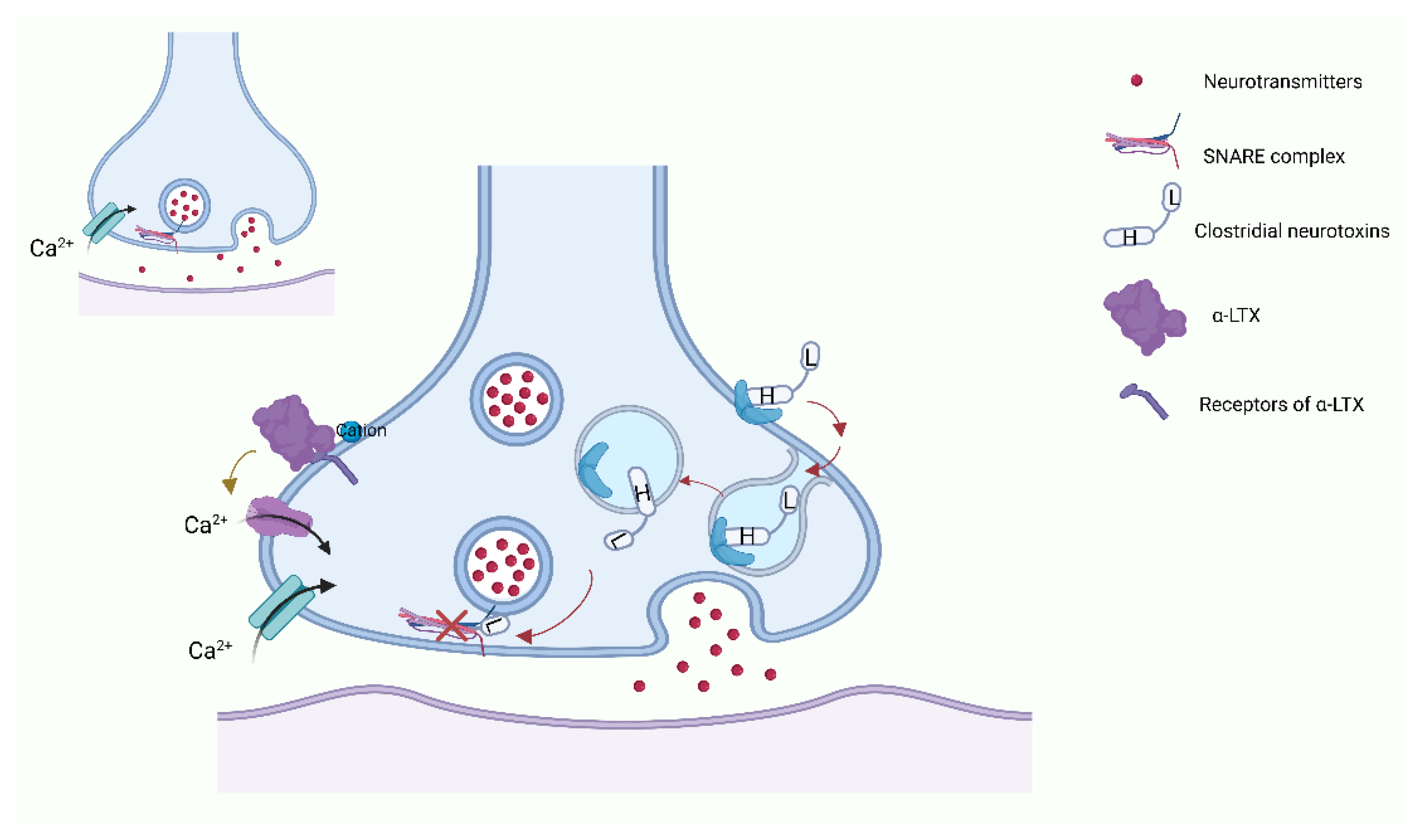{kind=link}
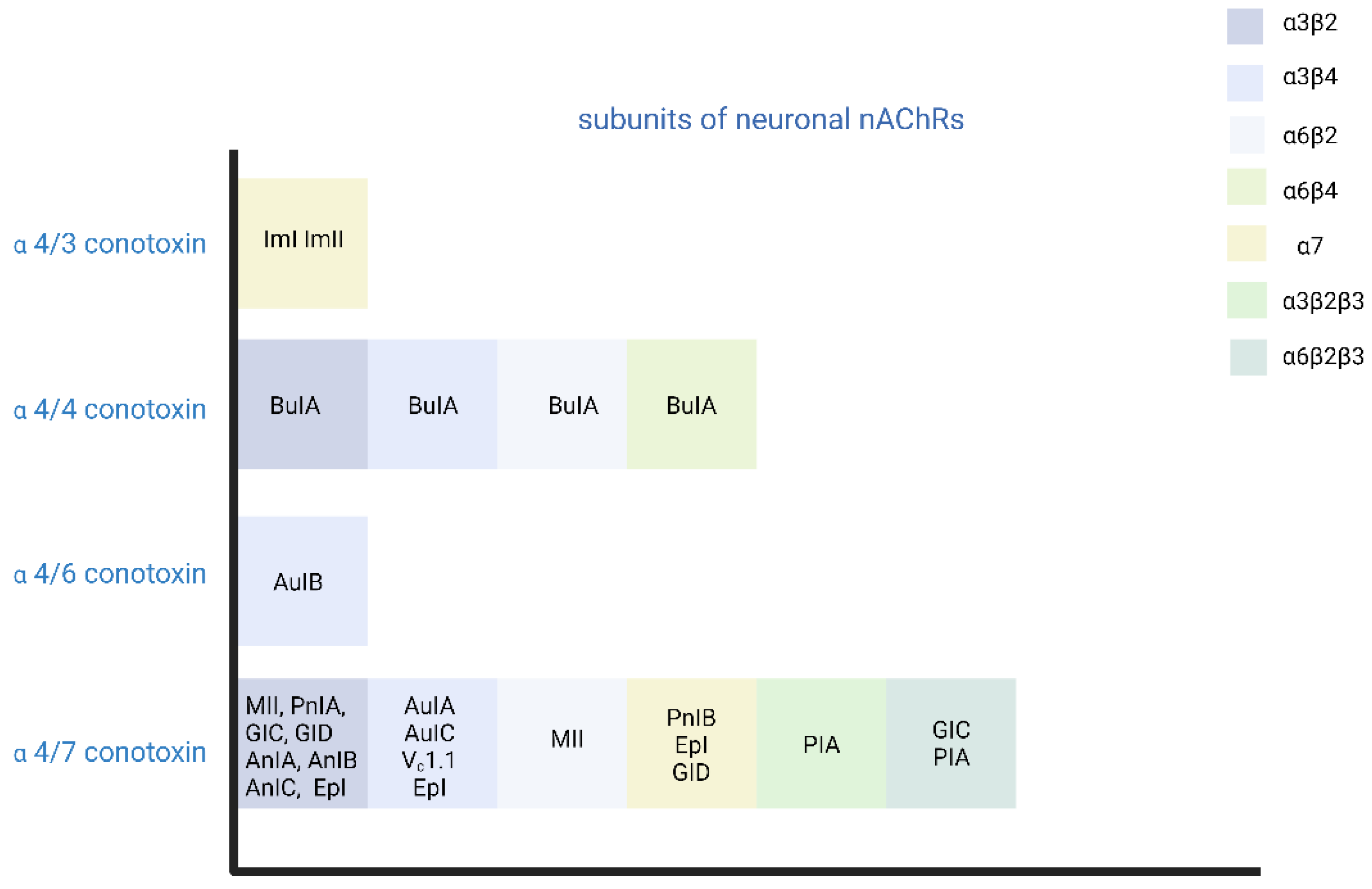{kind=link}
Table 1.
The summary of the clinical applications of neurotoxins (the “Indications” are found in the table).
Table 1.
The summary of the clinical applications of neurotoxins (the “Indications” are found in the table).
| Types of Neurotoxins | Trade Name (FDA Approved) | Indications (FDA Approved) | Clinical Applications | Reference | |
|---|---|---|---|---|---|
| BoNT | Botox (BoNT/A) Xeomin (BoNT/A) Dysport (BoNT/A) Myobloc/Neurobloc (BoNT/B) | Botox | Overactive bladder (OAB) with symptoms of urge urinary; Urinary incontinence due to detrusor overactivity associated with a neurologic condition (e.g., spinal cord injury (SCI), multiple sclerosis (MS)) in adults who have an inadequate response to or are intolerant of anticholinergic medication; Neurogenic detrusor overactivity (NDO) in pediatric patients 5 years of age and older who have an inadequate response to or are intolerant of anticholinergic medication; Prophylaxis of headaches in adult patients with chronic migraine (≥15 days per month with headache lasting 4 h a day or longer); Spasticity in patients 2 years of age and older; Cervical dystonia in adult patients; Severe axillary hyperhidrosis that is inadequately managed by topical agents in adult patients; Blepharospasm associated with dystonia in patients 12 years of age and older; Strabismus in patients 12 years of age and older. | Dystonic muscle contractions Neuropathic pain Neuroinflammation Depression (under investigation) Skin diseases Headache | [164] [155,156,165,166] [167] [168] [158] [169] |
| Xeomin | Chronic sialorrhea in patients 2 years of age and older; Upper limb spasticity in adults; Upper limb spasticity in pediatric patients 2 to 17 years of age, excluding spasticity caused by cerebral palsy; Cervical dystonia in adults; Blepharospasm in adults; Temporary improvement in the appearance of moderate-to-severe glabellar lines with corrugator and/or procerus muscle activity in adults. | ||||
| Dysport | Cervical dystonia in adults; Temporary improvement in the appearance of moderate-to-severe glabellar lines associated with procerus and corrugator muscle activity in adults < 65 years of age; Spasticity in patients 2 years of age and older. | ||||
| Myobloc/Neurobloc | Cervical dystonia to reduce the severity of abnormal head position and neck pain associated with cervical dystonia in adults; Chronic sialorrhea in adults. | ||||
| TeNT | Improve the motor functions Carrier to deliver into the CNS | [170] [171] | |||
| α-LTX | Type I diabetes (expected) | [172] | |||
| Snake Presynaptic Neurotoxins | Anticancer Antibacterial Antinociception | [173] [174] [175] | |||
| ω-agatoxin | Modulate the nociceptive process | [176] | |||
| Conotoxins | Prialt™ (Ziconitide) (a form of ω-conotoxin MVIIA) | Management of severe chronic pain in patients for whom intrathecal therapy is warranted and who are intolerant of or refractory to other treatment, such as systemic analgesics, adjunctive therapies, or intrathecal morphine. | Chronic pain (cancer- or AIDS-related neuropathy) Spinal cord injury | [177] [178] | |
| DTX | Diagnosis of neurodegenerative diseases (potential) | [179] [180] | |||
| Postsynaptic Neurotoxins | Cobratide | Disorders linked to NMJ dysfunction Anticancer Anti-inflammation Analgesic effect | [181] [182] [183] [183] | ||
Table 2.
Summary of the clinical applications of the FDA-approved neurotoxins.
| Applications | ||
|---|---|---|
| Botox | Muscle | Blepharospasm hemifacial spasm Strabismus cervical dystonia Upper limb and lower limb (adults) spasticity Bladder (neurogenic detrusor overactive (DO), overactive bladder (OB)) Forehead wrinkles |
| Other | Migraine | |
| Xeomin | Muscle | Cervical dystonia frown lines Blepharospasm upper limb spasticity |
| Other | Sialorrhea in adults | |
| Dysport | Muscle | Cervical dystonia Upper limb (adults) and lower limb (children + adults) spasticity Frown lines and wrinkles |
| Myobloc/Neurobloc | Cervical dystonia | |
| Prialt™ | Severe chronic pain | |
| Cobratide | Chronic pain | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhou, K.; Luo, W.; Liu, T.; Ni, Y.; Qin, Z. Neurotoxins Acting at Synaptic Sites: A Brief Review on Mechanisms and Clinical Applications. Toxins 2023, 15, 18. https://doi.org/10.3390/toxins15010018
AMA Style
Zhou K, Luo W, Liu T, Ni Y, Qin Z. Neurotoxins Acting at Synaptic Sites: A Brief Review on Mechanisms and Clinical Applications. Toxins. 2023; 15(1):18. https://doi.org/10.3390/toxins15010018
Chicago/Turabian StyleZhou, Kunming, Weifeng Luo, Tong Liu, Yong Ni, and Zhenghong Qin. 2023. "Neurotoxins Acting at Synaptic Sites: A Brief Review on Mechanisms and Clinical Applications" Toxins 15, no. 1: 18. https://doi.org/10.3390/toxins15010018
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.